Submitted:
14 August 2025
Posted:
15 August 2025
You are already at the latest version
Abstract
Non-alcoholic fatty liver disease (NAFLD) is characterized by the accumulation of hepatic lipids, inflammation, and fibrosis. Currently, more than 110 phytochemical constituents of Passiflora edulis have been reported, which have been attributed with biological activities such as: antihypertensive, antidiabetic, antitumor, hepatoprotective, an-ti-inflammatory, and hypolipidemic. The objective of this study was to evaluate the in vitro and in silico effect of the ethanolic extract of P. edulis leaves on the transcriptional factors PPARα and SREBP-1. To this end, HepG2 cells were induced to an NAFLD model with oleic acid, the hepatic content of triglycerides and cholesterol was quantified both intracellularly and extracellularly, and protein-ligand molecular docking was performed to analyze the binding af-finities of some components of the extract with the human proteins PPARα and SREBP-1. The extract was found to reduce hepatic lipid content. Meanwhile, the molecular docking results showed that the bioactive components of the extract had a higher affinity for PPARα (agonist) than for SREBP-1c (antagonist). In conclusion, the results indicate the important role of P. edulis in the prevention and treatment of NAFLD.
Keywords:
NAFLD
; PPARα
; SREBP-1c
; lipids
; molecular coupling
1. Introduction
Non-alcoholic fatty liver disease (NAFLD) is recognized as a hepatic manifestation of metabolic syndrome, and its prevalence has been estimated at between 25% and 30% in adults worldwide [1]. It is related to a spectrum of liver diseases ranging from hepatic steatosis to steatohepatitis, fibrosis, and cirrhosis [2]. A diet rich in sugars and saturated fats, as well as a lack of physical activity, are considered to negatively affect health and the development of NAFLD. However, the use of medicinal plants has come to play a vital role in the prevention and treatment of this disease. In this regard, whole plants or parts of them can be used for medicinal purposes; according to the World Health Organization (WHO), more than 80% of the world's population regularly uses traditional medicines to meet their primary health care needs [3]. Furthermore, more than 50% of new medicines developed and approved for marketing are derived from modified products or active components of medicinal plants [4].
In this regard, bioactive compounds such as flavonoids, saponins, tannins, and alkaloids have been the subject of recent research due to their recognized antioxidant and hepatoprotective activity [5]. Furthermore, epidemiological studies have highlighted that these bioactive molecules have the ability to modulate metabolic pathways involved in inflammatory processes and lipid metabolism [6]. In particular, carotenoids, in addition to their antioxidant capacity, interfere with intestinal cholesterol absorption, reducing its plasma levels, which supports their usefulness in the prevention of dyslipidemia [7].
An important factor in the early onset of NAFLD is the modulatory role played by the peroxisome proliferator-activated receptor alpha (PPAR-α). This belongs to the family of nuclear receptor transcription factors, which include three isoforms: PPARα, PPARβ/δ, and PPARγ, where PPARα is particularly abundant in the liver, but is expressed and active in many other tissues, including skeletal muscle, adipose tissue, intestine, kidneys, and heart, which contribute to fatty acid homeostasis [8,9] as the main regulator of lipid metabolism by regulating numerous genes involved in the uptake and activation of fatty acids, peroxisomal and mitochondrial fatty acid oxidation, ketogenesis, triglyceride turnover, lipid droplet biology, gluconeogenesis, and bile synthesis/secretion [10]. In addition, it has other implications, such as glucose metabolism and homeostasis, glycerol control for gluconeogenesis, proinflammatory and anti-inflammatory activity, and oxidative stress [11]. On the other hand, sterol regulatory element-binding protein (SREBP) regulates the expression of enzymes that catalyze the synthesis of fatty acids, cholesterol, triglycerides, and phospholipids [12]). SREBP1c, one of three members of the SREBP family of helix-loop-helix transcription factors, is essential for the genomic actions of insulin in carbohydrate and lipid metabolism and plays a central role in the progression of NAFLD due to the biosynthesis of triglycerides, fatty acids, and cholesterol through its target effectors, which include stearoyl-CoA desaturase 1 (SCD1), fatty acid synthase (FAS), and acetyl-CoA carboxylase (ACC) [13].
Ethnopharmacological information indicates that the Passiflora genus has been used to treat various conditions such as Alzheimer's disease, cancer, Parkinson's disease, and liver disease [14]. It has anti-inflammatory, antidiabetic, and sedative properties that have been used for anxiolytic purposes [15,16]. P. foetida leaf extract was found to have anticancer activity in colorectal cancer cell models, as well as showing intracellular lipid-lowering effects, suggesting its usefulness in metabolic disorders such as NAFLD [17,18]. Passiflora incarnata leaf extract has been used to treat dyslipidemia, hypertrophy, and hepatic oxidative stress. It has also been shown to reduce the effects of a high-fat diet by lowering TC and TG levels while increasing high-density lipoprotein levels [19].
In particular, Passiflora edulis, also known as passion fruit, has been evaluated in extracts, fruit juices, and isolated compounds, showing a variety of health effects and biological activities, such as antioxidant, antihypertensive, antitumor, antidiabetic, and hypolipidemic activities [20,21,22]. In addition, passion fruit seed extract prevented NAFLD by improving liver hypertrophy and hepatic histology in mice fed a high-fat diet [23]. For these reasons, the objective of this research was to evaluate the effect of P. edulis ethanolic extract on cell viability and on transcription factors that play an important role in lipid metabolism in a hepatic cell model.
2. Results
2.1. Phytochemical Characterization
In the phytochemical characterization of the ethanolic extract of P. edulis leaves (EE), phenols were present in higher proportions with 235.7 ± 9.7 mg EAG/g DE, and total alkaloids were present in lower proportions with 8.3 ± 1.1 mg EC/g ES (Table 1).
2.2. Effect of Ethanolic Extract from P. edulis Leaves on Cell Viability
The effect on cell viability was evaluated in HepG2 and HFF cells. In HepG2 cells, EE decreased viability; showing a relationship directly proportional to concentration and exposure time; this decrease was statistically significant (p<0.001) for treatments above 1500 µg/mL (75.9%). Likewise, at 48 hours, the decrease in viability was significant for treatments above 200 µg/mL (79%), with the 3000 µg/mL treatment (13.86%) showing the greatest reduction in viability in HepG2 cells (Figure 1A.). The IC50 determined for the viability of HepG2 cells exposed to EE at 24 hours was 6663 µg/mL and 1311 µg/mL for 48 hours.
EE treatments on HFF cells showed that, after 24 hours of treatment, viability decreased in a concentration-dependent manner, which was statistically significant for treatments above 500 µg/mL. However, above this concentration, the decrease in viability was not affected by further increases in concentration. On the other hand, at 48 hours, the decrease in viability percentage was significant for treatments above 200 µg/mL and stabilized at 1000 µg/mL. The estimated IC50 at both 24 and 48 hours was 6716 and 1914 µg/mL, respectively (Figure 1B). When calculating the selectivity index, values of 1.01 and 1.46 are obtained for the 24- and 48-hour treatments, respectively. It can therefore be observed that the extract at 24 hours of treatment does not have a differential effect between healthy and cancerous cells; however, at 48 hours, HepG2 cells showed greater sensitivity to the extract than healthy cells.
2.3. Effect of Ethanolic Extract from P. edulis Leaves on Intracellular and Extracellular Triglycerides
The effect of EE on total lipid accumulation in AO-induced HepG2 cells was monitored intracellularly and extracellularly at two incubation times corresponding to 24 and 48 hours.
As for intracellular TG, during the 24-hour incubation period, a statistically significant increase in TG concentration was observed in cells treated with AO (129.83 µg triglycerides/µg protein) compared to the concentration in control cells (87.10 µg triglycerides/µg protein); while in the treatments (cells exposed to AO and EE), a statistically significant reduction in TG concentration was observed, showing that all treatments maintain values similar to the control cells. Likewise, it was observed that cells treated only with the extract (200 µg/mL) and without AO showed lower TG values than the control cells, indicating that P. edulis has an effect on lipid metabolism (Figure 2A).
During the 48-hour incubation period, there was a statistically significant decrease in TG concentration in cells treated with the extract compared to cells containing only AO, demonstrating once again that P. edulis extract has the ability to regulate intracellular lipid accumulation in liver cells. A relevant result is that treatment at 200 µg/mL reduces TG concentration to values similar to those of the control cells (Figure 2A). The IC50 of the treatments was estimated at 24 and 48 hours of exposure, finding values of 1422.25 µg/mL and 174.28 µg/mL, indicating that the effect increased significantly over time.
On the other hand, when evaluating the concentrations of extracellular TG or TG released into the culture medium of HepG2 cells treated with EE from P. edulis leaves at both 24 and 48 hours, a significant decrease in TG released from EE-treated cells was observed compared to cells containing AO, showing that there were no significant differences between the values of the treatments and the control cells, even the cells treated only with EE at 200 µg/mL had lower values compared to the control cells (Figure 2B).
In the 48-hour incubation assay, behavior very similar to that observed in the 24-hour assay was observed, i.e., a statistically significant increase in TG release in AO-treated cells compared to the TG concentration in control cells. In addition, a significant decrease in TG concentration was observed in cells treated with EE compared to cells that only had AO (Figure 2B). Likewise, it was evident that the treatments with EE did not present significant differences compared to the control, and even lower values were recorded compared to the control cells, indicating an improvement in TG release into the extracellular medium. The IC50 of the treatments was estimated at 24 and 48 hours of exposure, finding values of 560.6 µg/mL and 40,6 µg/mL, indicating that the extract reduces the amount of TG released by HepG2 cells, and this effect is enhanced with increased exposure time.
2.4. Effect of Ethanolic Extract from P. edulis Leaves on Intracellular and Extracellular Cholesterol
With regard to intracellular cholesterol, during the 24-hour exposure period, statistically significant differences were observed in the EE treatments compared to cells containing only AO (Figure 3A). In addition, it was observed that the AO100 and AO200 treatments presented lower cholesterol values than those observed in the control cells, with these differences being statistically significant. When estimating the IC50 at 24 hours, it was 134.57 µg/mL, and at 48 hours, it was 199.91 µg/mL, indicating that the extract has the ability to reduce cholesterol accumulation within HepG2 cells.
After 48 hours of incubation, there was a statistically significant decrease in the cells treated with the extract and the cells that only had AO, showing that all treatments had lower cholesterol values than those observed in the Ct cells (Figure 3A), indicating that the EE had an effect on intracellular cholesterol metabolism. It is also important to note that cells treated only with EE without AO, both in the 24- and 48-hour incubation periods, showed the greatest decrease in cholesterol concentration, indicating that the effect of the extract lasted over time. Finally, the IC50 was estimated at 24 and 48 hours with values of 134.6 µg/mL and 199.9 µg/mL, respectively.
With regard to extracellular cholesterol, during the 24-hour incubation period, only the AO200 mixture showed significant differences compared to cells containing only AO (Figure 3B). Similarly, the AO200 mixture generated lower values compared to the control cells. In the assay performed at 48 hours, cholesterol levels in cells treated with the mixture of AO and EE from P. edulis leaves were close to the levels of cholesterol released into the medium by the control cells (Figure 3B). In addition, the IC50 was estimated at 434.36 µg/mL at 24 hours and 238.88 µg/mL at 48 hours.
2.5. Molecular Coupling of the Compounds Presents in Passiflora edulis Against SREBP-1 and PPARα
Figure 4 shows the affinity values of each compound for the two proteins of interest (SREBP-1 and PPARα). From the perspective of the average affinity or binding energy for each molecule, it can be seen that PPARα showed affinity with 64.28% of the compounds evaluated, with the compounds isoorientin, amygdalin, lonicerin, kaempferol, and luteolin showing the best affinities. On the other hand, the affinity of the compounds reported in P. edulis for SREBP-1 only includes 27.2% of the compounds evaluated, with cyclopassifolic acids and cyclopassiflosides, saponarin, and spinosin being the compounds with the highest affinity for this transcription factor.
In order to analyses which compounds would show the best effects in a cell assay, the Rank-by-Rank consensus method was implemented to combine the data from both proteins and both programmes. Table 2 shows the consensus results, with the compounds with the best rankings highlighted in green. In this vein, it would be expected that in an experiment using the compounds separately, the compounds indicated here would exhibit better biological effects. In general, it can be observed that the compounds with the best rankings are flavonoids and glycosides such as: 7-de-O-methylaciculatin, isoorientin, kaempferol, luteolin, saponarin, espinosin, vitexin and lonicerin, with the latter compound showing the best results for both proteins and both programmes.
3. Discussion
The differences found in the phytochemicals present in EE from P. edulis leaves may vary depending on the methodologies and standards used, as well as due to the fact that the production and quantity of secondary metabolites vary from one place to another and from one environment to another [24]. This could be due to genetic, environmental, and ecological factors, including latitude, longitude, precipitation, climate change, soil microbiome and nutrients, temperature, phytopathogens and cultural practices, post-harvest storage, processing, among others.
The EE of P. edulis leaves had a high total phenolic content of 235.7 mg EAG/g DE, compared to that reported by Aguillón et al, [25] and Guimarães & Modolo [26], who reported lower results for this same extract, with total phenolic content of 150.3 mg EAG and 122.0 mg EAG, respectively. However, when considering extracts obtained with other solvents, such as the aqueous extract of P. edulis leaves with phenolic contents of 10.45 mg EAG [27], this difference may be due to the different polarities that affect the efficiency of phenol extraction. Regarding the different types of phenols, it has been reported that chlorogenic acid and its derivatives have the ability to decrease TG accumulation in AO-induced HepG2 cells [28]; another study confirmed the protective effect of chlorogenic acid (80 µM and 50 µM) against the formation of lipid droplets induced by free fatty acids (FFA) and the elevation of ROS in HepG2 cells [29]. Chlorogenic acid also improved blood lipid metabolism by alleviating FFA, TG, and modulating the hepatic level of pACC through the AMPK pathway [30]. Additionally, this compound has been reported to increase AMPK and ACC phosphorylation and reduce the expression of SREBP-1C, ACC, and FAS in HepG2 cells [31].
Furthermore, anthocyanins have been found to activate AMPK in human HepG2 cells and can decrease the level of SREBP-1c [32]; in addition, other phenolic compounds that have been found, such as caffeic acid, catechins, and procyanidins, have the ability to improve the AMPK/ACC/SREBP-1c pathway, reducing hepatic steatosis due to FFA overload [33].
Regarding the total flavonoid content, Santos et al. [34] found 29.4 mg EC in the EE of P. edulis leaves, which is lower than the results reported in this study. Sotiropoulou et al. [35] mention that quercetin stands out for its hepatoprotective role, where in a study with rats treated with ethanol, quercetin proved to be beneficial in chronic ethanol-induced liver injury due to the increase in glutathione content. The inhibition of glucose uptake and cell proliferation was also evident, finding that quercetin is capable of improving insulin resistance, decreasing fat accumulation and increasing cell proliferation, while inhibiting IL-8 and TNF-α levels, with an increase in cellular glutathione. In addition, luteolin has been found to exhibit anti-inflammatory, antioxidant, antitumor and immune regulatory activity [36], where this flavonoid has the ability to significantly reduce body weight and liver index, restoring serum lipid levels, inhibiting hepatic lipid accumulation and improving insulin resistance in Wistar rats.
Regarding the total tannin content, Saravanan & Parimelazhagan [37] report a quantity of 67.90 ± 3.02 mg EAG (gallic acid) in the methanolic extract of Passiflora ligularis, which is higher than that reported in our research. De Huang et al [38] mention that epigallocatechin gallate significantly reduced the development of obesity, hyperglycemia, insulin resistance, inflammatory cytokines, hypercholesterolemia, and fatty liver in rats fed a high-fat diet. On the other hand, Li et al. [39] mention that tannic acid decreased the mRNA expression of genes related to lipogenesis and attenuated lipid accumulation in vivo, observing that the characteristics of NAFLD, including body weight, liver mass, fat mass, and serum lipid profile, improved with tannic acid supplementation in vivo.
Finally, with regard to the total alkaloid content, De Sousa et al. [40] mention that the leaves of P. edulis contain alkaloids such as harmine, harmalol, harmaline, and harman, a β-carboline alkaloid. Alkaloids have also been detected in extracts and fractions from the bark of P. edulis, where different indole alkaloids such as harmine, harmaline, and harmalol have been identified [41]. Various reports have mentioned a wide range of pharmacological activities, including anti-inflammatory [42], antioxidant, and anticancer activities [43], making it a possible candidate for NAFLD [44].
Cell viability assays showed that the EE from P. edulis leaves had an antiproliferative effect on HepG2 at 24 and 48 hours (IC50/24 hours = 6663 µg/mL – IC50/48 hours = 1311 µg/mL). In addition, the maximum inhibition percentage of EE was 27% at 24 hours and 86.1% at 48 hours. Similarly, EE was found to have a greater cytotoxic effect on HepG2 cells compared to the HFF cell line, indicating greater selectivity for the tumor cell line, which is favorable, as differential antiproliferative activity is sought. Similarly, in the study conducted by Villada et al., [45] they found an effect of cell viability on SW480 cells, dependent on the dose and exposure time. These effects could be related to the presence of certain metabolites that have been described in extracts of P. edulis, such as terpenes and glycosides, which have been found in both aqueous extracts of the fruit and ethanolic extracts of the leaves; such as ‘passiflorine’ and ‘passicapsin,’ alkaloids, tannins, quinones, flavonoids, carotenoids, anthocyanins, ascorbic acid, vitamins B6, vitamin A, riboflavin, and lactones [15], which have been found in different plant structures and could be involved in the biological effects described in this study.
Regarding other species from the Passiflora genus, it has been reported that Passiflora liguralis EE showed significant antiproliferative activity on human hepatocellular carcinoma cells (IC50: 50 mg/mL); however, no cytotoxic effect was observed in primary hepatocyte cultures [46]. On the other hand, Passiflora alata leaf extract has been reported to have cytotoxic or anticancer activities attributed to its constituent flavonoids and saponins, where the data obtained indicated that P. alata inhibits cell proliferation >75% against the four tumor cell lines (PC-3, K-562, HepG2, and S180) using the MTT method, and IC50 < 30 µg/mL [47].
On the other hand, the increase in lipid accumulation in HepG2 cells promoted by the presence of oleic acid triggers the generation of high levels of oxygen free radicals compared to control values [48], due to the high rate of lipid peroxidation that is generally associated with damage at the cellular level, which can lead to alterations in cell division controls, causing the cell to develop other pathological conditions, such as steatohepatitis, cirrhosis, or more alarming consequences such as cancer or death. In this study, HepG2 cells were treated with pathophysiological levels of oleic acid to mimic the entry of excess free fatty acids into hepatocytes, leading to steatosis, because hepatocytes activate enzymes associated with lipogenesis, resulting in increased TG synthesis and greater lipid accumulation in the liver [49], a situation that was evident in this research. However, in this study, TG accumulation was substantially reduced both intracellularly (50.34%) and extracellularly (56.65%) at 24 hours of incubation and at 48 hours of incubation, it was reduced by 62.74% and 48.68%, respectively, compared to cells that were induced to lipid accumulation with AO, demonstrating that P. edulis EE can attenuate NAFLD in silico and in vitro. Similarly, studies have reported that blackberry (Rubus sp.), wild blueberry (Vaccinium spp.), strawberry (Fragaria ananassa) and chokeberry (Aronia melanocarpa) have an inhibitory effect on oleic acid-induced TG accumulation in HepG2 cells [58]. Furthermore, in a study conducted by Kocaadam et al [51] using an animal model fed a high-fat diet until they developed fatty liver, the administration of Curcuma longa extracts reduced lipid accumulation. Similarly, Lonicera caerulea extract has been reported to show a significant decrease in intracellular TG in HepG2 cells that were induced to accumulate lipids with AGL for 24 hours [52]. With regard to TC levels, our study also showed a reduction both intracellularly (46.05%) and extracellularly (49.18%) after 24 hours of incubation, and after 48 hours of incubation, the reduction was 48.68% and 47.78%, respectively. Similarly, it has been reported that the ethanolic extract of Liriope platyphylla root significantly decreased serum CT levels in mice fed a high-fat diet [53].
With regard to the in-silico results of molecular docking, several reports have been published on the biological activity of certain compounds and their interaction with the PPARα and SREBP-1 proteins, such as luteolin. In the study conducted by Yin et al., [54] they found that treatment with luteolin can significantly inhibit body weight gain in mice and hepatic TG levels. This result is due to the fact that luteolin regulates hepatic lipid homeostasis by inhibiting LXR activation, which causes a decrease in lipid accumulation and suppresses LXR-SREBP-1 signalling. Additionally, kaempferol can improve insulin resistance to reduce hepatic fat overaccumulation and prevent the progression of NAFLD, as well as promote hepatic expression of CYP4A1 (cytochrome P450) secondary to direct upregulation of hepatic PPARα expression [55].
In the case of saponarin, it has been reported to inhibit the expression of SREBP-1 and PPARγ [56], which are associated with lipid metabolism, the expression of lipogenic enzymes such as FAS, and TG production. Consequently, saponarin suppresses TG accumulation by stimulating AMPK, which is an important energy regulator. Several natural products, including isoorientin, lonicerin, kaempferol, and luteolin, have shown promising results in preclinical studies through the activation of AMPK and the regulation of its proteins related to lipid metabolism [57,58,59]. AMPK directly inhibits the activation of SREBP-1c, a transcription factor that controls lipid metabolism, with SREBP-1c activating the expression of FAS and SCD-1, two proteins crucial in hepatic fatty acid synthesis, to promote de novo lipogenesis. Tie et al. [60] found in their work that treatment with kaempferol and kaempferide in HepG2 cells reduced the AO-induced increase in SREBP-1c, FAS, and SCD-1, suggesting that these flavonoids inhibit intracellular lipid accumulation by suppressing the expression of lipogenic proteins.
4. Materials and Methods
The plant material (P. edulis leaves) was collected in the village of Tierra Blanca (Roldanillo, Valle del Cauca, Colombia), located at the global geographic coordinates 4° 24´0 N, 75.8489° at an altitude of 939 meters above sea level. The samples were taken to the Biochemistry Laboratory of the Faculty of Health Sciences at the University of Quindío.
Preparation of the extract. The leaves of P. edulis were washed and then dried in a circulating air oven at a constant temperature of 40°C. The material was then pulverized and finally leached for 8 days using 96% ethanol, with the leachate being constantly recirculated. The chlorophylls were then separated by liquid-liquid extraction with ethanol-water (1:1) [61]. The ethanol was evaporated at a reduced pressure of 60 mbar and at a temperature below 30°C in a rotary evaporator (Heidolph). The resulting suspension was subjected to freeze-drying (Telstar LyoQuest) and the product obtained was stored protected from light at -20°C.
Determination of total phenol content. This was determined using the Folin Ciocalteu reagent method [62]. An aliquot of 50 µL of the ethanol extract (EE) (1 mg/mL) was taken and 2.5 mL of Folin Ciocalteu reagent (1:10 dilution) and 2 mL of 7.5% (w/v) sodium carbonate (NaCO3) were added, mixed until homogeneous, and incubated at 40°C for 15 minutes. A solution of 2 mL of Na2CO3 in 2 mL of distilled water was used as a blank, and the reading was taken at a wavelength of 765 nm. The results were expressed as mg equivalent of gallic acid per gram of dry extract (mg EAG/g DE).
Determination of total flavonoid content. To an aliquot of 150 µL of EE (1 mg/mL), 45 µL of 5% sodium nitrate (NaNO3) was added. At five and six minutes, 90 µL of 10% aluminum chloride (AlCl3) and 300 µL of 1 M sodium hydroxide (NaOH) were added, respectively. The final volume of the mixture was adjusted with distilled water to 1.5 mL. Finally, the mixture was allowed to stand for 10 minutes at room temperature and read at a wavelength of 510 nm. The results were expressed as mg catechin equivalents per gram of dry extract (mg CE/g DE) [63].
Determination of total polysaccharide content. This was determined using the phenol-sulfuric acid method [64]. To an aliquot of 100 µL of EE (1 mg/mL), 100 µL of 5% phenol and 500 µL of 95% sulfuric acid (H2SO4) were added, mixed until homogeneous, and incubated at room temperature for 15 minutes. The reading was taken at a wavelength of 490 nm, and distilled water plus H2SO4 was used as the blank. The results were expressed as mg glucose equivalents per gram of dry extract (mg GE/g DE).
Determination of total tannin content. To an aliquot of 250 µL of EE (1 mg/mL), 500 µL of bovine serum albumin solution in 0.2 M acetic buffer with pH 5.0 (pH adjusted with 0.17 M NaCl) was added, mixed thoroughly, and allowed to stand for 15 minutes. It was then centrifuged at 5000 rpm for 15 minutes. The supernatant was discarded and the precipitate was diluted with 1 mL of aqueous solution containing 1% sodium dodecyl sulfate (SDS) and 4% triethanolamine. Finally, 250 µL of 0.01 M ferric chloride (FeCl3) in 0.01 M hydrochloric acid (HCL) was added and allowed to stand for 30 minutes. The reading was taken at a wavelength of 510 nm. As a blank, 1 mL of aqueous solution (1% SDS and 4% triethanolamine) was used in 250 µL of the 0.01 M FeCl3 solution in 0.01 M HCL. The results were expressed as mg equivalent of tannic acid per gram of dry extract (mg EAT/g DE) [63].
Determination of total alkaloid content. This was determined using the bromocresol green method [65]. A 1 mL aliquot of EE (1 mg/mL) was taken and 5 mL of phosphate buffer (pH 4.7) and 5 mL of bromocresol green solution were added. Next, 2 mL of chloroform was added and mixed. This last step was repeated two more times. Finally, the extract was collected in a 10 mL volumetric flask and read at a wavelength of 470 nm. The results were expressed as mg quinine equivalents per gram of dry extract (mg EQ/g DE).
Cell culture. HepG2 and HFF cell lines were maintained and propagated in Dulbecco's Modified Eagle's Medium (DMEM) (Gibco) with 25 mM glucose and 2 mM L-glutamine, supplemented with 10% fetal bovine serum (FBS) (Gibco), 100 IU/mL penicillin, 100 µg/mL streptomycin (Gibco), and 1% nonessential amino acids (Gibco). They were incubated at 37°C in a humid atmosphere of 95% air and 5% CO2. The different biological assays were performed while the cells were in the exponential phase and had a confluence of more than 80%.
Cell viability. Cell viability assays were performed using the sulforhodamine B (SFB) method [66]. Ten thousand cells/well from each cell line were cultured in 96-well culture plates in a final volume of 200 μl of culture medium and incubated for 24 and 48 hours in the presence of EE from P. edulis leaves at different concentrations (50–3000 µg/mL). After the exposure period to the extract, 100 μL of cold trichloroacetic acid at a concentration of 15% (v/v) was added to each well. They were incubated at 4°C for 1 hour in the dark, the acid was removed, and four washes with water were performed. They were then left to dry at room temperature, and finally, 100 μL of SFB (0.4% w/v diluted in 1% acetic acid) was added per well. They were incubated for 30 minutes at room temperature, and then excess SRB was removed by washing the adhesion surface with acetic acid (1% v/v in distilled water). The plates were left to dry at room temperature for 24 hours. For absorbance reading, SRB was solubilized by adding 200 μl of Tris-HCl buffer (10 mM pH 10.5) to each well and shaking orbitally for 15 minutes. The optical density was read at 490 nm by spectrophotometry.
Three biological assays were performed, with each assay repeated three times for each treatment and control. Cell viability was calculated using the following equation:
% viability = (ABt/ABc) * 100
where, ABt = absorbance of treatments; ABC = absorbance of untreated cells.
The Selectivity Index (SI) of the treatments evaluated was defined as the ratio of cytotoxicity between non-cancerous cells and cancerous cells. Treatments reporting an SI value greater than 3 are considered to have high selectivity. The following equation was used:
SI = (IC50 in HFF cells)/(IC50 in HepG2 hepatocellular
cancer cells)
Effect of the extract on total lipids (cholesterol and triglycerides). 10,000 HepG2 cells/well were cultured in sterile 6-well plates in a final volume of 2,000 μl of culture medium until 90% confluence was reached. The EE of P. edulis (50, 100, and 200 µg/mL) and oleic acid (100 µM) were evaluated. The cells were incubated for 24 and 48 hours. The levels of cholesterol and triglycerides were quantified both intracellularly (cell lysate) and extracellularly (culture medium) for each treatment. As a positive control, cells were cultured with oleic acid only, and as a negative control, cells were cultured with culture medium only [67].
Once the exposure period of the extract had elapsed, the culture medium was removed and stored (for extraction and quantification of extracellular lipids). Subsequently, the monolayer was washed with 1X PBS, and the cells were lysed with 1000 µl of lysis buffer (20 mM Hepes, 420 mM NaCl, 1% Triton, 0.5% SDS, pH 7). Immediately, the boxes were incubated at room temperature with constant shaking for 10 minutes.
Normalization of lipid concentration. The assay was performed according to Walker [68]. After cell lysis, 50 µl of the lysate was taken from each of the treatments, controls, and standards, 500 µl of the BCA working reagent (50:1, bicinchoninic acid solution/copper sulfate pentahydrate solution 4%) was added, and immediately test tubes were covered and incubated at 37°C for 30 minutes. Finally, they were kept at room temperature for 10 minutes before measurement. The reading was taken at 562 nm. The calibration curve was performed with albumin.
Lipid extraction and quantification. Lipid extraction was performed according to the method established by Blight and Dyer [69]. The volume of cell lysate was normalized against the lowest protein concentration observed in either the treatments or controls. A normalized and measured volume of 400 µL of cell lysate was taken for the quantification of the extracted lipids using the commercial Liquicolor® (Human) kit for triglycerides and cholesterol, independently for each one. The same was done for the culture medium. Subsequently, 500 µL of chloroform and 1 mL of methanol were added, mixed well, and placed in orbital agitation at 180 rpm for 1 hour. Next, 500 µL of chloroform and 500 µL of water were added, forming two phases. It was then centrifuged at 3000 rpm for 3 minutes to achieve better separation, then the chloroform phase was removed to a new tube, and the chloroform was allowed to evaporate completely overnight. Finally, 500 µL of quantification reagent provided by the Human liquicolor® colorimetric kit was added to analyze either cholesterol or triglycerides, incubated for 10 minutes, and spectrophotometric reading was performed at a wavelength of 500 nm, also reading the absorbance of the standard provided by the kit.
The results were expressed as µg of lipid/µg of protein for each time point. To convert the absorbances to a concentration unit, the following equation recommended by the commercial Liquicolor® (Human) kit was used:
Lipid concentration = 200 x (ABt/ABe) (mg/dl)
ABt = Absorbance of treatments and controls; ABe= Absorbance of the standard, either cholesterol or triglyceride
Molecular docking calculations. Protein-ligand molecular docking calculations were performed for some components of the extract on the human proteins PPARα and SREBP-1. For this purpose, 10 different structures were used for the PPARα protein (Ensemble docking). The protein structures were obtained from the Protein Data Bank, PDB (https://www.rcsb.org/). For the PPARα protein, the structures with PDB codes 2ZNN [70], 3ET1 [71], 3SP6 [72], 3SP9 [72], 3VI8 [73], 6KB2 [74], 5HYK [75], 1I7G [76], 2NPA [77], and 6KBA [73]. For the SREBP-1 protein, there is only one structure deposited in the PDB with code 1AM9 [78]; however, in this crystal there are two repetitions of the protein, which were used as two conformations of that protein.
The preparation of the receptors consisted of the removal of water molecules and other co-crystallized ligands, followed by the addition of polar hydrogens and the assignment of Kollman charges. This was done using the Autodock Tools v1.5.6 program [79]. In the case of ligands, a list of 42 compounds present in P. edulis leaves was obtained from the Foodb database (https://foodb.ca/). Two programs were used, Autodock Vina [80] and Smina [81]. A total of 3024 molecular docking calculations were performed. All data obtained from this part were analyzed and organized according to the consensus method known as Rank by Rank. Using the equation:
where n is the total number of programs used (in this case, 2). Using the values obtained for binding affinities as a reference, a value r_i^j is assigned, which is equivalent to the location of compound i with respect to the other compounds. Thus, for example, if a compound occupies position 5 in the list, its score will be 5 for program j. Finally, the average of the results of the programs used is obtained.
Statistical design. Data were expressed as the mean ± standard deviation of the results of three determinations in triplicate for each assay. The IC50 (mean inhibitory concentration) was determined using linear regression. Statistical differences between three or more data sets were analyzed using GraphPad Prism 8.0 (GraphPad Software Inc., San Diego, CA) and SPSS v.25 statistical software for one- and two-factor analysis of variance (ANOVA) and a Tukey test for multiple comparisons. In addition, a Duncan test was performed to compare each treatment with the negative control. Values of p<0.05 were considered significant.
5. Conclusions
Our study showed that the EE from P. edulis leaves had a high content of phenols, polysaccharides, and flavonoids. In terms of cell viability, EE had a differential effect on the two cell lines, showing greater selectivity for the HepG2 cancer cell line. Likewise, cells treated with EE showed a statistically significant reduction in total intracellular and extracellular TG and CT content at the different concentrations evaluated. It was also determined in silico that five bioactive compounds, namely isoorientin, lonicerin, kaempferol, luteolin, and amygdalin, may be possible activators of the PPARα transcription factor and, consequently, will have effects on fatty acid oxidation. Similarly, compounds such as lonicerin, saponarin, espinosin, and cyclopasifloside VII can inhibit the transcription factor SREBP-1c, achieving inhibition of de novo lipogenesis and a decrease in lipid accumulation. Thus, the results of this research contribute to the knowledge of P. edulis as a potential species that attenuates or prevents NAFLD.
Author Contributions
Conceptualization, Johanny Aguillon and Jhon Sebastian León; methodology, Johanny Aguillon and Nelsy Loango; software, Jhon Sebastian León; validation, Johanny Aguillon and Jhon Sebastian León.; formal analysis, Johanny Aguillon and Nelsy Loango; investigation, Johanny Aguillon and Nelsy Loango; resources, Nelsy Loango; data curation, Jhon Sebastian León ; writing—original draft preparation, Johanny Aguillon; writing—review and editing, Johanny Aguillon and Nelsy Loango; visualization, Johanny Aguillon; Nelsy Loango; supervision, Nelsy Loango; project administration, Nelsy Loango; funding acquisition, Johanny Aguillon and Nelsy Loango. All authors have read and agreed to the published version of the manuscript.
Funding
This research has been funded by the University of Quindío.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki and did not require approval from a bioethics committee because the trials did not involve humans or experimental animals.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author due to the information is part of a degree project, it belongs to the University of Quindío, according to its intellectual property protocol.
Acknowledgments
The authors would like to thank the Master's Degree Program in Biomedical Sciences at the University of Quindío and Carlos Andres Rodriguez Salazar for their support and advice. We would also like to thank the members of the research group.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| NAFLD | Non-alcoholic fatty liver disease |
| PPARα | The peroxisome proliferator-activated receptor alpha |
| SREBP-1 | The sterol regulatory element-binding protein type 1 |
| HepG2 | Cell line exhibiting epithelial-like morphology that was isolated from a hepatocellular carcinoma |
| WHO | The World Health Organization |
| SCD1 | Stearoyl-CoA desaturase 1 |
| FAS | Fatty acid synthase |
| ACC | Acetyl-CoA carboxylase |
| EE | The ethanolic extract of P. edulis leaves |
| mg EAG/g DE | mg equivalent of gallic acid/g of dry extract |
| mg EC/g ES | mg equivalent of caffeine/g of dry extract |
| mgEQ | mg equivalent |
| HFF | Fibroblast cell that was isolated from the foreskin of a donor |
| AO | Oleic acid |
| TG | Triglycerides |
| CT | Cholesterol |
| IC50 | Medium Inhibitory Concentration |
| FFA | Free fatty acids |
| ROS | Reactive oxygen species |
| AMPK | AMP-activated protein kinase pathway |
| IL-8 | Interleukin 8 |
| TNF-α | Tumor necrosis factor alpha |
| CYP4A1 | Enzyme cytochrome P450 4A1 |
References
- Younossi ZM, Rinella ME, Sanyal AJ, Harrison SA, Brunt EM, Goodman Z, et al. Nonalcoholic fatty liver disease: assessment of variability in pathologic interpretations. Mod Pathol. 2020, 33, 776–85. [Google Scholar] [CrossRef]
- Cotter T, Rinella M. Nonalcoholic fatty liver disease 2020: the state of the disease. Gastroenterology, 2020; 1;158(7):1851-64.
- Wang W, Xu J, Fang H, Li Z, Li M. Advances and challenges in medicinal plant breeding. Plant Sci, 1: 301, 1105.
- Zhou Y, Li Y, Wang Y, Wang D, Li X, Zhang X, et al. CRISPR-Cas gene editing technology and its application prospect in medicinal plants. Chin Med. 2022, 17, 1–14. [Google Scholar] [CrossRef]
- Lugumira R, Tafire H, Vancoillie F, Ssepuuya G, Van Loey A. Nutrient and phytochemical composition of nine African leafy vegetables: a comparative study. Foods. 2025, 14, 1304. [Google Scholar] [CrossRef]
- Matsumoto, M. Prevention of metabolic syndrome by phytochemicals and vitamin D. Biomed Res Int, 2022, 2022 9917154. [Google Scholar] [CrossRef]
- El Omari N, Bakrim S, Khalid A, Abdalla AN, Iesa MAM, El Kadri K, et al. Unveiling the molecular mechanisms: dietary phytosterols as guardians against cardiovascular diseases. Nat Prod Bioprospect. 2024, 14, 27. [Google Scholar] [CrossRef]
- Tan Y, Wang M, Yang K. PPAR modulators as current and potential cancer treatments. Front Oncol, 2021: 11, 5999.
- Todisco S, Santarsiero A, Convertini P. PPAR Alpha as a Metabolic Modulator of the Liver: Role in the Pathogenesis of Nonalcoholic Steatohepatitis (NASH). Biology. 2022, 11, 792. [Google Scholar]
- Henderson, GC. Plasma free fatty acid concentration as a modifiable risk factor for metabolic disease. Nutrients. 2021, 13, 2590. [Google Scholar] [CrossRef]
- Kou Y, Geng F, Guo D. Lipid metabolism in glioblastoma: from de novo synthesis to storage. Biomedicines. 2022, 10, 1943. [Google Scholar]
- Ferré P, Phan F, Foufelle F. SREBP-1c and lipogenesis in the liver: an update. Biochem J. 2021, 478, 3723–39. [Google Scholar] [CrossRef]
- Chandrasekaran, P.; Weiskirchen, R. The Role of SCAP/SREBP as central regulators of lipid metabolism in hepatic steatosis. Int. J. Mol. Sci. 2024, 25, 1109. [Google Scholar] [CrossRef]
- Medeiros N, Almeida D, Lima J. In vitro antioxidant activity of Passiflora alata extract by different kinds of treatment on rat liver. Curr Bioact Compd 2018; 14:21–5.
- Smruthi R, Divya M, Archana K, Maddaly R. The active compounds of Passiflora spp. and their potential medicinal uses from both in vitro and in vivo evidence. J Adv Biomed Pharm Sci 2020; 4:45–55.
- Smilin G, Abbirami E, Sivasudha T, Ruckmani K. Passiflora caerulea L. fruit extract and its metabolites ameliorate epileptic seizure, cognitive deficit and oxidative stress in pilocarpine-induced epileptic mice. Metab Brain Dis 2020; 35:159–73.
- Dharmasiri PGNH, Ranasinghe P, Jayasooriya PT, Samarakoon KW. Antioxidant, anti-diabetic and anti-inflammatory activities of Passiflora foetida grown in Sri Lanka. Trop Agric Res. 2024, 35, 24–33. [Google Scholar] [CrossRef]
- Dharmasiri PGNH, Ranasinghe P, Jayasooriya PT, Samarakoon KW. Antioxidant, anti-diabetic and anti-inflammatory activities of Passiflora foetida grown in Sri Lanka. Trop Agric Res. 2024, 35, 24–33. [Google Scholar] [CrossRef]
- Michael HS, Mohammed NB, Ponnusamy S, Edward GN. A folk medicine: Passiflora incarnata L. phytochemical profile with antioxidant potency. Turk J Pharm Sci. 2022, 19, 287–92. [Google Scholar] [CrossRef] [PubMed]
- Hu Y, Jiao L. A new C-glycosyl flavone and a new neolignan glycoside from Passiflora edulis Sims peel. Nat Prod Res 2018; 32:2312–8.
- Silva LNS, Oliveira DM, Oliveira AS, Cunha VMN, Carvalho FAA, Costa JP, et al. Hypotensive and vasorelaxant effects of ethanolic extract from Passiflora edulis Sims leaves. J Ethnopharmacol 2017; 204:127–32. [CrossRef]
- Nerdy N, Ritarwan K. Hepatoprotective activity and nephroprotective activity of peel extract from three varieties of the passion fruit (Passiflora sp.) in the albino rat. Open Access Maced J Med Sci. 2019, 7, 536–42. [Google Scholar] [CrossRef] [PubMed]
- Jako P, Chonpathompikunlert P, Malakul W, Tunsophon S. Passiflora edulis extract ameliorates HFD-induced hepatic steatosis mediated through Nrf2 and IRS-1 activation, NFκB suppression, and hepatic lipid metabolism and bile acid modulation in obese rats. Biomed Pharmacother, 4752.
- Preetha P, Kamalabai S, Jayachandran S. Qualitative and quantitative phytochemical screening of Vitex negundo L. extract using chromatographic and spectroscopic methods. Qualitative and quantitative phytochemical screening of Vitex negundo L. extract using chromatographic and spectroscopic methods. Nat Volat Essent Oils J. 2021, 11949–61. [Google Scholar]
- Aguillón J, Arango S, Uribe F, Loango N. Cytotoxic and apoptotic activity of extracts from leaves and juice of Passiflora edulis. J Liver Res Disord Ther. 2018, 4, 67–71. [Google Scholar]
- Guimarães SF, Lima IM, Modolo LV. Phenolic content and antioxidant activity of parts of Passiflora edulis as a function of plant developmental stage. Acta Bot Bras. 2020, 34, 74–82. [Google Scholar] [CrossRef]
- Do Carmo L, Martins M, Magalhães R, Júnior M, Macedo A. Passiflora edulis leaf aqueous extract ameliorates intestinal epithelial barrier dysfunction and reverts inflammatory parameters in Caco-2 cells monolayer. Food Res Int, 9162.
- Pietrzyk N, Zaklos-Szyda M, Kołopulus L. Fruit phenolic compounds protect against FFA-induced steatosis of HepG2 cells via AMPK pathway. J Funct Foods, 2021: 80, 1044.
- Liu Y, Zhai T, Yu Q, Zhu J, Chen Y. Effect of high exposure of chlorogenic acid on lipid accumulation and oxidative stress in oleic acid-treated HepG2 cells. Chin Herb Med. 2018, 10, 199–205. [Google Scholar]
- Meng F, Song C, Liu J, Chen F, Zhu Y, Fang X, et al. Chlorogenic acid modulates autophagy by inhibiting the activity of ALKBH5 demethylase, thereby ameliorating hepatic steatosis. J Agric Food Chem. 2024, 72, 19257–68. [Google Scholar] [CrossRef]
- Wang Y, Li X, Zhang Y, Liu Y, Jiang Y. Chlorogenic acid and its isomers attenuate NAFLD by mitigating lipid accumulation in oleic acid-induced HepG2 cells and high-fat diet-fed zebrafish. J Food Biochem. 2024, 48, e13958. [Google Scholar]
- Yan F, Zheng X. Anthocyanin-rich mulberry fruit improves insulin resistance and protects hepatocytes against oxidative stress during hyperglycemia by regulating AMPK/ACC/mTOR pathway. J Funct Foods, 2017; 30:270–81.
- Zhang Y, Pan H, Ye X, Chen S. Proanthocyanidins from Chinese bayberry leaves reduce obesity and associated metabolic disorders in high-fat diet-induced obese mice through a combination of AMPK activation and alteration in gut microbiota. Food Funct. 2022, 13, 2295–305. [Google Scholar] [CrossRef]
- Zhang X, Wang C, Jin X, Hou Y, Zheng L. Optimization of extraction technology of total flavonoids from Passiflora edulis peel by ultrasonic assisted with complex enzyme and its antioxidant activity. Sci Technol Food Ind. 2022, 43, 215–22. [Google Scholar] [CrossRef]
- Sotiropoulou M, Katsaros I, Vailas M. Nonalcoholic fatty liver disease: the role of quercetin and its therapeutic implications. Saudi J Gastroenterol. 2021, 27, 319. [Google Scholar] [CrossRef] [PubMed]
- Liu X, Sun R, Li Z. Luteolin alleviates nonalcoholic fatty liver disease in rats via restoration of intestinal mucosal barrier and microbiota involving the gut-liver axis. Arch Biochem Biophys, 1: 711, 1090.
- Saravanan S, Parimelazhagan T. In vitro antioxidant, antimicrobial and anti-diabetic properties of polyphenols from Passiflora ligularis Juss. fruit pulp. Food Sci Hum Wellness. 2014, 3, 56–64. [Google Scholar] [CrossRef]
- Huang J, Li W, Liao W, Hao Q, Tang D, Wang D, Wang Y, Ge G. Green tea polyphenol epigallocatechin-3-gallate alleviates nonalcoholic fatty liver disease and ameliorates intestinal immunity in mice fed a high-fat diet. Food Funct. 2020, 11, 9924–9935. [Google Scholar] [CrossRef]
- Li Y, Wang Y, Wang Y, Wang Y, Zhang Y, Lu Y, et al. Tannic acid attenuates hepatic oxidative stress, apoptosis and inflammation in a high-fat diet-induced NAFLD rat model. Exp Ther Med. 2020, 20, 1–10. [Google Scholar] [CrossRef]
- De Souza Mesquita LM, de Souza EL, de Oliveira Lima E, de Freitas Araújo AA, de Almeida RN. Passiflora edulis: an insight into current researches on pharmacology and phytochemistry. Front Pharmacol 2020; 11:617. [CrossRef]
- Gonzalez L, Alvarez A, Murillo E, Guerra C, Mendez J. Potential uses of the peel and seed of Passiflora edulis f. edulis Sims (gulupa) based on chemical characterization, antioxidant, and antihypertensive properties. Asian J Pharm Clin Res. 2019, 12, 104–12. [Google Scholar]
- Wang Y, Li X, Zhang Y, Liu C, Zhang G. Harmine alleviates LPS-induced acute lung injury by inhibiting CSF3-mediated MAPK/NF-κB signaling pathway. Respir Res. 2023, 24, 65. [Google Scholar] [CrossRef]
- Zhang Y, Wang Y, Li X, Liu C, Zhang G. Alkaloids from traditional Chinese medicine against hepatocellular carcinoma: Mechanisms and perspectives. Biomed Pharmacother, 1: 137, 1113. [CrossRef]
- Sun S, Zhong H, Zhao Y. Indole alkaloids of Alstonia scholaris (L.) R. Br. alleviate nonalcoholic fatty liver disease in high-fat diet-fed mice. Nat Prod Bioprospect. 2022, 12, 1–11. [Google Scholar]
- Villada Ramos JA, Aguillón Osma J, Restrepo Cortes B, Loango Chamarro N, Maldonado Celis ME. Identification of potential bioactive compounds of Passiflora edulis leaf extract against colon adenocarcinoma cells. Biochem Biophys Rep 2023; 34:101453. [CrossRef]
- Maëlle Carraz, Cédric Lavergne, Valérie Jullian, Michel Wright, Jean Edouard Gairin, Mercedes Gonzales de la Cruz, Geneviève Bourdy. Antiproliferative activity and phenotypic modification induced by selected Peruvian medicinal plants on human hepatocellular carcinoma Hep3B cells, Journal of Ethnopharmacology, 2015, 166:185-199. doi.org/10.1016/j.jep.2015.02.028.
- Amaral G, Gomes V, Andrade N. Cytotoxic, antitumor and toxicological profile of Passiflora alata leaf extract. Molecules. 2020, 25, 4814. [Google Scholar] [CrossRef]
- Cui W, Chen S, Hu Q. Quantification and mechanisms of oleic acidinduced steatosis in HepG2 cells. American journal of translational research. 2010, 2, 95. [Google Scholar]
- Feng X, Zhang R, Yang Z, Zhang K, Xing J. Mechanism of metabolic dysfunction-associated steatotic liver disease: Important role of lipid metabolism. J Clin Transl Hepatol. 2024, 12, 815–826. [Google Scholar]
- Wang Y, Zhao L, Wang D, Huo Y, Ji B. Anthocyanin-rich extracts from blackberry, wild blueberry, strawberry, and chokeberry: antioxidant activity and inhibitory effect on oleic acid-induce d hepatic steatosis in vitro. J. Sci. Food Agric. 2016, 96, 2494–503. [Google Scholar] [CrossRef] [PubMed]
- Kocaadam B, Ozgurtas T, Ozcelik AO, Sezgin GC. Curcumin attenuates hepatic steatosis in high-fat diet-fed rats and oleic acid-treated HepG2 cells. Phytother Res. 2021, 35, 4382–92. [Google Scholar] [CrossRef]
- Park M, Yoo J, Lee Y, Lee J. Lonicera caerulea extract attenuates non-alcoholic fatty liver disease in free fatty acid-induced HepG2 hepatocytes and in high fat diet-fed mice. Nutrients. 2019, 11, 494.
- Le, H. , Choi, J., Jun, S. Ethanol extract of Liriope platyphylla root attenuates non-alcoholic fatty liver disease in high-fat diet-induced obese mice via regulation of lipogenesis and lipid uptake. Nutrients, 2021, 13, 3338. [Google Scholar] [CrossRef]
- Yin Y, Gao L, Lin H. improves non-alcoholic fatty liver disease in db/db mice by inhibition of liver X receptor activation to down-regulate expression of sterol regulatory element binding protein 1c. Biochemical and biophysical research communications. 2017, 482, 720–726. [Google Scholar] [CrossRef]
- Liu P, Wu P, Yang B. “et al”. Kaempferol prevents the progression from simple steatosis to non-alcoholic steatohepatitis by inhibiting the NF-κB pathway in oleic acid-induced HepG2 cells and high-fat diet-induced rats. Journal of Functional Foods. 2021, 85, 104655. [Google Scholar] [CrossRef]
- Kim J, Jeong E, Jo S, Park J, Kim J. Effects of light-controlled germination on saponarin content and lipid accumulation in HepG2 and 3T3-L1 cells using barley sprout extracts. Molecules. 2020, 25, 5349. [Google Scholar] [CrossRef]
- Han B, Yu Q, Yang L, Shen Y, Wang X. Kaempferol induces autophagic cell death of hepatocellular carcinoma cells via activating AMPK signaling. Oncotarget. 2017, 8, 86227–86239. [Google Scholar] [CrossRef]
- Zhu Y, Liu R, Shen Z, Cai G. Luteolin and lycopene protect against NAFLD via regulation of Sirt1/AMPK signaling. Life Sci, 2020: 256, 1179.
- Chang Y, Chen Y, Huang W, Liou C. Fucoxanthin attenuates fatty acid-induced lipid accumulation in FL83B hepatocytes through regulated Sirt1/AMPK signaling pathway. Biochemical and biophysical research communications. 2018, 495, 197–203. [Google Scholar] [CrossRef]
- Tie F, Ding J, Hu N. “et al”. Kaempferol and kaempferide attenuate oleic acid-induced lipid accumulation and oxidative stress in HepG2 cells. International journal of molecular sciences. 2021, 22, 8847. [Google Scholar] [CrossRef] [PubMed]
- Aguillón J, Maldonado M, Loango N. Antioxidant and antiproliferative activity of ethanolic and aqueous extracts from leaves and fruit juice of Passiflora edulis. Perspect Nutr Humana. 2013, 15, 13–25. [Google Scholar] [CrossRef]
- Muñoz O, Torres G, Nuñez J. New insights into the Folin-Ciocalteu reagent interaction with sugars during total polyphenol quantification. Rev Cienc Quím Biol. 2017, 20, 23–8. [Google Scholar]
- Armentano M, Bisaccia B, Miglionico R. Antioxidant and proapoptotic activities of Sclerocarya birrea [(A. Rich.) Hochst.] Methanolic root extract on the hepatocellular carcinoma cell line HepG2. BioMed Research International.
- López X, Taramuel A, Arboleda C, Segura F, Restrepo L. Comparación de métodos que utilizan ácido sulfúrico para la determinación de azucares totales. Revista cubana de Química, 2017; 29: 180-198.
- Prajapati D, Makvana A, Patel M, Chaudhary R, Patel R. Spectrophotometric method for the determination of total alkaloids in selected plant parts. J Pharmacogn Phytochem. 2024, 13, 491–494. [Google Scholar] [CrossRef]
- Vichai V, Kirtikara K. Sulforhodamine B colorimetric assay for cytotoxicity screening. Nature protocols, 2006, 1, 1112–1116. [Google Scholar] [CrossRef]
- Teixeira FS, Pimentel LL, Vidigal SSMP, Azevedo-Silva J, Pintado ME, Rodríguez-Alcalá LM. Differential lipid accumulation on HepG2 cells triggered by palmitic and linoleic fatty acids exposure. Molecules. 2023, 28, 2367. [Google Scholar] [CrossRef]
- Walker, J. The Bicinchoninic Acid (BCA) Assay for Protein Quantitation. The Protein Protocols Handbook. 2009, 11–15. [Google Scholar]
- Bligh G, Dyer W. A rapid method of total lipid extraction and purification. Canadian journal of biochemistry and physiology. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Oyama T, Toyota K, Waku T. “et al”. Adaptability and selectivity of human peroxisome proliferator-activated receptor (PPAR) pan agonists revealed from crystal structures. Acta Crystallographica Section D Biological Crystallography. 2009, 65, 786–795. [Google Scholar] [CrossRef]
- Artis R, Lin J, Zhang C. “et al”. Scaffold-based discovery of indeglitazar, a PPAR pan-active anti-diabetic agent. Proceedings of the National Academy of Sciences 2008. 106(1), 262–267.
- Jin L, Lin S, Rong H. “et al”. Structural Basis for iloprost as a dual peroxisome proliferator-activated receptor α/δ agonist. Journal of Biological Chemistry. 2011, 286, 31473–31479. [Google Scholar] [CrossRef]
- Kuwabara N, Oyama, T, Tomioka D. “et al”. Peroxisome proliferator-activated receptors (PPARs) have multiple binding points that accommodate ligands in various conformations: Phenylpropanoic acid-type PPAR ligands bind to PPAR in different conformations, depending on the subtype. Journal of Medicinal Chemistry, 2012. 55(2), 893–902.
- Kamata S, Oyama T, Saito K. “et al”. PPARα ligand-binding domain structures with endogenous fatty acids and fibrates. iScience. 2020, 23, 101727. [Google Scholar] [CrossRef]
- Capelli D, Cerchia C, Montanari R. Structural basis for PPAR partial or full activation revealed by a novel ligand binding mode. Scientific Reports. 2016, 6. [Google Scholar]
- Cronet P, Petersen W, Folmer R. “et al”. Structure of the PPARα and -γ ligand binding domain in complex with AZ 242; Ligand selectivity and agonist activation in the PPAR family. Structure. 2001, 9, 699–706. [Google Scholar] [CrossRef] [PubMed]
- Oon H, Hae S, Kim. “et al”. Design and synthesis of oxime ethers of α-acyl-β-phenylpropanoic acids as PPAR dual agonists. Bioorganic & Medicinal Chemistry Letters. 2007, 17, 937–941. [Google Scholar]
- Parraga A, Bellsolell L, Ferré-D’Amaré A, Burley K. Co-crystal structure of sterol regulatory element binding protein 1a at 2.3 å resolution. Structure. 1998, 6, 661–672. [Google Scholar] [CrossRef] [PubMed]
- Morris M, Huey R, Lindstrom W. “et al”. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. Journal of Computational Chemistry. 2009, 30, 2785–2791. [Google Scholar] [CrossRef]
- Trott O, Olson J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. Journal of Computational Chemistry.
- Koes R, Baumgartner P, Camacho J. Lessons learned in empirical scoring with smina from the CSAR 2011 Benchmarking Exercise. Journal of Chemical Information and Modeling. 2013, 53, 1893–1904. [Google Scholar] [CrossRef]
Figure 1.
Effect of ethanolic extract of P. edulis leaves on viability in HepG2 (A) and HFF (B) cells at 24 and 48 hours. Statistically significant differences between the control and treatment groups: *p<0,05; **p<0,01; ***p<0,001.
Figure 1.
Effect of ethanolic extract of P. edulis leaves on viability in HepG2 (A) and HFF (B) cells at 24 and 48 hours. Statistically significant differences between the control and treatment groups: *p<0,05; **p<0,01; ***p<0,001.
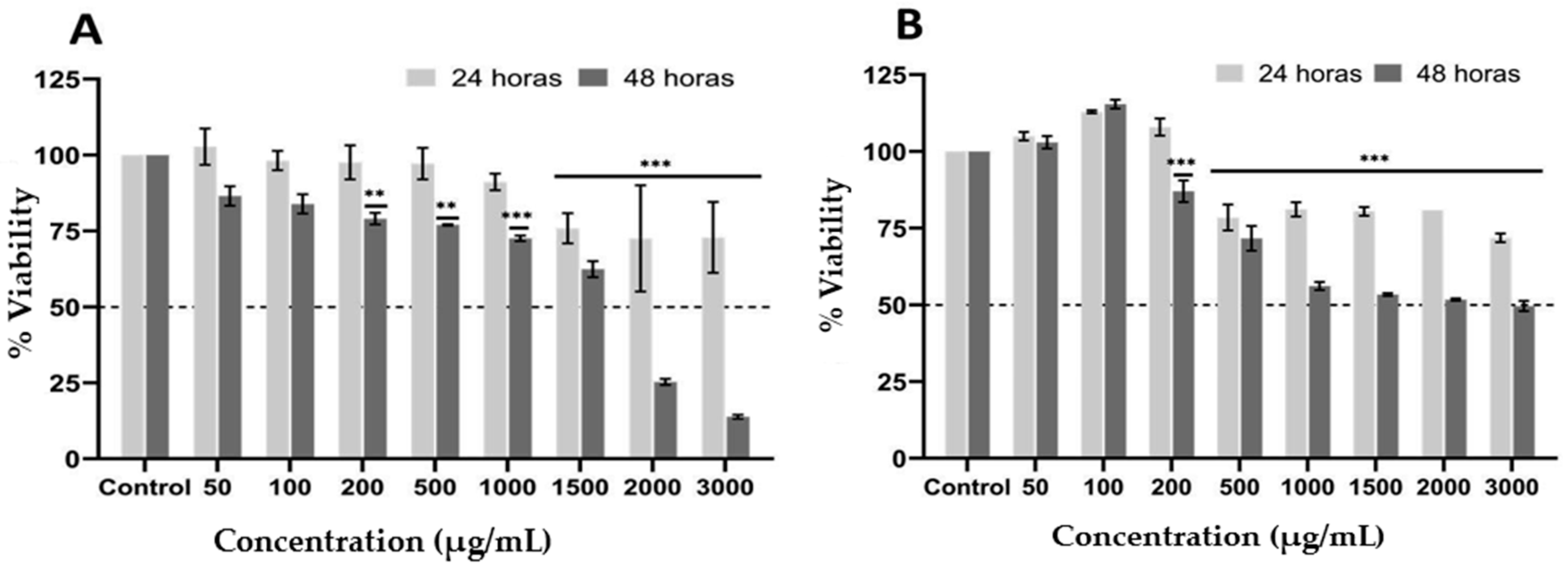
Figure 2.
Effect of ethanolic extract of P. edulis leaves on the total triglyceride content of both intracellular (A) and extracellular (B) HepG2 cells in response to 24- and 48-hour incubation. Control; untreated cells; AO, cells with oleic acid (100µM); AO50, oleic acid and ethanolic leaf extract 50 µg/mL; AO100, oleic acid and ethanolic leaf extract 100 µg/mL; AO200, oleic acid and ethanolic leaf extract 200 µg/mL; 200 EE, ethanolic leaf extract 200 µg/mL alone.
Figure 2.
Effect of ethanolic extract of P. edulis leaves on the total triglyceride content of both intracellular (A) and extracellular (B) HepG2 cells in response to 24- and 48-hour incubation. Control; untreated cells; AO, cells with oleic acid (100µM); AO50, oleic acid and ethanolic leaf extract 50 µg/mL; AO100, oleic acid and ethanolic leaf extract 100 µg/mL; AO200, oleic acid and ethanolic leaf extract 200 µg/mL; 200 EE, ethanolic leaf extract 200 µg/mL alone.
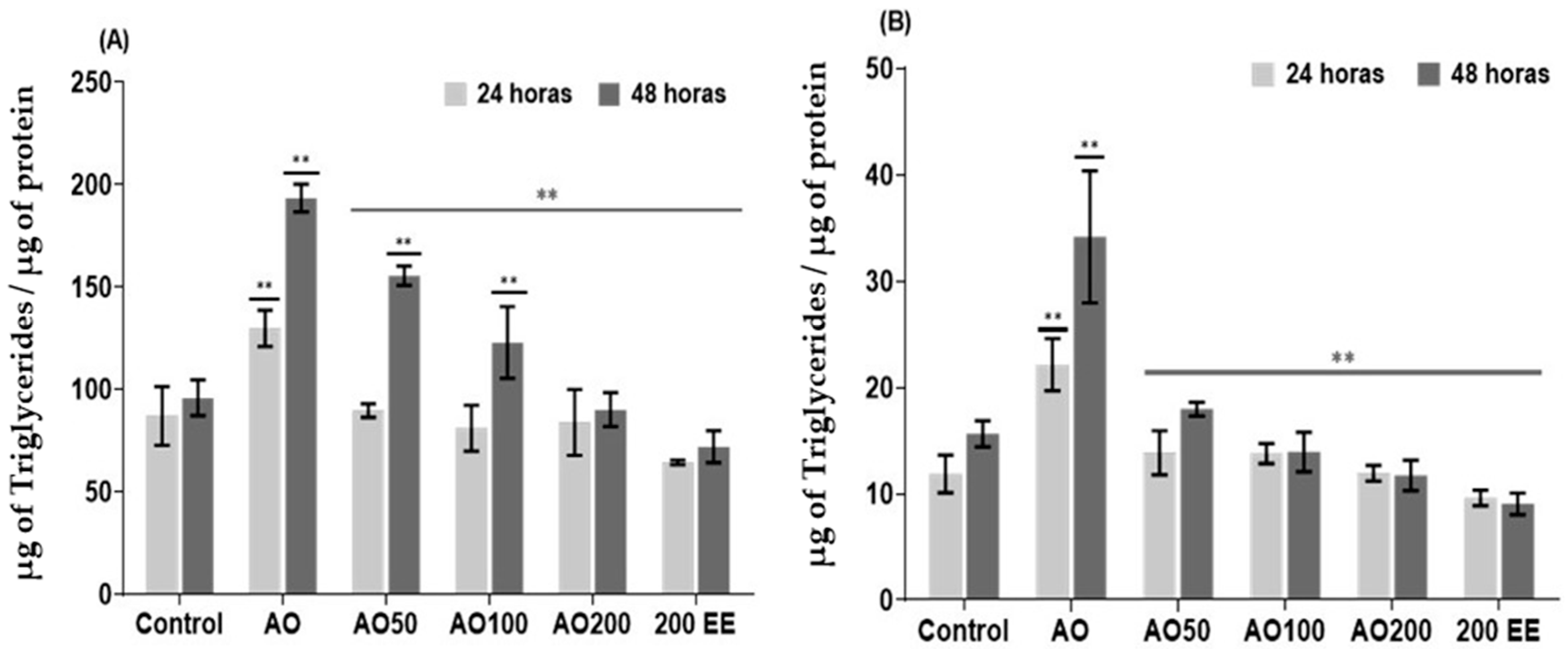
Figure 3.
Effect of ethanolic extract of P. edulis leaves on the total cholesterol content of both intracellular (A) and extracellular (B) HepG2 cells in response to 24- and 48-hour incubation. Control; untreated cells; AO, cells with oleic acid (100µM); AO50, oleic acid and ethanolic leaf extract 50 µg/mL; AO100, oleic acid and ethanolic leaf extract 100 µg/mL; AO200, oleic acid and ethanolic leaf extract 200 µg/mL; 200 EE, ethanolic leaf extract 200 µg/mL alone.
Figure 3.
Effect of ethanolic extract of P. edulis leaves on the total cholesterol content of both intracellular (A) and extracellular (B) HepG2 cells in response to 24- and 48-hour incubation. Control; untreated cells; AO, cells with oleic acid (100µM); AO50, oleic acid and ethanolic leaf extract 50 µg/mL; AO100, oleic acid and ethanolic leaf extract 100 µg/mL; AO200, oleic acid and ethanolic leaf extract 200 µg/mL; 200 EE, ethanolic leaf extract 200 µg/mL alone.
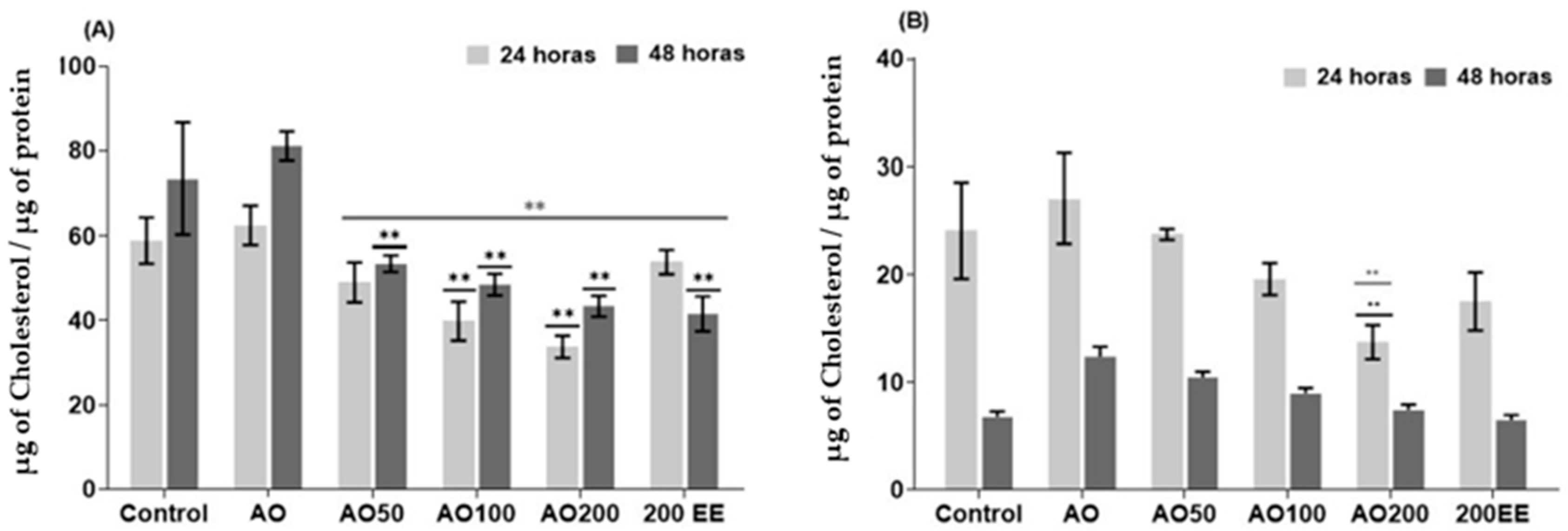
Figure 4.
Average binding affinities of each compound for each protein. (A) shows the results obtained by Autodock Vina and (B) by the Smina software. The compounds that showed the best results for PPARα and SREBP-1 are shaded in purple and green, respectively.
Figure 4.
Average binding affinities of each compound for each protein. (A) shows the results obtained by Autodock Vina and (B) by the Smina software. The compounds that showed the best results for PPARα and SREBP-1 are shaded in purple and green, respectively.
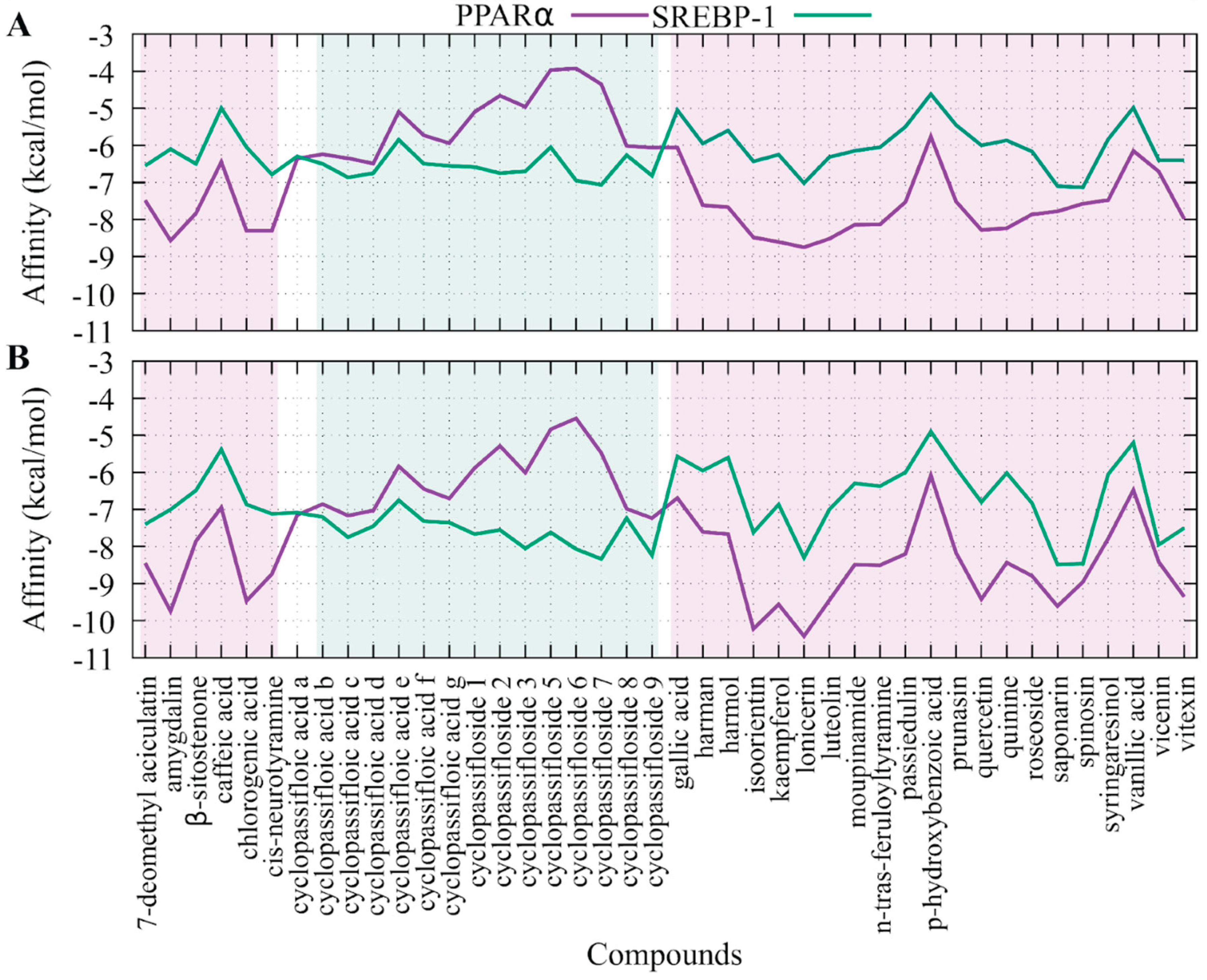

Table 1.
Phytochemical characterization of the EE of P. edulis.
| Metabolite type | Total metabolite content (mgEQ) |
|---|---|
| Phenols | 235.7 ± 9.7 |
| Polysaccharides | 221.9 ± 16.4 |
| Flavonoids | 182.4 ± 7.9 |
| Tannins | 45.1 ± 3.5 |
| Alkaloids | 8.3 ± 1.1 |
Table 2.
Final consensus of the results of molecular docking of the phytochemical of P. edulis compounds on transcription factors.
Table 2.
Final consensus of the results of molecular docking of the phytochemical of P. edulis compounds on transcription factors.
| Compound | PPARα | SREBP-1 | MIX |
|---|---|---|---|
| 7deomethylaciculatin | 16.70 | 13 | 14 |
| Amygdalin | 5.19 | 24 | 13.5 |
| chlorogenic acid | 8.38 | 27 | 16.5 |
| cisnfeuroyltyramine | 11.57 | 14 | 12 |
| cyclopassifloic acid c | 27.99 | 7.5 | 17 |
| cyclopassifloicacid d | 27.18 | 11.5 | 18 |
| cyclopassifloside1 | 35.27 | 11 | 23 |
| cyclopassifloside3 | 32.7 | 8.5 | 21 |
| cyclopassifloside6 | 40.85 | 5.5 | 23.5 |
| cyclopassifloside7 | 39.15 | 3 | 22 |
| cyclopassifloside9 | 27.05 | 6.5 | 15.5 |
| Isoorientin | 5.05 | 15 | 8 |
| Kaempferol | 6.85 | 24 | 15 |
| Lonicerín | 5.4 | 4.5 | 3.5 |
| Luteolin | 7.05 | 22 | 14 |
| Quercetin | 8.95 | 29.5 | 18.5 |
| Saponarin | 10.85 | 1 | 4.5 |
| Spinosin | 12.9 | 2 | 6.5 |
| Vitexin | 11.2 | 18 | 13.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



