
Submitted:
12 August 2025
Posted:
14 August 2025
You are already at the latest version
Abstract
A new species, Fragilaria longwania sp. nov., from Sihailongwan Maar Lake in northeastern China, is described on the basis of observations made using light microscopy (LM) and scanning electron microscopy (SEM) that reveal the distinctive features that separate this new taxon from the other species currently placed in the genus Fragilaria. Fragilaria longwania sp. nov. has relatively short valve length, high stria density, spindle-shaped valves with a swollen and sometimes deformed central area and capitate valve apices, short spines and weakly developed apical pore fields. This combination of character was found to be unique when the new species was compared with morphologically similar taxa. This species was found in various samples collected from Sihailongwan Maar Lake between 2003 and 2017. The analysis of the diatom assemblages collected in the sediment trap samples allowed an assessment of the cell size variation, seasonal occurrences and ecological requirements of this species. Fragilaria longwania appears to be a planktonic diatom associated with the spring season.
Keywords:
Fragilariaceae
; taxonomy
; planktonic diatom
1. Introduction
In the past few years, considerable progress has been made on the revision of the genus Fragilaria Lyngbye (1819: 185)[1], with the reanalysis of historic samples and type materials. These analyses have provided a better understanding for some of the most reported species belonging to Fragilaria as well as the description of numerous new species (e.g., [2,3,4,5]).
By contrast, few new species have been described from material collected in China. According to the list compiled by Kociolek et al. [6], only one new species of Fragilaria has been described from China between 2000 and 2019 (i.e. Fragilaria crenophila var. sinensis Rioual [7]). Since the publication of this list, Fragilaria huebeneri A. Schwarz, KJ Krahn et CE Wetzel was described from material collected from Lake Namco, on the Tibetan Plateau [8]. When compared with the large number of new species belonging to Ulnaria, a genus closely related to Fragilaria, that were recently described from China (e.g., [9]), it is very likely that the diversity of the genus Fragilaria in China has been understudied.
As part of an ongoing research project on the modern and down-core diatom assemblages of Sihailongwan Maar Lake (Northeast China), a species of Fragilaria, that was found mostly in sediment trap samples, could not be identified using the currently available literature. On the basis of observations made using light microscopy (LM) and scanning electron microscopy (SEM) and after comparisons with known species of the genus, this Fragilaria is described as new-to-science.
2. Materials and Methods
2.1. Study Site
The study site, Sihailongwan Maar Lake, is located in the Long Gang volcanic field Jilin Province, in Northeast China (42°17’N, 126°36’E, 790 m a.s.l., Figure 1). This lake has is a typical maar lake, i.e. a volcanic lake formed by a phreatomagmatic explosion. Its morphology is therefore shaped like a bowl, with steep rocky slopes and a flat bottom, a relatively small surface area (0.5 km2) but a deep water depth (maximum 50 m). Detailed information on the geology, recent climate and vegetation of the region around the lake as well as on the lake limnology can be found in previous studies on this lake [10,11,12].
As shown by the analyses on short cores and trap material, the sediment sequence of this lake is composed of seasonal laminations, or varves, which correspond to the alternation of allogenic atmospheric dust inputs that correspond to the winter/spring season and authigenic material, mainly composed of algal remains including diatoms, deposited during the spring, summer and autumn seasons [13,14]. The high quality of its sediment sequence, that store a record of environmental and human impacts at a continental and global scale over the past millennia [11], made this site particularly suitable for palaeolimnological investigations. Considering these characteristics, Lake Sihailongwan was selected as a candidate site to become a Global boundary Stratotype Section and Point (GSSP) for the demarcation of the Anthropocene [15].
2.2. Limnology of Lake Sihailongwan
Between August 2020 and November 2014, chemical variables such as pH, alkalinity (Gran titration), electrical conductivity and the concentrations for the main cations, anions and nutrients were measured on water samples collected at the same time as sediment trap samples were retrieved. On a few occasions, the depth of the photic zone was assessed using a Secchi disk. Details on the water chemistry methods that were used to measure these variables are provided in Rioual et al. [16] and a summary of the results is given in Table 1.
2.3. Diatom Sampling and Sample Preparation
Material for diatom analysis was obtained from sediment traps deployed at the center of Lake Sihailongwan between May 2003 and July 2017. The mooring was designed so that the traps collected all material, organic and inorganic, that settle into the lake water column but that the content of the trap would not be affected by possible resuspension from the bottom sediment. Samples were collected monthly between 2002 and 2005 and bimonthly from 2006. In total 187 sediment trap samples were collected and analyzed for this study. In addition to the trap samples, were also analyzed three periphyton samples scrapped from the surface of the buoy that maintain the trap mooring in the water column, two surface sediment samples (the top ~1cm, representing recent epipelic assemblage as well as several years of sedimentation), that were collected from the middle of the lake (~50 m water depth) on the 21st October 2005 and 14th August 2007, and a series of benthic samples (epilithon, epiphyton, epipelon and epixylon) collected in the littoral zone of the lake at different times during this study. Details of these samples are given in Table 3. The purpose of analyzing these additional samples was to determine the preferred habitat of the new species.
Diatom samples were prepared according to the standard procedure described in Battarbee et al. [17] using hydrogen peroxide. Slides for light microscopy were prepared by mounting the cleaned suspension in Naphrax®. LM observations were made at ×1000 magnification using a Leica DM 2500 equipped with immersion objectives (N Plan, 1.25 numerical aperture) and differential interference contrast (DIC). Photomicrographs were captured using a Leica DFC450 camera (Leica Microsystems Ltd). For scanning electron microscope analyses, aliquots of cleaned suspensions were filtered and rinsed through a 3-µm Isopore™ polycarbonate membrane (Merck, Darmstadt, Germany). The membrane filters were the mounted on aluminum stubs using double-coated carbon conductive adhesive tape and coated with gold using a Quorum Q150R S Plus sputter coater (Quorum Technologies, Laughton, UK). To illustrate the morphological variability of the species, we provide images captured from various samples collected at different locations in the lake (e.g. sediment trap, surface sediment from the deepest point, epipelon from the littoral zone of the lake) and in different years and season. The type of sample and the time of collection are given in the captions of the images. Through the years, different scanning electron microscopes were used. Most of the images reproduced in this paper were taken with a Phenom XL desktop SEM (Nanoscience Instruments, Phoenix, USA) at IGG-CAS (Beijing, China) operated at 10 kV.
3. Results
3.1. New Species Description
Division: Bacillariophyta
Class: Fragilariophyceae – F.E. Round 1990 [18].
Subclass: Fragilariophycidae – Round & Crawford
Order: Fragilariales – Silva, 1962 sensu emend. [19].
Family: Fragilariaceae – Greville 1833 [20].
Genus: Fragilaria Lyngbye 1819 [1].
Fragilaria longwania Rioual sp. nov.
Description: LM (Figure 2): Frustules in girdle view rectangular, solitary or occasionally two cells connected to each other (Figure 2V). Valves spindle-shaped to elongated, narrowly lanceolate with clearly convex, never parallel margins, tapering from a broadened central area towards protracted, distinctly capitate apices, often deflected. Teratological forms with irregularly undulating or bent valve outlines often observed (Figure 2B, Figure 2T). Valve dimensions: length 15—44 µm (n= 1078, LM and SEM), width 2.0—3.5 µm (from SEM images only, n=36). Sternum distinct but narrow, gradually widening towards the central area. Central area swollen, forming small, irregularly shape hyaline zone, occasionally absent. Striae alternating, parallel throughout entire valve, 21—25 in 10 µm. Areolae sometimes discernible in LM under differential interference contrast.
SEM (Figure 3Figure 4Figure 5Figure 6Figure 7 and Figure 8): Series of marginal spines, generally emerging from vimines, but sometimes from virgae, occurring more or less evenly from apex to apex (Figure 3C, Figure 5C, Figure 6C, Figure 7B). Spines acute, shaped like shark tooth (Figure 3B-C and Figure 8A). Up to three tiny spines present at apices (Figure 4B, Figure 5B and Figure 6B). Valve face flat with weakly raised virgae (Figure 3A, Figure 5c and Figure 7c). Central area with ghost striae (Figure 3D, Figure 5C, Figure 6C and Figure 7B). Striae uniseriate, composed of several (up to 6) rounded areolae (Figure 5C, Figure 6C, Figure 7B) becoming transapically elongated on the mantle (Figure 8A-B), 62—74 in 10µm (n= 35). Virgae much wider than the striae. Apical pore field of the ocellulimbus type, not well developed, composed of only one or two rows of small rounded poroids (Figure 3A and Figure 3E, Figure 4B-C, Figure 5B, Figure 5D and 8B-C). One rimoportula is present per valve, positioned at a distal end (Figure 3A, Figure 4A, Figure 5A and Figure 7A). Externally, rimoportula opening small, rounded but not rimmed, generally located at the centre of the last stria (Figure 3B, Figure 4B, Figure 5B, Figure 6B and Figure 7D). Internal rimoportula opening more or less elongated, oblique (Figure 4C). Girdle composed of only two or three open bands, ligulate, tapering and opening at the valve pole (Figure 8A-C). Each band with one row of poroids (Figure 4C and Figure 8A-C).
Holotype: 20 April 2005, collected by P. Rioual, slide number IGGDC-DB-SIHA-0504 (herb. IGGDC) partly shown here in Figure 2, holotype specimen: illustrated in Figure 2Z, located using England Finder J27/1.
Paratype: BM 101 673 in the Natural History Museum, London, UK. (Note: this is the same slide as for Staurosira longwanensis that was described in Rioual et al. [21].
Registration:http://phycobank.org/xxxxxxxxxx (pending)
Type locality: China, Lake Sihailongwan, Jilin Province (E: 126°35’51’’–126°36’24’’, N: 42°17’0’’–42°17’24’’; Elevation: 797 m above sea level)
Etymology: For naming this species we followed Dubois [22] who recommends keeping species names as simple and short as possible in order to be euphonious in all languages and easily memorable. The epithet of the new species refers to “Long wan” the Chinese name for the maar lakes of the Long Gang volcanic field in Jilin Province and which means “Dragon bowl” in English.
Distribution: So far only observed in the type locality.
3.2. Analysis of the Trap Samples
In Lake Sihailongwan, during the 15-year period of investigation, two centric diatoms, Discostella stelligeroides (Hustedt) Houk & Klee and Lindavia balatonis (Pantocsek) Nakov, Guillory, Julius, Theriot & Alverson largely dominated the sediment trap assemblages. Fragilaria longwania was present in 139 of the 187 trap samples analyzed, with a maximum abundance of 19.8% recorded at the beginning of the study in June 2003 and a mean abundance of 2.1%. Over the study period, a general decline in relative abundances is observed (Figure 9A). On the other hand, during the same period the mean valve length follows a positive trend (Figure 9B).
The seasonal distribution of F. longwania was investigated by plotting its relative abundances against Julian day on a year-to-year basis (Figure 10). In most years, such as 2003, 2004, 2007, 2008, 2009, 2010, 2011, 2103, 2014, abundances of F. longwania exhibit a peak during the spring period (April-May-June). In some years (2003, 2004, 2013), a second peak occurred in the autumn (September-October). Only in 2006, the seasonal distribution of F. longwania was characterized by a single autumnal peak. Generally, F. longwania had low abundances during the summer season, which is characterized not only by high temperature, but also high precipitation, low wind speed and low sunshine (due to high cloud cover).
3.3. Analysis of Additional Samples
Discostella stelligeroides and Lindavia balatonis, the two centric diatoms that dominate the assemblages collected in the sediment trap samples, are also present and often dominant in all the other types of samples collected in the lake. These include periphyton samples collected from the mooring buoy and in the various benthic samples from the littoral zone (Table 2).
For F. longwania, the highest percentages were found in sediment trap samples (up to 19.8%). Unsurprisingly considering its abundance in trap samples, F. longwania is also among the most abundant species in the surface sediment samples collected from the middle of the lake reaching 1.8% in October 2005 and 1.2% in August 2007 (Table 2). It is also found in the periphyton samples collected from the mooring buoy, although its abundance in this type of sample is also declining and therefore follows the same trends observed in the trap samples. F. longwania can be completely absent in the benthic samples collected from the littoral zone and when present this species never reaches 3% of the assemblages.
4. Discussion
4.1. Generic Placement of the New Species
As discussed by Williams [23], the genus Fragilaria is currently aphyletic, i.e. without identified synapormophy, and so far, recent attempts to separate new genera from Fragilaria have not been entirely convincing [7,24,25]. Nevertheless, F. longwania shares many characters with what Williams [23] refers as `classical’ species of Fragilaria, including the presence of one rimoportula on one apex, apical pore fields of the ocellulimbus type, uniseriate striae and open girdle bands. F. longwania is therefore best placed in that genus. Among the groups of Fragilaria species that Williams [23] recognized according to the occurrence and placement of their marginal spines, F. longwania is closest to the group 3, i.e. `classical’ species with spines that emerge from the vimines, even if some spines appear to emerge from virgae. In that group, are included Fragilaria tenera var. nanana and Fragilaria billingsi, two of the species that closely resemble F. longwania (see below).
4.2. Comparison with Other Species of the Genus Fragilaria
The new species described here is comparable to several other species of Fragilaria (Table 3), especially to species of the Fragilaria rumpens complex which has been recently reviewed by Van de Vijver et al. [3] who analyzed the type material of numerous species described in the 19th century.
One of the most similar species is Fragilaria campyla (Hilse) Van de Vijver, Kusber et DM Williams, which largely overlaps with F. longwania in terms of valve length (35—45 µm vs 15—44 µm) and width (2.5—3.0 µm vs 2.0—3.5 µm). F. campyla however lacks a swollen area and its valves are linear to linear-lanceolate, not spindle-shaped to elongated and narrowly lanceolate. In F. campyla the ocellulimbi are also larger than in F. longwania [3].
Fragilaria parva (Grunow) Tuji et DM Williams, is another species of the F.rumpens complex with similar ranges of valve dimensions compare to F. longwania. In addition, F. parva also possesses a swollen area and shark tooth shaped spines. However, F. parva differs by having linear to linear-lanceolate valve outline, a large hyaline rectangular zone in the central area and well-developed ocellulimbi [3].
Fragilaria metcalfeana Van de Vijver, DM Williams et Kusber, is a fossil species originally described from Neogene material collected in Dubravica (Croatia). This taxon has also a distinctly inflated central area, but its valves are much longer (60—120 µm) and characterized by a well-delimited hyaline central area and large ocellulimbi at the apices [3].
Fragilaria pseudofamiliaris Van de Vijver, TM Schuster, Kusber et DM Williams, has similar valve dimensions (length 30—50 µm and width 2.0—3.0 µm) and protracted subcapitate to capitate apices. However, this species differs from F. longwania by having elongated, narrowly lanceolate valve, with broad linking spines that help this species in producing long ribbon-like colonies, that we never observed in F. longwania. In addition, this species has a lower stria density (18—19 striae in 10 microns) and well-developed ocellulimbi composed of at least 4 rows of small poroids [3].
Last among the species of the Fragilaria rumpens complex that could be usefully compared to F. longwania is Fragilaria scotica (Grunow) Van de Vijver, CE Wetzel et Ector. This species also possesses a clearly inflated central area. Its outline is however not spindle-shaped, but linear-lanceolate with gradually narrowing margins and protracted, rostrate apices. In addition, F. scotica has much lower stria density than F. longwania with 15 instead of 21-25 striae in 10 microns [3].
Outside the F. rumpens complex, several more species can be compared to F. longwania. Fragilaria nanoides Lange-Bertalot, which was described from Julma Ölkky, an oligo-dystrophic lake from Finland, has a very similar silhouette, and overlapping ranges of stria density (22.5—23 vs 21—25 striae in 10 µm). However, F. nanoides has longer (40—90 µm) and narrower valves (1.8—2.4 µm) [26].
Fragilaria billingsi Wengrat, C.E.Wetzel et E.Morales, that was described from a reservoir in southern Brazil, is another species with an inflated central area, but its valves are longer (54—76 µm), have a lower stria density (17—20 striae in 10 microns), clearly raised costae and well-developed ocellulimbi [27].
Fragilaria huebeneri A.Schwarz, KJ Krahn et CE Wetzel, which was described from Nam Co, a large subsaline lake in Tibet, has spindle-shaped valves that are often curved, like what can be observed in F. longwania. However, in F. huebeneri the central area is less inflated, the valves are more narrowly tapering towards subcapitate apices. In addition, F. huebeneri has much longer valves (99—121 µm), possesses raised costae and spatula-shaped spines that allow this species to form ribbon-like colonies. The ocellulimbi are also well-developed with 3-5 rows of poroids [8].
Fragilaria stoermeriana Alexson, Reavie et Van de Vijver, described from plankton samples collected in the eastern basin of Lake Superior (Ontario, Canada), also shares some characteristics with F. longwania such as a distinctly swollen central area, protracted capitate ends, weakly raised virgae, acute spines irregularly scattered along the entire margin of the valve and small ocellulimbi at the apices. The two species have also similar stria density (20—24 vs 21—24 striae in 10 microns). However, F. stoermeriana is much longer (60—110 µm) and can form stellate colonies [28].
Finally, the apparently cosmopolitan Fragilaria tenera var. nanana (Lange-Bertalot) Lange-Bertalot et S.Ulrich partly overlaps with F. longwania in terms of valve dimensions as length and width of its valves range between 29—85 µm and 2.0—2.3 µm, respectively. However, the stria density of the variety nanana is lower (18.5—20 instead of 21—24 in 10 microns) as reported by Lange-Bertalot and Ulrich [29] and the outline of the valves, even for the short ones, is not spindle-shaped but lanceolate with a barely inflated central area.

Table 3.
Selected morphological characteristics of Fragilaria longwania sp. nov. and of other species of Fragilaria sharing a broadly similar outline.
Table 3.
Selected morphological characteristics of Fragilaria longwania sp. nov. and of other species of Fragilaria sharing a broadly similar outline.
| Taxon | Length (µm) | Width (µm) | Striae in 10 µm | Valve outline | Central area | Pore fields | Distribution | Reference |
| F.longwania | 15—44 | 2.0—3.5 | 21—25 | spindle shaped | clearly swollen | small, only one or two rows of small poroids | Lake Sihailongwan, NE China | This study |
| F.campyla | 35—45 | 2.5—3.0 | 19—20 | linear to linear-lanceolate | not swollen | well-developed, several rows of small poroids | Europe | [3] |
| F. parva | 20—40 | 3.0—3.5 | 19—21 | linear to linear-lanceolate | swollen | well-developed, 4 rows of large poroids | Europe | [3] |
| F. metcalfeana | 60—120 | 2.5—3.5 | ca. 18 | linear but strongly elongated | distinctly inflated | large, 2-3 rows of large poroids | Fossil, Neogene, Dubravica, Croatia | [3] |
| F. pseudofamiliaris | 30—50 | 2.0—3.0 | 18—19 | elongated, narrowly lanceolate | not swollen | well-developed, at least 4 rows of small poroids | Artesian well, Dresden, Germany | [3] |
| F. scotica | 65 | 4 | 15 | linear-lanceolate | clearly inflated | not observed | Loch Leven, Scotland | [3] |
| F. nanoides | 40—90 | 1.8—2.4 | 22.5—24 | linear-lanceolate | weakly inflated | not observed | Lake Julma Ölkky, Finland | [26] |
| F.billingsii | 54—76 | 2.0—2.5 | 17—20 | narrow lanceolate | bilaterally gibbous | well-developed | Reservoir, Sao Paulo State, Brazil | [27] |
| F.huebeneri | 99—121 | 2.0—4.0 | 17—21 | spindle to needle-shaped, narrowly lanceolate | indistinct | well-developed, 3-5 rows of poroids | Lake NamCo, Tibet,China | [8] |
| F.stoermeriana | 60—110 | 1.5—3.0 | 20—24 | long, lanceolate | distinctly swollen | small ocellulimbi | Lake Superior, Ontario, Canada | [28] |
| F. tenera var. nanana | 29—85 | 2.0—2.3 | 18.5—20 | linear to linear-lanceolate | barely inflated | well-developed, 3 rows of poroids | cosmopolitan | [29] |
4.3. Ecology, Seasonality, Cell-Size and Distribution
From the results of our water chemistry analyses (Table 1), Lake Sihailongwan has circumneutral, weakly alkaline water. Considering the median values for total phosphorus concentration and Secchi disk depth of ~15 µg/L and 3.8 m, respectively, the trophic status of Lake Sihailongwan can be considered as mesotrophic. The highest abundances of this species were observed in trap samples. It is also found in the periphyton attached to the mooring buoy and in the subfossil assemblages found in the surface sediment collected at the bottom of the water column. Together, these data on the occurrences and abundances in samples collected from the middle of the lake and the fact that it was never found in large abundance in the benthic samples collected from the littoral zone suggest that this species is part of the diatom plankton.
From the series of trap samples, F. longwania appears to be associated to the spring season (and in some years, to the autumn). In the region of Lake Sihailongwan meteorological conditions during spring are characterized by low temperatures, high wind speed and sunny conditions (Figure 10). During the 15 year-long period we investigated for this study, valve length of F. longwania tended to increase with decrease in valve abundance (Figure 9). A similar inverse relationship between cell size and density was observed for another species of Fragilaria by Soumya et al. [30] in their investigation of a natural planktonic population from Lake Biwa (Japan). According to these authors, this relationship indicates occurrence of cell division. If this is true for F. longwania in Lake Sihailongwan, it would suggest that conditions at the beginning of the study period (from 2003 to 2013) were more favorable for the growth of the population by vegetative cell division than at the end of the study period (2014-2017), when less numerous and longer cells were found, suggesting lower rates of cell division. Temperature and nutrients are thought to be key factors that affect such changes in cell size and density but our water chemistry data only cover a short part of the study period and a more detailed investigation would be required to unveil such relationships.
F. longwania is the second diatom species described from Lake Sihailongwan after Staurosira longwanensis Rioual, Morales & Ector [21]. So far, F. longwania has not been observed in other lakes from NE China or anywhere else but considering the relatively understudied diversity of the genus Fragilaria in China, it is probably premature to consider it as a species endemic to Lake Sihailongwan.
Finally, preliminary diatom analyses of the long sedimentary sequence of Lake Sihailongwan [11] indicate that Fragilaria longwanensis has been present in the diatom flora of this lake for at least the past 20,000 years. Its population however, appears to be declining as suggested by the trends we observed (Figure 9 and Figure 10) in the trap samples we collected over a 15 year-long period.
Author Contributions
Conceptualization P.R.; Methodology P.R.; Investigation P.R.; Resources, P.R.; Data curation, P.R.; Writing–original draft, P.R.; Writing–review & editing, P.R. and G.C.; Funding acquisition, G.C. and J.L. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the National Natural Science Foundation of China (grants no. 42488201 and 42030507).
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
The authors would like to thank Mr Xiaobo Yuan for maintaining the sediment trap mooring in Lake Sihailongwan for many years and collecting samples from it. We are grateful to Dr Bart Van de Vijver (Meise Botanic Garden, Belgium) for his constructive advice on the taxonomy of Fragilaria species.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| SEM | Scanning Electron Microscopy |
| LM | Light Microscopy |
References
- Lyngbye, H.C. Tentamen hydrophytologiae danicae continens omnia hydrophyta cryptogama Daniae, Holsatiae, Faeroae, Islandiae, Groenlandiae hucusque cognita, systematice disposita, descripta et iconibus illustrata, adjectis simul speciebus norvegicis. Hafniae [Copenhagen]: typis Schultzianis, in commissis Librariae Gyldendaliae. 1819, pp. [i]–xxxii, [1]–248, 70 pls. [CrossRef]
- Van de Vijver, B.; Williams, D.M.; Kelly, M.; Jarlman, A.; Wetzel, C.E.; Ector, L. Analysis of some species resembling Fragilaria capucina (Fragilariaceae, Bacillariophyta). Fottea 2021, 21, 128–151. [Google Scholar] [CrossRef]
- Van de Vijver, B.; Williams, D.M.; Schuster, T.M.; Kusber, W.-H.; Cantonati, M.; Wetzel, C.E.; Ector, L. Analysis of the Fragilaria rumpens complex (Fragilariaceae, Bacillariophyta) with the description of two new species. Fottea 2022, 22, 93–121. [Google Scholar] [CrossRef]
- Van de Vijver, B.; Williams, D.M.; Kusber, W.-H.; Cantonati, M.; Hamilton, P.B.; Wetzel, C.E.; Ector, L. Fragilaria radians (Kützing) D. M. Williams et Round, the correct name for F. gracilis (Fragilariaceae, Bacillariophyta): a critical analysis of this species complex in Europe. Fottea 2022, 22, 256–291. [Google Scholar] [CrossRef]
- Van de Vijver, B.; Schuster, T.M.; Jónsson, G.S.; Hansen, I.; Williams, D.M.; Kusber, W.-H.; Wetzel, C.E.; Ector, L. A critical analysis of the Fragilaria vaucheriae complex (Bacillariophyta) in Europe. Fottea 2023, 23, 62–96. [Google Scholar] [CrossRef]
- Kociolek, J.P.; You, Q.; Liu, Q.; Liu, Y.; Wang, Q. Continental diatom biodiversity discovery and description in China: 1848 through 2019. PhytoKeys 2020, 160, 45–97. [Google Scholar] [CrossRef]
- Rioual, P.; Flower, R.J.; Chu, G.; Lu, Y.; Zhang, Z.; Zhu, B.; Yang, X. Observations on a fragilarioid diatom found in inter-dune lakes of the Badain Jaran Desert (Inner Mongolia, China), with a discussion on the newly erected genus Williamsella Graeff, Kociolek & Rushforth. Phytotaxa 2017, 329, 28–50. [Google Scholar] [CrossRef]
- Krahn, K.J.; Schwarz, A.; Wetzel, C.E.; Cohuo-Durán, S.; Daut, G.; Macario-González, L.; Pérez, L.; Wang, J.; Schwalb, A. Three new needle-shaped Fragilaria species from Central America and the Tibetan Plateau. Phytotaxa 2021, 479, 1–22. [Google Scholar] [CrossRef]
- Liu, B. The diatom genus Ulnaria (Bacillariophyceae) in China. PhytoKeys 2023, 228, 1–118. [Google Scholar] [CrossRef]
- Stebich, M.; Rehfeld, K.; Schlütz, F.; Tarasov, P.; Liu, J.; Mingram, J. Holocene vegetation and climate dynamics of NE China based on the pollen record from Sihailongwan Maar Lake. Quaternary Science Reviews 2015, 124, 275–289. [Google Scholar] [CrossRef]
- Mingram, J.; Stebich, M.; Schettler, G.; Hu, Y.; Rioual, P.; Nowaczyk, N.; Dulski, P. , You, H.; Opitz, S.; Liu, Q.; Liu, J. Millennial-scale East Asian monsoon variability of the last glacial deduced from annually laminated sediments from Lake Sihailongwan, N. E. China. Quaternary Science Reviews 2018, 201, 57–76. [Google Scholar] [CrossRef]
- Li, P.; Chu, G.; Rioual, P.; Zhan, N.; Zhang, G.; Zhu, Z.; Qi, L.; Xie, M.; Sun, Q. Independent temperature records since the last deglaciation from the varved sediments of Sihailongwan maar lake, northeastern China. Quaternary Science Reviews 2025, 349, 109139. [Google Scholar] [CrossRef]
- Chu, G.; Liu, J.; Schettler, G.; Li, J.; Sun, Q.; Gu, Z.; Lu, H.; Liu, Q.; Liu, T. Sediment fluxes and varve formation in Sihailongwan, a maar lake from northeastern China. Journal of Paleolimnology 2005, 34, 311–324. [Google Scholar] [CrossRef]
- Schettler, G.; Liu, Q.; Mingram, J. , Negendank, J.F.W. Palaeovariations in the East-Asian monsoon regime geochemically recorded in varved sediments of Lake Sihailongwan (Northeast China, Jilin Province). Part 1: Hydrological conditions and dust flux. Journal of Paleolimnology 2006, 35, 239–270. [Google Scholar] [CrossRef]
- Han, Y.; An, Z.; Lei, D.; Zhou, W.; Zhang, L.; Zhao, X.; Yan, D.; Arimoto, R.; Rose, N.L.; Roberts, S.L.; Li, L.; Tang, Y.; Liu, X.; Fu, X.; Schneider, T.; Hou, X.; Lan, J.; Tan, L.; Liu, X.; Hu, J.; Cao, Y.; Liu, W.; Wu, F.; Wang, T.; Qiang, X.; Chen, N.; Cheng, P.; Hao, Y.; Wang, Q.; Chu, G.; Guo, M.; Han, M.; Tan, Z.; Wei, C.; Dusek, U. The Sihailongwan Maar Lake, northeastern China as a candidate Global Boundary Stratotype Section and point for the Anthropocene Series. The Anthropocene Review 2023, 10, 177–200. [Google Scholar] [CrossRef]
- Rioual, P.; Lu, Y.; Yang, H.; Scuderi, L.; Chu, G.; Holmes, J.; Zhu, B.; Yang, X. Diatom-environment relationships and a transfer function for conductivity in lakes of the Badain Jaran Desert, Inner Mongolia, China. Journal of Paleolimnology 2014, 50, 207–229. [Google Scholar] [CrossRef]
- Battarbee, R.W.; Jones, V.J.; Flower, R.J.; Cameron, N.G.; Bennion, H.; Carvalho, L.R.; Juggins, S. Diatoms. In Smol, J.P.; Birks, H.J.B., Ed.; Last, W.M. (Eds), Tracking environmental change using lake sediments, Volume 3: terrestrial, algal, and siliceous indicators. Kluwer Academic Publishers, Dordrecht, The Netherlands, 2001; pp. 155–202. [Google Scholar] [CrossRef]
- Round, F.E.; Crawford, R.M.; Mann, D.G. The Diatoms: Biology and Morphology of the Genera; Cambridge University Press: Cambridge, UK, 1990. [Google Scholar]
- Silva, P.C. Classification of algae. In Lewin, R.A. (ed.): Physiology and Biochemistry of Algae. Academic Press, New York & London, 1962; pp. 827–837.
- Greville, R.K. Div. IV. Diatomaceae. In: The English Flora of Sir James Edward Smith. Class XXIV. Cryptogamia. Vol. V. (or Vol. II of Dr. Hooker’s British flora). Part I. Comprising the Mosses, Hepaticae, Lichens, Characeae and Algae. (Hooker, W.J. Eds), London: Longman, Brown, Green & Longmans Paternoster-Row. 1833; pp. 401–415.
- Rioual, P.; Morales, E.A.; Chu, G.; Han, J.; Li, D.; Liu, J.; Liu, Q.; Mingram, J.; Ector, L. Staurosira longwanensis sp. nov., a new araphid diatom (Bacillariophyta) from Northeast China. Fottea 2014, 14, 91–100. [Google Scholar] [CrossRef]
- Dubois, A. Describing new species. Taprobanica 2010, 2, 6–24. [Google Scholar] [CrossRef]
- Williams, D.M. Spines and homologues in `araphid’ diatoms. Plant Ecology and Evolution 152, 150–162. [CrossRef]
- Graeff, C.L.; Kociolek, J.P.; Rushforth,S. R. New and interesting diatoms (Bacillariophyta) from Blue Lake Warm Springs, Tooele County, Utah. Phytotaxa 2013, 153, 1–38. [Google Scholar] [CrossRef]
- Al-Handal, A.Y.; Kociolek, J.P.; Abdullah, D.S. Williamsella iraqiensis sp. nov., a new diatom (Bacillariophyta, Fragilariophyceae) from Sawa lake, South Iraq. Phytotaxa 2016, 244, 289–297. [Google Scholar] [CrossRef]
- Lange-Bertalot, H.; Metzeltin, D. Indicators of oligotrophy. 800 taxa representative of three ecologically distinct lake types. Carbonate buffered - oligodystrophic - weakly buffered soft water. In: Lange-Bertalot, H. (ed.). Iconographia Diatomologica vol. 2. Koeltz Scientific Books, Königstein, Germany, 390 pp.
- Wengrat, S.; Morales, E.A.; Wetzel, C.E.; Almeida, P.D.; Ector, L.; Bicudo, D.C. Taxonomy and ecology of Fragilaria billingsii sp. nov. and analysis of type material of Synedra rumpens var. fusa (Fragilariaceae, Bacillariophyta) from Brazil. Phytotaxa 2016, 270, 191–202. [Google Scholar] [CrossRef]
- Alexson, E.E.; Reavie, E.D.; Van de Vijver, B.; Wetzel, C.E.; Ector, L.; Wellard Kelly, H.A.; Aliff, M.N.; Estepp, L.R. Revision of the needle-shaped Fragilaria species (Fragilariaceae, Bacillariophyta) in the Laurentian Great Lakes (United States of America, Canada). Journal of Great Lakes Research 2022, 48, 999–1020. [Google Scholar] [CrossRef]
- Lange-Bertalot, H.; Ulrich, S. Contributions to the taxonomy of needle-shaped Fragilaria and Ulnaria species. Lauterbornia 2014, 78, 1–73. [Google Scholar]
- Soumya, D.; Cai, J.; Hodoki, Y.; Goda, Y.; Akatsuka, T.; Nakano, S. Seasonal changes in the cell size and density of the diatom Fragilaria crotonensis Kitton in Lake Biwa. [CrossRef]
Figure 1.
(a) Map of north-east Asia showing the location of Lake Sihailongwan in Jilin Province (red asterisk), in NE China. (b) Map of Lake Sihailongwan, showing the morphology of its catchment and its bathymetry, the white asterisk shows the position of the sediment trap mooring in the middle of the lake (c) View of Lake Sihailongwan, taken from the southern shore of the lake (photo taken on 6th June 2025). The red line indicates the position of the mooring.
Figure 1.
(a) Map of north-east Asia showing the location of Lake Sihailongwan in Jilin Province (red asterisk), in NE China. (b) Map of Lake Sihailongwan, showing the morphology of its catchment and its bathymetry, the white asterisk shows the position of the sediment trap mooring in the middle of the lake (c) View of Lake Sihailongwan, taken from the southern shore of the lake (photo taken on 6th June 2025). The red line indicates the position of the mooring.
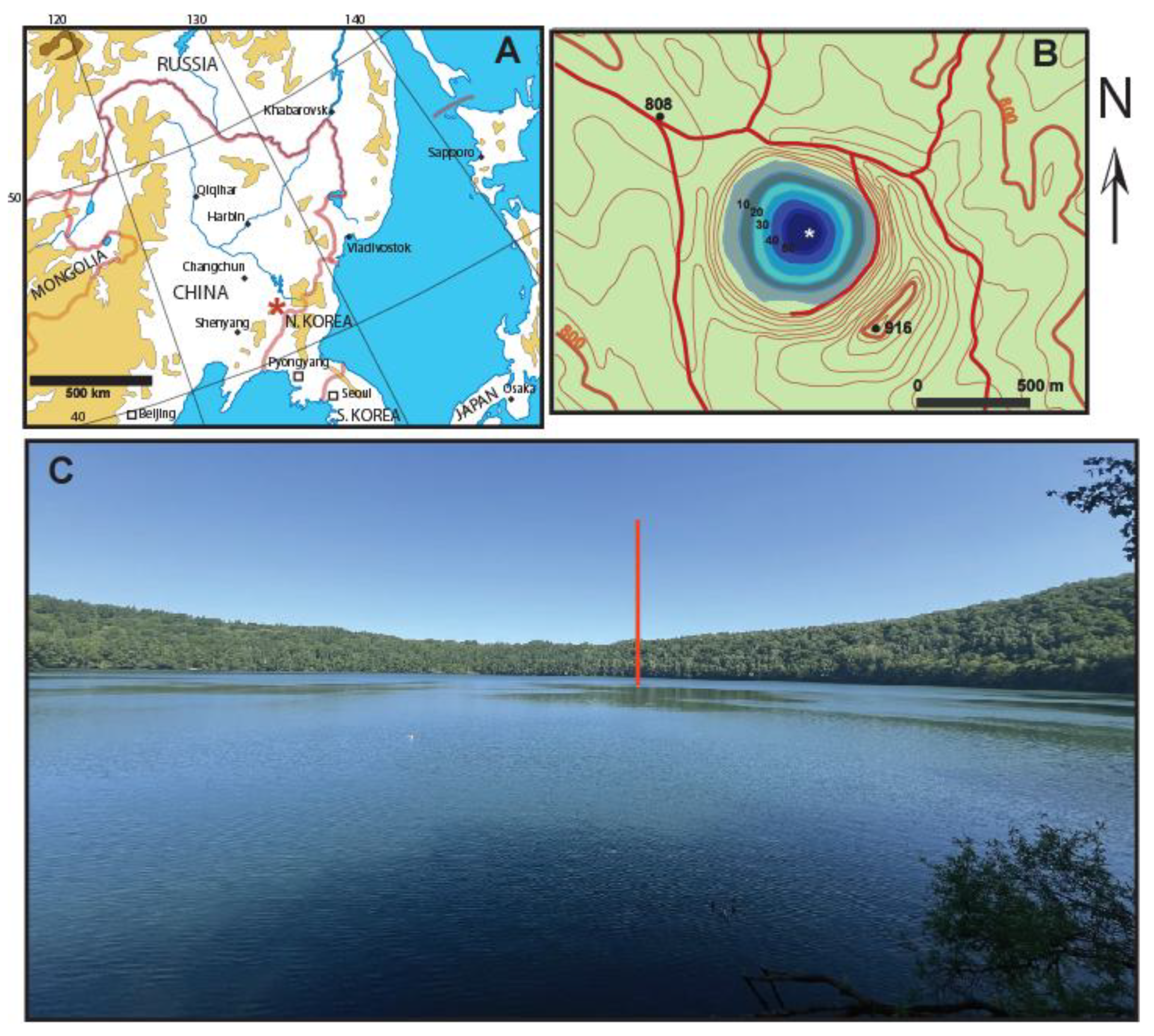
Figure 2.
(A ̶ AC) Fragilaria longwania sp. nov. from Lake Sihailongwan, northeast China. LM micrographs of valves and frustules showing the size diminution series. All material from sediment trap samples, in April 2005 for (A, C-E, I-J, N, Q-R, Z-AA), in May 2005 for (G-H, L-M, O-P), and in September 2006 for (B, K, S-Y, AB-AC). (D) Initial valve with a more linear outline and wider apices. (V) Two frustules in girdle view connected to each other. (AA) One solitary frustule in girdle view. Scale bar = 10 µm. Holotype specimen is Figure 2Z.
Figure 2.
(A ̶ AC) Fragilaria longwania sp. nov. from Lake Sihailongwan, northeast China. LM micrographs of valves and frustules showing the size diminution series. All material from sediment trap samples, in April 2005 for (A, C-E, I-J, N, Q-R, Z-AA), in May 2005 for (G-H, L-M, O-P), and in September 2006 for (B, K, S-Y, AB-AC). (D) Initial valve with a more linear outline and wider apices. (V) Two frustules in girdle view connected to each other. (AA) One solitary frustule in girdle view. Scale bar = 10 µm. Holotype specimen is Figure 2Z.
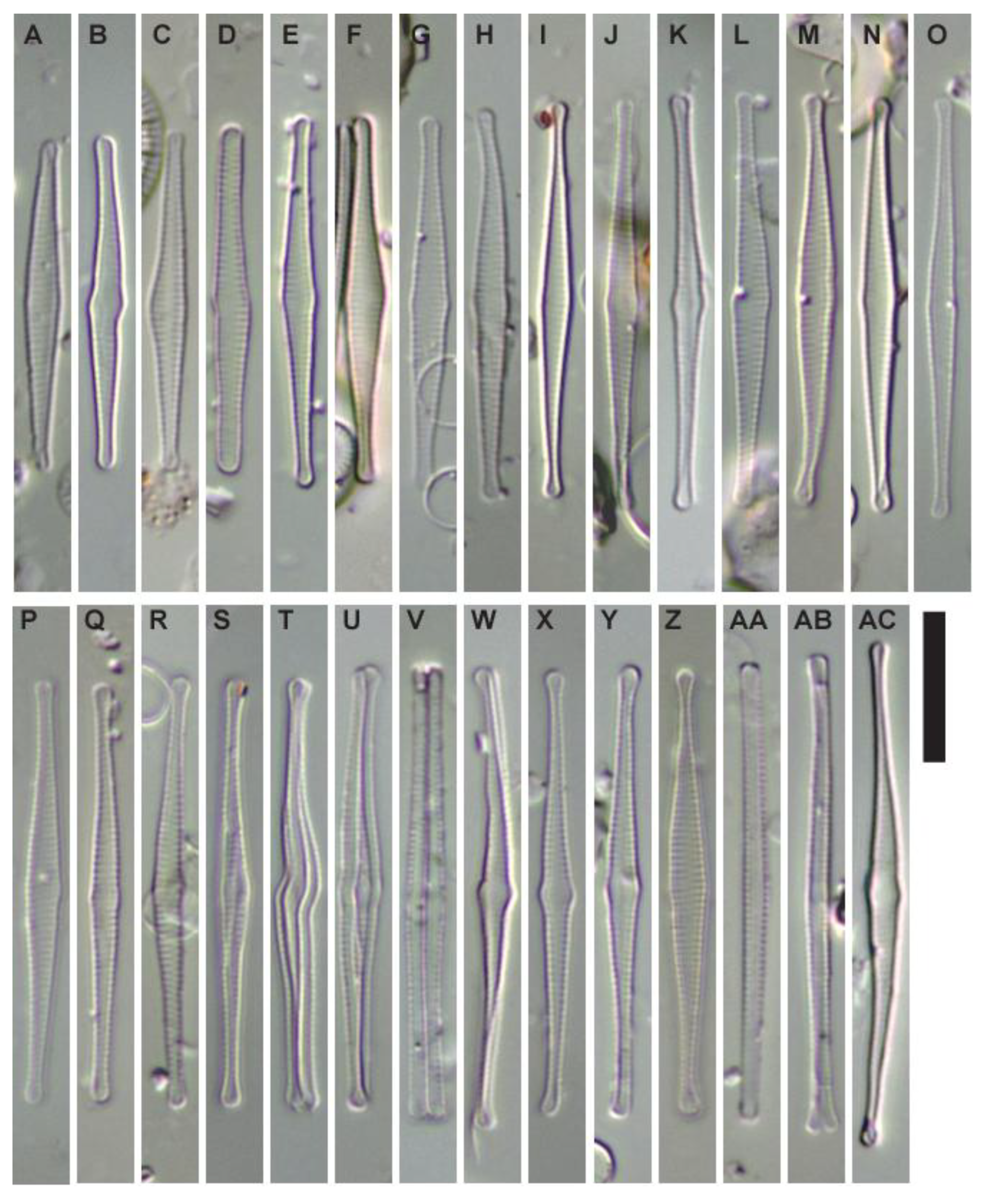
Figure 3.
(A-E) SEM micrographs of Fragilaria longwania sp. nov. from Lake Sihailongwan. (A-C) Material from an epipelon sample collected in November 2006 from 17m water depth (=paratype BM 101 673). (A) External view of a valve. (B) Same specimen as in A, close-up on the apex showing the external opening of a rimoportula (black arrow), the apical pore field (white arrow). (C) Marginal spines emerging from vimines (black arrows), more rarely from a virga (white arrow). (D-E) Material from an epipelon sample collected in April 2008 by 12 m water depth. (D) Internal view of the central area, note the ghost striae. (E) Same valve as in D, view of the apex without rimoportula, note the reduced apical pore field. Scale bars = 2 µm for (A); = 1 µm for (B-D).
Figure 3.
(A-E) SEM micrographs of Fragilaria longwania sp. nov. from Lake Sihailongwan. (A-C) Material from an epipelon sample collected in November 2006 from 17m water depth (=paratype BM 101 673). (A) External view of a valve. (B) Same specimen as in A, close-up on the apex showing the external opening of a rimoportula (black arrow), the apical pore field (white arrow). (C) Marginal spines emerging from vimines (black arrows), more rarely from a virga (white arrow). (D-E) Material from an epipelon sample collected in April 2008 by 12 m water depth. (D) Internal view of the central area, note the ghost striae. (E) Same valve as in D, view of the apex without rimoportula, note the reduced apical pore field. Scale bars = 2 µm for (A); = 1 µm for (B-D).

Figure 4.
(A-C) SEM micrographs of Fragilaria longwania sp. nov. from Lake Sihailongwan, sediment trap sample collected in July 2009. (A) Whole view of two valves from the same frustule. (B) Detail from A showing: on the right, the external view of an apex, with opening of the rimoportula (black arrow) and three short spines located above the apical pore field, on the left, the internal view of the apex of the other valve without rimoportula. (C) Detail from A showing the internal opening of a rimoportula (black arrow) and the apical pore field. The girdle bands on the left and the right of the valve are ornamented by one row of poroids. Scale bars = 10 µm for (A); = 2 µm for (B-C).
Figure 4.
(A-C) SEM micrographs of Fragilaria longwania sp. nov. from Lake Sihailongwan, sediment trap sample collected in July 2009. (A) Whole view of two valves from the same frustule. (B) Detail from A showing: on the right, the external view of an apex, with opening of the rimoportula (black arrow) and three short spines located above the apical pore field, on the left, the internal view of the apex of the other valve without rimoportula. (C) Detail from A showing the internal opening of a rimoportula (black arrow) and the apical pore field. The girdle bands on the left and the right of the valve are ornamented by one row of poroids. Scale bars = 10 µm for (A); = 2 µm for (B-C).
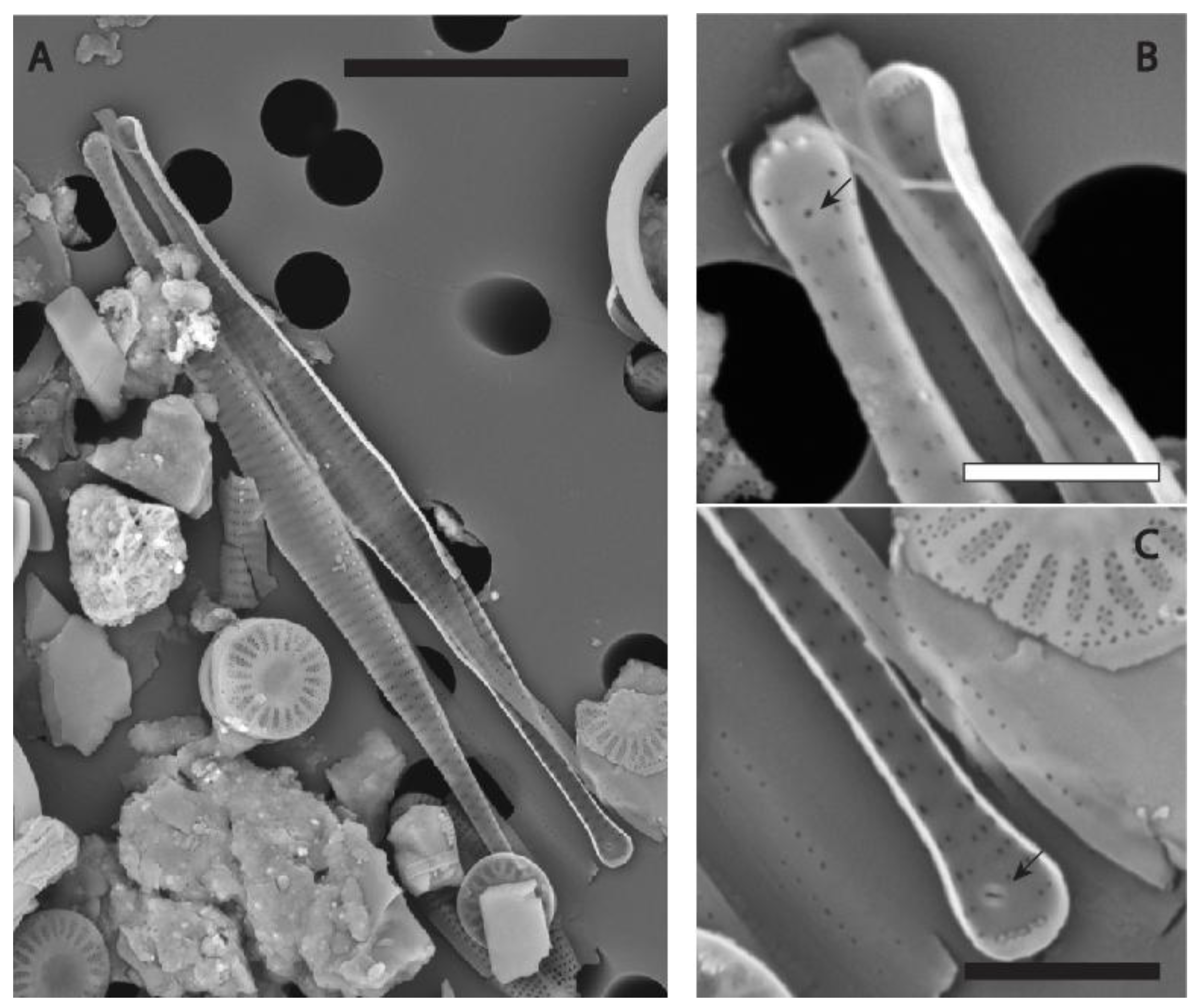
Figure 5.
(A-D) SEM micrographs of Fragilaria longwania sp. nov. from Lake Sihailongwan, sediment trap sample collected in August 2012. (A) External view of a whole valve. (B) Detail from A showing the apex with the opening of the rimoportula (black arrow) and three short spines located above the apical pore field. (C) Detail from A showing an asymmetrical central area, barely swollen, uniseriate striae composed of up to six small areolae. Marginal spines irregularly spaced, emerging from either the vimines or the virgae. (D) Detail from A showing the apex without rimoportula and the apical pore field composed of two rows of tiny poroids (black arrow). Scale bars = 8 µm for (A); = 3 µm for (B-D).
Figure 5.
(A-D) SEM micrographs of Fragilaria longwania sp. nov. from Lake Sihailongwan, sediment trap sample collected in August 2012. (A) External view of a whole valve. (B) Detail from A showing the apex with the opening of the rimoportula (black arrow) and three short spines located above the apical pore field. (C) Detail from A showing an asymmetrical central area, barely swollen, uniseriate striae composed of up to six small areolae. Marginal spines irregularly spaced, emerging from either the vimines or the virgae. (D) Detail from A showing the apex without rimoportula and the apical pore field composed of two rows of tiny poroids (black arrow). Scale bars = 8 µm for (A); = 3 µm for (B-D).
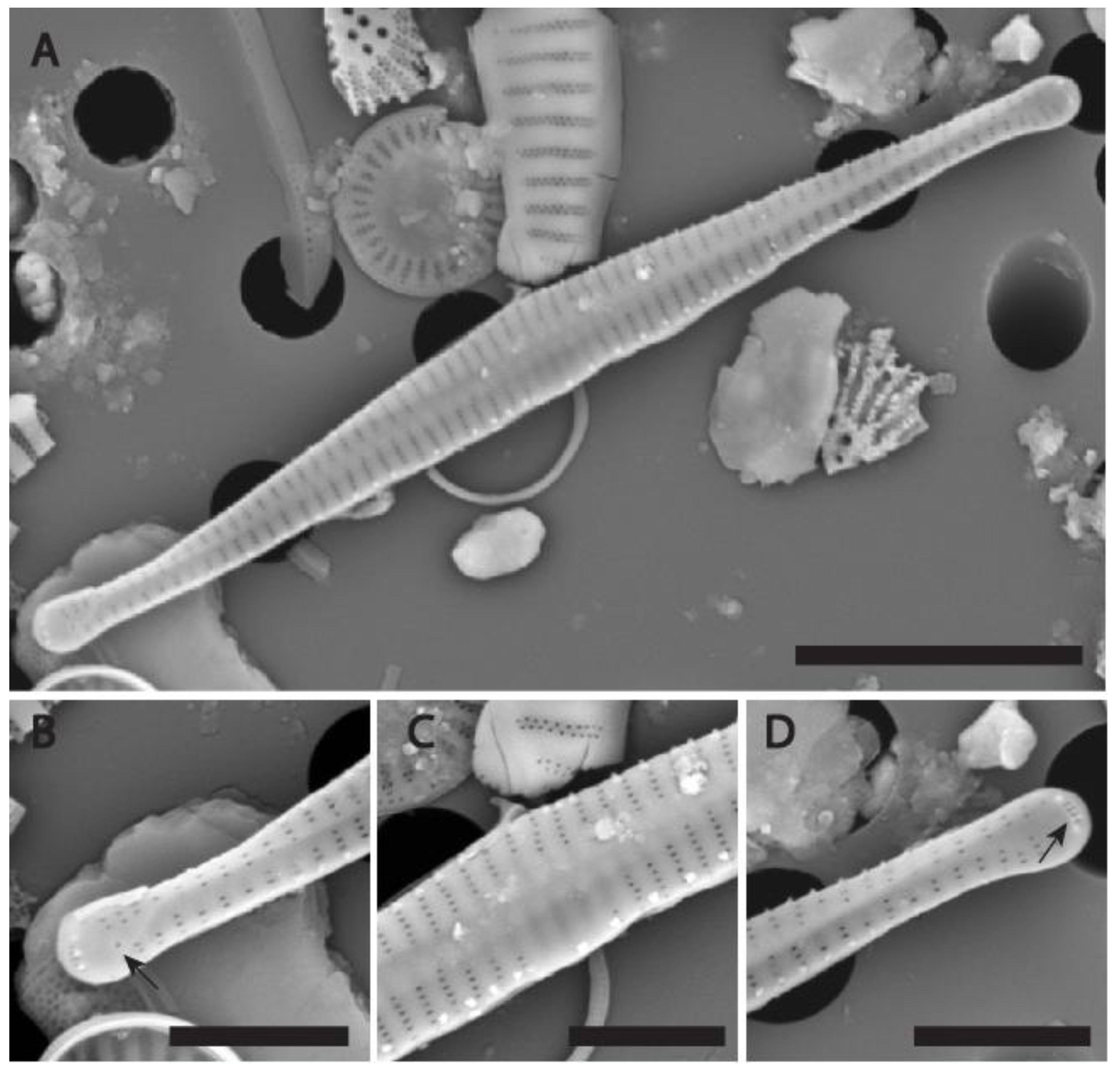
Figure 6.
(A-D) SEM micrographs of Fragilaria longwania sp. nov. from Lake Sihailongwan, sediment trap sample collected in August 2012. (A) External view of a whole frustule, twisted along the apical axis. (B) Detail from A showing the apex with the opening of the rimoportula (black arrow) and two spines located above the apical pore field. (C) Detail from A showing a clearly swollen, symmetrical central area with ghost striae on both sides of the sternum. Marginal spines mostly emerging from the vimines. (D) Detail from A showing an oblique view of the apex without rimoportula. Scale bars = 10 µm for (A); = 3 µm for (B-C); = 2 µm for (D).
Figure 6.
(A-D) SEM micrographs of Fragilaria longwania sp. nov. from Lake Sihailongwan, sediment trap sample collected in August 2012. (A) External view of a whole frustule, twisted along the apical axis. (B) Detail from A showing the apex with the opening of the rimoportula (black arrow) and two spines located above the apical pore field. (C) Detail from A showing a clearly swollen, symmetrical central area with ghost striae on both sides of the sternum. Marginal spines mostly emerging from the vimines. (D) Detail from A showing an oblique view of the apex without rimoportula. Scale bars = 10 µm for (A); = 3 µm for (B-C); = 2 µm for (D).
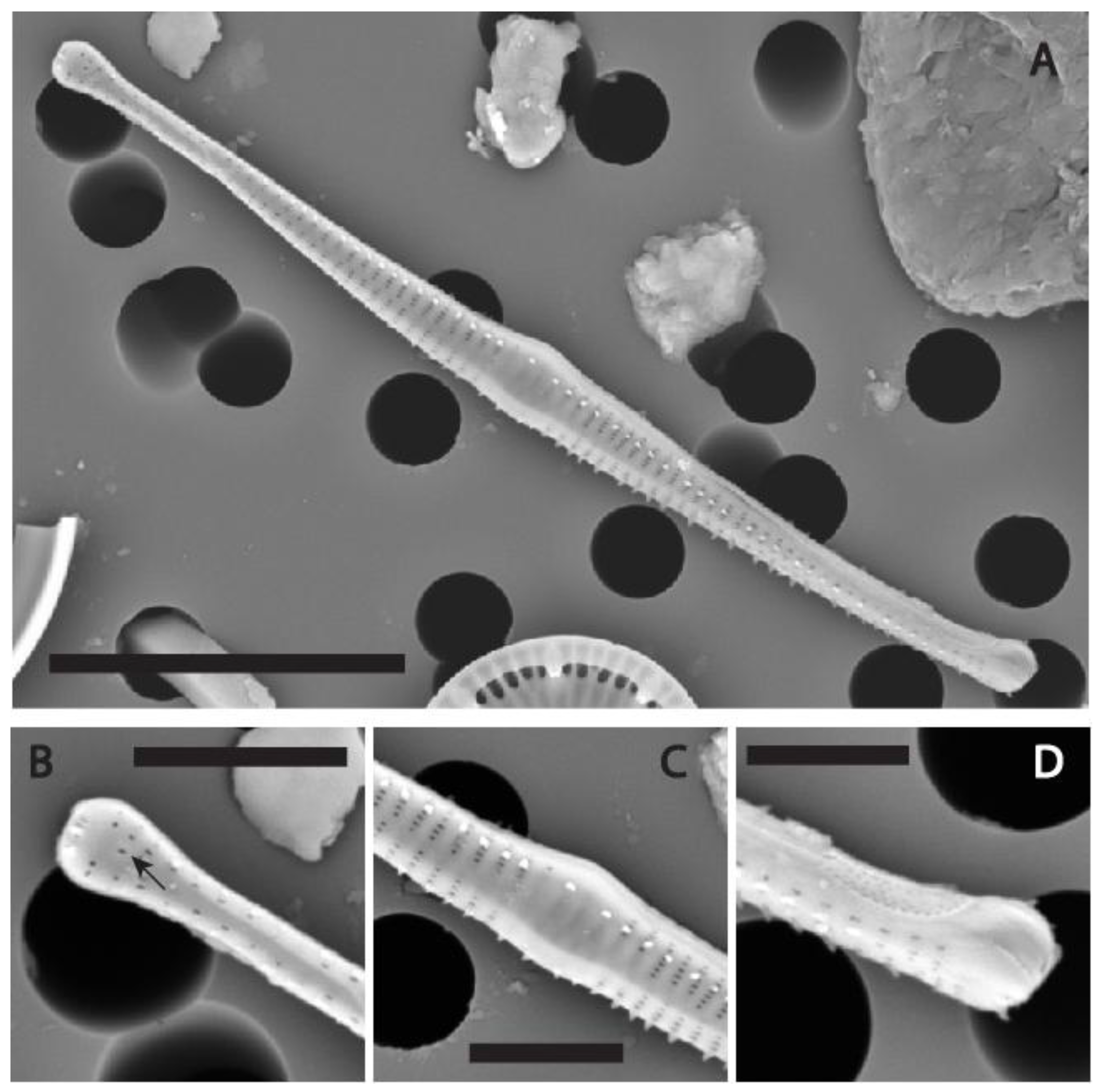
Figure 7.
(A-D) SEM micrographs of Fragilaria longwania sp. nov. from Lake Sihailongwan, sediment trap sample collected in June 2016. (A) External view of a whole valve showing the typical spindle-shaped outline, with protracted, distinctly capitate apices. (B) Detail from A showing a barely swollen, asymmetrical central area and irregularly spaced marginal spines emerging from either the vimines or the virgae. (C) Detail from A showing the apex without a rimoportula (D) Detail from A showing the apex with the opening of the rimoportula (black arrow). Scale bars = 5 µm for (A); = 2 µm for (B-D).
Figure 7.
(A-D) SEM micrographs of Fragilaria longwania sp. nov. from Lake Sihailongwan, sediment trap sample collected in June 2016. (A) External view of a whole valve showing the typical spindle-shaped outline, with protracted, distinctly capitate apices. (B) Detail from A showing a barely swollen, asymmetrical central area and irregularly spaced marginal spines emerging from either the vimines or the virgae. (C) Detail from A showing the apex without a rimoportula (D) Detail from A showing the apex with the opening of the rimoportula (black arrow). Scale bars = 5 µm for (A); = 2 µm for (B-D).
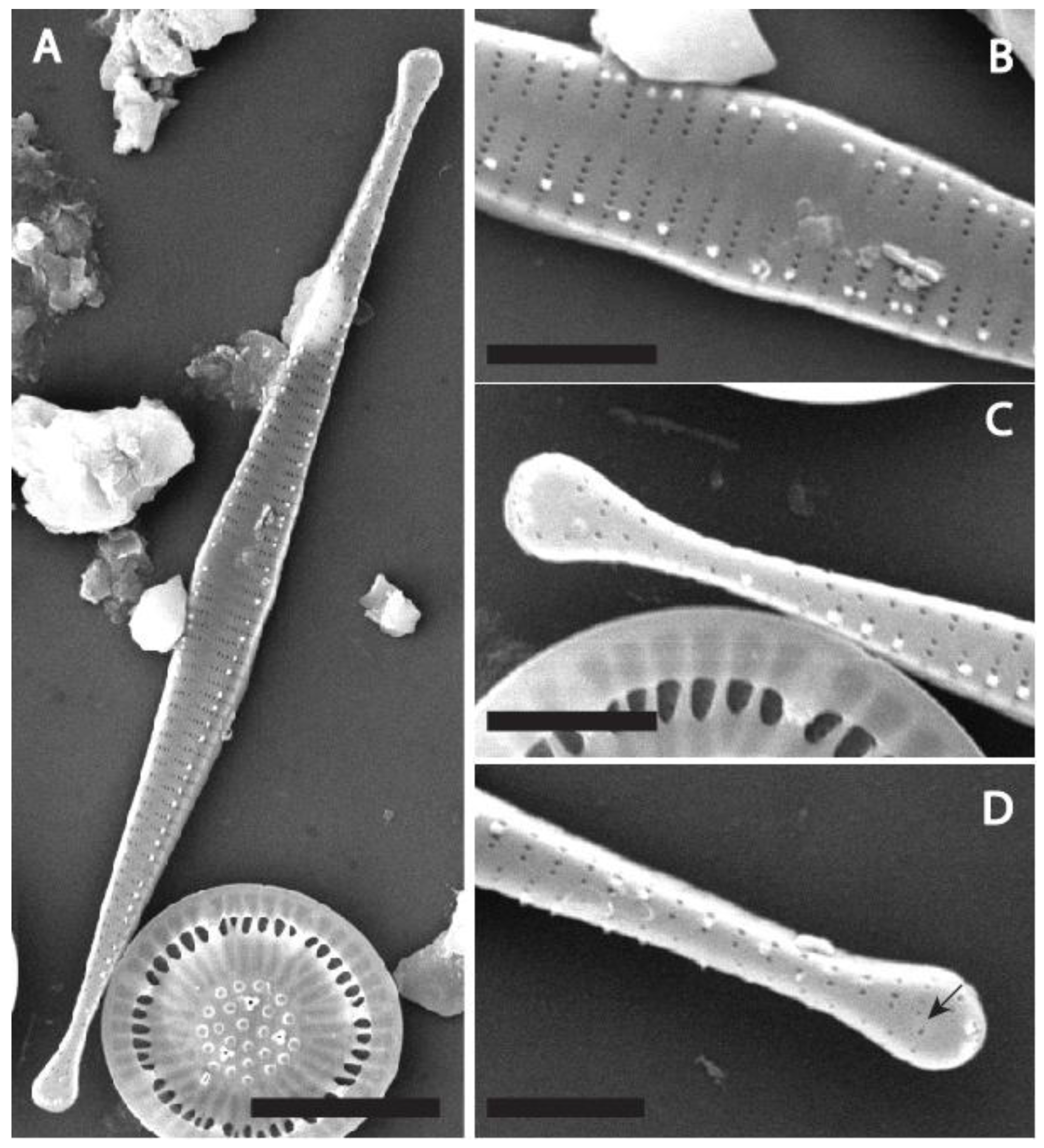
Figure 8.
(A-C) SEM micrographs of Fragilaria longwania sp. nov. from Lake Sihailongwan, sediment trap sample collected in August 2012 (A, B) and July 2017 (C). (A) Two frustules in girdle view, note the shark tooth shape of the spines. (B) Apex of a dividing cell in girdle view showing both epivalve and hypovalve with associated girdle bands (Labeled B1 to B3). (C) Apex of a frustule showing two ornamented girdle bands (B1 and B2) and very small apical pore fields.
Figure 8.
(A-C) SEM micrographs of Fragilaria longwania sp. nov. from Lake Sihailongwan, sediment trap sample collected in August 2012 (A, B) and July 2017 (C). (A) Two frustules in girdle view, note the shark tooth shape of the spines. (B) Apex of a dividing cell in girdle view showing both epivalve and hypovalve with associated girdle bands (Labeled B1 to B3). (C) Apex of a frustule showing two ornamented girdle bands (B1 and B2) and very small apical pore fields.

Figure 9.
(A) Relative percentages of Fragilaria longwania in sediment trap samples collected in Lake Sihailongwan between 2003 and 2017. The fitted linear regression (dashed line) shows a significant declining trend. (B). Valve length of Fragilaria longwania (in micrometer). The blue triangles symbolize the mean values, while the bars show the full range (minimum/maximum values). The linear regression (dashed line) indicates a significant increase in mean valve length.
Figure 9.
(A) Relative percentages of Fragilaria longwania in sediment trap samples collected in Lake Sihailongwan between 2003 and 2017. The fitted linear regression (dashed line) shows a significant declining trend. (B). Valve length of Fragilaria longwania (in micrometer). The blue triangles symbolize the mean values, while the bars show the full range (minimum/maximum values). The linear regression (dashed line) indicates a significant increase in mean valve length.
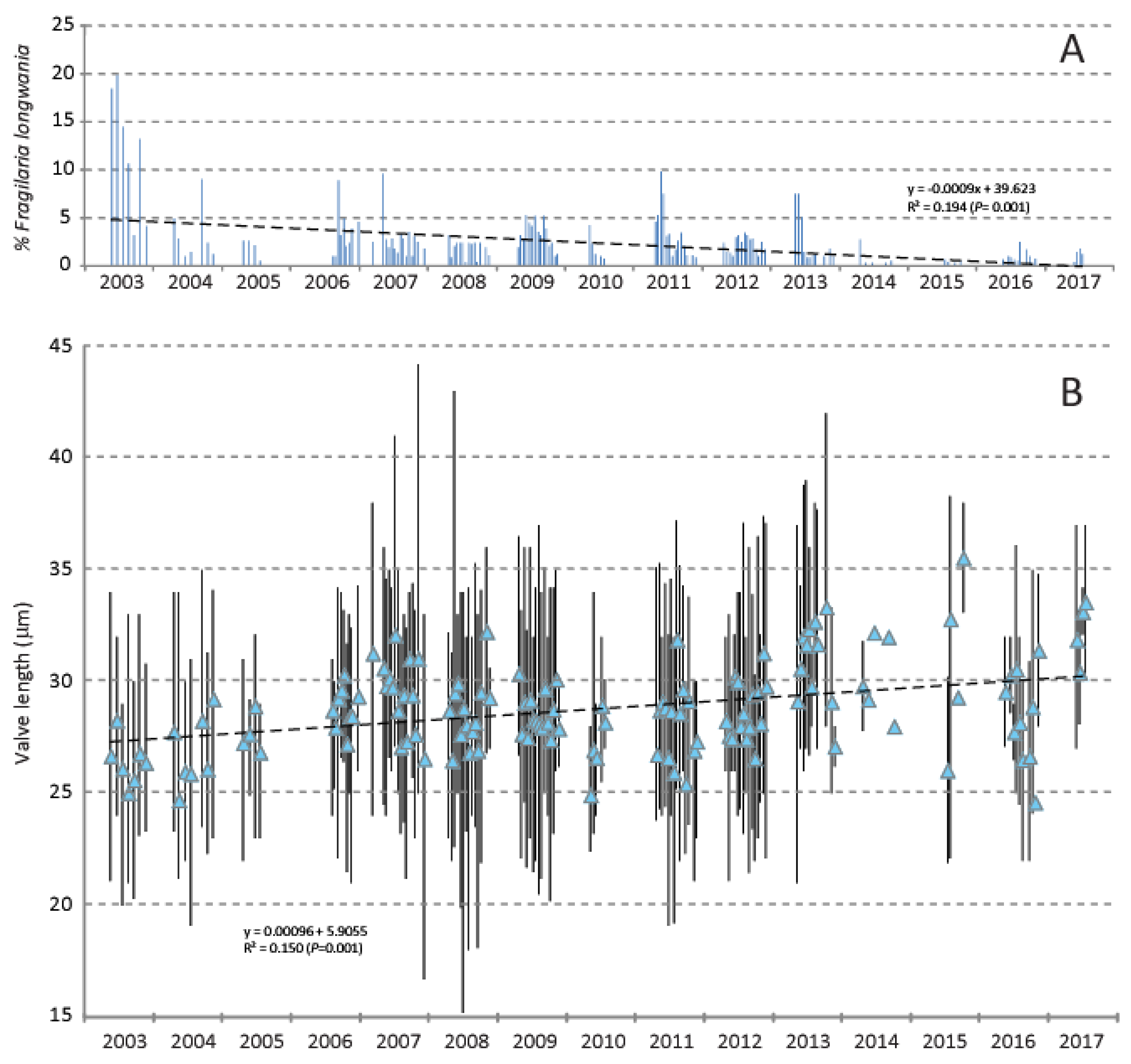
Figure 10.
Fragilaria longwania in sediment trap samples collected in Lake Sihailongwan in the 15 years of the study, from 2003 and 2017. The diatom data that include the relative percentages and mean valve length values of F. longwania are plotted year by year against the Julian day to better show the seasonal distribution in association with the meteorological data from Jingyu weather station (located about 20 km to the northeast of Lake Sihailongwan). The meteorological data correspond to the calculated daily mean values for the trap periods.
Figure 10.
Fragilaria longwania in sediment trap samples collected in Lake Sihailongwan in the 15 years of the study, from 2003 and 2017. The diatom data that include the relative percentages and mean valve length values of F. longwania are plotted year by year against the Julian day to better show the seasonal distribution in association with the meteorological data from Jingyu weather station (located about 20 km to the northeast of Lake Sihailongwan). The meteorological data correspond to the calculated daily mean values for the trap periods.
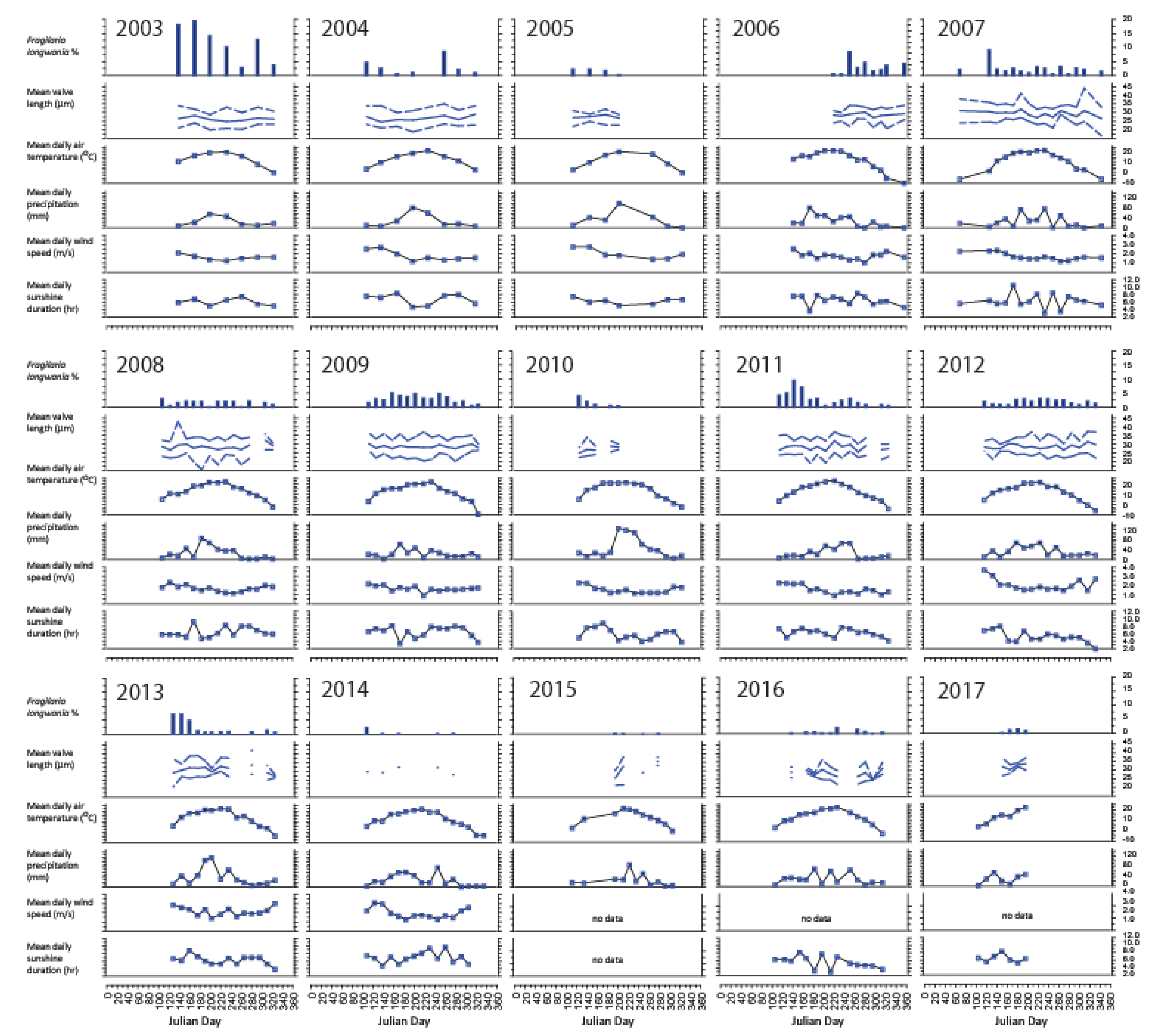
Table 1.
Summary of the chemical and limnological measurements made on water samples collected from Lake Sihailongwan between August 2010 and November 2014.
Table 1.
Summary of the chemical and limnological measurements made on water samples collected from Lake Sihailongwan between August 2010 and November 2014.
| Variable | unit | nb | Min. | Max. | Median | St. dev. |
| pH | 99 | 5.53 | 7.21 | 6.55 | 0.24 | |
| Elec. Conductivity | µS/L | 99 | 51 | 105 | 62 | 7 |
| Alkalinity | µeq/L | 99 | 338 | 660 | 478 | 52 |
| Tot. Phosphorus | µg/L | 99 | 5.0 | 88.6 | 14.9 | 14.4 |
| Tot. Nitrogen | µg/L | 97 | 7 | 2443 | 360 | 294 |
| Diss. Organic Carbon | mg/L | 92 | 0.2 | 10.0 | 1.7 | 1.2 |
| Cl | mg/L | 78 | 0.6 | 3.8 | 1.2 | 0.6 |
| NO3 | mg/L | 73 | 0 | 2.4 | 0.6 | 0.5 |
| SO4 | mg/L | 78 | 3.4 | 7.8 | 5.7 | 1.0 |
| Ca | mg/L | 78 | 2.7 | 28.8 | 5.3 | 5.3 |
| K | mg/L | 78 | 0.6 | 3.3 | 1.3 | 0.6 |
| Mg | mg/L | 78 | 1.2 | 3.6 | 2.3 | 0.6 |
| Na | mg/L | 78 | 1.2 | 4.4 | 2.2 | 0.7 |
| Si | mg/L | 99 | 0.1 | 0.8 | 0.4 | 0.2 |
| Secchi Disk depth | m | 22 | 2.4 | 7.6 | 3.8 | 1.3 |
Table 2.
Summary results of the diatom analysis of the samples collected for this study.
| Date | Sample | Dominant species | % Fragilaria longwania |
| From 19-May-03 to 15-Jul-17 | Sediment trap samples (centre of the lake, -45m water depth), (n =187) | Discostella stelligeroides (43.3, 0.4-92.0%), Lindavia balatonis (26.2, 1.7-76.3%)1 | 2.1 (0—19.8%)1 |
| 26-Apr-05 | Epilithon (small stones, near shore, <0.5m depth) | Lindavia balatonis (19.3%), Achnanthidium sieminskae (16.0%), Encyonopsis perborealis (7.6%), Discostella stelligeroides (5.9%), Encyonema silesiacum (5.0%) | Not observed |
| 21-Oct-05 | Surface-sediment (-50m water depth, lake centre) | Discostella stelligeroides (57.2%), Lindavia balatonis (17.6%), Pseudostaurosira parasitoides (3.2%), Nanofrustulum trainorii (2.2%), Fragilaria pectinalis (1.9%) | 1.8% |
| 18-Nov-06 | Periphyton on floating buoy (lake centre) | Achnanthidium sieminskae (50.6%), Brachysira neoexilis (15.2%), Encyonopsis subminuta (7.2%), Staurosirella ovata (5.9%), Discostella stelligeroides (4.6%) | 3.0% |
| 19-Nov-06 | Epilithon (small stones, near shore, <0.5m depth) | Nitzschia cf sociabilis (12.6%), Achnanthidium sieminskae (9.6%), Lindavia balatonis (9.1%), Nitzschia archibaldii (7.8%), Tabellaria hercynica (7.4%) | 2.6% |
| 19-Nov-06 | Epiphyton (-10m water depth, filamentous green) | Discostella stelligeroides (18.7%), Lindavia balatonis (9.6%), Pseudostaurosira pseudoconstruens (8.7%), Staurosirella ovata (6.5%), Sellaphora chistiakovae (3.0%) | 1.3% |
| 19-Nov-06 | Epipelon (-17m water depth) | Discostella stelligeroides (32.0%), Staurosirella ovata (15.4%), Lindavia balatonis (10.6%), Pseudostaurosira parasitoides (5.9%), Staurosira longwanensis (5.9%) | 1.2% |
| 14-Aug-07 | Surface-sediment (-51m water depth, lake centre) | Discostella stelligeroides (80.4%), Lindavia balatonis (6.3%), Fragilaria longwania (1.2%) | 1.2% |
| 22-Apr-08 | Periphyton on floating buoy (lake centre) | Achnanthidium sieminskae (38.5%), Discostella stelligeroides (16.3%) Brachysira neoexilis (14.4%), Encyonopsis perborealis (4.4%), Lindavia balatonis (3.0%), | 0.4% |
| 22-Apr-08 | Epilithon (small stones, near shore, <0.5m depth) | Encyonopsis perborealis (23.6%), Encyonema silesiacum (14.6%), Discostella stelligeroides (12.2%), Lindavia balatonis (10.6%), Achnanthidium sieminskae (8.9) | Present (<0.1%) |
| 3-Aug-16 | Periphyton on floating buoy (lake centre) | Achnanthidium sieminskae (27.5%), Encyonopsis perborealis (19.0%), Lindavia balatonis (14.2%), Eucocconeis laevis (13.3%), Rossithidium pusillum (7.3%) | Not observed |
| 3-Aug-16 | Epilithon (small stones, near shore, <0.5m depth) | Encyonopsis perborealis (29.3%), Achnanthidium sieminskae (9.30%), Lindavia balatonis (8.7%), Achnanthidium sp cf duriense (8.0%), Nitzschia lacuum (6.0%) | 1.3% |
| 3-Aug-16 | Epixylon (dead tree, near shore, <0.5m depth) | Encyonopsis perborealis (32.8%), Achnanthidium sieminskae (18.0%), Lindavia balatonis (14.8%), Nitzschia gessneri (6.9%), Brachysira neoexilis (4.8%) | Not observed |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



