Submitted:
31 July 2025
Posted:
31 July 2025
You are already at the latest version
Abstract
Lady beetle can utilize the alternative food sources to enhance their diets, other than essential prey. These alternative diets may enhance their developments, survival and fecundity. The study examined the effects of prey and alternative food sources on the development and performance of predaceous coccinellid Coelophora inaequalis (Fab-ricius) (Coleoptera: Coccinellidae), utilizing eggs of rice moth Corcyra cephalonica (Stainton) (Lepidoptera: Pyralidae) and bee pollen as alternative diets, while Aphis gossypii Glover (Hemiptera: Aphididae) served as the essential prey. Results revealed that 1st instar larvae of C. inaequalis are not able to maintain their development by feeding exclusively on an alternative diets, while other groups fed on A. gossypii and A. gossypii supplemented with bee pollen success to complete development. Although C. inaequalis that fed on A. gossypii supplemented with bee pollen experienced a shorter development time than those fed solely on A. gossypii, they exhibited decreased fecun-dity, lower survival rate and shorter adult longevity compared to the latter group. Adult of C. inaequalis fed exclusively on rice moth eggs or bee pollen or a combination of both can survive for 4-9 days. Rice moth eggs and bee pollen can serve as alternative food sources to enhance longevity of C. inaequalis adults.
Keywords:
bee pollen
; rice moth
; Aphis gossypii
1. Introduction
Lady beetles (Coleoptera: Coccinellidae) are major natural enemies of aphids throughout their life cycles. Naturally, lady beetles are generalist predators consume various small preys such as aphids, psyllids, mealybugs and scale insects in their habitats. Moreover, many reports indicated that with the exception of live prey, some lady beetle species consume alternative foods including pollen, extrafloral nectar, foliage and fruit [1,2,3,4]. One of the msost nutritious of non-prey foods consumed by coccinellid is pollen. [4] reported that pollen is quantitatively superior to prey in terms of energy, protein and carbohydrates, and is differed significantly in lipids content compared with prey. In addition, pollen also contains other beneficial nutrients needed for insects such as amino acids, sugars, sterols, vitamins, flavonoids, carotenoids, and minerals [5,6]. At least 39 species of predatory lady beetle have been recorded as consuming pollen [4]. Previous studies reported both beneficial and unbeneficial roles of pollen on the development and reproduction of coccinellid species. For certain coccinellid species, consuming pollen alongside aphids can enhance reproduction and overall performance [7,8,9]. Larvae of Adalia bipunctata (Coleoptera: Coccinellidae) fed only on pollen of Rosaceae successfully develop to adults [10]. A diet of bee pollen alone allowed 35–48% of the larvae of the field population of multicoloured Asian lady beetle Harmonia axyridis (Pallas) (Coleoptera: Coccinellidae), exotic species settle down in Europe, successfully reach adulthood and females able to produce a small number of viable eggs [11]. The exploitation pollen as a food to sustain development and reproduction in the absence of insect of H. axyridis supported competitive advantage over the native European lady beetles [11]. In contrast, [12] reported that pollen did not support the development time of immature stages of H. axyridis reared on canola pollen together with aphid Myzus persicae (Sulzer) (Hemiptera: Aphididae) when compared with other group fed only on M. persicae. Moreover, female fecundity of H. axyridis reared on mix diet were significantly lower than that of only fed M. persicae and this phenomenon found in Propylea japonica (Thunberg) (Coleoptera: Coccinellidae) also. Likewise, bee pollen did not support to produce mature oocytes of predatory lady beetle Brumoides foudraii (Mulsant) (Coleoptera: Coccinellidae) reared on the mix of mealybug Ferrisia dasylirii (Cockerell) (Hemiptera: Pseudococcidae) and pollen when compared to other group only fed on F. dasylirii [13].
Besides this, eggs of the rice moth Corcyra cephalonica (Stainton) (Lepidoptera: Pyralidae) served as an efficient host for mass-rearing various species of parasitoids and predators [14] were studied on the suitable as alternative food for coccinellids rearing. [15] reported the survival rates and predatory efficacy of the Serangium japonicum Chapin (Coleoptera: Coccinellidae) are similar to those feeding on natural host Bemesia tabaci Gennadius (Hemiptera: Aleyrodidae) when S. japanicum larvae from 2nd larval instar onwards reared on C. cephalonica eggs.
Coelophora inaequalis (Fabricius) (Coleoptera: Coccinellidae) is widely distributed across India (Andamans), Sri Lanka, Southeast Asia (Thailand, Malaysia, Indonesia, Philippines), New Guinea, New Caledonia, Micronesia and Australia [16]. Coelophora inaequalis was introduced to Hawaii and Florida as a biological control agent to control the yellow sugarcane aphid Sipha flava (Forbes) (Hemiptera: Aphididae) [17]. Recently, C. inaequalis are important lady beetle species commonly found in many regions of Thailand distributed cover Northern to Southern. Biology of C. inaequalis settled down in Thailand has not yet been studied. Previous studies of the life history of C. inaequalis reviewed that C. inaequalis fed on Aphis craccivora Koch (Hemiptera: Aphididae) has the total development period, including incubation, was 10-11 days. The life span of the female adults was 112-126, while that of the male adults was 111-126 days and the total consumption from egg hatching to adult death was 2,821-3,509 prey individuals for male and 2,959-4,779 prey individuals for female. The number of eggs laid by the female total 510-783 eggs for a period of 30 days [18]. Understanding the biology and possibility of mass rearing of C. inaequalis can be useful for biological control of insect pests and contribute to sustainable agricultural practices.
2. Materials and Methods
2.1. Mass Rearing of Corcyra cephalonica
Eggs of rice moth C. cephalonica supported by Surat Thani Agricultural Technology Promotion Center (Plant Protection), Surat Thani, Thailand were sprinkled on diets composed of broken rice, rice bran and sugar (1: 1: 0.1, W:W). The ratio of rice moth eggs and diet was 0.1 g/1 kg of diet and kept in 20×30×10 cm plastic box under 25±1 °C, 70±5% RH, and 12 hours photoperiod in the laboratory. After C. cephalonica larvae developed to adult, male and female of C. cephalonica were moved to net bags and put on black plastic cup for egg harvesting. Harvested rice moth eggs were used to carry out the experiment.
2.2. Mass Rearing of Coelophora inaequalis
Adults of C. inaequalis were collected from organic vegetable farm, Paron Subdistrict, Kanchanadit District, Surat Thani Province, Thailand. Each pair of male and female adults were reared on Aphis gossypii Glover (Hemiptera: Aphididae) ad libitum in 6×4.5×3 cm plastic box under 25±1 °C, 70±5% RH, and 12 hours photoperiod in the laboratory. Water was supplied using moistened cotton ball put on 80 ml plastic cup. After female laid eggs, male and female of C. inaequalis were moved to new 6×4.5×3 cm plastic box for protection of egg cannibalism. Early hatched C. inaequalis larvae were used in this study.
Experiments I Effect of prey and alternative food sources on C. inaequalis development
First instar larvae of C. inaequalis were released in 6×4.5×3 cm plastic box and fed on tested diets: rice moth eggs, bee pollen, A. gossypii with bee pollen, and only A. gossypii as control. Previous observation found that 1st instar larvae of C. inaequalis only consumed A. gossypii and ignored rice moth eggs when two preys were offered simultaneously. Then, A. gossypii with rice moth eggs treatment was cut out from the experiment. Twenty larvae were used in each diets, and three replicates were established. Diets were supplied to C. inaequalis larvae ad libitum. Water was provided to larvae as same as adults under 25±1 °C, 70±5% RH, and 12 hours photoperiod in the laboratory. Developmental period and the number of consumed prey of each C. inaequalis stage and adult longevity were observed. The number of laid egg through female’s lifespans and percent of egg hatch were recorded.
Experiment II Effect of alternative food sources on longevity of C. inaequalis adult
To determine the effect of prey and alternative food sources on performance of C. inaequalis adult, newly emerged male and female C. inaequalis adults were fed on three different diets: rice moth eggs, bee pollen and rice moth eggs with bee pollen. Twenty male and female adults were fed per diet, and three replicates were established. All other experimental conditions were as for the experiment I. Adult longevity were observed.
2.3. Data Analysis
Two-way ANOVA was used to test for effect of C. inaequalis groups of each replicate, diet, and their interactions. If there was no significant interaction effect then the dependent data were pooled across individual group of each repeated experiment times (replicate) and effect of diet was tested using one-way ANOVA or t-test analysis. If a significant interaction effect was detected, then the effect of diet on the dependent data was analyzed by one-way ANOVA or t-test for each repeated experiment times. Data were transformed, if required, to meet the assumptions of ANOVA or t-test, and then back-transformed for graphical presentation of the number of prey and for the development time table. Response variables analyzed were the duration of larva-to-adult period, the number of consumed prey, adult longevity, the number of egg laid by female throughout their lifespan and percent of egg hatch. Post-hoc, pairwise comparisons of means were made using Tukey tests. The data were analyzed using SPSS statistics 17.0.
3. Results
3.1. Effect of Prey and Alternative Food Sources on C. inaequalis Development and Reproduction
Diets effected the development and survival of C. inaequalis. First instar larvae of C. inaequalis did not consume bee pollen and rice moth eggs, and all of larvae died on second days of the experiment. Then, C. inaequalis larvae fed on A. gossypii with bee pollen, and A. gossypii alone were observed their developments. There was no significant interaction effect between C. inaequalis groups of each repeated experiment times and diets in the developmental periods of C. inaequalis (ANOVA: 1st instar F2, 114 = 0.400, P = 0.671; 2nd instar F2, 114 = 0.282, P = 0.754; 3rd F2, 114 = 0.049, P = 0.952; 4th instar F2, 114 = 0.337, P = 0.715; pupal stage F2, 103 = 1.059, P = 0.351; adult stage; F2, 93 = 0.302, P = 0.740; male adult F2, 39 = 0.497, P = 0.612; female adult F2, 48 = 1.762, P = 0.183), so data were pooled across replicates. Results presented that with the exception of 3rd larval instar, C. inaequalis fed on A. gossypii supplemented with bee pollen seemed to develop in each stage faster than that of C. inaequalis fed on A. gossypii alone (Table 1). The higher mortality rate occurred on C. inaequalis fed on A. gossypii aphids with bee pollen as a supplementary food source. Finally, the number of survivable adult and adult longevity of C. inaequalis fed exclusively on A. gossypii was higher than that of C. inaequalis group fed on A. gossypii supplemented with bee pollen (Table 1). There was no significant difference in male longevity between the two diets. Females, on the other hand, demonstrate significantly longer lifespans when fed exclusively on A. gossypii. (Table 1).
There were no significant interaction effect between C. inaequalis groups of each repeated experiment times and diets in the number of laid eggs and percentage of egg hatch (ANOVA: laid eggs F2, 41 = 0.089, P = 0.915; percentage of egg hatch F2, 41 = 0.086, P = 0.918), then data were pooled across repeated experiment times. The research indicates that when C. inaequalis females fed only on A. gossypii, they produced significantly more eggs and a higher percentage of those eggs hatched compared to females fed on A. gossypii supplemented with bee pollen (t-test: laid eggs t = 6.815, P < 0.0001; percentage of egg hatch t = 6.031, P < 0.0001). Specifically, C. inaequalis females fed on A. gossypii alone laid 349.46±16.86 eggs with an 80.83±1.51% hatch rate, while those with the pollen supplement laid 214.10±8.96 eggs and had a 62.87±2.42% hatch rate.

Table 1.
Developmental period of C. inaequalis fed on different diets.
| Life stage | Development time (days, Mean±SE) | t-test | |
| A. gossypii | A. gossypii + bee pollen | ||
| 1st larva | 3.68±0.09 | 3.20±0.05 | t = -4.634, P<0.0001 |
| n | 60 | 60 | |
| 2nd larva | 2.87±0.11 | 2.67±0.09 | t = -1.289, P = 0.200 |
| n | 60 | 60 | |
| 3rd larva | 2.15±0.05 | 2.27±0.06 | t = 1.577, P = 0.118 |
| n | 60 | 60 | |
| 4th larva | 2.98±0.07 | 2.60±0.06 | t = -3.820, P<0.0001 |
| n | 60 | 60 | |
| pupa | 4.53±0.10 | 4.13±0.34 | t = -3.584, P = 0.001 |
| n | 57 | 52 | |
| Overall immature | 16.26±0.18 | 14.98±0.11 | t = 6.046, P<0.0001 |
| n | 57 | 52 | |
| Adult | 108.04±0.87 | 104.91±0.86 | t = 2.497, P=0.014 |
| n | 53 | 46 | |
| Male adult | 103.88±1.18 | 102.70±0.10 | t = 0.719, P = 0.476 |
| n | 25 | 20 | |
| Female adult | 111.75±0.76 | 106.62±1.24 | t = 3.560, P = 0.001 |
| n | 28 | 26 | |
n = number of C. inaequalis.
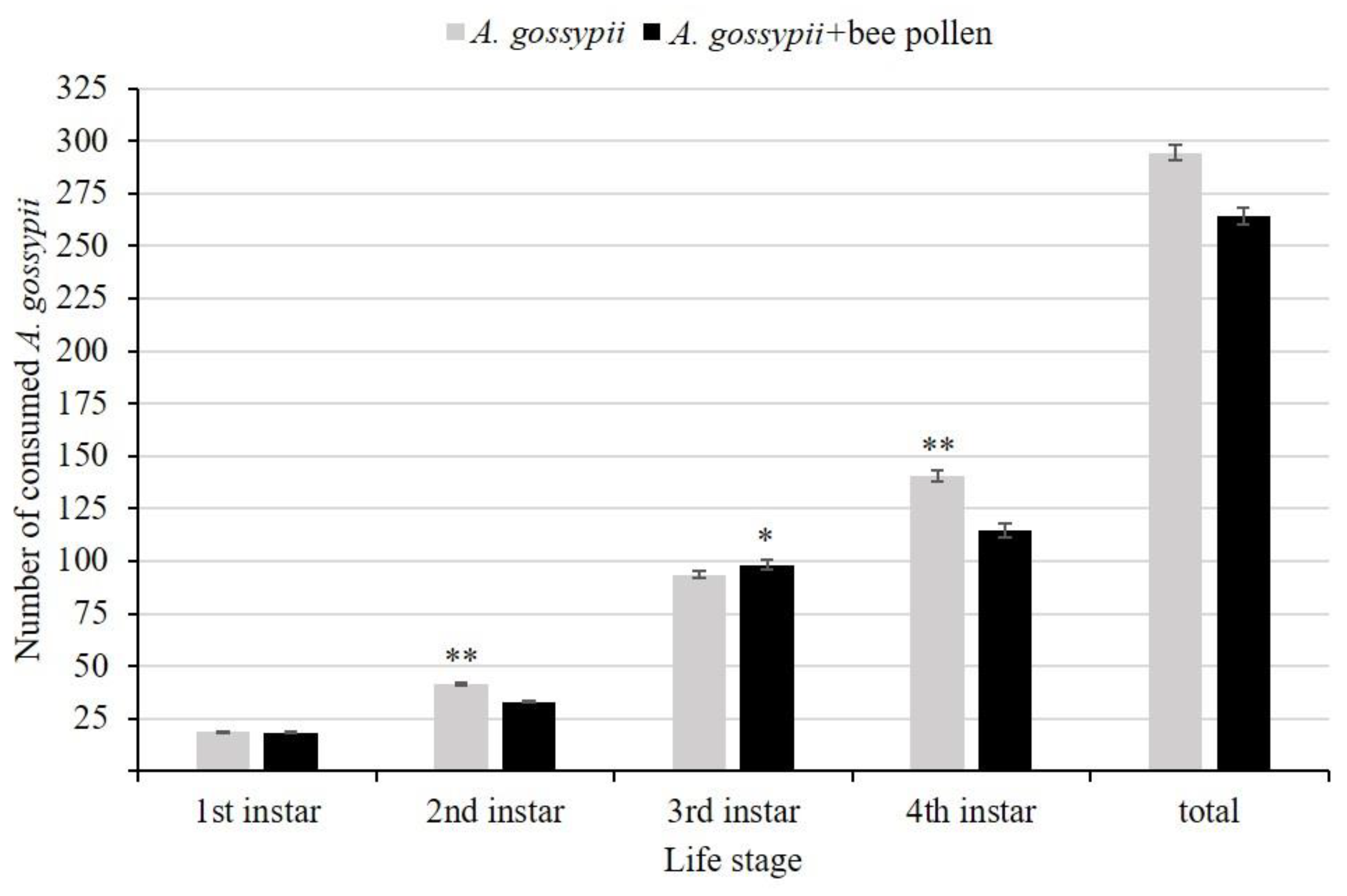
There was no significant interaction effect between C. inaequalis groups of each repeated experiment times and diets in the number of prey consumed by C. inaequalis (ANOVA: 1st instar F2, 114 = 0.164, P = 0.849; 2nd instar F2, 114 = 0.132, P = 0.877; 3rd F2, 114 = 0.688, P = 0.504; 4th instar F2, 114 = 0.352, P = 0.704), so data were pooled across experiment times. Results presented that with the exception of 1st instar larvae (t-test: t = 0.728, P = 0.468), there was the significant different in the number of consumed prey among different diet of each larval stage (Figure 1). The number of consumed A. gossypii was higher in C. inaequalis fed only on A. gossypii than that of C. inaequalis fed on A. gossypii supplemented with bee pollen in 2nd and 4th instar larvae and also all larval stages (t-test: 2nd instar t = 15.923, P < 0.0001; 3rd instar t = -2.851, P = 0.005; 4th instar t = 9.098, P < 0.0001; total t = 8.115, P < 0.0001) (Figure 1).
Figure 1.
Mean number (±SE) of prey consumed by C. inaequalis of each larval stages. Statistical significance different among treatments presented with one asterisk (Tukey test: P < 0.05) or two asterisks (Tukey test: P < 0.0001). Significance is based on log (x + 1)-transformed data, non-transformed data are plotted.
Figure 1.
Mean number (±SE) of prey consumed by C. inaequalis of each larval stages. Statistical significance different among treatments presented with one asterisk (Tukey test: P < 0.05) or two asterisks (Tukey test: P < 0.0001). Significance is based on log (x + 1)-transformed data, non-transformed data are plotted.
3.2. Effect of Alternative Food Sources on Longevity of C. inaequalis Adult
Results indicated that there were no significant interaction effect between C. inaequalis groups of both sexes of each repeated experiment times and diets among the longevity period of C. inaequalis male and female adults (ANOVA: male F4,171 = 2.163, P = 0.075; female F4,171 = 1.332, P = 0.260), then data were pooled across experiment time. The lifespan of male and female adults that were exclusively fed rice moth eggs was significantly shorter than that of groups fed bee pollen or a combination of rice moth eggs and bee pollen (Table 2). Notably, all females on these alternative diets did not lay any eggs throughout their lives.
Table 2.
The longevity of C. inaequalis adults fed on different diets.
| Prey | Longevity (day) | |
| male | female | |
| Bee pollen | 8.88±0.27a | 9.38±0.27a |
| Rice moth egg | 4.45±0.16b | 4.05±0.12b |
| Rice moth egg + bee pollen | 8.84±0.26a | 9.34±0.23a |
| F-test | F2,177 = 237.392, P < 0.0001 | F2,177 = 448.443, P < 0.0001 |
Values (mean ± SE) in the same column followed by a different letter are statistically different based on Tukey-test for longevity of each sexes (60 individuals of each diets) at P < 0.05. Significance is based on log (x + 1)-transformed data, non-transformed data are presented.
4. Discussion
Previous researches indicated that some important lady beetle species used for biological control survived and reproduced when reared on non-natural preys (pollen, nectar, eggs, larvae and pupae of other insects, etc.) [1,2,3,4]. However, results of this study presented that C. inaequalis larvae fed only on bee pollen or rice moth eggs could not survived from 1st instar to 2nd instar stage.
Interestingly, C. inaequalis adult could survived for 4-5 days and 8-9 days even only consumed rice moth eggs and bee pollen, respectively but did not lay eggs during that time. These results are partially explained by the different of prey suitability for larval and adult coccinellids. [19] noted that adult coccinellids possess stronger mandibles and more advanced digestive systems compared to their larval counterparts. Consequently, C. inaequalis larvae tend to reject non-prey foods due to their limited physiological capabilities. In this study, the 1st instar larvae of C. inaequalis were unable to survive when exclusively fed bee pollen or rice moth eggs, exhibiting the unacceptable behavior of not consuming them. Previous study also indicated the unacceptable on rice moth eggs of the 1st instar larvae of S. japonicum [20] but S. japanicum larvae from 2nd larval instar could consume rice moth eggs and completely develop to adults [19]. Although C. inaequalis larvae reared on A. gossypii supplemented with bee pollen had shorter development time, the survival rate was lower than that of C. inaequalis groups fed on A. gossypii solely. Thus, bee pollen may give some beneficial nutrients that support the development of C. inaequalis. In addition, result of experiment I indicated that C. inaequalis larvae reared on A. gossypii supplemented with bee pollen tend to consumed A. gossypii fewer than that of C. inaequalis fed on A. gossypii alone. This phenomenon was found in H. axyridis and P. japonica with decreasing in predation when pollen was provide as additional food [12]. They suggested that when pollen is readily available, H. axyridis and P. japonica may not need to rely as heavily on predation to obtain food. This can lead to a decrease in the number of insects they consume. The reduced consumption of A. gossypii by C. inaequalis larvae results in insufficient nutrient intake for their performance, leading to a higher mortality rate compared to the group that was exclusively fed A. gossypii. For results of diets tests in adults, bee pollen and rice moth eggs may provide essential nutrients for C. inaequalis adults but they did not lay eggs during their lifespans, regardless of these food sources. This implies that the C. inaequalis adults require specific food sources for survival but do not reproduce when given these limited diets. This suggests that C. inaequalis has specific nutritional requirements for reproduction that neither rice moth eggs nor bee pollen can fulfill. This could be due to a lack of certain essential nutrients, insufficient quantities of necessary nutrients, or a combination of both as [21] indicated that coccinellids need to consume more nutrients than required for maintenance to reproduce. For example, [22] demonstrated that adult female H. axyridis were unable to produce viable eggs when fed exclusively on a diet of Aphis spiraecola Patch (Hemiptera: Aphididae), despite the fact that this aphid species can support the complete development of larvae into adults. In this study, C. inaequalis could complete their developments and reproduce successfully when feeding on A. gossypii or A. gossypii supplemented with bee pollen. Aphids are the natural preys of C. inaequalis that support their success on development and reproduction [18]. Nevertheless, the fecundity of C. inaequalis adults fed on A. gossypii supplemented with bee pollen was significantly lower than that of other group fed on A. gossypii alone. In addition, results of previous studies indicated that pollen or bee pollen did not support the reproduction of all coccinellid species but generally they can prolong larval and adult longevity when prey are absent [13,23]. [12,23] proposed that pollen from specific plant species might contain secondary metabolites that are toxic to insects or that inhibit their oviposition. Hence, result of this study also confirmed that bee pollen did not increase the fecundity of C. inaequalis. For the Coccinellidae as a group, it has been proposed to classify prey types as “essential” if they support both completed larval development and successful adult reproduction, or “alternative” if they serve only as an energy source to extend adult longevity [24]. Therefore, results of this study confirm that A. gossypii was identified as an essential prey, while bee pollen and rice moth eggs were classified as alternative prey for C. inaequalis.
5. Conclusions
Bee pollen and rice moth eggs serve as alternative prey for adult C. inaequalis, providing temporary sustenance; however, exclusive consumption of these diets results in unsuccessful fecundity. Although C. inaequalis shows a greater acceptance of bee pollen as a food source compared to rice moth eggs, C. inaequalis adults that were fed on A. gossypii supplemented with bee pollen exhibited a decreased in fecundity and higher death rate, compared to those fed solely on A. gossypii. Furthermore, the 1st instar larvae of C. inaequalis showed no acceptance of bee pollen or rice moth eggs. This presents an intriguing opportunity for further research on the potential effects of offering bee pollen or rice moth eggs to other larval instars of C. inaequalis and possibility of applying these diets as artificial food for C. inaequalis mass rearing.
Author Contributions
Conceptualization, W.R.; methodology, W.R. and M.T.; formal analysis, W.R. and J.R.; writing—original draft preparation, W.R., J.R. and M.T.; writing— review and editing, W.R.; visualization, W.R.; supervision, W.R.; funding acquisition, W.R. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Prince of Songkla University, Surat Thani Campus.
Data Availability Statement
All datasets supporting the conclusion of this article are include in the article. Data will not be shared in any other source.
Acknowledgments
The authors would like to express their gratitude to Mr. Uthit Petchdaeng for his assistance with insect rearing.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Pemberton, R.W.; Vandenberg, N.J. Extrafloral nectar feeding by ladybird beetles (Coleoptera: Coccinellidae). Proceedings of the Entomological Society of Washington. 1993, 95, 139–151. [Google Scholar]
- Koch, R.L.; Burkness, E.C.; Wold Burkness, S.J.; Hutchinson, W.D. Phytophagous preferences of the multicolored Asian ladybeetle (Coleoptera: Coccinellidae) from autumn-riping fruit. J. Econ. Entomol. 2004, 97, 539–544. [Google Scholar] [CrossRef] [PubMed]
- Moser, S.E.; Harwood, J.D.; Obrycki, J.J. Larval feeding on Bt hybrid and non-Btcorn seedlings by Harmonia axyridis (Coleoptera: Coccinellidae) and Coleomegilla maculata (Coleoptera: Coccinellidae). Environ. Entomol. 2008, 37, 525–533. [Google Scholar] [PubMed]
- Lundgren, J.G. Nutritional aspects of non-prey foods in the life histories of predaceous Coccinellidae. Biol. Control. 2009, 51, 294–305. [Google Scholar] [CrossRef]
- Atrouse, O.M.; Oran, O.M.; AL-Abbadi, S.Y. Chemical analysis and identification of pollen grains from different Jordanian honey samples. Int. J. Food Sci. Tech. 2004, 39, 413–417. [Google Scholar] [CrossRef]
- Lundgren, J.G.; Wiedenmann, R.N. Nutritional suitability of field corn pollen as food for the predator, Coleomegilla magulata and Harmonia axyridis (Coleoptera: Coccinellidae) during anthesis in an Illinois cornfield. Environ. Entomol. 2004, 33, 958–963. [Google Scholar] [CrossRef]
- De Clercq, P.; Bonte, M.; Vanspebroeck, K.; Bolckmans, K.; Deforce, K. Development and reproduction of Adalia bipunctata (Coleoptera Coccinellidae) on eggs of Ephestia kuehniella (Lepidoptera: Phycitidae) and pollen. Pest Manag. Sci. 2005, 61, 1129–1132. [Google Scholar] [CrossRef] [PubMed]
- Omkar. Suitability of different foods for a generalist ladybird, Micraspis discolor (Coleoptera: Coccinellidae). Int. J. Trop. Insect Sci. 2006, 26, 35–40. [Google Scholar] [CrossRef]
- Triltsch, H. Food remains in the guts of Coccinella septempunctata (Coleoptera: Coccinellidae) adults and larvae. Eur. J. Entomol. 1999, 96, 355–364. [Google Scholar]
- Hemptinne, J.L.; Desprets, A. Pollen as a spring food for Adalia bipunctata. In Ecology of Aphidophaga; Hodek, I., Ed.; Academia, Prague and Dr. W. Junk, Dordrecht, 1986; pp. 29–35.
- Berkvens, N.; Bonte, J.; Berkvens, D.; Deforce, K.; Tirry, L.; Clercq, P.D. Pollen as an alternative food for Harmonia axyridis. BioControl. 2008, 53, 201–210. [Google Scholar] [CrossRef]
- Xia, P.-L.; Wang, X.-F.; Feng, Y.; Huang, Y.; Ji, F.; Cui, W.-X.; Qiao, B.-M. 2024. Effect of pollen consumption on development and intraguild predation of two predatory coccinellidae. J. Asia-Pacific Entomol. 2024, 27. [Google Scholar] [CrossRef]
- Lima, M.S.; Pontes, W.J.T.; Nobrega, R.L. Pollen did not provide suitable nutrients for ovary development in a ladybird Brumoides foudraii (Coleoptera: Coccinellidae). Diversitas J. 2020, 5, 1486–1494. [Google Scholar] [CrossRef]
- Manjunath, T.M. Rice moth, Corcyra cephalonica (Lepidoptera, Pyralidae) – A boon for biocontrol as a factitious host for mass production of parasitoids and predators. J. Biol. Control, 2023, 37, 01–05. [Google Scholar] [CrossRef]
- Ali, S.; Peng, J.; Liang, J.F.; Huang, C.; Xie, Y.-H.; Wang, X. Changes in life history parameters and transcriptome profile of Serangium japonicum associated with feeding on natural prey (Bemisia tabaci) and alternate host (Corcyra cephalonica eggs). BMC Genomics. 2023, 24, 112. [Google Scholar] [CrossRef] [PubMed]
- Poorani, J. Coelophora inaequalis. In An illustrated guide to lady beetles (Coleoptera: Coccinellidae) of the Indian Subcontinent. Part 1. Tribe Coccinellini, pp. 1–307 in Zootaxa (Vol. 5332, Number 1, pp. 106–107). Zenodo. 2023.
- Peck, S.; Thomas, M. A distributional checklist of the beetles (Coleoptera) of Florida. Arthropods of Florida and Neighboring Land Areas. Volume 16. Florida Department of Agriculture and Consumer Services, Gainesville. 1998, 180 pp.
- Mora, J.G.; Gapud, V.P.; Velasco, L.R.I. Life history and voracity of Coelophora inaequalis (Fabricius) (Coleoptera: Coccinellidae) on Aphis craccivora Koch (Hemiptera: Aphididae). Philippine-Entomologist (Philippines). 1995, 9, 523–553. [Google Scholar]
- Michaud, J.P. On the assessment of prey suitability in aphidophagous coccinellidae. European J. Entomol. 2005, 102, 385–390. [Google Scholar] [CrossRef]
- Ding, X.L.; Yao, F.L.; Zheng, Y.; Lu, X.S.; He, Y.X. Effect of two alternative prey species on development and fecundity of Serangium japonicum (Coleoptera: Coccinellidae). Chin. J. Biol. Cont. 2019, 35, 855–860. [Google Scholar]
- Michaud, J.P. Development and reproduction of lady beetles (Coleoptera: Coccinellidae) on the citrus aphids Aphis spiraecola Patch and Toxoptera citricida (Kirkaldy) (Homoptera: Aphididae). Biol. Control. 2000, 18, 287–297. [Google Scholar] [CrossRef]
- He, X.; Sigsgaard, L. A floral diet increases the longevity of the coccinellid Adalia bipunctata but does not allow molting or reproduction. Front. Ecol. Evo. 2019, 7, 6. [Google Scholar] [CrossRef]
- Seagraves, M.P. Lady beetle oviposition behavior in response to the trophic environment. Biol. Control. 2009, 51, 313–322. [Google Scholar] [CrossRef]
- Hodek, I.; Honek, A. Ecology of Coccinellidae. Kluwer Academic Publishers, Dordrecht. 1996. [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



