
Submitted:
26 August 2025
Posted:
28 August 2025
You are already at the latest version
Abstract
Over the past few years, consumers’ preference for wines with a distinct regional identity has shifted producers to adopt traditional grape varieties and has increased interest in the indigenous grapevine microbiome, due to its potential influence on the wine quality characteristics. In the present study, indigenous Saccharomyces cerevisiae strains were isolated from three local grape varieties (“Keratsouda”, “Karnachalas”, and “Bogialamas”) of the Soufli region in Northern Greece. After molecular identification, all strains were evaluated for key oenological traits (ethanol resistance, SO₂ resistance, H₂S production, and amino‑acid decarboxylation). Three representative strains (one per grape variety) were further evaluated for their long-term survival after freeze-drying and their fermentation potential at 12 °C and 22 °C. Ethanol content ranged in similar values (11.9–12.5%) for all wines, but kinetic parameters were significantly affected by the strain used and the fermentation temperature. Principal Component Analysis revealed that the volatile composition as well as practical wine attributes were affected by both the strain used and the fermentation temperature and resulted in distinct sensory profiles in the new products.
Keywords:
indigenous S. cerevisiae strains
; regional grape varieties
; yeast technological characterization
; wine aromatic profile
1. Introduction
Industrial winemaking practices rely on the extensive use of a few selected commercial Saccharomyces cerevisiae strains often promoted under different product names and brands [1,2]. The dynamic nature of these starter cultures ensures the effective completion of the alcoholic fermentation by outcompeting the native grape microflora and also provides specific sensory characteristics to the final product [3]. However, as a direct consequence of the decrease of microbial communities involved in fermentations, a series of constrains has risen for the wine sector. Reduction of wine profiles’ uniqueness, general flattening of the bouquet and organoleptic uniformity of products, even across different vitivinicultural regions, are the most common malfunctions observed [4,5].
Nevertheless, based on the idea that indigenous yeasts may provide unique flavors that preserve and reflect the regional characteristics of a vineyard, many winemakers have shifted again towards spontaneous fermentations [3,6]. In fact, a vineyard’s microflora is an interactive ecosystem, unique for each grape variety, that is highly affected by the geographical location, the climate, as well as the vitivinicultural practices applied. This results in significant species diversity that represent and highlight the regional character of the vineyard, simply called ‘terroir’ [7-11].
This aspect has led to a growing market and industrial interest in isolating and exploiting indigenous yeasts [12] in an effort to produce unique wines, but also guarantee consistent fermentation outcomes across vintages [7,13]. Various indigenous strains of S. cerevisiae have been isolated the past years from various grape varieties (e.g. Cabernet Sauvignon, Malbec, “Gioia del Coll” DOC, Merlot, Garnacha and others) worldwide [9]. These strains have developed to regional-specific starter cultures showing quick adaptation to their specific environments, enhanced fermentation efficiency and production of unique aroma profiles [11,14,15].
Greek vineyards represent a complex mixture of international and domestic varieties. Specifically, in the Soufli region of Northern Greece “Keratsouda”, a highly productive rose variety with bright pink color, is grown [16]. Similarly, “Karnachalas” and “Bogialamas” are two genetically close varieties that are typically used as a mixture by local winemakers [17]. The indigenous yeast communities of these varieties have not been yet studied, but in an effort to promote the whole region they could be carefully exploited and result in signature wines with high market potential [17,18].
In this context, the aim of the present study was the isolation and of new S. cerevisiae strains from the local varieties “Karnachalas”, “Keratsouda”, and “Biogialamas” of the Soufli region in Northern Greece. All strains were selected according to their technological characteristics and evaluated concerning their impact to wine fermentation and quality attributes. Thus, the isolated yeast strains were assessed as potential fermentation starters in order to enhance the expression of the regional characteristics of the produced wines.
2. Materials and Methods
2.1. Isolation of New Yeast Strains from Local Grape Varieties
Grape samples from the 3 local varieties (~2 kg for each), “Karnachalas”, “Keratsouda”, and “Bogialamas” were provided by the company “Winery – Distillery Gr Bellas L.P.” (coordinates: 41.183865, 26.260774) during the 2023 vintage, in late October.
For each variety, stems along with any bruised or rotten grapes were removed by hand and the remaining grapes were crushed in an iMix bag mixer (Interlab, Mourjou, France). The extracted grape must was then transferred to batch bioreactors (0.5 L) and left to perform spontaneous alcoholic fermentation at ~25 °C. Fermentations were monitored daily (by Be° measurements) until full sugar utilization occurred or no further activity was observed. After completion, samples were taken from the mid-zone of each bioreactor, decimally diluted and plated on sterile YPD agar (yeast extract 10 g/L, peptone 20 g/L, dextrose 20 g/L, agar 20 g/L). Agar plates were incubated at 30 °C for 48-72 h and the dilutions on which yeast colonies were clearly distinguishable were chosen for re-streaking. All yeast strains were routinely kept in Malt broth and Malt agar plates (Condalab, Madrid, Spain) at 4 °C until further use. Stock samples were kept at -80 °C by inoculating fully grown cultures in 30% v/v glycerol tubes [6].
The new strains were observed under an optical microscope. After examining their cellular morphology, 10 strains in total, resembling S. cerevisiae cells, were selected for molecular identification.
2.2. Molecular Identification of the New Strains
Total DNA of the new isolated strains, was extracted using the Nucleospin® DNA Yeast kit (Macherey-Nagel GmbH & Co. KG, Düren, Germany), following the manufacturer’s instructions. The highly variable Internal Transcribed Spacer (ITS) regions of the ITS1 and ITS2 sequences were amplified with the primers ITS1-4.
Polymerase Chain Reaction (PCR) amplification was performed using the HotStarTaq Plus Master Mix Kit (Qiagen, Germatown, MD, USA), consisting of 35 cycles with the following steps: 94 °C for 3 min, 35 cycles of 94 °C for 30 s, 53 °C for 40 s, 72 °C for 90 s, and the final elongation step at 72 °C for 5 min. PCR products were then subjected to electrophoresis in 2% agarose gel to confirm amplification and to determine the relative intensity of bands. Then, the amplicons were purified using Ampure PB beads (Pacific Biosciences).
PCR products were subjected to electrophoresis on 2% agarose gels to confirm amplification and assess band intensity. Subsequently, a purification step was performed using Ampure XP beads (Beckman Coulter, Brea, California, USA).
Sequences obtained by MR DNA (www. mrdnalab.com, Shallowater, TX, USA) were taxonomically classified using the BLASTn algorithm against the NCBI reference database (www.ncbi.nlm.nih.gov).
2.3. Technological Characterization of the New Strains
• Ethanol resistance: Each yeast strain was spot-inoculated on grape must-agar plates containing increasing concentrations of ethanol (up to 14% vol). Resistance to ethanol was evaluated on the basis of positive growth after 72 h at 26 °C [19].
• SO2 resistance: Each yeast strain was spot-inoculated on grape must-agar plates containing increasing concentrations of SO2 in the form of the commercially available potassium metabisulfite. Plates were incubated at 26 °C for 48 h and the colony growth size was assessed (no growth, small growth, medium growth, high growth) [20].
• H2S production: Each yeast strain was spot-inoculated on Bismuth Sulfite Glucose Glycine Yeast (BIGGY) agar and incubated at 26 °C for 48 h. H2S production was assessed based on the color formation of the colonies: white (no production); yellow (low production); dark yellow (moderate production); brown or black (high production), as previously described [20].
• Decarboxylation of amino acids: Each yeast strain was spot-inoculated on modified YPG agar plates (5 g/L bacteriological peptone; 5 g/L yeast extract; 1 g/L glucose; 20 g/L agar), as recently described [21]. The medium was supplemented with 10 g/L of each amino acid and 6 mL of a 1% ethanol/water solution of bromocresol purple indicator at a final concentration of 0.5%. Before sterilization, the pH was adjusted to 5.2 in all cases. The plates were incubated at 26 °C for 3-5 days. The formation of a purple ring around the grown colonies indicated positive reaction of the strain.
The strains evaluated for their technological characteristics were grown overnight at Malt broth at 30 °C. Prior to each analysis, they were diluted using sterile ¼ Ringer’s solution (VWR International GmbH, Radnor, PA, USA) at a concentration of 105 cfu/mL. All analyses were performed in triplicate.
2.4. Freeze-Drying and Monitoring of Yeast Cell Viability During Storage
Three yeast strains (one for each variety) were selected based on their technological characteristics, and examined for cell survival during long-term storage.
Each yeast strain was initially grown overnight at 1 L flask with Malt broth at 30 °C. Yeast cells were collected by centrifugation and subsequently freeze-dried on a BenchTop Pro (Virtis, SP Scientific, Warminster, PA, USA) [22]. The freeze-dried cultures were then stored at 20-25 oC, 4 oC and -18 oC for a period up to 9 months. Cell survival was monitored in frequent intervals (1, 3, 6, and 9 months). 1 g of freeze-dried yeast cells biomass was transferred to 9 mL of sterile ¼ Ringer’s solution, serial decimal dilutions were performed, and plate counting on Malt agar plates after incubation at 30 °C for 72 h. All analyses were performed in triplicate and the survival rate % at each time interval was calculated as logcfu/g, divided by the initial yeast cell counts (logcfu/g), and multiplied by 100, as previously described [23].
2.5. Fermentation Efficiency of the New Strains
Fermentation efficiency of the 3 new yeast strains was evaluated in batch fermentations (500 mL) at 22 °C and 12 °C, based on the region’s climate during the winemaking season. Fresh varietal grape musts: “Keratsouda” must (total sugars content: 250±0.5 g/L), and “Karnachalas”/’Bogialamas” must as a mixture (total sugars content: 210±0.5 g/L) were provided by the company “Winery – Distillery Gr Bellas L.P.” immediately after pressing (2024 vintage).
Fermentation inoculums were prepared as previously described [6]. Fully grown yeast strains were firstly inoculated to the grape must of its origin (50 mL) and subsequently re-inoculated to the bioreactors (500 mL) reaching a final concentration of 106 cfu/mL.
All fermentations were performed in three independent replicates. Wine samples were collected after completion of each fermentation and subjected to chemical analyses.
2.6. Chemical and Sensory Analyses
2.6.1. pH, Total Acidity, and Volatile Acidity
pH was determined with a pH-330i pH meter (WTW GmbH, Germany) following the manufacturer’s guidelines. Total acidity was estimated by titration with 4 g/L NaOH solution. Volatile acidity was determined by titration (with 4 g/L NaOH) after steam distillation with an Electronic Distiller (DUALSTILL Exacta + Optech Labcenter S.p.a., Italy) [24]. All analyses were performed in triplicate.
2.6.2. Ε. thanol, Glycerol, Residual Sugars, and Organic Acid Content
Εthanol, glycerol, residual sugars, and organic acid content were determined on a Shimadzu chromatography system (Shimadzu Corp., Germany) equipped with a Nucleogel ION 300 OA column (300 x 7.8mm) (Macherey-Nagel, Germany). Peak separation was performed at 85 °C and H2SO4 (0.049 g/L) was used as the mobile phase at 0.3 mL/min. All concentrations were calculated using standard curves prepared by standard solutions (R2 ≥ 0.99). Fermentation parameters were calculated as previously described [24]. All analyses were performed in triplicate.
2.6.3. Major Volatiles
Gas Chromatography system [MASTER GC Fast Gas Chromatograph (DANI Instruments S.p.a., Italy)], equipped with a CP-Wax 57 CB column (50 m x 0.32 mm i.d., 0.20 μm film thickness) (Chrompack), was used for the determination of wine major volatiles (acetaldehyde, ethyl acetate, 1-propanol, isobutanol, 1-hexanol, amyl alcohol, isoamyl alcohol, and methanol), as previously described [24]. Wine samples (1 μL) were injected at 250 °C in 1:20 split mode. Oven temperature was initially set at 40 °C (with a 5 min hold), followed by a raise to 180 °C at a rate of 3 °C/min. All analyses were performed in triplicate.
Clarity MASTER GC Communication integrated software (DANI Instruments S.p.a.) was used for data processing. Concentration of volatile compounds were calculated using standard curves prepared by standard solutions (R2 ≥ 0.99).
2.6.4. Minor Volatiles
HS-SPME GC/MS analysis [6890N GC, 5973NetworkedMS MSD (Agilent Technologies, USA), HP-5MS column (30m, 0.25mm i.d., 0.25 μm film thickness)] was used for the determination of minor volatiles content. For each wine, 10 mL of sample and 3 g of NaCl were transferred into a 20-mL headspace vial and 4-methyl-2-pentanol was added as internal standard. The vial was thermostated at 60 °C for 45 min and minor volatiles were isolated by a 2-cm fibre (50/30 mm DVB/CAR/PDMS) (Supelco, USA). Compounds were desorbed at 240 °C (injector temperature) at splitless mode for 3 min and a ramp program was followed for their analysis. Specifically, column temperature was initially set at 35 °C (with a 6 min hold), increased to 60 °C at a rate of 2 °C/min (with a 5 min hold), increased again to 200 °C at a rate of 5 °C/min, and finally increased to 250 °C at a rate of 25 °C/min (with a final 6 min hold). Helium was used as the carrier gas (linear velocity of 1.8 mL/min). All analyses were performed in triplicate.
ChemStation integrated software (Agilent Technologies) was used for data processing and all compounds were identified by comparing mass spectra with spectra from the NBS75K and Wiley275 reference libraries, by spectra from standard compounds (in-house libraries) and by determining the Kovats retention indexes and comparing them with those reported in the literature. All volatile compounds were semi-quantified and expressed as mg/L, as previously described [25].
2.6.5. Preliminary Sensory Evaluation
Aroma and taste of the produced wines were assessed by a mixed panel of 10 tasters (males and females equally represented), as previously described [26]. All participants were young enthusiasts (age up to 35 years old) and frequent wine consumers, but had no official wine-tasting training and were not involved in the project at any way. For this reason, detailed instructions and training was provided on basic aspects regarding wine-glass handling, wine swirling, wine smelling, and wine tasting. In order to avoid confusion, expected aroma (fruity, winelike, spirituous, yeasty, piquant) and taste attributes (saltness, sweetness, sourness, bitterness) were explained prior to the evaluation. All sensory trials were blind and randomized and wines were served in a colored glass, covered with a glass cup (to avoid aroma dispersion), under low light at 12–15 °C. The participants were asked to identify the aroma and taste characteristics of each sample and evaluate their perception on a 0–5 scale (0: unacceptable, 5: wonderful). Additional information regarding aftertaste, body, and overall quality were also collected. The tasters were reminded to rinse their palate with water and eat a cracker between samples.
2.7. Statistical Analysis
Statistical significance of oenological parameters was examined by 2-way analysis of variance. Duncan’s multiple range test was used to determine significant differences (P < 0.05) among results. Statistical significance at P < 0.05, coefficients and ANOVA tables were computed by Statistica v.12.0 (Stat Soft Inc., Tulsa, OK, USA).
Principal Component Analysis was computed by XLSTAT 2015.1 (Addinsoft, Paris, France).
3. Results and Discussion
3.1. Yeast Strains Isolation and Characterization
Isolation of indigenous yeast strains was performed after the spontaneous alcoholic fermentations for each grape variety must were completed. Samples were plated on YPD agar and after incubation colonies were selected (from the dish with the greater dilution) based on their characteristics under microscope. Ten colonies, strongly resempling Saccharomyces species morphology, proceeded to molecular identification and further evaluation.
The results confirmed that all indigenous strains exhibited ≥ 99% sequence similarity to Saccharomyces cerevisiae. The findings confirmed the dominance of S. cerevisiae among the yeast microbiota of the studied grape varieties and are consistent with previous studies emphasizing the prevalence of this species during spontaneous alcoholic fermentation [27,28].
All 10 selected indigenous strains showed resistance to SO2 concentrations ranging 200-300 mg/L (Table 1), as previously documented for indigenous strains [8,29]. All new strains were capable of growing at 14% (v/v) ethanol with the adaptation rate ranging from “low” to “high”, although even higher ethanol resistance has been documented in previous studies [7,29,30].
Hydrogen sulfide is naturally produced by yeasts as a result of the reduction of inorganic sulfur compounds [31]. It may play a metabolic and protective role during the early phase of fermetation and potentially participate in other signalling processes, too [32,33]. All our strains (with the exception of BO-Y1) exhibited high H2S production (Table 1), which is not uncommon for indigenous S. cerevisiae strains [20]. However, overproduction of H2S often results in off-odours, so application of mitigation strategies like copper-fining or purging wine in a reductive state with inert gas is commonly advised [34]. Other factors, like the presence of residual sulfur in the grapes), assimilable nitrogen levels of must, or even the aeration effect during fermentation may also be responsible for high H2S values and thus should be carefully adjusted by winemakers [34-36].
Amino acids are known to contribute to various yeast metabolic pathways. In come cases however, their decarboxylation by certain yeast strains may result to the production of biogenic amines and raise safety concerns for the consumer [37]. In our case (Table 1), arginine and tyrosine was universally metabolized, as previously shown for indigenous strains [20]. Specifically, arginine is one of the most important sources for yeasts as it is involved in both anabolic and catabolic pathways [38] and tyrosine may be degraded in order to produce important aroma compounds, like fusel alcohols [39]. Tryptophane and leucine were also metabolized with different effectiveness rates among strains, while no response was acquired for histidine, lysine, and phenylalanine.
It is widely known that maintaining fresh microbial cultures for extended periods is a significant challenge for the industry. Thus, the storage of starter cultures between winemaking seasons is highly important and represents a viable and sustainable solution [40]. Microbial preservation mainly relies on application of drying techniques like freeze-drying, thermal-drying and others that offer protection against contamination, maintenance of cell viability through long preservation times, as well as storage and handling convenience [41].
In order to investigate the long-term cell viability of the newly isolated yeast strains, freeze-drying was applied and the freeze-dried cells were stored at ambient (20-25 °C), refrigerator (4 °C), and freezing conditions (-18 °C). In frequent time intervals (1, 3, 6, and 9 months), samples were analyzed and cell viability was determined. The results are summarized in Figure 1. As expected, low storage temperatures (4 & -18 °C) resulted in significantly higher (Ρ < 0.05) survival rates compared to ambient temperatures, at all cases. Low temperatures are strongly preferred in order to effectively prolong microbial survival [42,43], but still a small drop in yeast counts was inevitable through storage duration, but not significant in all cases. Interestingly, survival rates documented (at 4 & -18 °C) ranged in levels similar or better that previously documented for freeze-dried yeast cultures [44].
In any case, the survival of dried microorganisms during long-term storage is a dynamic process that depends on various factors such as microbial growth conditions, initial cell concentration, the drying medium, and more [45,46]. Protective agents have shown to further enhance survival of microorganisms [47], but carry-over effect and matters of cost-effectiveness advise against their use [22,48]. However, it should be noted that strain-specific characteristics that may affect the ability of microorganisms to withstand freeze-drying and storage cannot be excluded [49], even amongst different strains of the same species [40].
3.2. Fermentations
All new yeast strains were evaluated regarding their fermentation efficiency in batch fermentations at 22 oC and 12 oC. The temperature selection was based on the region’s climate during the winemaking season (October and November), according to the Hellenic National Meteorological Service [50]. Fermentation kinetic data and other important parameters are presented in Table 2.
Both the strain used and the fermentation temperature affected significantly (P < 0.05) the fermentation duration, the ethanol productivity, the residual sugars content and sugar conversion, the organic acids and glycerol concentration. Total acidity and volatile acidity values were significantly affected (Ρ < 0.05) only by the strain used. Strong interactions (P < 0.05) were noted in all the above cases. On the contrary, no significant differences were observed for ethanol content, ethanol production yield, and pH values.
As expected, fermentations at low temperatures (12 °C) led to increased fermentation duration [22,24]. Significantly higher (P < 0.05) residual sugars occurred in all cases at 12 °C, ranging 17.9-38.2 g/L (in mean values), that later resulted in significantly lower conversion values comparing to 22 °C. Notably though, the higher residual sugars content is not considered a fault and may be utilized in order to provide a discreet flavor sweetness to the final product [26,51,52]. Interestingly, and despite of the differences noted in sugar consumption among fermentations performed at 12 and 22 °C (Table 2), all final products presented similar ethanol concentration (11.9-12.5 % vol). Temperature (high or low) is one of the main factors influencing yeast metabolism and directly affecting wine fermentation. Specifically, fermentations at low temperatures have been suggested to favor the growth of non-Saccharomyces populations rather than that of S. cerevisiae strains, thus affecting consumption of available carbon sources [51,53]. Given the nature of the fresh varietal musts used in this experiment, and despite the inoculum applied (final concentration of 106 cfu/mL), the potential presence and action of non-Saccharomyces populations at the early stages of fermentation can not be excluded. Additionally, improved ethanol production yield was recorded at 12 °C, but not significant in all cases. In fact, fermentation performance of yeasts is known to be strain-specific and similar findings have been previously documented in wine production with commercial and indigenous S. cerevisiae strains in lower temperatures (< 15 °C) [53,54]. Regarding ethanol productivities, values up to 10.5 g/(Ld) were noted in fermentations at 22 °C, but even at 12 °C the observed values were higher or similar to those usually found in traditional winemaking [55,56].
Glycerol is a fermentation-derived secondary metabolite, mainly known for contributing to the wine's flavor by adding sweetness and “fullness”, ultimately providing a “rounder” mouthfeel [57]. Glycerol production is widely affected by the fermenting yeast strain and the temperature applied, but other factors like sulfur dioxide levels, agitation, or even the must’s pH might also influence its final content [51]. In our samples, the highest (P < 0.05) glycerol concentration (11.0±0.8 g/L) was observed in wines fermented by KA-Y1 at 12 °C (Table 2). In the rest of the cases, however, intermediate values (5.7–7.0 g/L), typically encountered in fermentations with indigenous S. cerevisiae strains, were observed [8,9].
Regarding organic acids, acetic acid is produced naturally in wine as a result of fermenting microorganisms’ metabolism [51]. In low concentrations (up to 0.3 g/L), it may add to the wine’s complexity, but certain products (like Canadian ice wines) rely on high concentrations (often exceeding 2.0 g/L) for their special sensory character [58]. In our samples, acetic acid was detected in high quantities only in wines fermented with KE-Y1 strain. Nevertheless, differences (as well as accumulation) of acetic acid is not uncommon in fermentations performed by indigenous S. cerevisiae strains and has been previously associated to potentially diminished acetyl-CoA synthetase capabilities [59]. On the other hand, lactic acid was found in levels ranging 0.3-2.3 g/L, but also is widely known to be highly depended on the wine type, the occurrence of malolactic fermentation, the grape cultivar and the viticultural practices applied [60,61].
Total acidity and pH was found in values typical for wines [22,51,52]. On the contrary, elevated volatile acidities were found at fermentations performed at 12 °C, but not significantly in all cases [25]. Notably, the highest (P < 0.05) volatile acidities were detected in wines fermented with the KE-Y1 strain, probably a direct consequence of high acetic acid content (Table 2). According to OIV, volatile acidity should not exceed values of 1.2 g of acetic acid/L [62]. The application of physicochemical methods (e.g. reverse osmosis), the use of biological deacidification (by re-fermentation of the final product), or blending with low volatile acidity wines are some of the strategies that may effectively address this issue [63,64]. It should be noted though, that in special occasions, high volatile acidity (> 2 g/L) is lawfully permitted and may be utilized as a tool by experienced winemakers in order to develop complex organoleptic attributes [65]. In any case, no fault (like vinegar taint or any other) was detected in our samples during the preliminary sensory evaluation.
3.4.1. Major Wine Volatiles
Μajor wine volatiles detected are shown in Table 3. Acetaldehyde, ethyl acetate, isobutanol, 1-hexanol, and methanol concentrations were significantly (P < 0.05) affected by both the indigenous strain used and the fermentation temperature. Amyl and isoamyl alcohols, and 1-propanol concentrations were significantly (P < 0.05) affected only by the strain used. Strong interactions (P < 0.05) were observed between the factors in almost all cases (with the exception of acetaldehyde).
Acetaldehyde is one of the major wine constituents, which when found in low concentrations is known to provide fruit scents [66]. Ιn wines fermented with the KA-Y1 strain, it was detected in significantly higher (P < 0.05) concentrations exceeding 200 mg/L. In general, its presence in concentrations up to 125 mg/L is acceptable in wines [64], while in higher levels it may be considered responsible for the “oxidised” [67] or “flat character” of the product [63]. Nevertheless, concentrations over 220 mg/L are also essential for the special organoleptic character found in products like fortified wines [63]. In any case, no implications were observed in our samples during the preliminary sensory evaluation (subsection 3.5).
Production of ethyl acetate is known to be affected by the fermentation temperature, as well as by the strain of S. cerevisiae used, with ‘natural yeast’ fermentations often tending to result in concentrations higher than average [68]. In all our cases however, ethyl acetate levels ranged between 31-127 mg/L (in mean values). Typically, ethyl acetate values up to 150 mg/L are considered to provide a positive fragrance complexity to the final products [51].
Higher alcohols (amyl alcohol, isoamyl alcohol, isobutanol, and 1-propanol) consist a group of compounds that is considered to have the strongest impact on the overall aroma of wine [69]. In low concentrations, they add to the product’s aromatic complexity by adding fruity notes [63,70], but on higher levels, they might confer a masking effect on the character of the wine [71]. It should be noted, however, that whether the overall contribution of higher alcohols is positive or negative is highly variable and depends heavily on the total aromatic context of the product [72]. In our samples, higher alcohols were found in high concentrations for all strains, but never exceeded 550 mg/L (in mean values) [63], and no unpleasant scents were detected.
Methanol and 1-hexanol, on the contrary, were detected in low concentrations < 50 mg/L in all wines.
Wines fermented by indigenous S. cerevisiae strains at 12 & 22 °C were subjected to HS-SPME GC/MS analysis, in order to determine the minor volatiles responsible for the aroma. According to the results (Table 4), both the indigenous strain used and the fermentation temperature affected significantly (P < 0.05) all the volatile groups of interest (esters, alcohols, organic acids, miscellaneous, and total volatile content) and strong interactions (P < 0.05) were observed between the two factors in all cases.
The largest group of minor volatiles detected in our samples (as shown in Table 4 and Figure 2) was esters. Their concentration was significantly higher (P < 0.05) in wines produced at 22 °C comparing to wines produced at 12 °C (Table 4). Esters are fermentation-derived compounds that originate enzymatically from higher alcohols or plain ethanol and many of them are associated with fruity, floral, sweet, honey and other pleasing scents contributing to the wine aroma profile [73,74].
Alcohols were the second most abundant group of volatile compounds identified (Table 4). Similarly to esters, their concentrations were elevated at 22 °C, but not significantly in all cases (and not for all strains). On the contrary, very low concentrations of organic acids and carbonyls occurred in both fermentation temperatures (Table 4). Organic acids are mainly known for their low odor threshold limit and their potential impact on wine flavor by contributing to the product’s “freshness” [75,76]. Except for their aromatic contributions, in the case of increased concentrations the synthesis of their corresponding esters might also be favored [76]. Aldehydes and ketones are produced naturally during fermentation or occur as oxidation byproducts, although in many cases their exact source of origin is difficult to discern [77]. They present low odor thresholds and are known to enhance aroma and flavor complexity [78].
In total, 41 volatile compounds were identified. As shown in Figure 2 (heatmap), variations were observed among samples for all volatile compounds detected, potentially determining the final aromatic complexity of each wine. Specifically, characteristic ester subgroups like acetate esters and ethyl fatty acid esters were identified in all samples, but in greater concentrations in wines fermented at 22 °C. Specifically, ethyl propanoate may be responsible for fresh notes detected in wine, while ethyl hexanoate, ethyl octanoate, ethyl decanoate, and 2-phenylethyl acetate usually add common fruity notes [79]. Distinct banana-like scents, peer-like scents or apple-peel notes may be attributed to 3-methylbutyl acetate, 2-methylbutyl acetate, and ethyl butyrate, respectively. On the contrary, compounds like ethyl dodecanoate may be responsible for more smokey, toasty and earthy aromas resulting in the dry character of the final product [51,80-82]. In any case, it should be noted that excessive ester amounts may produce pungent odor and foul taste and impact negatively the wine quality.
Key alcohol compounds like phenylethyl alcohol and 1-hexanol were present in all samples (Figure 2) contributing mostly rose aroma and floral-fruity scents, respectively. Similarly, citronellol (adding citrus and sweet notes) was identified in most cases [83-85]. Eucalyptol, characterized by a distinct green aroma, was found in all samples and is believed to be adding “fresh”, and “cool” notes to the product [86]. On the other hand, compounds like 2,3-butanediol, known for adding bitter-sweet notes) [51,87] and 1-heptanol (present in most of the samples), may be considered to be of low to none sensory importance for wine, but their presence has been previously associated with the discriminant classification of regional wines [88].
Regarding organic acids, octanoic acid was detected in all samples (Figure 2). Hexanoic and decanoic acid were also detected in low concentrations in most of the wines. As of carbonyl compounds, only β-damascenone, known for contributing to the varietal scent with intense, exotic, flower or rose-like notes was detected in most of the samples produced at 12 °C (Figure 2). A few miscellaneous compounds were also observed in most of the samples. 1,1-Diethoxyethane, which may indeed contribute to the wine bouquet [51,81], was detected in both fermentations temperatures (12 & 22 °C) (Figure 2), and is primarily known for its fruity-green notes and refreshing character.
3.4.3. Chemometrics
HS-SPME GC/MS results and practical wine attributes (residual sugars, glycerol and ethanol concentration, organic acids content, total and volatile acidity, and pH) were also subjected to Principal Component Analysis. As shown in Figure 3, the two principal components accounted for 65.06% of the total variance with ethanol concentration, residual sugars, lactic and acetic acid content, volatile acidity, and pH contributing mostly to PC1, while minor volatile compounds (esters, higher alcohols, and miscellaneous compounds) contributed mostly to PC2. Both the strain used and the fermentation temperature applied seem to have affected significantly the volatile composition and the oenological parameters in the new wines.
Specifically, wines produced with the KE_Y1 strain were located to the top right quadrant of the graph and (as shown previously in Table 2) were characterized by significantly higher (P < 0.05) residual sugars and acetic acid content, as well as significantly higher (P < 0.05) volatile acidity and pH values comparing to wines fermented with the other strains. Similarly, wines produced by the KA-Y1 strain gathered to the top left quadrant and were associated with high concentration of esters, alcohols, carbonyls, and miscellaneous compounds. Interestingly, wines fermented with the BO-Y1 strain at 22 °C were located close to wines fermented with the KA-Y1 strain, indicating broadly similar aromatic characteristics between these strains at 22 °C (as shown previously in Table 3). In fact, fermentation temperature is a crucial factor regarding the aromatic complexity, as fermentation at 22 °C correlated positively on PC2 for most samples (especially BO-Y1). Notably, KA-Y1_12 and KA-Y1_22 samples were closely grouped in the plot indicating relatively stable performance of the strain in a wide temperature range (12 and 22 °C), an attribute of high importance for the wine sector.
3.5. Preliminary Sensory Evaluation
All wines fermented with indigenous S. cerevisiae strains were evaluated for their sensory aroma and flavor characteristics. According to the results (Table S1), all products were mostly characterized as light-bodied with a predominant wine-like aroma, a mostly sour taste, and a mild aftertaste. A strong aftertaste, however, was noted in wines fermented with the KA-Y1 strain, possibly as a result of increased acetaldehyde levels. Nevertheless, the flavor outcome was not unpleasant, but characterized as interesting and piquant-like mostly contributing to the character of the product. Notably, no vinegar or other repulsive scents were detected in any of the wines produced with the KE-Y1 strain (or any other strain), despite the significantly high (P < 0.05) content of acetic acid and high volatile acidity values (as shown previously in Table 2).
3.6. Technological Considerations and Current Limitations
In this work, indigenous S. cerevisiae strains were isolated from local grape varieties (“Karnachalas”, “Keratsouda”, and “Bogialamas”) and subsequently used to ferment the grape must (of their origin). Technological advantages derived from this strategy include, among others, the enhanced strain adaptability to each must’s conditions, better regulation of the native microbial flora and improved control of alcoholic fermentation. At the same time, the utilization of lower fermentation temperatures for enhancing the production of aromatic compounds and improving the wine quality is well known between experienced winemakers. These combined actions aim to showcase the unique regional terroir characteristics in the final product [89-91]. Thus, our findings offer local winemakers the opportunity to create and promote 'signature wines' by following the example of several Spanish wineries that have already obtained the 'Pago' designation. This term constitutes a form of Spanish Geographical Indication [9,92] and is used to describe wines coming from distinct regions with specific soil and microclimate characteristics (like Soufli), produced with traditional vineyard and winemaking techniques and most importantly fermented by the indigenous yeast strains each winery has in possession [93,94].
However, it should be noted that the proposed methodology cannot be adopted uncritically. Future experiments should primarily focus on confirming and replicating the obtained results. Isolation of more strains may facilitate the creation of a native yeasts collection able to produce wines with more typical wine characteristics (e.g. lower H2S production, lower volatile acidity values, etc) and should be a priority, as a single winemaking season is not sufficiently representative of the region’s terroir. Prioritizing the effective combination of yeast strains for enhanced aroma development, along with the isolation and inoculation of malolactic bacteria, is essential. Generally, simple parameters like temperature affect the fermentation process and influence the microbial metabolic pathways [95]. Thus, their impact over multiple native strains is a variable that may be effectively utilized for the formation of unique sensory attributes and should be further explored in future experiments. Innovative technologies such as immobilization on natural supports should also be examined, as they have been shown to boost culture survival during storage, enhance fermentation activity, lower volatile acidity, and contribute positively to the wine’s character [22,23,48,96]. Finally, the performance of the selected strains should be evaluated in industrial conditions. Apart from the above, current findings may be used as a roadmap and provide useful insights for the targeted isolation of native yeasts (from different grape varieties) in the region or across other geographical regions. Advanced omics methodologies may be applied in a later stage in order to combine all available data in an effort to predict the functional outcomes of native fermentations. In any case, the detailed technological characterization of the indigenous microbial flora and the aromatic profile of the wines produced (in each grapevine region, like Soufli) is the vital step in order to facilitate scale up and generate in the future positive outcomes not only for the local winemakers, but also for the region, the consumers and wine-industry in general.
4. Conclusions
In the current study, 10 indigenous S. cerevisiae strains (ITS sequence similarity ≥ 99%) were isolated from local grape varieties in the Soufli region in Northern Greece. All strains exhibited satisfactory ethanol resistance (up to 14% vol), variable SO₂ resistance (200–300 mg/L), predominantly high H₂S production, and distinct amino acid decarboxylation profiles. Long-term survival of 3 freeze-dried selected strains was examined and rates ≥ 75% even after 9 months at low storage temperatures (4 & -18 °C) were recorded. Fermentation parameters were significantly influenced by both the strain used and the fermentation temperature (12 or 22 °C), but high ethanol content [11.9–12.5% (v/v)] and ethanol productivities higher or similar to those usually found in traditional winemaking were observed. PCA revealed that both the strain used and the fermentation temperature seem to have affected significantly the volatile composition and practical wine attributes resulting in products with distinct sensory profiles. In conclusion, the utilization of indigenous S. cerevisiae strains could present a viable strategy for the preservation and promotion of the regional terroir, enabling production of potentially high quality “signature wines”, in various temperatures.
Author Contributions
Conceptualization, A.N. and Yi.K.; data curation, A.N., I.P. and N.P.; funding acquisition, Yi.K. and Yo.K.; investigation, A.N., I.K. and V.K.; methodology, A.N. and I.P.; project administration, Yi.K. and Yo.K.; supervision, Yi.K.; writing—original draft, A.N.; writing—review and editing, A.N. and Yi.K. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the project “INNOVATION IN LOCAL WINE VARIETIES” [MIS Μ16ΣΥΝ2-00141], RURAL DEVELOPMENT PROGRAMME (RDP) 2014 – 2020, Sub-measure 16.1 – 16.2 - Action 2, co-funded by the European Union and Greek National Funds.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Fernández-Espinar, M.T.; López, V.; Ramón, D.; Bartra, E.; Querol, A. Study of the authenticity of commercial wine yeast strains by molecular techniques. Int. J. Food Microbiol. 2001, 70, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Vigentini, I.; Fracassetti, D.; Picozzi, C.; Foschino, R. Polymorphisms of Saccharomyces cerevisiae genes involved in wine production. Curr. Microbiol. 2009, 58, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Whiteley, L.E.; Rieckh, G.; Diggle, F.L.; Alaga, Z.M.; Nachbaur, E.H.; Nachbaur, W.T.; Whiteley, M. Use of commercial or indigenous yeast impacts the S. cerevisiae transcriptome during wine fermentation. Microbiol. Spectr. 2024, 12. [Google Scholar] [CrossRef] [PubMed]
- Csoma, H.; Zakany, N.; Capece, A.; Romano, P.; Sipiczki, M. Biological diversity of Saccharomyces yeasts of spontaneously fermenting wines in four wine regions: Comparative genotypic and phenotypic analysis. Int. J. Food Microbiol. 2010, 140, 239–248. [Google Scholar] [CrossRef]
- Di Maio, S.; Polizzotto, G.; Di Gangi, E.; Foresta, G.; Genna, G.; Verzera, A.; Scacco, A.; Amore, G.; Oliva, D. Biodiversity of Indigenous Saccharomyces Populations from Old Wineries of South-Eastern Sicily (Italy): Preservation and Economic Potential. PLoS ONE 2012, 7, e30428. [Google Scholar] [CrossRef]
- Kontogiannatos, D.; Troianou, V.; Dimopoulou, M.; Hatzopoulos, P.; Kotseridis, Y. Oenological Potential of Autochthonous Saccharomyces cerevisiae Yeast Strains from the Greek Varieties of Agiorgitiko and Moschofilero. Beverages 2021, 7, 27. [Google Scholar] [CrossRef]
- Capece, A.; Pietrafesa, R.; Siesto, G.; Romaniello, R.; Condelli, N.; Romano, P. Selected Indigenous Saccharomyces cerevisiae Strains as Profitable Strategy to Preserve Typical Traits of Primitivo Wine. Fermentation 2019, 5, 87. [Google Scholar] [CrossRef]
- Liu, J.; Li, R.; Li, Y.; Sun, Y. Selection of indigenous Saccharomyces cerevisiae strains with good oenological and aroma characteristics for winemaking in Ningxia China, Food Chem. : X. 2024, 23, 101693. [Google Scholar] [CrossRef]
- Berbegal, C.; Ferrer, S.; Polo, L.; Pardo, I.; García-Esparza, M.J.; Andrés, L.; Álvarez, I.; Lizama, V. Diversity of Indigenous Saccharomyces cerevisiae Yeast Strains in Cabernet Sauvignon Fermentations from Utiel-Requena Region (Spain) as a Resource to Improve Wine Distinctiveness. Fermentation 2023, 9, 654. [Google Scholar] [CrossRef]
- Costantini, A.; Cravero, M.C.; Panero, L.; Bonello, F.; Vaudano, E.; Pulcini, L.; Garcia-Moruno, E. Wine Fermentation Performance of Indigenous Saccharomyces cerevisiae and Saccharomyces paradoxus Strains Isolated in a Piedmont Vineyard. Beverages 2021, 7, 30. [Google Scholar] [CrossRef]
- Castellanos, E.; Pico, J.; Moore, J.; Dumayne, M.; Coletta, C.; Castellarin, S.; Measday, V. Suitability of indigenous Saccharomyces cerevisiae strains isolated from spontaneous fermentations as wine fermentation starter cultures in the Okanagan Valley wine region in Canada. OENO One 2025, 59. [Google Scholar] [CrossRef]
- Chen, Y.; Jiang, J.; Song, Y.; Zang, X.; Wang, G.; Pei, Y.; Song, Y.; Qin, Y.; Liu, Y. Yeast Diversity during Spontaneous Fermentations and Oenological Characterisation of Indigenous Saccharomyces cerevisiae for Potential as Wine Starter Cultures. Microorganisms 2022, 10, 1455. [Google Scholar] [CrossRef] [PubMed]
- Mas, A.; Padilla, B.; Esteve-Zarzoso, B.; Beltran, G.; Reguant, C.; Bordons, A. Taking advantage of natural biodiversity for wine making: The WILDWINE Project. Agric. Agric. Sci. Procedia 2016, 8, 4–9. [Google Scholar] [CrossRef]
- Aponte, M.; Romano, R.; Villano, C.; Blaiotta, G. Dominance of S. cerevisiae Commercial Starter Strains during Greco di Tufo and Aglianico Wine Fermentations and Evaluation of Oenological Performances of Some Indigenous/Residential Strains. Foods 2020, 9, 1549. [Google Scholar] [CrossRef]
- Zabukovec, P.; Cadez, N.; Cus, F. Isolation and Identification of Indigenous Wine Yeasts and Their Use in Alcoholic Fermentation. Food Technol. Biotechnol. 2020, 58(3), 337–347. [Google Scholar] [CrossRef]
- Biniari, K.; Stavrakaki, M. Genetic study of native grapevine varieties of Northern, Western and Central Greece with the use of ampelographic and molecular methods. Not. Bot. Horti Agrobot. 2019, 47, 46–53. [Google Scholar] [CrossRef]
- Miliordos, D.E.; Merkouropoulos, G.; Kogkou, C.; Arseniou, S.; Alatzas, A.; Proxenia, N.; Hatzopoulos, P.; Kotseridis, Y. Explore the Rare—Molecular Identification and Wine Evaluation of Two Autochthonous Greek Varieties: “Karnachalades” and “Bogialamades”. Plants 2021, 10, 1556. [Google Scholar] [CrossRef]
- Molinet, J.; Cubillos, F.A. Wild yeast for the future: Exploring the use of wild strains for wine and beer fermentation. Front. Genet. 2020, 11, 589350. [Google Scholar] [CrossRef]
- Mauriello, G.; Capece, A.; D’ Auria, M.; Garde-Cerdan, T.; Romano, P. SPME–GC method as a tool to differentiate VOC profiles in Saccharomyces cerevisiae wine yeasts. Food Microbiol. 2009, 26, 246–252. [Google Scholar] [CrossRef]
- Banilas, G.; Sgouros, G.; Nisiotou, A. Development of microsatellite markers for Lachancea thermotolerans typing and population structure of wine-associated isolates. Microbiol. Res. 2016, 193, 1–10. [Google Scholar] [CrossRef]
- Siesto, G.; Corbo, M.R.; Pietrafesa, R.; Sinigaglia, M.; Romano, P.; Bevilacqua, A. Screening of Saccharomyces and Non-Saccharomyces Wine Yeasts for Their Decarboxylase Activity of Amino Acids. Foods 2022, 11, 3587. [Google Scholar] [CrossRef] [PubMed]
- Nikolaou, A.; Sgouros, G.; Mitropoulou, G.; Santarmaki, V.; Kourkoutas, Y. Freeze-dried Immobilized Kefir Culture in Low Alcohol Wine-Making. Foods 2020, 9, 115. [Google Scholar] [CrossRef] [PubMed]
- Prapa, I.; Nikolaou, A.; Panas, P.; Tassou, C.; Kourkoutas, Y. Developing Stable Freeze-Dried Functional Ingredients Containing Wild-Type Presumptive Probiotic Strains for Food Systems. Appl. Sci. 2023, 13, 630. [Google Scholar] [CrossRef]
- Nikolaou, A.; Galanis, A.; Kanellaki, M.; Tassou, C.; Akrida-Demertzi, K.; Kourkoutas, Y. Assessment of free and immobilized kefir culture in simultaneous alcoholic and malolactic cider fermentations. LWT-Food Sci. Technol. 2017, 76, 67–78. [Google Scholar] [CrossRef]
- Nikolaou, A.; Tsakiris, A.; Kanellaki, M.; Bezirtzoglou, E.; Akrida-Demertzi, K.; Kourkoutas, Y. Wine production using free and immobilized kefir culture on natural supports. Food Chem. 2019, 272, 39–48. [Google Scholar] [CrossRef]
- Nikolaou, A.; Kourkoutas, Y. High-Temperature Semi-Dry and Sweet Low AlcoholWine-Making Using Immobilized Kefir Culture. Fermentation 2021, 7, 45. [Google Scholar] [CrossRef]
- Agarbati, A.; Comitini, F.; Ciani, M.; Canonico, L. Occurrence and Persistence of Saccharomyces cerevisiae Population in Spontaneous Fermentation and the Relation with “Winery Effect”. Microorganisms 2024, 12, 1494. [Google Scholar] [CrossRef]
- Suzuki, N.; Iwahashi, H.; Watanabe, D. Yeast diversity during the spontaneous fermentation of wine in a winery and in a laboratory. Int. J. Food Microbiol. 2024, 411, 110345. [Google Scholar] [CrossRef]
- Feng, L.; Jia, H.; Wang, J.M.; Qin, Y.; Liu, Y.L.; Song, Y.Y. Selection of indigenous Saccharomyces cerevisiae strains for winemaking in northwest China. Am. J. Enol. Vitic. 2019, 70, 115–126. [Google Scholar] [CrossRef]
- Suranská, H.; Vránová, D.; Omelková, J. Isolation, identification and characterization of regional indigenous Saccharomyces cerevisiae strains. Braz. J. Microbiol. 2016, 47, 181–190. [Google Scholar] [CrossRef]
- Swiegers, J.H.; Pretorius, I.S. Modulation of volatile sulfur compounds by wine yeast. Appl. Microbiol. Biotechnol. 2007, 74, 954–960. [Google Scholar] [CrossRef]
- Huang, C.-W.; Walker, M.E.; Fedrizzi, B.; Gardner, R.C.; Jiranek, V. Hydrogen sulfide and its roles in Saccharomyces cerevisiae in a winemaking context. FEMS Yeast Res. 2017, 17. [Google Scholar] [CrossRef]
- Hou, R.; Jelley, R.E.; van Leeuwen, K.A.; Pinu, F.R.; Fedrizzi, B.; Deed, R.C. Hydrogen Sulfide Production during Early Yeast Fermentation Correlates with Volatile Sulfur Compound Biogenesis but Not Thiol Release. FEMS Yeast Res. 2023, 23, foad031. [Google Scholar] [CrossRef]
- Sánchez-Gimeno, D.; Vela, E.; Ferreira, V.; Ontañón, I. Alternative strategies for eliminating hydrogen sulfide and methanethiol from wine: results and learnings. OENO One 2024, 58. [Google Scholar] [CrossRef]
- Bekker, M.Z.; Day, M.P.; Holt, H.; Wilkes, E.; Smith, P.A. Effect of oxygen exposure during fermentation on volatile sulfur compounds in Shiraz wine and a comparison of strategies for remediation of reductive character. Aust. J. Grape Wine Res. 2016, 22, 24–35. [Google Scholar] [CrossRef]
- Jastrzembski, J.A.; Allison, R. B.; Friedberg, E.; Sacks, G.L. Role of Elemental Sulfur in Forming Latent Precursors of H2S in Wine. J. Agric. Food Chem. 2017, 65, 10542–10549. [Google Scholar] [CrossRef] [PubMed]
- Speranza, B.; Campaniello, D.; Petruzzi, L.; Sinigaglia, M.; Corbo, M.R.; Bevilacqua, A. Preliminary characterization of yeasts from Bombino Bianco, a grape variety of Apulian Region, and selection of an isolate as a potential starter. Fermentation 2019, 5, 102. [Google Scholar] [CrossRef]
- Messenguy, F.; Dubois, E. Regulation of Arginine Metabolism in Saccharomyces cerevisiae: A Network of Specific and Pleiotropic Proteins in Response to Multiple Environmental Signals. Food Technol. Biotechnol. 2000, 38, 277–285. [Google Scholar]
- Ter Schure, E.G.; Flikweert, M.T.; van Dijken, J.P.; Pronk, J.T.; Verrips, C.T. Pyruvate decarboxylase catalyzes decarboxylation of branched-chain 2-oxo acids but is not essential for fusel alcohol production by Saccharomyces cerevisiae. Appl. Environ. Microbiol. 1998, 64, 1303–1307. [Google Scholar] [CrossRef]
- N'Guessan, F.K.; Coulibaly, H.W.; Alloue-Boraud, M.W.A.; Cot, M.; Djè, K.M. Production of freeze-dried yeast culture for the brewing of traditional sorghum beer, tchapalo. Food Sci. Nutr. 2016, 4, 34–41. [Google Scholar] [CrossRef]
- Miyamoto-Shinohara, Y.; Sukenobe, J.; Imaizumi, T.; Nakahara, T. Survival curves for microbial species stored by freeze-drying. Cryobiology 2006, 52, 27–32. [Google Scholar] [CrossRef]
- Tripathi, M.K.; Giri, S.K. Probiotic functional foods: Survival of probiotics during processing and storage. J. Funct. Foods 2014, 9, 225–241. [Google Scholar] [CrossRef]
- Jofré, A.; Aymerich, T.; Garriga, M. Impact of Different Cryoprotectants on the Survival of Freeze-Dried Lactobacillus Rhamnosus and Lactobacillus Casei/Paracasei during Long-Term Storage. Benef. Microbes 2015, 6, 381–386. [Google Scholar] [CrossRef] [PubMed]
- Nyanga, L.K.; Nout, M.J.; Smid, E.J.; Boekhout, T.; Zwietering, M.H. Yeasts preservation: alternatives for lyophilisation. World J. Microbiol. Biotechnol. 2012, 28, 3239–3244. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, A.S.; Silva, J.; Ho, P.; Teixeira, P.; Malcata, F.X.; Gibbs, P. Survival of freeze- dried Lactobacillus plantarum and Lactobacillus rhamnosus during storage in the presence of protectants. Biotechnol. Lett. 2002, 24, 1587–1591. [Google Scholar] [CrossRef]
- Morgan, C.A.; Herman, N.; White, P.A.; Vesey, G. Preservation of microorganisms by drying; a review. J. Microbiol. Methods 2006, 66, 183–193. [Google Scholar] [CrossRef]
- Hubalek, Z. Protectants used in the cryopreservation of microorganisms. Cryobiology 2003, 46, 205–229. [Google Scholar] [CrossRef]
- Nikolaou, A.; Nelios, G.; Kanellaki, M.; Kourkoutas, Y. Freeze-dried immobilized kefir culture in cider-making. J. Sci. Food Agric. 2020, 100, 3319–3327. [Google Scholar] [CrossRef]
- Nikolaou, A.; Mitropoulou, G.; Nelios, G.; Kourkoutas, Y. Novel Functional Grape Juices Fortified with Free or Immobilized Lacticaseibacillus rhamnosus OLXAL-1. Microorganisms 2023, 11, 646. [Google Scholar] [CrossRef]
- Hellenic National Meteorological Service. https://emy.gr/en/climatic-data?tab=statistics-tab (Accessed on July 2025).
- Jackson, R.S. Wine Science: Principles and Applications, 4th ed.; Academic Press Inc.: San Diego, CA, USA, 2014. [Google Scholar]
- Mallios, P.; Kourkoutas, Y.; Iconomopoulou, M.; Koutinas, A.A.; Psarianos, C.; Marchant, R.; Banat, I.M. Low-temperature wine-making using yeast immobilized on pear pieces. J. Sci. Food Agric. 2004, 84, 1615–1623. [Google Scholar] [CrossRef]
- Torija, M.J.; Rozès, N.; Poblet, M.; Guillamón, J.M.; Mas, A. Effects of fermentation temperature on the strain population of Saccharomyces cerevisiae. Int. J. Food Microbiol. 2003, 80, 47–53. [Google Scholar] [CrossRef]
- Gao, Y.-T.; Zhang, Y.-S.; Wen, X.; Song, X.-W.; Meng, D.; Li, B.-J.; Wang, M.-Y.; Tao, Y.-Q.; Zhao, H.; Guan, W.-Q.; Du, G. The glycerol and ethanol production kinetics in low-temperature wine fermentation using Saccharomyces cerevisiae yeast strains. Int. J. Food Sci. Technol. 2019, 54, 102–110. [Google Scholar] [CrossRef]
- Kopsahelis, N.; Bosnea, L.; Kanellaki, M.; Koutinas, A.A. Volatiles Formation from Grape Must Fermentation Using a Cryophilic and Thermotolerant Yeast. Appl. Biochem. Biotechnol. 2012, 167, 1183–1198. [Google Scholar] [CrossRef] [PubMed]
- Sipsas, V.; Kolokythas, G.; Kourkoutas, Y.; Plessas, S.; Nedovic, V.A.; Kanellaki, M. Comparative study of batch and continuous multi-stage fixed-bed tower (MFBT) bioreactor during wine-making using freeze-dried immobilized cells. J. Food Eng. 2009, 90, 495–503. [Google Scholar] [CrossRef]
- Scanes, K.T.; Hohrnann, S.; Prior, B.A. Glycerol Production by the Yeast Saccharomyces cerevisiae and its Relevance to Wine: A Review. S. Afr. J. Enol. Vitic. 1998, 19, 17–24. [Google Scholar] [CrossRef]
- Nurgel, C.; Pickering, G.J.; Inglis, D.L. Sensory and chemical characteristics of Canadian ice wines. J. Sci. Food Agric. 2004, 84, 1675–1684. [Google Scholar] [CrossRef]
- Rodicio, R.; Heinisch, J.J. Sugar metabolism in yeasts. In Biology of Microorganisms on Grapes, in Must and in Wine; König, H., Unden, G., Frölich, J., Eds.; Springer International Publishing AG: Cham, Switzerland, 2017; pp. 189–213. [Google Scholar]
- Heras-Roger, J.; Díaz-Romero, C.; Darias-Rosales, J.; Darias-Martín, J. Organic Acids in Varietal Red Wines: Influence of Grape Cultivar, Geographical Origin, and Aging. Beverages 2025, 11, 102. [Google Scholar] [CrossRef]
- Volschenk, H.; Van Vuuren, H.; Viljoen-Bloom, M. Malic acid in wine: Origin, function and metabolism during vinification. S. Afr. J. Enol. Vitic. 2006, 27, 123–136. [Google Scholar] [CrossRef]
- Organisation International de la Vigneet du Vin. OIV-MA-AS4-01 Type IV. Microbiological Analysis of Wines and Musts Methodm, Compendium of International Methods of Analysis-OIV: Paris, France, 2010.
- Ribereau-Gayon, P.; Glories, Y.; Maujean, A.; Dubourdieu, D. The microbiology of wine and vinifications. In Handbook of Enology; Wiley: Chichester, UK, 2006; Volume 1. [Google Scholar]
- Zoecklein, W.B.; Fugelsang, C.K.; Gump, H.B.; Nury, S.F. Wine analysis and production; Chapman and Hall Inc.: New York, USA, 1995. [Google Scholar]
- Cliff, M.A.; Pickering, G.J. Determination of odour detection thresholds for acetic acid and ethyl acetate in ice wine. J. Wine Res. 2006, 17, 45–52. [Google Scholar] [CrossRef]
- Arias-Pérez, I.; Sáenz-Navajas, M.P.; de-la-Fuente-Blanco, A.; Ferreira, V.; Escudero, A. Insights on the role of acetaldehyde and other aldehydes in the odour and tactile nasal perception of red wine. Food Chem. 2021, 361, 130081. [Google Scholar] [CrossRef]
- Noble, A.C.; Arnold, R.A.; Buechsenstein, J.; Leach, E.J.; Schmidt, J.O.; Stern, P.M. Modification of a standardized system of wine aroma terminology. Am. J. Enol. Vitic. 1987, 38, 143–146. [Google Scholar] [CrossRef]
- Grainger, K. Excessive Volatile Acidity and Ethyl Acetate. In Wine Faults and Flaws; John Wiley & Sons Ltd: Chichester, West Sussex, UK, 2021. [Google Scholar] [CrossRef]
- Rigou, P.; Mekoue, J.; Sieczkowski, N.; Doco, T.; Vernhet, A. Impact of industrial yeast derivative products on the modification of wine aroma compounds and sensorial profile. A review. Food Chem. 2021, 358, 129760. [Google Scholar] [CrossRef]
- Bartowsky, E.J.; Pretorius, I.S. Microbial formation and modification of flavor and off-flavor compounds in wine. In Biology of Microorganisms on Grapes, in Must and in Wine; König, H., Unden, G., Fröhlich, J., Eds.; Springer: Heidelberg/Berlin, Germany, 2009; pp. 209–231. [Google Scholar]
- Cameleyre, M.; Lytra, G.; Tempere, S.; Barbe, J.C. Olfactory Impact of Higher Alcohols on Red Wine Fruity Ester Aroma Expression in Model Solution. J. Agric. Food Chem. 2015, 63, 9777–9788. [Google Scholar] [CrossRef]
- de-la-Fuente-Blanco, A.; Sáenz-Navajas, M.P.; Ferreira, V. On the effects of higher alcohols on red wine aroma. Food Chem. 2016, 210, 107–114. [Google Scholar] [CrossRef]
- Jeromel, A.; Korenika, A.M.J.; Tomaz, I. 6—an influence of different yeast species on wine aroma composition. In Fermented Beverages; Grumezescu, A.M., Holban, A.M., Eds.; Woodhead Publishing: Cambridge, UK, 2019. [Google Scholar] [CrossRef]
- Perestrelo, R.; Silva, C.; Gonçalves, C.; Castillo, M.; Câmara, J.S. An approach of the madeira wine chemistry. Beverages 2020, 6. [Google Scholar] [CrossRef]
- Lan, Y.B.; Xiang, X.F.; Qian, X.; Wang, J.M.; Ling, M.Q.; Zhu, B.Q.; Liu, T.; Sun, L.B.; Shi, Y. , Reynolds, A.G.; Duan, C.Q. Characterization and differentiation of key odor-active compounds of 'Beibinghong' icewine and dry wine by gas chromatography-olfactometry and aroma reconstitution. Food Chem. 2019, 287, 186–196. [Google Scholar] [CrossRef] [PubMed]
- Ma, T.; Sam, F.E.; Didi, D.A.; Atuna, R.A.; Amagloh, F.K.; Zhang, B. Contribution of edible flowers on the aroma profile of dealcoholized pinot noir rose wine. LWT-Food Sci. Technol. 2022, 170, 114034. [Google Scholar] [CrossRef]
- Waterhouse, A.L.; Sacks, G.L; Jeffery, D.W. Aldehydes, Ketones, and Related Compounds. In Understanding Wine Chemistry; John Wiley & Sons Ltd: Chichester, West Sussex, UK, 2016. [Google Scholar] [CrossRef]
- Januszek, M.; Satora, P.; Wajda, Ł.; Tarko, T. Saccharomyces bayanus Enhances Volatile Profile of Apple Brandies. Molecules 2020, 25, 3127. [Google Scholar] [CrossRef] [PubMed]
- Meilgaard, M.C. Effects on flavor of innovations in brewery equipment and processing: A review. J. Inst. Brew. 2001, 107, 271–286. [Google Scholar] [CrossRef]
- Rajoka, M.I.; Ferhan, M.; Khalid, A.M. Kinetics and thermodynamics of ethanol production by a thermotolerant mutant of Saccharomyces cerevisiae in a microprocessor-controlled bioreactor. Lett. Appl. Microbiol. 2005, 40, 316–321. [Google Scholar] [CrossRef]
- Etiévant, X.P. Wine. In Volatile Compounds in Foods and Beverages; Maarse, H., Ed.; Marcel Dekker: New York, NY, USA, 1991; pp. 483–533. [Google Scholar]
- Miranda-Lopez, R.; Libbey, L.M.; Watson, B.T.; McDaniel, M.R. Odor analysis of pinot noir wines from grapes of different maturities by a Gas chromatography-olfactometry technique (Osme). J. Food Sci. 1992, 57, 985–993. [Google Scholar] [CrossRef]
- Pena-Alvarez, A.; Capella, S.; Juarez, R.; Labastida, C. Determination of terpenes in tequila by solid phase microextraction-gas chromatography-mass spectrometry. J. Chromatogr. A 2006, 1134, 291–297. [Google Scholar] [CrossRef]
- Vilanova, M.; Sieiro, C. Determination of free and bound terpene compounds in Albarino wine. J. Food Compos. Anal. 2006, 19, 694–697. [Google Scholar] [CrossRef]
- Zhou, Y.; Chao, Y.; Huang, C.; Li, X.; Yi, Z.; Zhu, Z.; Yan, L.; Ding, Y.; Peng, Y.; Xie, C. Influence of Lactiplantibacillus plantarum and Saccharomyces cerevisiae Individual and Collaborative Inoculation on Flavor Characteristics of Rose Fermented Beverage. Foods 2025, 14, 1868. [Google Scholar] [CrossRef] [PubMed]
- Capone, D.L.; Van Leeuwen, K.; Taylor, D.K.; Jeffery, D.W.; Pardon, K.H.; Elsey, G.M.; Sefton, M.A. Evolution and occurrence of 1,8-cineole (eucalyptol) in Australian wine. J. Agric. Food Chem. 2011, 59, 953–959. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Arribas, M.V.; Polo, M.C. Wine Chemistry and Biochemistry; Springer: New York, NY, USA, 2009. [Google Scholar]
- Zhang, L.; Liu, Q.; Li, Y.; Liu, S.; Tu, Q.; Yuan, C. Characterization of wine volatile compounds from different regions and varieties by HS-SPME/GC-MS coupled with chemometrics. Curr. Res. Food Sci. 2022, 6, 100418. [Google Scholar] [CrossRef]
- Lopes, C.A.; Rodriguez, M.E.; Sangorrin, M.; Querol, A.; Caballero, A.C. Patagonian wines: The selection of an indigenous yeast starter. J. Ind. Microbiol. Biotechnol. 2007, 34, 539–546. [Google Scholar] [CrossRef]
- Suranska, H.; Vranova, D.; Omelkova, J. Isolation, identification and characterization of regional indigenous Saccharomyces cerevisiae strains. Braz. J. Microbiol. 2016, 47, 181–190. [Google Scholar] [CrossRef]
- Could Better Adapt to the Physicochemical Conditions and Natural Microbial Ecology of Prince Grape Must Compared with Commercial Saccharomyces cerevisiae FX10. Molecules 2022, 27, 6892. [CrossRef]
- OIV. Resolution OIV/VITI 333/2010, Definition of Vitivinicultural “TERROIR”, 2010. Available online: https://www.oiv.int/public/medias/379/viti-2010-1-en.pdf (accessed on July 2025).
- Ut, C.; Berbegal, C.; Lizama, V.; Polo, L.; García, M.J.; Andrés, L.; Pardo, I.; Álvarez, I. Isolation and characterisation of autochthonous Saccharomyces cerevisiae from ‘Pago’ Merlot wines of Utiel-Requena (Spain) Origin. Aust. J. Grape Wine Res. 2021, 28, 330–346. [Google Scholar] [CrossRef]
- Berbegal, C.; Polo, L.; Lizama, V.; Álvarez, I.; Ferrer, S.; Pardo, I.; García-Esparza, M.J. Influence of native S. cerevisiae strains on the final characteristics of “Pago” Garnacha wines from East Spain. Beverages 2023, 9, 17. [Google Scholar] [CrossRef]
- Zhou, Y.; Chao, Y.; Huang, C.; Li, X.; Yi, Z.; Zhu, Z.; Yan, L.; Ding, Y.; Peng, Y.; Xie, C. Influence of Lactiplantibacillus plantarum and Saccharomyces cerevisiae Individual and Collaborative Inoculation on Flavor Characteristics of Rose Fermented Beverage. Foods 2025, 14, 1868. [Google Scholar] [CrossRef] [PubMed]
- Nikolaou, A.; Sgouros, G.; Santarmaki, V.; Mitropoulou, G.; Kourkoutas, Y. Preliminary Evaluation of the Use of Thermally-Dried Immobilized Kefir Cells in Low Alcohol Winemaking. Appl. Sci. 2022, 12, 6176. [Google Scholar] [CrossRef]
Figure 1.
Survival rate % of freeze-dried indigenous S. cerevisiae strains during long-term storage at different conditions. Strain codes: KA-Y1, S. cerevisiae strain isolated from “Karnachalas” grapes; KE-Y1, S. cerevisiae strain isolated from “Keratsouda” grapes; BO-Y1, S. cerevisiae strain isolated from “Bogialamas” grapes. Each bar represents the mean value of three independent replicates, with error bars representing the Standard Deviation.
Figure 1.
Survival rate % of freeze-dried indigenous S. cerevisiae strains during long-term storage at different conditions. Strain codes: KA-Y1, S. cerevisiae strain isolated from “Karnachalas” grapes; KE-Y1, S. cerevisiae strain isolated from “Keratsouda” grapes; BO-Y1, S. cerevisiae strain isolated from “Bogialamas” grapes. Each bar represents the mean value of three independent replicates, with error bars representing the Standard Deviation.
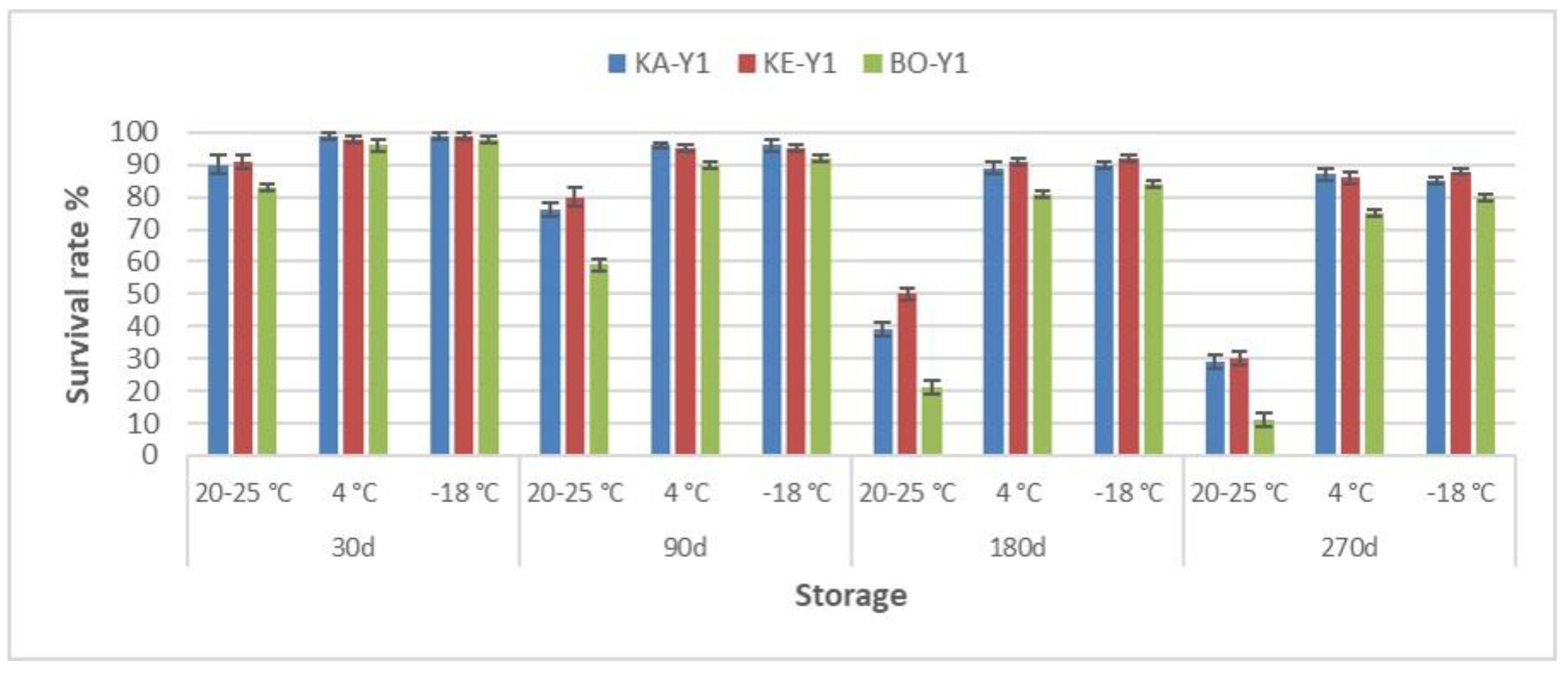
Figure 2.
Heatmap of minor volatiles in wines fermented by indigenous S. cerevisiae strains at 12 & 22 °C. Differences in volatile concentrations are shown for each compound in blue (min) and red (max) shades. KA-Y1, wine fermented by S. cerevisiae strain isolated from “Karnachalas” grapes; KE-Y1, wine fermented by S. cerevisiae strain isolated from “Keratsouda” grapes; BO-Y1, wine fermented by S. cerevisiae strain isolated from “Bogialamas” grapes.
Figure 2.
Heatmap of minor volatiles in wines fermented by indigenous S. cerevisiae strains at 12 & 22 °C. Differences in volatile concentrations are shown for each compound in blue (min) and red (max) shades. KA-Y1, wine fermented by S. cerevisiae strain isolated from “Karnachalas” grapes; KE-Y1, wine fermented by S. cerevisiae strain isolated from “Keratsouda” grapes; BO-Y1, wine fermented by S. cerevisiae strain isolated from “Bogialamas” grapes.
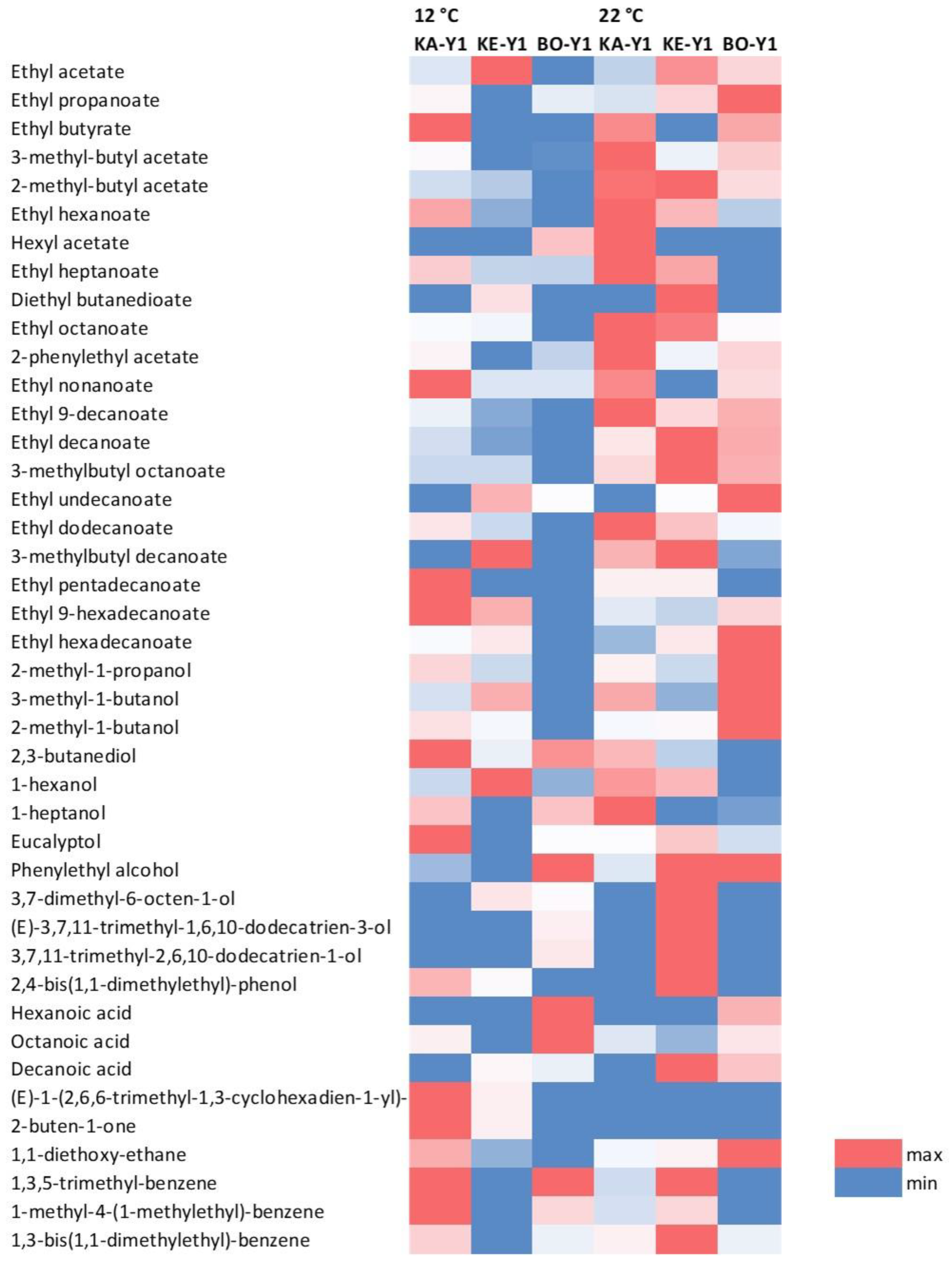
Figure 3.
PCA plot of minor volatiles and practical fermentation attributes of wines produced with indigenous yeast strains at 12 & 22 °C. KA-Y1, wine fermented by S. cerevisiae strain isolated from “Karnachalas” grapes; KE-Y1, wine fermented by S. cerevisiae strain isolated from “Keratsouda” grapes; BO-Y1, wine fermented by S. cerevisiae strain isolated from “Bogialamas” grapes. The fermentation temperature is indicated at the end of each sample code.
Figure 3.
PCA plot of minor volatiles and practical fermentation attributes of wines produced with indigenous yeast strains at 12 & 22 °C. KA-Y1, wine fermented by S. cerevisiae strain isolated from “Karnachalas” grapes; KE-Y1, wine fermented by S. cerevisiae strain isolated from “Keratsouda” grapes; BO-Y1, wine fermented by S. cerevisiae strain isolated from “Bogialamas” grapes. The fermentation temperature is indicated at the end of each sample code.
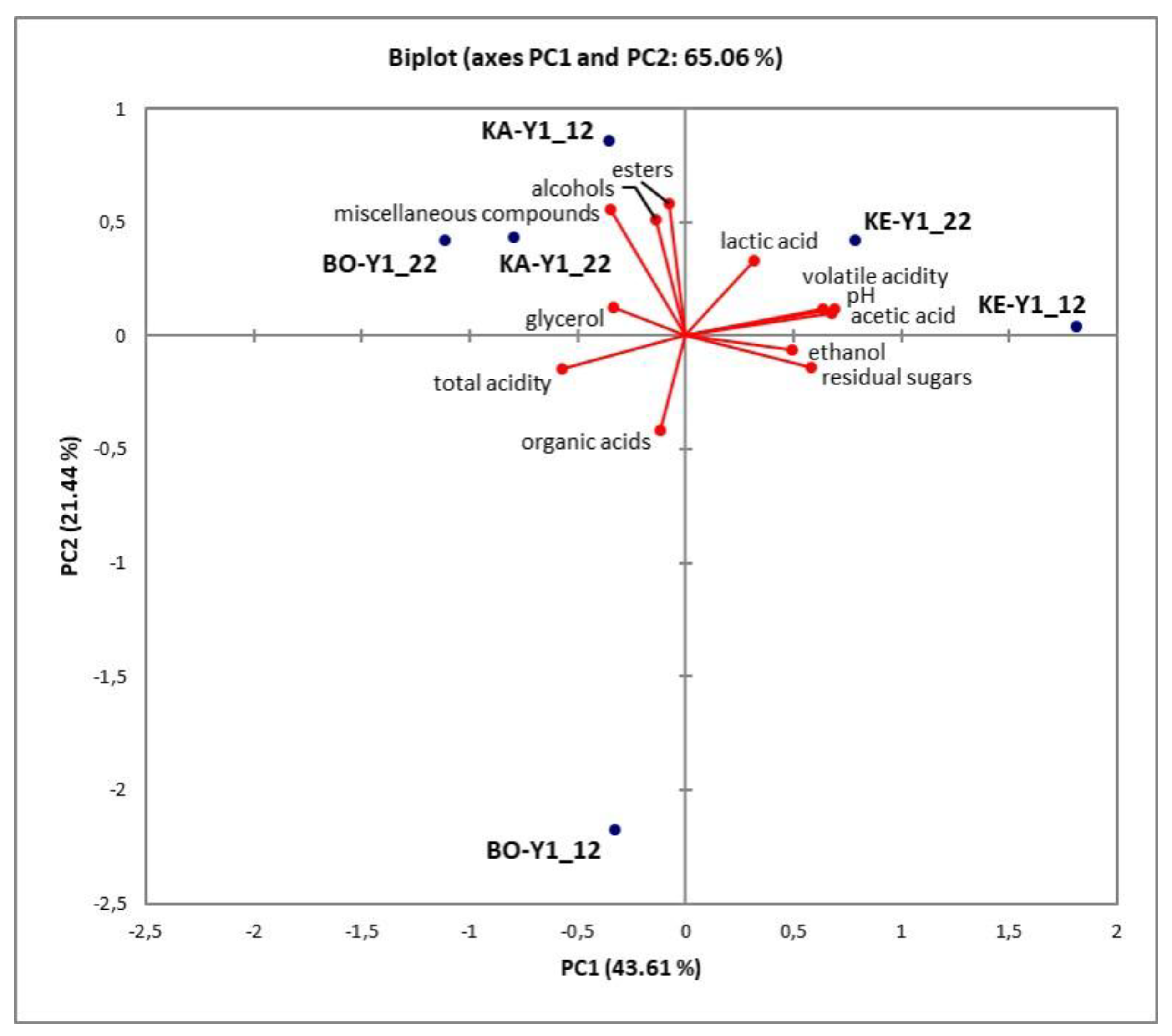

Table 1.
Technological characterization of indigenous S. cerevisiae strains.
| Grape variety | Yeast strain |
Ethanol resistance (14% vol) |
*SO2 resistance | H2S production | Decarboxylation of amino acids | |||
| Arginine | Tyrosine | Tryptophane | Leucine | |||||
| Karnachalas | ΚA-Y1 | High | 200 | High | + | + | - | - |
| ΚA-Υ3 | High | 300 | High | + | + | + | + | |
| ΚA-Υ4 | Low | 300 | High | + | + | + | + | |
| KA-YL2 | Medium | 300 | High | + | + | + | + | |
| KA-YL4 | Medium | 300 | High | + | + | + | + | |
| Keratsouda | KE-Y1 | Low | 200 | High | + | + | - | - |
| KE-Y2 | Low | 300 | High | + | + | - | + | |
| Bogialamas | BO-Y1 | Medium | 200 | Medium | + | + | + | + |
| BO-YL1 | Medium | 300 | High | + | + | + | + | |
| BO-YL2 | High | 250 | High | + | + | + | + | |
*: Expressed as mg L-1 of potassium metabisulfite; +/-: Positive/negative decarboxylation activity, respectively; Yeast strain codes: KA-Y1, S. cerevisiae strain isolated from “Karnachalas” grapes; KE-Y1, S. cerevisiae strain isolated from “Keratsouda” grapes; BO-Y1, S. cerevisiae strain isolated from “Bogialamas” grapes.
Table 2.
Kinetic parameters and organic acid profile of wines fermented by indigenous S. cerevisiae strains at 12 & 22 °C.
Table 2.
Kinetic parameters and organic acid profile of wines fermented by indigenous S. cerevisiae strains at 12 & 22 °C.
| Fermentation temperature | Yeast strain |
Fermentation time (d) |
Ethanol concentration (% vol) |
Residual sugars (g L-1) |
Ethanol productivity (g (L d)-1) | Ethanol production yield |
Conversion (%) |
Glycerol (g L-1) |
Lactic acid (g L-1) |
Acetic acid (g L-1) |
Total acidity (g tartaric L-1) |
Volatile acidity (g acetic L-1) |
pH |
| 12 °C | KA-Y1 | 21±2b | 11.9±0.5a | 17.9±1.0b | 4.5±0.1e | 0.49±0.02a,b | 91.5±0.6c | 11.0±0.8a | 2.3±0.2a | nd | 6.5±0.4b,c | 1.17±0.06c | 3.4±0.2a |
| KE-Y1 | 47±3a | 12.5±0.7a | 38.2±4.1a | 2.1±0.1f | 0.47±0.03a,b | 84.7±1.5d | 5.7±0.4c | 2.1±0.1a | 2.5±0.2a | 6.0±0.4c | 1.92±0.15b | 3.8±0.3a | |
| BO-Y1 | 19±1b,c | 12.0±0.8a | 18.4±0.9b | 5.0±0.1d | 0.49±0.02a | 91.2±0.4c | 6.8±0.5b,c | 0.3±0.1c | nd | 7.5±0.5a,b | 0.99±0.06c,d | 3.3±0.1a | |
| 22 °C | KA-Y1 | 9±0.5d | 12.0±0.5a | 4.9±0.5d | 10.5±0.1a | 0.46±0.03a,b | 97.7±0.2a | 6.8±0.6b,c | 0.3±0.1c | nd | 7.8±0.6a | 0.90±0.03d | 3.3±0.1a |
| KE-Y1 | 16±1c | 11.9±0.7a | 12.0±2.1c | 5.9±0.1c | 0.40±0.03b | 95.2±0.5b | 3.8±0.4d | 1.4±0.1b | 1.8±0.1b | 5.2±0.3c | 2.19±0.15a | 3.7±0.2a | |
| BO-Y1 | 10±0.5d | 12.0±0.5a | 7.3±06d | 9.5±0.1b | 0.47±0.03a,b | 96.5±0.2a,b | 7.0±0.5b | 0.3±0.1c | nd | 8.5±0.6a | 0.75±0.03d | 3.3±0.1a |
Yeast strain codes: KA-Y1, S. cerevisiae strain isolated from “Karnachalas” grapes; KE-Y1, S. cerevisiae strain isolated from “Keratsouda” grapes; BO-Y1, S. cerevisiae strain isolated from “Bogialamas” grapes. Data shown are mean values of three independent fermentations ± Standard Deviation. Different lowercase letters within a column indicate significant differences among values (2-way ANOVA: Duncan’s multiple range test, P < 0.05). nd: not detected.3.4. Volatiles.
Table 3.
Μajor volatiles of wines fermented by indigenous S. cerevisiae strains at 12 & 22 °C.
|
Fermentation temperature |
Yeast strain | Acetaldehyde (mg L-1) |
Ethyl acetate (mg L-1) |
1-Propanol (mg L-1) | Isobutanol (mg L-1) | 1-Hexanol (mg L-1) | Amyl alcohol (mg L-1) |
Isoamyl alcohol (mg L-1) |
Methanol (mg L-1) |
| 12 °C | KA-Y1 | 206±18a | 73±6c | 42±3c | 81±6c,d | 9±1c | 71±5c | 194±19b | 25±3a |
| KE-Y1 | 67±7d | 80±6c | 7±1d | 54±5e | 2±1d | 49±3d | 147±16c | 9±1d | |
| BO-Y1 | 121±12c | 71±8c | 75±8a | 112±10b | 8±1c | 102±9a | 251±18a | 19±2b | |
| 22 °C | KA-Y1 | 168±11b | 111±12b | 36±2c | 158±13a | 11±1b | 96±7a,b | 249±25a | 23±3a |
| KE-Y1 | 34±3e | 127±11a | 61±4b | 86±9c | 14±2a | 84±8b,c | 231±16a | 15±2c | |
| BO-Y1 | 62±4d | 31±2d | 35±4c | 64±4d,e | 2±1d | 50±5d | 104±11d | 6±1e |
Yeast strain codes: KA-Y1, S. cerevisiae strain isolated from “Karnachalas” grapes; KE-Y1, S. cerevisiae strain isolated from “Keratsouda” grapes; BO-Y1, S. cerevisiae strain isolated from “Bogialamas” grapes. Data shown are mean values of three independent fermentations ± Standard Deviation. Different lowercase letters within a column indicate significant differences among values (2-way ANOVA: Duncan’s multiple range test, P < 0.05). 3.4.2. Minor Volatiles.
Table 4.
Μinor volatiles of wines fermented by indigenous S. cerevisiae strains at 12 & 22 °C.
|
Fermentation temperature |
Yeast strain |
Esters content (mg L-1) |
Alcohols content (mg L-1) |
Organic acids content (mg L-1) |
Carbonyl compounds content (mg L-1) |
Miscellaneous compounds content (mg L-1) |
Total volatiles (mg L-1) |
| 12 °C | KA-Y1 | 82.3b | 35.8c | 0.7c | 0.1a | 5.1a,b | 124.0b |
| KE-Y1 | 72.1b | 42.1b,c | 0.6c | <0.1b | 2.8d | 117.6b | |
| BO-Y1 | 39.2c | 12.8e | 2.1a | nd | 2.4d | 56.5c | |
| 22 °C | KA-Y1 | 106.0a | 43.7b | 0.5c | nd | 4.1c | 154.4a |
| KE-Y1 | 114.9a | 26.7d | 1.8b | nd | 4.5b,c | 147.9a | |
| BO-Y1 | 99.0a | 54.5a | 1.7b | nd | 5.4a | 160.6a |
Yeast strain codes: KA-Y1, S. cerevisiae strain isolated from “Karnachalas” grapes; KE-Y1, S. cerevisiae strain isolated from “Keratsouda” grapes; BO-Y1, S. cerevisiae strain isolated from “Bogialamas” grapes. Data shown are mean values of three independent fermentations ± Standard Deviation (standard deviation was ±5% for all values). Different lowercase letters within a column indicate significant differences among values (2-way ANOVA: Duncan’s multiple range test, P < 0.05). nd: not detected.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



