
Submitted:
31 July 2025
Posted:
31 July 2025
You are already at the latest version
Abstract
The objective was to analyze the phylogeny and morphology of the Glomeromycetes on the order level hitherto consisting of Glomerales, Entrophosporales, Diversisporales and Gigasporales. The updated data sets show that Pacisporaceae is near to Gigasporales, but without support to place this family in Gigasporales or Diversisporales, while the Acaulosporaceae and Sacculosporaceae clades clearly separate from Diversisporaceae. The phylogeny indicates that Pacisporaceae should be transferred here to the new order Pacisporales. Morphologically, Pacisporales differ from all families in the Gigasporales by type of spore formation (on subtending hyphae instead on suspensors (= bulbi), by formation of vesicular-arbuscular mycorrhiza, and by germination characteristics. The phylogenetic analysis supports the separation of Acaulosporaceae and Sacculosporaceae from Diversisporales, from each other, and from all other Glomeromycetes orders. These two families are here transferred to Acaulosporales and Sacculosporales. Acaulosporales are characterized by spore formation in a clear distance of 50-250 µm from globose termini of sporiferous saccules and by the diagnostic, ‘beaded’ inner wall. In Sacculosporales, there is no distance between the spores and the saccule termini, and the inner wall is never ‘beaded’. Additionally, four new families are separated within the Glomerales, and one new family within the Diversisporales.
Keywords:
molecular phylogeny
; systematics
; classification
; arbuscular mycorrhiza
; vesicular-arbuscular
; VAM
; AM fungi
1. Introduction
The Glomeromycetes Caval.-Sm. were the first class described in the arbuscular mycorrhizal fungi (AMF), initially covering not only the typical AM fungi whose intraradical fungal structures stain intensively blue in staining reagents like Trypan blue or ink [1,2]. The most ancestral AM fungal clades e.g. [3], however, were separated and have been counted within the Archaeosporomycetes Sieverd., G.A. Silva, B.T. Goto & Oehl and Paraglomeromycetes Oehl, G.A. Silva, B.T. Goto & Sieverd. [4,5]. These three fungal classes constitute together the subphylum Glomeromycotina Spatafora & Stajich [6] and the phylum Glomeromycota C. Walker & A. Schüssler [7] of the Mucoromyceta [8]. The Glomeromycetes currently comprise four fungal orders: Glomerales J.B. Morton & Benny [9,10], Entrophosporales Błaszk., Sánchez-García, B.T. Goto & Magurno [11], Diversisporales C. Walker & A. Schüssler [12] and Gigasporales Sieverd., G.A. Silva, B.T. Goto & Oehl [4]. Archaeosporales C. Walker & A. Schüssler and Paraglomerales C. Walker & A. Schüssler complete the six AMF orders currently widely accepted within the Glomeromycota [5,7]. Furthermore, the Glomerales currently comprise five fungal families: Glomeraceae [13], Septoglomeraceae, Sclerocystaceae, Kamienskiaceae and Dominikiaceae [14]. Entrophosporales comprise the single family Entrophosporaceae [11,15], Gigasporales comprise the families Gigasporaceae, Intraornatosporaceae, Dentiscutataceae, Scutellosporaceae and Racocetraceae [4,5], and Diversisporales the families Diversisporaceae, Acaulosporaceae, Sacculosporaceae and Pacisporaceae [5,7]. Finally, the order Archaeosporales comprises the families Archaeosporaceae, Ambisporaceae, Geosiphonaceae and Polonosporaceae [3,16,17], while Paraglomerales comprise the two families Paraglomeraceae and Pervetustaceae [3,18].
So far, the greatest diversity of morphological characters in the Glomeromycetes were described within the Diversisporales of all six described AMF orders [e.g, 4,7], which was initially also expressed in the name of the order. On the one hand, these great differences were clearest for AM fungi forming spores on sporogeneous cells [9,19,20]. Phylogenetic dissimilarities have been the most prominent argument to firstly separate this fungal group from the Diversisporales [4]. On the other hand, several other, i.e. glomoid, acaulosporoid and entrophosporoid AM fungal clades have hitherto continued within the Diversisporales, on various phylogenetic positions, although also for a few of these clades prominent phylogenetic dissimilarities have increasingly been recognized. Morphological key characters can be named for specific AM fungal groups, such as the spore formation in the Sacculosporaceae, the diagnostic beaded germinal wall in the Acaulosporaceae or the typical subtending hyphal features of Pacispora, Diversispora and Redeckera [20,21,22,23,24,25].
The aim of this study was to double-check the data bases and the different major phylogeny clades, in order to investigate, if there is enough substance i) to separate Acaulosporaceae, Sacculosporaceae and Pacisporaceae from the Diversisporales, and ii) to continue within this order with multiple families originating from the type family Diversisporaceae. Therefore, we reviewed the phylogeny and the morphological major characters of all these taxa. Hereafter, firstly the phylogenetic differences are presented between all major clades of the Glomeromycetes. Secondly, the morphological characters and major differences are highlighted for all the major clades within this fungal class. Finally, in the Taxonomy section, new taxa are presented, where justified by molecular phylogeny, and when additionally, these taxa are supported by morphological arguments.
2. Materials and Methods
2.1. Phylogenetic Analyses
The phylogeny was reconstructed using an alignment (dataset) generated with AM fungal sequences from species of Glomeromycetes (Supplementary Material, Spreadsheet S1). The dataset was used to reconstruct the Glomeromycetes phylogeny based on partial SSU, 5.8S, and partial LSU nrDNA (SSU-5.8S-LSU). Paraglomus brasilianum (Spain & J. Miranda) J.B. Morton & D. Redecker was included as outgroup. In the alignment were used sequences with at least the 5.8S, and partial LSU nrDNA. The dataset was aligned in Mafft [26], using the default parameters. Bayesian (BI) analysis (two runs over 1 × 107 generations, with a sample frequency of 1,000 and a burnin value of 25%) was performed in MrBayes 3.2.7a [27], launched from Cipres Science Gateway portal [28]. Maximum likelihood (ML) analysis (1000 bootstrap) was performed in RaxML-NG [29,30]. Prior to phylogenetic analyses, the model of nucleotide substitution was estimated using ModelTest-NG [31]. The best model for both analyses was GTR + I + G.
2.2. Morphological Analyses
We morphologically analyzed all species belonging to Glomeromycetes deposited in those mycological herbaria, which harbor the majority of the type material of AM fungi, i.e. OSC (types deposited between 1968 to 2005), Z + ZT (types deposited between 2002 and 2024), and URM (types deposited between 2008 and 2024); additionally, AM fungal type material were analyzed from private collections generated by Sieverding, Oehl, Trappe, Błaszkowski and McGee. The Hall and Abbott [32] photographic slide collection was also reviewed. Older specimens (mounted on microscopic slides prior to 1990) were mostly mounted in lactophenol, while others were fixed with polyvinyl alcohol–lactic acid–glycerol (PVLG) or in a mixture of PVLG + Melzer’s reagent, which after 1990 are the principal fixing media [33]. Newly mounted spores and sporocarps from collections or from cultures were fixed using the latter two fixing media or occasionally also in a mixture of 1:1 lactic acid to water, in Melzer’s reagent and in water [34]. When available, spores freshly isolated from soils or bait cultures were also mounted and analyzed. All spore observations and all information on spore characteristics are based on spores extracted from soil, from trap cultures or from single or multiple spore-derived pure cultures. No information is provided from in vitro cultured materials. Spore wall terminology follows the nomenclature of Walker [35], Stürmer and Morton [36], and Błaszkowski [37]. Analyses of the spore walls, germination structures and all other mycorrhizal structures were performed using compound microscopes at 100–1000× magnification (Zeiss Axioplan, Oberkochen, Germany; Leika DFC 295; Wetzlar, Germany). For this paper, all original species descriptions and published species emendations were also considered and thoroughly studied.
3. Results
3.1. Molecular Phylogeny
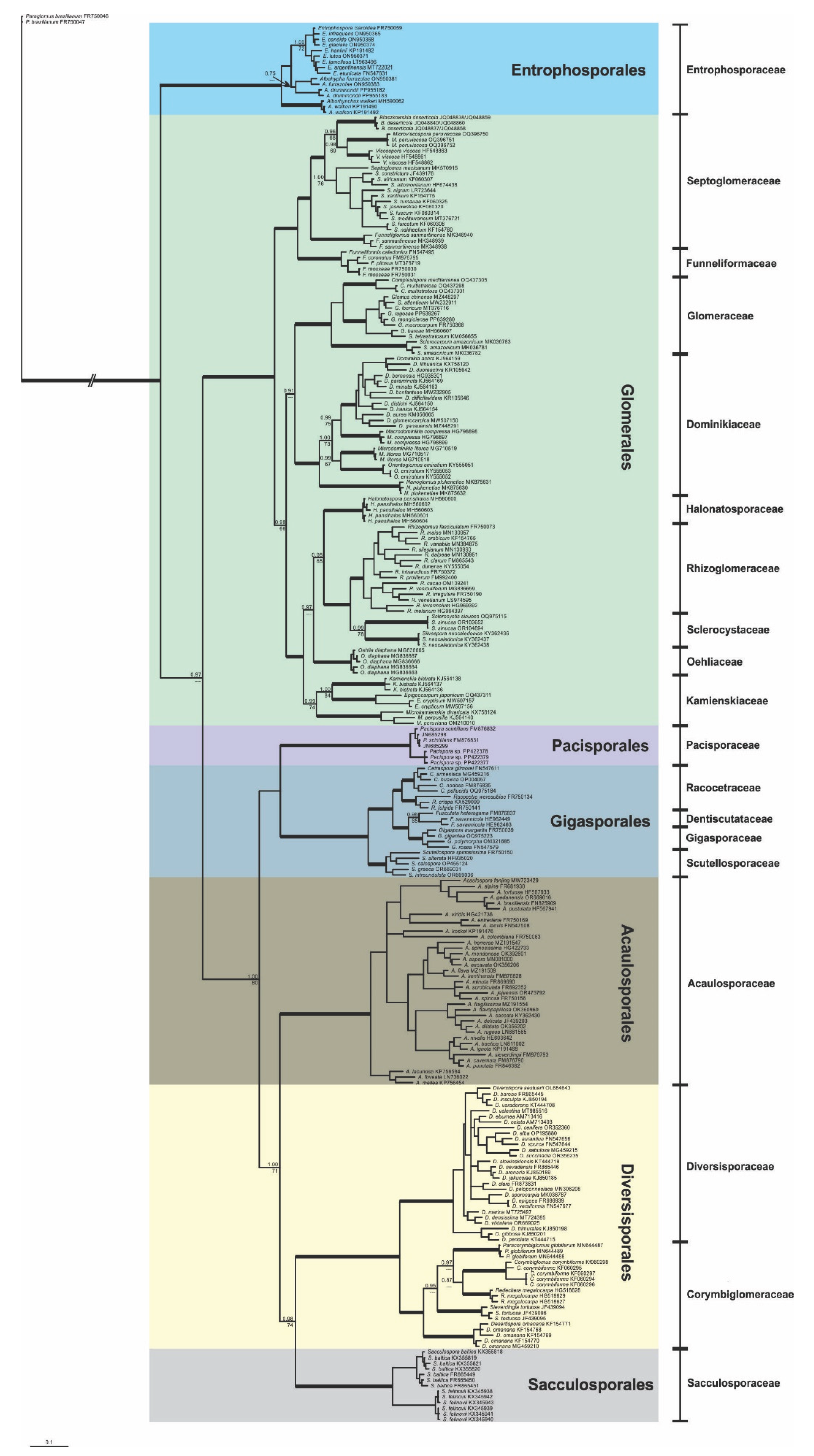
The phylogenetic tree for the Glomeromycetes showed seven different clades (Figure 1) with full support for BI and ML analyses for the four well-established orders: Diversisporales, Entrophosporales, Gigasporales, and Glomerales, and three new orders, Acaulosporales, Pacisporales, and Sacculosporales. From these orders, three have just a single monogeneric family (Acaulosporales - Acaulosporaceae – Acaulospora; Pacisporales - Pacisporaceae - Pacispora; and Sacculosporales - Sacculosporaceae - Sacculospora).
Our results confirm Entrophosporales with a single family (Entrophosporaceae), and three genera (Albahypha, Alborhynchus, and Entrophospora). However, there was just a low BI support to maintain Albahypha drummonddi and A. furrazolae together. The families and genera from Gigasporales, represented in our tree, were confirmed phylogenetically by our results with strong support (≥0.95 for BI and ≥90% for ML analyses), except for ML analysis to Dentiscutataceae and Fuscutata, with just 65% of support. Two well supported clades, representing family level, were found in Diversisporales (Diversisporaceae and a new family, Corymbiglomeraceae). Diversisporaceae has a single genus (Diversispora), and showed full support in our phylogenetic analysis. Corymbiglomeraceae was fully supported by BI and presented a ML support of 94%. The phylogeny recovered five genera in Corymbiglomeraceae (Corymbiglomus, Desertispora, Paracorymbiglomus, Redeckera, and Sieverdingia) with full support in all phylogenetic analyses. Glomerales showed nine clades strongly supported on the family level (Dominikiaceae, Glomeraceae, Kamienskiaceae, Sclerocystaceae, Septoglomeraceae, and four new families, Funneliformaceae, Halonatosporaceae, Rhizoglomeraceae, and Oehliaceae). All families and all genera in Glomerales, represented in our tree, presented support ≥0.99 for BI and ≥90% for ML analyses, except for ML analysis in Kamienskiaceae (74%), Sclerocystaceae (78%), and Septoglomus (76%). Considering our phylogenetic findings, three new orders and five new families needed to be described in Glomeromycetes.
Figure 1.
Phylogenetic tree from partial SSU, 5.8s and partial LSU nrDNA sequences of Glomeromycetes. Sequences are labeled with their database accession numbers. Support values from Bayesian inference (BI) and maximum likelihood (ML) are shown only at the genera level or above. Only support values of at least 65% are shown. Thick branches represent clades with more than 90% support in all analyses. The tree was rooted by Paraglomus brasilianum.
Figure 1.
Phylogenetic tree from partial SSU, 5.8s and partial LSU nrDNA sequences of Glomeromycetes. Sequences are labeled with their database accession numbers. Support values from Bayesian inference (BI) and maximum likelihood (ML) are shown only at the genera level or above. Only support values of at least 65% are shown. Thick branches represent clades with more than 90% support in all analyses. The tree was rooted by Paraglomus brasilianum.
3.2. Morphological Analyses
In this study, no new morphological findings are reported in the Glomeromycetes, since all described genera and families have been well defined in the last two decades during their first descriptions already or, when their descriptions were emended based on the current knowledge and state-of-the-art. Here, some outstanding characters of genera, families or orders are highlighted.
The phylogenetic analyses suggested seven AM fungal orders in Glomeromycetes. Three of them form exclusively (Glomerales and Pacisporales) or almost exclusively (Entrophosporales) spores on subtending hyphae. Glomerales and Entrophosporales differentiate mono-walled, so-called glomoid spores (SW), while Pacispora form bi-walled spores, an outer spore wall and an inner germinal wall (OW & IW). Glomerales form spores, which generally are in same or similar color as the terminal or intercalary formed spores, while the subtending hyphae in Entrophosporales are always hyaline, subhyaline or white independent of the spore color. In Entrophosporales, so far only the type species has been known to form not only glomoid, but within the neck of sporiferous saccules also so-called entrophosporoid, bi-walled spores, with OW & IW (Table S1).
Two orders differentiate exclusively spores on or within the neck of sporiferous saccules, with two or three walls (Sacculosporales and Acaulosporales, respectively). Acaulosporales species form spores on or within the neck of sporiferous saccules, in more or less variable distance, which are ca. 1-2 fold as long as the spores size formed in/on the neck. Acaulosporales species have three spore walls (OW & IW, and an additional middle wall, MW), of which IW differentiate a typical, granular, so-called ´beaded´ wall layer on its outer surface. Sacculosporales species exclusively form spores within sporiferous saccules, but without distance directly beneath the saccule terminus, and the spores lack the granular, beaded wall layer.
Gigasporales differentiate spores exclusively on sporogenous cells. Species of this order differentiate genus-specifically 1-4 spore walls (OW, MW, (rarely MW2) & IW), and genus-specifically germinate through germ warts (if solely mono-walled) or through prominent, pigment-less to light-yellow or well-pigmented brown germination shields, consisting of one, two, a few or multiple germination lobes/compartments (if > one spore wall). Pacisporales and Gigasporales share one important morphological character, which they do not share with any of the other glomeromycotan orders, i.e. the formation of extraradical and, more rarely, also intraradical auxiliary cells e.g. [37]. This is one outstanding argument to separate the Pacisporaceae from the Diversisporales. Pacisporales and Gigasporales mainly separate by spore formation type on subtending hyphae or on sporogenous cells, by spore germination through the germination shield center (perm pore) or through visible germ tube initiations in the periphery of prominent germination shields, respectively, if germination is not very different as through germ warts direct from the spore wall in the special case of Gigaspora. Moreover, according to Błaszkowski [37], Pacisporales form typical versicular-arbuscular mycorrhiza, while Gigasporales solely have arbuscular mycorrhiza, but intraradical vesicle formation is not (yet) known.
Lastly, Diversisporales continue to be the most diverse order forming glomoid spores (glomoid = on subtending hyphae) with genus-specifically one or two walls, or rarely acaulosporoid or entrophosporoid spores with two walls. Importantly, the specific characteristics of all the genera within this order can be easily distinguished from all other genera within the Glomeromycota [14,15,24,38,39].
The phylogenetic advances suggested a separation of the Diversisporales into two families, based on the two major clades recognized within this order (Figure 1). The newly revised family Diversisporaceae comprise now solely the diversisporoid species sensu stricto, and two species with formation of bi-walled spores on or within the neck of sporiferous saccules, similar, but different to those of Sacculospora and Entrophospora infrequens [5,25]. In the new family Corymbiglomeraceae, besides the type genus Corymbiglomus, also Paracorymbiglomus, Desertispora, Redeckera, and Sieverdingia are included [39]. However, the phylogenetic and morphological data bases of these taxa are still exiguous, so that a clearer separation of these AMF genera hitherto is not yet feasible.
Finally, within the Glomerales, also the further separation of four new families, based on the phylogenetic findings, is supported by morphology. Moreover, Silva et al. [14] have already suggested Halonatosporaceae, Oehliaceae and Rhizoglomeraceae as possible new families. Shortly, Oehliaceae and Rhizoglomeraceae frequently form spore agglomerations within the roots. Oehliaceae species generally form a septum closing the spore base, while in Rhizoglomeraceae the spore pores are not closed at the base, but often have several septa within long subtending hyphae in rather long distance from the spore base (> 25 µm). Halonatosporaceae can be characterized by spores forming a halo with radiate columns. Finally, Funneliformaceae separate easily from all other species in the Septoglomeraceae and other families in the Glomerales and Glomeromycetes, by i) their spore formation on funnel-shaped subtending hyphae, thus ii) wide pores at the spore bases, iii) and wide, broad septa positioned within a short distance from the spore base or rarely also at the spore base, and iv) spore formation in clusters or small sporocarps, which is not known for the morphologically and phylogenetically members of the most closely related family (Septoglomeraceae), especially to the morphologically most closely related genus Funneliglomus.
Glomeromycetes now comprise twenty families of which nine belong to the Glomerales and five to the Gigasporales. Two families are separated within the Diversisporales, while the lasting four orders are represented each by one family (Table S2).
3.3. Taxonomy of the Glomeromycetes
Glomeromycetes Caval.-Sm., Biol. Rev. Cambridge Philos. Soc. 73: 246. 1998.
Mycobank MB 90168
Emended description: see Oehl et al. [4]
Type order: Glomerales J.B. Morton and Benny, emend. Błaszk., B.T. Goto, and Magurno
Other orders:
Acaulosporales Sieverd., Silva G.A. & Oehl,
Entrophosporales Błaszk., Sánchez-García, B.T. Goto & Magurno,
Gigasporales Sieverd., G.A. Silva, B.T. Goto & Oehl,
Pacisporales Oehl, Sieverd., G.A. Silva,
Sacculosporales Silva G.A., Sieverd. & Oehl.
Glomerales J.B. Morton and Benny, Mycotaxon 37: 473. 1990.
MycoBank MB 90425
Type family: Glomeraceae Piroz. & Dalpé emend. Oehl, G.A. Silva & Sieverd.
Other families:
Dominikiaceae G.A. Silva, Sieverd. & Oehl,
Funneliformaceae Oehl, Sieverd. & G.A. Silva,
Halonatosporaceae Oehl, Sieverd. & G.A. Silva,
Kamienskiaceae G.A. Silva, Sieverd. & Oehl,
Oehliaceae Sieverd. & G.A. Silva,
Rhizoglomeraceae Sieverd., G.A. Silva & Oehl,
Sclerocystaceae Oehl, G.A. Silva & Sieverd.,
Septoglomeraceae Oehl, G.A. Silva & Sieverd.
Glomeraceae Piroz. & Dalpé, Symbiosis 7: 19. 1989.
MycoBank MB 82026
Type genus: Glomus Tul. & C. Tul.
Other genera:
Complexispora Błaszk., B.T. Goto, Niezgoda & Magurno,
Sclerocarpum B.T. Goto, Błaszk., Niezgoda, Kozłowska & Jobim,
Simiglomus Sieverd., G.A. Silva & Oehl.
Glomus Tul. & C. Tul., G. bot. ital. 2(1): 63. 1845. [1844].
MycoBank MB 20244
Description: see Silva et al. [14]
Type species: Glomus macrocarpum Tul & C. Tul., G. bot. ital. 2(1): 63. 1845. [1844].
MycoBank MB 240247
≡ Endogone macrocarpa (Tul. & C. Tul.) Tul. & C. Tul., Fungi Hypog.: 182. 1851.
MycoBank MB 218537
≡ Endogone guttulata E. Fisch., Ber. Schweiz. Bot. Ges. 32: 13. 1923.
MycoBank MB 266684
≡ Endogone nuda Petch., Ann. R. Bot. Gdns Peradeniya 9: 322. 1925.
MycoBank MB 223365
≡ Endogone pampaloniana Bacc., Nuovo Giorn. Bot. Ital., n.s. 10: 90. 1903.
MycoBank MB 223215
≡ Paurocotylis fulva var. zealandica Cooke, Grevillea 8: 59. 1879.
MycoBank MB 522257
Complexispora Błaszk., B.T. Goto, Niezgoda & Magurno, Mycol. Prog. 22(5, no. 34): 7. 2023.
MycoBank MB 847607
Type species: Complexispora multistratosa Błaszk., B.T. Goto, Niezgoda & Magurno,
Mycol. Prog. 22(5, no. 34): 6. 2023.
MycoBank MB 847608
Sclerocarpum B.T. Goto, Błaszk., Niezgoda, Kozłowska & Jobim, Mycol. Prog. 18(3): 375. 2019.
MycoBank MB 828316
Type species: Sclerocarpum amazonicum B.T. Goto, Błaszk., Niezgoda, Kozłowska & Jobim, Mycol. Prog. 18: 377. 2019.
MycoBank MB 828317
Simiglomus Sieverd., G.A. Silva & Oehl, Mycotaxon 116: 104. 2011.
MycoBank MB 518435
Description: see Oehl et al. [24]
Type species: Simiglomus hoi (S.M. Berch & Trappe) G.A. Silva, Oehl & Sieverd., Mycotaxon 116: 104 (2011).
MycoBank MB 518461
Basionym: Glomus hoi S.M. Berch & Trappe, Mycologia 77: 654. 1985.
MycoBank MB 105333
Dominikiaceae G.A. Silva, Sieverd. & Oehl, Taxonomy 4: 773. 2024.
MycoBank MB 855469
Description: see Silva et al. [14]
Type genus: Dominikia Błaszk., Chwat & Kovács
Other genera:
Delicatispora Błaszk., Niezgoda & B.T. Goto,
Macrodominikia Oehl, Sieverd. & G.A. Silva,
Microdominikia Oehl, Corazon-Guivin & G.A. Silva,
Nanoglomus Corazon-Guivin, G.A. Silva & Oehl,
Orientoglomus G.A. Silva, Oehl & Corazon-Guivin.
Dominikia Błaszk., Chwat & Kovács, Nova Hedwigia 100(1–2): 228. 2014 [2015].
MycoBank MB 808255
Emended description: see Silva et al. [14]
Type species: Dominikia minuta (Błaszk., Tadych & Madej) Błaszk., Chwat & Kovács,
Nova Hedwigia 100: 230. 2014 [2015].
MycoBank MB 808256
Basionym: Glomus minutum Błaszk., Tadych & Madej. Mycotaxon 76: 189. 2000.
Delicatispora Błaszk., Niezgoda & B.T. Goto, MycoKeys 112: 261. 2025.
Mycobank MB 856187
Description: see Błaszkowski et al. [38]
Type species: Delicatispora indica (Błaszk., Wubet & Harikumar) Błaszk., Niezgoda & B.T. Goto, MycoKeys 112: 261. 2025.
Mycobank MB 856189
Basionym: Glomus indicum Błaszk., Wubet & Harikumar, Botany 88: 134. 2010.
Mycobank MB 515245.
≡ Dominikia indica (Błaszk., Wubet & Harikumar) Błaszk., G.A. Silva & Oehl, Nova Hedwigia 101 (1-2): 71. 2014.
Mycobank MB 808258
Macrodominikia Oehl, Sieverd. & G.A. Silva, Taxonomy 4: 774. 2024.
MycoBank 855470
Type species: Macrodominikia compressa (Sieverd., Oehl, Palenz., Sánchez-Castro & G.A. Silva) Oehl, Sieverd. & G.A. Silva, Taxonomy 4: 774. 2024.
MycoBank MB 855471
Basionym: Glomus compressum Sieverd., Oehl, Palenz., Sánchez-Castro & G.A. Silva. Nova Hedwigia 99: 433. 2014.
MycoBank MB 807530
≡ Dominikia compressa (Sieverd., Oehl, Palenz., Sánchez-Castro & G.A. Silva) Oehl, Palenz., Sánchez-Castro & G.A. Silva. Nova Hedwigia. 101: 71. 2014 [2015].
MycoBank MB 809861
Microdominikia Oehl, Corazon-Guivin & G.A. Silva, Mycol. Prog. 18(12): 1400. 2019.
MycoBank MB 831098
Description: see Silva et al. [14]
Type species: Microdominikia litorea (Błaszk. & Kozłowska) Oehl, Corazon-Guivin & G.A. Silva, Mycol. Prog. 18(12): 1400. 2019.
MycoBank MB 831099
Basionym: Dominikia litorea Błaszk. & Kozłowska, Phytotaxa 338(3): 246. 2018.
MycoBank MB 823832
Nanoglomus Corazon-Guivin, G.A. Silva & Oehl, Mycol. Prog. 18(12): 1398. 2019.
Description: see Silva et al. [14]
MycoBank MB 831096
Type species: Nanoglomus plukenetiae Corazon-Guivin, G.A. Silva & Oehl, Mycol.
Prog. 18(12): 1398. 2019.
MycoBank MB 831097
Orientoglomus G.A. Silva, Oehl & Corazon-Guivin, Mycol. Prog. 18(12): 1400. 2019.
MycoBank MB 831100
Description: see Silva et al. [14]
Type species: Orientoglomus emiratium (Błaszk., Kozłowska, Mullath, AlDhaheri & Al-Yahya’ei) G.A. Silva, Oehl & Corazon-Guivin, Mycol. Prog. 18(12): 1403. 2019.
MycoBank MB 831101
Basionym: Dominikia emiratia Błaszk., Kozłowska, Mullath, AlDhaheri & Al-Yahya’ei, Botany 95(7): 632. 2017.
MycoBank MB 819815
Funneliformaceae Oehl, Sieverd. & G.A. Silva, fam. nov.
Mycobank MB 860129
Description: Spores formed within soil or rarely roots, singly or sometimes in sporocarps with one, a few to several spores per sporocarp only; the conspicuous SH is concolorous with spore wall color (or slightly lighter in color), SH is species-specific and generally funnel-shaped to rarely cylindrical. Wall differentiation and pigmentation may continue over long distances from the spore base (often > 50–250 μm), then mycelium generally is hyaline. Pore regularly closed by a conspicuous septum in some distance from spore base, or with some genera at spore base. Septum arises species-specifically from the structural wall layer, from an additional adherent innermost, (semi-)flexible lamina, or from both but not by introverted wall thickening, which is lacking. Forming typical vesicular-arbuscular mycorrhiza, with mycorrhizal structures that stain blue to dark blue with trypan blue.
Type genus: Funneliformis C. Walker & A. Schüssler, emend. Oehl, G.A. Silva & Sieverd.
Funneliformis C. Walker & A. Schüssler, The Glomeromycota—a species list with new families and genera: 13. 2010.
MycoBank MB 542894
Type species: Funneliformis mosseae (T.H. Nicolson & Gerd.) C. Walker & A. Schüssler, The Glomeromycota—a species list with new families and genera: 13. 2010.
MycoBank MB 542895
Basionym: Endogone mosseae T.H. Nicolson & Gerd., Mycologia 60: 314. 1968.
MycoBank MB 330367
≡ Glomus mosseae (T.H. Nicolson & Gerd.) Gerd. & Trappe, Mycol. Mem. 5: 40. 1974.
MycoBank MB 314604
Halonatosporaceae G.A. Silva, Oehl & Sieverd., fam. nov.
Mycobank MB 860134
Description: Spores formed in loose clusters or singly in soil and frequently in roots. SH wall continuous with the SW and for a certain distance concolorous with the SW or slightly lighter in color. Outer SW layer strongly swells in PVLG, forming a halo with radiate columns. SH cylindrical or slightly flared, sometimes slightly constricted at spore base, straight or slightly curved. Pore open or closed by a septum at spore base. SW shows more than one, generally two to three (and up to five) distinct layers. Forming vesicular–arbuscular mycorrhizae, whose fungal structures stain blue to dark blue with trypan blue.
Type genus: Halonatospora Błaszk., Niezgoda, B.T. Goto & Kozłowska
Halonatospora Błaszk., Niezgoda, B.T. Goto & Kozłowska, Botany 96(11): 743. 2018.
MycoBank MB 826963
Type species: Halonatospora pansihalos (S.M. Berch & Koske) Błaszk., Niezgoda, B.T. Goto & Kozłowska, Can. J. Bot. 96(11): 743 (2018)
MycoBank MB 826964
Basionym: Glomus pansihalos S.M. Berch & Koske. Mycologia 78: 832, 1986.
MycoBank MB 358213
Kamienskiaceae G.A. Silva, Sieverd. & Oehl, Taxonomy 4: 772. 2024.
MycoBank MB 855468
Description: see Silva et al. [14]
Type genus: Kamienskia Błaszk., Chwat & Kovács
Other genera:
Epigeocarpum Błaszk., B.T. Goto, Jobim, Niezgoda & Marguno,
Microkamienskia Corazon-Guivin, G.A. Silva & Oehl
Kamienskia Błaszk., Chwat & Kovács, Nova Hedwigia 100(1–2): 230. 2014 [2015].
MycoBank MB 808260
Description: see Silva et al. [14]
Type species: Kamienskia bistrata (Błaszk., D. Redecker, Koegel, Symanczik, Oehl & Kovács) Błaszk., Chwat & Kovács, Nova Hedwigia 100(1–2): 230. 2014 [2015].
MycoBank MB 808261
Basionym: Glomus bistratum Błaszk., D. Redecker, Koegel, Symanczik, Oehl & Kovács,
Botany 87: 267. 2009.
MycoBank MB 512540
Epigeocarpum Błaszk., B.T. Goto, Jobim, Niezgoda & Marguno, Frontiers in Microbiology 12(no. 655910): 10. 2021.
MycoBank MB 838879
Type species: Epigeocarpum crypticum Jobim, Błaszk., Niezgoda, Magurno & B.T. Goto, Frontiers Microbiol. 12(no. 655910): 14. 2021.
MycoBank MB 838880
Microkamienskia Corazon-Guivin, G.A. Silva & Oehl, Nova Hedwigia 109: 359. 2019. MycoBank MB 830814
Description: see Silva et al. [14]
Type species: Microkamienskia perpusilla (Błaszk. & Kovács) Corazon-Guivin, G.A. Silva & Oehl. Nova Hedwigia 109: 361. 2019.
MycoBank MB 830815
Basionym: Glomus perpusillum Błaszk. & Kovács. Mycologia 101: 249. 2009.
MycoBank MB 512346
≡ Kamienskia perpusilla (Błaszk. & Kovács) Błaszk., Chwat & Kovács. Nova Hedwigia 100: 231. 2015.
MycoBank MB 808264
Oehliaceae Sieverd. & G.A. Silva, fam. nov.
Mycobank MB 860132
Description: Spores formed in loose sporocarps, in clusters or singly in soil and frequently also intraradical singly or in loose sporocarps and clusters. When formed in sporocarps, they are not organized around a central plexus of hyphae. SH wall continuous with the SW and for a certain distance concolorous with the SW or slightly lighter in color. SH cylindrical or seldom slightly funnel-shaped at spore base. Pore at spore base regularly closed by a septum. SW shows more than one, generally two to three (and up to five) distinct layers. Forming vesicular–arbuscular mycorrhizae, fungal structures stain blue to dark blue with trypan blue.
Type genus: Oehlia Błaszk., Kozłowska & Dalpe
Oehlia Błaszk., Kozłowska & Dalpe, Nova Hedwigia 107(3–4): 507. 2018.
MycoBank MB 824689
Type species: Oehlia diaphana (J.B. Morton & C. Walker) Błaszk., Kozłowska & Dalpe,
Nova Hedwigia 107(3–4): 507. 2018.
MycoBank MB 824693
Basionym: Glomus diaphanum J.B. Morton & C. Walker. Mycotaxon 21: 433, 1984.
MycoBank MB 106161
Rhizoglomeraceae Sieverd., G.A. Silva & Oehl, fam. nov.
Mycobank MB 860131
Description: Spores formed in loose sporocarps, in clusters or singly in soil and frequently also intraradical in loose sporocarps and clusters. When formed in compact sporocarps, they are not organized around a central plexus of hyphae. SH wall continuous with the SW and for a certain distance concolorous with the SW or slightly lighter in color. SH cylindrical or seldom slightly funnel-shaped at spore base. Pore at spore base regularly open, rarely closed by a septum. SW shows more than one, generally two to three (and up to five) distinct layers, of which one or several of the outermost layers may easily separate when pressure is applied to spores. Forming vesicular–arbuscular mycorrhizae, the fungal structures of which stain blue to dark blue in trypan blue.
Type genus: Rhizoglomus Sieverd., G.A. Silva & Oehl
Rhizoglomus Sieverd., G.A. Silva & Oehl, Mycotaxon 129: 377. 2015.
MycoBank MB 803191
Description: see Rhizoglomeraceae and Silva et al. [14]
Type species: Rhizoglomus intraradices (N.C. Schenck & G.S. Sm.) Sieverd., G.A. Silva & Oehl, Mycotaxon 129(2): 378. 2015 [2014].
MycoBank MB 803192
Basionym: Glomus intraradices N.C. Schenck & G.S. Sm., Mycologia 74: 78. 1982.
MycoBank MB 110704
≡ Rhizophagus intraradices (N.C. Schenck & G.S. Sm.) C. Walker & A. Schüssler, The Glomeromycota—a species list with new families and genera: 19. 2010.
MycoBank MB 542910
Sclerocystaceae Oehl, G.A. Silva, & Sieverd., Taxonomy 4: 770. 2024.
MycoBank MB 855467
Description: see Silva et al. [14]
Type genus: Sclerocystis Berk. & Broome
Other genera:
Silvaspora Błaszk., Niezgoda, B.T. Goto, Crossay & Magurno,
Parvocarpum Magurno.
Sclerocystis Berk. & Broome, J. Linn. Soc., Bot. 14(no. 74): 137. 1873. [1875].
MycoBank MB20512
Emended description: see Silva et al. [14]
Type species: Sclerocystis coremioides Berk. & Broome, J. Linn. Soc., Bot. 14(no. 74):
137. 1873. [1875].
MycoBank MB 213141
≡ Glomus coremioides (Berk. & Broome) D. Redecker & J.B. Morton, Mycologia 92:
284. 2000.
MycoBank MB 464612
Parvocarpum Magurno, MycoKeys 107: 283. 2024.
Emended description: see Silva et al. [14]
Type species: Parvocarpum badium (Oehl, Redecker & Sieverd.) Magurno, MycoKeys 107: 284. 2024.
MycoBank MB 853560
Basionym: Glomus badium Oehl, D. Redecker & Sieverd., J. Appl. Bot. Food Qual. 79: 39. 2005.
MycoBank MB 341387
≡ Funneliformis badius (Oehl, D. Redecker & Sieverd.) C. Walker & A. Schüssler, The Glomeromycota—a species list with new families and genera: 13. 2010.
MycoBank MB 541897
Silvaspora Błaszk., Niezgoda, B.T. Goto, Crossay & Magurno, Frontiers in Microbiology 12(no. 655910): 14. 2021.
MycoBank 838881
Description: see Silva et al. [14]
Type species: Silvaspora neocaledonica (D. Redecker, Crossay & Cilia) Błaszk., Niezgoda, B.T. Goto, Crossay & Magurno, Frontiers in Microbiology 12(no. 655910): 14 (2021)
MycoBank MB 838882
Basionym: Rhizophagus neocaledonicus D. Redecker, Crossay & Cilia, Mycol. Prog. 17: 739. 2018.
MycoBank MB 820537
≡ Rhizoglomus neocaledonicum (D. Redecker, Crossay & Cilia) Oehl, A. Turrini & Giovann., Mycol. Prog. 17: 1216 (2018).
MycoBank MB 827095
Septoglomeraceae Oehl, G.A. Silva, Sieverd., Taxonomy 4: 768. 2024.
Mycobank MB 855466
Emended description: Spores formed in soil and sometimes in roots, terminally on or intercalarily in hyphae, either singly, in loose spore clusters or in sporocarps. Spores with one mono- to multiple-layered wall. SH wall conspicuously continuous and concolorous with the SW or slightly lighter in color than the SW; SH funnel-shaped, cylindrical or constricted, rarely inflated; straight, curved or flared; forming typical vesicular–arbuscular mycorrhiza with mycorrhizal structures that stain blue to dark blue with trypan blue.
Type genus: Septoglomus Sieverd., G.A. Silva & Oehl
Other genera:
Blaszkowskia G.A. Silva & Oehl,
Funneliglomus Corazon-Guivin, G.A. Silva & Oehl,
Melanoglomus B.T. Goto, Błaszk., Sieverd., G.A. Silva & Oehl,
Microviscospora Oehl, Corazon-Guivin, Błaszk., B.T. Goto, Sieverd. & G.A. Silva,
Viscospora Sieverd., Oehl & G.A. Silva.
Septoglomus Sieverd., G.A. Silva & Oehl, Mycotaxon 116: 105. 2011.
MycoBank MB 518436
Emended description: see Oehl et al. [24]
Type species: Septoglomus constrictum (Trappe) Sieverd., G.A. Silva & Oehl. Mycotaxon 116: 105. 2011.
MycoBank MB 518462
Basionym: Glomus constrictum Trappe, Mycotaxon 6: 361. 1977.
MycoBank MB 314589
≡ Funneliformis constrictus (Trappe) C. Walker & A. Schüssler, The Glomeromycota— a species list with new families and genera: 14. 2010.
MycoBank MB 542904
Blaszkowskia G.A. Silva & Oehl, Mycol. Prog. 22(11, no. 74): 5. 2023.
MycoBank MB 847414
Description: see Silva et al. [14]
Type species: Blaszkowskia deserticola (Trappe, Bloss & J.A. Menge) Oehl & G.A.
Silva. Mycol. Prog. 22(11, no. 74): 5. 2023.
MycoBank MB 847415
Basionym: Glomus deserticola Trappe, Bloss & J.A. Menge, Mycotaxon 20: 123. 1984.
MycoBank MB 106847
≡ Septoglomus deserticola (Trappe, Bloss & J.A. Menge) G.A. Silva, Oehl & Sieverd.
Mycotaxon 116: 106. 2011.
MycoBank MB 518463
Funneliglomus Corazon-Guivin, G.A. Silva & Oehl, Sydowia 71: 19. 2019.
MycoBank MB 829266
Description: see Silva et al. [14]
Type species: Funneliglomus sanmartinense Corazon-Guivin, G.A. Silva & Oehl, Sydowia 71: 21. 2019
MycoBank MB 830216
Melanoglomus B.T. Goto, Błaszk., Sieverd., G.A. Silva & Oehl, Mycol. Prog. 2025. In review.
MycoBank MB 857542
Description: see Goto et al. [45]
Type species: Melanoglomus titan (B.T. Goto & G.A. Silva) B.T. Goto, Błaszk., Sieverd., G.A. Silva & Oehl
MycoBank MB 857543
Basionym: Septoglomus titan B.T. Goto & G.A. Silva, Mycotaxon 124: 105. 2013.
MycoBank MB564321
Microviscospora Oehl, Corazon-Guivin, Błaszk., B.T. Goto, Sieverd. & G.A. Silva, Mycol. Prog. 2025. In review.
MycoBank MB 857544
Description: see Goto et al. [45]
Type species: Microviscospora peruviscosa Oehl, Corazon-Guivin, Błaszk., B.T. Goto, Sieverd. & G.A. Silva
MycoBank MB 857544
Basionym: Viscospora peruviscosa Corazon-Guivin, G.A. Silva & Oehl, J. Appl. Bot. Food Quality - Angew. Botan. 96: 118. 2023.
MycoBank MB 857545
Viscospora Sieverd., Oehl & G.A. Silva, Mycotaxon 116: 108. 2011.
MycoBank MB 518439
Emended description: see Goto et al. [45]
Type species: Viscospora viscosa (T.H Nicolson) Sieverd., Oehl & G.A. Silva, Mycotaxon
116: 108. 2011.
MycoBank MB 518471
Basionym: Glomus viscosum T.H. Nicolson, Mycol. Res. 99: 1502. 1995.
MycoBank MB 413125
≡ Septoglomus viscosum (T.H. Nicolson) C. Walker, D. Redecker, D. Stiller & A. Schüssler, Mycorrhiza 23: 524. 2013
MycoBank MB 550089
Acaulosporales Sieverd., Silva G.A. & Oehl, ord. nov.
Mycobank MB 860126
Description: Spores formed laterally on or intrahyphally within hyphal stalk of a terminally or intercalary formed sporiferous saccule, in same distance to the sporiferous saccule termini, or rarely even within the inflated sporiferous saccule terminus; spores have three walls: an outer spore wall, a middle wall and an inner wall. One or a few layers of the outer spore wall are continuous with the wall of the stalk and the sporiferous saccule. The outer hyaline to subhyaline layers are often evanescent, the inner layers of the outer spore wall are permanent. When the connections of the hyphal stalk break off, the spore appears with one to two cicatrices which are closed by the permanent sublayers of the inner layers of the outer spore wall. The middle wall is thin and flexible. The inner wall consists of several thin layers of which the outer layer is ornamented having a characteristic' beaded' appearance. The second layer of the inner wall often stains deep purple in Melzer's reagent. The inner wall functions as germinal wall, a germination orb may be formed on the outer surface. Forming vesicular-arbuscular mycorrhizae whose fungal structures in the roots stain significantly blue with trypan blue.
Type family: Acaulosporaceae J.B. Morton & Benny, Mycotaxon 37: 479. 1990.
Mycobank MB 82037
Emended description; see description for the order here, directly above.
Type genus: Acaulospora Gerd. & Trappe
Other genus:
Kuklospora Oehl & Sieverd., J. Appl. Bot. Food Qual. 80: 74. 2006.
Mycobank MB 29042, current synonym of Acaulospora
Acaulospora Gerd. & Trappe, Mycologia Memoirs 5: 31. 1974.
Mycobank MB 20003
Emended description; see description for the order here directly above.
Type species: Acaulospora laevis Gerd. & Trappe, Mycologia Memoirs 5: 33. 1974.
MycoBank MB 308078
Diversisporales C. Walker & A. Schüssler, Mycological Research 108 (9): 981. 2004.
Mycobank MB 90593
Emended description: Spore formation diversisporoid, otosporoid, or tricisporoid, redeckeroid or (para-)corymbiglomoid. Diversisporoid spores formed singly, in clusters, or in disorganized sporocarps with up to very high spore numbers (up to several hundreds). In pigmented spores, SH conspicuously change color, becoming hyaline to white behind the septum, (immediately or at a very short distance from this septum); SH generally straight, cylindrical, in some species constricted or inflated. Spores with 13 wall layers; pore often closed with a septum that may arise from innermost wall lamina, an overlaying laminate layer, or from both; SH pore rarely open. Otosporoid and tricisporoid spores with two multiple-layered walls; otosporoid spores formed laterally on the persistent neck of a terminal or intercalary sporiferous saccule at some distance from the saccule terminus; spore pore generally closed by a septum at spore base. Tricisporoid spores formed within the evanescent neck of a tightly attached terminal or intercalary sporiferous saccule, closely attached to the saccule terminus which is often smaller in size than the mature spores attached, rarely equal in size; tricisporoid spores with two cicatrices formed by the outer wall pigmented structural layer. Para-)corymbiglomoid spores include several generic clades of Glomeromycetes fungi, such as Corymbiglomus, Paracorymbiglomus and Sieverdingia species forming mono-to-bi-walled glomoid-like spores, either corymbiforme or with a dense hyphal mantle on cylindrical SH, or Desertispora, forming glomoid-diversisporoid-like spores staining purple on the spore wall, which is not known for Diversispora species, or redeckeroid spores, which form mono-walled spores disorganized in large and compact sporocarps, containing hundreds to thousands of spores per sporocarp; spores, with 2 to rarely 3 wall layers; SH generally broad at spore base and with a conspicuous, thick and broad septum that arises from the inner lamina of the structural wall layer; structural layer generally continue over very short distances into SH; SWL1 fragile, usually inflating in a short distance to the spore base where SWL2 becomes invisible in the SH (= characteristically ‘redeckeroid’).
Type family: Diversisporaceae C. Walker & A. Schüssler
Other family: Corymbiglomeraceae G.A. Silva, Sieverd. & Oehl.
Diversisporaceae C. Walker & A. Schüssler, Mycological Research 108 (9): 981. 2004.
Mycobank MB 82125
Emended description: Spore formation diversisporoid, otosporoid, or tricisporoid. Diversisporoid spores formed singly, in clusters, or in disorganized sporocarps with up to very high spore numbers (up to several hundreds). In pigmented spores, SH conspicuously change color, becoming hyaline to white behind the septum, (immediately or at a very short distance from this septum); SH generally straight, cylindrical, in some species constricted or inflated. Spores with 1–3 wall layers; pore often closed with a septum that may arise from innermost wall lamina, an overlaying laminate layer, or from both; SH pore rarely open. Otosporoid and tricisporoid spores with two multiple-layered walls; otosporoid spores formed laterally on the persistent neck of a terminal or intercalary sporiferous saccule at some distance from the saccule terminus; spore pore generally closed by a septum at spore base. Tricisporoid spores formed within the evanescent neck of a tightly attached terminal or intercalary sporiferous saccule, closely attached to the saccule terminus which is often smaller in size than the mature spores attached, rarely equal in size; tricisporoid spores with two cicatrices formed by the outer wall pigmented structural layer.
Type genus: Diversispora C. Walker & A. Schüssler
Other genera:
Otospora Oehl, Palenzuela & N. Ferrol, Mycologia 100(2): 297. 2008.
Mycobank MB 506746, current synonym of Diversispora
Tricispora Oehl, Sieverd., G.A. Silva & Palenz., Mycotaxon 117: 310. 2012.
Mycobank MB 561642, current synonym of Diversispora
Diversispora C. Walker & A. Schüssler, Mycol. Res. 108 (9): 982. 2004.
Mycobank MB 28884
Emended description: see description for the family Diversisporaceae here, directly above.
Type species: Diversispora spurca (C.M. Pfeiff., C. Walker & Bloss) C. Walker & A. Schüssler, Mycological Research 108 (9): 982. 2004.
Mycobank MB 487795
Basionym: Glomus spurcum C.M. Pfeiff., C. Walker & Bloss, Mycotaxon 59: 374. 1996.
Mycobank MB 415789
Corymbiglomeraceae G.A. Silva, Sieverd. & Oehl fam. nov.
Mycobank MB 860135
Description: This new family includes several generic clades of Glomeromycetes fungi, such as Corymbiglomus, Paracorymbiglomus and Sieverdingia species forming mono-to-bi-walled glomoid-like spores, either corymbiforme or with a dense hyphal mantle on cylindrical SH, or Desertispora, forming glomoid-diversisporoid-like spores staining purple on the spore wall, which is not known for Diversispora species, or Redeckera species, which form mono-walled spores disorganized in large and compact sporocarps, containing hundreds to thousands of spores per sporocarp; spores, with 2 to rarely 3 wall layers; SH generally broad at spore base and with a conspicuous, thick and broad septum that arises from the inner lamina of the structural wall layer; structural layer generally continue over very short distances into SH; SWL1 fragile, usually inflating in a short distance to the spore base where SWL2 becomes invisible in the SH (= characteristically ‘redeckeroid’).
Type genus: Corymbiglomus (Błaszk. & Chwat) emend. Błaszk., Niezgoda & B.T.Goto
Other genera:
Desertispora Błaszk., Kozłowska, Ryszka, Al-Yahya’ei & Symanczik,
Paracorymbiglomus Błaszk., Niezgoda & B.T. Goto,
Redeckera C. Walker & A. Schüssler,
Sieverdingia Błaszk., Niezgoda & B.T. Goto.
Corymbiglomus Błaszk. & Chwat, Glomeromycota 1: 272. 2012.
Mycobank MB 564566
Description: see Błaszkowski [46],
Type species: Corymbiglomus corymbiforme (Blaszk.) Błaszk. & Chwat, The Glomeromycota 1: 274. 2012.
Mycobank MB 564567
Basionym: Glomus corymbiforme Blaszk., Mycologia 87(5): 732. 1995.
Mycobank MB 413122
Desertispora Błaszk., Kozłowska, Ryszka, Al-Yahya’ei & Symanczik, Mycological Progress 17 (4): 444. 2018.
Mycobank MB 823563
Description: see Symanczik et al. [47]
Type species: Desertispora omanana (Symanczik, Błaszk. & Al-Yahya’ei) Symanczik, Błaszk., Kozłowska & Al-Yahya’ei, Mycological Progress 17 (4): 444. 2018.
Mycobank MB 830800
Basionym: Diversispora omanana Symanczik, Błaszk. & Al-Yahya’ei, Mycologia 106 (2): 247. 2014.
Mycobank MB 830799
Paracorymbiglomus Błaszk., Niezgoda & B.T. Goto, Mycokeys 117: 178. 2025.
Mycobank MB 858391
Description: see Błaszkowski et al. [39]
Type species: Paracorymbiglomus globiferum (Koske & C. Walker) Błaszk., Niezgoda & B.T. Goto, MycoKeys 117: 178. 2025.
MycoBank No: 858393
Basionym: Glomus globiferum Koske & C. Walker, Mycotaxon 26. 133. 1986.
Synonym: Corymbiglomus globiferum (Koske & C. Walker) Błaszk. & Chwat, Acta Mycol. 48 (1): 99. 2013.
MycoBank No 622179
Redeckera C. Walker & A. Schüssler, The Glomeromycota—a species list with new families and new genera: 44. 2010.
MycoBank MB 542402
Description: see Oehl et al. [24]
Type species: Redeckera megalocarpa (D. Redecker) C. Walker & A. Schüssler, The Glomeromycota—a species list with new families and new genera: 44. 2010.
MycoBank MB 542403
Basionym: Glomus megalocarpum D. Redecker, Mycological Progress 6 (1): 38. 2007.
Mycobank MB 529715
Sieverdingia Błaszk., Niezgoda & B.T. Goto, Mycological Progress 18 (11): 1368. 2019.
Mycobank MB 832298
Description: see Błaszkowski et al. [48]
Type species: Sieverdingia tortuosa (N.C. Schenck & G.S. Sm.) Błaszk., Niezgoda & B.T. Goto, Mycological Progress 18 (11): 1369. 2019.
MycoBank MB 832299
Basionym: Glomus tortuosum N.C. Schenck & G.S. Sm., Mycologia 74 (1): 83. 1982.
MycoBank MB 110706
Entrophosporales Błaszk., Sánchez-García, B.T. Goto & Magurno, Frontiers in Microbiology 13 (no. 962856): 10. 2022.
MycoBank MB 846043
Type family: Entrophosporaceae Oehl & Sieverd.
Entrophosporaceae Oehl & Sieverd., J. Appl. Bot. Food Qual. 80: 73. 2006.
MycoBank MB 521877
Type genus: Entrophospora R.N. Ames & R.W. Schneid.
Other genera:
Albahypha Oehl, G.A. Silva, B.T. Goto & Sieverd.,
Alborhynchus Oehl, B.T. Goto, Corazon-Guivin, Sieverd. & G.A. Silva
Entrophospora R.N. Ames & R.W. Schneid., Mycotaxon 8 (2): 347. 1979.
MycoBank MB 20223
Type species: Entrophospora infrequens (I.R. Hall) R.N. Ames & R.W. Schneid., Mycotaxon 8 (2): 348. 1979.
Mycobank MB 313899
Basionym: Glomus infrequens I.R. Hall, Transactions of the British Mycological Society 68 (3): 345. 1977.
MB 314599
Albahypha Oehl, G.A. Silva, B.T. Goto & Sieverd., Mycotaxon 117: 308. 2011.
Mycobank MB 561639
Emended description: see Silva et al. [15]
Type species: Albahypha drummondii (Blaszk. & Renker) Sieverd., Oehl, B.T. Goto & G.A. Silva, Mycotaxon 117: 308. 2012.
Mycobank MB 561640
Basionym: Glomus drummondii Blaszk. & Renker, Mycological Research 110 (5): 559. 2006.
Mycobank MB 510049
≡ Entrophospora drummondii (Błaszk. & Renker) Błaszk., Niezgoda, B.T. Goto & Magurno, Frontiers in Microbiology 13 (no. 962856): 13. 2022.
Mycobank MB 836247
Alborhynchus Oehl, B.T. Goto, Corazon-Guivin, Sieverd. & G.A. Silva, J Fungi 11(2, no 97): 6. 2025.
Mycobank MB 856948
Description: see Silva et al. [15]
Type species: Alborhynchus walkeri (Blaszk. & Renker) Oehl, B.T. Goto, Corazon-Guivin, Sieverd. & G.A. Silva, J Fungi 11(2, no 97): 7. 2025.
Mycobank MB 856949
Basionym: Glomus walkeri Blaszk. & Renker, Mycological Research 110 (5): 563. 2006.
Mycobank MB 510050
≡ Entrophospora walkeri (Błaszk. & Renker) Błaszk., Niezgoda, B.T. Goto & Magurno, Frontiers in Microbiology 13 (no. 962856): 13. 2022.
Mycobank MB 836252
Gigasporales Sieverd., G.A. Silva, B.T. Goto & Oehl, Mycotaxon 116: 373. 2011.
MycoBank MB 519688
Description: see Oehl et al. [4]
Type family: Gigasporaceae Morton & Benny
Other families:
Dentiscutataceae F.A. Souza, Oehl & Sieverd.,
Intraornatosporaceae B.T. Goto & Oehl,
Racocetraceae Oehl, Sieverd. & F.A. Souza,
Scutellosporaceae Sieverd., F.A. Souza & Oehl.
Gigasporaceae Morton & Benny, Mycotaxon 37: 483. 1990.
MycoBank MB 82038
Description: see Morton & Benny [9]
Emended description: see Oehl et al. [49] for Gigasporaceae
Type genus: Gigaspora Gerd. & Trappe, Mycologia Memoirs 5: 25. 1974.
MycoBank MB 20239
Type species: Gigaspora gigantea (T.H. Nicolson & Gerd.) Gerd. & Trappe, Mycologia Memoirs 5: 29. 1974.
MycoBank MB 314484
Basionym: Endogone gigantea T.H. Nicolson & Gerd., Mycologia 60 (2): 321. 1968.
MycoBank MB 330364
Scutellosporaceae Sieverd., F.A. Souza & Oehl, Mycotaxon 106. 330. 2009 [2008].
Mycobank MB 511945
Emended description: Sporocarps are unknown. Spores generally singly formed on bulbous sporogenous cells that are formed terminally on subtending hyphae (=sporogenous hyphae; SH) that arise from mycelia hyphae in soil. Spores have 3 walls, an outer, a middle and an inner, germinal wall. Outer wall with 3 or several layers, middle wall with 1–2 layers, and inner wall with 2-4. A single germination shield, rarely two shields, are formed on the outer surface or between the outer and the subsequent layer of the inner wall. Germination shields are transparent, or hyaline to subhyaline, seldom light yellow, generally bi-lobed, sometimes mono-lobed; often violin-shaped or oval to ovoid, rarely cardioid, circular or coiled; only a few folds cover the shield surface where in general one germ tube initiation (gti) is formed per lobe. Generally from one gti, seldom from both gti, a germ tube arises and penetrates the outer spore walls. SH form one to several septa in some distance to the sporogenous cells. Forming typical arbuscular mycorrhizae without vesicle formation in roots; knobby extraradical auxiliary cells branching from the hyphal mycelium without spines on the surface.
Type genus: Scutellospora C. Walker & F.E. Sanders
Other genera:
Bulbospora Oehl & G.A. Silva,
Orbispora Oehl, G.A. Silva & D.K. Silva.
Scutellospora C. Walker & F.E. Sanders, Mycotaxon 27: 179. 1986.
Mycobank MB 25074
Emended description: Sporocarps unknown. Spores formed on sporogenous cells that form terminally on a hypha which arises from mycelia hyphae in soil. Outer spore wall generally is three-layered and continuous with the wall of the sporogenous cell. Outer layer of the outer spore wall generally rigid, second layer laminate and third layer thin, often membranous, tightly adherent to the laminate layer and thus, often difficult to observe. Pore between the spore and sporogenous cell is narrow and usually closed by a plug formed by spore wall material. Two hyaline walls (‘MW’ and ‘IW’) form de novo during spore formation and have 1–2 and 2–4 layers, respectively. The IW is two to three-layered forming a germination shield on its outer surface or between the outer and the subsequent layer of IW. Germination shield is transparent, or hyaline to subhyaline, seldom light yellow, bi-lobed; often violin-shaped to oval to ovoid to more rarely cardioids or coiled and then, either circular or apparently broad ellipsoid to irregular; only a few folds cover the shield surface where 1–2 rounded germ tube initiations (‘gti’, about 2–4 μm in diam) are visible from where the germ tubes arise which penetrate the outer spore wall layers. Sometimes one to a few so-called false lobes are visible, which never bear a gti, and usually develop artificially by pressure on the spore, e.g., through pressure on the cover slides under the microscope; mycelia hyphae form one to several septa in some distance to the sporogenous cells. Auxiliary cells in the hyphal mycelium, as far as known, are knobby without spines on the surface. Forming typical arbuscular mycorrhizae without vesicle formation in roots; knobby extraradical auxiliary cells branching from the hyphal mycelium without spines on the surface.
Type species: Scutellospora calospora (T.H. Nicolson & Gerd.) C. Walker & F.E. Sanders, Mycotaxon 27: 180. 1986.
Mycobank MB 128413
Basionym: Endogone calospora T.H. Nicolson & Gerd., Mycologia 60 (2): 322. 1968.
Mycobank MB MB 330363
≡ Gigaspora calospora (T.H. Nicolson & Gerd.) Gerd. & Trappe, Mycologia Memoirs 5: 28. 1974.
Mycobank MB 314482
Bulbospora Oehl & G.A. Silva, Sydowia 66. 315. 2014.
Mycobank MB 809231
Description: see Marinho et al. [50]
Type species: Bulbospora minima Oehl, Marinho, B. T. Goto & G. A. Silva, Sydowia 66. 315. 2014.
MycoBank MB 809232
Orbispora Oehl, G.A. Silva & D.K. Silva, Mycotaxon 116. 163. 2011.
MycoBank MB 519533
Emended description: Sporocarps unknown. Spores 100-300 µm, formed on sporogenous cells that form terminally on hyphae arising from mycelia in soil. Outer spore wall (OW) generally (2–)3-layered and continuous with the wall of the sporogenous cell. Two hyaline walls (‘MW’ and ‘IW’) form de novo during spore formation and have 1–2 and 2–3 layers, respectively. A germination orb is formed on the outer IW surface or between the outer and the subsequent layer of IW. Germination orb is transparent, or hyaline to subhyaline, seldom light yellow, mono-lobed; coiled and then, either circular or apparently broad ellipsoid to rarely irregular; with one rounded germ tube initiation in the outer periphery of the lobe. One (rarely two) germ tube arises from this gti to penetrate the outer spore wall layers.
Type species: Orbispora pernambucana (Oehl, D.K. Silva, N. Freitas & L.C. Maia) Oehl, G.A. Silva & D.K. Silva, Mycotaxon 116. 166. 2011.
MycoBank MB 519535
Basionym: Scutellospora pernambucana Oehl, D.K. Silva, N. Freitas & L.C. Maia, Mycotaxon 106: 363. 2009 [2008].
MycoBank MB 512130
Racocetraceae Oehl, Sieverd. & F.A. Souza, Mycotaxon 106: 333. 2009 [2008].
MB 511946
Description: see Oehl et al. [49]
Type genus: Racocetra Oehl, F.A. Souza & Sieverd.
Other genus: Cetraspora Oehl, F.A. Souza & Sieverd.
Racocetra Oehl, F.A. Souza & Sieverd., Mycotaxon 106: 334. 2009 [2008].
MB 511947
Description: see Oehl et al. [49]
Type species: Racocetra coralloidea (Trappe, Gerd. & I. Ho) Oehl, F.A. Souza & Sieverd., Mycotaxon 106: 336. 2009 [2008].
MB 511948
Basionym: Gigaspora coralloidea Trappe, Gerd. & I. Ho, Mycologia Memoir No. 5:
30. 1974.
≡ Scutellospora coralloidea (Trappe, Gerd. & I. Ho) C. Walker & F.E. Sanders, Mycotaxon 27: 181. 1986.
Cetraspora Oehl, F.A. Souza & Sieverd., Mycotaxon 106: 337. 2009 [2008].
MB 511957
Description: see Oehl et al. [49]
Type species: Cetraspora gilmorei (Trappe & Gerd.) Oehl, F.A. Souza & Sieverd., Mycotaxon 106: 338. 2009 [2008].
MycoBank MB 511958
Basionym: Gigaspora gilmorei Trappe & Gerd., Mycologia Memoir No. 5: 27. 1974.
≡ Scutellospora gilmorei (Trappe & Gerd.) C. Walker & F.E. Sanders, Mycotaxon 27: 181. 1986.
Dentiscutataceae F.A. Souza, Oehl & Sieverd., Mycotaxon 106. 340. 2009 [2008].
MycoBank MB 511962
Description: see Oehl et al. [49]
Type genus: Dentiscutata Sieverd., F.A. Souza & Oehl
Other genera:
Fuscutata Oehl, F.A. Souza & Sieverd.,
Quatunica F.A. Souza, Sieverd. & Oehl.
Dentiscutata Sieverd., F.A. Souza & Oehl, Mycotaxon 106. 340. 2009 [2008].
MycoBank MB 511968
Description: see Oehl et al. [49]
Type species: Dentiscutata nigra (J.F. Redhead) Sieverd., F.A. Souza & Oehl, Mycotaxon 106. 342. 2009 [2008].
MycoBank MB 511969
Basionym: Gigaspora nigra J.F. Redhead, Mycologia 71: 187. 1979.
MycoBank MB 314489
≡ Scutellospora nigra (J.F. Redhead) C. Walker & F.E. Sanders, Mycotaxon 27: 181. 1986.
MycoBank MB 128421
Fuscutata Oehl, F.A. Souza & Sieverd., Mycotaxon 106. 342. 2009 [2008].
MycoBank MB 511963
Description: see Oehl et al. [49]
Type species: Fuscutata heterogama Oehl, F.A. Souza, L.C. Maia & Sieverd., Mycotaxon 106. 342. 2009 [2008].
MycoBank MB 511964
Quatunica F.A. Souza, Sieverd. & Oehl, Mycotaxon 106. 347. 2009 [2008].
MycoBank MB 511976
Description: see Oehl et al. [49]
Type species: Quatunica erythropus (Koske & C. Walker) F.A. Souza, Sieverd. & Oehl, Mycotaxon 106. 348. 2009 [2008].
MycoBank MB 511977
Basionym: Gigaspora erythropus Koske & C. Walker, Mycologia 76: 250. 1984.
MycoBank MB 318956
≡ Scutellospora erythropus (Koske & C. Walker) C. Walker & F.E. Sanders, Mycotaxon 27: 181. 1986.
MycoBank MB318960
Intraornatosporaceae B.T. Goto & Oehl, Mycotaxon 119: 121. 2012.
MycoBank MB 563599
Description: see Goto et al. (2012)
Type genus: Intraornatospora B.T. Goto, Oehl & G.A. Silva, Mycotaxon 119: 122. 2012.
Other genus: Paradentiscutata B.T. Goto, Oehl & G.A. Silva
Intraornatospora B.T. Goto, Oehl & G.A. Silva, Mycotaxon 119: 122. 2012.
MycoBank MB 563600
Description: see Goto et al. [51]
Type species: Intraornatospora intraornata (B.T. Goto & Oehl) B.T. Goto, Oehl & G.A. Silva, Mycotaxon 119: 122. 2012.
MycoBank MB 563601
Basionym: Racocetra intraornata B.T. Goto & Oehl, Mycotaxon 109: 485. 2009.
MycoBank MB 513428
Paradentiscutata B.T. Goto, Oehl & G.A. Silva, Mycotaxon 119: 122. 2012.
MycoBank MB 563602
Description: see Goto et al. [51]
Type species: Paradentiscutata bahiana Oehl, Magna, B.T. Goto & G.A. Silva, Mycotaxon 119: 123. 2012.
MycoBank MB 563603
Pacisporales Oehl, Sieverd., G.A. Silva, ord. nov.
MycoBank MB 860128
Description: Spores formed singly in soils or rarely in roots, terminally on SH, with two spore walls (OW & IW), while only the outer wall is continuous with the mycelia and SH, but the inner wall forms de novo during spore differentiation. Spore pore usually is closed by a septum at the spore base arising from the inner wall layers of the outer spore wall. IW functions as germinal wall; germ tubes emerge from the germinal wall directly or from the center of a multiple-lobed germination structure consisting of several lobes positioned around this center. Forming typical vesicular-arbuscular mycorrhizal fungi, whose fungal structures in the roots stain significantly blue with trypan blue, and additionally extraradical auxiliary cells in the rhizosphere of mycorrhizal roots, rarely intraradically, as also known for Gigasporales.
Type family: Pacisporaceae C. Walker, Błaszk., A. Schüssler & Schwarzott, Mycol. Res. 108 (9): 981. 2004.
MycoBank MB 82126
Emended description: see above for Pacisporales
Type genus: Pacispora Sieverd. & Oehl
Pacispora Sieverd. & Oehl, J. Appl. Bot. 78: 74. 2004.
MycoBank MB 28842
Emended description: see above for Pacisporales
Type species: Pacispora scintillans (S.L. Rose & Trappe) Sieverd. & Oehl ex C. Walker, Vestberg & A. Schüssler, Mycological Research 111 (3): 254. 2007.
MycoBank MB 510506
Basionym: Glomus scintillans S.L. Rose & Trappe, Mycotaxon 10 (2): 417. 1980.
MycoBank MB 113776
Sacculosporales Silva G.A., Sieverd. & Oehl, ord. nov
Mycobank MB 860127
Description: Sporocarps unknown. Spores formed within the hyphal neck of closely adherent terminal or intercalary formed sporiferous saccules. Spores have two walls: outer and inner. At least two layers (including the outer wall structural layer) are continuous with the sporiferous saccule wall. After the hyphal neck connections break off, spores show two, often opposite, cicatrices that are closed by the permanent sublayers of the outer wall structural layer. Inner wall forms de novo. Inner wall consists of several layers, none of which has a granular, 'beaded' appearance, and does not stain in Melzer’s reagent.
Type family: Sacculosporaceae Oehl, Sieverd., G.A. Silva, B.T. Goto, I.C. Sánchez & Palenzuela, Mycotaxon 117: 310. 2012.
MycoBank MB 561645
Emended description: see above for the order Sacculosporales and Willis et al. [52]
Type genus: Sacculospora Oehl, Sieverd., G.A. Silva, B.T. Goto, I.C. Sánchez & Palenzuela
Sacculospora Oehl, Sieverd., G.A. Silva, B.T. Goto, I.C. Sánchez & Palenzuela, Mycotaxon 117: 311. 2011.
MycoBank MB 561646
Description: see above for the order Sacculosporales and Willis et al. [52]
Type species: Sacculospora baltica (Blaszk., Madej & Tadych) Oehl, Palenzuela, I.C. Sánchez, B.T. Goto, G.A. Silva & Sieverd., Mycotaxon 117: 311. 2012.
MycoBank MB 561647
Basionym: Entrophospora baltica Blaszk., Madej & Tadych, Mycotaxon 68: 167. 1998.
Mycobank MB 444868
4. Discussion
Our results show seven major clades at order level in Glomeromycetes [sensu 4] with full support in all analyses (Figure 1). Tedersoo et al. [53] and Silva et al. [14] have obtained the same seven clades, fully supported for this class. Although the topology of our tree, considering the relationships among the orders, is relatively similar to that obtained by Silva et al. [14], it is different from the tree obtained by Tedersoo et al. [53]. In Tedersoo et al. [53], Sacculosporales is near to Pacisporales, but with low IQ-TREE support (88.1/78%), while in our tree Sacculosporales is a sister group to Diversisporales (0.95 for BI and 90% for ML), and Pacisporales is near to Gigasporales, but with no support. In our tree, Acaulosporales is a sister group to a clade with Sacculosporales and Diversisporales together (1.00 for BI and 71% for ML), while the tree generated by Tedersoo et al. [53] showed Acaulosporales as a sister group of Diversisporales (44.4/99%). Considering the topology variation, in order level, found between different trees, and considering the low or no support to place the order Pacisporales with other clades, we are confident that it is not possible to put Pacisporales together with Gigasporales or Sacculosporales, nor Diversisporales together with Sacculosporales or Acaulosporales in a same taxon, respectively. Thus, it was necessary to describe three new orders.
Silva et al. [14] described four new families in Glomerales. These authors suggested that future studies could describe new families in this order, citing specifically Halonatospora, Oehlia, and Rhizoglomus as three genera types for future new families. The nine family clades in Glomerales, here generated, are strongly supported (>0.99 for BI and >90% for ML), except for ML analysis of Kamienskiaceae and Sclerocystaceae (74 and 78%, respectively), justifying the erection of four new families. In our tree, Halonatosporaceae is close to Rhizoglomeraceae and Sclerocystaceae, and Oehliaceae is near to the clade composed by these three families. The studies from Tedersoo et al. [53] show Halonatosporaceae near to Kamienskiaceae and Oehliaceae near to Sclerocystaceae and Rhizoglomeraceae, but with no good support to maintain these groups in any of these families. Our tree and that from Tedersoo et al. [53] give support to erect Funneliformaceae in a new family. The data by Tedersoo et al. [53] had no support to erect Rhizoglomeraceae, however our phylogenetic tree well separates this family. Considering our analysis and the morphology of the different groups, we are convinced that Rhizoglomeraceae represent a strong new family in Glomerales.
Entrophosporales present a single family with three genera [15], however there was no ML support in our tree to place Albahypha drummondi and A. furrazolae together, and the BI support, to group these species, was low (0.75). Acaulosporales, Pacisporales, and Sacculosporales possess monogeneric families. In relation to Gigasporales, the five families and eleven genera represented in our tree are well supported, and our findings are similar to that obtained by Silva et al. [14], except for the ML analysis for Dentiscutataceae and Fuscutata (0.96 for BI and 65% for ML).
In Diversisporales, two clades were found with strong support. To our opinion these clades represent the current families in this order (Figure 1). These clades are present in the tree obtained by Tedersoo et al. [53], and the groups representing Diversisporaceae and Corymbiglomeraceae have a strong IQ-TREE support (99.9/100 and 99.8/99, respectively). Diversisporaceae is monogeneric with Diversispora, nonetheless Corymbiglomeraceae is composed by five well supported genera. In Tedersoo et al. [53], the genus Redeckera is not well defined. This problem was partially solved by Blaszkowski et al. [39] by the description of Paracorymbiglomus, however future work will be necessary to clarify the phylogenetic relationship of this group.
Morphologically, no new data are created in this study. Several major clades have been well known as clearly distinct, such as the Acaulosporaceae, Sacculosporaceae and Pacisporaceae e.g. [9,25,37,52). The existing data were summarized here (see Table S1). Based on the advances in the phylogenetic analyses, these morphological clades are here separating on the order level.
The newly revised family Diversisporaceae comprise now solely the diversisporoid species sensu stricto, and the recently recombined species, formerly attributed to Otospora and Tricispora [5,25]. In the new family Corymbiglomeraceae, besides the type genus Corymbiglomus, also Paracorymbiglomus, Desertispora, Redeckera and Sieverdingia are included. However, the phylogenetic and morphological data bases of these taxa are still scarce, so that a clearer separation of these AMF genera hitherto is not yet feasible. Concerning morphology, Redeckera is the next genus, which should be separated on the family level, as soon as more species are analyzed on the molecular-biological level. This clade of fungi seems to be as clearly separable morphologically, as for instance Acaulosporaceae, Gigasporaceae and Rhizoglomeraceae e.g. [4,9,54].
Funneliformis, Halonatospora, Oehlia and Rhizoglomus are morphologically well-defined taxa, which were now presenting also taxa on the family level. The earlier described families are also separating readily [14], which recently was also used as a useful base to separate ecological groups within Glomerales [55]. In Glomerales, one well-known family is still analyzed on a low level. This is Sclerocystaceae, in which only for a few species molecular analyses have been available. In this family, we expect a clearer separation on the genus level, when more sequences are available, for instance for early described species like S. clavispora, S. rubiformis and S. taiwanensis e.g. [56–59). Interestingly, species of Sclerocystaceae were in earlier times recombined with the genus Glomus [60].
In conclusion, the present revision of the Glomeromycetes is, to our opinion, a strong step forward in the taxonomy and phylogeny of the Glomeromycetes. There is now a strong correlation between phylogenetic and morphological clades, much more than ever before. It will be interesting, how this fungal class and the whole phylum will be further explored on the taxonomic, but also beyond on the ecological level. The most ancestral clades have not anymore been included in Glomeromycetes since a while, also based on concomitant phylogenetic and morphological data. These ancestral clades of AM fungi in Archaeosporomycetes and Paraglomeromycetes currently remain the larger mystery, since hyphae and spores are often very short-lived and also the intraradical structures are not easily to detect in nature or even after staining procedures. However, when thinking about small-spored, hyaline and short-lived spores of Viscospora, Kamienskia or Dominikia [61,62], or even easily visible sporocarps of Sclerocystaceae [63,64,65] or big spores in Dentiscutataceae [66,67] and Gigasporaceae e.g. [68], it becomes also clear again that much research work has still to be done also in Glomeromycetes.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Spreadsheet S1: GenBank accession numbers for the sequences used in this study; Table S1: Morphological key characteristics of the seven orders in Glomeromycetes; Table S2: Current orders, families and genera in the Glomeromycetes.
Author Contributions
All authors contributed to the study’s conception and design. Phylogenetic analyses were performed by G.A.d.S. and morphological analyses by E.S. and F.O. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data (sequences) presented in this article are in GenBank (https://www.ncbi.nlm.nih.gov).
Acknowledgments
Daniele Magna Azevedo de Assis and Thays Gabrielle Lins de Oliveira thanks the Fundação de Amparo à Ciência e Tecnologia do Estado de Pernambuco (FACEPE) for providing fellowship. Gladstone A. Silva has a fellowship from the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) (Proc. 312606/2022-2, approval date: 16 February 2023).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Cavalier-Smith, T. A revised six-kingdom system of life. Biol. Rev. Camb. Philos. Soc. 1998, 73, 203–266.
- Stürmer, S.L. A history of the taxonomy and systematics of arbuscular mycorrhizal fungi belonging to the phylum Glomeromycota. Mycorrhiza 2012, 22(4), 247–258. [CrossRef]
- Morton, J.B.; Redecker, D. Two new families of Glomales, Archaeosporaceae and Paraglomaceae, with two new genera Archaeospora and Paraglomus, based on concordant molecular and morphological characters. Mycologia 2001, 93, 181–195.
- Oehl, F.; Silva, G.A.; Goto, B.T.; Maia, L.C.; Sieverding, E. Glomeromycota: two new classes and a new order. Mycotaxon 2011a, 116(1), 365–379. [CrossRef]
- Hyde, K.D.; Noorabadi, M.T.; Thiyagaraja, V.; He, M.Q.; Johnston, P.R.; Wijesinghe, S.N.; Armand, A.; Biketova, A.Y.; Chethana K.W.T.; Erdoğdu, M.; et al. The 2024 Outline of Fungi and fungus-like taxa. Mycosphere 2024, 15(1), 5146–6239. [CrossRef]
- Spatafora, J. W.; Chang, Y.; Benny, G.L.; Lazarus, K.; Smith, M.E.; Berbee, M.L.; Bonito, G.; Corradi, N.; Grigoriev, I.; Gryganskyi, A.; et al. A phylum-level phylogenetic classification of zygomycete fungi based on genome-scale data. Mycologia 2016, 108(5), 1028–1046. [CrossRef]
- Schüßler, A.; Schwarzott, D.; Walker, C. A new fungal phylum, the Glomeromycota: Phylogeny and evolution. Mycol. Res. 2001, 105, 1413–1421. [CrossRef]
- Tedersoo, L.; Sánchez-Ramírez, S.; Kõljalg, U.; Bahram, M.; Döring, M.; Schigel, D.; May, T.; Ryberg, M.; Abarenkov, K. High-level classification of the Fungi and a tool for evolutionary ecological analyses. Fungal Divers. 2018, 90, 135–159. [CrossRef]
- Morton, J.B.; Benny, G.L. Revised classification of arbuscular mycorrhizal fungi (Zygomycetes): A new order, Glomales, two new suborders, Glomineae and Gigasporinae, and two families, Acaulosporaceae and Gigasporaceae, with an emendation of Glomaceae. Mycotaxon 1990, 37, 471–491.
- Schwarzott, D.; Walker, C.; Schüßler, A. Glomus, the largest genus of the arbuscular mycorrhizal fungi (Glomales), is nonmonophyletic. Mol. Phylogenet Evol. 2001 21(2), 190-197. [CrossRef]
- Błaszkowski, J.; Sánchez-García, M.; Niezgoda, P.; Zubek, S.; Fernández, F.; Vila, A.; Al-Yahya’ei, M.N.; Symanczik, S.; Milczarski, P.; Malinowski, R.; et al. A new order, Entrophosporales, and three new Entrophospora species in Glomeromycota. Front. Microbiol. 2022, 13, 962856. [CrossRef]
- Walker, C.; Schüßler, A. Nomenclatural clarifications and new taxa in the Glomeromycota. Mycol. Res. 2004, 108(9), 981–982. [CrossRef]
- Pirozynski, K.A.; Dalpé, Y. Geological history of the Glomaceae with particular reference to mycorrhizal symbiosis. Symbiosis 1989, 7: 1–36.
- Silva, G.A.; Assis, D.M.A.; Sieverding, E.; Oehl, F. Four new families of arbuscular mycorrhizal fungi within the order Glomerales. Taxonomy 2024, 4(4), 761-779. [CrossRef]
- Silva, G.A.; Sieverding, E.; Assis, D.M.A.; Goto, B.T.; Corazon-Guivin, M.A.; Oehl, F. Revision of Entrophosporales, with three genera and an identification key for all species currently attributed to this order. Journal of Fungi 2025, 11(2), 97. [CrossRef]
- Błaszkowski, J.; Niezgoda, P.; Meller, E.; Milczarski, P.; Zubek, S.; Malicka, M.; Uszok, S.; Casieri, L.; Goto, B.T.; Magurno, F. New taxa in Glomeromycota: Polonosporaceae fam. nov., Polonospora gen. nov., and P. polonica comb. nov. Mycol. Prog. 2021a, 20, 941–951. [CrossRef]
- Schüßler, A. Molecular phylogeny, taxonomy, and evolution of Geosiphon pyriformis and arbuscular mycorrhizal fungi. Plant soil 2002 244(1), 75–83. [CrossRef]
- Błaszkowski, J.; Kozłowska, A.; Crossay, T.; Symanczik, S.; Al-Yahya’ei, M.N. A new family, Pervetustaceae with a new genus, Pervetustus, and P. simplex sp. nov. (Paraglomerales), and a new genus, Innospora with I. majewskii comb. nov. (Paraglomeraceae) in the Glomeromycotina. Nova Hedw. 2017, 105, 397–410. [CrossRef]
- Nicolson, T.H.; Gerdemann, J.W. Mycorrhizal Endogone species. Mycologia 1968, 60(2), 313–325. [CrossRef]
- Gerdemann, J.W.; Trappe, J.M. The Endogonaceae of the Pacific Northwest. Mycol. Mem. 1974, 5, 1–76.
- Velazquez, M.S.; Cabello, M.N.; Irrazabal, G.B.; Godeas, A.M. Acaulosporaceae from El Palmar National Park, Entre Ríos, Argentina. Mycotaxon 2008, 103, 171–187.
- Błaszkowski, J.; Madej, T.; Tadych, M. Entrophospora baltica sp. nov. and Glomus fuegianum, two species in the Glomales from Poland. Mycotaxon 1998, 68, 165–184.
- Oehl, F.; Sieverding, E. Pacispora, a new vesicular arbuscular mycorrhizal fungal genus in the Glomeromycetes. J. Appl. Bot. Food Qual. 2004, 78, 72–82.
- Oehl, F.; Silva, G.A.; Goto, B.T.; Sieverding, E. Glomeromycota: Three new genera and glomoid species reorganized. Mycotaxon 2011b, 116, 75–120. [CrossRef]
- Oehl, F.; Silva, G.A.; Sánchez-Castro, I.; Goto, B.T.; Maia, L.C.; Vieira, H.E.E.; Barea, J.-M.; Sieverding, E.; Palenzuela, J. Revision of Glomeromycetes with entrophosporoid and glomoid spore formation with three new genera. Mycotaxon 2011c, 117, 297–316. [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: efficient Bayesian phylogenetic inference and model choice across a large model space. Syst Biol. 2012, 61, 539–42. [CrossRef]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; pp. 1–8. [CrossRef]
- Kozlov, A.M.; Darriba, D.; Flouri, T.; Morel, B.; Stamatakis, A. RAxML-NG: a fast, scalable and user-friendly tool for maximum likelihood phylogenetic inference. Bioinformatics 2019, 35: 4453–4455. [CrossRef]
- Edler, D.; Klein, J.; Antonelli, A.; Silvestro, D. RaxmlGUI 2.0: A graphical interface and toolkit for phylogenetic analyses using RAxML. Methods Ecol Evol. 2021, 12, 373–377. [CrossRef]
- Darriba, D.; Posada, D.; Kozlov, A.M.; Stamatakis, A.; Morel, B.; Flouri, T. ModelTest-NG: A New and Scalable Tool for the Selection of DNA and Protein Evolutionary Models. Molecular Biology and Evolution 2020, 37, 291–294. [CrossRef]
- Hall, I.R.; Abbott, L.K. Photographic Slide Collection Illustrating Features of the Endogonaceae, 3rd ed.; Invery Agricultural Research Centre and Soil Science Department, University ofWestern Australia: Crawley, Australia, 1979; pp. 1–27.
- Brundrett, M.; Melville, L.; Peterson, L. Practical Methods in Mycorrhizal Research; Mycologue Publications, University of Guelph: Guelph, ON, Canada, 1994.
- Spain, J.L. Emendation of Archaeospora and of its type species, Archaeospora trappei. Mycotaxon 2003, 87, 109–112.
- Walker, C. Taxonomic concepts in the Endogonaceae: Spore wall characteristics in species descriptions. Mycotaxon 1983, 18, 443–455.
- Stürmer, S.L.; Morton, J.B. Developmental patterns defining morphological characters in spores of four species in Glomus. Mycologia 1997, 89, 72–81. [CrossRef]
- Błaszkowski, J. Glomeromycota; W. Szafer Institute of Botany, Polish Academy of Sciences: Kraków, Poland, 2012; p. 303, ISBN 978-83-89648-82-2.
- Błaszkowski, J.; Zubek, S.; Milczarski, P.; Malinowski, R.; Niezgoda, P.; Goto, B.T. New taxa and a combination in Glomerales (Glomeromycota, Glomeromycetes). MycoKeys 2025a, 112, 253. [CrossRef]
- Błaszkowski, J.; Goto, B.T.; Zubek, S.; Milczarski, P.; Malinowski, R.; Niezgoda, P. Błaszkowski, T. Paracorymbiglomus gen. nov., Diversispora conica sp. nov., and new combinations in Diversisporaceae (Glomeromycota). MycoKeys 2025b, 117, 171–190. [CrossRef]
- Błaszkowski, J.; Yamato, M.; Niezgoda, P.; Zubek, S.; Milczarski, P.; Malinowski, R.; Goto, B.T.; Uszok, S.; Casieri, L.; Magurno, F. A new genus, Complexispora, with two new species, C. multistratosa and C. mediterranea, and Epigeocarpum japonicum sp. nov. Mycol. Prog. 2023, 22, 1–15. [CrossRef]
- Jobim, K.; Błaszkowski, J.; Niezgoda, P.; Kozłowska, A.; Zubek, S.; Mleczko, P.; Chachuła, P.; Ishikawa, N.K.; Goto, B.T. New sporocarpic taxa in the phylum Glomeromycota: Sclerocarpum amazonicum gen. et sp. nov. in the family Glomeraceae (Glomerales) and Diversispora sporocarpia sp. nov. in the Diversisporaceae (Diversisporales). Mycol. Prog. 2019, 18, 369-384. [CrossRef]
- Błaszkowski, J.; Niezgoda, P.; Goto, B.T.; Kozłowska, A. Halonatospora gen. nov. with H. pansihalos comb. nov. and Glomus bareae sp. nov. (Glomeromycota; Glomeraceae). Botany 2018, 96(11), 737–748. [CrossRef]
- Błaszkowski, J.; Jobim, K.; Niezgoda, P.; Meller, E.; Malinowski, R.; Milczarski, P.; Zubek, S.; Magurno, F.; Casieri, L.; Bierza, W.; Błaszkowski, T.; Crossay, T.; Goto, B.T. New glomeromycotan taxa, Dominikia glomerocarpica sp. nov. and Epigeocarpum crypticum gen. nov. et sp. nov. from Brazil, and Silvaspora gen. nov. from New Caledonia. Front. Microbiol. 2021b, 12, 655910. [CrossRef]
- Błaszkowski, J.; Kozłowska, A.; Niezgoda, P.; Goto, B. T.; Dalpé, Y. A new genus, Oehlia with Oehlia diaphana comb. nov. and an emended description of Rhizoglomus vesiculiferum comb. nov. in the Glomeromycotina. Nova Hedwigia, 2018, 107(3-4), 501–518.
- Goto, B.T.; Silva, G.A.; Błaszkowski, J.; Niezgoda, P.; Corazon-Guivin, M.A.; Oliveira, N.V.L.; Sieverding, E.; Oehl, F. Phylogeny and morphology of two new arbuscular mycorrhizal fungal genera, with an identification key for all genera within the order Glomerales. Mycol. Prog. 2025, in review, re-submitted after minor revision.
- Blaszkowski, J. Glomus corymbiforme, a new species in Glomales from poland. Mycologia 1995, 87, 732–737. [CrossRef]
- Symanczik, S.; Al-Yahya’ei, M.N.; Kozłowska, A.; Ryszka, P.; Błaszkowski, J. A new genus, Desertispora, and a new species, Diversispora sabulosa, in the family Diversisporaceae (order Diversisporales, subphylum Glomeromycotina). Mycol. Prog. 2018, 17, 437–449. [CrossRef]
- Błaszkowski, J.; Niezgoda, P.; de Paiva, J.N.; da Silva, K.J.G.; Theodoro, R.C.; Jobim, K.; Orfanoudakis, M.; Goto, B.T. Sieverdingia gen. nov., S. tortuosa comb. nov., and Diversispora peloponnesiaca sp. nov. in the Diversisporaceae (Glomeromycota). Mycol. Prog. 2019, 18(11), 1363–1382. [CrossRef]
- Oehl, F.; de Souza, F.A.; Sieverding, E. Revision of Scutellospora and description of five new genera and three new families in the arbuscular mycorrhiza-forming Glomeromycetes. Mycotaxon 2008, 106(1), 311-360.
- Marinho, F.; Silva, G.A.; Ferreira, A.C.A.; Veras, J.S.N.; Sousa, N.M.F.; Goto, B.T.; Maia, L.C.; Oehl, F. Bulbospora minima, a new genus and a new species in the Glomeromycetes from semi-arid Northeast Brazil. Sydowia 2014, 66(2), 313–323. [CrossRef]
- Goto, B.T.; Silva, G.A.; Assis, D.M.A.; Silva, D.K.A.; Souza, R.G.; Ferreira, A.C.A.; Jobim, K.; Mello, C.M.A.; Vieira, H.E.E.; Maia, L.C.; Oehl, F. Intraornatosporaceae (Gigasporales), a new family with two new genera and two new species. Mycotaxon 2012, 119(1), 117–132. [CrossRef]
- Willis, A.; Błaszkowski, J.; Prabhu, T.; Chwat, G.; Góralska, A.; Sashidhar, B.; Harris, P.; D’Souza, J.; Vaingankar, J.; Adholeya, A. Sacculospora felinovii, a novel arbuscular mycorrhizal fungal species (Glomeromycota) from dunes on the west coast of India. Mycol. Prog. 2016, 15(7), 791-798. [CrossRef]
- Tedersoo, L.; Magurno, F.; Alkahtani, S.; Mikryukov, V. Phylogenetic classification of arbuscular mycorrhizal fungi: New species and higher-ranking taxa in Glomeromycota and Mucoromycota (class Endogonomycetes). MycoKeys 2024, 107, 249–271. [CrossRef]
- Sieverding, E.; Silva, G.A.; Berndt, R.; Oehl, F. Rhizoglomus, a new genus of the Glomeraceae. Mycotaxon 2015, 129(2), 373–386. [CrossRef]
- Buil, P.A.; Jansa, J.; Rozmoš, M.; Kotianová, M.; Bukovská, P.; Grilli, G.; Marro, N.; Janoušková, M. Soil cropping selects for nutrient efficient but more costly indigenous mycorrhizal fungal communities. Biol. Fertil. Soils 2025 61(5), 841–859. [CrossRef]
- Schenck, N.C.; Perez, Y. Manual for the identification of VA mycorrhizal fungi; Gainesville. USA: INVAM, University of Florida. 1990.
- Almeida, R.T.; Schenck, N.C. A revision of the genus Sclerocystis (Glomaceae, Glomales). Mycologia 1990, 82(6), 703–714. [CrossRef]
- Wu, C.-G. Glomales of Taiwan: III. A comparative spore ontogeny in Sclerocystis (Glomaceae, Glomales). Mycotaxon 1993, 47: 25-39.
- Wu, C.-G. Glomales of Taiwan: IV. A monograph of Sclerocystis (Glomaceae). Mycotaxon 1993, 49, 327–349.
- Redecker, D.; Morton, J.B.; Bruns, T.D. Molecular phylogeny of the arbuscular mycorrhizal fungi Glomus sinuosum and Sclerocystis coremioides. Mycologia 2000, 92(2), 282–285. [CrossRef]
- Walker, C.; Giovannetti, M.; Avio, L.; Citernesi, A.S.; Nicolson, T.H. A new fungal species forming arbuscular mycorrhizas: Glomus viscosum. Mycol. Res. 1995, 99(12), 1500–1506. [CrossRef]
- Błaszkowski, J.; Chwat, G.; Góralska, A.; Ryszka, P.; Kovács, G. M. Two new genera, Dominikia and Kamienskia, and D. disticha sp. nov. in Glomeromycota. Nova Hedwigia 2015, 100(1–2), 225–238. [CrossRef]
- Berkeley, M.J.; Broome, C.E. Enumeration of the fungi of Ceylon. Part II. J. Linn. Soc. London Bot. 1873, 14:137.
- Wu, C.G.; Chen, Z.C. The Endogonaceae of Taiwan Ⅱ. Two New Species of Sclerocystis from Taiwan. Trans. Mycol. Soc. Republic China 1987, 2(2), 73–83.
- Goto, B.T., Bezerra, J.L.; Maia, L.C. Sclerocystis coremioides (Glomeromycota) forming epigeous sporocarps on organic substrates of cocoa plantations in the Atlantic Forest of Bahia. Agrotrópica 2016, 28(1), 23–28.
- Spain, J. L.; Miranda, J.D. Scutellospora cerradensis: an ornamented species in the Gigasporaceae (Glomales) from the Cerrado region of Brazil. Mycotaxon 1996, 60, 129–136.
- De Souza, F.A.; Declerck, S.; Kowalschuk, G.A. Morphological, ontogenetic and molecular characterization of Scutellospora reticulata (Glomeromycota). Mycol. Res. 2005, 109(6), 697–706. [CrossRef]
- Manawasinghe, I.S.; Hyde, K.D.; Wanasinghe, D.N.; Karunarathna, S.C.; Maharachchikumbura, S.S.; Samarakoon, M.C.; Voglmayr, H.; Pang, K.-L.; Chiang, M. W.-L.; Jones, E.B.G.; et al. Fungal diversity notes 1818–1918: taxonomic and phylogenetic contributions on genera and species of fungi. Fungal Divers. 2025, 130(1), 1–261. [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



