
Submitted:
31 July 2025
Posted:
04 August 2025
You are already at the latest version
Abstract
Secondary metabolites isolated from medicinal plants are of immense therapeutic, nutraceutical, and market importance. Unfortunately, natural biosynthesis of these compounds is generally low, dependent on factors that include the environment and physiology. Over the past few years, biotechnological innovation has opened up new and sustainable approaches to meet the requirement for secondary metabolite production in vitro. This review identifies recent biotechnological interventions such as plant tissue culture, elicitation, metabolic engineering, and synthetic biology that have been successfully utilized to enhance the biosynthesis of bioactive phytochemicals. An emphasis is given to the mechanisms, benefits, and disadvantages of these strategies, with specific reference to prominent medicinal plant species. Future directions are to interface omics tools with AI-powered metabolic pathway modeling for targeted metabolite improvement.
Keywords:
secondary metabolites
; medicinal plants
; tissue culture
; elicitation
; metabolic engineering
; synthetic biology
; hairy root
; plant biotechnology
1. Introduction
Medicinal plants have been employed for thousands of years in conventional as well as contemporary medicine because they possess a rich treasure trove of bioactive secondary metabolites like alkaloids, flavonoids, terpenoids, and phenolics (Twaij and Hasan, 2022; Rauf et al., 2025). These metabolites are synthesized through complex biosynthetic pathways, often at low levels. Environmental stress, growth stage, and genetics play a huge role in their accumulation. For these reasons, an increasing interest exists in applying biotechnological means to overcome these challenges and provide sustainable production (Figure 1).
1.1. Role of Plant Tissue Culture
Plant tissue culture (Karuppusamy, 2009; Fatima et al., 2023 is an exemplary technology that offers a platform in controlled conditions for secondary metabolite production (Nordine, 2025). Platforms most frequently used are callus and suspension cultures, shoot cultures, and hairy root cultures (Liu et al., 2025) (Figure 2).
- Callus and Cell Suspension Cultures: Sufficient for the production of uniform biomass. e.g., Taxus spp. cell cultures for the production of paclitaxel (Tomilova et al., 2023).
- Hairy Root Cultures: They are obtained from Agrobacterium rhizogenes and have genetic stability and high biosynthetic potential. Withania somnifera and Rauwolfia serpentine are a few among them (Singh et al., 2022; Srivastava et al., 2016).
- Shoot/Root Organ Cultures: Sustain site-specific biosynthesis, which generally yields more than undifferentiated cells (Kikowska et al., 2025; Sánchez-López et al., 2025).
1.2. Elicitation Strategies
Elicitors are biotic or abiotic stress-inducing compounds that induce the formation of secondary metabolites by triggering plant defense mechanisms (Khan et al., 2025; Malu et al., 2025).
- Biotic Elicitors: Consist of fungal or bacterial cell wall materials e.g., chitosan (Javed et al., 2025), yeast extract (Lescano et al., 2025).
- Abiotic Elicitors Salicylic acid (Rithichai et al., 2024), jasmonic acid (Rasheed et al., 2017 silver nanoparticles (Bernela et al.,2023; Verma et al., 2024 and UV light are some of them.
These stimuli trigger cascades of signaling and transcription factors that upregulate the essential biosynthetic genes.
1.3. Precursor Feeding
The major metabolic precursors as Carbohydrates (Mukherjee et al., 2025) serve as basic inputs for the plant’s biosynthesis of varying secondary metabolites (Figure 3).
Supplementation of biosynthetic precursors on cultures, e.g., shikimic acid, mevolunic acid, phenylalanine (Delfi et al., 2025) has been proven to boost the yields of metabolites by controlling the Pathways of the metabolites (Figure 3).
1.4. Metabolic
Metabolic refers to the alteration of genes that code for key enzymes in a pathway. Other approaches that have been used are:
- Overexpression of pathway-specific genes (e.g., PAL, CHS). Research has also been found to reveal that Salicylic acid and Gibberellic acid induce the expression of the phenylalanine ammonia-lyase (PAL) synthesis genes responsible for an increase in phenolics (e.g., rosmarinic acid) and falvanoid as well in Salvia officinalis ( Moreira et al., 2022).
- Silencing of competitive pathways. The sterol pathway is a competitive pathway of artemisinin biosynthesis in A. annua. Four competitive branch pathway genes β-caryophyllene synthase gene (CPS), β-farnesene synthase gene (BFS), germacrene A synthase gene (GAS) and SQS were further down-regulated independently by the antisense method in A. annua. The content of artemisinin and dihydroartemisinic acid (DHAA) were increased significantly in different transgenic lines.
- Engineering transcription factors to co-upregulate multiple genes (Shi et al., 2024) (Figure 5).
Synthetic biology tools like CRISPR/Cas and synthetic promoters provide unadulterated control of metabolite flux (Gao et al., 2025) but along synthetic promoters, offer precise control over metabolite flux as GABA, with hypotensive properties enhanced in Tomato fruit and rice grains using this technology (Das et al., 2024)
1.5. Omics and Systems Biology Approaches
Multiomics integration i.e., Genomics, Transcriptomics, proteomics, and metabolomics give information about the regulatory networks (Figure 4) of secondary metabolism (Karalija et al., 2025; Wu et al., 2025).
This gives a picture of how the confluence of genomics, transcriptomics, proteomics, metabolomics, and single-cell omics allows deciphering and control of specialized metabolic processes in medicinal plants. Despite integration offering a general view of secondary metabolite biosynthesis, primary bottlenecks include handling data, paucity of pathway information, dynamic nature of metabolism, and impossibility of easy translation of complicated information. To counter these, solutions include high-end bioinformatics platforms, real-time omics techniques, synthetic biology integration, and high-performance data analysis software. In all, there is a need for multi-omics integration to enhance medicinal compound discovery and production (Liu et al., 2025b).
Multi-omics data integration facilitates:
- Selection of critical metabolic bottlenecks.
- Marker-assisted selection for cell lines with high yields.
- In silico modeling of biosynthetic networks for targeted intervention.
1.6. Limitations and Challenges
- Somaclonal variation in long-term cultures.
- Problems of scalability for commercial cultivation.
- Regulatory problems with genetically modified products.
- Limited knowledge of intricate metabolic controls in non-model plants (Dsouza et al., 2025; Karalija et al., 2025; Singh et al., 2025).
2. Methodology
We reviewed using the systematic literature analysis approach to gather, analyze, and synthesize ongoing research on biotechnological interventions in the increase of secondary metabolite production in medicinal plants. The steps involved in the methodology were:
2.1. Literature Search Strategy
A comprehensive literature search of peer-reviewed research papers, review articles, and conference papers was carried out using the following databases:
- PubMed
- ScienceDirect
- Scopus
- Google Scholar
- SpringerLink
The search covered journals from the period January 2010 to July 2025 but accentuating recent and most impactful developments post-2015. One article citation from bioRxiv repository on Artemsia database for revealing mul-tiomics research secrets was included (https://artemisia-db.com/).
2.2. Search Terms and Keywords
For seeking relevant articles, Boolean combinations of the following keywords were utilized:
- “Secondary metabolites”
- “Medicinal plants”
- “Plant tissue culture”
- “Hairy root culture”
- “Elicitation”
- “Metabolic engineering”
- “Synthetic biology”
- “Biotechnological enhancement”
- “Plant-derived compounds”
- “Omics approaches in plants”
2.3. Inclusion and Exclusion Criteria
Inclusion Criteria:
- Articles published in the English language.
- functor web Studies that report experimental or review data on biotechnological enhancement of secondary metabolites.
- functor web Research on plant in vitro culture methods (e.g., callus, hairy root, suspension culture).
- functor web Articles discussing elicitor-based, molecular, or omics-level intervention.
Exclusion Criteria:
- Studies not focused on medicinal plants.
- Articles with poor methodological description.
- Descriptions of primary metabolites or non-target plant species.
- Articles in non-peer-reviewed journals or without available full texts.
2.4. Organization and Data Extraction
Relevant data were categorized and grouped based on:
- Type of biotechnological intervention (e.g., elicitation, genetic modification, tissue culture)
- Plant species studied
- Targeted enhanced metabolite(s)
- Mechanism or pathway targeted
- Experimental outcome and yield improvements
Tables and figures were prepared to integrate interventions, mark interesting studies, and compare improvement strategies across plant systems.
2.5. Review Process and Quality Evaluation
Each article was critically examined for:
- Scientific quality and reproducibility
- Novelty in methodology
- Relevance to upgrading of pharmaceutically important compounds
Only well-documented, high-quality studies were used to give the review reliability.
2.6. Synthesis of Results
The information was synthesised thematically and classified into:
- In vitro tissue culture techniques
- Elicitor-based improvement
- Synthetic biology and metabolic engineering
- Indirect tissue culture strategies (miRNA, Ploidy)
- Omics and systems biology platforms
The final draft of the review was prepared to paint a homogeneous and coherent picture of the existing status and future prognosis of biotechnological interventions in this sector.
3. Results
A meta-analysis of more than 100 peer-reviewed articles demonstrated a broad range of biotechnological interventions effectively used to improve secondary metabolite yield in medicinal plants. The results have been grouped into major intervention strategies: in vitro tissue culture, elicitation, metabolic engineering, and omics-based strategies. The rundown of significant outcomes is given below.
3.1. Tissue Culture-Based Enhancement
Several plant species showed enhanced metabolite production through callus, suspension, or hairy root cultures (Table 1):
3.2. Elicitor-Mediated Enhancement
Elicitors like jasmonic acid (JA), salicylic acid (SA), yeast extract, and silver nanoparticles (AgNPs) showed an excellent effect on the secondary metabolite biosynthesis (Table 2).
3.3. Genetic Modification and Metabolic Engineering
Metabolic pathway engineering by overexpression of dominant biosynthetic genes and inhibition of unwanted pathways resulted in remarkable enhancement of metabolite accumulation (Table 3):
3.4. Indirect Methods of Plant Tissue Culture Use
3.4.1. MicroRNA (amiRNA, eTMs)
Plant microRNAs (miRNAs), a family of roughly 21-nucleotide-long small noncoding RNAs (ncRNAs), typically function as important regulators of their target genes by directing mRNA cleavage or translational repression (Jiang et al., 2021; Ghosh et al., 2024). In contrast, miRNAs are directly implicated in the biosynthesis regulation of Secondary metabolites like alkaloids, flavonoids, and terpenes that are responsible for the therapeutic activity (Yang et al., 2025).
3.4.2. Ploidy Engineering
In medicinal plants, secondary metabolite production per unit of biomass has vast economic importance. Therefore, doubling the genome of cells promotes genome multiplication, protein synthesis increase, and secondary metabolite yield increase (Ghosh et al., 2024).
3.5. Omics and Systems Biology Contributions
Omics data integration (transcriptomics, proteomics, metabolomics) gave information about pathway regulation e.g., biosynthesis of artemsinin (Shi et al., 2024) in A. annua (Figure 5) and revealed new targets for intervention. The use of CRISPR/Cas9 and other gene-editing technologies may enable the metabolic engineering of medicinal plants for boosting specialized metabolite production Integrated multi-omics offers a promising route for boosting our comprehension of interactions among genes, proteins, and metabolites, potentially solving for new specialized metabolites of therapeutic interest (Shi et al., 2024; Wang et al., 2024).
4. Discussion
The improvement of secondary metabolite production in medicinal plants is a center of attention in plant biotechnology because of the worldwide demand for natural therapeutic products. The paper has highlighted the wide range of biotechnological interventions ranging from multiomics inetgartion with in vitro culture systems to elicitor applications and genetic engineering interventions that have been proved effective in improving yield and quality of bioactive compounds (Karalija et al., 2025; Shi et al., 2024; Wang et al., 2024).
In vitro culture systems, such as callus culture, cell suspension, hairy roots, and organ cultures, provide controlled and scalable systems for metabolite production, irrespective of environmental oscillations. Other studies, such as those on Withania somnifera, Artemisia annua, and Taxus spp., have recorded significant fold increases in the accumulation of secondary metabolites under optimized culture conditions.These platforms are also useful assets in biosynthetic pathway and regulatory process research (Al-Khayri et al., 22; Fett-Neto & DiCosmo, 2020; Haider & Ghosh, 2024; Zhao et al., 2023).
The use of elicitors, such as jasmonic acid, salicylic acid, and yeast extract, has been especially effective in eliciting stress-like reactions in plant cells, thus upregulating the genes of interest associated with secondary metabolite biosynthesis. For instance, the elicitation of Catharanthus roseus with jasmonic acid has been reported to promote vincristine and vinblastine production, both of which possess pivotal anticancer activity. The above results highlight the prospect of elicitation as an affordable and scalable means of boosting metabolite production (Lala et al., 2021; Kochan et al., 2017; Rasheed et al., 2017; Rithichai et al., 2024).
Genetic manipulation has also propagated this discipline by allowing for targeted manipulation of biosynthetic genes and regulatory transcription factors. Methods such as overexpression of chalcone synthase in Glycyrrhiza uralensis or co-expression of ORCA3 and MYC2 transcription factors in Catharanthus roseus have led to the immense upregulation of flavonoids and terpenoid indole alkaloids, respectively. These therapies not only increase metabolite quantity but also increase the reproducibility and stability of the production (Paul et al., 2017; Yin et al., 2020; Zhao et al., 2022; Zunun-Pérez et al., 2017).
4.1. Refinement of Tissue Culture Based
The data presented (Table 1) exhibit very clearly the potential of biotechnological in vitro culture methods to augment greatly the yield of valuable secondary metabolites from medicinal plants. A major antimalarial metabolite, artemisinin, was increased 7.8 times by organ cultures of Artemisia annua, demonstrating the possibility of tissue-specific growth under controlled conditions for metabolite enhancement. Concurrently, callus culture of Rauwolfia serpentina reflected a 2-fold enhancement in reserpine content, demonstrating the capability of dedifferentiated plant tissues to serve as biofactories for alkaloid production. Cell suspension culture of Taxus brevifolia and T. cuspidata are notable in having reached levels of commercial paclitaxel production, an anticancer drug of efficacy, demonstrating the scalability of this technology. Besides, Withania somnifera hairy root cultures established through Agrobacterium rhizogenes-mediated transformation yielded a remarkably 11.49-fold greater withanolide accumulation, underscoring the relevance of Agrobacterium rhizogenes-mediated transformation in high-yielding and stable metabolite biosynthesis platforms. Together, these examples support the applicability of in vitro biotechnologies as effective and sustainable alternative means to classical harvesting from natural plant populations for conservation and development of pharmaceutical compounds (Karalija et al., 2025; Shi et al., 2024; Wang et al., 2024).
4.2. Elicitor Mediated Enhancement
The results (Table 2) reveal the heterogeneous and high-order impacts of various elicitors on inducing secondary metabolite biosynthesis in medicinally important plants, exemplifying a planned strategy towards plant biotechnology. Salicylic acid of the tested elicitors showed a very high stimulation with a 282.96-fold increase in eugenol production in Ocimum sanctum (Rithichai et al., 2024), suggesting intensive activation of the phenylpropanoid pathway. In the same way, the biotic elicitor cellulase caused an 8.6-fold enhancement of glycyrrhizin content in Glycyrrhiza glabra, showing its function in induction of metabolic flow possibly through cell wall breakdown and increased precursor supply (Srivastava et al., 2019). Silver nanoparticles (AgNPs), as abiotic nanomaterial-based elicitors, enhanced essential oil production by 2.3 times in Achillea millefolium, demonstrating their role in metabolite accumulation in response to oxidative stress (Lala et al., 2021). Conversely, yeast extract, an intricate biotic elicitor, increased ginsenosides moderately by 1.57-fold in Panax ginseng (Kochan et al., 2017), pointing to its impact on terpenoid biosynthetic pathways. Surprisingly, jasmonic acid, well known for its role in secondary metabolism regulation, modestly enhanced vinblastine and vincristine content in Catharanthus roseus by only 0.6× (Rasheed et al., 2017), indicating that the efficiency of elicitors tends to be species- and pathway-specific. These observations individually highlight the promise of elicitation strategies to maximize metabolite productivity, albeit with effectiveness being strongly associated with the plant species, elicitor dose, and exposure regime (Lala et al., 2021; Kochan et al., 2017; Rasheed et al., 2017; Rithichai et al., 2024).
4.3. Metabolic Engineering and Genetic Modification
The information (Table 3) reveals a variety of genetic and signaling-driven engineering tools aimed at the enhancement of secondary metabolite production in medicinal plants, emphasizing the precision and efficacy of manipulation of metabolic pathways. Among the key strategies is gene co-expression, e.g., HMGR (3-hydroxy-3-methylglutaryl-CoA reductase) and ADS (amorpha-4,11-diene synthase) overexpression simultaneously in Artemisia annua, which showed 8.65-fold increased artemisinin yield (Zhao et al., 2022). This notable enhancement reflects precursor supply (via HMGR) and step-specific catalysis (via ADS) strategic enhancement in terpenoid biosynthesis.
Likewise, co-expression of ADS with FPPS (farnesyl diphosphate synthase) and FPPS with CYP71AV1(cytochrome P450 monooxygenase) and CPR (cytochrome P450 reductase) also augmented artemisinin yields 2.6× and 3.6×, respectively, demonstrating the significance of synergistically enhancing both precursor production and downstream oxidative processes (Zhao et al., 2022).
In Catharanthus roseus, co-expression of transcription factors MYC2 and ORCA3 activated the terpenoid indole alkaloid (TIA) pathway, resulting in enhanced biosynthesis of anticancer metabolites like vincristine and ajmalicine (Paul et al., 2017). This is a sign of the potential of the transcription factors to control the various biosynthetic genes globally and therefore are hopeful targets for general metabolic enhancement.
As a second example, CHS overexpression in Glycyrrhizia uralensis activated flavonoid production, demonstrating the efficacy of rate-limiting enzyme targeting within the phenylpropanoid pathway (Yin et al., 2020). Furthermore, treatment with salicylic acid and hydrogen peroxide in Capsicum annuum elicited phenylalanine ammonia-lyase (PAL) activity, leading to enhanced levels of capsiate, suggesting that signaling molecules could regulate defense pathways to promote metabolite accumulation (Paul et al., 2017; Yin et al., 2020; Zhao et al., 2022; Zúnún-Pérez et al., 2017).
Together, these findings demonstrate that gene stacking, transcription factor manipulation, and induction of the signaling are robust pathway engineering tools that can be modulated to overproduce particular kinds of secondary metabolites in a species- and pathway-dependent manner.
4.4. Ploidy Engineering
The information (Table 4) illustrates the effect of agents inducing polyploidy mainly colchicine and oryzalin on the production of secondary metabolites in medicinal plants. Polyploidy, through doubling the chromosomes, tends to result in increased gene dosage, increased nuclear content, and metabolic changes, which can activate biosynthetic pathways. For instance, induction of tetraploidy in Ajuga reptans with oryzalin increased 20-hydroxecdysone production, a major phytoecdysteroid (Navrátilová et al., 2022; Svecarova et al., 2019). Likewise, colchicine treatment doubled Catharanthus roseus anticancer alkaloid vincristine yields (Fathimunnissa, 2011), and increased 4-fold bacoside deposition in Bacopa monnieri (Kharde et al., 2017), which indicates that autotetraploidy can immensely increase secondary metabolite yields in certain species. However, not all the events were successful Artemisia annua and Echinacea purpurea showed suboptimal or slight increments in artemisinin, cichoric acid, and chlorogenic acid after colchicine-induced tetraploidy (Lin et al., 2011; Abdoli et al., 2013), It described the response as species-specific and metabolite-specific. Further, colchicine-induced polyploidy in Linum album increased podophyllotoxin production by 1.39× (Javadian et al., 2017), and, thus indicated potential in the optimization of lignan pathways. These findings in general indicate that although polyploidy is a useful approach for metabolite improvement, its efficacy is plant genotype-, target compound-, and regulatory network complexity-dependent (Navrátilová et al., 2022; Svecarova et al., 2019; Fathimunnissa, 2011; Kharde et al., 2017; Lin et al., 2011; Abdoli et al., 2013; Javadian et al., 2017).
4.5. Engineering Strategies to Enhancing Artemisinin Synthesis in Artemisia annua
Artemisinin, a sesquiterpene lactone that has been purified from Artemisia annua, is in huge market demand because of its strong antimalarial effect (Yuan et al., 2023). The integrated model for the biosynthesis pathway of artemisinin in Artemisia annua (Figure 5), showing the ubiquitous nature of transcription factors (TFs) to regulate the crucial enzymic steps.The biosynthesis is started by photosynthesis-driven glucose production that is diverted along the mevalonate (MVA) and methylerythritol phosphate (MEP) pathways to produce the precursors IPP (isopentenyl diphosphate) and DMAPP (dimethylallyl diphosphate). These are then decreased by farnesyl diphosphate synthase (FPS) to become FPP (farnesyl diphosphate) the universal substrate of sesquiterpenoids (Hassani et al., 2020).
The first committed step of artemisinin biosynthesis is catalyzed by amorphadiene synthase (ADS), which produces amorpha-4,11-diene from FPP. The intermediate then goes through a series of oxidative and reductive conversions by cytochrome P450 enzyme CYP71AV1, which converts it sequentially into artemisinic alcohol, artemisinic aldehyde, and ultimately to dihydroartemisinic aldehyde by the action of DBR2 (double bond reductase 2). Secondary conversion by ALDH1 (aldehyde dehydrogenase 1) produces dihydroartemisinic acid, which, through spontaneous photooxidation, produces artemisinin in glandular trichomes. Four genes in artemisinin-specific biosynthetic pathway namely ADS, CYP71AV1, DBR2 and ALDH1 were expressed at greater levels in MeJasmonic acid/JA treatment (Hassani et al., 2020; Yan et al., 2017; Xie et al., 2021; Xiang et al., 2015).
Figure 5.
Simplified Integrating of Transcription Factor Dynamics into Artemisinin Biosynthesis Models in A. annua (Modified from Shi et al., 2024) Note: The blue boxes show enzymes, JA=Jasmonic acid and SA= Salicylic acid, ADS= amorpha-4,11-diene synthase, ALDH1= aldehyde dehydrogenase 1, CYP71AV1= cytochrome P450-dependent hydroxylase, DBR2=double bond reductase 2, DMAPP= dimethylallyl diphosphate, FPP= farnesyl diphosphate, FPS= farnesyl diphosphate synthase, IP=, isopentenyl diphosphate, IPPI= isopentenyl diphosphate isomerase, MVA= mevalonate; MEP, methyl-D-erythritol 4-phosphate. Notably, this number highlights the pivotal regulatory function of transcription factors, portrayed as red arrows that affect almost every enzymatic step.
Figure 5.
Simplified Integrating of Transcription Factor Dynamics into Artemisinin Biosynthesis Models in A. annua (Modified from Shi et al., 2024) Note: The blue boxes show enzymes, JA=Jasmonic acid and SA= Salicylic acid, ADS= amorpha-4,11-diene synthase, ALDH1= aldehyde dehydrogenase 1, CYP71AV1= cytochrome P450-dependent hydroxylase, DBR2=double bond reductase 2, DMAPP= dimethylallyl diphosphate, FPP= farnesyl diphosphate, FPS= farnesyl diphosphate synthase, IP=, isopentenyl diphosphate, IPPI= isopentenyl diphosphate isomerase, MVA= mevalonate; MEP, methyl-D-erythritol 4-phosphate. Notably, this number highlights the pivotal regulatory function of transcription factors, portrayed as red arrows that affect almost every enzymatic step.
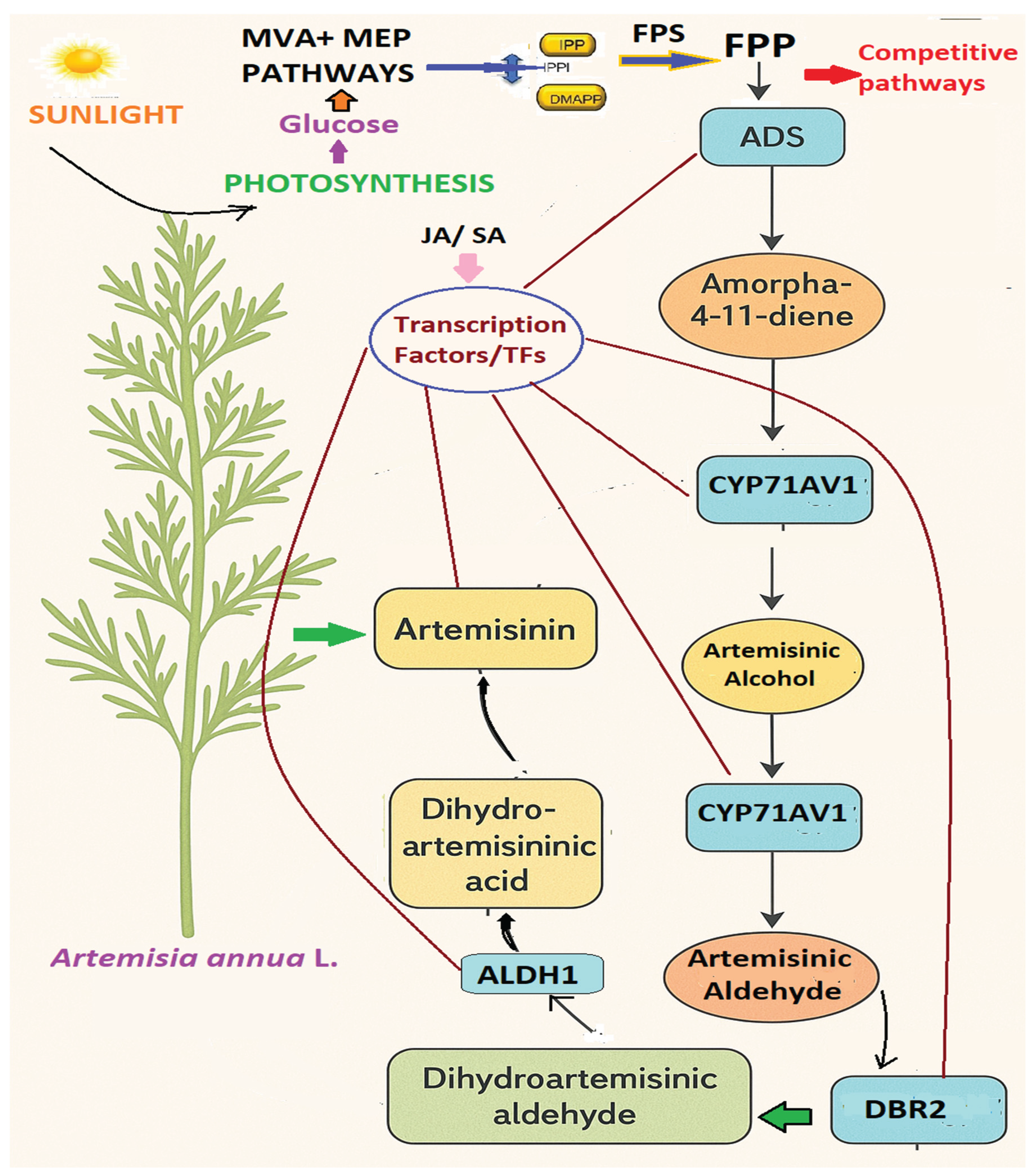
TFs regulate the level of structural genes for enzymes like ADS, CYP71AV1, DBR2, and ALDH1, thus fine-tuning the metabolic flow in the pathway.
Environmental conditions such as sunlight also have a crucial role, both in initiating photosynthesis and in the terminal non-enzymatic photooxidation step to produce artemisinin. The research result (Figure 6) of Yuan et al., (2023) illustrates that Jasmonic acid (JA) is a major elicitor, triggering transcription factors signaling cascade like AabHLH112, AaERF1, and AabHLH113, which indirectly regulate the expression of core biosynthetic genes like ADS, CYP71AV1, DBR2, and ALDH1 upregulate encoded upstream enzymes. Jasmonic acid induces the activation of transcription factors AabHLH112 and AaERF1, which induce the upregulation of upstream ADS and CYP71AV1 enzymes catalyzing early steps from farnesyl diphosphate to artemisinic aldehyde. At the same time, AabHLH112 interact AabHLH113, which control DBR2 and ALDH1 enzymes catalyzing conversion of artemisinic aldehyde to dihydroartemisinic acid, a direct intermediate of artemisinin. This two-pathway convergence emphasizes the transcriptional control sites necessary for metabolic engineering for increasing The biosynthesis of artemisinin A useful antimalarial sesquiterpene lactone is a highly regulated Jasmonic acid(JA)-responsive metabolic process in Artemisia annua (Rasheed et al., 2017; Xiang et al.,2019; Yuan et al., 2025).
These differential patterns reveal spatial regulation of artemisinin pathway genes and can guide tissue-targeted metabolic engineering. To further explore gene annotations, expression matrices, and visualization tools, the Artemisia Database is an exhaustive platform for transcriptomic analysis and functional genomics.
These differential patterns based on Artmesia database heatmap (Figure 7) reflect the spatial regulation of artemisinin pathway genes in five organs of Artemisia annua (Leaf, Petiole, Flower, Root, Stem) and have the potential to inform tissue-targeted metabolic engineering strategies. Of particular note, mikado.chr4G1338 (ADS) and mikado.chr3G1536 (CYP71AV1) are found to have maximum expression intensities in leaf and flower tissues, followed by stem and petiole, as highlighted by the red coloration. The heatmap scale indicates expression values from 0 (dark blue, lowest) to 3 (red, highest). The Gene IDs and their respective gene names are mikado.chr4G1338=ADS, mikado.chr3G1536=CYP71AV1, mikado.chr2G1956=DBR2, and mikado.chr8G609=ALDH1. For more exploration of gene annotations, expression matrices, and visualization tools, the Artemisia Database offers an extensive platform for transcriptomic analysis and functional genomics (https://artemisia-db.com/).
Therefore, this combined model not only traces out the metabolic pathway but also captures the intricate interaction between metabolic enzymes and transcriptional regulation, providing a useful platform for metabolic engineering approaches to boost artemisinin productivity in A. annua (Kumari et al., 2025;Taheri et al., 2025).
5. Future Prospects
The future of the production of secondary metabolites from medicinal plants is in precision bioengineering, multi-omics-driven design, and scalable and sustainable production platforms. With responsible innovation and interdisciplinary collaboration, there is the potential for transforming natural product biosynthesis to respond to the increasing global need for safe, effective, and affordable plant-derived therapeutics. Future approaches such as AI-assisted metabolic flux analysis, genome editing, and machine-learning models have the potential to revolutionize secondary metabolite enhancement. Combination with vertical farming and bioreactors can result in commercial-scale production with low environmental impact.
6. Conclusions
Biotechnological methods offer not just a practical replacement for conventional extraction techniques but also an environmentally friendly and scalable means to fulfilling international needs for plant-based therapeutics. DNA Biotechnological applications offer promising, sustainable, and scalable methods towards enhancing secondary metabolites in medicinal plants. Continued advancement of molecular biology, systems biology, and synthetic biology will give the foundation for efficient production of such vital compounds to meet global pharmaceutical and nutraceutical demands. Additional interdisciplinarity research and technological fusion will be the key to fully exploiting the potential of medicinal plants during the 21st century.
Acknowledgments
We are very much thankful to Farooq, Agriculture Research Institute, University of Agriculture, North, Mingora, Khyber Pukhtunkhwa, Pakistan. Special thanks to Shah Meer Ali Khan (BS Computer Science, National College of Business Administration & Ecnomics, Lahore, Punjab, Pakistan).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Abdoli, M., Moieni, A., & Naghdi Badi, H. (2013). Morphological, physiological, cytological and phytochemical studies in diploid and colchicine-induced tetraploid plants of Echinacea purpurea (L.). Acta Physiologiae Plantarum, 35(7), 2075-2083. https://link.springer.com/article/10.1007/s11738-013-1242-9. [CrossRef]
- Al-Khayri, J. M., Sudheer, W. N., Lakshmaiah, V. V., Mukherjee, E., Nizam, A., Thiruvengadam, M., M., Nagella, P., Alessa, F.M., Al-Mssallem, M.Q., Rezk, A.A. and Shehata, W.F., & Attimarad, M. (2022). Biotechnological approaches for production of artemisinin, an anti-malarial drug from Artemisia annua L. Molecules, 27(9), 3040. https://www.mdpi.com/1420-3049/27/9/3040. [CrossRef]
- Das, S., Kwon, M., & Kim, J. Y. (2024). Enhancement of specialized metabolites using CRISPR/Cas gene editing technology in medicinal plants. Frontiers in plant science, 15, 1279738. https://www.frontiersin.org/journals/plant-science/articles/10.3389/fpls.2024.1279738/full. [CrossRef]
- Delfi, S. A., Putri, S. I., Santoso, P., & Idris, M. (2025). Twenty-Five Years Research on Micropropagation of Stevia and Curcuma sp. and Improving Secondary Metabolites using Precursor-elicitor in vitro: A Review. Jurnal Biologi Tropis, 25(2), 1921-1940. [CrossRef]
- Dsouza, A., Dixon, M., Shukla, M., & Graham, T. (2025). Harnessing controlled-environment systems for enhanced production of medicinal plants. Journal of Experimental Botany, 76(1), 76-93. [CrossRef]
- Fathimunnissa Begum, F. B. (2011). Augmented production of vincristine in induced tetraploids of Agrobacterium transformed shooty teratomas of Catharanthus roseus. Medicinal Plants, International journal of Phytomedicines and Related Industries. 3(1).59-64. https://www.cabidigitallibrary.org/doi/full/10.5555/20123177175, https://www.indianjournals.com/ijor.aspx?target=ijor:mpijpri&volume=3&issue=1&article=008. [CrossRef]
- Fatima, I., Akram, M., Mukhtar, H., Gohar, U. F., Sajid, Z. A., & Hameed, U. (2023). Tissue Culture of Medicinal Plants. In Essentials of Medicinal and Aromatic Crops. 1-32. Cham: Springer International Publishing. https://link.springer.com/chapter/10.1007/978-3-031-35403-8_1.
- Fett-Neto, A. G., & DiCosmo, F. (2020). Production of paclitaxel and related taxoids in cell cultures of Taxus cuspidata: perspectives for industrial applications. In Plant Cell Culture Secondary Metabolism Toward Industrial Application, 139-166. CRC Press. https://www.taylorfrancis.com/chapters/edit/10.1201/9780138743208-6/production-paclitaxel-related-taxoids-cell-cultures-taxus-cuspidata-perspectives-industrial-applications-arthur-fett-neto-frank-dicosmo.
- Gantait, S., Das, S., Mahanta, M., & Banerjee, M. (2023). Influence of in vitro culture age on morphology, antioxidant activities, reserpine production, and genetic fidelity in Indian snakeroot (Rauvolfia serpentina (L.) Benth. ex Kurz.). South African Journal of Botany, 162, 864-872. https://www.sciencedirect.com/science/article/abs/pii/S0254629923006002. [CrossRef]
- Gao, H., Pei, X., Song, X., Wang, S., Yang, Z., Zhu, J., Lin, Q., Zhu, Q., & Yang, X. (2025). Application and development of CRISPR technology in the secondary metabolic pathway of the active ingredients of phytopharmaceuticals. Frontiers in Plant Science, 15, 1477894. https://www.frontiersin.org/journals/plant-science/articles/10.3389/fpls.2024.1477894/full. [CrossRef]
- Halder, T., & Ghosh, B. (2024). Withania somnifera (L.) Dunal: Enhanced production of withanolides and phenolic acids from hairy root culture after application of elicitors. Journal of Biotechnology, 388, 59-71. https://doi.org/10.1016/j.jbiotec.2024.04.010, https://www.sciencedirect.com/science/article/abs/pii/S0168165624001093. [CrossRef]
- Hassani, D., Fu, X., Shen, Q., Khalid, M., Rose, J. K., & Tang, K. (2020). Parallel transcriptional regulation of artemisinin and flavonoid biosynthesis. Trends in plant science, 25(5), 466-476. https://www.cell.com/trends/plant-science/abstract/S1360-1385(20)30015-7?dgcid=raven_jbs_etoc_email.
- Javadian, N., Karimzadeh, G., Sharifi, M., Moieni, A., & Behmanesh, M. (2017). In vitro polyploidy induction: changes in morphology, podophyllotoxin biosynthesis, and expression of the related genes in Linum album (Linaceae). Planta, 245(6), 1165-1178. https://link.springer.com/article/10.1007/s00425-017-2671-2. [CrossRef]
- Jiang, S., Cui, J. L., & Li, X. K. (2021). MicroRNA-mediated gene regulation of secondary metabolism in plants. Critical Reviews in Plant Sciences, 40(6), 459-478. https://www.tandfonline.com/doi/abs/10.1080/07352689.2022.2031674. [CrossRef]
- Kajla, M., Roy, A., Singh, I. K., & Singh, A. (2023). Regulation of the regulators: transcription factors controlling biosynthesis of plant secondary metabolites during biotic stresses and their regulation by miRNAs. Frontiers in Plant Science, 14, 1126567. https://www.frontiersin.org/journals/plant-science/articles/10.3389/fpls.2023.1126567/full. [CrossRef]
- Karalija, E., Macanović, A., & Ibragić, S. (2025). Revisiting Traditional Medicinal Plants: Integrating Multiomics, In Vitro Culture, and Elicitation to Unlock Bioactive Potential. Plants, 14(13), 2029. https://pmc.ncbi.nlm.nih.gov/articles/PMC12252194/. [CrossRef]
- Karuppusamy, S. (2009). A review on trends in production of secondary metabolites from higher plants by in vitro tissue, organ and cell cultures. J Med Plants Res, 3(13), 1222-1239. https://academicjournals.org/article/article1380530836_Karuppusamy.pdf.
- Khan, A., Kanwal, F., Ullah, S., Fahad, M., Tariq, L., Altaf, M. T., Riaz, A., & Zhang, G. (2025). Plant secondary metabolites—Central regulators against abiotic and biotic stresses. Metabolites, 15(4), 276. https://www.mdpi.com/2218-1989/15/4/276. [CrossRef]
- Kharde, A. V., Chavan, N. S., Chandre, M. A., Autade, R. H., & Khetmalas, M. B. (2017). In vitro enhancement of bacoside in brahmi (Bacopa monnieri) using colchicine. J plant biochem physiol, 5(1), 1-6. https://www.researchgate.net/profile/Autade-Haribhau/publication/312165383_In_Vitro_Enhancement_of_Bacoside_in_Brahmi_Bacopa_monnieri_Using_Colchicine/links/5873afdf08ae8fce4924bc61/In-Vitro-Enhancement-of-Bacoside-in-Brahmi-Bacopa-monnieri-Using-Colchicine.pdf?origin=journalDetail&_tp=eyJwYWdlIjoiam91cm5hbERldGFpbCJ9.
- Kikowska, M., Urbańska, M., Hermosaningtyas, A., Gornowicz-Porowska, J., Budzianowska, A., Siwulski, M., Blicharska, E., & Thiem, B. (2025). Biotechnological stimulation of phenolics in Lychnis flos-cuculi shoot cultures. Plant Cell, Tissue and Organ Culture (PCTOC), 162(1), 2. https://link.springer.com/article/10.1007/s11240-025-03117-z. [CrossRef]
- Kochan, E., Szymczyk, P., Kuźma, Ł., Lipert, A., & Szymańska, G. (2017). Yeast extract stimulates ginsenoside production in hairy root cultures of American ginseng cultivated in shake flasks and nutrient sprinkle bioreactors. Molecules, 22(6), 880. https://www.mdpi.com/1420-3049/22/6/880. [CrossRef]
- Kumari, S., Rai, N., Singh, S., Saha, P., Bisen, M. S., & Pandey-Rai, S. (2025). Functional identification of AaDREB-9 transcription factor in Artemisia annua L. and deciphering its role in secondary metabolism under PEG-induced osmotic stress. Plant Physiology Reports, 1-12. https://link.springer.com/article/10.1007/s40502-025-00857-0. [CrossRef]
- Lescano, L., Cziáky, Z., Custódio, L., & Rodrigues, M. J. (2025). Yeast extract elicitation enhances growth and metabolite production in Limonium algarvense callus cultures. Plant Cell, Tissue and Organ Culture (PCTOC), 160(2), 45. https://link.springer.com/article/10.1007/s11240-025-02991-x. [CrossRef]
- Lin, X., Zhou, Y., Zhang, J., Lu, X., Zhang, F., Shen, Q., Q., Wu, S., Chen, Y., Wang, T., & Tang, K. (2011). Enhancement of artemisinin content in tetraploid Artemisia annua plants by modulating the expression of genes in artemisinin biosynthetic pathway. Biotechnology and applied biochemistry, 58(1), 50-57. https://iubmb.onlinelibrary.wiley.com/doi/abs/10.1002/bab.13. [CrossRef]
- Liu, C., Ahmad, N., Tao, Y., Hussain, H., Chang, Y., Umar, A. W., & Liu, X. (2025a). Reprogramming Hairy Root Cultures: A Synthetic Biology Framework for Precision Metabolite Biosynthesis. Plants, 14(13), 1928. https://www.mdpi.com/2223-7747/14/13/1928. [CrossRef]
- Liu, Y. H., Zeng, W. Q., Tao, S. F., Du, Y. N., Yu, F. H., Qi, Z. C., & Yang, D. F. (2025b). SmilODB: a multi-omics database for the medicinal plant danshen (Salvia miltiorrhiza, Lamiaceae). Frontiers in Plant Science, 16, 1586268. https://www.frontiersin.org/journals/plant-science/articles/10.3389/fpls.2025.1586268/full. [CrossRef]
- Malu, M., Chatterjee, J., Choudhary, D., Ramakrishna, W., & Kumar, R. (2025). Biotic, abiotic, and genetic elicitors as a new paradigm for enhancing alkaloid production for pharmaceutical applications. South African Journal of Botany, 177, 579-597. https://www.sciencedirect.com/science/article/abs/pii/S0254629924008019. [CrossRef]
- Moreira, G. C., Carneiro, C. N., Dos Anjos, G. L., da Silva, F., Santos, J. L., & Dias, F. D. S. (2022). Support vector machine and PCA for the exploratory analysis of Salvia officinalis samples treated with growth regulators based in the agronomic parameters and multielement composition. Food Chemistry, 373, 131345. https://www.sciencedirect.com/science/article/abs/pii/S0308814621023517. [CrossRef]
- Mukherjee, S., Biswas, D., & Ghosh, B. (2025). Carbohydrates acting as enhancers of secondary metabolites synthesis in in vitro cultures of Centella asiatica (L.) Urb. Plant Cell, Tissue and Organ Culture (PCTOC), 161(2), 1-16. https://link.springer.com/article/10.1007/s11240-025-03055-w. [CrossRef]
- Navrátilová, B., Ondřej, V., Vrchotová, N., Tříska, J., Horník, Š., & Pavela, R. (2022). Impact of Artificial Polyploidization in Ajuga reptans on content of selected biologically active glycosides and Phytoecdysone. Horticulturae, 8(7), 581. https://www.mdpi.com/2311-7524/8/7/581. [CrossRef]
- Nordine, A. (2025). Trends in plant tissue culture, production, and secondary metabolites enhancement of medicinal plants: a case study of thyme. Planta, 261(4), 84. https://link.springer.com/article/10.1007/s00425-025-04655-8. [CrossRef]
- Paul, P., Singh, S. K., Patra, B., Sui, X., Pattanaik, S., & Yuan, L. (2017). A differentially regulated AP 2/ERF transcription factor gene cluster acts downstream of a MAP kinase cascade to modulate terpenoid indole alkaloid biosynthesis in Catharanthus roseus. New Phytologist, 213(3), 1107-1123. https://nph.onlinelibrary.wiley.com/doi/full/10.1111/nph.14252. [CrossRef]
- Rasheed, F., Naeem, M., Uddin, M., & Khan, M. (2017). Synergistic effect of irradiated sodium alginate and methyl jasmonate on anticancer alkaloids production in periwinkle (Catharanthus roseus L.). https://www.indianjournals.com/ijor.aspx?target=ijor:jfeb&volume=7&issue=2&article=002. [CrossRef]
- Rauf, A. Rauf, A., Naz, S., Khan, M. U., Zahid, T., Ahmad, Z., Akram, Z., ... & Alhomrani, M. (2025). Aldose reductase inhibitory evaluation and in Silico studies of bioactive secondary metabolites isolated from Fernandoa. Adenophylla (Wall. Ex G. Don). Journal of Molecular Structure, 1328, 141308. https://www.sciencedirect.com/science/article/abs/pii/S0022286024038134. [CrossRef]
- Rithichai, P., Jirakiattikul, Y., Nambuddee, R., & Itharat, A. (2024). Effect of salicylic acid foliar application on bioactive compounds and antioxidant activity in holy basil (Ocimum sanctum L.). International Journal of Agronomy, 2024(1), 8159886. https://onlinelibrary.wiley.com/doi/full/10.1155/2024/8159886. [CrossRef]
- Sánchez-López, G. C., Carranza-Ojeda, D., Pérez-Picaso, L., Martínez-Pascual, R., Viñas-Bravo, O., López-Torres, A., Molphe-Bach, E.P., García-Ríos, E., Morales-Serna, J.A., & Villalobos-Amador, E. (2025). Establishment of in vitro root cultures and hairy roots of Dioscorea composita for diosgenin production. Plant Cell, Tissue and Organ Culture (PCTOC), 161(1), 1-22. https://link.springer.com/article/10.1007/s11240-025-03022-5. [CrossRef]
- Shi, M., Zhang, S., Zheng, Z., Maoz, I., Zhang, L., & Kai, G. (2024). Molecular regulation of the key specialized metabolism pathways in medicinal plants. Journal of Integrative Plant Biology, 66(3), 510-531. https://onlinelibrary.wiley.com/doi/full/10.1111/jipb.13634. [CrossRef]
- Singh, M., Suyal, D. C., Dinkar, N., Joshi, S., Srivastava, N., Maurya, V. K., Agnihotri, A., & Agrawal, S. (2023). Agrobacterium rhizogenes as molecular tool for the production of hairy roots in Withania somnifera. J. Appl. Biol. Biotechnol., 11, 1-10. https://d1wqtxts1xzle7.cloudfront.net/113878469/870_pdf-libre.pdf?1714297075=&response-content-disposition=inline%3B+filename%3DAgrobacterium_rhizogenes_as_molecular_to.pdf&Expires=1753888727&Signature=SLb2izVSl8HHUO9MjO~94lfbfFjfaBXG9wAt96q453~jE5A7QNcWRWctJBRi5yAjKkytRV0fIJst4fCgxQ1zKROymZINaWjO~mQG0CI9YvLSLZ7qCS0SDxwvGdPtRHq4japnEFojb9AIXJag4EFXnwxiOjU7e85zsRa0Mqbg~B2u3jtW338pe5wCV7gF-wOoG7w-MIbixF8ebhr5uHvxcSy~kUQWTDf-JnB-hOiluWM75Z2cIljV7gvHNduXhTgOEWKyRmvMI5Xmt69zwZ4BIK~iHJ5nT4mnRMgXvb391C4LBMz95-QgC~j-K5~V3MY3iv4DiVYjBK7i1KLNDiRWfg__&Key-Pair-Id=APKAJLOHF5GGSLRBV4ZA.
- Singh, R., Yadav, R., & Fatima, S. K. (2025). The engineering of medicinal plants: Future prospects and limitations. In Medicinal Biotechnology (pp. 79-102). Academic Press. https://www.sciencedirect.com/science/article/abs/pii/B9780443222641000050.
- Srivastava, M., Sharma, S., & Misra, P. (2016). Elicitation based enhancement of secondary metabolites in Rauwolfia serpentina and Solanum khasianum hairy root cultures. Pharmacognosy magazine, 12(Suppl 3), S315. https://pmc.ncbi.nlm.nih.gov/articles/PMC4971950/. [CrossRef]
- Srivastava, M., Singh, G., Sharma, S., Shukla, S., & Misra, P. (2019). Elicitation enhanced the yield of glycyrrhizin and antioxidant activities in hairy root cultures of Glycyrrhiza glabra L. Journal of Plant Growth Regulation, 38(2), 373-384. https://link.springer.com/article/10.1007/s00344-018-9847-2. [CrossRef]
- Strzemski, M., Dresler, S., Hawrylak-Nowak, B., Tkaczyk, P., Kulinowska, M., Feldo, M., Maggi, F., & Hanaka, A. (2025). Chitosan elicitation enhances biomass and secondary metabolite production in Carlina acaulis L. Scientific Reports, 15(1), 23411. https://www.nature.com/articles/s41598-025-07085-4. [CrossRef]
- Svecarova, M., Navrátilová, B., Hasler, P., & Ondrej, V. (2019). Artificial induction of tetraploidy in Humulus lupulus L. using oryzalin. Acta Agrobotanica, 72(2). https://scholar.google.com/scholar_lookup?title=Artificial+induction+of+tetraploidy+in+Humulus+lupulus+L.+using+oryzalin&author=%C5%A0v%C3%A9carov%C3%A1,+M.&author=Navr%C3%A1tilov%C3%A1,+B.&author=Ha%C5%A1ler,+P.&author=Ond%C5%99ej,+V.&publication_year=2019&journal=Acta+Agrobot.&volume=72&pages=1764&doi=10.5586/aa.1764. [CrossRef]
- Swain, H. , Gantait, S., & Mandal, N. (2023). Developments in biotechnological tools and techniques for production of reserpine. Applied microbiology and biotechnology 107(13), 4153–4164. [CrossRef] [PubMed]
- Taheri, A., Almeida-Silva, F., Zhang, Y., Fu, X., Li, L., Wang, Y., & Tang, K. (2025). Artemisia Database: A Comprehensive Resource for Gene Expression and Functional Insights in Artemisia annua. bioRxiv, 2025-05. https://www.biorxiv.org/content/10.1101/2025.05.21.655314v1.abstract.
- Tomilova, S. V., Globa, E. B., Demidova, E. V., & Nosov, A. M. (2023). Secondary metabolism in Taxus spp. plant cell culture in vitro. Russian Journal of Plant Physiology, 70(3), 23. https://link.springer.com/article/10.1134/S102144372270008X. [CrossRef]
- Twaij, B. M., & Hasan, M. N. (2022). Bioactive secondary metabolites from plant sources: types, synthesis, and their therapeutic uses. International Journal of Plant Biology, 13(1), 4-14. https://www.mdpi.com/2037-0164/13/1/3.
- Verma, P., Dixit, J., Singh, C., Singh, A. N., Singh, A., Tiwari, K. N., Muthu, M.S., Nath, G., & Mishra, S. K. (2024). Preparation of hydrogel from the hydroalcoholic root extract of Premna integrifolia L. and its mediated green synthesis of silver nanoparticles for wound healing efficacy. Materials Today Communications, 41, 110228. https://www.sciencedirect.com/science/article/abs/pii/S2352492824022098. [CrossRef]
- Wang, M., Zhang, S., Li, R., & Zhao, Q. (2024). Unraveling the specialized metabolic pathways in medicinal plant genomes: A review. Frontiers in Plant Science, 15, 1459533. https://www.frontiersin.org/journals/plant-science/articles/10.3389/fpls.2024.1459533/full. [CrossRef]
- Wu, Z., Hu, Y., Hao, R., Li, R., Lu, X., Itale, M. W., Yuan, Y., Zhu, X., Zhang, J., Wang, L. and Sun, M., & Hou, X. (2025). Research Progress of Genomics Applications in Secondary Metabolites of Medicinal Plants: A Case Study in Safflower. International Journal of Molecular Sciences, 26(8), 3867. https://www.mdpi.com/1422-0067/26/8/3867. [CrossRef]
- Xiang, L. Xiang, L., Zhu, S., Zhao, T., Zhang, M., Liu, W., Chen, M., Lan, X., & Liao, Z. (2015). Enhancement of artemisinin content and relative expression of genes of artemisinin biosynthesis in Artemisia annua by exogenous MeJA treatment. Plant growth regulation, 75(2), 435-441. https://link.springer.com/article/10.1007/s10725-014-0004-z. [CrossRef]
- Xiang, L., Jian, D., Zhang, F., Yang, C., Bai, G., Lan, X., Chen, M., Tang, K., & Liao, Z. (2019). The cold-induced transcription factor bHLH112 promotes artemisinin biosynthesis indirectly via ERF1 in Artemisia annua. Journal of experimental botany, 70(18), 4835-4848. https://academic.oup.com/jxb/article/70/18/4835/5489275. [CrossRef]
- Xie, L., Yan, T., Li, L., Chen, M., Hassani, D., Li, Y., Qin, W., Liu, H., Chen, T., Fu, X. and Shen, Q., & Tang, K. (2021). An HD-ZIP-MYB complex regulates glandular secretory trichome initiation in Artemisia annua. New Phytologist, 231(5), 2050-2064. https://nph.onlinelibrary.wiley.com/doi/full/10.1111/nph.17514. [CrossRef]
- Yan, T., Chen, M., Shen, Q., Li, L., Fu, X., Pan, Q., Shi, P., Lv, Z., Jiang, W. and Ma, Y.N., & Tang, K. (2017). HOMEODOMAIN PROTEIN 1 is required for jasmonate-mediated glandular trichome initiation in Artemisia annua. New Phytologist, 213(3), 1145-1155. https://nph.onlinelibrary.wiley.com/doi/full/10.1111/nph.14205. [CrossRef]
- Yang, Y., Chen, W., Mei, Z., & Liu, Y. (2025). The multifaceted role of microRNA in medicinal plants. Medicinal Plant Biology, 4(1). https://maxapress.com/article/doi/10.48130/mpb-0025-0006. [CrossRef]
- Yin, Y. C., Hou, J. M., Tian, S. K., Yang, L., Zhang, Z. X., Li, W. D., & Liu, Y. (2020). Overexpressing chalcone synthase (CHS) gene enhanced flavonoids accumulation in Glycyrrhiza uralensis hairy roots. Botany Letters, 167(2), 219-231. https://www.tandfonline.com/doi/abs/10.1080/23818107.2019.1702896. [CrossRef]
- Yuan, M., Shu, G., Zhou, J., He, P., Xiang, L., Yang, C., Chen, M., Liao, Z., & Zhang, F. (2023). AabHLH113 integrates jasmonic acid and abscisic acid signaling to positively regulate artemisinin biosynthesis in Artemisia annua. New Phytologist, 237(3), 885-899. https://nph.onlinelibrary.wiley.com/doi/full/10.1111/nph.18567. [CrossRef]
- Yuan, M., Sheng, Y., Bao, J., Wu, W., Nie, G., Wang, L., & Cao, J. (2025). AaMYC3 bridges the regulation of glandular trichome density and artemisinin biosynthesis in Artemisia annua. Plant Biotechnology Journal, 23(2), 315-332. https://onlinelibrary.wiley.com/doi/full/10.1111/pbi.14449. [CrossRef]
- Zhao, L., Zhu, Y., Jia, H., Han, Y., Zheng, X., Wang, M., & Feng, W. (2022). From plant to yeast—Advances in biosynthesis of artemisinin. Molecules, 27(20), 6888. https://www.mdpi.com/1420-3049/27/20/6888. [CrossRef]
- Zhao, Z., Zhang, Y., Li, W., Tang, Y., & Wang, S. (2023). Transcriptomics and physiological analyses reveal changes in paclitaxel production and physiological properties in Taxus cuspidata suspension cells in response to elicitors. Plants, 12(22), 3817. https://www.mdpi.com/2223-7747/12/22/3817, https://www.sciencedirect.com/science/article/abs/pii/S0168165624001093. [CrossRef]
- Zunun-Pérez, A. Y., Guevara-Figueroa, T., Jimenez-Garcia, S. N., Feregrino-Pérez, A. A., Gautier, F., & Guevara-González, R. G. (2017). Effect of foliar application of salicylic acid, hydrogen peroxide and a xyloglucan oligosaccharide on capsiate content and gene expression associated with capsinoids synthesis in Capsicum annuum L. Journal of Biosciences, 42(2), 245-250. https://link.springer.com/article/10.1007/s12038-017-9682-9. [CrossRef]
Figure 1.
Biotechnological Strategies in Secondary Metabolite Enhancement.
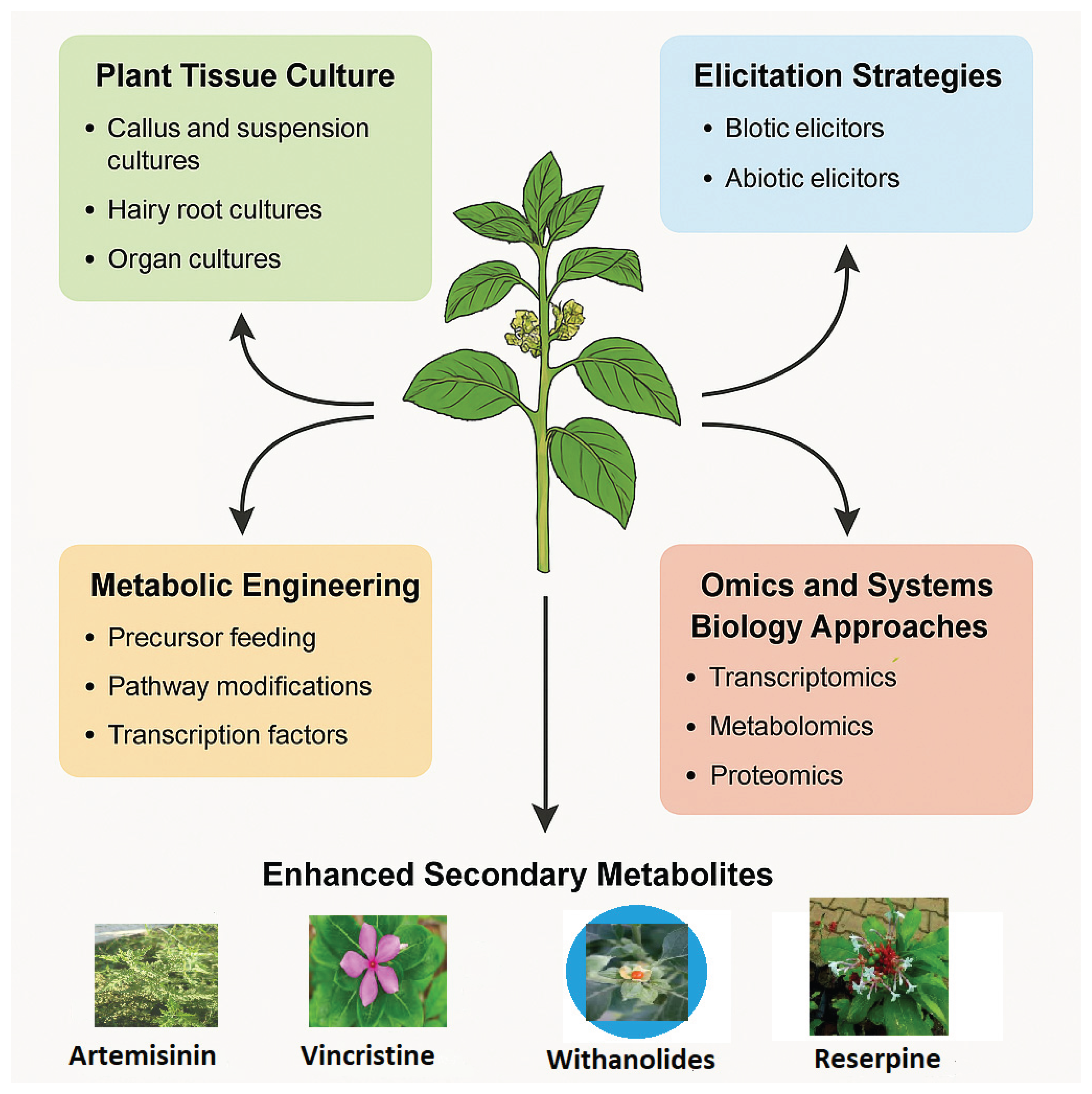
Figure 2.
Innovative Pathways for Secondary Metabolite Enrichment in Medicinal Plants (Modified from Ghosh et al., 2024).
Figure 2.
Innovative Pathways for Secondary Metabolite Enrichment in Medicinal Plants (Modified from Ghosh et al., 2024).
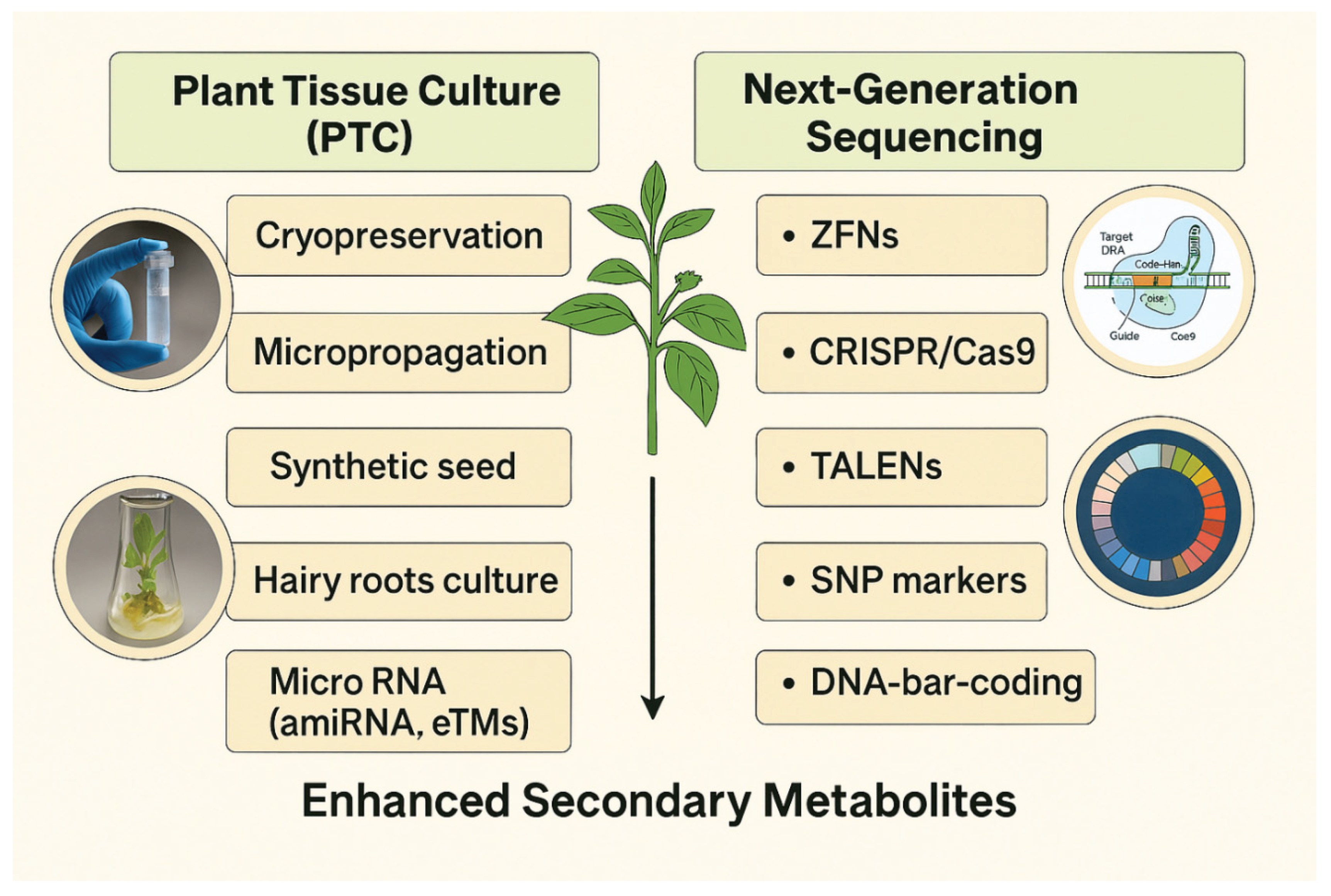
Figure 3.
Flowchart of Plant Secondary Metabolite Biosynthetic Pathways (Modified from Kajla et al., 2023).
Figure 3.
Flowchart of Plant Secondary Metabolite Biosynthetic Pathways (Modified from Kajla et al., 2023).
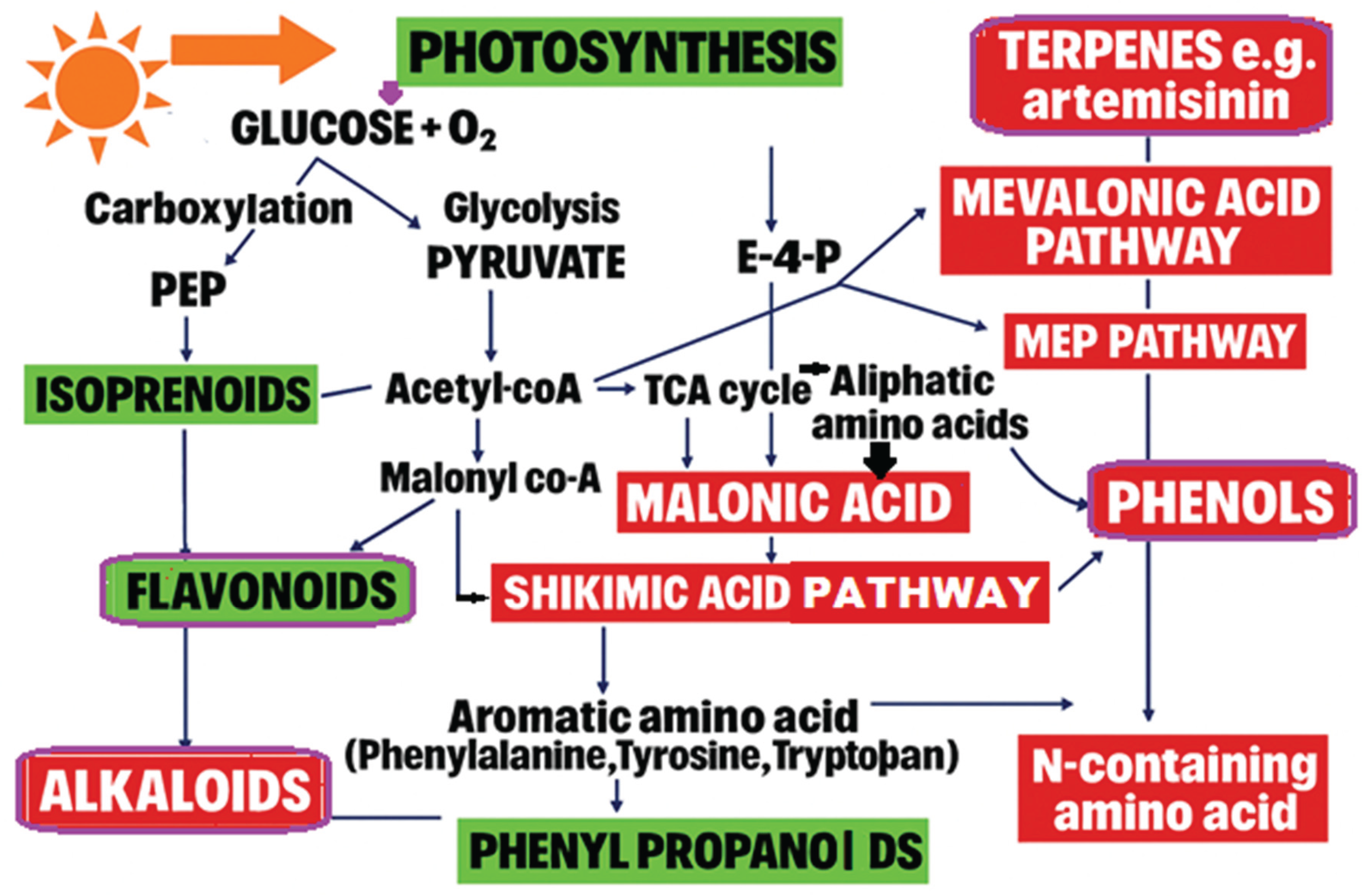
Figure 4.
Multi-Omics integration in Medicinal Plants Metabolic Pathways, Challenges and Solutions (Modified from Wang et al., 2024).
Figure 4.
Multi-Omics integration in Medicinal Plants Metabolic Pathways, Challenges and Solutions (Modified from Wang et al., 2024).
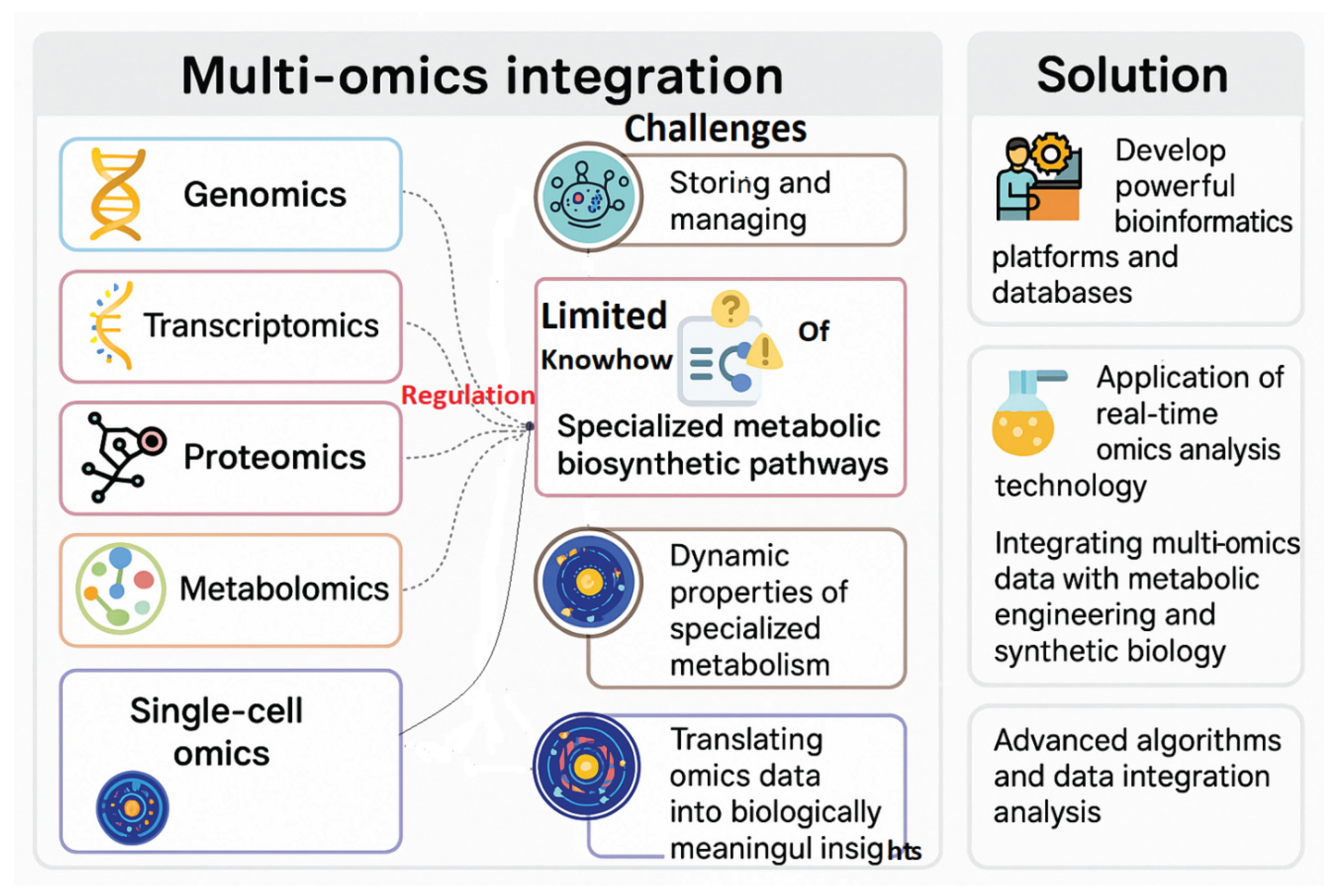
Figure 6.
Transcriptional Regulation of Artemisinin Biosynthesis via JA Signaling Pathways in Artemisia annua (Modified from Yuan et al., 2023) Note: The large green boxes show Transcription Factors/TFs,Yellow boxes= Enzymes, JA=Jasmonic acid, ADS= amorpha-4,11-diene synthase, CYP71AV1= cytochrome P450-dependent hydroxylase, , DBR2=double bond reductase 2, ALDH1= aldehyde dehydrogenase 1.
Figure 6.
Transcriptional Regulation of Artemisinin Biosynthesis via JA Signaling Pathways in Artemisia annua (Modified from Yuan et al., 2023) Note: The large green boxes show Transcription Factors/TFs,Yellow boxes= Enzymes, JA=Jasmonic acid, ADS= amorpha-4,11-diene synthase, CYP71AV1= cytochrome P450-dependent hydroxylase, , DBR2=double bond reductase 2, ALDH1= aldehyde dehydrogenase 1.
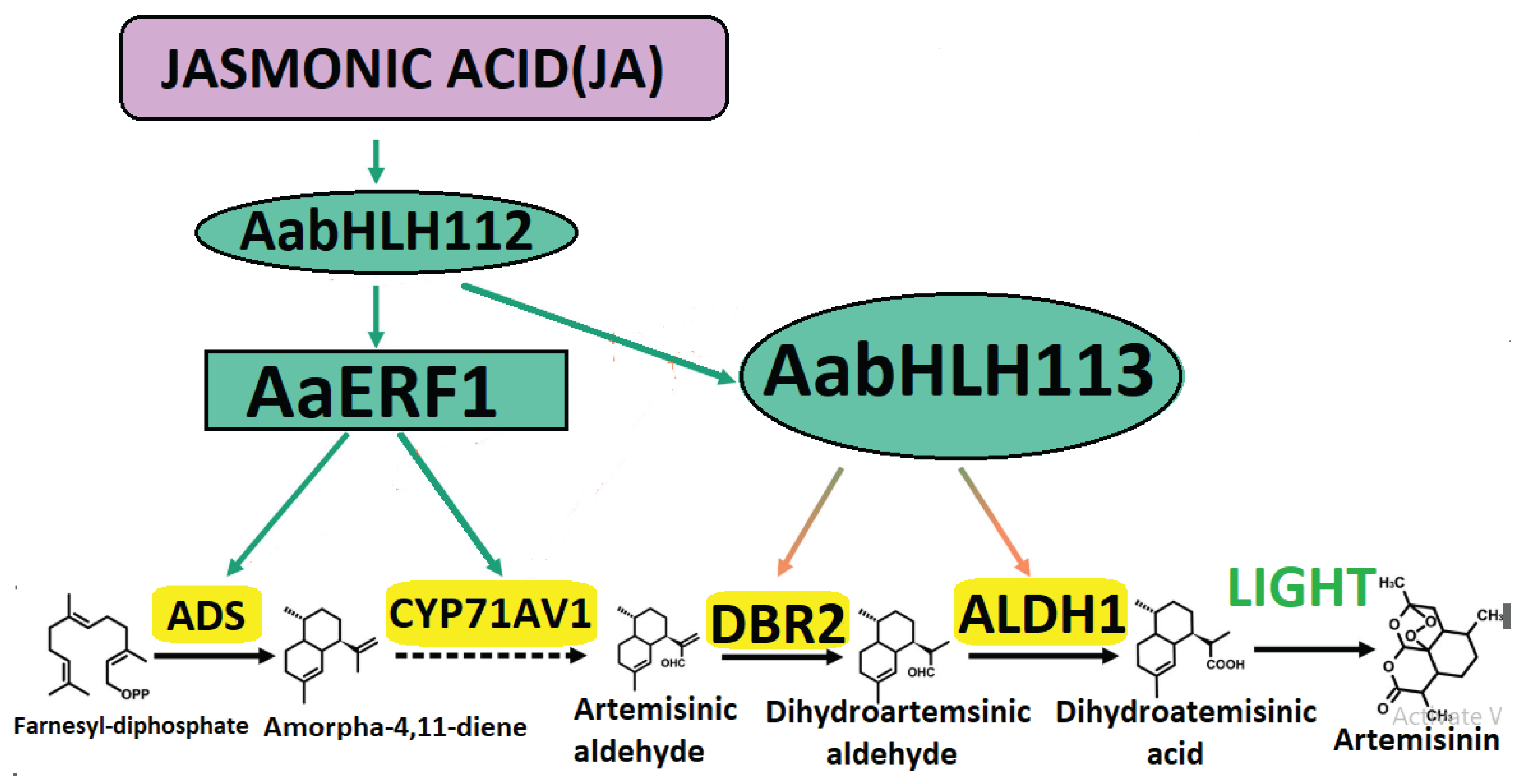
Figure 7.
The Artemisinin database output analysis of Tissue-Specific Gene Expression Heatmap Landscapes of Artemisinin Biosynthetic Genes in Artemisia annua: Insights from Spatial Transcriptomics and Functional Genomics. Note: Gene IDs and the consecutive genes names are mikado.chr4G1338=ADS, mikado.chr3G1536=CYP71AV1, mikado.chr2G1956=DBR2, mikado.chr8G609=ALDH1. (https://artemisia-db.com/).
Figure 7.
The Artemisinin database output analysis of Tissue-Specific Gene Expression Heatmap Landscapes of Artemisinin Biosynthetic Genes in Artemisia annua: Insights from Spatial Transcriptomics and Functional Genomics. Note: Gene IDs and the consecutive genes names are mikado.chr4G1338=ADS, mikado.chr3G1536=CYP71AV1, mikado.chr2G1956=DBR2, mikado.chr8G609=ALDH1. (https://artemisia-db.com/).
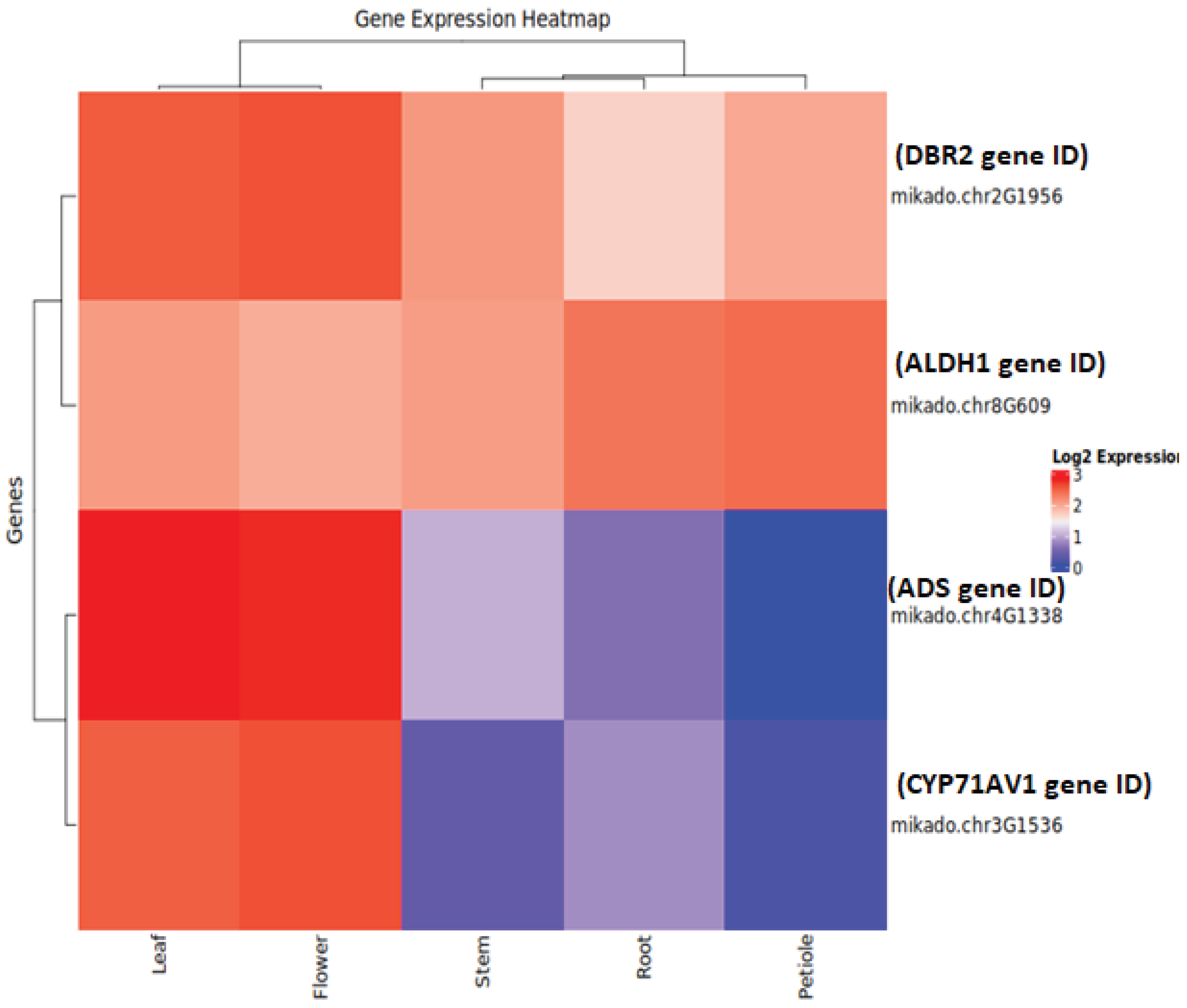

Table 1.
Biotechnological In Vitro Culture Strategies and Approaches for Enhanced Secondary Metabolite Production in Medicinal Plants.
Table 1.
Biotechnological In Vitro Culture Strategies and Approaches for Enhanced Secondary Metabolite Production in Medicinal Plants.
| Plant Species | Culture Type | Target Metabolite | Yield Increase | Reference |
|---|---|---|---|---|
| Artemisia annua | Organ culture | Artemisinin | 7.8 fold | Al-Khayri et al., 2022 |
| Rauwolfia serpentina | Callus culture | Reserpine | 2 fold | Gantet et al., 2023 |
| Taxus brevifolia and Taxus cuspidata | Cell suspension | Paclitaxel | Commercial large scale | Fett-Neto & DiCosmo, 2020; Zhao et al., 2023 |
| Withania somnifera | Hairy roots | Withanolides | 11.49 fold | Haider and Gosh, 2024 |
Table 2.
Influence of Different Elicitors on Secondary Metabolite Production in Selected Medicinal Plants.
Table 2.
Influence of Different Elicitors on Secondary Metabolite Production in Selected Medicinal Plants.
| Elicitor Type | Plant Model | Metabolite | Observed Effect | Reference |
|---|---|---|---|---|
| AgNPs | Achillea millefolium | Essential oil | 2.3× increase | Lala et al., 2021 |
| Cellulase | Glycyrrhiza glabra | Glycyrrhizin | 8.6× increase | Srivastava et al., 2019 |
| Jasmonic Acid | Catharanthus roseus | Vinblastine, Vincristine | 0.6× increase | Rasheed et al., 2017 |
| Salicylic Acid | Ocimum sanctum | Eugenol | 282.96× increase | Rithichai et al., 2024 |
| Yeast Extract | Panax ginseng | Ginsenosides | 1.57× increase | Kochan et al., 2017 |
Table 3.
Genetic and Signaling-Based Approaches for the Improvement of Secondary Metabolite Production in Medicinal Plants.
Table 3.
Genetic and Signaling-Based Approaches for the Improvement of Secondary Metabolite Production in Medicinal Plants.
| Modification Strategy | Plant | Gene Targeted | Result | Reference |
|---|---|---|---|---|
| 3-hydroxy-3-methylglutaryl coenzyme and amorpha-4,11-diene synthase coexpression | Artemisia annua | HMGR and ADS genes | ↑Artemisinin 8.65× increase |
Zhao et al., 2022 |
| ADS-FPPS enzymes co-expression | Artemisia annua | ADS-FPPS genes | ↑Artemisinin 2.6× increase |
Zhao et al., 2022 |
| Co-expression of TFs ORCA3 and MYC2 | Catharanthus roseus | Transcriptional regulators | ↑ TIA Biosynthesis as Vincristine and ajmalicine | Paul et al., 2017 |
| FPPS, CYP71AV1 and CPR enzymes co-expression | Artemisia annua |
FPPS, CYP71AV1 and CPR genes |
↑Artemisinin 3.6× increase |
Zhao et al., 2022 |
| Overexpression of CHS | Glycyrrhizia uralensis | Chalcone synthase | ↑ Flavonoid content | Yin et al., 2020 |
| salicylic acid/H2O2 | Capsicum annuum | Phenylalanine ammonia-lyase | ↑ Capsiate production | Zunun-Pérez et al., 2017 |
Table 4.
Impact of Polyploidy-Inducing Agents on Medicinal Plant Secondary Metabolite Yield.
| Agent | Plant Model | Mechanism | Metabolite | Observed Effect | Reference |
|---|---|---|---|---|---|
| Oryzalin | Ajuga reptans | Tetraploidy | 20-hydroxecdysone | Increased content | Navrátilová et al., 2022; Svecarova et al., 2019 |
| Colchicine | Catharanthus roseus | Autoploidy (chromosome doubling) | vincristine | 2× increase | Fathimunnissa, 2011 |
| Colchicine | Bacopa monnieri | Autoploidy (diploidy) | bacoside | 4× increase | Kharde et al., 2017 |
| Colchicine | Artemisia annua | Tetraploidy | artemisinin | 0.7× increase | Lin et al., 2011 |
| Colchicine | Echinacea purpurea | Tetraploidy | cichoric acid and chlorogenic acid | 0.45× and 0.71× increase respectively | Abdoli et al., 2013 |
| Colchicine | Linum album | Polyploidy | podophyllotoxin | 1.39× increase | Javadian et al., 2017 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



