
Submitted:
24 July 2025
Posted:
24 July 2025
You are already at the latest version
Abstract
To prepare a boosted sterile insect technique (SIT) field trial in Saint-Joseph, Reunion island, we compared the attractiveness of two adult traps for Aedes albopictus and Aedes aegypti. In addition, we explored the co-occurrence of these species in their usual larval habitats. Two traps were compared with two conditions each using Latin square design: BG Sentinel trap baited with carbon dioxide (CO2) with/without addition of BG Lure and ovi-sticky trap with/without hay. Ovi-sticky were inefficient. In both Aedes species, CO2-baited traps were equally effective for catching females when baited or not with the lure. In contrast, they were more attractive for males than for females with the lure. Aedes aegypti larvae were found in 4 of 6 vacoas (Pandanus utilis), and 1 of 4 anthropic breeding sites. In vacoas, the densities of Aedes albopictus and Aedes aegypti larvae were negatively correlated, whereas the correlation was positive between chironomids and Aedes aegypti. The abundance of adults and larvae varied with weather conditions. Finally, CO2-baited traps will be used without lure for the entomological monitoring to assess the effectiveness of the area-wide boosted SIT intervention, to reduce costs and logistics.
Keywords:
mosquito monitoring
; pyriproxyfen
; boosted sterile insect technique
; breeding site
; chironomids
1. Introduction
The repeated occurrence of arbovirus outbreaks in La Reunion highlights the need for effective tools to monitor Aedes mosquito populations. Two Aedes species considered arbovirus vectors are found in La Reunion: Aedes albopictus and Aedes aegypti [1]. Aedes albopictus is widely distributed in urban and rural areas, from coastal zones to elevations of up to 1,200 meters, and in both natural and artificial breeding sites. In contrast, Ae. aegypti populations are confined to a few natural environments, especially ravines in the south and west of the island. This limited distribution is attributed to ecological competition, as Ae. albopictus out-competes Ae. aegypti in urban areas [2]. Recurrent dengue outbreaks have been recorded in La Reunion in recent decades, with 29,577 confirmed cases and 33 deaths reported in 2021. The transmission of the dengue virus is now considered endemic [3,4]. In 2005-2006, during a large chikungunya epidemic, more than 300,000 cases were reported, with more than 200 deaths [5]. Furthermore, new cases of chikungunya were reported in August 2024, preceding a large outbreak of chikungunya during the austral summer of 2025‘ [6]. Although vaccines are now available against dengue and chikungunya viruses, mosquito control remains an important component of the prevention and mitigation of mosquito-borne infections. The Sterile Insect Technique (SIT) is a promising and environmentally friendly approach [7]. It involves the mass-rearing, sterilization, and release of male mosquitoes that compete with wild males to mating with wild females. Because most female mosquitoes only mate once in their life, less offspring are produced when SIT is implemented. A derived method, SIT boost, combines the release of sterile males with the application of a biocide to their body [8]. Treated males transfer the biocide to homospecific females during mating or other physical contacts. Subsequently, the contaminated females deposit the biocide in breeding sites, thereby preventing larval development and the emergence of adult mosquitoes. Cage trials showed that sterile males coated with pyriproxyfen can contaminate breeding sites directly, even in the absence of females [9]. Because the two species partially share the same breeding sites, both can be suppressed at the same time. This was observed during the pilot suppression trial conducted at the same site, where the release of sterile male Ae. aegypti treated with pyriproxyfen led to the suppression of populations of this species by more than 90%, as well as Ae. albopictus by approximately 60% [10].
In any case (SIT or boosted SIT), an accurate monitoring of targeted mosquito populations is essential to assess suppression and provide useful indicators for the control program, such as the sterile-to-wild male ratio. Following SIT and boosted SIT small area field trials, a phase III boosted SIT study has started in 2025 in Saint-Joseph (South of the island) with releases of sterile male Ae. albopictus treated with pyriproxyfen. Before starting the control trial, we implemented this study to refine the settings of the Biogents Sentinel traps commonly used to monitor adult Aedes populations in La Reunion [11], and to assess the possibility of using ovi-sticky traps, widely used in Singapore to monitor both Ae. aegypti and Ae. albopictus [12,13]. We also carried out a survey to assess the presence of larvae of these two species and better predict the likely impact of releasing sterile male Ae. albopictus treated with pyriproxyfen on Ae. aegypti populations.
2. Materials and Methods
2.1. Traps
BG-Sentinel traps (Biogents, Germany) consist of a collapsible, dark blue fabric container and a white lid with holes covering its opening. Air is sucked into the trap through a black catch pipe by an electrical fan, through a black netting acting as a cage. They were baited with carbon dioxide released from a bottle at 0.2 L / 24 h as previously described [11]. We evaluated whether the attractiveness of this basic setting improved when BG Lure was added, a commercial attractant made of pellets that emit an odor that mimics human sweat (https://sea.biogents.com/attractants/bg-lure-attractant/). Fresh pellets were left out for one week prior to their first use to avoid a repellent effect. Ovi-sticky taps are a small plastic black jar baited with tap water, with a sticky pad covering its inner side, thus catching any insect entering the trap and resting on the sticky paper. We evaluated whether the attractiveness of this basic setting improved when trap water was replaced with a hay infusion prepared with 50 g of hay infused in 5 liters of water for one week and diluting the resulting solution with tap water in a 1:5 ratio.
2.2. Design
The four trap settings described above were compared with the Latin square design [14]. Four traps with these settings were placed in four different positions and simultaneously rotated at each trapping occasion, so that each trap occupied each position once at the end of a replicate. The positional and time variations were therefore the same for each trap. Two protocols were used, each with a different set of two locations (Figure 1):
- Protocol 1: a ravine located in the eastern part of the study site, and an orchard located in its western part;
- Protocol 2: two close sets located in an urban area located in the northern part of the study site.
Trap rotations were performed every 24 hours for protocol 1. Based on the results and to optimize captures, they were changed to rotate ovi-stisky traps every week for Protocol 2. Each week, the BG Sentinel traps were placed for 48 hours only at each site, to accommodate their future use within the control program. At each rotation, the captured adult insects were poured into a plastic container and stored at -20°C to kill them. Then, they were poured into Petri dishes for identification using a binocular and a morphological key adapted to local mosquitoes. Individuals of Ae. aegypti and Ae. albopictus were counted by species and sex.
2.3. Larval Habitat
To assess the distribution of Aedes larvae across different habitat types, larval samples were collected from two different environments (i) leaves stipes in the canopy of vacoa trees (Pandanus utilis, an endemic shrub) or (ii) domestic containers in private gardens such as flower pots, rainwater collection tanks and pet bowls. The sampling sites were located at various distances from ravines or rivers where the populations of Ae. aegypti are considered restricted. The samples were collected during four field sessions, one in February (before the devastating cyclone Garance), once in March, and two in April 2025. These time points were spaced for at least 15 days to allow the development of new cohorts and to assess potential temporal variations in the distribution of species related to environmental or meteorological conditions. The larvae were collected with 20 ml Pasteur pipettes. Five leaves per vacoa were sampled by inserting the pipette in the leaf stripe, where rainwater accumulates. For domestic containers, larvae were collected from the surface of stagnant water. Once collected, the larvae were transferred to Petri dishes containing a small volume of water and frozen at -20°C until identification. The larvae in stage L3 were identified with a binocular and a key adapted to the local mosquito species [15].
2.4. Data Analysis
We made separate analyses for Ae. aegypti and Ae. albopictus, under the assumption of different drivers for their population dynamics, including different host patterns and contrasted breeding sites. We used a Bayesian mixed effect Poisson model of mosquito density to assess the effect of BG Lure as an additional attractant to CO2-baited traps, conditionally depending on the sex of the mosquitoes. This model has two fixed effects, coded as two main effects and their interaction, two dummy variables, coded as 0 (‘without lure’, or ‘female’) or 1 (‘with lure’, or ‘male’). In addition, an observation-level identifier was included as a random effect associated with the model intercept to account for the overdispersion in the data [16]. For observation i with count and expected mean , the linear predictor was:
with
where was the random effect with a Gamma prior (precision 0.0005), and , , and were the coefficients of the fixed effect. The full model was as follows:
with , i.e., a Poisson distribution with mean , and was the error.
The model coefficients were fitted using an integrated nested Laplace approximation of the Markov chain process, implemented in the INLA add-in package for R [17,18]. Then 10,000 simulated fitted values were drawn in the posterior distribution of the model coefficients and stored in a matrix M with 96 rows (2 locations * 4 traps * 4 replicates + 2 locations * 4 traps * 8 replicates) and 10,000 columns (one column for each simulated dataset). Its rows were aggregated by covariate pattern (observed combinations of ‘sex’ and ‘lure’). Estimates of and its credible lower and upper 95% limits were obtained taking the mean and quantiles 2.5% and 97.5% of the aggregated rows.
The matrix M was also used to estimate the relative densities and their 95% credible intervals:
where was the density in the reference pattern (e.g., average count of females in traps without lure CO2. The 95% credible intervals for were used to assess the statistical significance of the additional attractiveness (that is, higher density) associated using lure with CO2-baited traps (): should this interval cover 1.00, the null hypothesis of no additional attractiveness is accepted; otherwise, it is rejected.
In addition, the random effect was used to explore the variability of attractiveness according to the location of the trap, as well as the trend in time in the dynamics of the mosquito population.
To assess the joint distribution of larvae in vacoa samples, their count was modeled with a negative binomial regression with two qualitative covariates: ’species’ (three categories, reference = Ae. albopictus), and ’month’ (month when the sample was made, three categories February, March and April, reference = February). Firstly, we considered the two main effects and their interaction, then we tried to find a better submodel with a stepwise procedure based on the Akaike information criterion corrected for small samples (AICc) [19].
3. Results
3.1. Insect Catches
In total, 733 mosquitoes were caught during 96 trapping sessions from 2025-02-03 to 2025-04-29, including 447 Aedes albopictus, 187 Aedes aegypti, 66 unidentified Aedes spp. (apparent to one of the two previous species but too damaged to be identified), 26 Culex quinquefesciatus and 7 chironomids (Figure 2). Among the catches, only 3 mosquitoes (female Ae. albopictus) were caught in ovi-sticky traps. We concluded that this kind of ovi-sticky trap was not adapted for mosquito monitoring in La Reunion. For the remainder of the analysis, we focused on the evaluation of the effect of adding BG Lure to CO2-baited traps on their attractiveness.
3.2. Effect of Lure on CO2-Baited Traps Attractiveness
The results are shown in Figure 3 (1) Ae. albopictus and (2) Ae. aegypti). The same comments apply for both species, with similar numeric values for attractiveness:
- Without lure, the attractiveness of the CO2-baited trap was the same for the females and the males.
- With lure, the attractiveness of the CO2-baited trap was unchanged for the females. In contrast, it was 3 times higher for male than for female Ae. albopictus: relative attractiveness , 95% credible interval [1.6; 5.5], and 2 times higher for male than for female Ae. aegypti: [1.1; 3.5].
3.3. Variation in Space and Time
For the Protocol 1 (Figure 4a), the estimated random effect was lower in the ravine than in the orchard (t test, difference d = -1.0, t = -2.4, p = 0.02) and higher for Ae. aegypti than for Ae. albopictus (t test, d = 1, t = 2.4, p = 0.02). In both mosquito species, there was a sharp decrease in in 6 of 8 traps between the start and the end of the survey (one week, early February 2025). More generally, there were large variations in between and within the traps, for both species.
The random effect patterns for the data collected with Protocol 2 (Figure 5) were clearer than with Protocol 1, in relation to the longer time period, as well as different and less contrasted locations (Figure 1). On average, was not significantly different for Ae. albopictus and Ae. aegypti (t test: d=-0.1, t=-0.3, p=0.76). However, the variations between and within the trap in were smaller for Ae. albopictus than for Ae. aegypti. For the latter, sharp and positive peaks were observed on the same dates for 3 of 8 traps (in 2 different locations). In addition, appeared to decrease with time, with less variability for Ae. albopictus than for Ae. aegypti.
A decreasing linear-time trend was observed in the distribution of the estimated random effect of the density Poisson models of Ae. albopictus, strongly correlated with the minimum daily temperature (Figure 5c): bilateral Spearman correlation, , , .
The random effect pattern was very different for Ae. aegypti. It started close to 0, then it consistently increased during one month, that is, shortly after the occurrence of the Garance cyclone. Then, it started to decrease in parallel with the minimum daily temperature.
3.4. Larvae
The larvae were collected during 4 sampling sessions between 14 February and 22 April 2025. A total of 101 larvae were collected from 10 breeding sites (4 domestic containers and 6 vacoas): 72 Ae. albopictus, 6 Ae. aegypti and 23 chironomids. Aedes albopictus was found in 9 of 10 sites (5 of 6 in vacoas), Ae. aegypti in 5 of 10 sites (4 of 5 in vacoas), both were present together in 4 of 10 sites (3 of 6 in vacoas), and Ae. aegypti was found alone in 1 of 10 sites (1 of 6 vacoas). No Culex larvae were collected. All chironomids were found in vacoa breeding sites, as well as 5 of 6 Ae. aegypti larvae. Thus, Ae. albopictus was the highly predominant species in domestic containers. Furthermore, the number of collected larvae decreased with time: 54 in February (65% in vacoas, 33 samples), 37 in March (35% in vacoas, 33 samples) and 13 in April (85% in vacoas, 66 samples). Of the 10 sampling sites, Ae. albopictus was found at 5 sites (1 of 4 domestic containers, 4 of 6 vacoas). Aedes albopictus was detected at 4 of 5 sites where Ae. aegypti was found. Chrironomids were found in 4 of 6 vacoas. The number of larvae collected varied greatly among the samples. Up to 12 Ae. albopictus larvae were found in a sample, 2 Ae. aegypti larvae, and 5 chironomid larvae.
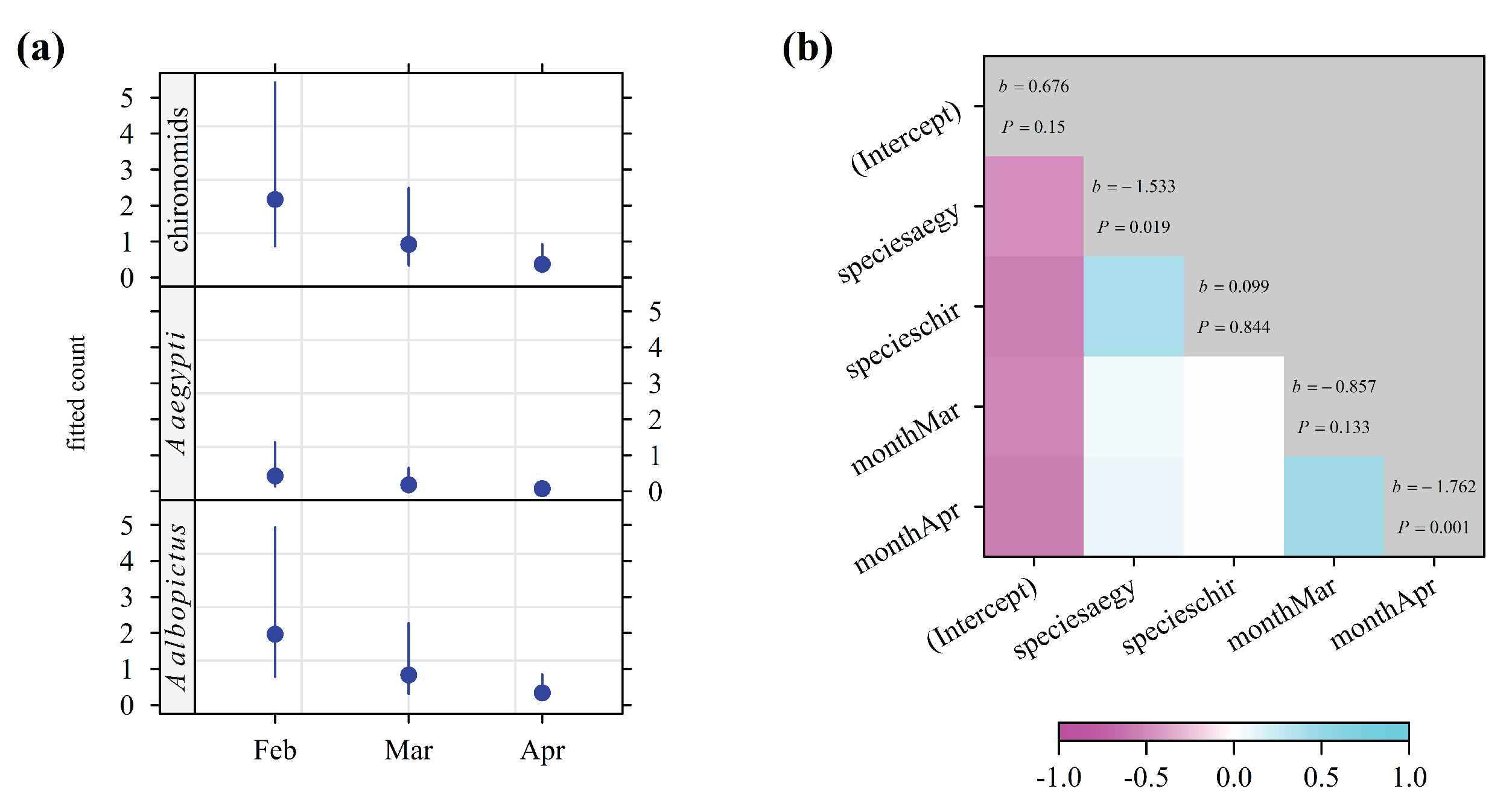
The results of the correlation analysis of the co-occurrence of larvae by insect species in the vacoas are shown in Figure 6. The model with the lowest AICc was the additive model. The density of Ae. albopictus larvae in vacoa samples was negatively correlated with the same index for chironomids and Ae. aegypti, in March and April. In contrast, the densities of chironomids and Ae. aegypti larvae were positively correlated, as well as the densities in March and April. The density of larvae in vacoa samples was lower for Ae. aegypti than for Ae. albopictus (P = 0.019), as well as in April compared to March (P = 0.001).
4. Discussion
The main objective of this study was to evaluate the efficiency of different trap types and baiting strategies for collecting Ae. albopictus and Ae. aegypti mosquitoes in various environments in Saint-Joseph. We compared BG traps baited with CO2 alone versusCO2 combined with BG-lure, and ovi-sticky traps filled with hay infusion or water. BG traps without any bait were excluded from the trial, as previous work conducted in the same area had demonstrated their low efficiency compared to baited traps [11,14]. The Latin square results provided evidence that the ovi-sticky traps were ineffective under local conditions, despite attempts to optimize their effectiveness. This contrasts with the results of Singapore, where stick traps with ovi have been used successfully for many years [12,13]. This can be explained by differences in management and vector control strategies between the two islands. In Singapore, the prevention and elimination of mosquito breeding sites is strictly regulated. Both private individuals and professionals must ensure that there are no potentially attractive breeding sites for mosquitoes, including the risk of being fired if a breeding site is discovered by competent bodies. However, in La Reunion, although the official vector control strategy promotes community involvement to prevent and eliminate mosquito breeding sites, many domestic breeding sites remain widespread and difficult to access, resulting in competition with ovi-sticky traps and reducing their attractiveness.
Adding BG-lure to CO2-baited traps did not improve their attractiveness for female mosquitoes, regardless of their species. This contrasts with other studies [20]. Such discrepancies could be explained by bio-ecological differences among mosquito populations or by environmental heterogeneity. Given the financial and logistic costs of using BG-Lure, and in the absence of strong statistical evidence, the best option for long-term monitoring implemented in the OpTIS project remains the use of BG traps baited with CO2 alone.
We observed considerable variability in trap performance depending on local weather and location within the study area. For example, before the Garance cyclone (27-28 February 2025), 169 adult mosquitoes were captured in the Langevin orchard, while fewer than 10 were collected indoors during heavy rains.
A secondary objective of this study was to investigate the local distribution of adult Ae. albopictus and Ae. aegypti and their breeding sites in Saint-Joseph. Identification of adult mosquitoes confirmed the predominance of Ae. albopictus, even in this part of the island. This is consistent with previous findings [21]. More surprisingly, the presence of Ae. aegypti was also documented in urban domestic environments. On some occasions, activity peaks were observed (Figure 4), when its density outnumbered Ae. albopictus density. These findings are of interest to public health, highlighting a possible strengthening effect of Ae. aegypti during outbreaks of arboviruses.
Rainfall patterns could also play a role in the relative abundance of Ae. aegypti and Ae. albopictus (Figure 6). The mosquito productivity of natural breeding sites of Ae. aegypti (e.g., vacoas) is likely to depend more on rainfall than on human activity, in contrast to the mosquito productivity of artificial containers for which Ae. albopictus is better adapted than Ae. aegypti in La Reunion. Larval collections further supported this assumption, revealing the low noise presence of Ae. aegypti in domestic breeding sites and confirming Ae. albopictus as the dominant species as described in previous studies [11,14]. Interestingly, in Vincendo vacoas, located close to a ravine, fewer larvae were collected than in the Langevin orchard, although they were 130 m from a river. More research is needed to clarify the ecological or microclimatic factors that underlie these spatial differences.
Non-target species such as chironomid larvae and adult Culex quinquefasciatus, a potential vector of Rift Valley fever and West Nile viruses, were also captured [22]. It is not surprising that Ae. aegypti and the chironomid larvae were positively correlated, since the Ae. aegypti strain from La Reunion is selvatic [1]. Although not central to this study, their presence provides valuable information for future environmental impact assessments related to the release of sterile males treated with pyriproxyfen as part of the OpTIS project.
The sudden drop in Ae. aegypti density after the Garance cyclone suggests that extreme weather events may significantly affect their populations, especially considering their typical larval habitats in ravines, which consist mainly of rock hollows (data not presented in this study) [1,23]. This was also observed in Mauritius during a pilot SIT trial [24].
The distribution data for Ae. Aegypti remain preliminary but warrant further investigation. Longitudinal studies covering different seasons and habitats (e.g., ravines vs. households) are needed to better understand the spatial dynamics of both Aedes species. Such data would allow for better tailoring of control measures to local ecological contexts. Furthermore, future research comparing the vector competence of Ae. aegypti and Ae. albopictus under the La Reunion conditions would be valuable [25].
Regarding the future control effort using boosted SIT using sterile male Ae. albopictus to carry pyriproxyfen, the fact that it was found in 4 of 5 breeding habitats where we recorded Ae. aegypti is promising regarding the cross-efficacy of this technology. Recently, we proposed to use the hetero-specific efficacy of the boosted SIT to prevent the invasion of new territories by invasive species.
This study has several limitations. First, the limited number of Latin square repetitions, which may restrict the generalization of the findings. A longer sampling period covering different seasons would provide more representative and robust data. Second, unmeasured factors, such as fine-scale meteorological fluctuations [26] or human activity patterns, could have influenced the efficacy of the trap and the presence of species.
5. Conclusions
This study highlighted the importance of considering local specificities when selecting trapping methods. BG traps baited with CO2 alone will be used for mosquito monitoring in the OpTIS project. The findings on the distribution of Ae. aegypti populations remain preliminary data. It would be both interesting and necessary to study their distribution not only near ravines but also within households, taking into account weather conditions during a longitudinal study that spans different seasons, and to compare it with the distribution of Ae. albopictus populations. In fact, by gaining a better understanding of the spatial distribution and seasonal dynamics of local target species, control strategies can be better adapted and therefore more effective.
Author Contributions
Conceptualization, C.B., T.B., R.L. and J.B.; methodology, C.B., T.B., R.L. and J.B.; software, R.L.; validation, J.B.; formal analysis, R.L. and J.B.; investigation, C.V., R.B., A.H., M.W., C. B. and J.B.; data curation, R.L. and J.B.; writing—original draft preparation, C.V., R.B., R.L. and J.B.; writing—review and editing, R.B., C.B., R.L. and J.B.; visualization, R.L.; supervision, T.B. and J.B.; project administration, J.B.; funding acquisition, T.B. and J.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research has been supported by the OpTIS project, co-financed through the FEDER program by the European Union and the Réunion Region. Europe is committed to La Réunion through the FEDER program.
Data Availability Statement
The original data and R code presented in the study are openly available in Figshare at https://figshare.com/s/3b71db503502dfca6fae, DOI 10.6084/m9.figshare.29616800.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| Ae. aegypti | Aedes aegypti |
| Ae. albopictus | Aedes albopictus |
| BG | Biogents, Germany |
| BGS | Biogents Sentinel, Germany |
| CO2 | carbon dioxide |
| DOAJ | Directory of open access journals |
| FEDER | Fonds Européen de Développement Régional |
| LD | Linear dichroism |
| MDPI | Multidisciplinary Digital Publishing Institute |
| OpTIS | Opérationalisation de la technique de l’insecte stérile |
| SIT | Sterile Insect Technique |
| TLA | Three letter acronym |
References
- Salvan, M.; Mouchet, J. Aedes albopictus et Aedes aegypti à l’Ile de La Réunion [Aedes albopictus and Aedes aegypti at Ile de la Réunion]. Ann. Soc. Belg. Med. Trop. 1994, 74, 323–326. [Google Scholar] [PubMed]
- Delatte, H.; Dehecq, J.S.; Thiria, J.; Domerg, C.; Paupy, C.; Fontenille, D. Geographic Distribution and Developmental Sites of Aedes albopictus (Diptera: Culicidae) During a Chikungunya Epidemic Event. Vector-Borne Zoonotic Dis. 2008, 8, 25–34. [Google Scholar] [CrossRef]
- Vincent, M.; Larrieu, S.; Vilain, P.; Etienne, A.; Solet, J.L.; François, C.; Roquebert, B.; Bandjee, M.C.J.; Filleul, L.; Menudier, L. From the threat to the large outbreak: dengue on Reunion Island, 2015 to 2018. Euro Surveill. 2019, 24. [Google Scholar] [CrossRef]
- Vincent, M.; Paty, M.C.; Gerardin, P.; Balleydier, E.; Etienne, A.; Daoudi, J.; Thouillot, F.; Jaffar-Bandjee, M.C.; Menudier, L. From dengue outbreaks to endemicity: Reunion Island, France, 2018 to 2021. Euro Surveill. 2023, 28. [Google Scholar] [CrossRef]
- Soumahoro, M.K.; Boelle, P.Y.; Gaüzere, B.A.; Atsou, K.; Pelat, C.; Lambert, B.; La Ruche, G.; Gastellu-Etchegorry, M.; Renault, P.; Sarazin, M.; et al. The Chikungunya epidemic on La Réunion Island in 2005-2006: a cost-of-illness study. PLoS Neglect.Trop. Dis. 2011, 5, e1197. [Google Scholar] [CrossRef]
- Frumence, E.; Piorkowski, G.; Traversier, N.; Amaral, R.; Vincent, M.; Mercier, A.; Ayhan, N.; Souply, L.; Pezzi, L.; Lier, C.; et al. Genomic insights into the re-emergence of chikungunya virus on Réunion Island, France, 2024 to 2025. Euro surveillance: bulletin Europeen sur les maladies transmissibles = European communicable disease bulletin 2025, 30. [Google Scholar] [CrossRef]
- Bouyer, J. Current status of the sterile insect technique for the suppression of mosquito populations on a global scale. Infectious Diseases of Poverty 2024, 13. [Google Scholar] [CrossRef] [PubMed]
- Bouyer, J.; Lefrançois, T. Boosting the sterile insect technique to control mosquitoes. Trends Parasitol. 2014, 30, 271–273. [Google Scholar] [CrossRef]
- Dupraz, M.; Lancelot, R.; Diouf, G.; Malfacini, M.; Marquereau, L.; Gouagna, L.C.; Rossignol, M.; Chandre, F.; Baldet, T.; Bouyer, J. Comparison of the standard and boosted sterile insect techniques for the suppression of Aedes albopictus populations under semi-field conditions. Parasites 2025. [Google Scholar] [CrossRef]
- Bouyer, J.; Almenar Gil, D.; Pla Mora, I.; Dalmau Sorli, V.; Maïga, H.; Mamai, W.; Claudel, I.; Brouazin, R.; Yamada, H.; Gouagna, L.C.; et al. Suppression of Aedes mosquito populations with the boosted sterile insect technique in tropical and Mediterranean urban areas. Scientific Reports 2025, 15. [Google Scholar] [CrossRef]
- Claudel, I.; Brouazin, R.; Lancelot, R.; Gouagna, L.C.; Dupraz, M.; Baldet, T.; Bouyer, J. Optimization of adult mosquito trap settings to monitor populations of Aedes and Culex mosquitoes, vectors of arboviruses in La Reunion. Sci. Rep. 2022, 12, 19544. [Google Scholar] [CrossRef]
- Ho, S.H.; Lim, J.T.; Ong, J.; Hapuarachchi, H.C.; Sim, S.; Ng, L.C. Singapore’s 5 decades of dengue prevention and control-Implications for global dengue control. PLOS Neglected Tropical Diseases 2023, 17, e0011400. [Google Scholar] [CrossRef] [PubMed]
- Bansal, S.; Lim, J.T.; Chong, C.S.; Dickens, B.; Ng, Y.; Deng, L.; Lee, C.; Tan, L.Y.; Kakani, E.G.; Yoong, Y.; et al. Effectiveness of Wolbachia-mediated sterility coupled with sterile insect technique to suppress adult Aedes aegypti populations in Singapore: a synthetic control study. The Lancet Planetary Health 2024, 8, e617–e628. [Google Scholar] [CrossRef] [PubMed]
- Brouazin, R.; Claudel, I.; Lancelot, R.; Dupuy, G.; Gouagna, L.C.; Dupraz, M.; Baldet, T.; Bouyer, J. Optimization of oviposition trap settings to monitor populations of Aedes mosquitoes, vectors of arboviruses in La Reunion. Sci. Rep. 2022, 12, 18450. [Google Scholar] [CrossRef] [PubMed]
- ARS Réunion. Clé d’identification des moustiques de La Réunion aux stades adulte et larvaires. Technical report, ARS Réunion, 2025.
- Harrison, X.A. Using observation-level random effects to model overdispersion in count data in ecology and evolution. PeerJ 2014, 2, e616. [Google Scholar] [CrossRef]
- Rue, H.; Riebler, A.I.; Sørbye, S.H.; Illian, J.B.; Simpson, D.P.; Lindgren, F.K. Bayesian computing with INLA: A review. Annual Reviews of Statistics and Its Applications 2017, 4, 395–421. [Google Scholar] [CrossRef]
- R Core Team. In R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2025.
- Burnham, K.; Anderson, D. Kullback-Leibler information as a basis for strong inference in ecological studies. Wildlife Research 2001, 28, 111–120. [Google Scholar] [CrossRef]
- Wilke, A.B.B.; Carvajal, A.; Medina, J.; Anderson, M.; Nieves, V.J.; Ramirez, M.; Vasquez, C.; Petrie, W.; Cardenas, G.; Beier, J.C. Assessment of the effectiveness of BG-Sentinel traps baited with CO2 and BG-Lure for the surveillance of vector mosquitoes in Miami-Dade County, Florida. PLOS ONE 2019, 14, e0212688. [Google Scholar] [CrossRef]
- Delatte, H.; Toty, C.; Boyer, S.; Bouetard, A.; Bastien, F.; Fontenille, D. Evidence of habitat structuring Aedes albopictus populations in Réunion Island. PLoS Negl. Trop Dis. 2013, 7, e2111. [Google Scholar] [CrossRef]
- Cardinale, E.; Bernard, C.; Lecollinet, S.; Rakotoharinome, V.; Ravaomanana, J.; Roger, M.; Olive, M.; Meenowa, D.; Jaumally, M.; Melanie, J.; et al. West Nile virus infection in horses, Indian Ocean. Comparative Immunology, Microbiology and Infectious Diseases 2017, 53, 45–49. [Google Scholar] [CrossRef]
- Bagny, L.; Delatte, H.; Quilici, S.; Fontenille, D. Progressive Decrease in Aedes aegypti Distribution in Reunion Island Since the 1900s. J. Med. Entomol. 2009, 46, 1541–1545. [Google Scholar] [CrossRef] [PubMed]
- Iyaloo, D.P.; Bouyer, J.; Facknath, S.; Bheecarry, A. Pilot Suppression trial of Aedes albopictus mosquitoes through an Integrated Vector Management strategy including the Sterile Insect Technique in Mauritius. bioRxiv 2020, 2020, https. [Google Scholar] [CrossRef]
- Gomard, Y.; Lebon, C.; Mavingui, P.; Atyame, C.M. Contrasted transmission efficiency of Zika virus strains by mosquito species Aedes aegypti, Aedes albopictus and Culex quinquefasciatus from Reunion Island. Parasite. Vector. 2020, 13. [Google Scholar] [CrossRef]
- Bidlingmayer, W.L. A comparison of trapping methods for adult mosquitoes: species response and environmental influence. Journal of medical entomology 1967, 4, 200–220. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Trap location set (i) for a field experiment with a Latin square design, and (ii) for a survey of larval habitat, both conducted in Saint-Joseph (La Reunion) from February to May 2025.
Figure 1.
Trap location set (i) for a field experiment with a Latin square design, and (ii) for a survey of larval habitat, both conducted in Saint-Joseph (La Reunion) from February to May 2025.
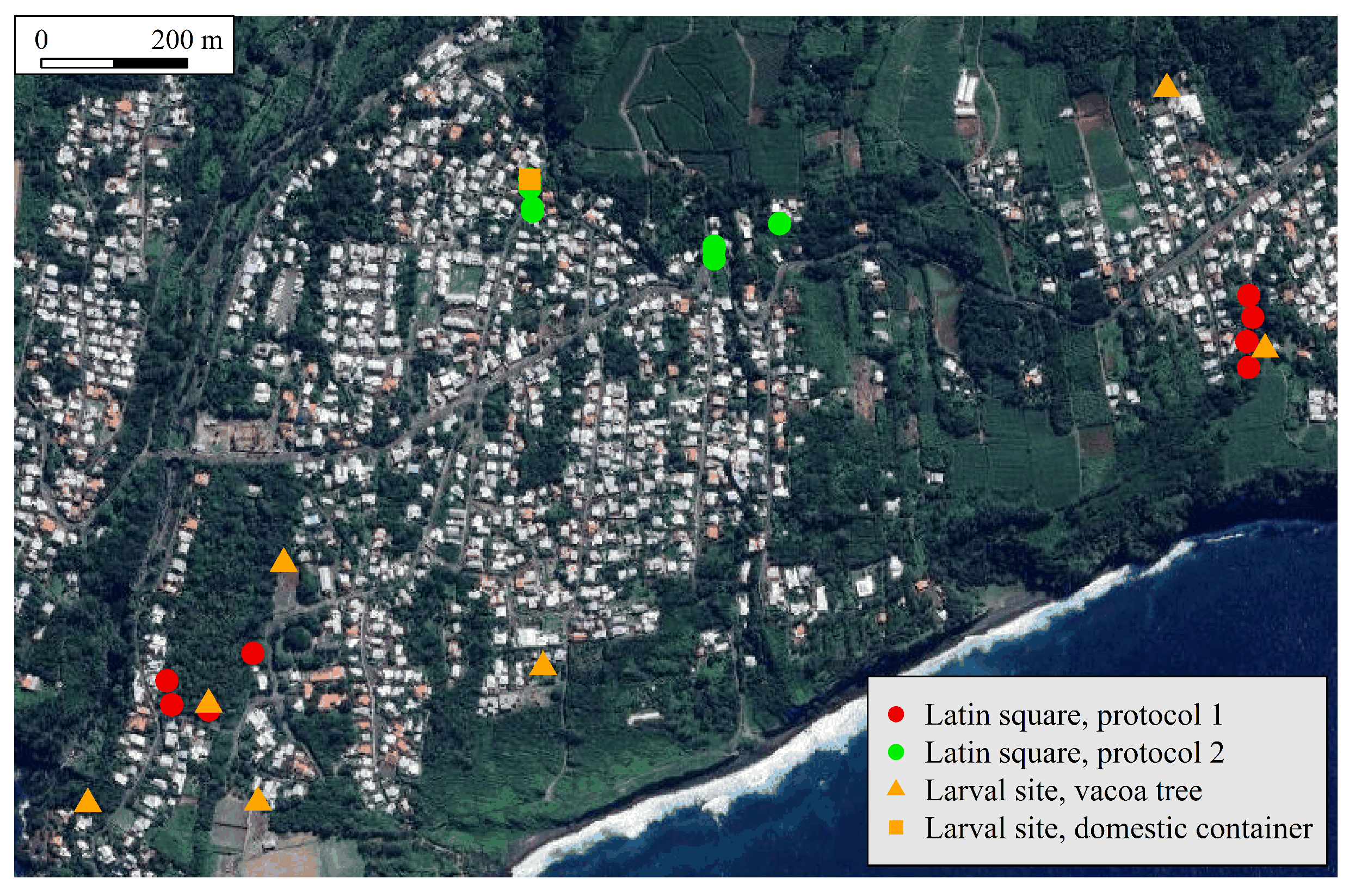
Figure 2.
Observed density (count/trap/d) for a selection of insects of interest collected during a field experiment implemented in Saint-Joseph (La Reunion) from February to May 2025. The violin plot in the background of each series of points is an estimate of the probability density of this series.
Figure 2.
Observed density (count/trap/d) for a selection of insects of interest collected during a field experiment implemented in Saint-Joseph (La Reunion) from February to May 2025. The violin plot in the background of each series of points is an estimate of the probability density of this series.
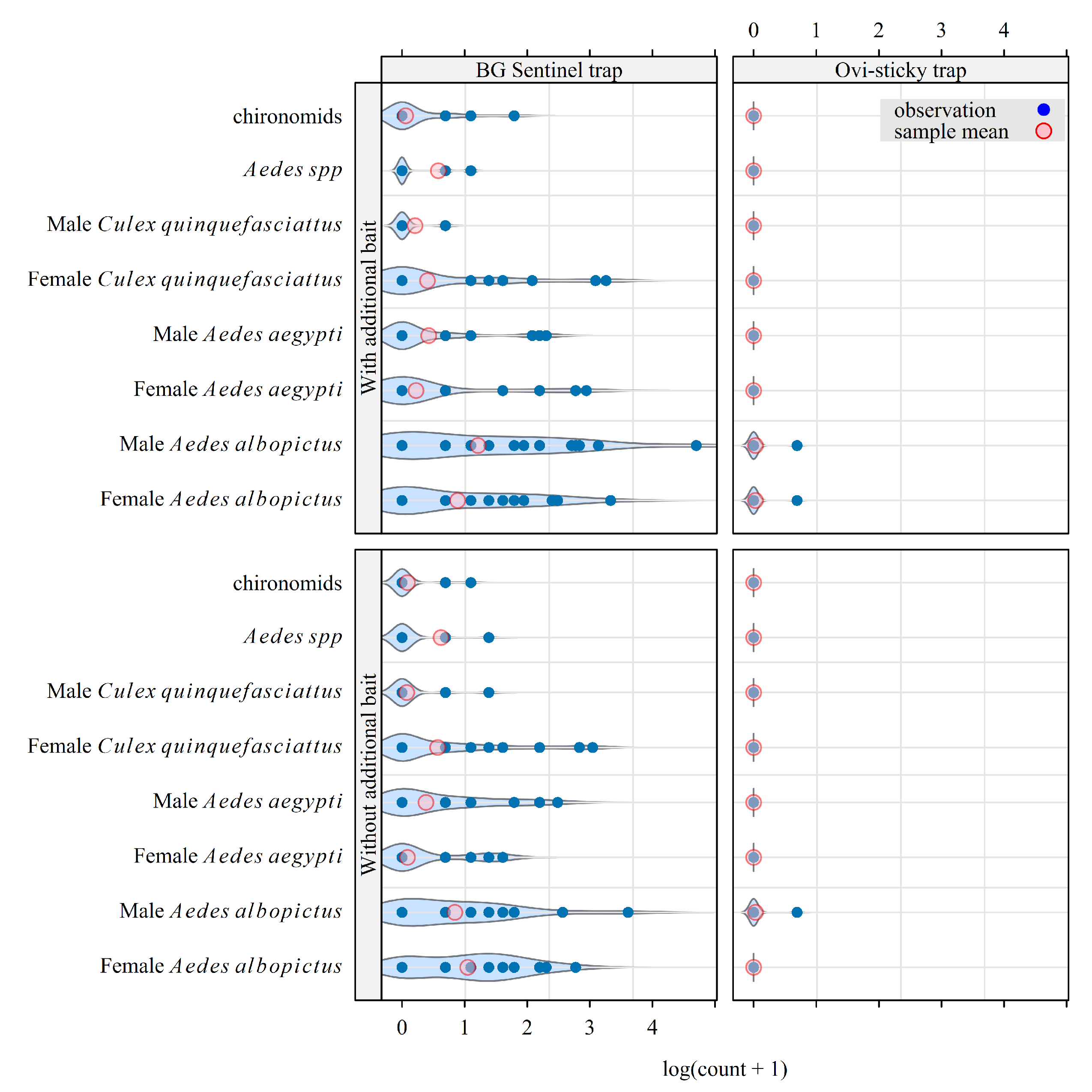
Figure 3.
Attractiveness and relative attractiveness of CO2-baited traps fitted with a Bayesian mixed-effect Poisson model for Ae. albopictus (plots a and b, left panel) and Ae. aegypti (plots a and b, right panel) in a field experiment implemented in Saint-Joseph (La Reunion) from February to May 2025. A Latin square design (two protocols, each with two sets of four traps in two locations four and eight replicates/traps in protocols 1 and 2, respectively) to evaluate the attractiveness gained with the BG lure. The violin plot in the background of each series of points is an estimate of the probability density of this series.
Figure 3.
Attractiveness and relative attractiveness of CO2-baited traps fitted with a Bayesian mixed-effect Poisson model for Ae. albopictus (plots a and b, left panel) and Ae. aegypti (plots a and b, right panel) in a field experiment implemented in Saint-Joseph (La Reunion) from February to May 2025. A Latin square design (two protocols, each with two sets of four traps in two locations four and eight replicates/traps in protocols 1 and 2, respectively) to evaluate the attractiveness gained with the BG lure. The violin plot in the background of each series of points is an estimate of the probability density of this series.
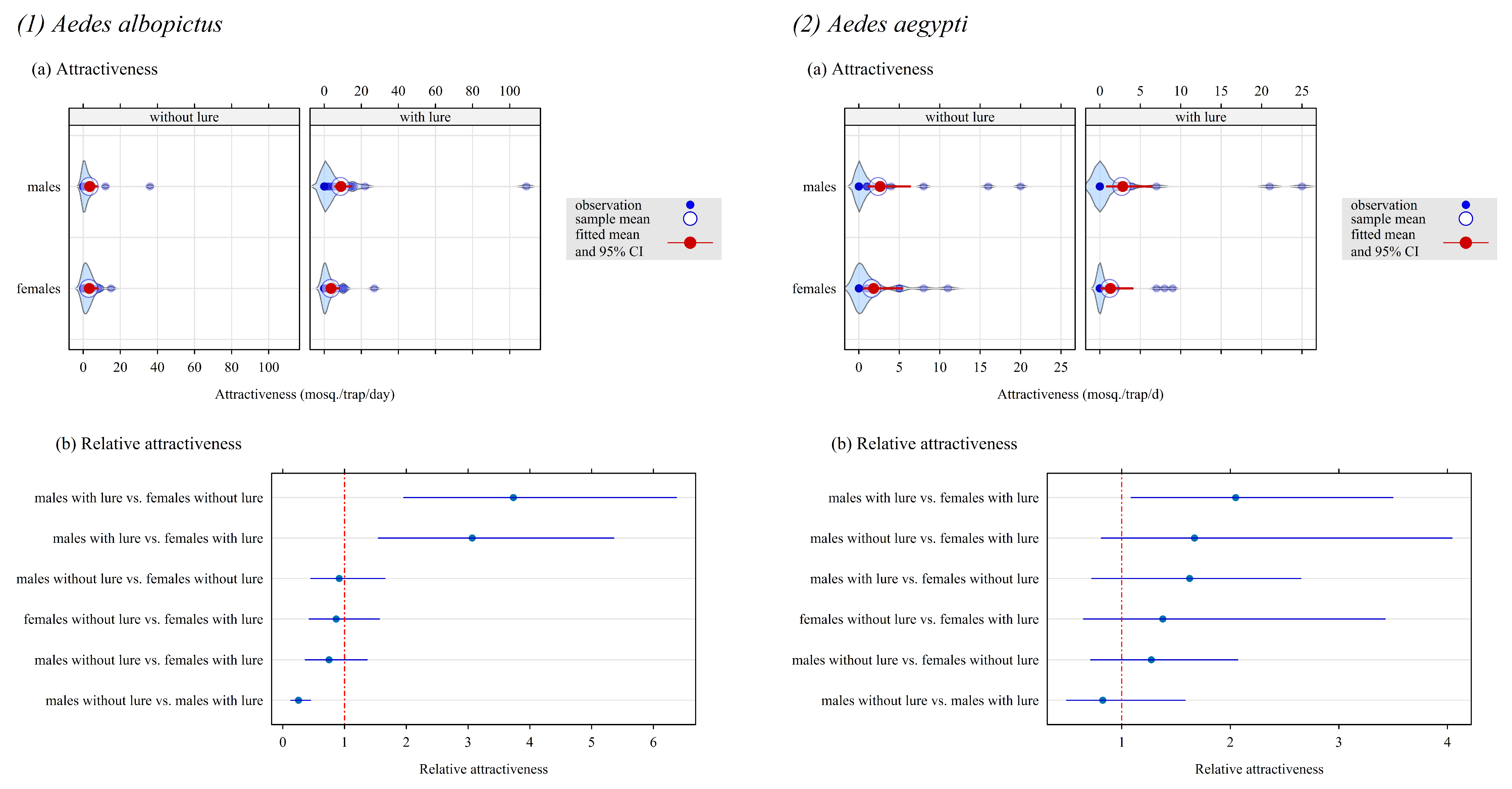
Figure 4.
Observation-level random effect estimated with a Bayesian mixed-effect Poisson model of trap attractiveness for Aedes aegypti and Aedes albopictus in a field experiment implemented in Saint-Joseph (La Reunion) from February to May 2025. Two distinct protocols were used, each with a Latin square design (a) protocol 1: two sets of four traps in two contrasted locations, four replicates and (b) protocol 2: two sets of four traps in two similar locations, eight replicates/trap.
Figure 4.
Observation-level random effect estimated with a Bayesian mixed-effect Poisson model of trap attractiveness for Aedes aegypti and Aedes albopictus in a field experiment implemented in Saint-Joseph (La Reunion) from February to May 2025. Two distinct protocols were used, each with a Latin square design (a) protocol 1: two sets of four traps in two contrasted locations, four replicates and (b) protocol 2: two sets of four traps in two similar locations, eight replicates/trap.
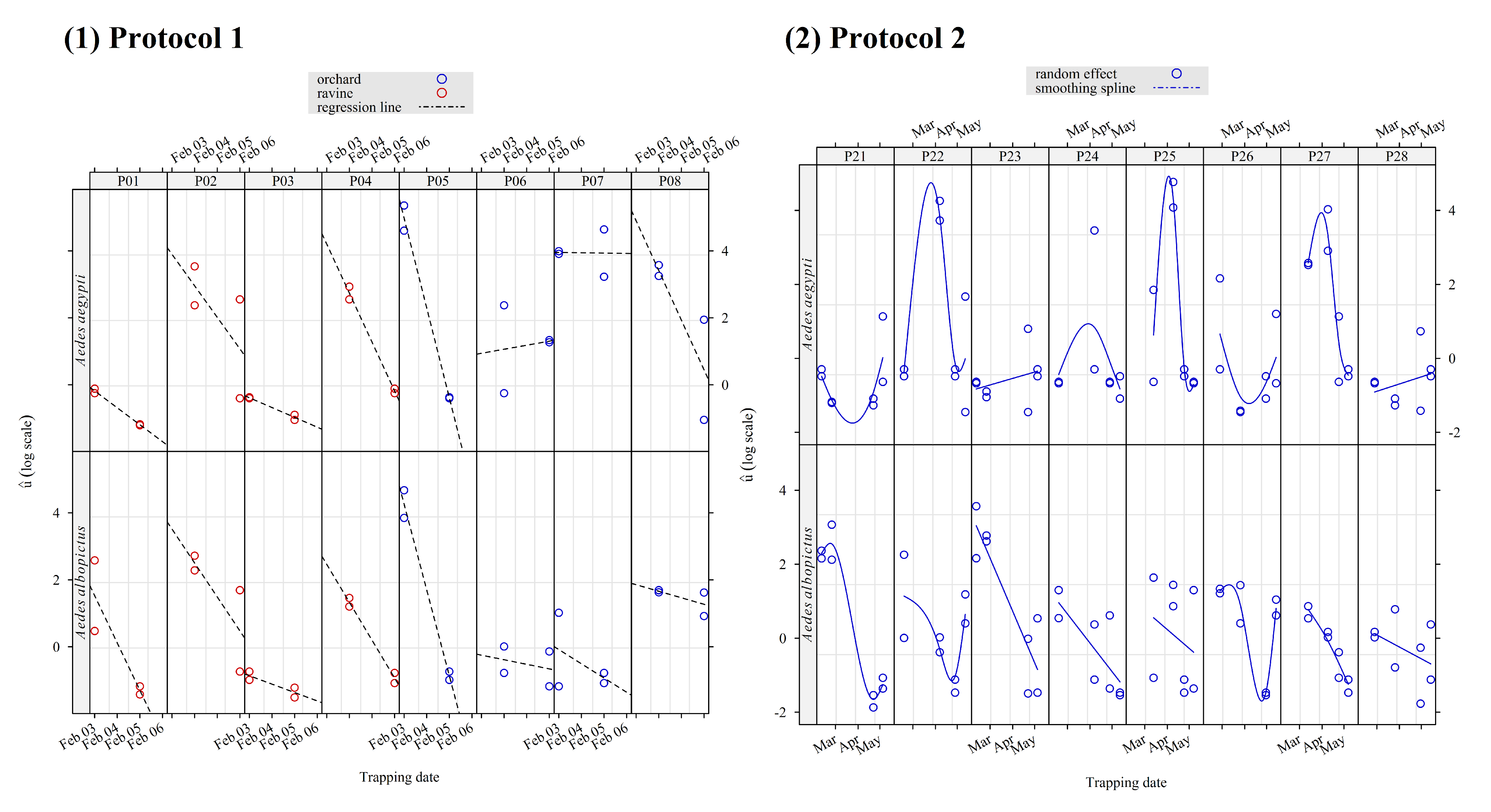
Figure 5.
Meteorological conditions ((a) minimum daily temperature; (b) daily rainfall) and time trend in the random effect ((c) Ae. albopictus; (d) Ae. aegypti) estimated with a Bayesian mixed-effect Poisson model of adult-mosquito attractiveness for CO2-baited BGS traps. The random effect shown on this plot was estimated with data from protocol 2: Latin square design, two sets of four traps in two similar locations, 8 replicates from March to May 2025. This plot was made with data collected during protocol 2. Meteorological data were provided by Meteo-France. They were taken from the station located in Saint-Joseph.
Figure 5.
Meteorological conditions ((a) minimum daily temperature; (b) daily rainfall) and time trend in the random effect ((c) Ae. albopictus; (d) Ae. aegypti) estimated with a Bayesian mixed-effect Poisson model of adult-mosquito attractiveness for CO2-baited BGS traps. The random effect shown on this plot was estimated with data from protocol 2: Latin square design, two sets of four traps in two similar locations, 8 replicates from March to May 2025. This plot was made with data collected during protocol 2. Meteorological data were provided by Meteo-France. They were taken from the station located in Saint-Joseph.
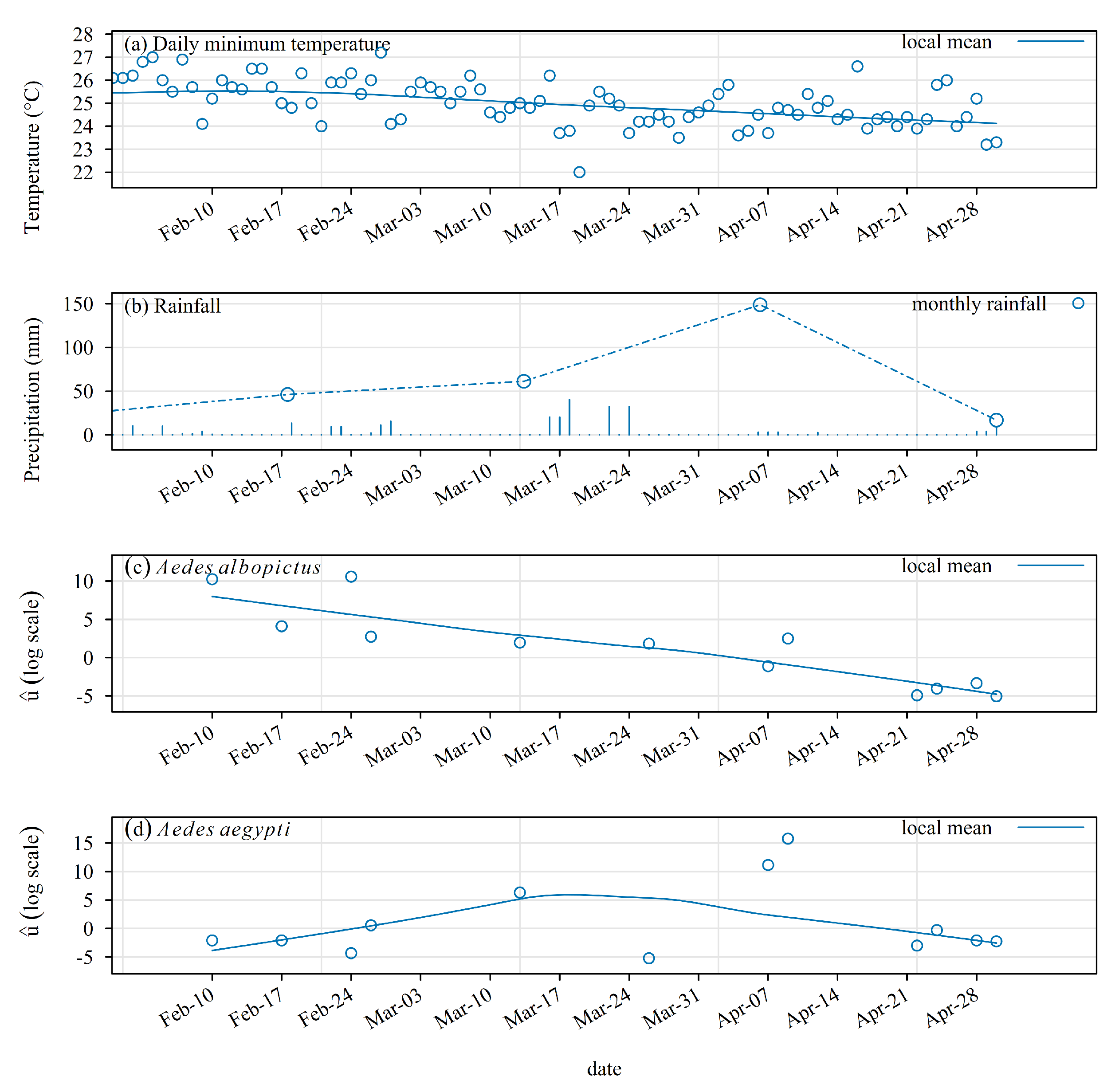
Figure 6.
Density of larvae in the water sampled in the stipes of vacoa trees in Saint-Joseph from February 2025 to April 2025. (a) density fitted with a negative binomial count model (b) correlation matrix of model coefficients: the correspondence with correlation is provided by the color key; on the diagonal, b is the fitted coefficient, and P is the p-value of the Wald test with H0: b = 0. The intercept is the log of fitted density of Ae. albopictus in February.
Figure 6.
Density of larvae in the water sampled in the stipes of vacoa trees in Saint-Joseph from February 2025 to April 2025. (a) density fitted with a negative binomial count model (b) correlation matrix of model coefficients: the correspondence with correlation is provided by the color key; on the diagonal, b is the fitted coefficient, and P is the p-value of the Wald test with H0: b = 0. The intercept is the log of fitted density of Ae. albopictus in February.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



