
Submitted:
19 February 2025
Posted:
20 February 2025
You are already at the latest version
Abstract
Drosophila suzukii, commonly known as spotted-wing drosophila, has emerged as a highly destructive pests in global fruit and wine production. The effectiveness of chemical control is significantly compromised by rapid resistance development and limited range of insec-ticide options. Biological control presents a promising sustainable alternative., Our pre-vious work suggested the La Jolla virus (LJV) as a suitable candidate for the development of an insect virus-based control option. Here we characterized the natural transmission and pathology of the virus. We tested various modes of horizontal transmission, includ-ing airborne, venereal and oral, and fecal routes. To understand LJV pathology in infected flies, we studied feeding behavior and demonstrated changes in the food absorption compared to non-infected flies. We also investigated the impact on fecundity and egg-to-adult success rate. Altogether, these results collectively improve our understanding of LJV transmission in natural populations and the implication of infected flies in food ingestion and overall fitness.
Keywords:
Drosophila suzukii
; biological pest control
; La Jolla virus
; Iflavirus
; Transmission
; food intake
1. Introduction
Since the beginning of its invasion more than a decade ago (Garcia et al., 2022), Drosophila suzukii has spread across much of the globe (Hause, 2011; Depra et al.,2014), resulting in severe yield losses in many crops, such as cherries, berries, and grapes. Originally described in Japan and endemic to East Asia, this pest is now found on nearly every continent causing annual damages in the millions (Lee et al., 2011; Knapp et al., 2021). Whereas most other Drosophila species feed and oviposit on decaying fruit, D. suzukii females have a preference to lay eggs in intact ripening and ripe fruit with their specialized serrated ovipositor. D. suzukii has an exceptionally broad range of host plants targeting soft and ripening fruit across cultivated and wild environments (Fernández-Moreno et al., 2016; Bieńkowski & Orlova-Bienkowskaja, 2020). With a high reproduction rate of ~400 eggs per female and a rapid life cycle of 8 days, the species demonstrates extraordinary adaptability. Its ability to tolerate diverse environmental factors, adjust reproductive strategies based on temperature and photoperiod makes it a highly resilient pest capable of causing significant agricultural devastation if left unchecked (Little et al., 2020).
Traditional control methods for D. suzukii have relied heavily on conventional insecticides, including synthetic pyrethroids, organophosphates, spinosyns, and neonicotinoids. However, the application of synthetic insecticides comes with many disadvantages, including a high risk of insecticide resistance development (Haviland et al., 2012) and harmful effect on the environment and non-target organisms (Connolly, 2013; Aktar et al., 2009; Farooq and Pisante, 2019; Schetelig et al., 2017). Moreover, the D. suzukii larvae hatching inside the fruit are well protected from the environment and due to the timing of application prior to harvesting, difficult to treat. Thus, new and sustainable methods to control D. suzukii are urgently needed.
A promising strategy is the use of insect-specific viruses to control insect pests due to their generally high host specificity (Raj et al., 2022; Lee & Vilcinskas, 2017; Schetelig et al., 2017). In previous studies, we identified a potential virus candidate, the La Jolla Virus (LJV), a positive sense single stranded RNA virus and member of the Iflaviridae family (Webster et al., 2015; Carrau et al., 2018; Carrau et al; 2021). Initially discovered in a metagenomic approach in Drosophila melanogaster (Webster et al., 2015), LJV was subsequently isolated from moribund D. suzukii specimens in Germany. Intrathoracic injection of LJV caused significant mortality among adult D. suzukii flies (Carrau et al., 2018). Similarly, oral administration of LJV also led to significant mortality in a concentration dependent manner (Linscheid et al., 2022). LJV demonstrates superior efficacy as an RNA virus against Drosophilids compared to Drosophila C Virus (DCV), particularly when administered orally. While DCV causes only 10-25% mortality in Drosophila melanogaster via oral infection, even at high titers (Ferreira et al, 2014; Wong et al., 2015), LJV significantly reduces the lifespan of infected flies with all LJV exposed flies succumbing earlier than controls.
Our research aimed to elucidate the transmission dynamics of LJV in D. suzukii. Building on previous studies that demonstrated oral infection and vertical transmission (Linscheid et al., 2022), we conducted experiments to further characterize horizontal, vertical, and potential vector-mediated transmission routes (Cory, 2015). We investigated the impact of LJV epidemics on D. suzukii populations and examined how viral infection affects the pest's feeding behavior. These findings contribute to a more comprehensive understanding of LJV-host interactions, crucial for developing effective biocontrol strategies. A key question remains: can a chronic or natural LJV infection significantly impact wild populations of D. suzukii? This study aims to address this critical question by investigating whether persistent viral infections can influence population dynamics, behavior, and overall fitness of D. suzukii in natural settings, providing valuable insights into the potential of LJV as a biocontrol agent.
2. Materials and Methods
2.1. Drosophila suzukii Cultures Maintenance
The D. suzukii fly strain was derived from a laboratory colony established in the summer of 2012 in Ontario, Canada (Jakobs et al., 2015). From this line, we produced one LJV negative and one LJV positive subline. The LJV negative line was maintained in a climate chamber (26 °C, 60% relative humidity, 12-h photoperiod) and checked regularly for LJV as described in 2.2, while the infected flies were kept in an incubator (Binder, Neckarsulm, Germany) with the same ambient conditions (75 cm height x 60 cm width x 45cm depth). All flies were maintained on a standard fly diet composed of 60 g L−1 cornmeal, 8 g L−1 soy-bean flour, 18 g L−1 baker’s yeast, 60 g L−1 malt, 22 g/L molasses, 6.25 mL L−1 propionic acid, and 2 × 0.8 g nipagin, which is poured in 50-ml drosophila vials. All experiments were carried out on flies aged 3-7 days post eclosion.
2.2. Extraction and Quantification of La Jolla Virus in Flies
RNA was extracted using TRIZOL reagent (Thermo Fisher Scientific, Waltham, MA, USA) from the collected flies, 50 ng/µl aliquots were then prepared for the quantification. Quantification was done to assess the degree and progress of the infection in flies. Quantification was done via StepOne real-time PCR system (Applied Biosystems, Waltham, MA, USA) with the Luna Universal Probe One-Step RT-qPCR Kit (New England Biolabs, Ipswich, MA, USA), amplification was done via probes and primers described in Table 1. The Real time PCR was done with the following steps: an initial heating to 55 °C for 10 minutes then 95 °C for 1 minute. This was proceeded by 40 cycles of 95 ◦C for 10 seconds for denaturation and 60 °C for 30 seconds for annealing position 64–95 bp within the 128 bp product.
Viral infection levels were quantified using real-time PCR, with cycle threshold (Ct) values serving as the primary metric. Uninfected control flies consistently exhibited Ct values of approximately 30, while the highest infection levels observed corresponded to Ct values of around 10. This range of Ct values provided a robust measure of infection intensity across experimental conditions.
2.3. Transmission Assays
We arranged several experiments to determine if the transmission of the virus in a population of flies is airborne, venereal, oral, or fecal. For the airborne infection assay we placed two vials of uninfected flies on shelf at the top of the incubator in a 5 cm distance from the infected fly culture and two vials at the very bottom of the infected fly culture in a 40 cm distance from the infected fly culture. Flies were regularly collected every 3 days (5 flies per interval and location). To test the hypothesis of venereal transmission of LJV within a population, different combinations of 10 virgin females were paired with 10 males. The combinations were as follows: As controls, we paired infected males and females (IVF × IM) and non-infected males and females (NIM x NIVF) as positive and negative baselines, as test condition Non-Infected Virgin Female and Infected male (IM × NIVF), Infected Virgin female and Non-Infected male (NIM x IVF). As internal control, we set up pairings with same sex conditions to check for the requirement of the mating itself, or if factors such as close contact including oral or fecal route are sufficient for transmission. The pairings went as follows Non-infected Virgin Female x Infected Virgin Female (NIVF × IVF) and Non-Infected Male × Infected Male (NIM × IM). All flies were allowed to pair for 3 days before collecting 5 of each sex. To differentiate the LJV negative flies from the infected, we cut off the wings of the infected flies. For the oral transmission we placed 50 infected males on a vial of food and allowed them to feed for 3 days, then we removed the flies, flooded the vial with CO2, and used a UV cross linker (Stratagene Stratalinker 2400, U.S.A.) to surface sterilize the vial and the food within it. After this, 50 females and 50 males from the LJV negative flies were introduced and allowed to feed on the fly food. We then proceeded to collect 5 flies from both sexes every 3 days. For the fecal transmission, we placed a cotton ball saturated with 2 ml of 100 mM sucrose solution on parafilm in a completely empty drosophila vial. We then introduced 50 infected males and allowed them to feed and defecate on the walls of the vial for 3 days. After this we removed the flies, the cotton ball, and the parafilm. Then we thoroughly flooded the vial with CO2, added a fresh cotton ball on parafilm, introduced 50 LJV negative females and males to the vial, and allowed them to feed. Flies were collected every 3 days and the cotton ball was exchanged as necessary.
2.4. Fecundity and Egg to Adult Viability
To determine the influence of the virus on a fly population, we monitored fecundity and egg-to-adult viability of LJV positive flies compared to LJV negative flies. In short, we placed flies on grape juice agar plates (30% grape juice, 1% agar), transferring 100 eggs on a new agar plate. We then transferred the agar and placed it in a drosophila vial with food, then counted the larva after 6 days, the number of pupae, and the number of emerged adults.
2.5. The Feeding Behavior of Infected Flies
To assess the impact of viral infection on feeding behavior, we employed the flyPAD (fly Proboscis and Activity Detector) system, an automated high-resolution behavioral monitoring tool that uses capacitive sensors to detect and analyze feeding behavior in Drosophila (Itskov et al., 2014). The flyPAD system allows for the extraction of various feeding metrics, including the number of activity bouts, total duration of activity bouts, and number of sips, all of which correlate with food intake. The system's sensitivity enables the estimation of the volume of food consumed per sip, providing a comprehensive view of feeding dynamics in individual flies.
We adopted a slightly modified protocol based on Itskov et al. (2014). The experiment consisted of three runs each for females and males, comparing 24 La Jolla Virus (LJV) infected flies to 24 uninfected control flies per run. Prior to the experiment, flies were wet starved (females for 24 hours and males for 18 hours) to standardize hunger levels. Individual flies were then placed in the feeding arena of the flyPAD system. For each assay, we added 4 µl of either 5 mM or 50 mM sucrose solution to the reading electrode. Flies were allowed to feed for 1 hour, during which their interactions with the food were continuously monitored by the flyPAD system. The system recorded multiple behavioral metrics, including the number of activity bouts (approaches to food), total duration of activity bouts, and the number of sips. These parameters were used to estimate food intake and analyze feeding dynamics.
2.6. Statistical Analysis and Graph Design
All analysis was carried out via GraphPad Prism v9.1.2 for Windows (GraphPad Software, San Diego, CA, USA) for all the transmission experiments, a 2way ANOVA test was run with multiple comparisons against an alpha of 0.05. The life stage experiment was done via 2way ANOVA and the comparison was done via Šídák's multiple comparisons and compared to alphas of 0.05. The results and graphs of the behavior test are all calculated and plotted via the flyPAD program.
3. Results
3.1. Airborne Transmission
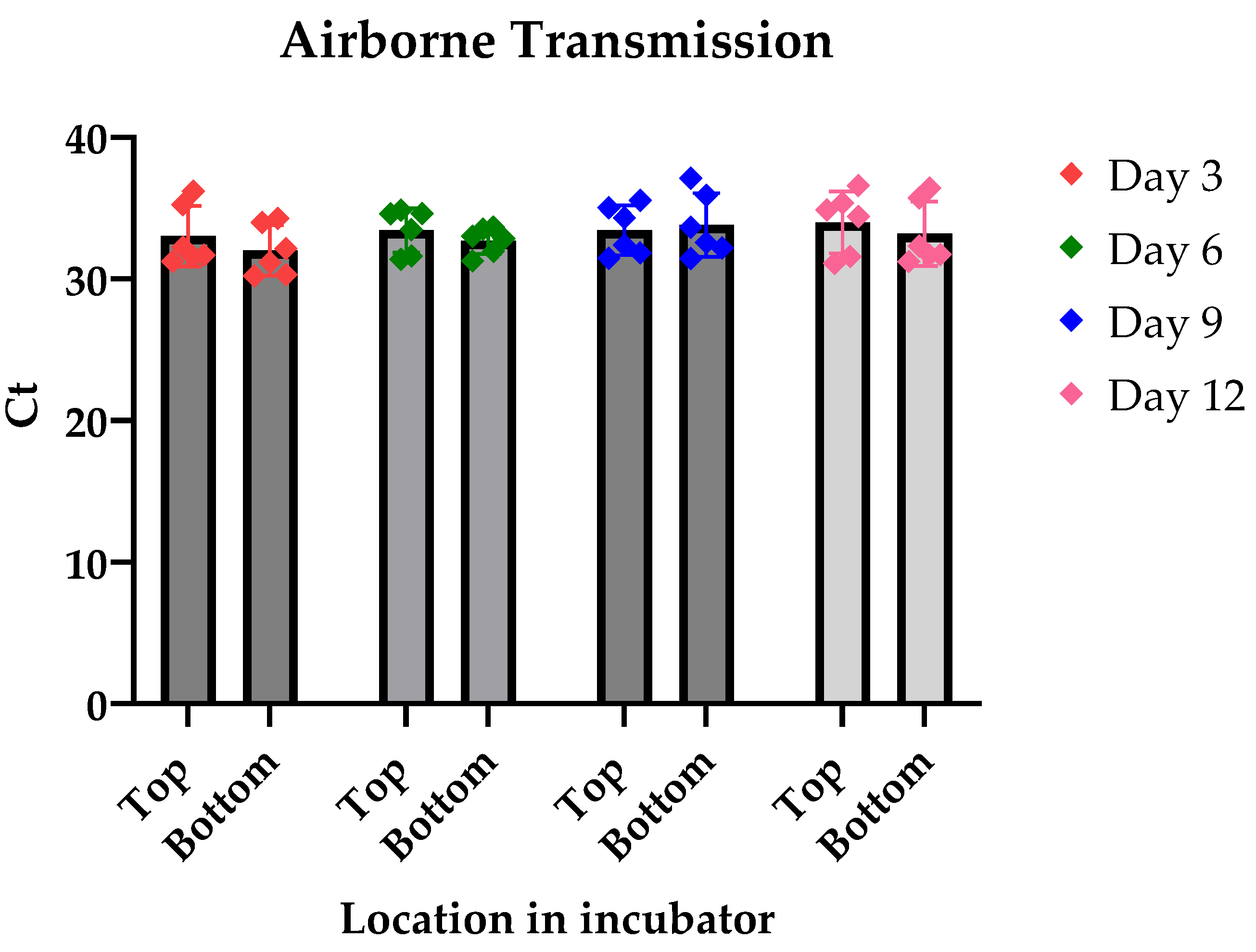
To investigate whether LJV is airborne transmitted, we raised non-infected close together with LJV-infected flies in a closed environment, i.e., an incubator with constant temperature and humidity settings. Our findings revealed that infected flies exhibited high titers of LJV, with a baseline mean infection Ct value of 9.9. In contrast, the adjacent non-infected flies maintained their uninfected status throughout the observation period, from day 3 to day 12 post-exposure. The Ct values for these non-infected flies remained relatively stable and significantly higher than the established baseline threshold, confirming their uninfected state.
Figure 1.
The location of the test tubes (top and bottom shelves) and the proximity to the LJV positive stock collection (white box).
Figure 1.
The location of the test tubes (top and bottom shelves) and the proximity to the LJV positive stock collection (white box).

Figure 2.
Figure shows the results of the Realtime PCR after several days of the flies being in the vicinity of an infected fly culture. The graph demonstrates no significant change in the Ct of the extracted samples concluding there was no infection. And result show no significant difference in the infection levels as the days went on. P-value = 0.38 Alpha = 0.05.
Figure 2.
Figure shows the results of the Realtime PCR after several days of the flies being in the vicinity of an infected fly culture. The graph demonstrates no significant change in the Ct of the extracted samples concluding there was no infection. And result show no significant difference in the infection levels as the days went on. P-value = 0.38 Alpha = 0.05.
3.2. Venereal Transmission
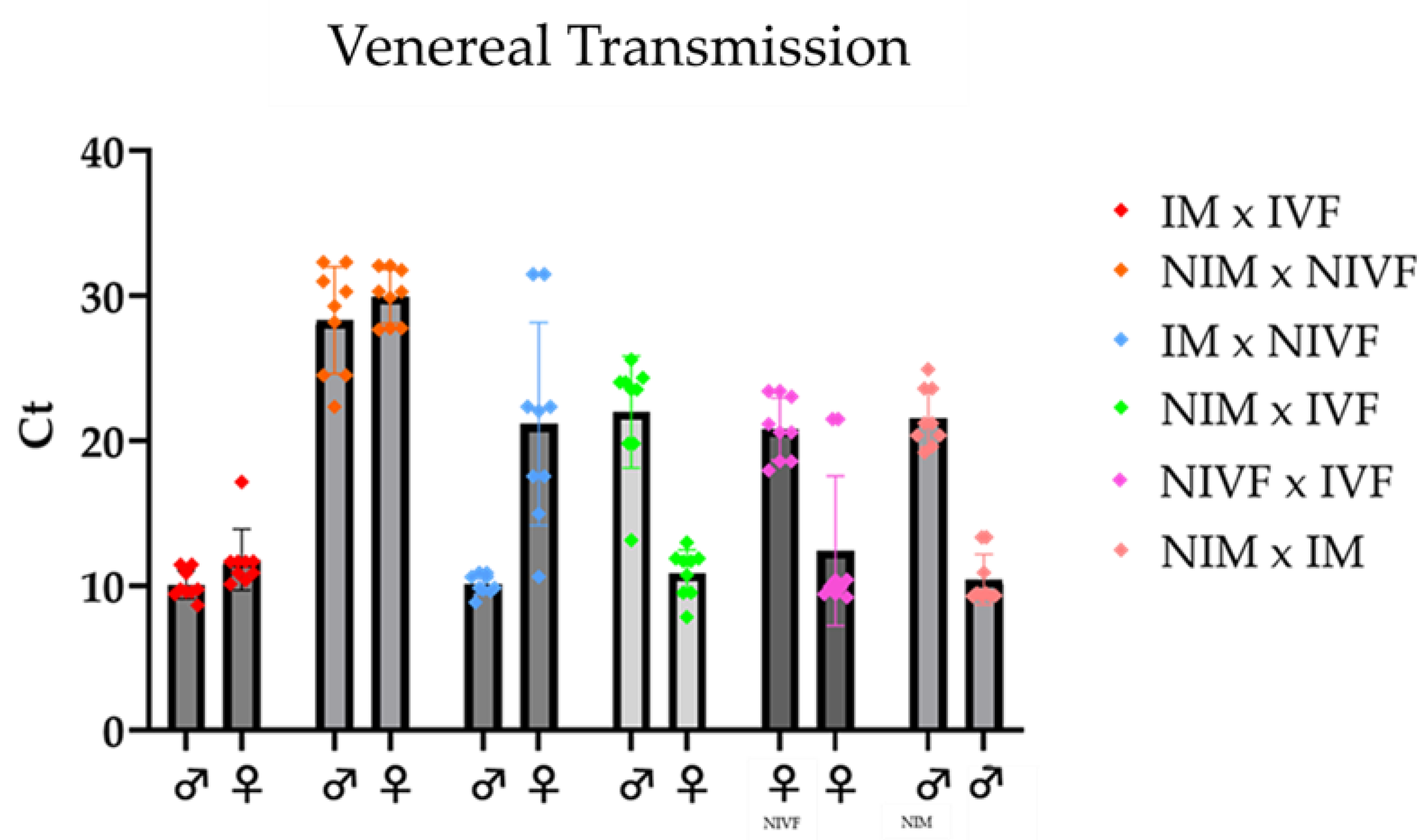
Pairing experiments revealed distinct viral transmission patterns. Non-infected fly pairs maintained their uninfected status, as evidenced by high Ct values (approximately 30) in RT-qPCR analysis. In contrast, pairings involving infected flies resulted in high levels of LJV transmission, indicated by low Ct values (around 10). Notably, same-sex pairings showed transmission rates comparable to female-male pairings. This similarity suggests that virus transmission occurs primarily through contact, likely via fecal or oral routes, rather than through venereal transmission.
Figure 3.
The infection levels after the flies were allowed to mate for 3 days. The first two sets are the positive (IM×IVF, red dots) and negative controls (NIM×NIVF, orange dots), followed by the different combinations tested. All infection levels are significant compared to the negative control (NIVF + NIM) alpha = 0.05.
Figure 3.
The infection levels after the flies were allowed to mate for 3 days. The first two sets are the positive (IM×IVF, red dots) and negative controls (NIM×NIVF, orange dots), followed by the different combinations tested. All infection levels are significant compared to the negative control (NIVF + NIM) alpha = 0.05.
3.3. The Transmission of the Virus Occurs by Oral and Fecal Contamination
The experimental outcomes for fecal and oral transmission routes showed striking similarities when assessing infection rates after several days of exposure (Figure 4 a & b). Control flies maintained their non-infected status, as evidenced by high Ct values (approximately 30) in RT-qPCR analysis. In contrast, flies exposed to either feces or food from LJV-infected individuals exhibited significantly lower Ct values (approximately 20), indicating successful virus transmission through these routes. Notably, the transmission efficiency appeared to follow a time-dependent trend, with higher transmission rates observed after longer incubation periods. This temporal pattern suggests a gradual accumulation of viral particles in the exposed flies, potentially leading to more robust infections over time. These results collectively suggest the effectiveness of both fecal and oral routes in LJV transmission among D. suzukii populations, highlighting the virus's capacity for efficient horizontal spread through environmental contamination.
3.4. Chronic LJV Infection Decreases Egg-to-Adult Success Rate
Our results demonstrate a significant reduction in the percentage of emerged adults due to the impact of the virus (Figure 5). While all eggs successfully hatched, differences began to emerge at the larval stage, particularly when larvae reached the L3 stage. The virus seems to disrupt normal larval growth, resulting in a gradually widening gap in survival rates when compared to uninfected controls. Although significant differences were observed across all developmental stages, the most pronounced effect was seen during adult emergence. At this stage, the virus caused a substantial decline of around 40% in the number of adults successfully completing pupation and emerging.
3.5. LJV Affects the Feeding Behavior of Female Flies
We employed the flyPAD (fly Proboscis and Activity Detector) system to assess whether LJV infection alters the feeding behavior of D. suzukii. Our results reveal significant changes in the feeding patterns of infected females compared to uninfected controls.
The feeding behavior of non-infected and the infected flies was evaluated for both genders (Figure 6A and 6B) using the cumulative duration of activity bouts. This metric, representing the total time engaged in physical activity over multiple bouts during a one-hour period (x-axis in Figure 6A & B) and represents in the FlyPad system the sum of all individual activity bout durations recorded for a fly during the observation period), is calculated in the FlyPad system by summing all individual activity bout durations for each fly. This measure effectively captures the full spectrum of fly movement patterns, from brief exploratory actions to extended locomotion periods, making it valuable for assessing overall activity levels. While female flies exhibited a notable difference, males showed no significant variation.
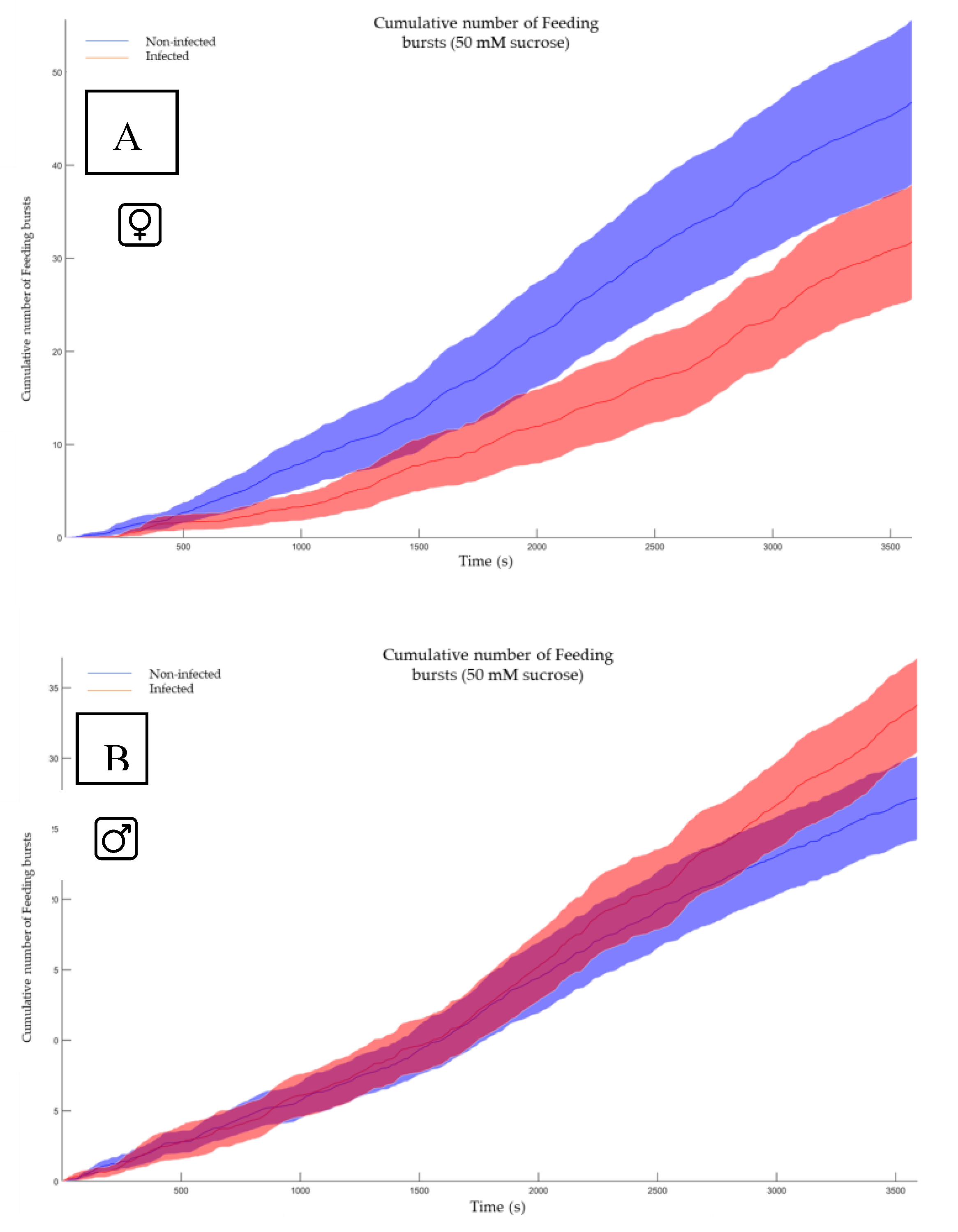
In contrast, the cumulative number of feeding bursts represents the total count of discrete feeding events recorded by the FlyPad system over a specified observation period. In Drosophila studies using FlyPad technology, a feeding burst is typically defined as a short, intense period of proboscis extension and food interaction. This metric provides a quantitative measure of feeding frequency, offering insights into the flies' overall food consumption patterns and feeding behavior. By summing these individual feeding events, the total feeding activity, can be assessed which may be influenced by factors such as the LJV infection status. The cumulative number of feeding bursts serves as an indicator of feeding drive and can be used to compare feeding behaviors across different experimental conditions. In our investigation, no significant difference (p-value 0.068 for females; p-value 0.12 for males) was observed between noninfected and infected flies, suggesting no substantial variation in feeding uptake.
Figure 6.
Cumulative duration of activity bouts. A) The feeding activity of the non-infected females (blue) vs the infected females (red). The software conducted a Mann-Whitney test and shows a p-value of 0.036. B) The feeding activity of the non-infected males (blue) vs the infected males (red) The software conducted a Mann-Whitney test and shows a p-value of 0.93.
Figure 6.
Cumulative duration of activity bouts. A) The feeding activity of the non-infected females (blue) vs the infected females (red). The software conducted a Mann-Whitney test and shows a p-value of 0.036. B) The feeding activity of the non-infected males (blue) vs the infected males (red) The software conducted a Mann-Whitney test and shows a p-value of 0.93.
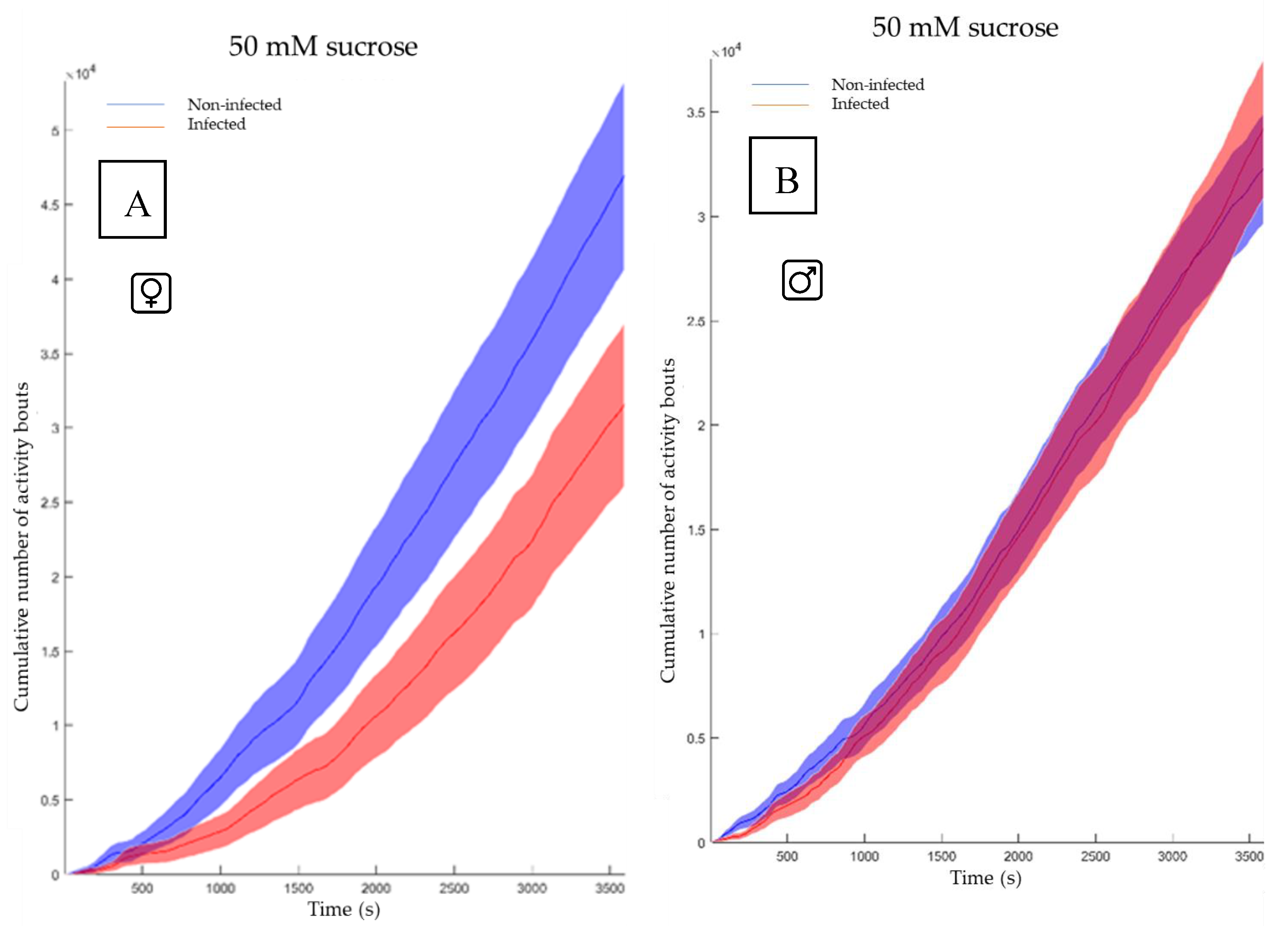
Figure 7.
Shows the cumulative number of feeding bursts. A) The feeding activity of the non-infected females (blue) vs the infected females (red). The software conducted a Mann-Whitney test and shows a p-value of 0.068. B) The feeding activity of the non-infected males (blue) vs the infected males (red) The software conducted a Mann-Whitney test and shows a p-value of 0.12.
Figure 7.
Shows the cumulative number of feeding bursts. A) The feeding activity of the non-infected females (blue) vs the infected females (red). The software conducted a Mann-Whitney test and shows a p-value of 0.068. B) The feeding activity of the non-infected males (blue) vs the infected males (red) The software conducted a Mann-Whitney test and shows a p-value of 0.12.
4. Discussion
We studied the transmission routes and pathologies of the La Jolla virus (LJV) in its host Drosophila suzukii. Although insect viruses typically do not spread through the air, it is worth noting that certain human respiratory viruses, such as coronaviruses, influenza viruses, and rhinoviruses (Shao et al., 2021; Chang et al., 2023) can be transmitted via airborne routes. Our research suggests that airborne transmission does not occur, at least under the specific laboratory conditions we used. The study’s experimental findings validate that LJV primarily spreads through the oral-fecal route, with venereal transmission likely playing a secondary role. This finding is consistent with prior studies on iflaviruses, which have demonstrated diverse transmission strategies, including horizontal (oral-fecal) and vertical routes, depending on the host species and environmental conditions. For example, Euscelidius variegatus iflavirus 1 (EVV-1) primarily spreads via fecal-oral transmission, as observed in Nilaparvata lugens honeydew virus-1 and Nora Virus in other insect species (Ottati et al., 2020; Habayeb et al., 2009). Deformed Wing Virus (DWV), an Iflaviridae family member akin to LJV, illustrates the complex transmission dynamics and infection effects seen in insect RNA viruses (de Miranda & Genersch, 2010). The transmission routes and infection outcomes of DWV are among the most extensively studied within the Iflaviridae family, offering valuable insights into the ecology of these viruses. The entoparasitic mite Varroa destructor serves as a crucial vector for DWV, substantially influencing its virulence. The concurrent presence of DWV and Varroa mites results in symptoms like pupal mortality and the characteristic deformed wings of worker bees (Lanzi et al., 2006). Although LJV appears to infect without a vector, similar symptoms such as pupal death can be observed in D. suzukii LJV infections. A common effect is the severely shortened adult life span in an acute DWV infection. In mite-free conditions, DWV develops into a persistent, covert (chronic-asymptomatic) infection. In contrast, LJV seems to establish a persistent and chronic symptomatic in the absence of a vector, significantly impacting population dynamics by reducing offspring numbers and fitness of the emerging flies. This outcome is very important, since an application of LJV as biocontrol agent would also be able to spread chronically within a population and maintaining their virulence.
Next, we explored the possibility of venereal transmission of the virus. Venereal transmission of arboviruses has been documented for several cases (Heinig-Hartberger et al., 2023; Ovenden & Mahon, 1984; Shroyer, 1990; De Miranda & Fries, 2008). To test this hypothesis, we paired infected flies with their mates and monitored for infections. All flies paired with infected partners showed initial signs of infection, which was a promising indicator. However, infection was also observed in same-sex pairings, suggesting potential environmental transmission rather than direct sexual contact. To confirm sexual transmission conclusively, future studies should involve dissecting and examining sexual organs for viral presence. If confirmed, sexual transmission could be a valuable mechanism, especially when combined with other techniques, such as the Sterile Insect Technique (SIT). Sterile infected males, that have been sterilized with eco-friendly methods (Abdelhafiz et al., 2023), could be released as viral vectors to disseminate the infection throughout the population. While venereal transmission cannot be excluded for LJV, its significance appears to be secondary to the oral-fecal route. Studies on other insect-specific viruses, such as Culex flavivirus (CxFV), have shown that venereal transmission plays a minor role compared to vertical and horizontal routes (Bolling et al., 2012). Similarly, recent research on medfly-associated iflaviruses indicate that while vertical transmission via females is predominant, male-mediated venereal transmission is possible but less efficient (Hernandez-Pelegrín et al., 2024). Studies comparing iflaviruses have revealed diverse transmission methods within this viral family. While some iflaviruses, like the Antheraea pernyi Vomit Disease virus (Geng et al., 2017) and Spodoptera exigua iflavirus (Virto, 2014), primarily transmit vertically through eggs or larvae, this study did not focus on vertical transmission. However, its potential role in LJV epidemiology deserves further exploration.
The predominance of oral-fecal transmission has significant implications for understanding host-virus interactions and population-level dynamics. Insects with gregarious feeding behaviors or those inhabiting densely populated environments are particularly susceptible to rapid viral spread via contaminated food or substrates. The systemic nature of infection ensures that all life stages contribute to environmental contamination. This continuous viral shedding increases the chances of sustained viral circulation within host populations, even in the absence of overt disease symptoms. Additionally, investigating how environmental factors like temperature or humidity affect fecal-oral transmission efficiency could provide valuable context for understanding seasonal variations in infection rates. Future research should also investigate potential interactions between LJV and other pathogens or microbiota within the host gut. As demonstrated by SeIV1's interaction with baculoviruses, such associations may modulate viral infectivity and persistence.
We subsequently evaluated the complete life cycle of both uninfected and infected flies to determine their egg-to-adult survival rate. This investigation builds upon a previous study (Linscheid et al., 2022) which demonstrated that oral infection with LJV negatively affects fly emergence. Our aim was to verify these findings and explore the potential impact on a chronically infected fly population, thereby demonstrating the potential of LJV as a biocontrol agent. Our findings revealed that LJV significantly impairs the adult emergence rate in fly populations. This is in contrast to the picorna-like Nora virus, which can establish infections in laboratory fly strains and persist for several years, without causing any significant pathological effects (Habayeb et al., 2006). The persistent presence of LJV in the environment could serve as a continuous control measure, which, when combined with other complementary methods, may lead to population suppression. The results indicate that the virus has its most significant effect during the later developmental stages, possibly affecting essential physiological or metabolic functions required for pupation and adult emergence. This underscores the virus's capacity to influence population dynamics by decreasing reproductive success and adult survival rates in D. suzukii, even in cases of long-term, chronic infection within the population.
Finally, we examined the virus's effect on fly feeding behavior. Our findings indicate that the virus has a more pronounced impact on female flies, with infected females taking more time to find food compared to their uninfected counterparts. This suggests a reduced drive or motivation to seek out food sources among infected females. This observation is particularly relevant in natural environments where food is likely to be more dispersed and limited, potentially affecting fly survival rates. Interestingly, male flies do not exhibit the same response to the virus, showing no significant differences in feeding behavior.
Our study reveals that while infected flies spend more time locating food, their overall food consumption remains unaffected. Although the statistical difference was marginal (p-value slightly exceeding the 0.05 alpha threshold), these findings could have significant implications in natural settings, especially in areas with scarce food resources. In field conditions with regular maintenance and fewer hiding spots for flies, we anticipate observable effects, particularly given the infected flies' reduced motivation to search for food. Our research also highlights opportunities for further investigation into how the virus influences D. suzukii feeding behavior. Future studies could explore whether the virus affects specific receptors and if so, which ones. Additionally, research could examine if the virus impacts fly mobility and whether different food types alter the foraging behavior of infected flies.
Viruses offer a promising alternative due to their specificity and effectiveness, and LJV has shown significant antagonistic effects against D. suzukii. However, like other insect viruses, the application of LJV presents its own set of challenges. Previous research has demonstrated that, once infected, the virus can kill the insect within 5-8 days (Linscheid et al., 2022). This relatively slow action may limit its immediate impact in controlling infestations. To address these challenges, studying the transmission dynamics of LJV within D. suzukii populations is a prerequisite for a controlled release in natural environments. Conducting field trials is crucial for gaining insights in natural settings, as laboratory conditions often fail to capture the complexities of real-world environments. Furthermore, developing viral formulations to improve stability and delivery, or employing strategies like continuously infecting food sources with viral particles, could strengthen the practical application of LJV as a biocontrol agent. Additionally, care must be taken to minimize off-target effects, as members of the Iflaviridae (Chen et al., 2012) are known antagonists to bees, raising ecological concerns. Combining LJV with other management techniques such as SIT could maximize its efficacy and reduce the likelihood of resistance developing in D. suzukii population.
In summary, our study highlights the critical importance of the oral-fecal route as the primary mechanism for transmitting LJV and the long-term effects of chronic infection including feeding behavior change on population level. These findings align with existing literature on iflaviruses and provide a foundation for further exploration into host-virus dynamics and ecological implications. Understanding these transmission pathways is essential for developing targeted strategies to manage infections caused by LJV and related viruses in insect populations.
Author Contributions
Conceptualization, K.-Z.L. and A.V.; methodology, K.-Z.L., I.A., and T.K.; validation, K.-Z.L., and I.A.; formal analysis, I.A.; investigation, I.A.; writing—original draft preparation, I.A.; writing—review and editing, I.A., K.-Z.L. and A.V.; visualization, I.A.; supervision, K.-Z.L. and A.V.; project administration, K.-Z.L.; funding acquisition, K.-Z.L. All authors have read and agreed to the published version of the manuscript.
Funding
The project was supported by the Federal Ministry of Food and Agriculture (BMEL) based on a decision of the Parliament of the Federal Republic of Germany via the Federal Office for Agriculture and Food (BLE) under the innovation support program number 2818807A19.
Institutional Review Board Statement
Not applicable
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
We would like to thank Pavel M. Itskov (Lisbon, Portugal) for support in the development of the food intake assay and fruitful discussions.
Conflicts of Interest
The authors declare no conflict of interest
Abbreviations
The following abbreviations are used in this manuscript:
| LJV | La Jolla Virus |
| DWV DCV |
Deformed Wing Virus Drosophila C Virus |
References
- Garcia, F. R. M.; Lasa, R.; Funes, C. F.; Buzzetti, K. (2022). Drosophila suzukii Management in Latin America: Current Status and Perspectives. Journal of Economic Entomology, 115(4), 1008–1023. [CrossRef]
- Hauser, M. (2011). A historic account of the invasion of Drosophila suzukii (Matsumura) (Diptera: Drosophilidae) in the continental United States, with remarks on their identification. Pest Management Science, 67(11), 1352–1357. [CrossRef]
- Deprá, M.; Poppe, J. L.; Schmitz, H. J.; De Toni, D. C.; Valente, V. L. S. (2014). The first records of the invasive pest Drosophila suzukii in the South American continent. Journal of Pest Science, 87(3), 379–383. [CrossRef]
- Lee, J. C.; Bruck, D. J.; Dreves, A. J.; Ioriatti, C.; Vogt, H.; Baufeld, P. (2011). In Focus: Spotted wing drosophila, Drosophila suzukii, across perspectives. Pest Management Science, 67(11), 1349–1351. [CrossRef]
- Knapp, L.; Mazzi, D.; Finger, R. (2021). The economic impact of Drosophila suzukii: perceived costs and revenue losses of Swiss cherry, plum and grape growers. Pest Management Science, 77(2), 978–1000. [CrossRef]
- Fernández-Moreno, M.A.; Farr, C.L.; Kaguni, L.S.; Garesse, R. (2007). Drosophila melanogaster as a Model System to Study Mitochondrial Biology. In: Leister, D., Herrmann, J.M. (eds) Mitochondria. Methods in Molecular Biology™, vol 372. Humana Press. [CrossRef]
- Bieńkowski, A. O.; Orlova-Bienkowskaja, M. J. (2020). Invasive agricultural pest Drosophila suzukii (Diptera, drosophilidae) appeared in the russian caucasus. Insects, 11(11), 1–7. [CrossRef]
- Little, C. M.; Little, C. M.; Chapman, T. W.; Hillier, N. K. (2020). Plasticity Is Key to Success of Drosophila suzukii (Diptera: Drosophilidae) Invasion. Journal of Insect Science, 20(3). [CrossRef]
- Haviland, D. R.; Beers, E. H. (2012). Chemical control programs for Drosophila suzukii that comply with international limitations on pesticide residues for exported sweet cherries. Journal of Integrated Pest Management, 3(2), 1–6. [CrossRef]
- Connolly, C. (2013). The risk of insecticides to pollinating insects. Communicative & Integrative Biology, 6(5), e25074. [CrossRef]
- Aktar, W.; Sengupta, D.; Chowdhury, A. (2009). Impact of pesticides use in agriculture: Their benefits and hazards. Interdisciplinary Toxicology, 2(1), 1–12. [CrossRef]
- Farooq, M.; Pisante, M. (2019). Innovations in sustainable agriculture. In Innovations in Sustainable Agriculture. [CrossRef]
- Schetelig, M. F.; Lee, K. Z.; Otto, S.; Talmann, L.; Stökl, J.; Degenkolb, T.; Halitschke, R. (2017). Environmentally sustainable pest control options for Drosophila suzukii. Journal of Applied Entomology, 142(1–2), 3–17. [CrossRef]
- Nikhil Raj, M.; Samal, I.; Paschapur, A.; Subbanna, A. R. N. S. (2022). Entomopathogenic viruses and their potential role in sustainable pest management. New and Future Developments in Microbial Biotechnology and Bioengineering: Sustainable Agriculture: Revitalization through Organic Products, September, 47–72. [CrossRef]
- Lee, K. Z.; Vilcinskas, A. (2017). Analysis of virus susceptibility in the invasive insect pest Drosophila suzukii. Journal of invertebrate pathology, 148, 138–141. [CrossRef]
- Webster; C. L.; Waldron, F. M.; Robertson, S.; Crowson, D.; Ferrari, G.; Quintana, J. F.; Brouqui, J. M.; Bayne, E. H.; Longdon, B.; Buck, A. H.; Lazzaro, B. P.; Akorli, J.; Haddrill, P. R.; Obbard, D. J. (2015). The discovery, distribution, and evolution of viruses associated with Drosophila melanogaster. PLoS Biology, 13(7), 1–33. [CrossRef]
- Carrau, T.; Hiebert, N.; Vilcinskas, A.; Lee, K.Z. (2018). Identification and characterization of natural viruses associated with the invasive insect pest Drosophila suzukii. Journal of Invertebrate Pathology. 154. [CrossRef]
- Carrau, T.; Lamp, B.; Reuscher, C. M.; Vilcinskas, A.; Lee, K. Z. (2021). Organization of the structural protein region of la jolla virus isolated from the invasive pest insect Drosophila suzukii. Viruses, 13(5). [CrossRef]
- Linscheid, Y.; Kessel, T.; Vilcinskas, A.; Lee, K. Z. (2022). Pathogenicity of La Jolla Virus in Drosophila suzukii following Oral Administration. Viruses, 14(10), 2158. [CrossRef]
- Ferreira, Á. G.; Naylor, H.; Esteves, S. S.; Pais, I. S.; Martins, N. E.; Teixeira. (2014). The Toll-Dorsal Pathway Is Required for Resistance to Viral Oral Infection in Drosophila. PLoS Pathogens, 10(12). [CrossRef]
- Wong, Z. S.; Brownlie, J. C.; Johnson, K. N. (2016). Impact of ERK activation on fly survival and Wolbachia-mediated protection during virus infection. Journal of General Virology, 97(6), 1446–1452. [CrossRef]
- Cory, J. S. (2015). Insect virus transmission: Different routes to persistence. Current Opinion in Insect Science, 8, 130–135. [CrossRef]
- Jakobs, R.; Gariepy, T. D.; Sinclair, B. J. (2015). Adult plasticity of cold tolerance in a continental-temperate population of Drosophila suzukii. Journal of insect physiology, 79, 1–9. [CrossRef]
- Itskov, P. M.; Moreira, J. M.; Vinnik, E.; Lopes, G.; Safarik, S.; Dickinson, M. H.; Ribeiro, C. (2014). Automated monitoring and quantitative analysis of feeding behaviour in Drosophila. Nature Communications, 5. [CrossRef]
- Shao, L.; Ge, S.; Jones, T.; Santosh, M.; Silva, L; F. O.; Cao, Y.; Oliveira, M. L. S.; Zhang, M.; BéruBé, K. (2021). The role of airborne particles and environmental considerations in the transmission of SARS-CoV-2. Geoscience frontiers, 12(5), 101189. [CrossRef]
- Chang, Y.; Wang, Y.; Li, W.; Wei, Z.; Tang, S.; Chen, R. Mechanisms, Techniques and Devices of Airborne Virus Detection: A Review. Int. J. Environ. Res. Public Health 2023, 20, 5471. [CrossRef]
- Ottati S, Persico A; Rossi M; Bosco D; Vallino M; Abbà S; Molinatto G; Palmano S; Balestrini R; Galetto L; Marzachì C. Biological characterization of Euscelidius variegatus iflavirus 1. J Invertebr Pathol. 2020 ;173:107370. [CrossRef]
- Heinig-Hartberger, M.; Hellhammer, F.; Zöller, D.D.J.A.; Dornbusch, S.; Bergmann, S.; Vocadlova, K.; Junglen, S.; Stern, M.; Lee, K.-Z.; Becker, S.C. Culex Y Virus: A Native Virus of Culex Species Characterized In Vivo. Viruses 2023, 15, 235. [CrossRef]
- Ovenden, J. R.; Mahon, R. J. (1984). Venereal transmission of Sindbis virus between individuals of Aedes australis (Diptera: Culicidae). Journal of medical entomology, 21(3), 292–295. [CrossRef]
- Shroyer D. A. (1990). Venereal transmission of St. Louis encephalitis virus by Culex quinquefasciatus males (Diptera: Culicidae). Journal of medical entomology, 27(3), 334–337. [CrossRef]
- Habayeb, M. S.; Cantera, R.; Casanova, G.; Ekström, J. O.; Albright, S.; Hultmark, D. (2009). The Drosophila Nora virus is an enteric virus; transmitted via feces. Journal of Invertebrate Pathology, 101(1), 29–33. [CrossRef]
- de Miranda, J. R.; Genersch, E. (2010). Deformed wing virus. Journal of Invertebrate Pathology, 103(SUPPL. 1), S48–S61. [CrossRef]
- Lanzi, G.; de Miranda, J. R.; Boniotti, M. B.; Cameron, C. E.; Lavazza, A.; Capucci, L.; Camazine, S. M.; Rossi, C. (2006). Molecular and Biological Characterization of Deformed Wing Virus of Honeybees (Apis mellifera L.). Journal of Virology, 80(10), 4998–5009. [CrossRef]
- de Miranda, J. R.; Fries, I. (2008). Venereal and vertical transmission of deformed wing virus in honeybees (Apis mellifera L.). Journal of Invertebrate Pathology, 98(2), 184–189. [CrossRef]
- Abdelhafiz, I.; Gerth, S.; Claussen, J.; Weule, M.; Hufnagel, E.; Vilcinskas, A.; Lee, K. Z. (2024). Radioactivity and GMO-Free Sterile Insect Technology for the Sustainable Control of the Invasive Pest Drosophila suzukii. Advanced biology, 8(7), e2400100. [CrossRef]
- Bolling, B. G.; Olea-Popelka, F. J.; Eisen, L.; Moore, C. G.; Blair, C. D. (2012). Transmission dynamics of an insect-specific flavivirus in a naturally infected Culex pipiens laboratory colony and effects of co-infection on vector competence for West Nile virus. Virology, 427(2), 90–97. [CrossRef]
- Hernández-Pelegrín, L.; Huditz, H.-I.; García-Castillo, P.; de Ruijter, N. C. A.; van Oers, M. M.; Herrero, S.; Ros, V. I. D. (2024). Covert RNA viruses in medflies differ in their mode of transmission and tissue tropism. Journal of Virology, 98(6), e0010824. [CrossRef]
- Geng, P.; Li, W.; de Miranda, J. R.; Qian, Z.; An, L.; Terenius, O. (2017). Studies on the transmission and tissue distribution of Antheraea pernyi iflavirus in the Chinese oak silkmoth Antheraea pernyi. Virology, 502, 171–175. [CrossRef]
- Virto, C.; Navarro, D.; Tellez, M. M.; Herrero, S.; Williams, T.; Murillo, R.; Caballero, P. (2014). Natural populations of Spodoptera exigua are infected by multiple viruses that are transmitted to their offspring. Journal of invertebrate pathology, 122, 22–27. [CrossRef]
- Habayeb, M. S.; Ekengren, S. K.; Hultmark, D. (2006). Nora virus, a persistent virus in Drosophila, defines a new picorna-like virus family. Journal of General Virology, 87(10), 3045–3051. [CrossRef]
- Chen, Y. P.; Becnel, J. J.; Valles, S. M. (2012). RNA Viruses Infecting Pest Insects. In Insect Pathology, Second Edition (Second Edition). Elsevier Inc. [CrossRef]
Figure 4.
The infection levels in the flies (A. Fecal transmission B. Oral Transmission) after several days of infection. The figure shows a steady infection rate in both experiments. infected flies showed a significant difference to the non-infected control (P-value < 0.0001 in 4a and 4b).
Figure 4.
The infection levels in the flies (A. Fecal transmission B. Oral Transmission) after several days of infection. The figure shows a steady infection rate in both experiments. infected flies showed a significant difference to the non-infected control (P-value < 0.0001 in 4a and 4b).
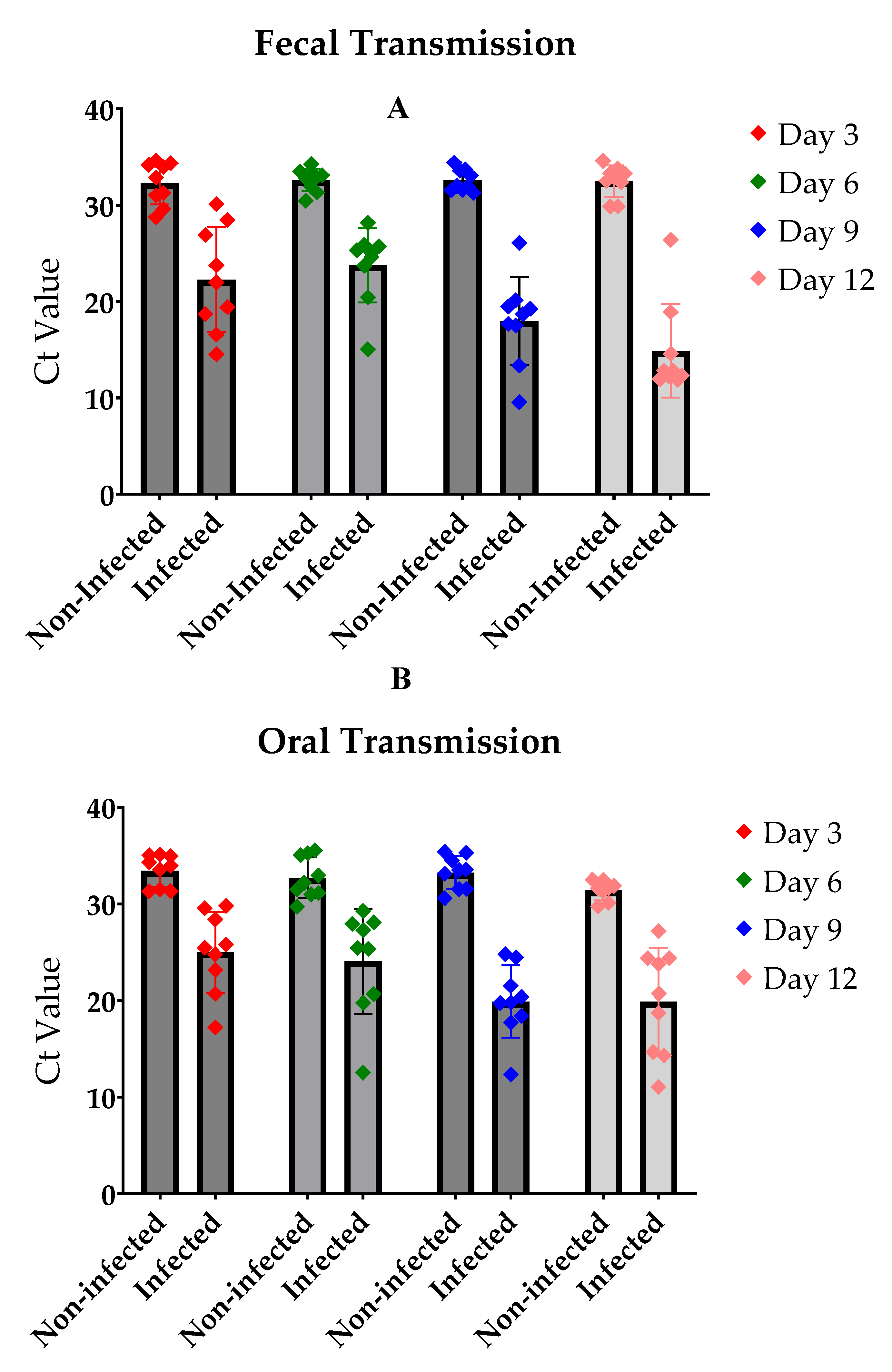
Figure 5.
Egg-to-adult success rate of non-infected versus infected flies for larva (yellow bars), pupa (orange bars), and adult (grey bars). All differences are significant when α=0.05. SE of difference = 3. 274.
Figure 5.
Egg-to-adult success rate of non-infected versus infected flies for larva (yellow bars), pupa (orange bars), and adult (grey bars). All differences are significant when α=0.05. SE of difference = 3. 274.
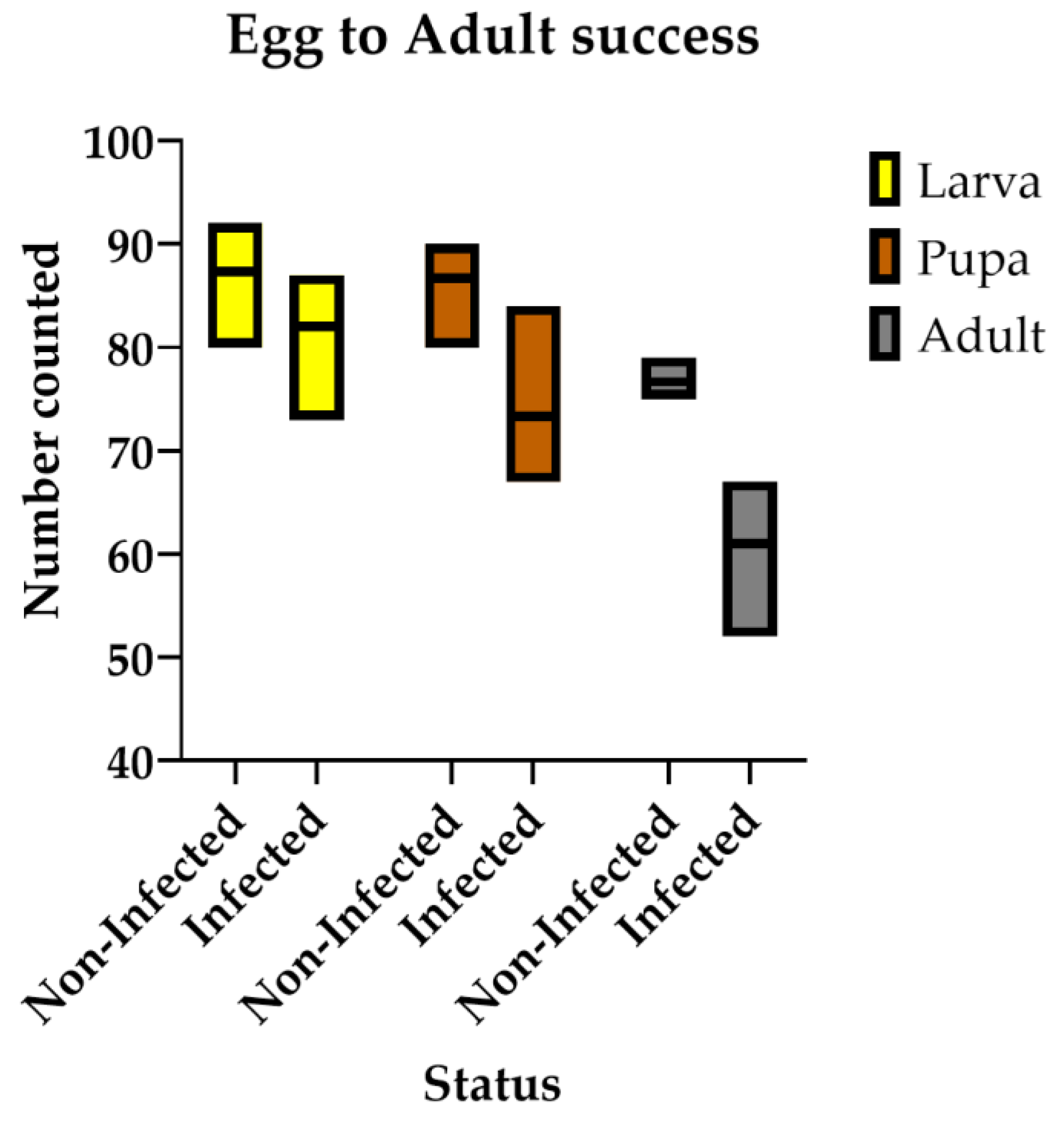

Table 1.
shows rhe sequence of the primers and probes used in this study.
| Description | Sequence | Product Length (bp) | |
|---|---|---|---|
| LJV specific probe * | 5‘-ACTCGGCGTTATCGTTACAACCGCACATATC-3‘ | ||
| LJV forward primer | 5’-CAACACGTTGTGCTGCCTGA-3’ | 128 | |
| LJV reverse primer | 5’-TCCATCCAAACTCCACCTCC-3’ | 128 | |
* Labeled on 50 with fluorescent reporter dye FAM, on 30 with fluorescent quencher TAMRA. The probe is at.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



