Submitted:
04 August 2025
Posted:
05 August 2025
You are already at the latest version
Abstract
Tartrazine (TZ, also known as FD&C Yellow No. 5 or E102) is a synthetic, water-soluble yellow food dye widely used in the food and pharmaceutical industries. Some studies have associated TZ with allergic reactions, especially among people with dye sensitivities or pre-existing allergies. Recent research also suggests a possible link between TZ consumption and the worsening of behavioral disorders, especially in children, including symptoms such as hyperactivity, irritability, restlessness, and sleep disturbances. Experimental studies in laboratory animals have highlighted potential risks associated with prolonged or high-dose exposure, including toxic effects on the nervous system and liver function. In addition, increasing evidence indicates that TZ can induce oxidative stress (OS) by increasing the production of reactive oxygen species (ROS), which can contribute to cellular damage and inflammation. Although the evidence remains inconclusive, there are recommendations to limit the intake of syn-thetic food dyes especially in children’s diets. The purpose of this review is to examine the potential toxic effects on health of tartrazine by analyzing findings from experimental studies in cell cultures and laboratory animals, as well as correlations observed in humans. We focus on documented adverse reactions, including possible neurotoxic, hepatotoxic, oxidative, and behavioral effects. Through this, we aim to contribute to a more comprehensive understanding of the risks associated with exposure to this syn-thetic food dye.
Keywords:
tartrazine
; health
; zebrafish
; toxicity
; hyperactivity
; oxidative stress
1. Introduction
Tartrazine is an azo dye commonly used to impart color to food products [1,2,3,4,5]. Approximately 65% of azo dyes are used as food additives and are found in various products such as candies, jams, and soft drinks, as well as in pharmaceuticals, cosmetics, and textiles. Although TZ adds no nutritional benefit and has no nutritional value, it remains the most widely used food dye, with a significant presence in the global food industry [6,7,8,9]. According to the World Health Organization (WHO), since 2016, the acceptable daily intake (ADI) has been set at 0–10 mg/kg body weight [10]. Although it was one of the first food additives to raise concerns regarding the possibility of causing adverse reactions, it is still used in the EU and other parts of the world [11].
Moreover, some research has suggested a possible association between TZ exposure and allergic reactions such as urticaria, asthma, migraines, blurred vision, itching, or sleep and behavioral disorders, including hyperactivity in children [12,13,14,15,16,17]. In particular, children appear to be more sensitive to this additive, with possible links reported between food dyes and behavioral disorders such as attention deficit hyperactivity (ADHD), and even mutagenic or carcinogenic pathologies [18,19]. In a double-blind placebo-controlled study, a group of children was exposed to several doses of TZ. Some of them exhibited clear behavioral reactions such as irritability, restlessness, and sleep disturbances [20].
In recent decades, numerous experimental studies, both in vitro and on animal models, have investigated the effects of TZ on cellular functions, tissue integrity, and behavior. The results suggest this food additive may be involved in inducing OS through the generation of ROS [21,22], activating the inflammatory response through the overproduction of proinflammatory cytokines interleukin-1 (IL-1) and interleukin-6 (IL-6) [23,24], modifying enzymatic activity, and contributing to the onset of neurobehavioral disorders [25], including impairments in learning and memory processes [26,27]. From the perspective of the biological mechanisms involved, OS has been identified as a pathway through which TZ exerts its toxic effects, contributing to the imbalance between pro-oxidants and antioxidants in cells [28,29]. This imbalance has been reflected in elevated levels of malondialdehyde (MDA), a byproduct of lipid peroxidation, indicating enhanced oxidative damage. Concurrently, a significant reduction in the activity of endogenous antioxidant enzymes such as superoxide dismutase (SOD) and catalase (CAT) has been observed, further supporting the pro-oxidant shift induced by TZ exposure [30].
However, the data available in scientific literature are heterogeneous, and translating results from preclinical experiments into the context of human exposure remains challenging. At the same time, the food industry is increasingly exploring natural alternatives to mitigate the health side effects of synthetic dyes. Therefore, replacing them with natural alternatives represents a much better option [19]. Some well-known natural alternatives include turmeric, carotenoids, annatto, saffron, and paprika extracts. For example, the azo dye TZ can potentially be replaced with carotenoids extracted from pequi (Caryocar brasiliense) using high-performance ionic liquids as extractants, a modern and sustainable method that offers both safety and efficiency benefits [31,32].
The aim of this review is to provide a critical and integrative analysis of experimental data from in vitro and animal model studies regarding the effects of TZ exposure, with a focus on the toxicological mechanisms involved (including OS, neurotoxicity, and inflammatory response). By correlating these findings with existing observations of side effects in humans, the paper aims to offer a comprehensive perspective on the potential risks to human health. Thus, the study contributes to clarifying a current issue regarding the safety of synthetic food additives and supports the need to reassess TZ exposure, particularly among children.
2. Health Implications of Tartrazine Exposure
TZ is a synthetic yellow dye widely used in food, pharmaceutical, and non-food products (Table 1). Although it enhances the appearance of products, several studies have reported its potential to trigger allergic reactions, asthma, or skin hypersensitivity. These adverse effects have led to growing interest in natural alternatives such as annatto or beta-carotene [2,25,33,34,35,36,37,38,39,40,41,42,43,44,45].
From a chemical perspective, TZ is an anionic acid dye that, in an acidic environment, is converted into tartrazic acid [46]. In the body, TZ is reduced to an aromatic amine, a structure that is strongly stabilized due to its nitroso derivative [47]. This compound can trigger a wide range of immunological reactions, including fatigue, irritability, clinical depression, headaches, and sleep disturbances. Figure 1 illustrates the main pathways through which TZ may affect human health: OS, chronic inflammation, genotoxicity, and neurobehavioral disorders. These mechanisms are especially triggered under repeated or chronic exposure and may contribute to the development of systemic conditions, particularly in children and sensitive individuals. Both ingestion and simple skin contact with products containing this substance may cause hypersensitivity reactions. Some researchers suggest that symptoms may occur even at low exposure levels, with manifestations appearing up to 72 hours after contact [45].
Although there are no conclusive studies proving that TZ directly causes anxiety in humans, there is sufficient evidence from animal studies indicating that this additive may increase OS, negatively affect liver and kidney function, alter lipid levels, and consequently raise the risk of cardiovascular disease. The results regarding its impact on behavior and neurotoxicity are often contradictory or inconclusive.
TZ has been associated with various adverse reactions in some individuals, including migraines, agitation, asthma attacks, blurred vision, eczema, and other skin rashes. Additionally, some studies have suggested a possible increased risk of thyroid cancer [47,48]. In a randomized, double-blind, placebo-controlled study, the administration of a mixture of food colorants (5 mg E110, 2.5 mg E122, 7.5 mg E102, 5 mg E124) along with 45 mg E211 was linked to negative effects on concentration, hyperactive behavior, and attention in children aged 3 and 8–9 years [49]. Another study reported that TZ was responsible for approximately 1% of acute urticaria and/or angioedema cases [50].
The effects of TZ on the behavior of children with suspected hyperactivity included irritability, restlessness, and sleep disturbances, and these reactions intensified with increasing dosage, indicating a dose-response relationship [20]. The administration of TZ in atopic patients triggered allergic reactions, predominantly cutaneous and respiratory, which were significantly more frequent compared to placebo. These included angioedema, nasal congestion, rhinorrhea, wheezing, skin rashes, and pruritus, and approximately 5% of participants showed elevated levels of TZ-specific IgE [51].
Data from the literature on the cytotoxic, mutagenic, and genotoxic effects of TZ remain unclear and often contradictory. Although Soares et al. [52] did not identify significant cytotoxic effects, the authors highlighted the potential health risk posed by TZ, suggesting that prolonged use may contribute to carcinogenic process.
3. Oxidative Stress Induced by Tartrazine
Several studies on rats have correlated the action of TZ with the induction of OS [21,48,53,54,55,56] (Table 2). OS represents an imbalance between the production of ROS and the capacity of endogenous antioxidant systems to neutralize them, leading to damage to cellular components such as lipids, proteins, and DNA [57,58].
Exposure to TZ, even at concentrations considered environmentally relevant (50 mg/L), has been shown to cause significant biochemical changes, such as increased levels of MDA and nitric oxide (NO), along with a decrease in antioxidant enzyme activities (SOD, CAT, glutathione (GSH)) and acetylcholinesterase (AChE), suggesting neurobiochemical dysfunctions [66]. Daily administration of higher doses, such as 320 mg/kg for four weeks, exacerbates oxidative imbalance in the brain by reducing glutathione peroxidase (GPx) activity and increasing lipid peroxidation, reflected in elevated MDA levels [59]. Even significantly lower doses (7.5 mg/kg), administered over a longer period (90 days), induce severe OS, associated with increased MDA and reduced activities of endogenous antioxidant enzymes (CAT, SOD, GPx) and GSH levels [30].
Moreover, the combination of TZ and erythrosine has demonstrated synergistic effects in inducing OS in brain structures such as the striatum, where increases in MDA, AChE, and nitrites, along with reductions in GSH and CAT, suggest disruption of cerebral redox balance and a potential role in neurotoxicity [60].
Repeated exposure to TZ at doses ranging from 10–75 mg/kg over periods of up to 8 weeks has been associated with marked alterations in oxidative balance in key organs such as the liver, kidneys, and pancreas. This occurs through decreased levels of endogenous antioxidants (GSH, SOD, CAT, glutathione reductase (GR)) and increased MDA, reflecting a significant contribution to metabolic and endocrine dysfunctions [1,61,62].
Additional studies have shown that high doses (200–500 mg/kg) exacerbate systemic OS, affecting the liver, kidneys, spleen, and blood. Marked increases in MDA [29,55,56,63,64,65] were observed, along with decreases in GSH, CAT, and SOD [29,56,63,65], TOS, and TAS [65], as well as alterations in histopathological parameters [29,55,56,63,65]. However, Golli et al. [55] reported increases in CAT and GST levels. These imbalances are correlated with marked histopathological changes in the liver, kidneys, and intestines, supporting the idea of systemic toxicity via oxidative mechanisms.
Also, the studies by Amin et al. [54] and Gao et al. [48] highlight the impact of doses ranging from 15 to 700 mg/kg on the liver and brain, through the inhibition of GSH, SOD, CAT, and the increase of lipid peroxidation [48,54], reflecting the impairment of cognitive functions and potential neurotoxicity. These effects are attributed to the formation of free radicals during the metabolism of TZ, which promotes the activation of inflammatory pathways and increased expression of IL-1 and IL-6, reinforcing the role of chronic inflammation in the toxicity of this substance [48].
4. In Vitro Studies on Tartrazine Toxicity
In vitro studies provide a controlled experimental framework to evaluate the direct effects of TZ on cells, thus allowing the identification of the molecular mechanisms of toxicity. Several in vitro studies have assessed the cytotoxic and genotoxic effects of TZ in different types of human cells and experimental models (Table 3). In human lymphocytes exposed to concentrations ranging from 0.25 to 64.0 mM, TZ did not show cytotoxicity, but induced significant genotoxic effects at all tested concentrations, with only partial DNA repair observed [52]. Similarly, human leukocytes exposed to TZ at 5–500 μg/mL showed no cytotoxic or mutagenic effects; however, DNA damage was observed at concentrations ≥70 μg/mL, consistent with in silico toxicity predictions [67].
An in vitro study on human foreskin fibroblasts evaluated the effects of several dyes, including TZ. Testing revealed that TZ did not induce significant cytotoxic effects, even at high concentrations. In contrast, indigo carmine and chlorophyllin exhibited marked cytotoxicity at elevated doses. Moreover, chlorophyllin also triggered increased ROS production, indicating potential OS. These findings underscore the importance of further research into the safety of dyes commonly used in pharmaceutical and cosmetic formulations [68].
Investigations into the estrogenic, cytotoxic, and genotoxic potential of TZ compared to phytoestrogens, using yeast assays and MCF-7 breast cancer cells, demonstrated that TZ possesses moderate estrogenic activity, with low cytotoxicity and no genotoxicity detected in short-term exposures [69]. Finally, TZ tested at concentrations of 2.5, 5, and 10 mM again showed no genotoxicity and negligible cytotoxicity in human lymphocytes but confirmed significant cytotoxic effects in melanoma cells [70] Exposure to TZ was associated with increased activation of enzymes involved in epigenetic regulation in human cells such as HaCaT, HepG2, and A549. These changes may influence cell proliferation and survival processes, favoring the activation of oncogenic pathways. Additionally, significant DNA fragmentation was observed, indicating a potential genotoxic effect. These results suggest that TZ may contribute to epigenetic disruptions and increased cancer risk [71].
These studies suggest that, although TZ presents limited cytotoxicity in normal human cells, it may induce genotoxic effects and exhibit notable cytotoxicity in certain cancer cell lines. The cell type-specific responses emphasize the importance of using diverse cellular models in assessing the safety profile of this widely used azo dye.
5. Tartrazine Toxicity in Experimental Animal Models
Experimental studies conducted on experimental models indicate a significant toxic potential of TZ, especially under chronic exposure or at high doses (Table 4). Data obtained from Danio rerio show increased sensitivity to TZ during embryonic development. According to the study by Joshi and Katti [72], exposure to concentrations ≥10 mM was associated with the occurrence of deformities, edema, cardiovascular dysfunctions, delayed hatching, and increased mortality. At concentrations ≥75 mM, embryonic development was completely inhibited, highlighting the significant toxic potential of the substance on embryonic development. Similar results were reported by Jiang et al. [73], who noted reduction in embryonic survival, delayed hatching, swelling of the heart, and deformities along the body axis, with estimated effective concentration for 50% of the population and lethal concentration for 50% of the population values of 42.66 millimolar and 47.10 millimolar, respectively. Furthermore, Gupta et al. [74] observed that exposure to TZ in embryonic water, especially at concentrations of 0.5% and above, caused a significant increase in hatching rate and changes in SOD1 gene expression during early developmental stages. Additionally, Thanh et al. [75] identified a vascular toxicity profile, manifested by hemorrhages, edema, and abnormal vessel branching, correlated with altered migration and proliferation of endothelial cells. Adverse effects were also observed under chronic exposure conditions. Linskens et al. [76] demonstrated that prolonged administration of TZ (22 μM) caused cognitive deficits in adult fish, reflected by decreased learning capacity and cognitive flexibility. These changes were not present with limited exposure during the early post-embryonic period, suggesting a duration-dependent manifestation of neurotoxicity.
In rodents, oral administration of TZ induces significant structural and functional changes. Meena et al. [77] highlighted hematological and hepatic alterations at doses of 100–200 mg/kg, manifested by changes in organ weights and biochemical imbalances. Similarly, Arefin et al. [78] reported evidence of hepatotoxicity and nephrotoxicity at doses of 200–400 mg/kg, including increases in bilirubin, creatinine, and body weight. TZ significantly affects glucose homeostasis and pancreatic endocrine function, being associated with increased blood glucose levels and lipase activity, as well as decreased insulin, calcium, and magnesium levels after 30 days of administration [1]. Other histopathological effects include dilation of perineural spaces and severe degeneration of Purkinje cells, edema, and increased astrocyte populations. Subchronic exposure to TZ has been correlated with alterations in hepatic and renal parameters, evidenced by increases in alanine aminotransferase (ALT), aspartate aminotransferase (AST), urea, and total protein [64], as well as transaminases, lactate dehydrogenase (LDH), creatinine, and uric acid, with decreases in total proteins, albumin, and globulins [48,59,60].
At the neurobehavioral level, TZ induces hyperactivity, anxiety, and cognitive deficits in behavioral tests, alongside cerebral histological changes including neuronal apoptosis and vascular congestion, highlighting its neurotoxic effect [48,59,60]. In a complementary study, Kamel and El-Lathey [79] reported anxious and depressive effects, as well as decreased social interaction in male Wistar rats, suggesting a cumulative negative impact on mental health.
Chronic exposure (7.5–500 mg/kg) has been correlated with increases in hepatic enzymes (ALT, AST, alkaline phosphatase (ALP)) [26,30,54,61,63,80,81,84], renal markers (urea, creatinine, uric acid) [61,63,80,81,84], and lipid parameters (low-density lipoprotein (LDL), triglycerides) [30,61,65,80,81]. These changes were accompanied by histopathological lesions in vital organs such as the liver, kidneys, testes, stomach, colon, heart, and central nervous system [30,56,61,63,80,81,83,85,86]. Hematological and immunological alterations included leukocytosis, increased tumor necrosis factor alpha (TNF-α), and splenic changes, alongside decreases in hemoglobin and platelets [85,87]. Endocrine disorders, electrolyte imbalances, and neuroinflammation were also reported [60].
Altinoz et al. [56] documented severe intestinal lesions with losses of intestinal antioxidants and increases in lipid peroxidation. Additionally, cardiovascular impairment was suggested by elevated cardiac troponin, creatine kinase, and non- high-density lipoprotein cholesterol (HDL) cholesterol [83], indicating a myocardial risk in cases of prolonged exposure.
A significant number of studies have analyzed the protective potential of natural compounds. Curcumin, crocin, Nigella sativa oil, chlorophyll, and spinach fruit extract have proven effective in reducing OS, normalizing biochemical parameters, and alleviating tissue damage [30,56,61,62,63,65,81,82,85,86,89]. Al-Seeni et al. [61] reported in a study with doses of 10 mg/kg over 8 weeks hepatic and renal dysfunctions, histopathological lesions, and alterations in lipid profile and oxidative markers, with partial improvement following administration of Nigella sativa oil.
Available data consistently highlight the systemic toxic effects of TZ, manifested through hepatic, renal, hematological, metabolic, and neurological dysfunctions, confirmed by both biochemical changes and histopathological lesions. Prolonged exposure also induces hematological and immunological disorders, disturbances in glucose and electrolyte metabolism, as well as negative effects on the central nervous system, including anxiety, depression, reduced locomotor activity, and neuroinflammation [30,60,63,80,85,87,88].
6. Conclusions
Tartrazine remains a widely used synthetic food additive, yet accumulating scientific evidence raises serious concerns about its safety. Preclinical studies consistently report toxic effects such as oxidative stress, inflammation, neurotoxicity, metabolic disruption, hepatotoxicity, and potential genotoxicity. While these findings are mostly from animal models, the underlying mechanisms, including antioxidant system impairment and inflammatory pathway activation, are relevant to human disease. Although direct clinical evidence is limited, epidemiological data suggest associations between tartrazine exposure and adverse effects like hyperactivity, allergies, migraines, and behavioral issues, particularly in children. Given the potential for cumulative exposure, individual variability, and unclear thresholds for chronic toxicity, a re-evaluation of tartrazine’s safety is both timely and necessary. Replacing tartrazine with safer, natural alternatives should be prioritized to protect public health, especially for sensitive populations.
7. Future Perspectives
For a comprehensive understanding of the risks associated with tartrazine, long-term clinical studies, identification of specific biomarkers, and clarification of the involved epigenetic mechanisms are needed. In parallel, the development of safe natural alternatives and the reevaluation of current regulations become essential priorities for protecting public health.
Author Contributions
Conceptualization, M.V., A.S. and A.C.; methodology, M.V., V.R. and C.A.; validation, V.B., M.H. and A.C.; formal analysis, M.V. and A.S.; investigation, resources, data curation, M.V., A.S., V.R., V.B., G.P., C.I., A.C., C.A., M.H; writing—original draft preparation, M.V., A.S. V.R. and I.-M.B; writing—review and editing, G.P. and C.A; visualization, V.R., C.I. and I.-M.B.; supervision, V.B. and M.H. All authors have read and agreed to the published version of the manuscript.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Rehman, K.; Ashraf, A.; Azam, F.; Hamid Akash, M.S. Effect of Food Azo-Dye Tartrazine on Physiological Functions of Pancreas and Glucose Homeostasis. Turk. J. Biochem. 2019, 44, 197–206. [Google Scholar] [CrossRef]
- Leulescu, M.; Rotaru, A.; Pălărie, I.; Moanţă, A.; Cioateră, N.; Popescu, M.; Morîntale, E.; Bubulică, M.V.; Florian, G.; Hărăbor, A.; et al. Tartrazine: Physical, Thermal and Biophysical Properties of the Most Widely Employed Synthetic Yellow Food-Colouring Azo Dye. J Therm Anal Calorim 2018, 134, 209–231. [Google Scholar] [CrossRef]
- Chung, K.T. Azo Dyes and Human Health: A Review. J Environ Sci Health C Environ Carcinog Ecotoxicol Rev 2016, 34, 233–261. [Google Scholar] [CrossRef]
- Ambroziewicz, Z.M.; Siemiątkowski, R.; Łata, M.; Dowgiert, S.; Sikorska, M.; Kamiński, J.; Więcław, K.; Grabowska, H.; Chruściel, J.; Mąsior, G. Long-Term Health Effects of Artificially Colored Foods in Adults and Children: A Review of Scientific Literature on Attention Deficits, Carcinogenicity, and Allergy Risks. J. Educ. Health Sport 2024, 76, 56522. [Google Scholar] [CrossRef]
- Kamat, P.; Shetti, P.P.; Paranjape, R.; Shetti, P.P. Techniques for Detection and Measurement of Tartrazine in Food Products 2008 to 2022: A Review. J Chem Health Risks 2024, 14, 721–729. [Google Scholar]
- Mehedi, N.; Ainad-Tabet, S.; Mokrane, N.; Addou, S.; Zaoui, C.; Kheroua, O.; Saidi, D. Reproductive Toxicology of Tartrazine (FD and C Yellow No. 5) in Swiss Albino Mice. Am J Pharmacol Toxicol 2009, 4, 130–135. [Google Scholar] [CrossRef]
- Barciela, P.; Perez-Vazquez, A.; Prieto, M.A. Azo Dyes in the Food Industry: Features, Classification, Toxicity, Alternatives, and Regulation. Food Chem. Toxicol 2023, 178, 113935. [Google Scholar] [CrossRef]
- Alshehrei, F. Role of Microorganisms in Biodegradation of Food Additive Azo Dyes: A Review. Afr J Biotechnol 2020, 19, 799–805. [Google Scholar] [CrossRef]
- Aali, R.; Yari, A.R.; Ghafuri, Y.; Behnamipour, S. Health Risk Assessment of Synthetic Tartrazine Dye in Some Food Products in Qom Province (Iran). Curr Nutr Food Sci 2024, 20, 726–733. [Google Scholar] [CrossRef]
- World Health Organization. Available online: https://apps.who.int/food-additives-contaminants-jecfa-database/Home/Chemical/3885 (accessed on 11 July 2025).
- Vander Leek, T.K. Food Additives and Reactions: Antioxidants, Benzoates, Parabens, Colorings, Flavorings, Natural Protein-Based Additives. Encycl. Food Allergy 2024, 862–881. [Google Scholar] [CrossRef]
- Wróblewska, B. Influence of Food Additives and Contaminants (Nickel and Chromium) on Hypersensitivity and Other Adverse Health Reactions - A Review. Pol J Food Nutr Sci 2009, 59, 287–294. Available online: https://journal.pan.olsztyn.pl/pdf-98219-30948?filename=30948.pdf (accessed on 11 July 2025).
- Pestana, S.; Moreira, M.; Olej, B. Safety of Ingestion of Yellow Tartrazine by Double-Blind Placebo Controlled Challenge in 26 Atopic Adults. Allergol Immunopathol (Madr) 2010, 38, 142–146. [Google Scholar] [CrossRef] [PubMed]
- Banc, R.; Filip, L.; Cozma-Petruț, A.; Ciobârcă, D.; Miere, D. Yellow and Red Synthetic Food Dyes and Potential Health Hazards: A Mini Review. Bulletin of University of Agricultural Sciences and Veterinary Medicine Cluj-Napoca. Food Science and Technology 2024, 81, 1–17. [Google Scholar] [CrossRef]
- Bateman, B.J. The Behaviour of Three Year Olds in Relation to Allergy and Exposure to Artificial Additives. University of Southampton, Doctoral Thesis, 2004. Available online: https://eprints.soton.ac.uk/465625/ (accessed on 11 July 2025).
- Tuormaa, T.E. The Adverse Effects of Food Additives on Health: A Review of the Literature with Special Emphasis on Childhood Hyperactivity. J. Orthomol. Med 1994, 9, 225–243. Available online: https://orthomolecular.org/library/jom/1994/articles/1994-v09n04-p225.shtml (accessed on 21 July 2025).
- Feketea, G.; Tsabouri, S. Common Food Colorants and Allergic Reactions in Children: Myth or Reality? Food Chem 2017, 230, 578–588. [Google Scholar] [CrossRef]
- Arnold, L.E.; Lofthouse, N.; Hurt, E. Artificial Food Colors and Attention-Deficit/Hyperactivity Symptoms: Conclusions to Dye For. Neurotherapeutics 2012, 9, 599–609. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.M.; Reboredo, F.H.; Lidon, F.C. Food Colour Additives: A Synoptical Overview on Their Chemical Properties, Applications in Food Products and Health Side Effects. Foods 2022, 11, 379. [Google Scholar] [CrossRef] [PubMed]
- Rowe, K.S.; Rowe, K.J. Synthetic Food Coloring and Behavior: A Dose Response Effect in a Double-Blind, Placebo-Controlled, Repeated-Measures Study. J Pediatr 1994, 125, 691–698. [Google Scholar] [CrossRef] [PubMed]
- Visweswaran, B. Oxidative Stress by Tartrazine in the Testis of Wistar Rats. IOSR J Pharm Biol Sci 2012, 2, 44–49. [Google Scholar] [CrossRef]
- El-Desoky, G.E.; Wabaidur, S.M.; Alothman, Z.A.; Habila, M.A. Regulatory Role of Nano-Curcumin against Tartrazine-Induced Oxidative Stress, Apoptosis-Related Genes Expression, and Genotoxicity in Rats. Molecules 2020, 25, 5801. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Lv, X.; Zhang, Y.; Xin, Q.; Zou, Y.; Li, X. Tartrazine Exposure Results in Histological Damage, Oxidative Stress, Immune Disorders and Gut Microbiota Dysbiosis in Juvenile Crucian Carp (Carassius Carassius). Aquatic Toxicology 2021, 241, 105998. [Google Scholar] [CrossRef]
- Wu, L.; Xu, Y.; Lv, X.; Chang, X.; Ma, X.; Tian, X.; Shi, X.; Li, X.; Kong, X. Impacts of an Azo Food Dye Tartrazine Uptake on Intestinal Barrier, Oxidative Stress, Inflammatory Response and Intestinal Microbiome in Crucian Carp (Carassius Auratus). Ecotoxicol Environ Saf 2021, 223, 112551. [Google Scholar] [CrossRef]
- Elhkim, M.O.; Héraud, F.; Bemrah, N.; Gauchard, F.; Lorino, T.; Lambré, C.; Frémy, J.M.; Poul, J.M. New Con-siderations Regarding the Risk Assessment on Tartrazine. An Update Toxicological Assessment, Intolerance Reactions and Maximum Theoretical Daily Intake in France. Regul Toxicol Pharmacol. 2007, 47, 308–316. [Google Scholar] [CrossRef]
- Khayyat, L.; Essawy, A.; Sorour, J.; Soffar, A. Tartrazine Induces Structural and Functional Aberrations and Genotoxic Effects in Vivo. PeerJ 2017, 5, e3041. [Google Scholar] [CrossRef]
- Tanaka, T. Reproductive and Neurobehavioural Toxicity Study of Tartrazine Administered to Mice in the Diet. Food Chem Toxicol 2006, 44, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Mandelker, L. Oxidative Stress, Free Radicals, and Cellular Damage. Studies in Veterinary Medicine 2011, 1–17. [Google Scholar] [CrossRef]
- Aldaamy, A.M.Z.; Al-Zubiady, N.M.H. Study on Toxic Effect of Tartrazine Pigment on Oxidative Stress in Male Albino Rats. Biochem Cell Arch 2021, 21, 1021–1026. [Google Scholar]
- El-Desoky, G.E.; Abdel-Ghaffar, A.; Al-Othman, Z.A.; Habila, M.A.; Al-Sheikh, Y.A.; Ghneim, H.K.; Giesy, J.P.; Aboul-Soud, M.A.M. Curcumin Protects against Tartrazine-Mediated Oxidative Stress and Hepatotoxicity in Male Rats. Eur Rev Med Pharmacol Sci 2017, 21, 635–645. Available online: https://pubmed.ncbi.nlm.nih.gov/28239801/ (accessed on 18 July 2025). [PubMed]
- Ramos-Souza, C.; Nass, P.; Jacob-Lopes, E.; Zepka, L.Q.; Braga, A.R.C.; De Rosso, V.V. Changing Despicable Me: Potential Replacement of Azo Dye Yellow Tartrazine for Pequi Carotenoids Employing Ionic Liquids as High-Performance Extractors. Int. Food Res. J 2023, 174, 113593. [Google Scholar] [CrossRef]
- Amchova, P.; Siska, F.; Ruda-Kucerova, J. Safety of Tartrazine in the Food Industry and Potential Protective Factors. Heliyon 2024, 10, e38111. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.; Fondriest Gentry, A.; Schwartz, R.; Bauman, J. Tartrazine-Containing Drugs. Drug Intell Clin Pharm 1981, 15, 782–788. [Google Scholar] [CrossRef]
- Miller, K. Sensitivity to Tartrazine. Br Med J 1982, 285, 1597–1598. [Google Scholar] [CrossRef]
- Pennington, C.R. Diet and Disease. J. Nutr. Ther. 1988, 226–262. [Google Scholar] [CrossRef]
- Moutinho, I.L.D.; Bertges, L.C.; Assis, R.V.C. Prolonged Use of the Food Dye Tartrazine (FD&C Yellow N° 5) and Its Effects on the Gastric Mucosa of Wistar Rats. Braz J. Biol 2007, 67, 141–145. [Google Scholar] [CrossRef] [PubMed]
- Skypala, I. Other Causes of Food Hypersensitivity. In Food Hypersensitivity: Diagnosing and Managing Food Allergies and Intolerance; 2009; Available online: https://onlinelibrary.wiley.com/doi/book/10.1002/9781444312119#page=219 (accessed on 18 July 2025).
- Bush, R.K.; Taylor, S.L. Reactions to Food and Drug Additives. Middleton’s Allergy: Principles and Practice: Eighth Edition 2014, 2–2, 1340–1356. [Google Scholar] [CrossRef]
- Breitkreutz, J.; Tuleu, C. Pediatric and Geriatric Pharmaceutics and Formulatio. Modern Pharmaceutics: Fifth Edition, 2016; 2–2. Available online: https://www.taylorfrancis.com/chapters/edit/10.1201/b14445-32/pediatric-geriatric-pharmaceutics-formulation (accessed on 18 July 2025).
- Rovina, K.; Siddiquee, S.; Shaarani, S.M. A Review of Extraction and Analytical Methods for the Determination of Tartrazine (E 102) in Foodstuffs. Crit Rev Anal Chem 2017, 47, 309–324. [Google Scholar] [CrossRef] [PubMed]
- Sezgin, A. Food Additives : Colorants Food Additives : Colorants. In Science Within Food: Up- to - Date Advance s on Research and Educational Ideas. Researchgate 2018, 122, 87–94. Available online: https://www.researchgate.net/publication/322404775_Food_Additives_Colorants (accessed on 28 July 2025).
- Sharma, R.; Bamola, S.; Kumar Verma, S. Effects of Acid Yellow 23 Food Dye on Environment and Its Removal on Various Surfaces-A Mini Review. IRJET 2020, 7, 4550–4573. [Google Scholar]
- Koop, B.L.; Maciel, A.G.; Soares, L.S.; Monteiro, A.R.; Valencia, G.A. Natural Colorants. Natural Additives in Foods. 2022, pp. 87–122. Available online: https://link.springer.com/chapter/10.1007/978-3-031-17346-2_4 (accessed on 18 July 2025).
- Athira, N.; Jaya, D.S. Effects of Tartrazine on Growth and Brain Biochemistry of Indian Major Carps on Long-Term Exposure. Int. J. Adv. Biochem. Res 2022, 6, 17–25. [Google Scholar] [CrossRef]
- Hegazy, A.A.; Awad Hegazy, A.; Haliem, W.A.; Haliem, R.A.; Mamdouh El-Bestawy, E.; Mohammad, G.; Ali, E. Brief Overview about Tartrazine Effects on Health. Eur. Chem. Bull 2023, 12, 4698–4707. [Google Scholar]
- Pay, R.; Sharrock, A. V.; Elder, R.; Maré, A.; Bracegirdle, J.; Torres, D.; Malone, N.; Vorster, J.; Kelly, L.; Ryan, A.; et al. Preparation, Analysis and Toxicity Characterisation of the Redox Metabolites of the Azo Food Dye Tartrazine. Food Chem Toxicol 2023, 182, 114193. [Google Scholar] [CrossRef] [PubMed]
- Kamal, A.A.; Fawzia, S.E.-S. Toxicological and Safety Assessment of Tartrazine as a Synthetic Food Additive on Health Biomarkers: A Review. Afr J Biotechnol 2018, 17, 139–149. [Google Scholar] [CrossRef]
- Gao, Y.; Li, C.; Shen, J.; Yin, H.; An, X.; Jin, H. Effect of Food Azo Dye Tartrazine on Learning and Memory Functions in Mice and Rats, and the Possible Mechanisms Involved. J Food Sci 2011, 76, T125–T129. [Google Scholar] [CrossRef]
- McCann, D.; Barrett, A.; Cooper, A.; Crumpler, D.; Dalen, L.; Grimshaw, K.; Kitchin, E.; Lok, K.; Porteous, L.; Prince, E.; et al. Food Additives and Hyperactive Behaviour in 3-Year-Old and 8/9-Year-Old Children in the Community: A Randomised, Double-Blinded, Placebo-Controlled Trial. Lancet 2007, 370, 1560–1567. [Google Scholar] [CrossRef] [PubMed]
- Nettis, E.; Colanardi, M.C.; Ferrannini, A.; Tursi, A. Suspected Tartrazine-Induced Acute Urticar-ia/Angioedema Is Only Rarely Reproducible by Oral Rechallenge. Clin Exp Allergy 2003, 33, 1725–1729. [Google Scholar] [CrossRef]
- Araújo Caldas, L. De; Caldas Marmo, F.; da Costa, P.V.; Viana Jacobson, L.S. Hormesis in Tartrazine Allergic Responses of Atopic Patients: An Overview of Clinical Trials and a Raw Data Revision. Environ Dis 2020, 5, 59. [Google Scholar] [CrossRef]
- Soares, B.M.; Araújo, T.M.T.; Ramos, J.A.B.; Pinto, L.C.; Khayat, B.M.; De Oliveira Bahia, M.; Montenegro, R.C.; Burbano, R.M.R.G.; Khayat, A.S. Effects on DNA Repair in Human Lymphocytes Exposed to the Food Dye Tartrazine Yellow. Anticancer Res 2015, 35, 1465–1474. Available online: https://pubmed.ncbi.nlm.nih.gov/25750299/ (accessed on 18 July 2025).
- Boussada, M.; Lamine, J.A.; Bini, I.; Abidi, N.; Lasrem, M.; El-Fazaa, S.; El-Golli, N. Assessment of a Sub-Chronic Consumption of Tartrazine (E102) on Sperm and Oxidative Stress Features in Wistar Rat. Int Food Res J 2017, 24, 1473–1481. [Google Scholar]
- Amin, K.A.; Abdel Hameid, H.; Abd Elsttar, A.H. Effect of Food Azo Dyes Tartrazine and Carmoisine on Bi-ochemical Parameters Related to Renal, Hepatic Function and Oxidative Stress Biomarkers in Young Male Rats. Food Chem Toxicol, 2010, 48, 2994–2999. [Google Scholar] [CrossRef]
- El Golli, N.; Bini-Dhouib, I.; Jrad, A.; Boudali, I.; Nasri, B.; Belhadjhmida, N.; El Fazaa, S. Toxicity Induced after Subchronic Administration of the Synthetic Food Dye Tartrazine in Adult Rats, Role of Oxidative Stress. Recent Adv Biol Med 2016, 02, 20. [Google Scholar] [CrossRef]
- Altinoz, E.; Erdemli, M.E.; Gül, M.; Erdemli, Z.; Gül, S.; Turkoz, Y. Prevention of Toxic Effects of Orally Administered Tartrazine by Crocin in Wistar Rats. Toxicol Environ Chem 2021, 103, 184–198. [Google Scholar] [CrossRef]
- Ozougwu, J.C. Physiology of the Liver. IJRPB, 2017, 4, 13–24. Available online: https://www.academia.edu/38598619/Physiology_of_the_liver?auto=download (accessed on 18 July 2025).
- Kiran, T.R.; Otlu, O.; Karabulut, A.B. Oxidative Stress and Antioxidants in Health and Disease. J. Lab. Med 2023, 47, 1–11. [Google Scholar] [CrossRef]
- Hosieny, N.A.; Mona, E.; Ahmed, S.M.; Zayed, M. Toxic Effects of Food Azo Dye Tartrazine on the Brain of Young Male Albino Rats: Role of Oxidative Stress. Zagazig J Forensic Med 2020, 19, 60–73. [Google Scholar] [CrossRef]
- Wopara, I.; Adebayo, O.G.; Umoren, E.B.; Aduema, W.; Iwueke, A. V.; Etim, O.E.; Pius, E.A.; James, W.B.; Wodo, J. Involvement of Striatal Oxido-Inflammatory, Nitrosative and Decreased Cholinergic Activity in Neurobehavioral Alteration in Adult Rat Model with Oral Co-Exposure to Erythrosine and Tartrazine. Heliyon 2021, 7, e08454. [Google Scholar] [CrossRef]
- Al-Seeni, M.N.; El Rabey, H.A.; Al-Hamed, A.M.; Zamazami, M.A. Nigella Sativa Oil Protects against Tartrazine Toxicity in Male Rats. Toxicol Rep 2018, 5, 146–155. [Google Scholar] [CrossRef]
- Abd-Elhakim, Y.M.; Moustafa, G.G.; Hashem, M.M.; Ali, H.A.; Abo-EL-Sooud, K.; El-Metwally, A.E. Influence of the Long-Term Exposure to Tartrazine and Chlorophyll on the Fibrogenic Signalling Pathway in Liver and Kidney of Rats: The Expression Patterns of Collagen 1-α, TGFβ-1, Fibronectin, and Caspase-3 Genes. Environ Sci Pollut Res Int 2019, 26, 12368–12378. [Google Scholar] [CrossRef]
- Velioglu, C.; Erdemli, M.E.; Gul, M.; Erdemli, Z.; Zayman, E.; Bag, H.G.; Altinoz, E. Protective Effect of Crocin on Food Azo Dye Tartrazine-Induced Hepatic Damage by Improving Biochemical Parameters and Oxidative Stress Biomarkers in Rats. Gen Physiol Biophys 2019, 38, 73–82. [Google Scholar] [CrossRef]
- Abdelgayed, S.S. Toxicological and Histopathological Studies on the Effect of Tartrazine in Male Albino Rats. IJABE 2016, 10, 469–474. [Google Scholar]
- Erdemli, Z.; Altinoz, E.; Erdemli, M.E.; Gul, M.; Bag, H.G.; Gul, S. Ameliorative Effects of Crocin on Tartrazine Dye–Induced Pancreatic Adverse Effects: A Biochemical and Histological Study. Environ Sci Pollut Res Int 2021, 28, 2209–2218. [Google Scholar] [CrossRef] [PubMed]
- Haridevamuthu, B.; Murugan, R.; Seenivasan, B.; Meenatchi, R.; Pachaiappan, R.; Almutairi, B.O.; Arokiyaraj, S.; M. K, K.; Arockiaraj, J. Synthetic Azo-Dye, Tartrazine Induces Neurodevelopmental Toxicity via Mitochondria-Mediated Apoptosis in Zebrafish Embryos. J Hazard Mater 2024, 461, 132524. [CrossRef]
- Floriano, J.M.; Rosa, E. da; do Amaral, Q.D.F.; Zuravski, L.; Chaves, P.E.E.; Machado, M.M.; De Oliveira, L.F.S. Is Tartrazine Really Safe? In Silico and Ex Vivo Toxicological Studies in Human Leukocytes: A Question of Dose. Toxicol Res (Camb) 2018, 7, 1128–1134. [Google Scholar] [CrossRef]
- Pasdaran, A.; Azarpira, N.; Heidari, R.; Nourinejad, S.; Zare, M.; Hamedi, A. Effects of Some Cosmetic Dyes and Pigments on the Proliferation of Human Foreskin Fibroblasts and Cellular Oxidative Stress; Potential Cytotoxicity of Chlorophyllin and Indigo Carmine on Fibroblasts. J Cosmet Dermatol 2022, 21, 3979–3985. [Google Scholar] [CrossRef]
- Nasri, A.; Pohjanvirta, R. In Vitro Estrogenic, Cytotoxic, and Genotoxic Profiles of the Xenoestrogens 8-Prenylnaringenine, Genistein and Tartrazine. Environ. Sci. Pollut. Res. 2021, 28, 27988–27997. [Google Scholar] [CrossRef] [PubMed]
- Haverić, A.; Inajetović, D.; Vareškić, A.; Hadžić, M.; Haverić, S. In vitro Analysis of Tartrazine Genotoxicity and Cytotoxicity. G& A 2017, 1, 37–43. [Google Scholar] [CrossRef]
- Zand, A.; Macharia, J.M.; Szabó, I.; Gerencsér, G.; Molnár, Á.; Raposa, B.L.; Varjas, T. The Impact of Tartrazine on DNA Methylation, Histone Deacetylation, and Genomic Stability in Human Cell Lines. Nutrients 2025, 17, 913. [Google Scholar] [CrossRef] [PubMed]
- Joshi, V.; Katti, P. Developmental Toxicity Assay for Food Additive Tartrazine Using Zebrafish (Danio Rerio) Embryo Cultures. Int J Toxicol 2018, 37, 38–44. [Google Scholar] [CrossRef]
- Jiang, L.L.; Li, K.; Yan, D.L.; Yang, M.F.; Ma, L.; Xie, L.Z. Toxicity Assessment of 4 Azo Dyes in Zebrafish Embryos. Int J Toxicol 2020, 39, 115–123. [Google Scholar] [CrossRef]
- Gupta, R.; Ranjan, S.; Yadav, A.; Verma, B.; Malhotra, K.; Madan, M.; Chopra, O.; Jain, S.; Gupta, S.; Joshi, A.; et al. Toxic Effects of Food Colorants Erythrosine and Tartrazine on Zebrafish Embryo Development. Curr Res Nutr Food Sci 2019, 7, 876–885. [Google Scholar] [CrossRef]
- Thanh, D.D.; Bich-Ngoc, N.; Paques, C.; Christian, A.; Herkenne, S.; Struman, I.; Muller, M. The Food Dye Tartrazine Disrupts Vascular Formation Both in Zebrafish Larvae and in Human Primary Endothelial Cells. Scientific Reports 2024, 14, 1–10. [Google Scholar] [CrossRef]
- Linskens, A. The Long Term Effects of Tartrazine (FD&C Yellow No. 5) on Learning, Cognitive Flexibility, and Memory of Zebrafish (Danio Rerio) Embryos into Adulthood. Available online: https://cpb-us-w2.wpmucdn.com/sites.uwm.edu/dist/8/202/files/2018/06/Linskens_paper_LM_Seymour_2018-1lxiur8.pdf (accessed on 24 Jul 2025).
- Meena, B.; Sharma, S.; Sharma, S. Effects of Sub-Chronic Exposure of the Food Dye Amaranth on the Biochemistry of Swiss Albino Mice (Mus Musculus L.). IJISRT 2017, 21, 535–545. [Google Scholar] [CrossRef]
- Arefin, S.; Hossain, M.S.; Neshe, S.A.; Rashid, Md.M.O.; Tohidul Amin, M.; Hussain, Md.S. Tartrazine Induced Changes in Physiological and Biochemical Parameters in Swiss Albino Mice, Mus Musculus. Marmara Pharm J 2017, 21, 564–569. [Google Scholar] [CrossRef]
- Kamel, M.M.; El-Lethey, H.S. The Potential Health Hazard of Tartrazine and Levels of Hyperactivity, Anxiety-Like Symptoms, Depression and Anti-Social Behaviour in Rats. Am. J. Sci 2011, 7, 1211–1218. [Google Scholar]
- Usman, J.N.; Muhammad, G.A. Sub-Acute Toxicity Study on Tartrazine in Male Albino Rats. DUJOPAS 2022, 8, 97–105. [Google Scholar] [CrossRef]
- El-Desoky, G.E.; Wabaidur, S.M.; AlOthman, Z.A.; Habila, M.A. Evaluation of Nano-Curcumin Effects against Tartrazine-Induced Abnormalities in Liver and Kidney Histology and Other Biochemical Parameters. Food Sci Nutr 2022, 10, 1344–1356. [Google Scholar] [CrossRef]
- Erdemli, M.E.; Gul, M.; Altinoz, E.; Zayman, E.; Aksungur, Z.; Bag, H.G. The Protective Role of Crocin in Tartrazine Induced Nephrotoxicity in Wistar Rats. Biomed Pharmacother 2017, 96, 930–935. [Google Scholar] [CrossRef]
- Iroh, G.; Weli, B.O.; Adele, U.A.; Briggs, O.N.; Waribo, H.A.; Elekima, I. Assessment of Atherogenic Indices and Markers of Cardiac Injury in Albino Rats Orally Administered with Tartrazine Azo Dye. J Adv Med Pharm Sci 2020, 22, 51–61. [Google Scholar] [CrossRef]
- Aldaamy, A.; Merza Hamza, N.; Masikh Zebalah Al-Daamy, A.; Marza Hamza Al-Zubiady, N. Study of The Toxic Effect of Tartrazine Dye on Some Biochemical Parameters in Male Albino Rats. Sci. J. Med. Res. 2020, 4, 111–117. [Google Scholar]
- Abd-Elhakim, Y.M.; Hashem, M.M.; El-Metwally, A.E.; Anwar, A.; Abo-EL-Sooud, K.; Moustafa, G.G.; Ali, H.A. Comparative Haemato-Immunotoxic Impacts of Long-Term Exposure to Tartrazine and Chlorophyll in Rats. Int Immunopharmacol 2018, 63, 145–154. [Google Scholar] [CrossRef]
- Balta, I.; Sevastre, B.; Mireşan, V.; Taulescu, M.; Raducu, C.; Longodor, A.L.; Marchiş, Z.; Mariş, C.S.; Coroian, A. Protective Effect of Blackthorn Fruits (Prunus Spinosa) against Tartrazine Toxicity Development in Albino Wistar Rats. BMC Chem 2019, 13, 104. [Google Scholar] [CrossRef] [PubMed]
- Himri, I.; Bellahcen, S.; Souna, F.; Belmakki, F.; Aziz, M.; Bnouham, M.; Zoheir, J.; Berkia, Z.; Mekhfi, H.; Saalaoui, E. A 90-Day Oral Toxicity Study of Tartrazine, a Synthetic Food Dye, in Wistar Rats. Int J Pharm Pharm Sci 2011, 3, 159–169. [Google Scholar]
- El-Sakhawy, M.A.; Mohamed, D.W.; Ahmed, Y.H. Histological and Immunohistochemical Evaluation of the Effect of Tartrazine on the Cerebellum, Submandibular Glands, and Kidneys of Adult Male Albino Rats. Environ Sci Pollut Res Int. 2019, 26, 9574–9584. [Google Scholar] [CrossRef] [PubMed]
- Abd, B.E.; Naby, E.; Shalaby, R.A.; Fouda, F.M.; Ebiya, R.A. Evaluation of biochemical effects of tartrazine and curcumin in male albino rats. World J. Pharm. Res 2022, 11, 1472–1486. [Google Scholar] [CrossRef]
Figure 1.
Toxic Mechanisms Involved in Tartrazine Exposure (Figure partially created using BioRender resources)
Figure 1.
Toxic Mechanisms Involved in Tartrazine Exposure (Figure partially created using BioRender resources)
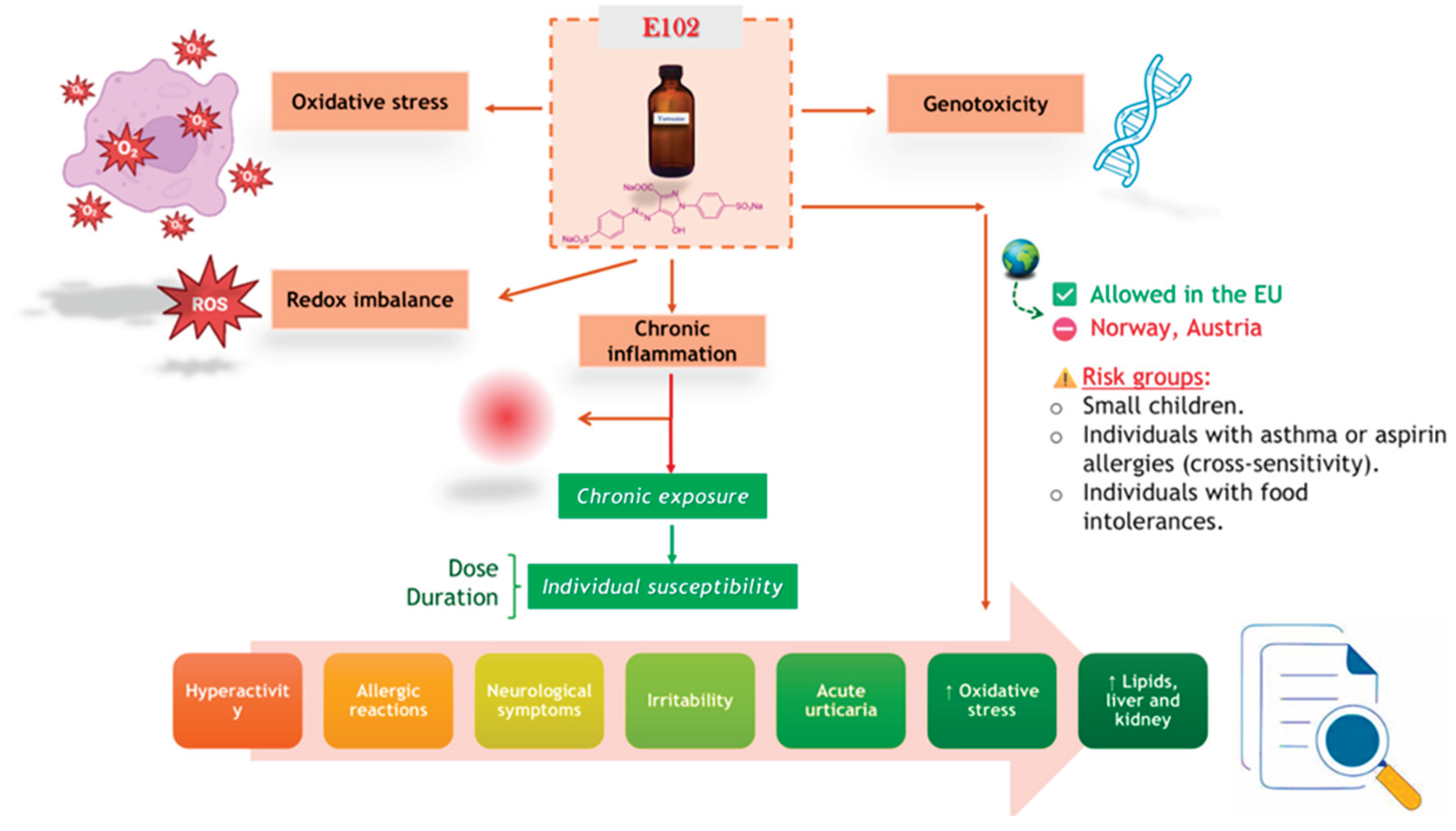

Table 1.
Common Uses of Tartrazine and Relevant Toxicological Concerns.
| Category | Product examples | References |
| Food products | TZ is widely used to impart an intense yellow color in various food products such as bread, beverages, cereals, peanuts, candies, jellies, chewing gum, flavored chips, creams, ice cream, yogurts, cakes, instant desserts, soups, sauces, jam, flavored rice, and pasta, but due to its potential side effects, there is a growing tendency to replace it with natural pigments like annatto or beta-carotene. |
[2,35,37,40,41,42,43,45] |
| Pharmaceuticals | TZ is used as a coloring excipient in multivitamins, gelatin capsules, tablets, syrups, and pediatric medicines, but in some cases it may cause allergic reactions or asthma in sensitive individuals. | [33,36,38,39,40] |
| Non-food products | TZ is also present in non-food products such as soaps, cosmetics, shampoos, hair conditioners, pastels, crayons, and stamp dyes. It may cause hypersensitivity reactions upon skin contact. | [25,34,44,45] |
Table 2.
Evidence of Oxidative Imbalance Following Tartrazine Exposure in Rodent Models.
| Experimental organism | n= | About species | Dose | Time | Method of administration | Sample | Effect | References |
| Rats |
18 | Young male albino rats (28 days, 60–80 g) | 320 mg/kg tartrazine in 1 ml distilled water, daily | 4 weeks | oral gavage | Brain | ↓ GPx ↑ MDA |
[59] |
| 50 | Wistar male albino rats | 7.5 mg/kg | 90 days | Diets containing dry mass | Liver | ↑ MDA ↓ GSH |
[30] | |
| Serum | ↓ SOD ↓ CAT ↓ GPx |
|||||||
| 24 | Male Wistar rats (10–12 weeks, 180–200 g) | 2, 6, 10 mg/kg (erythrosine + TZ 50:50 mix) | 6 weeks | oral gavage | Brain tissue | ↑ MDA ↓ GSH ↓ CAT ↑ ACHe. |
[60] | |
| 18 | Male albino rats | 10 mg/kg (+3.75 mg/kg sulfanilic acid) | 8 weeks | oral administration | Serum, liver and kidney tissue homogenate | ↑ MDA ↓GSH ↓ SOD ↓ CAT ↓ GR |
[61] | |
| 18 | White albino rats of either sex | Low (10 mg/kg) and high (50 mg/kg) doses | 15 and 30 days | oral administration | Serum | ↓ SOD |
[1] | |
| 30 | Sprague-Dawley male albino rats (150-200 g) | 75 mg/kg | 90 days | Oral administration by orogastric gavage | Hepatic and renal tissue homogenate | ↑ MDA ↓ GSH ↓ SOD ↓ CAT |
[62] | |
| 18 | Male albino rats (10–15 weeks, 190–250 g) | 400 mg/kg | 30 days | oral administration | Serum | ↑ MDA ↓ GSH ↓ SOD ↓ CAT ↓ GPx |
[29] | |
| 40 | Female Wistar albino rats (225–250 g) | 500 mg/kg | 21 days | oral gavage | Tissue homogen ates |
↑ MDA ↑ SOD ↑ TOS ↓ GSH ↓ CAT ↓ TAS |
[63] | |
| 40 | Adult female Wistar rats (225–250 g, 8–10 weeks) | 500 mg/kg | 3 weeks | oral gavage | Tissue homogenates | ↑ MDA ↑ TOS, ↓ GSH ↓ SOD ↓ CAT ↓ TAS |
[56] | |
| 30 | Adult male Sprague Dawley rats (120-150 g) | 200 mg/kg | 60 days | oral administration | Tissue homogenate | ↑ MDA |
[64] | |
| 40 | Female Wistar rats, (225–250 g) | 500 mg/kg | 3 weeks | oral gavage | Tissue homogenate | ↑ MDA ↑ TOS ↓ GSH ↓ TAS ↓ SOD ↓ CAT. |
[65] | |
| 20 | Male Wistar rats, (130 ± 40 g) |
300 mg | 30 days | oral administration | Tissue homogenate | ↑MDA ↑CAT ↑GST |
[55] | |
| 36 | Young male albino rats (60–80 g) | low doses of TZ 15 mg/kg bw | 30 days | oral administration | Liver tissue homogenate | ↑ MDA ↓ CAT ↓ SOD |
[54] | |
| high doses were 500 mg/kg bw | ↑ MDA ↓ CAT ↓ SOD ↓ GSH |
|||||||
| 40 | Sprague–Dawley rats (70 ± 10 g) |
175, 350, and 700 mg/kg bw | 30 days | oral gavage | Brain tissue | ↓GSH ↓SOD ↑MDA |
[48] |
ACHe – Acetylcholinesterase; bw – Body Weight; CAT – Catalase; GPx – Glutathione peroxidase; GR – Glutathione reductase; GSH – Glutathione; GST – Glutathione S-transferase; MDA – Malondialdehyde; SOD – Superoxide dismutase; TAS – Total Antioxidant Status; TOS – Total Oxidant Status; TZ – Tartrazine, ↑ - significantly increased; ↓ - significantly decreased.
Table 3.
Summary of Experimental Studies Assessing the Cytotoxic and Genotoxic Effects of Tartrazine in Different Human Cell Types and Assays.
Table 3.
Summary of Experimental Studies Assessing the Cytotoxic and Genotoxic Effects of Tartrazine in Different Human Cell Types and Assays.
| Cell Type | Concentration Tested | Tests Performed | Key Findings | References |
| Human lymphocytes | 0.25 – 64.0 mM | MTT assay, alkaline comet assay | No cytotoxicity; genotoxic at all doses; partial DNA repair. | [52] |
| Human leukocytes | 5 – 500 μg/mL | Trypan Blue viability, Micronucleus test, Comet assay, Cytogenetics, In silico | No cytotoxicity/mutagenicity; DNA damage at ≥70 μg/mL; supported by in silico models. | [67] |
| Human foreskin fibroblasts | 10, 100, 250, 500, 1000, and 2000 μg/ml for various dyes | MTT assay, ROS, lipid peroxidation, LDH | TZ: no effect; indigo carmine/chlorophyllin cytotoxic at high doses | [68] |
| Yeast assay, MCF-7 breast cancer cells | Not specified (short-term) | Estrogenic activity, LDH release, micronucleus test | 8-PN showed the strongest estrogenic effect, followed by TZ and genistein; all exhibited low cytotoxicity and no genotoxicity. | [69] |
| Human lymphocytes, GR-M melanoma cells | 2.5, 5and 10 mM | Chromosome aberration, CBMN assay, trypan blue test | No genotoxicity in lymphocytes; low cytotoxicity in lymphocytes; high cytotoxicity in melanoma cells | [70] |
| HaCaT | 20 µM, 40 µM și 80 µM | qRT-PCR Alkaline Comet Assay |
Upregulated DNMT and HDAC genes with increased DNA fragmentation, indicating epigenetic and genotoxic effects. | [71] |
| HepG2 | ||||
| A549 |
8-PN – 8-Prenylnaringenin; CBMN assay – Cytokinesis-Block Micronucleus Assay; DNA - Deoxyribonucleic Acid; DNMT – DNA Methyltransferase; HDAC – Histone Deacetylase; HepG2 – Human liver cancer cell line; LDH - Lactate Dehydrogenase; MTT - Methyl Thiazolyl Tetrazolium; qRT-PCR – Quantitative Real-Time Polymerase Chain Reaction; ROS - reactive oxygen species; TZ – Tartrazine.
Table 4.
Effects of Tartrazine on Behavior and Biochemistry in Animal Models
| Experimental model | n= | Method of administration | Time | Dose | Analysis | Effect | Ref. |
| Zebrafish | |||||||
| Zebrafish embryo |
20/concentration | Exposed to E3 medium with varying TZ concentrations in Petri dishes | 24, 48, 72, 96, 120, 144, 168 hpf |
0, 0.1, 1, 2, 3, 4, 5, 10, 20, 30, 40, 50, 75, 100 mM | Developmental anomalies (heart rate, edema, tail distortion, hatching, mortality) observed via bright field microscopy. | Control embryos hatched normally; ≥10 mM caused early hatching with deformities and ≥40 mM increased mortality. | [72] |
| 25 embryos/well | Embryos exposed in 6-well plates with E3 medium supplemented with TZ | 3-4 h post-fertilization to 4 dpf | 0, 5, 10, 20, 50 g/L | Zebrafish embryo toxicity and vascular defects. | Dose-dependent vascular defects: hemorrhage, edema, small eye, vessel abnormalities. | [75] | |
| 20 | Exposed in E3 medium | 72 hours (hpf) | 5-100 mM (various concentrations) | Developmental and cardiac toxicity parameters | TZ caused dose-dependent drops in survival, hatching, cardiac/yolk sac edema, spinal defects, and heart rate. | [73] | |
| 9 | Exposure via aquatic media | 6 months to a year | 22 μM | Behavioral tests: T-maze test, cognitive flexibility, memory, learning, perseverance, consistency in choices. | Learning, memory, and flexibility impaired; task completion and perseverance reduced. | [76] | |
| 100 | Exposed to varying erythrosine and TZ levels in embryo water. | Up to 10 dpf | Erythrosine: 0.001–0.1%; TZ: 0.01–0.5% | Biochemical and genetic analyses | High TZ (≥0.5%) boosted hatching (55% at 48 hpf, 100% at 72 hpf) and triggered SOD1 expression via OS. | [74] | |
| Mice | |||||||
| KunMing mouse (20 ± 2 g) | 40 | Oral gavage | 30 days | 175–700 mg/kg body mass | Behavioral (Step-through, Morris maze) and biochemical tests | TZ negatively affects learning and memory in mice, increasing escape time and reducing reaction time in tests. | [48] |
| Male Swiss albino mice (4 weeks) | 15 | Oral administration | 72 days | 100 mg/kg | Hematological analyses | Increased red blood cell count, WBC count, and hemoglobin | [77] |
| 200 mg/kg | 200 mg/kg | Increased red and WBC counts, hemoglobin, platelet volume, and MCV | |||||
| 200 mg/kg | |||||||
| Swiss albino mice (25-30g) |
15 | Oral administration | 25 days | 200mg/kg | Physiological and biochemical analyses | No effect on cholesterol; increased bilirubin and creatinine | [78] |
| 400 mg/kg | Increased total cholesterol, triglycerides, bilirubin, and creatinine | ||||||
| Rats | |||||||
| Sprague–Dawley rats (70 ± 10 g) |
40 | Oral gavage | 30 days | 175, 350, 700 mg/kg body mass | Behavioral tests: Open-field test. Biochemical analyses |
TZ increases activity and anxiety in rats, also causing histopathological changes in the brain. | [48] |
| Male Wistar rats (40-50 g) | 45 | Dissolved in tap drinking water | 16 weeks | %, 1% (low dose) and 2.5 % (high dose) | Behavioral tests: Open field behaviour test Elevated plus maze test Light-Dark transition task Forced swim test Social interaction test |
The study highlights the harmful effects of TZ on anxiety and depression, highlighting the risks of long-term exposure to food dyes on mental health. | [79] |
| Young male albino rats (28 days old, 60-80g) | 18 | Oral gavage | 4 weeks | 320 mg/kg TZ in 1 ml distilled water, once daily. | Neurobiological and histological analysis: Brains were harvested and analyzed for histological changes. |
TZ has a neurotoxic effect, evidenced by histological changes such as neuronal apoptosis and vascular congestion. | [59] |
| White albino rats of either sex | 18 | Oral administration | 15 and 30 days | Low dose: 10 mg/kg High dose: 50 mg/kg |
Biochemical, hormonal and histological analyses | TZ disrupts glucose balance, damages pancreas, alters endocrine function. Increases glucose, lipase; decreases insulin, Ca, Mg |
[1] |
| Male albino rats | 18 | Oral administration | 8 weeks | 10 mg/kg (+3.75 mg/kg sulfanilic acid) | Biochemical and histological | Caused liver and kidney dysfunction with lesions. Increased cholesterol, triglycerides, LDL, VLDL, ALT, AST, ALP, bilirubin, creatinine, urea, uric acid. Decreased HDL, total protein. |
[61] |
| Wistar male albino rats | 50 | Diets containing dry mass | 90 days | 7.5 mg/kg | Biochemical and histological analyses | TZ raised lipids, liver enzymes, kidney function. Increased total cholesterol, triglycerides, LDL, ALT, AST, ALP, LDH. |
[30] |
| Female Wistar albino rats (225–250 g) | 40 | Oral gavage | 21 days | 500 mg/kg | Biochemical analyses and histopathological examinations | Increased AST, ALT, ALP indicating liver damage. | [63] |
| Male albino rats (65–80 g) | 12 | Oral administration | 7 weeks | 7.5 and 75 mg/kg | Biochemical and histopathological analyses | Study showed harmful lipid, biochemical changes and liver-kidney damage. Increased cholesterol, triglycerides, LDL, VLDL, ALT, AST, ALP, creatinine, urea, uric acid. |
[80] |
| Male Wistar albino rats (200–250 g) | 40 | Oral administration | 50 days | 7.5 mg/kg | Biochemical and histopathological analyses | TZ impaired liver/kidney, altered histology, lipids, glucose. Increased ALT, AST, ALP, GGT, urea, uric acid, creatinine, protein, cholesterol, triglycerides, LDL; decreased HDL. |
[81] |
| Adult female Wistar rats (225–250 g, 8–10 weeks old) | 40 | Oral gavage | 3 weeks | 500 mg/kg | Biochemical and histopathological analyses | TZ caused degenerative and metaplastic changes in ileum and colon epithelium. | [56] |
| Wistar albino rats (146–153 g) | 20 | Dissolved in 1 ml of distilled water | 30 days | 7.5 mg/kg b.wt. | Biochemical, histological and ultrastructural analyses | TZ raised AST, ALT, ALP, uric acid, urea, creatinine, reduced antioxidants, and caused liver and kidney damage. | [26] |
| Male Wistar rats (10–12 weeks old, 180–200 g) | 24 | Oral gavage | 6 weeks | 2, 6, 10 mg/kg (50:50 erythrosine-TZ) | Behavioral (open field test, forced swimming test, tail suspension test), biochemical and enzymatic analyses | Increased nitrite, TNF-α; worsened anxiety and depression. | [60] |
| Sprague-Dawley male albino rats (150-200 g) | 30 | Oral administration by orogastric gavage | 90 days | 75 mg/kg | Biochemical, genetic, immunohistochemical, histology analyses | Increased AST, ALT, urea, creatinine; liver and kidney damage. | [62] |
| Female Wistar albino rats (225–250 g) | 40 | Oral gavage | 21 days | 500 mg/kg | Biochemical and histopathological analyses | TZ caused kidney glomerular collapse, inflammation, congestion. | [82] |
| Albino rats (~0.2 kg) | 63 | Oral administration | 30 and 60 days | 7.5 mg/kg | Biochemical and histopathological analyses | TZdamages heart, raises nHDL and creatine kinase, increasing cardiovascular risk. | [83] |
| Male rats (10–15 weeks old, 190–250 g) | 18 | Oral administration | 30 days | 400 mg/kg | Biochemical analyses | Increased ALT, AST, ALP, urea, uric acid, creatinine; decreased Na, K, Ca. | [84] |
| Adult male Sprague Dawley albino rats | 30 | Oral administration | 90 days | 1.35 mg/kg | Hematological, immunological, and histopathological analyses | Decreased hemoglobin, RBC, PCV%, platelets; increased WBC, neutrophils, lymphocytes, monocytes. | [85] |
| Female Wistar rats (225–250 g) | 40 | Oral gavage | 3 weeks | 500 mg/kg | Biochemical and histopathological analyses | Increased total cholesterol, glucose, triglycerides, LDL, VLDL; decreased HDL. | [65] |
| Albino Wistar rats | 20 | Oral administration | 7 weeks | 75 mg/250 mL water 100 mg/250 mL water |
Biochemical, hematological and histopathological analyses | TZ damaged liver, kidneys, spleen; no change in cholesterol, triglycerides, ALT. Increased AST, creatinine, WBC, neutrophils, lymphocytes. |
[86] |
| Adult male Sprague Dawley rats (120–150 g) | 30 | Oral administration | 60 days | 200 mg/kg | Biochemical, histological and physiological analyses | Subchronic TZ affects liver and kidney parameters and induces OS. Increased ALT, AST, urea, total protein. |
[64] |
| Male and female Wistar rats (170–200 g) | 30 | Oral administration | 13 weeks |
5 mg/kg | Hematological and histopathological analyses | no effect | [87] |
|
7.5 mg/kg |
Decreased platelets; increased neutrophils, basophils, and mean platelet volume. | ||||||
| 10 mg/kg | no effect | ||||||
| Adult male albino rats (120–150 g) | 40 | Oral gavage | 30 days | 7.5 mg/kg bw |
Histopathological and Immunohistochemical analyses | TZ causes structural damage in cerebellum, glands, kidneys, with edema, congestion, neuron vacuolization, and cell deformation. | [88] |
| 15 mg/kg bw |
ChatGPT a spus: Edema, dilated perineural spaces, and degenerating Purkinje cells. |
||||||
| 100 mg/kg bw | Severe Purkinje cell degeneration, gray matter vacuolization, edema, nuclear pyknosis, vessel engorgement, increased astrocytes. |
||||||
| Male Wistar rats (130 ± 40 g) | 20 | Oral administration | 30 days | 300 mg | Biochemical and histopathological analyses | Increased transaminases, LDH, creatinine, uric acid, kidney proteins; decreased total protein, albumin, globulin; HDL unchanged. | [55] |
| Young male albino rats (Rattus norvegicus), 60–80 g | 36 | Oral administration | 30 days | Low dose: 15 mg/kg bw | Biochemical analyses | Increased ALT, AST, ALP, total protein, albumin, globulin, creatinine, urea; decreased serum cholesterol. | [54] |
| High dose: 500 mg/kg bw | Increased ALP, total protein, albumin, creatinine, urea | ||||||
ALP – Alkaline Phosphatase; ALT – Alanine Aminotransferase; AST – Aspartate Aminotransferase; b.wt. – Body weight; Ca – Calcium; dpf – Days Post Fertilization, GGT – Gamma-Glutamyl Transferase; HDL – High-Density Lipoprotein; hpf – Hours Post Fertilization; LDH – Lactate Dehydrogenase; LDL – Low-Density Lipoprotein; MCV – Mean Corpuscular Volume; MPV – Mean Platelet Volume; Na – Sodium; OS – Oxidative Stress; PCV% – Packed Cell Volume Percentage (hematocrit); RBC – Red Blood Cells; SOD1 – Superoxide Dismutase 1; TNF-α – Tumor Necrosis Factor-alpha; TZ – Tartrazine; VLDL – Very Low-Density Lipoprotein; WBC – White Blood Cells.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



