Submitted:
27 July 2025
Posted:
28 July 2025
You are already at the latest version
Abstract
Food enzymes are essential to modern food processing, with many produced industrially through microbial fermentation. However, bacterial production strains can generate secondary metabolites, including trace amounts of antibiotics. Subtilisin, a widely used protease, is commonly produced using Bacillus paralicheniformis, a strain known to naturally synthesize bacitracin. This has raised concerns by the European Food Safety Authority (EFSA) regarding the potential contribution of subtilisin preparations to antimicrobial resistance (AMR).This review evaluates published evidence and quantitative data to assess whether to assess whether trace levels in subtilisin preparations pose an AMR risk. First, analytical data indicate that any residual bacitracin in the final enzyme preparation is extremely low and further diluted during food processing, resulting in dietary exposure well below that from permitted sources such as meat and dairy. Second, these concentrations fall far below established thresholds for selective pressure. Third, there is no robust evidence of cross-resistance to medically important antibiotics or of transferable resistance elements associated with bacitracin. Finally, bacitracin resistance genes are already widespread in the environment, including in drinking water, further diminishing the relative impact of trace exposures from food enzymes. Taken together, the evidence does not support the view that trace bacitracin in subtilisin poses a meaningful risk for the development or spread of AMR.
Keywords:
subtilisin
; bacitracin
; antimicrobial resistance
; EFSA
; Bacillus paralicheniformis
; food safety
Bacitracin is a polypeptide antibiotic composed of a mixture of structurally related cyclic peptides [1]. These peptides act primarily against Gram-positive bacteria by disrupting cell wall and peptidoglycan synthesis. Specifically, bacitracin inhibits bacterial cell wall formation by blocking the dephosphorylation of C55-isoprenyl pyrophosphate (also known as bactoprenol pyrophosphate), a lipid carrier responsible for transporting peptidoglycan precursors across the inner membrane [2]. Owing to its antimicrobial properties, bacitracin is widely used in consumer products such as ophthalmic and topical ointments, as well as in cosmetics, for the treatment of minor skin infections.
Subtilisin is a serine protease commonly produced by Bacillus species and widely used as a food enzyme for protein hydrolysis [3]. When subtilisin is produced using Bacillus paralicheniformis, trace amounts of bacitracin may be co-produced. This has prompted the European Food Safety Authority (EFSA) to evaluate whether the presence of bacitracin in such enzyme preparations poses a significant antimicrobial resistance (AMR) risk.
Bacillus paralicheniformis was formally recognized in 2015 as a species distinct from B. licheniformis, based on its ability to produce secondary metabolites such as bacitracin, fengycin, and a lantipeptide [4,5]. As a result, EFSA requires that subtilisin preparations derived from B. paralicheniformis, within the broader Bacillus subtilis group, undergo additional safety qualifications, including confirmation via whole-genome sequencing (WGS) that the production strain does not synthesize bacitracin.
EFSA has issued several scientific opinions concerning the safety of subtilisin produced by Bacillus paralicheniformis [6,7,8]. In one assessment involving strain AP-01 (Nagase [Europa] GmbH), bacitracin was not detected in any of the tested enzyme batches. However, the EFSA
panel considered the limit of detection too high to definitively rule out the presence of bacitracin at concentrations potentially relevant to the development of AMR, particularly due to concerns about cross-resistance to colistin, a highest priority critically important antimicrobial [8].
A similar rationale was applied in the evaluation of subtilisin produced by strain DP-Dzx96 (Genencor), in which bacitracin was detected in the enzyme preparation. As a result, EFSA determined that the product could not be considered safe for use in food processing [6].
Likewise, in a third case involving strain LMG S-30155 (ENMEX), EFSA identified genes involved in bacitracin biosynthesis and confirmed the presence of bacitracin in the enzyme preparation, leading to the same safety concern and conclusion [7].
Notably, this precautionary approach was consistently applied across all three cases, regardless of whether bacitracin was detected analytically or inferred from genomic data. However, this position warrants further scrutiny, as it appears to conflict with a substantial body of scientific evidence.
In the following section, I review the available evidence and data to assess whether trace levels of bacitracin in subtilisin preparations pose a meaningful AMR risk. This is supported by the following three key arguments:
1. Extremely Low Exposure Levels and Regulatory Comparisons
Subtilisin is not consumed directly but is used as a processing aid in food production. EFSA estimated dietary exposure to the enzyme based on Total Organic Solids (TOS) intake among high-consuming population groups. Although subtilisin preparations may contain trace amounts of bacitracin, the enzyme undergoes dilution by several orders of magnitude during food manufacturing. Notably, EFSA’s risk assessment does not appear to have fully accounted for this dilution factor.
As a result, the levels of bacitracin potentially entering the food chain are substantially lower than the maximum residue limits established by the Joint FAO/WHO Expert Committee on Food Additives (JECFA) [9] and by EU Regulation 37/2010 on pharmacologically active substances in foodstuffs of animal origin [10]. These international standards permit bacitracin residues in foods such as milk, meat, and eggs at concentrations significantly higher than those that could arise from the use of subtilisin in food processing. (See Table 1 for a comparison of bacitracin concentrations across regulatory thresholds and environmental sources).
2. Trace-Level Concentrations Are Below Selection Thresholds
The minimum inhibitory concentration (MIC) of bacitracin is approximately 2000 ng/ml [11]. In contrast, EFSA cites a 'Minimal Selective Concentration' (MSC) of 8 ng/mL, derived from a theoretical model that is not specific to bacitracin. This value lacks practical relevance for food safety assessments, as residual bacitracin levels in the final subtilisin preparations are at least 4000-fold lower than this MSC.
Moreover, polypeptide antibiotics like bacitracin are unlikely to induce secondary resistance when present at subinhibitory concentrations [12]. Consequently, the development of AMR under these conditions is highly improbable.
3. Lack of Evidence for Cross-Resistance or Transferability
EFSA has expressed concern that bacitracin may contribute to cross-resistance to colistin, referencing the emergence of plasmid-mediated colistin resistance (mcr-1) in Enterobacteriaceae [13]. However, the cited study reported mcr-1 in the context of general antibiotic use in livestock and did not identify bacitracin as a selective agent. The high-dose therapeutic use of multiple antibiotics in animal production is not comparable to the trace bacitracin levels present in food enzyme preparations, making selection for mcr-1 under these conditions highly improbable.
Over more than five decades of use as a feed additive and growth promoter, bacitracin has not been linked to increased resistance in clinically relevant pathogens such as Clostridium perfringens, staphylococci, or streptococci [14]. Furthermore, multiple studies have found no evidence of cross-resistance between bacitracin and medically important antibiotics used in human therapy [15,16,17,18,19]. Sub-therapeutic exposure to bacitracin has also been shown not to increase the frequency of multidrug resistance [20,21].
In addition, current data suggest that bacitracin resistance is not readily transferable via plasmids or other mobile genetic elements. Several investigations have failed to demonstrate such transferability [19,22], and some studies even indicate that bacitracin may inhibit plasmid transfer, thereby reducing the incidence of resistance to unrelated antibiotics [23,24].
Additional Considerations
Bacitracin also confers several secondary benefits, including reduced gut inflammation, enhanced absorption of vitamins and nutrients, inhibition of toxin production, increased phagocytic activity, and improved tolerance to physiological stress. Its low toxicity has supported its use in clinical settings, such as in skin grafting procedures [25], as well as in oral treatments for colitis and Clostridium difficile-associated diarrhea [26].
Further supporting the evidence outlined above, bacitracin occurs naturally in various environmental compartments, including groundwater, surface water, and soil—often at concentrations significantly higher than those found in subtilisin preparations [27,28]. Notably, no adverse health effects have been associated with such environmental exposure. This broader context reinforces the conclusion that trace levels of bacitracin in food enzyme preparations are unlikely to pose a risk to public health.
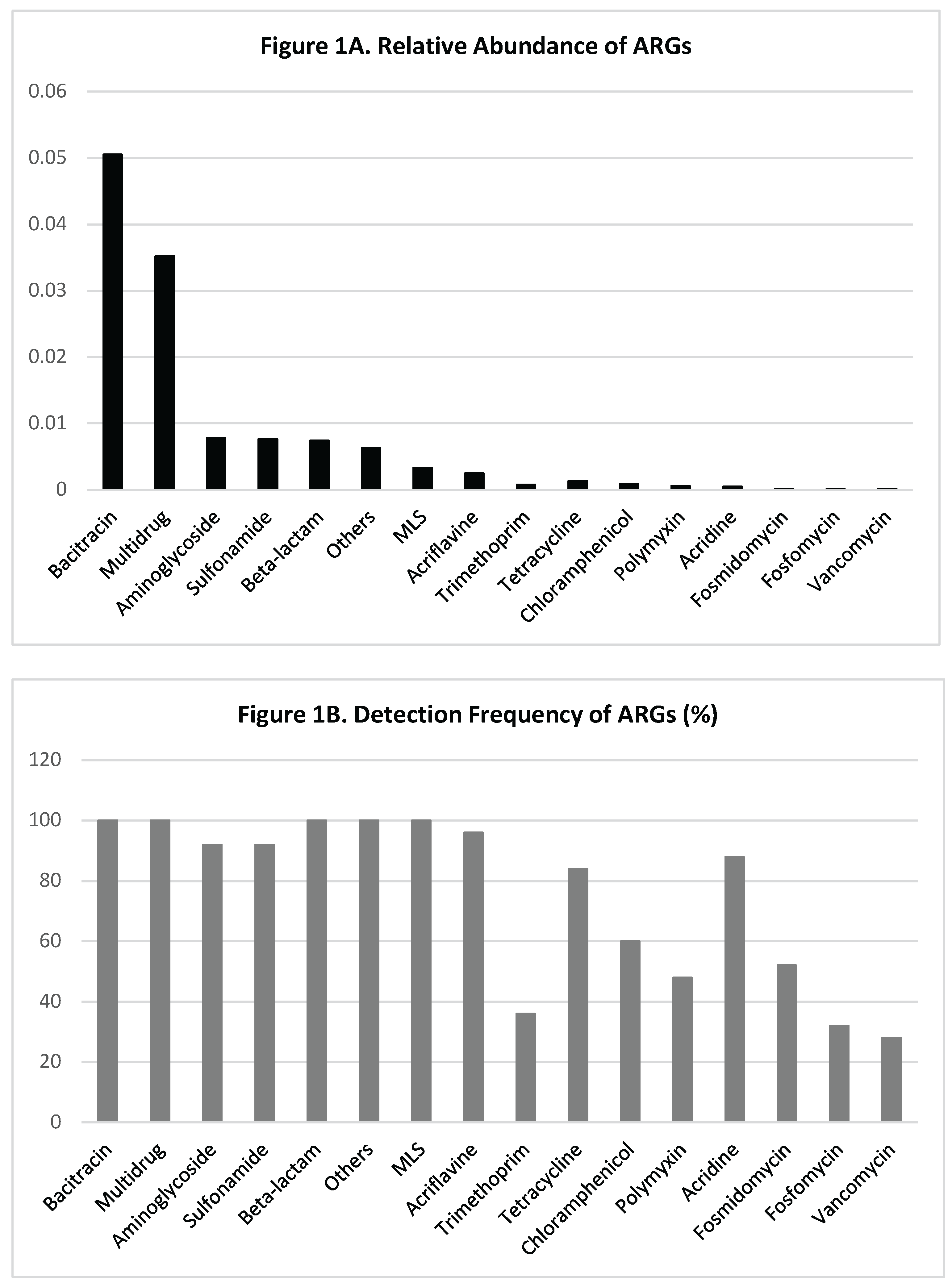
As illustrated in Figure 1, data on the abundance of bacitracin resistance genes (ARGs) in drinking water samples confirm their frequent detection without any documented association with increased AMR risk [27,28].
In summary, this manuscript demonstrates that:
- The levels of bacitracin present in subtilisin preparations do not raise safety concerns.
- Antimicrobial resistance (AMR) cannot develop at bacitracin concentrations below its minimum inhibitory concentration (MIC); levels in subtilisin are nearly 60-fold below the MIC and approximately 4000-fold below the minimal selective concentration (MSC) referenced by EFSA.
- Bacitracin does not induce cross-resistance with human medicinal antibiotics or other marketed antimicrobials.
- The transferability of bacitracin resistance via plasmids or other mobile genetic elements is unlikely.
- Environmental concentrations of bacitracin in groundwater and soil are significantly higher than those found in subtilisin preparations.
- Acceptable residue limits for bacitracin in meat, eggs, and milk, as established by regulatory bodies, are far higher than the trace levels present in food processed with subtilisin.
- Therefore, the likelihood that the use of subtilisin in food processing contributes to AMR development is considerably lower than other existing exposure pathways.

Table 1.
Comparative Bacitracin Levels in Food and Environmental Sources vs. Relevant Thresholds.
| Source or Threshold | Bacitracin Concentration (ng/mL or ng/g) |
Reference / Note |
|---|---|---|
| Subtilisin-processed food (estimated) | ≤ 0.002 | 4000× below MSC; 60× below MIC |
| Minimal Selective Concentration (MSC) | 8 | Hypothetical threshold (EFSA) |
| Minimum Inhibitory Concentration (MIC) | 2000 | Effective bacterial inhibition [11] |
| EU MRL in milk (‡) | 50,400 | 1.2 IU/mL × 42 (†)[9,10] |
| EU MRL in meat | 29,400 | 0.7 IU/g × 42 [9,10] |
| EU MRL in eggs | 201,600 | 4.8 IU/g × 42 [9,10] |
| Groundwater/Drinking water (natural presence) | Up to 10 | Detected environmental levels [27,28] |
| Animal feed (therapeutic use) | 100,000–200,000 | Used as additive [14] |
‡ MRL (Maximum Residue Limit): the highest legally permitted concentration of a veterinary drug residue in food, established to ensure consumer safety. † Conversion based on JECFA: 1 IU of bacitracin = 42 ng.
Figure 1.
Abundance and Detection Frequency of Antibiotic Resistance Genes (ARGs) in Drinking Water Samples. Top (A): Average relative abundance of ARG types, expressed as copies per average bacterial cell (capc). Bottom (B): Detection frequency of each ARG type across 25 drinking water samples. Bacitracin resistance genes are the most abundant and universally detected, while other ARGs vary in both abundance and prevalence. Data adapted from [28], with modifications to the presentation and graph style.
Figure 1.
Abundance and Detection Frequency of Antibiotic Resistance Genes (ARGs) in Drinking Water Samples. Top (A): Average relative abundance of ARG types, expressed as copies per average bacterial cell (capc). Bottom (B): Detection frequency of each ARG type across 25 drinking water samples. Bacitracin resistance genes are the most abundant and universally detected, while other ARGs vary in both abundance and prevalence. Data adapted from [28], with modifications to the presentation and graph style.
Acknowledgments
The author gratefully acknowledges the use of artificial intelligence (ChatGPT, OpenAI) to improve the language, readability, and clarity of the manuscript. All content, ideas, interpretations, and conclusions are the author’s own.
References
- Buijs, N.P.; Vlaming, H.C.; Kotsogianni, I.; Arts, M.; Willemse, J.; Duan, Y.; Alexander, F.M.; Cochrane, S.A.; Schneider, T.; Martin, N.I. A Classic Antibiotic Reimagined: Rationally Designed Bacitracin Variants Exhibit Potent Activity against Vancomycin-Resistant Pathogens. Proc Natl Acad Sci U S A 2024, 121, e2315310121. [Google Scholar] [CrossRef] [PubMed]
- Stone, K.J.; Strominger, J.L. Mechanism of Action of Bacitracin: Complexation with Metal Ion and C55-Isoprenyl Pyrophosphate. Proc Natl Acad Sci U S A 1971, 68, 3223. [Google Scholar] [CrossRef] [PubMed]
- Khan, Z.; Shafique, M.; Saleem, F.; Azim, M.K.; Jabeen, N.; Naz, S.A. Characterization of the Genome and Serine Protease of a Novel Bacillus Subtilis Isolate. Antonie van Leeuwenhoek, International Journal of General and Molecular Microbiology 2022, 115, 281–295. [Google Scholar] [CrossRef] [PubMed]
- Barat Baviera, J.M.; Bolognesi, C.; Chesson, A.; Cocconcelli, P.S.; Crebelli, R.; Gott, D.M.; Grob, K.; Lambré, C.; Lampi, E.; Mengelers, M.; et al. Taxonomic Identity of the Bacillus Licheniformis Strains Used to Produce Food Enzymes Evaluated in Published EFSA Opinions. EFSA Journal 2024, 22, e8770. [Google Scholar] [CrossRef] [PubMed]
- Dunlap, C.A.; Kwon, S.W.; Rooney, A.P.; Kim, S.J. Bacillus Paralicheniformis Sp. Nov., Isolated from Fermented Soybean Paste. Int J Syst Evol Microbiol 2015, 65, 3487–3492. [Google Scholar] [CrossRef] [PubMed]
- Lambré, C.; Barat Baviera, J.M.; Bolognesi, C.; Cocconcelli, P.S.; Crebelli, R.; Gott, D.M.; Grob, K.; Lampi, E.; Mengelers, M.; Mortensen, A.; et al. Safety Evaluation of the Food Enzyme Subtilisin from the Non-genetically Modified Bacillus Paralicheniformis Strain DP-Dzx96. EFSA Journal 2023, 21, e08155. [Google Scholar] [CrossRef] [PubMed]
- Lambré, C.; Barat Baviera, J.M.; Bolognesi, C.; Cocconcelli, P.S.; Crebelli, R.; Gott, D.M.; Grob, K.; Lampi, E.; Mengelers, M.; Mortensen, A.; et al. Safety Evaluation of the Food Enzyme Subtilisin from the Non-genetically Modified Bacillus Paralicheniformis Strain LMG S-30155. EFSA Journal 2023, 21, e07910. [Google Scholar] [CrossRef] [PubMed]
- Lambré, C.; Barat Baviera, J.M.; Bolognesi, C.; Cocconcelli, P.S.; Crebelli, R.; Gott, D.M.; Grob, K.; Lampi, E.; Mengelers, M.; Mortensen, A.; et al. Safety Evaluation of the Food Enzyme Subtilisin from the Non-genetically Modified Bacillus Paralicheniformis Strain AP-01. EFSA Journal 2024, 22, e8873. [Google Scholar] [CrossRef] [PubMed]
- Joint FAO/WHO Expert Committee on Food Additives (JECFA) Bacitracin: JECFA Evaluation Summary – Food Safety and Quality; 1968.
- European Commission EU Regulation 37/2010 on Pharmacologically Active Substances and Their Classification Regarding Maximum Residue Limits in Foodstuffs of Animal Origin; Brussels, 2010.
- Gu, Y.; Song, S.; Zhu, Q.; Jiao, R.; Lin, X.; Yang, F.; Veen, S. van der Bacitracin Enhances Ceftriaxone Susceptibility of the High-Level Ceftriaxone-Resistant Gonococcal FC428 Clone. Microbiol Spectr 2023, 11, e02449-23. [Google Scholar] [CrossRef] [PubMed]
- Axelsen, P.H. A Chaotic Pore Model of Polypeptide Antibiotic Action. Biophys J 2008, 94, 1549. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.Y.; Wang, Y.; Walsh, T.R.; Yi, L.X.; Zhang, R.; Spencer, J.; Doi, Y.; Tian, G.; Dong, B.; Huang, X.; et al. Emergence of Plasmid-Mediated Colistin Resistance Mechanism MCR-1 in Animals and Human Beings in China: A Microbiological and Molecular Biological Study. Lancet Infect Dis 2016, 16, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Phillips, I. The Use of Bacitracin as a Growth Promoter in Animals Produces No Risk to Human Health. Journal of Antimicrobial Chemotherapy 1999, 44, 725–728. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.N.; Li, Q.; Kohut, B.; Biedenbach, D.J.; Bell, J.; Turnidge, J.D. Contemporary Antimicrobial Activity of Triple Antibiotic Ointment: A Multiphased Study of Recent Clinical Isolates in the United States and Australia. Diagn Microbiol Infect Dis 2006, 54, 63–71. [Google Scholar] [CrossRef] [PubMed]
- A.L. Laboratories, Inc. Environmental Assessment for BMD® 514.1(b)10. Supplement to 21 CFR 558.76.; 1989.
- Alpharma AS, A.A.H.D. Bacitracin Zinc – New Evidence: Resistance and Safety to Human Health. Internal Report 1999.
- Alpharma Animal Health Division Public Health Impact of the Use of Bacitracin Zinc in Animals. Scientific Dossier for the Re-Examination of the Decision in Reg. (EC) 2821/98 Regarding the Use of Zinc Bacitracin in Animal Feeds. Internal Report 2000.
- Mathers, J. Bacitracin — Natural Peptide with Minimal Resistance Issues: A Comprehensive Overview Including Its Efficacy Against Clostridium Perfringens. In Proceedings of the Proceedings of Alpharma’s Swine Enteric Health Symposium; Alpharma Animal Health, 2008.
- Thibodeau, A.; Quessy, S.; Guévremont, E.; Houde, A.; Topp, E.; Diarra, M.S.; Letellier, A. Antibiotic Resistance in Escherichia Coli and Enterococcus Spp. Isolates from Commercial Broiler Chickens Receiving Growth-Promoting Doses of Bacitracin or Virginiamycin. Canadian Journal of Veterinary Research 2008, 72, 129. [Google Scholar] [PubMed]
- Walton, J.R.; Laerdal, O.A. The Effect of Dietary Zinc Bacitracin on the Resistance Status of Porcine Strains of Escherichia Coli. In Proceedings of the Proceedings of the 1980 International Pig Veterinary Society Congress; International Pig Veterinary Society , 1980.
- Helmuth, R.; Bulling, E. Proceedings of the Symposium “Criteria and Methods for the Microbiological Evaluation of Growth Promotors in Animal Feeds.” In Proceedings of the VetMed-Hefte; 1989.
- Walton, J.R. The Effect of Dietary Zinc Bacitracin on the Resistance Status of Intestinal Escherichia Coli and Enterococci from Broiler Chickens. Zentralblatt für Veterinärmedizin Reihe B 1984, 31, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Mathers, J.J. Individual and Combined in Vitro Effects of Bacitracin and Chlortetracycline in Swine-Derived Clostridium Spp. In Proceedings of the Abstracts of the American Society for Microbiology Annual Meeting (ASM Ann Meet); American Society for Microbiology, 2004; p. “Poster A-150”.
- Nguyen, R.; Khanna, N.R.; Safadi, A.O.; Patel, P.; Sun, Y. Bacitracin Topical. StatPearls 2024. [Google Scholar]
- Chang, T.-W.; Gorbach, S.L.; Bartlett, J.G.; Saginur, R. Bacitracin Treatment of Antibiotic-Associated Colitis and Diarrhea Caused by Clostridium Dilficile Toxin. Gastroenterology 1980, 78, 1584–1586. [Google Scholar] [CrossRef] [PubMed]
- Fu, C.; Xu, B.; Chen, H.; Zhao, X.; Li, G.; Zheng, Y.; Qiu, W.; Zheng, C.; Duan, L.; Wang, W. Occurrence and Distribution of Antibiotics in Groundwater, Surface Water, and Sediment in Xiong’an New Area, China, and Their Relationship with Antibiotic Resistance Genes. Science of The Total Environment 2022, 807, 151011. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Li, B.; Jiang, X.T.; Wang, Y.L.; Xia, Y.; Li, A.D.; Zhang, T. Catalogue of Antibiotic Resistome and Host-Tracking in Drinking Water Deciphered by a Large Scale Survey. Microbiome 2017, 5, 154. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



