
Submitted:
21 July 2025
Posted:
23 July 2025
You are already at the latest version
Abstract
Phytophagous beetles, particularly those within the superfamilies Chrysomeloidea and Curculionoidea, constitute one of the most diverse and ecologically significant groups of insect herbivores. Within this group, the subfamily Bruchinae is especially notable for its close association with leguminous plant seeds. As most Bruchinae species do not feed during the adult stage, the timing and regulation of vitellogenesis remain unclear. Previous studies suggest that vitellogenesis may be triggered by volatile organic compounds emitted by host seeds, which promote juvenile hormone (JH) synthesis. This increase in JH is hypothesized to stimulate vitellogenesis, enhance female attractiveness, and ultimately facilitate fertilization and oviposition. To explore this hypothesis, we investigated the external cues regulating reproductive physiology in the capital breeder Zabrotes subfasciatus. Specifically, we examined the effects of host seeds and male presence on oviposition dynamics, fecundity, ovary activation, and the expression of vitellogenic genes (vg and vgR) throughout adult life. Our results show that females initiate vitellogenesis during the final phases of adult development, enabling oviposition to begin as early as the first day after emergence. Oviposition remains at basal levels throughout adult life unless both host seeds and males are present (p < 0.0001). This oviposition pattern is consistent with ovary activation dynamics, which reveal that vitellogenesis peaks early in the oviposition period and is prolonged by the presence of seeds and males (p < 0.05). Notably, vg and vgR gene expression respond differentially to these cues (p < 0.05). We integrate our findings with previous literature to propose a working model for the regulation of oviposition in the Bruchinae beetle Z. subfasciatus.
Keywords:
ovary activation
; vitellogenesis
; oviposition
; vitellogenin
; juvenile hormone
; host seed
; VOCs
; bean weevil
; Zabrotes subfasciatus
; Bruchinae
1. Introduction
Phytophagous beetles, particularly those from the superfamilies Chrysomeloidea and Curculionoidea, along with lepidopterans and hemipterans, constitute the majority of insects that feed on plants. With approximately 135,000 species [1], these insects have achieved significant biological success, utilizing seeds, leaves, trunks, and wood as their primary food sources. This success is largely attributed to the acquisition of genes encoding enzymes capable of degrading plant cell wall compounds, which were obtained through horizontal gene transfer from bacteria and fungi [2,3].
The family Chrysomelidae, specifically the subfamily Bruchinae (with approximately 1,700 species) [4], is particularly notable for its association with plants in the Fabaceae (Leguminosae) family [5,6]. Species from the following genera exemplify this relationship: Acanthoscelides, Callosobruchus, Stator, Bruchus, Bruchidius, Caryedon, and Zabrotes [7]. The association between Bruchinae beetles and dry legume seeds appears to be ancestral, likely favoured by long periods of drought and the availability of stored seeds [8,9]. In these insects, eggs are deposited onto the pods or directly onto the seeds. Upon hatching, the larvae burrow into the walls of the pods and/or seeds, feeding on the interior of the seeds as they grow and develop until reaching the pupal stage (which lasts between 7 and 28 days). The larvae then emerge as adults, mate, and complete a new cycle. In some species, the pupal development occurs outside the seeds, with the cocoon either attached to the seeds or directly to the soil [6]. Most adult beetles, particularly those of greater economic concern due to their infestation of stored seeds, do not feed. However, some species have the ability to consume food during their adult life [10], and in those inhabiting more “natural” niches, food intake seems to enhance reproductive capacity [6].
As the eggs deposited on seeds give rise to immatures that develop within them, members of this subfamily are considered pests [9]. Infestations by these insects lead to significant losses, with up to 20% reduction in the value of grains during pre-processing and packaging [11,12,13]. However, paradoxically, despite the economic importance of beetles that prey on legume seeds, and aside from ecological and evolutionary studies that use some reproductive parameters [14,15,16,17], the developmental biology of Bruchinae species remains poorly understood. Zabrotes subfasciatus, for instance, has been used to explore molecular and physiological aspects of digestion [18,19] and the adaptation of phytophagous insects to new hosts [20,21,22]. With the availability of its genomic sequence (Accession: PRJNA1192986), Z. subfasciatus has the potential to become a key biological model for molecular studies in Bruchinae. This could be beneficial both from an experimental manipulation standpoint -its natural niche is easily replicable in the lab- and from a representational perspective, as insights gained from studying Z. subfasciatus could inform the understanding and management of other Bruchinae species [23]. Additionally, using Z. subfasciatus as a model offers several advantages, such as the ease of distinguishing individuals by sex (females and males) and the ability to identify seeds that have been oviposited upon, whether fertilized or not, allowing for precise and efficient analysis [24,25].
Z. subfasciatus Boheman (1833) [26] (Coleoptera; Chrysomeloidea; Chrysomelidae; Bruchinae; Schoch et al., 2020) is a beetle native to Central and South America, from where it spread worldwide, primarily to tropical regions that cultivate Fabaceae [24,27]. This species can prey on a variety of Fabaceae, but it prefers varieties of common beans (Phaseolus vulgaris), where it demonstrates comparatively high fitness [22,28,29,30]. As with other Bruchinae species, females deposit and attach their eggs to the surface of a host seed. After the embryo develops, the larva hatches, penetrates the seed coat, and feeds on the cotyledons and embryonic regions of the seed. After completing pupal development, the adults emerge -typically after 26 days of development at 30-35°C [10] - undergo a brief 10-day reproductive period, and then die [22].
The predominant mechanism by which insects attempt to optimize their reproductive capacity in response to environmental conditions is likely the modulation of oocyte maturation rates [31]. In Orthoptera, Blattodea, Hemiptera, and Coleoptera, where reproduction depends on adult nutrition or mating, vitellogenesis is strictly regulated by juvenile hormone (JH) [32,33,34]. Many different external stimuli (mediated by hormones, as JH) are known to modulate oocyte maturation rates in various insects: temperature, humidity, food availability, social regulation, etc. [31]. In Z. subfasciatus, as in other studied Bruchinae species (e.g., A. obtectus, Bruchidius atrolineatus, Callosobruchus maculatus, and Caryedon serratus), ovarian development is facilitated only when the host plants are at, or nearly at, the phenological stage suitable for first-instar larvae to feed [13,35,36,37,38]. Behavioural assays suggest that males are attracted to females for mating, but only after the females are exposed to oviposition substrates [38,39,40], what increases ovary development [41] so that females are capable of ovipositing starting from the first day of adulthood [22].
Considering Z. subfasciatus adults do not usually feed, it is unclear whether vitellogenesis occurs during post-embryonic development (e.g., in the later phases of the pupal period), or whether its induction in adults depends on the presence of a male (and subsequent mating), similar to what occurs in Drosophila and Anastrepha [42,43,44,45]. Alternatively, does vitellogenesis depend on the presence of oviposition substrates, such as Fabaceae seeds, as seen in other beetles like the burying beetle (Nicrophorus orbicollis), or is it a combination of both factors, as suggested by Pimbert and Pierre [41]? In N. orbicollis (Polyphaga, Silphidae), which does not feed on the carcass where it lays eggs, JH levels increase dramatically within 20 minutes of contact with the substrate, inducing vitellogenesis, which then occurs in the following 18 hours [46,47]. If vitellogenesis in Z. subfasciatus depends on a factor derived from seeds (a chemosensory factor), the events must also be rapid, simultaneously promoting vitellogenesis, enhancing female attractiveness, and allowing fertilization. A system that could facilitate these events might involve volatile organic compounds (VOCs) emitted by seeds [21] and/or the involvement of maxillary palps [48], JH production to promote vitellogenesis, and pheromone release to attract males. In this study, we aimed to comprehensively explore the external cues that regulate the dynamics of the reproductive physiology in Bruchinae by examining the effects of development substrate (host seeds) and the presence of males on oviposition and fecundity profiles, ovary activation, and the expression of vitellogenic genes [49] during the adult life of the bean weevil Z. subfasciatus.
2. Materials and Methods
2.1. Weevil Husbandry
The Z. subfasciatus individuals used in this study were derived from a stock population of approximately 6,000 individuals, initially collected from bean seeds in Ribeirão Preto, São Paulo, Brazil, and maintained on bean seeds in the laboratory since 1997. According to Souza et al. [50], Brazilian populations of Z. subfasciatus exhibit low genetic differentiation and weak geographic structure. To mitigate potential inbreeding effects, individuals collected from bean seeds in Poços de Caldas and Alfenas, Minas Gerais, Brazil, have been incorporated into the stock population approximately every three months since 2007. The stock population was maintained on bean seeds at 22 ± 5°C and 70 ± 15% relative humidity, under dark conditions. Additional details regarding the biology of this insect can be found in many studies [25,51,52,53].
2.2. Histological Processing of Pupal and Adult Ovaries
To determine whether Z. subfasciatus females emerge with activated ovaries or if this activation occurs only in newly emerged individuals, we collected weevils maintained on beans from the stock. Ovaries dissected under a stereomicroscope (Carl Zeiss-Stemi 305) from adult and pupae stage individuals were fixed in Karnovsky’s solution [54] at 4°C for 48 hours. Both the pupae and the adult ovaries were dehydrated in a series of ethanol concentrations: 70%, 80%, 90%, and 95% (2 changes of 30 minutes each at room temperature). They were then gradually embedded in resin (Historesin embedding kit, Leica Biosystems), starting with a 2:1 (v/v) ratio of 95% ethanol to resin, followed by 1:1 and 1:2 (1 hour each at room temperature). Finally, the specimens were incubated in pure resin for 48 hours at 4°C. Blocks were prepared according to the kit’s instructions. Sections were made using a Leica RM2135 rotary microtome, at a thickness of 5 μm, with high-profile blades (Leica 818). The material was stained with Harris’ hematoxylin and eosin [55]. Micrographs were captured using an Olympus microscope, equipped with a digital camera (AmScope – MU1003) and AmLite imaging software.
2.3. Experimental Group Allocation: Groups A, B, C, and D
Virgin females were selected from individual infested bean seeds, identified by the presence of darkened, unbroken opercula. These seeds were placed in 10 mL test tubes sealed with cotton and maintained under controlled environmental conditions of 70% humidity and 30ºC temperature in a BOD incubator (Eletrolab model EL101/03), considered optimal for species development [28,30,56].
After emergence, individuals were distributed into acrylic tubes (3.5 cm in height, 2 cm at the base, and 3 cm in upper diameter), sealed with fine-mesh tulle fabric secured with an elastic band, and allocated into four experimental groups: only females (Group A), females with males but without host seeds (Group B), females with host seeds but without males (Group C), and females with both males and host seeds (Group D). The experiment was set up with 18 replicates per experimental group. The tubes from groups C and D included 10 bean seeds to ensure enough oviposition substrate thus avoiding competition [17,57].
2.3.1. Assessment of the Oviposition and Fecundity Profiles
To ensure accurate daily oviposition records, the tubes were replaced every 24 hours with new ones containing an equal number of host seeds. Ten days after the final replacement -when the eggs became visible- egg counts were conducted on both the inner surfaces of the tubes and the host seeds. In the groups where insect emergence was possible (B and D), adult emergence was recorded between days 25 and 26 following the last replacement. These data were used to estimate fecundity under the experimental conditions.
2.3.2. Dissection of Adult Ovaries, Fluorescent Staining, and Scoring of Ovariole Activation
To assess the impact of host seed availability and mate presence on Z. subfasciatus ovary activation, newly emerged females (0–24 hours old), as well as females aged 4 and 8 days from each experimental group (10 replicates per day per group), were dissected under a stereomicroscope (Carl Zeiss-Stemi 305) to obtain ovaries. Ovaries were dissected in 1× phosphate-buffered saline (PBS; Sigma Aldrich, UK: 137 mM NaCl, 2.7 mM KCl, 10 mM Na2HPO4, 1.8 mM KH2PO4) and fixed for 24 hours at 4°C in 200 µL of n-heptane (Vetec-920178) and 37% formaldehyde (Proquímicos-10/0010).
After fixation, ovaries were washed twice (5 minutes each) in PBS containing 0.5% Triton X-100 (PBST) and incubated at room temperature for 20 minutes in a solution of phalloidin conjugated to Alexa 546 (1:500 v/v ratio in PBST; Invitrogen) to stain F-actin, primarily highlighting cytoplasmic structures. Subsequently, samples were washed three times (5 minutes each) in PBST and incubated for 4 minutes in a DAPI solution at a 1:2000 v/v ratio in PBST. After a final wash (three times, 10 minutes each) in PBST, ovaries were mounted in 80% glycerol and examined under a fluorescence microscope (Nikon Eclipse 80i) and a confocal microscope C2+ (Nikon). Ovariole activation was assessed based on the degree of vitellogenesis in the oocytes, and females were classified into two stages: Stage I, comprising individuals with primarily early-stage oocytes, and Stage II, comprising those with predominantly mature, yolk-rich oocytes.
2.3.3. Quantification of Vitellogenic Genes Transcript Levels Using RT-qPCR
Total RNA was extracted using TRIzol® (Ambion) from pools (4 pools each) of three whole bodies and five pairs of ovaries and processed as previously reported [20]. Briefly, to eliminate contaminant DNA, total RNA was incubated at 37ºC with 1 unit of RQ1 RNase-free DNase (Sigma) for 30 minutes, followed by enzyme inactivation at 65ºC for 10 minutes using 1 µL of Stop DNase Solution (Sigma). First-strand cDNA was synthesized from 1 μg of total RNA via reverse transcription using SuperScript® IV Reverse Transcriptase (Invitrogen) and Oligo dT12-18 primers (Invitrogen). Negative control reactions, lacking the enzyme, were conducted in parallel. To quantitatively compare gene transcript levels between populations, a real-time qPCR assay was performed using a QuantStudio 3 PCR System (Applied Biosystems). Amplifications were carried out in 10 μL reaction volumes, each containing 5 μL SYBR Green Master Mix 2X (Applied Biosystems), 1 μL first-strand cDNA (diluted 1:4 v/v in water), 3.2 μL water, and 0.4 μL of each specific primer (from a 10 μM stock solution; Table S1). The gene encoding Z. subfasciatus ribosomal protein L32 (rpl32) was used as the reference gene [21]. Negative controls included reactions without reverse transcriptase or template. PCR conditions were set as follows: 50ºC for 2 minutes, 95ºC for 10 minutes, followed by 40 cycles of 95ºC for 15 seconds and 60ºC for 1 minute. To ensure reproducibility, each SYBR Green assay was conducted in technical duplicates, with at least three biological replicates [58], from the four pools per tissue, as mentioned above.
2.4. Data Analysis
Statistical analyses began with the Shapiro-Wilk test to assess the normality of all datasets. Since the total number of eggs laid across treatments followed a normal distribution, comparisons among groups were conducted using one-way ANOVA, followed by Tukey’s multiple comparisons test to identify significant differences. To examine the number of emerged adults over time in the group of females with males and host seeds (Group D), a two-way repeated measures ANOVA was applied, as the same individuals were monitored across multiple time points. Šídák’s multiple comparisons test was used to assess differences between days. On day 2 of the experiment, differences among the four experimental groups were analysed using the Kruskal-Wallis’s test followed by Dunn’s multiple comparisons test. This non-parametric approach was chosen due to the non-normal distribution of the data at this specific time point. To evaluate the level of ovarian activation (stages I and II) under different experimental conditions, categorical data (presence or absence of oocyte stages) were analysed using Fisher’s exact test.
For the quantification of vitellogenic gene transcript levels by RT-qPCR, raw data were normalized to the reference gene rpl32, and relative transcript quantities were calculated using the 2^-ΔΔCt method with efficiency correction and a calibrator sample [21,59] (Applied Biosystems User Bulletin #2). Since these data did not follow a normal distribution, the Kruskal -Wallis test was again employed, followed by Dunn’s multiple comparisons test. However, for comparisons between pupae and the other experimental groups, the Mann -Whitney U test was applied, as the pupa group had a small sample size and was not included in the main experimental design. All statistical analyses and graph construction were performed using GraphPad Prism version 10.5.0 (GraphPad Software, Boston, Massachusetts, USA, www.graphpad.com) [60]. Differences were considered statistically significant when p < 0.05.
3. Results
3.1. Female Zabrotes subfasciatus Begin Vitellogenesis During the Final Phases of Adult Development
Histological analysis of newly emerged Z. subfasciatus ovaries confirmed the presence of three ovarioles per ovary, exhibiting a telotrophic meroistic organization (Figure 1A-C). Additionally, examination of ovarian activation revealed that vitellogenesis is initiated during the final phases of adult development (Figure S1 and Figure 1). During the larval phases and the early phase of pupal development - specifically in white-eyed pupae-, the ovarioles consist solely of a mass of germ and somatic cells with no signs of oocyte development (Figure S1 and Figure 1D). In early adult development ovarioles contain immature oocytes, with no visible signs of vitellogenin uptake (Figure 1E). As development advances, oocytes begin to accumulate yolk in defined regions (Figure 1F), progressively resembling the fully developed oocytes observed in newly emerged females (Figure 1G).
3.2. Oviposition in Zabrotes subfasciatus Begins Shortly After Adult Emergence and Is Enhanced by the Availability of Seeds and Males
In ideal conditions (with seeds and males), Z. subfasciatus females began ovipositing on the first day after emergence, reaching a peak on the second day with 7 ± 3.5 eggs (mean ± SD; Kruskal-Wallis’s test followed by Dunn’s multiple comparisons test p < 0.0001). After this peak, oviposition gradually declined until day 10 (Figure 2AB; Table S1, Table S2, Table S3 and Table S4). The number of eggs laid generally corresponded to the number of emerging individuals, except on day 3, when a significant difference was observed (Figure 2A; two-way repeated measures ANOVA, followed by Šídák’s multiple comparisons test p < 0.05; Table S2 and S3). Females without access to an oviposition substrate and/or males laid a total of 3 ± 0.2 eggs (mean ± SD) throughout their lifespan, whereas those provided with both substrate and males laid significantly more (60 ± 44 eggs mean ± SD; ordinary one-way ANOVA followed by Tukey’s multiple comparisons test p < 0.0001; Figure 2B; Table S4). This pattern was further validated by daily oviposition records. Notably, only females from experimental group D (with both oviposition substrate and males) exhibited significantly higher oviposition rates compared to all other groups (Figure 2CD; Table S4). Females in group C exhibited a slight, though non-significant, increase in egg-laying on days 3 and 4 compared to those in groups A and B. Conversely, on day 1, females in groups A and B laid slightly more eggs than those in group C, although this difference was also not statistically significant (Figure 2D).
3.3. Vitellogenesis Is Elevated During the Early Phase of the Oviposition Period in Zabrotes subfasciatus, and Its Duration Is Extended by the Presence of Seeds and Males
To investigate how the experimental environment -specifically host and male presence- affects the reproductive physiology of Z. subfasciatus, we assessed ovarian activation in females from the four experimental groups (Figure 3AB; Table S5 and S6). Across all groups, a consistent pattern emerged: females were predominantly in Stage I at the beginning of the oviposition period (notably on day 1), and in Stage II by the end of the period (day 8), indicating a significant temporal shift in ovarian development (Fisher’s exact test, p < 0.05; Figure 3CD; Table S7 and S8). Moreover, by day 4, group D exhibited a higher proportion of females in Stage I and a lower proportion in Stage II compared to the other groups (Figure 3CD). However, the difference was statistically significant only between the mean of groups D and C regarding females in Stage I (Fisher’s exact test p = 0.0007). The mean values between females from the groups A and C regarding Stage I were also statistically different (Fisher’s exact test p = 0.023).
3.4. Male Presence Enhances Vitellogenin (vg) Expression, While Seeds Boost Vitellogenin Receptor (vgR) Expression in the Ovaries of Zabrotes subfasciatus Females
All analysed tissues from adult Z. subfasciatus females exhibited higher levels of vitellogenic genes [61] transcription compared to pharate-adult females, in which transcription was nearly undetectable. Both tested genes (vg and vgR) were expressed in the carcass and ovaries across all female age groups and experimental conditions (Figure 4; Table S9, Table S10, Table S11, Table S12 and Table S13). Median levels of vg gene transcripts in the carcass were generally consistent among all experimental groups throughout the study period, except on day 1. On this day, females from group D showed significantly higher vg expression than those from the other groups, with a statistically significant difference observed compared to group B (Figure 4A; Kruskal-Wallis’s test followed by Dunn’s multiple comparisons test, p < 0.05; Table S14). Expression of the vgR gene in carcass tissue showed more variability. On day 1, females from group C had significantly higher vgR transcript levels than those in group B, and on day 8, they exceeded those in group A (Figure 4B; Kruskal-Wallis’s test followed by Dunn’s multiple comparisons test, p < 0.05; p < 0.01; Table S14). Additionally, group D females showed significantly greater vgR expression than group B on day 4 (Figure 4B; Kruskal-Wallis’s test followed by Dunn’s multiple comparisons test, p < 0.05; Table S14). In ovarian tissues, vg transcript levels were consistently higher in group D females, though the difference reached statistical significance only on day 4 compared to group B (Figure 4C; Kruskal-Wallis’s test followed by Dunn’s multiple comparisons test, p < 0.05; Table S15). Lastly, vgR expression also varied among groups. The highest median expression levels were consistently observed in females from group C, with statistically significant differences compared to group B on day 1 and group A on day 4 (Figure 4D; Kruskal-Wallis’s test followed by Dunn’s multiple comparisons test, p < 0.05; p < 0.01; Table S15).
4. Discussion
We first confirmed that adults Z. subfasciatus carry three ovarioles per ovary, each with telotrophic meroistic organization, and showed that vitellogenesis initiates during the final phases of adult development. Under ideal conditions, females began ovipositing on the first day after emergence, reaching a peak on the second day, after which the oviposition gradually declines until day 10. Oviposition is at basal low levels during the whole adult life of the females unless both seeds and males are present. The observed oviposition profile was further confirmed by the ovary activation (vitellin accumulation) dynamics which revealed vitellogenesis is elevated during the early phase of the oviposition period and its duration is extended by the presence of seeds and males, which affects differentially vitellogenin and vitellogenin receptor expression.
Ovariole number is genetically determined and ovary type is phylogenetically predicted. Some of the ovary characteristics, though may suffer alterations from cellular perturbations during the later phases of larval development and can be modified by the diet of the immature stages [62]. As reported previously for other Bruchinae [63,64,65], females Z. subfasciatus have ovaries with three telotrophic meroistic ovarioles. Each telotrophic ovariole has an anterior tropharium, in which nurse cells remain, synthesize and contribute their products finally into all growing oocytes via nutritive cords [66,67].
As predicted by the concept of capital breeding [68], based on reports on the nutritional biology of Bruchinae [6], we showed that Z. subfasciatus begin vitellogenesis during the final phases of adult development. Since there is no food intake during the adult development, the result suggests that during the larval feeding period, females accumulate nutrients in their fat body cells, which contributes to the development of oocytes during later post-embryonic phases and early days of adult life.
Our results show that oviposition, as a result of ovary activation and vitellogenesis, concentrates during the early days after adult emergence, with a peak already in the second day of adult life, thus partially agreeing with the finding reported in Teixeira et al. [22]. These authors showed that the weevils reared on beans have an oviposition maximum already on the first day after emergence. However, the authors also showed that variations in the host seed types may shift the observed oviposition peak.
Females Z. subfasciatus show basal low levels of oviposition during the whole adult life unless both seeds (host) and males are present. This result indicates that both factors are essential for promoting oviposition levels compatible with populational maintenance and growth, as suggested in an early work by Pimbert and Pierre [41]. Our results also show that the high levels of oviposition registered during the first days of adult life are fuelled by high levels of oocyte development, as evidenced by the high proportion of females in the Stage I of ovary activation, including developing and fully-developed oocytes. This condition was registered to last during the following days in the females reared in the presence of seed and males. During the last part of the adult life, however, females from all experimental groups have their ovaries in Stage II, i.e., mainly with fully developed oocytes. Interestingly, the females reared in the ideal condition showed the lowest proportion of fully developed oocytes retained within their reproductive system, at least before reaching the final days of the adult life. This result suggests that vitellogenesis might not depend directly on external factors and egg laying (evidenced by the proportionally low level of retained fully-developed oocytes) is enhanced by the presence of males in females with available host seeds (the latter also suggested by Pimbert and Pierre [41]). Moreover, the observation that females from all experimental groups retain Stage II ovaries until the end of their lives is noteworthy and warrants further investigation. This may indicate an inability to lay eggs, potentially linked to aging processes in females, males, or even to social factors. The latter is particularly plausible given that, in natural settings, females can encounter other females and may mate with multiple males [69] - conditions that could influence oviposition behaviour [70].
Vitellogenin has been shown to be a pleiotropic protein, performing multiple functions beyond its traditional role in yolk-protein formation in insects [71]. In line with this, we found that the vitellogenin gene -and that of its receptor, vgR- is expressed in all analysed tissues, including both the carcass and ovary. The expression of vgR in the carcass appears to be regulated in a complex manner by external factors, also warranting further investigation. This pattern of widespread expression is consistent with findings in the bumblebee [72], and notably includes ovarian expression, as also observed in the Chagas disease vector Rhodnius prolixus [73]. Interestingly, although vg is expressed at very low levels in pharate adults, its presence at this stage aligns with the detection of vitellogenic oocytes, suggesting an early onset of reproductive activity. These findings collectively indicate that basal vg transcription and its incorporation into oocytes are regulated by internal physiological signals -likely hormonal- and that the production of these cues may be further amplified by external environmental factors.
The expression of vg in the carcass (which includes the fat body) and ovaries is enhanced -at least at certain points during the oviposition period- by the presence of males when host seeds are available. In contrast, vgR expression in the ovaries appears to depend primarily on the availability of an oviposition substrate, suggesting that vg expression in the fat body and vg uptake by oocytes are physiological processes regulated by distinct external cues. As reported in other insect species, vg gene expression can be induced by JH [33]. vgR, however, has been described as being expressed in a relatively autonomous manner in some insects [74,75], a pattern that also appears to apply to the basal expression levels observed in Z. subfasciatus. Similar to findings in the beet armyworm Spodoptera exigua [76], vgR expression in the bean weevil is strongly modulated by the presence of an oviposition substrate- in this case, host seeds.
5. Conclusions
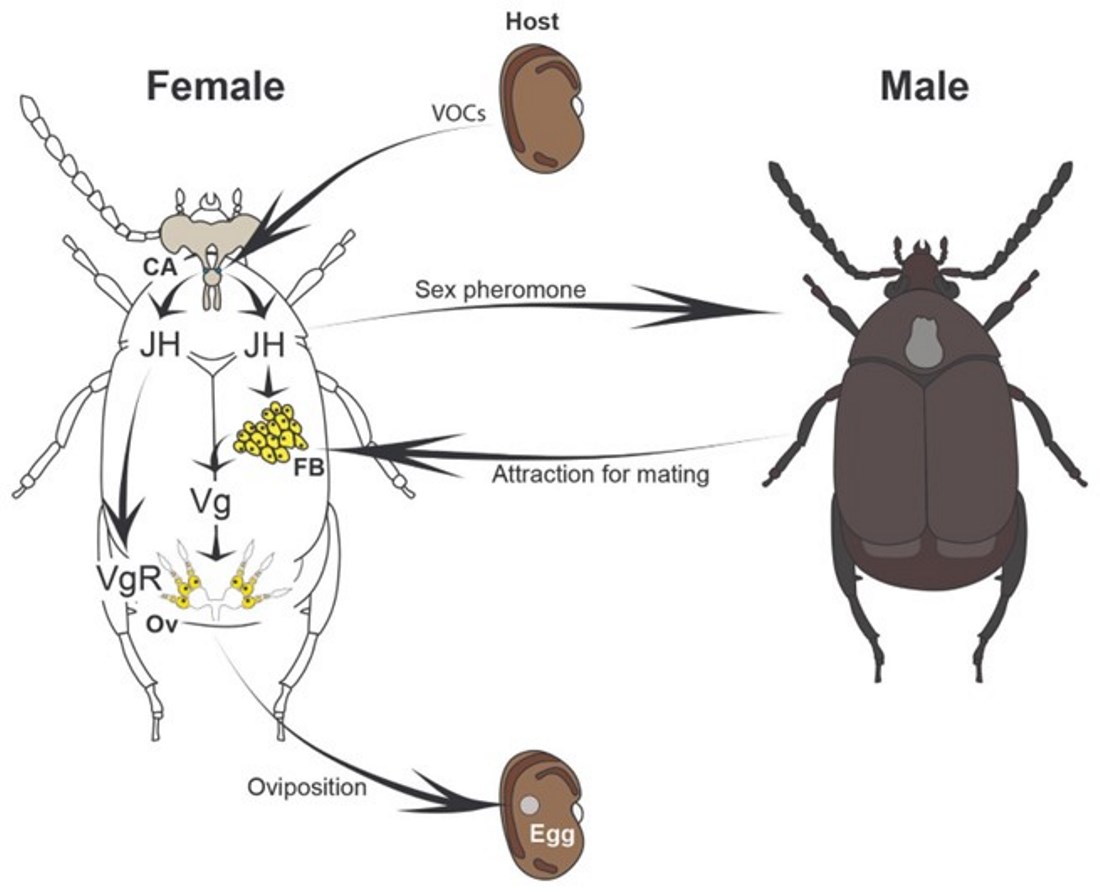
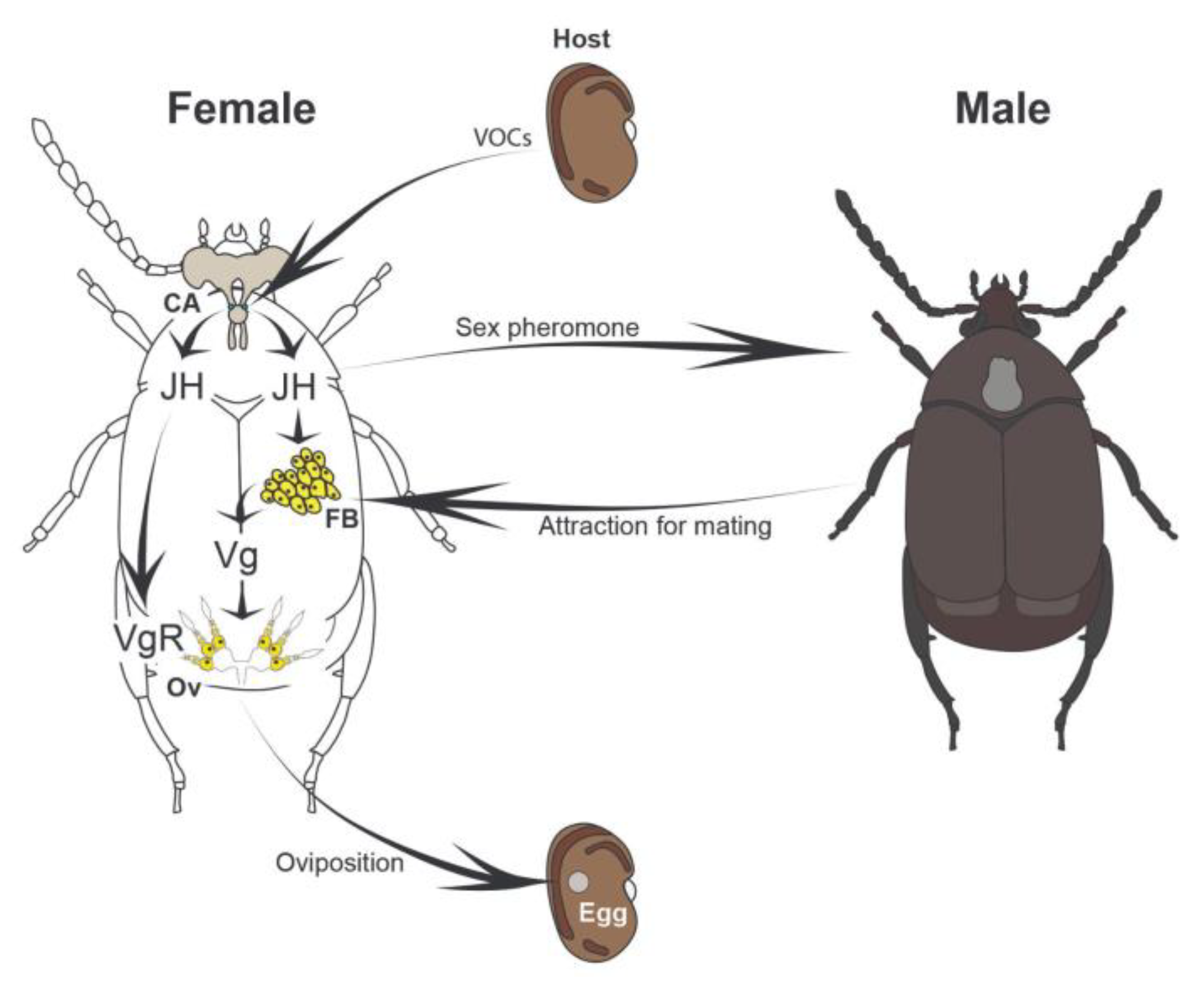
Our results indicate that the capital breeder Z. subfasciatus initiates vitellogenesis during the final phases of adult development. This process is likely fuelled by nutrients stored in the fat body cells during the larval stage. Consequently, the female’s physiological system is primed to begin oviposition shortly after adult emergence. Based on our findings and previous literature, we propose a model in which host seeds emit VOCs [77] that enhance JH synthesis in females [38,40]. Elevated JH levels, in turn, trigger the production and release of sex pheromones [40,41,78], promoting male attraction and facilitating mating. JH also induces the expression of the vitellogenin receptor (vgR) in the ovaries (and vg in fat bodies?), while the increased mating activity further modulates vitellogenin (vg) expression and promotes oviposition (Figure 5).
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Tables S1-S16.
Author Contributions
Conceptualization, SOM, IRVT and ARB; Methodology, SOM, IRVT, and ARB; Software, WOM; Validation, SOM and ARB.; Formal Analysis, SOM, BOC, and WOM; Investigation, SOM, BOC, JRM, ESCN, and LMRM; Resources, SOM, IRVT, and ARB; Data Curation, SOM; Writing – Original Draft Preparation, SOM and ARB; Writing – Review & Editing, SOM, JRM, IRVT, and ARB; Visualization, SOM, BOC, and JRM; Supervision, TSM and ARB; Project Administration, ARB; Funding Acquisition, IRVT and ARB.
Funding
Financial support was provided by the Fundação de Amparo à Pesquisa do Estado de Minas Gerais (FAPEMIG Grant Proc. # APQ-00325-22), Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq Grant Proc. # 404696/2021-0; CNPq Fellowship Proc. # 307847/2021-7), and the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior - Brasil (CAPES) - Finance Code 001. We are also grateful to institutional grants provided by FINEP – Financiadora de Estudos e Projetos. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Data Availability Statement
The original contributions presented in this study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author.
Acknowledgments
In this section, you can acknowledge any support given which is not covered by the author contribution or funding sections. This may include administrative and technical support, or donations in kind (e.g., materials used for experiments). Where GenAI has been used for purposes such as generating text, data, or graphics, or for study design, data collection, analysis, or interpretation of data, please add “During the preparation of this manuscript/study, the author(s) used [tool name, version information] for the purposes of [description of use]. The authors have reviewed and edited the output and take full responsibility for the content of this publication.”
Conflicts of Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- Lawrence, J.F. Coleoptera. In Synopsis and Classification of Living Organism; Parker, S.P., Ed.; MacGraw-Hill, 1982; Vol. 2, pp. 482–553.
- McKenna, D.D.; Shin, S.; Ahrens, D.; Balke, M.; Beza-Beza, C.; Clarke, D.J.; Donath, A.; Escalona, H.E.; Friedrich, F.; Letsch, H.; et al. The Evolution and Genomic Basis of Beetle Diversity. Proceedings of the National Academy of Sciences 2019, 116, 24729–24737. [Google Scholar] [CrossRef] [PubMed]
- McKenna, D.D.; Scully, E.D.; Pauchet, Y.; Hoover, K.; Kirsch, R.; Geib, S.M.; Mitchell, R.F.; Waterhouse, R.M.; Ahn, S.-J.; Arsala, D.; et al. Genome of the Asian Longhorned Beetle (Anoplophora Glabripennis), a Globally Significant Invasive Species, Reveals Key Functional and Evolutionary Innovations at the Beetle–Plant Interface. Genome Biology 2016, 17, 227. [Google Scholar] [CrossRef] [PubMed]
- Morse, G. Bruchinae Latereille. In Arthropoda: Insecta: Coleoptera.; Handbook of Zoology; Vol. 3, pp. 189–197.
- Johnson, C.D. Seed Beetle Host Specificity and the Systematics of the Leguminosae. In Advances in Legume Systematics Part 2; Ponhill, R.M., Ed.; 1981.
- Southgate, B.J. Biology of the Bruchidae. Ann. Rev. Entomol 1979, 24, 449–473. [Google Scholar] [CrossRef]
- Kingsolver, J.M. Handbook of the Bruchidae of the United States and Canada; United States Department of Agriculture, 2004.
- Diamond, J. Guns, Germs, and Steel: The Fates of Human Societies; W. W. Norton & Company, 1997.
- Tuda, M. Evolutionary Diversification of Bruchine Beetles: Climate-Dependent Traits and Development Associated with Pest Status. Bulletin of Entomological Research 2011, 101, 415–422. [Google Scholar] [CrossRef]
- Corrêa, C.P.; Parreiras, S.S.; Beijo, L.A.; Ávila, P.M. de; Teixeira, I.R.V.; Barchuk, A.R. Life History Trait Response to Ambient Temperature and Food Availability Variations in the Bean Weevil Zabrotes subfasciatus. Physiological Entomology 2021, 46, 189–199. [Google Scholar] [CrossRef]
- Mesterházy, Á.; Oláh, J.; Popp, J. Losses in the Grain Supply Chain: Causes and Solutions. Sustainability 2020, 12, 2342. [Google Scholar] [CrossRef]
- Pimbert, M. A Model of Host Plant Change of Zabrotes subfasciatus Boh. (Coleoptera: Bruchidae) in a Traditional Bean Cropping System in Costa Rica. Biological Agriculture & Horticulture 1985, 3, 39–54. [Google Scholar] [CrossRef]
- Pimbert, M. Reproduction and Oviposition Preferences of Zabrotes subfasciatus Stocks Reared from Two Host Plant Species. Entomologia Experimentalis et Applicata 1985, 38, 273–276. [Google Scholar] [CrossRef]
- Fox, C.W. The Effect of Inbreeding on Natural Selection in a Seed-feeding Beetle. Journal of Evolutionary Biology 2013, 26, 88–93. [Google Scholar] [CrossRef]
- Fox, C.W.; Waddell, K.J.; Mousseau, T.A. Host-Associated Fitness Variation in a Seed Beetle (Coleoptera: Bruchidae): Evidence for Local Adaptation to a Poor Quality Host. Oecologia 1994, 99, 329–336. [Google Scholar] [CrossRef]
- Sayadi, A.; Barrio, A.M.; Immonen, E.; Dainat, J.; Berger, D.; Tellgren-Roth, C.; Nystedt, B.; Arnqvist, G. The Genomic Footprint of Sexual Conflict. Nature Ecology & Evolution 2019, 3, 1725–1730. [Google Scholar] [CrossRef]
- Teixeira, I.R. do V. ; Beijo, L.A.; Barchuk, A.R. Behavioral and Physiological Responses of the Bean Weevil Zabrotes subfasciatus to Intraspecific Competition. Journal of Stored Products Research 2016, 69, 51–57. [Google Scholar] [CrossRef]
- Ibrahim, S.P.; Dias, R.O.; Ferreira, C.; Silva, C.P.; Terra, W.R. Histochemistry and Transcriptomics of Mucins and Peritrophic Membrane (PM) Proteins along the Midgut of a Beetle with Incomplete PM and Their Complementary Function. Insect Biochemistry and Molecular Biology 2023, 162, 104027. [Google Scholar] [CrossRef] [PubMed]
- Silva, C.P.; Terra, W.R.; Sá, M.F.G. de; Samuels, R.I.; Isejima, E.M.; Bifano, T.D.; Almeida, J.S. Induction of Digestive α-Amylases in Larvae of Zabrotes subfasciatus (Coleoptera: Bruchidae) in Response to Ingestion of Common Bean α-Amylase Inhibitor 1. Journal of Insect Physiology 2001, 47, 1283–1290. [Google Scholar] [CrossRef] [PubMed]
- Cruz, B. de O.; Miranda, S. de O.; Benedito, E.R.C.; Martins, J.R.; Beijo, L.A.B.; Nogueira, E.S.C.; Mazzoni, T.S.; Moda, L.M.R.; Teixeira, I.R. do V.; Barchuk, A.R. Early Adaptation to an Unusual Host in the Bean Weevil Zabrotes subfasciatus Is Associated with Changes in Body Size and Reproductive Physiology. Physiological Entomology 2025.
- Rodrigues, P.A. da P.; Martins, J.R.; Capizzani, B.C.; Hamasaki, L.T.A.; Simões, Z.L.P.; Teixeira, I.R. do V.; Barchuk, A.R. Transcriptional Signature of Host Shift in the Seed Beetle Zabrotes subfasciatus. Genetics and Molecular Biology 2024, 47. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, I.R. do V.; Barchuk, A.R.; Zucoloto, F.S. Host Preference of the Bean Weevil Zabrotes subfasciatus. Insect Science 2008, 15, 335–341. [CrossRef]
- Love, A.C.; Yoshida, Y. Reflections on Model Organisms in Evolutionary Developmental Biology. In; 2019; pp. 3–20.
- Valencia, C.A.; Mejía, C.C.; Schoonhoven, A. van Main Insect Pests of Stored Beans and Their Control [Tutorial Unit].; Centro Internacional de Agricultura Tropical (CIAT), 1986.
- Teixeira, I.R. do V.; Gris, C.F. Genetic Diversity of Grains, Storage Pests and Their Effects on the Worldwide Bean Supply. In Beans: Nutrition, Consumption and Health; Popescu, E., Golubev, I., Eds.; 2011.
- Boheman, C.H. Genera et Species Curculionidum, Cum Synonymia Hujus Familiae; Schoenherr, C.J., Ed.; Synonymia insectorum, oder: Versuch einer Synonymie aller bisher bekannten Insecten; nach Fabricii Systema Eleutheratorum &c. geordnet; Roret: Paris, France, 1833.
- Lucas, M.H. Spermophagus Semifasicatus. In Proceedings of the Bulletins trimestriels de la Société Entomologique de France; Annales de la Société Entomologique de France, 1858.
- Corrêa, C.P.; Capizzani, B.C.; Beijo, L.A.; Ávila, P.M. de; Teixeira, I.R. do V.; Barchuk, A.R. Adult Feeding and Host Type Modulate the Life History Traits of the Capital Breeder Zabrotes subfasciatus. Physiological Entomology 2020, 45, 120–128. [Google Scholar] [CrossRef]
- Meik, J.; Dobie, P. The Ability of Zabrotes subfasciatus to Attack Cowpeas. Entomologia Experimentalis et Applicata 1986, 42, 151–158. [Google Scholar] [CrossRef]
- Teixeira, I.R.V.; Zucoloto, F.S. Seed Suitability and Oviposition Behaviour of Wild and Selected Populations of Zabrotes subfasciatus (Boheman) (Coleoptera: Bruchidae) on Different Hosts. Journal of Stored Products Research 2003, 39, 131–140. [Google Scholar] [CrossRef]
- Hodin, J. She Shapes Events as They Come: Plasticity in Female Insect Reproduction. In Phenotypic Plasticity of Insects: Mechanisms and Consequences; Science Publishers: Enfield, NH, USA, 2009; pp. 423–521. [Google Scholar]
- Soller, M.; Bownes, M.; Kubli, E. Control of Oocyte Maturation in Sexually Mature Drosophila Females. Developmental Biology 1999, 208, 337–351. [Google Scholar] [CrossRef]
- Wu, Z.; Yang, L.; He, Q.; Zhou, S. Regulatory Mechanisms of Vitellogenesis in Insects. Frontiers in Cell and Developmental Biology 2021, 8. [Google Scholar] [CrossRef]
- Leyria, J. Endocrine Factors Modulating Vitellogenesis and Oogenesis in Insects: An Update. Molecular and Cellular Endocrinology 2024, 587, 112211. [Google Scholar] [CrossRef]
- Delobel, A. Effect of Groundnut Pods (Arachis hypogaea) and Imaginal Feeding on Oogenesis, Mating and Oviposition in the Seed Beetle, Caryedon Serratus. Entomol. Exp. Appl. 1989, 52, 281–289. [Google Scholar] [CrossRef]
- Pouzat, J.; Bilal, H.; Nammour, D.; Pimbert, M. A Comparative Study of the Host Plant’s Influence on the Sex Pheromone Dynamics of Three Bruchid Species. Acta Oecologica, Oecologia Generalis 1989, 10, 401–410. [Google Scholar]
- Papaj, D.R. Ovarian Dynamics and Host Use. Annual Review of Entomology 2000, 45, 423–448. [Google Scholar] [CrossRef] [PubMed]
- Pimbert, M.; Pouzat, J. Electroantennogram Responses of Zabrotes subfasciatus to Odours of the Sexual Partner. Entomologia Experimentalis et Applicata 1988, 47, 49–53. [Google Scholar] [CrossRef]
- Parsons, D.M.J.; Credland, P.F. Determinants of Oviposition in Acanthoscelides obtectus: A Nonconformist Bruchid. Physiological Entomology 2003, 28, 221–231. [Google Scholar] [CrossRef]
- Analysis in a Tubular Olfactometer of the Influence of Different Olfactory Stimuli on the Search for a Sexual Partner by Zabrotes subfasciatus. Entomologia Experimentalis et Applicata 1987.
- Pimbert, M.; Pierre, D. Ecophysiological Aspects of Bruchid Reproduction: The Influence of Pod Maturity and Seeds of Phaseolus vulgaris and the Influence of Insemination on the Reproductive Activity of Zabrotes subfasciatus. Ecological Entomology 1983, 8, 87–94. [Google Scholar] [CrossRef]
- Craddock, E.M.; Boake, C.R.B. Onset of Vitellogenesis in Female Drosophila silvestris Is Accelerated in the Presence of Sexually Mature Males. Journal of Insect Physiology 1992, 38, 643–650. [Google Scholar] [CrossRef]
- Boake, C.R.B.; Moore, S. Male Acceleration of Ovarian Development in Drosophila silvestris (Diptera, Drosophilidae): What Is the Stimulus? Journal of Insect Physiology 1996, 42, 649–655. [Google Scholar] [CrossRef]
- Knapp, R.A.; Norman, V.C.; Rouse, J.L.; Duncan, E.J. Environmentally Responsive Reproduction: Neuroendocrine Signalling and the Evolution of Eusociality. Current Opinion in Insect Science 2022, 53, 100951. [Google Scholar] [CrossRef]
- Pereira, R.; Teal, P.E.A.; Sivinski, J.; Dueben, B.D. Influence of Male Presence on Sexual Maturation in Female Caribbean Fruit Fly, Anastrepha suspensa (Diptera: Tephritidae). Journal of Insect Behavior 2006, 19, 31–43. [Google Scholar] [CrossRef]
- Trumbo, S.T. Juvenile Hormone-Mediated Reproduction in Burying Beetles: From Behavior to Physiology. Archives of Insect Biochemistry and Physiology 1997, 35, 479–490. [Google Scholar] [CrossRef]
- Trumbo, S.T.; Borst, D.W.; Robinson, G.E. Rapid Elevation of Juvenile Hormone Titer during Behavioral Assessment of the Breeding Resource by the Burying Beetle, Nicrophorus orbicollis. Journal of Insect Physiology 1995, 41, 535–543. [Google Scholar] [CrossRef]
- Pouzat, J. Host Plant Chemosensory Influence on Oogenesis in the Bean Weevil Acanthoscelides obtectus (Coleoptera: Bruchidae). Entomologia Experimentalis et Applicata 1978, 24, 601–608. [Google Scholar] [CrossRef]
- Li, H.-L.; Wang, X.-Y.; Zheng, X.-L.; Lu, W. Research Progress on Oviposition-Related Genes in Insects. Journal of Insect Science 2020, 20. [Google Scholar] [CrossRef]
- Souza, G.A. de; Carvalho, M.R. de O.; Martins, E.R.; Guedes, R.N.C.; Oliveira, L.O. de Diversidade Genética Estimada Com Marcadores ISSR Em Populações Brasileiras de Zabrotes subfasciatus. Pesquisa Agropecuária Brasileira 2008, 43, 843–849. [CrossRef]
- Bondar, G.G. Notas Biologicas Sobre Bruchideros Observados No Brasil. Tip. do Jornal do Commercio 1937.
- Teixeira, I.R. do V.; Barchuk, A.R.; Medeiros, L.; Zucoloto, F.S. Females of the Weevil Zabrotes subfasciatus Manipulate the Size and Number of Eggs According to the Host Seed Availability. Physiological Entomology 2009, 34, 246–250. [CrossRef]
- Teixeira, I.R.V.; Zucoloto, F.S. Intraspecific Competition in Zabrotes subfasciatus: Physiological and Behavioral Adaptations to Different Amounts of Host. Insect Science 2012, 19, 102–111. [Google Scholar] [CrossRef]
- Karnovsky, M.J. A Formaldehyde Glutaraldehyde Fixative of High Osmolality for Use in Electron Microscopy. The Journal of Cell Biology 1965, 27, 1A–149A. [Google Scholar]
- Harris, H.F. On the Rapid Conversion of Haematoxylin into Haematein in Staining Reactions. Journal of Applied Microscopic Laboratory Methods 1900, 3, 777. [Google Scholar]
- Howe, R.W.; Currie, J.E. Some Laboratory Observations on the Rates of Development, Mortality and Oviposition of Several Species of Bruchidae Breeding in Stored Pulses. Bulletin of Entomological Research 1964, 55, 437–477. [Google Scholar] [CrossRef]
- Dendy, J.; Credland, P.F. Development, Fecundity and Egg Dispersion of Zabrotes subfasciatus. Entomologia Experimentalis et Applicata 1991, 59, 9–17. [Google Scholar] [CrossRef]
- Silva, J.A. da; Farder-Gomes, C.F.; Barchuk, A.R.; Malaspina, O.; Nocelli, R.C.F. Sublethal Exposure to Thiamethoxam and Pyraclostrobin Affects the Midgut and Malpighian Tubules of the Stingless Bee Frieseomelitta varia (Hymenoptera: Apidae: Meliponini). Ecotoxicology 2024, 33, 875–883. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- GraphPad Software. GraphPad Prism, Version 10.5.0; GraphPad Software: Boston, MA, USA, 2023; Available online: https://www.graphpad.com/ (accessed on 17 July 2025).
- Sappington, T.W.; Raikhel, A.S. Molecular Characteristics of Insect Vitellogenins and Vitellogenin Receptors. Insect Biochemistry and Molecular Biology 1998, 28, 277–300. [Google Scholar] [CrossRef]
- Klowden, M.J.; Pallai, S.R. Physiological Systems in Insects; Fourth edition.; Academic Press, an imprint of Elsevier: London ; San Diego, CA, 2023; ISBN 978-0-12-820359-0.
- Mohamed, M.I.; Khaled, A.S.; Fattah, H.M.A.; Hussein, M.A.; Salem, D.A.M.; Fawki, S. Ultrastructure and Histopathological Alteration in the Ovaries of Callosobruchus maculatus (F.) (Coleoptera, Chrysomelidae) Induced by the Solar Radiation. The Journal of Basic & Applied Zoology 2015, 68, 19–32. [Google Scholar] [CrossRef]
- Aggarwal, S.K. Morphological and Histochemical Studies on Oogenesis in Callosobruchus analis Fabr. (Bruchidae-Coleoptera). Journal of Morphology 1967, 122, 19–33. [Google Scholar] [CrossRef]
- Büning, J. The Telotrophic Nature of Ovarioles of Polyphage Coleoptera. Zoomorphologie 1979, 93, 51–57. [Google Scholar] [CrossRef]
- Büning, J. Ovariole Structure Supports Sistergroup Relationship of Neuroptera and Coleoptera. ASP 2006, 64, 115–126. [Google Scholar] [CrossRef]
- Belles, X.; Piulachs, M.-D. Ecdysone Signalling and Ovarian Development in Insects: From Stem Cells to Ovarian Follicle Formation. Biochimica et Biophysica Acta (BBA) - Gene Regulatory Mechanisms 2015, 1849, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Jönsson, K.I.; Jonsson, K.I. Capital and Income Breeding as Alternative Tactics of Resource Use in Reproduction. Oikos 1997, 78, 57. [Google Scholar] [CrossRef]
- Katvala, M.; Rönn, J.L.; Arnqvist, G. Correlated Evolution between Male Ejaculate Allocation and Female Remating Behaviour in Seed Beetles (Bruchidae). Journal of Evolutionary Biology 2008, 21, 471–479. [Google Scholar] [CrossRef] [PubMed]
- Bailly, T.P.M.; Kohlmeier, P.; Etienne, R.S.; Wertheim, B.; Billeter, J.-C. Social Modulation of Oogenesis and Egg Laying in Drosophila melanogaster. Current Biology 2023, 33, 2865–2877. [Google Scholar] [CrossRef]
- Montserrat-Canals, M.; Schnelle, K.; Leipart, V.; Halskau, Ø.; Amdam, G.V.; Moeller, A.; Cunha, E.S.; Luecke, H. Cryo-EM Structure of Native Honey Bee Vitellogenin. Nature Communications 2025, 16, 5736. [Google Scholar] [CrossRef] [PubMed]
- Du, L.; Wang, M.; Li, J.; He, S.; Huang, J.; Wu, J. Characterization of a Vitellogenin Receptor in the Bumblebee, Bombus lantschouensis (Hymenoptera, Apidae). Insects 2019, 10, 445. [Google Scholar] [CrossRef]
- Melo, A.C.A.; Valle, D.; Machado, E.A.; Salerno, A.P.; Paiva-Silva, G.O.; Silva, N.L.C.E.; Souza, W. de; Masuda, H. Synthesis of Vitellogenin by the Follicle Cells of Rhodnius prolixus. Insect Biochemistry and Molecular Biology 2000, 30, 549–557. [Google Scholar] [CrossRef] [PubMed]
- Schonbaum, C.P.; Perrino, J.J.; Mahowald, A.P. Regulation of the Vitellogenin Receptor during Drosophila melanogaster Oogenesis. Molecular Biology of the Cell 2000, 11, 511–521. [Google Scholar] [CrossRef] [PubMed]
- Tufail, M.; Takeda, M. Molecular Cloning, Characterization and Regulation of the Cockroach Vitellogenin Receptor during Oogenesis. Insect Molecular Biology 2005, 14, 389–401. [Google Scholar] [CrossRef]
- Zhao, J.; Sun, Y.; Xiao, L.; Tan, Y.; Jiang, Y.; Bai, L. Vitellogenin and Vitellogenin Receptor Gene Expression Profiles in Spodoptera exigua Are Related to Host Plant Suitability. Pest Management Science 2018, 74, 950–958. [Google Scholar] [CrossRef]
- Capizzani, B.C.; Rainho, H.L.; Miranda, S. de O.; Rosa, V.D. de S.; Beijo, L.A.; Teixeira, I.R. do V.; Bento, J.M.; Barchuk, A.R. Contrasting Responses to Ethenylbenzene (Styrene) and 2-Ethyl-1-Hexanol Suggest Their Role as Chemical Cues in Host Selection by the Seed Beetle Zabrotes subfasciatus (Coleoptera: Chrysomelidae: Bruchinae). Neotropical Entomology 2024, 54, 9. [Google Scholar] [CrossRef]
- Zhang, X.; Li, S.; Liu, S. Juvenile Hormone Studies in Drosophila melanogaster. Frontiers in Physiology 2022, 12. [Google Scholar] [CrossRef]
Figure 1.
Ovary activation in Zabrotes subfasciatus occurs during the pharate-adult stage, prior to adult emergence. The ovaries (except that in C) were embedded in Historesin, sectioned to a thickness of 5 μm, and stained with Harris’ hematoxylin and eosin [55]. Micrographs were captured using a Nikon Eclipse 80i microscope equipped with a digital camera and Nis-Element 3.1 imaging software. For the image in (C), ovary was processed for 4’,6-diamidino-2-phenylindole (DAPI) and phalloidin conjugated to Alexa 546 and the micrograph captured in a confocal microscope C2+ Nikon. A) The diagram illustrates the ovary of an adult insect. B) and C) Histological and confocal images display a telotrophic ovariole highlighting nutritive cords. D-G: Representative images of females and ovariole sections; D) white-eyed pupa; E) and F) final pharate-adult phase; G) one-day-old adult female.
Figure 1.
Ovary activation in Zabrotes subfasciatus occurs during the pharate-adult stage, prior to adult emergence. The ovaries (except that in C) were embedded in Historesin, sectioned to a thickness of 5 μm, and stained with Harris’ hematoxylin and eosin [55]. Micrographs were captured using a Nikon Eclipse 80i microscope equipped with a digital camera and Nis-Element 3.1 imaging software. For the image in (C), ovary was processed for 4’,6-diamidino-2-phenylindole (DAPI) and phalloidin conjugated to Alexa 546 and the micrograph captured in a confocal microscope C2+ Nikon. A) The diagram illustrates the ovary of an adult insect. B) and C) Histological and confocal images display a telotrophic ovariole highlighting nutritive cords. D-G: Representative images of females and ovariole sections; D) white-eyed pupa; E) and F) final pharate-adult phase; G) one-day-old adult female.
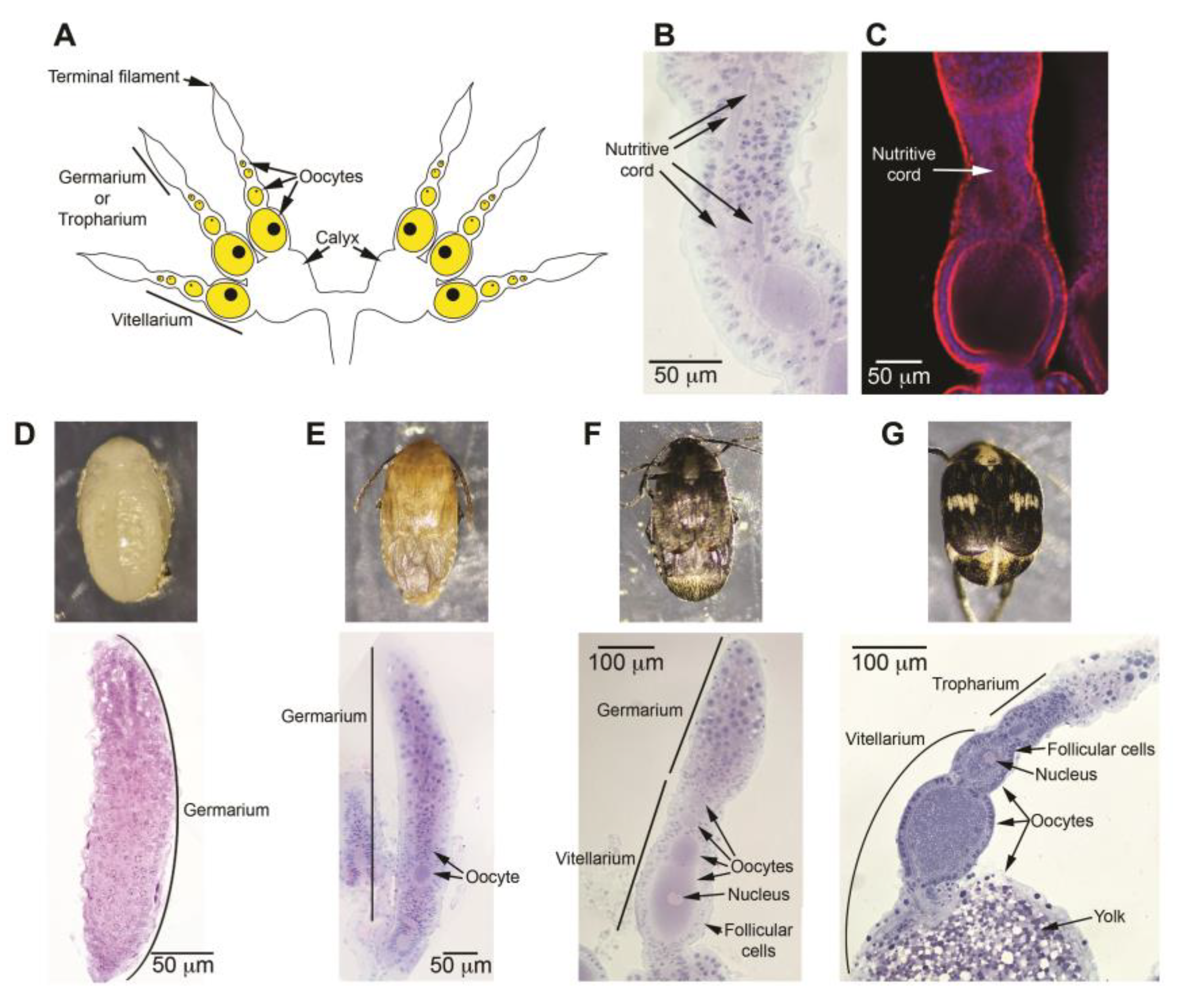
Figure 2.
Impact of host seed availability and mate presence on oviposition behaviour and fecundity of Zabrotes subfasciatus females. Eighteen newly emerged females per experimental group were individually placed in acrylic containers. Group A: females only; Group B: Females and males; Group C: females and seeds; Group D: females, males, and seeds. The number of laid eggs was registered daily as well as the number of adults emerged from each of the laid egg during the 10 days of the oviposition period. A) Oviposition and adult emergence profiles of the females from experimental group D: The number of emerged adults represents the individuals that developed from eggs laid on each specific day during the oviposition period. Different bold letters on the bars indicate statistically significant differences in the mean number of eggs laid during the oviposition period and different non-bold letters indicate statistically significant differences in the mean number of emergents (comparing the means of the different days). Two-way repeated measures ANOVA, followed by Šídák’s multiple comparisons test, **p < 0.01. B) Total number of eggs laid per female during the oviposition period (mean + SD). Ordinary one-way ANOVA followed Tukey’s multiple comparisons test, *p < 0.0001. C) Heatmap depicting the mean number of eggs laid per female in each of the 10-day oviposition period. D) Oviposition profile per female during the 10-day oviposition period (mean + SD). For statistical details, see Table S1, Table S2, Table S3 and Table S4.
Figure 2.
Impact of host seed availability and mate presence on oviposition behaviour and fecundity of Zabrotes subfasciatus females. Eighteen newly emerged females per experimental group were individually placed in acrylic containers. Group A: females only; Group B: Females and males; Group C: females and seeds; Group D: females, males, and seeds. The number of laid eggs was registered daily as well as the number of adults emerged from each of the laid egg during the 10 days of the oviposition period. A) Oviposition and adult emergence profiles of the females from experimental group D: The number of emerged adults represents the individuals that developed from eggs laid on each specific day during the oviposition period. Different bold letters on the bars indicate statistically significant differences in the mean number of eggs laid during the oviposition period and different non-bold letters indicate statistically significant differences in the mean number of emergents (comparing the means of the different days). Two-way repeated measures ANOVA, followed by Šídák’s multiple comparisons test, **p < 0.01. B) Total number of eggs laid per female during the oviposition period (mean + SD). Ordinary one-way ANOVA followed Tukey’s multiple comparisons test, *p < 0.0001. C) Heatmap depicting the mean number of eggs laid per female in each of the 10-day oviposition period. D) Oviposition profile per female during the 10-day oviposition period (mean + SD). For statistical details, see Table S1, Table S2, Table S3 and Table S4.
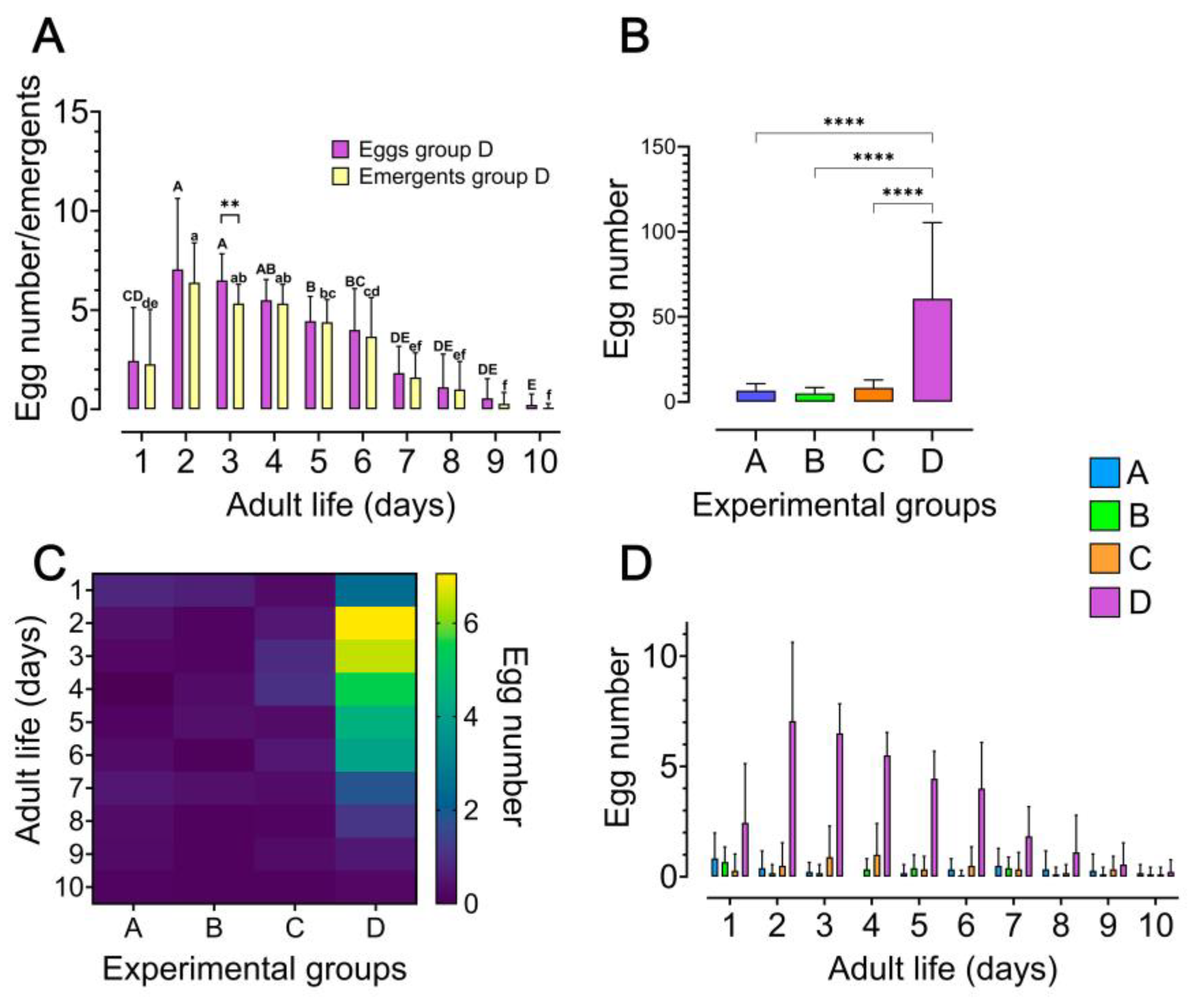
Figure 3.
Impact of host seed availability and mate presence on the ovary activation of adult Zabrotes subfasciatus females. Eighteen newly emerged females per experimental group were individually placed in acrylic containers. Group A: females only; Group B: Females and males; Group C: females and seeds; Group D: females, males, and seeds. The upper images depict representative photographs of ovarioles with oocytes in different levels of development (A, Stage I ovarioles) and an ovary with fully developed oocytes (B, Stage II ovarioles). Ten females from each experimental group were dissected and the activation level of their ovarioles was registered as corresponding to Stage I (C) or Stage II (D). Different letters on the columns with values from the same experimental group (colour) over the time studied indicate statistically significant differences (p < 0.05). Data are presented as mean + SD. Fisher’s exact test, *p < 0.01 and ***p < 0.001. For statistical details, see Table S5, Table S6 and Table S7, and S8.
Figure 3.
Impact of host seed availability and mate presence on the ovary activation of adult Zabrotes subfasciatus females. Eighteen newly emerged females per experimental group were individually placed in acrylic containers. Group A: females only; Group B: Females and males; Group C: females and seeds; Group D: females, males, and seeds. The upper images depict representative photographs of ovarioles with oocytes in different levels of development (A, Stage I ovarioles) and an ovary with fully developed oocytes (B, Stage II ovarioles). Ten females from each experimental group were dissected and the activation level of their ovarioles was registered as corresponding to Stage I (C) or Stage II (D). Different letters on the columns with values from the same experimental group (colour) over the time studied indicate statistically significant differences (p < 0.05). Data are presented as mean + SD. Fisher’s exact test, *p < 0.01 and ***p < 0.001. For statistical details, see Table S5, Table S6 and Table S7, and S8.
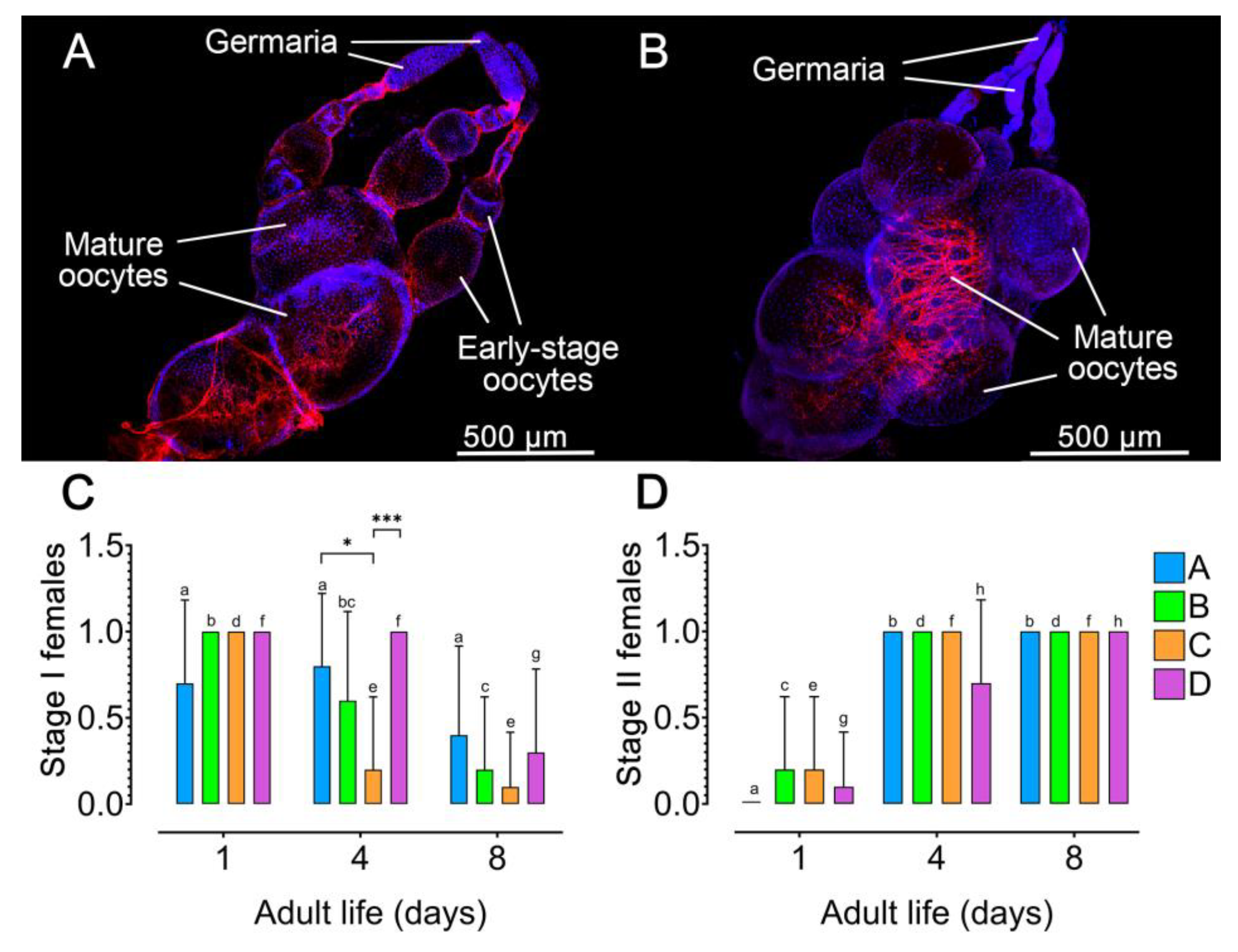
Figure 4.
Impact of host seed availability and mate presence on the expression of vitellogenic genes in Zabrotes subfasciatus females. Eighteen newly emerged females per experimental group were individually placed in acrylic containers. Group A: females only; Group B: Females and males; Group C: females and seeds; Group D: females, males, and seeds. Tissues from adult females were dissected and 4 RNA pools from 5 individuals were subjected to RT-qPCR (whole body of pupae were processed). Raw data were first normalized to the reference gene (rpl32) and the relative quantities of transcripts were calculated using the 2-ΔΔCt method with efficiency correction and a control sample for calibration [59] (Applied Biosystems User bulletin #2). Y axes: 2^–ΔΔCt values. Comparisons with the pupal stage, as well as between experimental groups and days, were performed using the Mann Whitney and test Kruskal-Wallis’s test followed by Dunn’s multiple comparisons test. Median and interquartile range (25th-75th percentiles), with minimum and maximum values; *p < 0.05; **p < 0.01. A) Vg gene transcript levels in carcass tissues, which include fat body. B) VgR gene transcript levels in carcass tissues. C) Vg gene transcript levels in ovary tissues. D) VgR gene transcript levels in ovary tissues. For statistical details, see Table S9, Table S10, Table S11, Table S12, Table S13, Table S14, Table S15 and Table S16.
Figure 4.
Impact of host seed availability and mate presence on the expression of vitellogenic genes in Zabrotes subfasciatus females. Eighteen newly emerged females per experimental group were individually placed in acrylic containers. Group A: females only; Group B: Females and males; Group C: females and seeds; Group D: females, males, and seeds. Tissues from adult females were dissected and 4 RNA pools from 5 individuals were subjected to RT-qPCR (whole body of pupae were processed). Raw data were first normalized to the reference gene (rpl32) and the relative quantities of transcripts were calculated using the 2-ΔΔCt method with efficiency correction and a control sample for calibration [59] (Applied Biosystems User bulletin #2). Y axes: 2^–ΔΔCt values. Comparisons with the pupal stage, as well as between experimental groups and days, were performed using the Mann Whitney and test Kruskal-Wallis’s test followed by Dunn’s multiple comparisons test. Median and interquartile range (25th-75th percentiles), with minimum and maximum values; *p < 0.05; **p < 0.01. A) Vg gene transcript levels in carcass tissues, which include fat body. B) VgR gene transcript levels in carcass tissues. C) Vg gene transcript levels in ovary tissues. D) VgR gene transcript levels in ovary tissues. For statistical details, see Table S9, Table S10, Table S11, Table S12, Table S13, Table S14, Table S15 and Table S16.
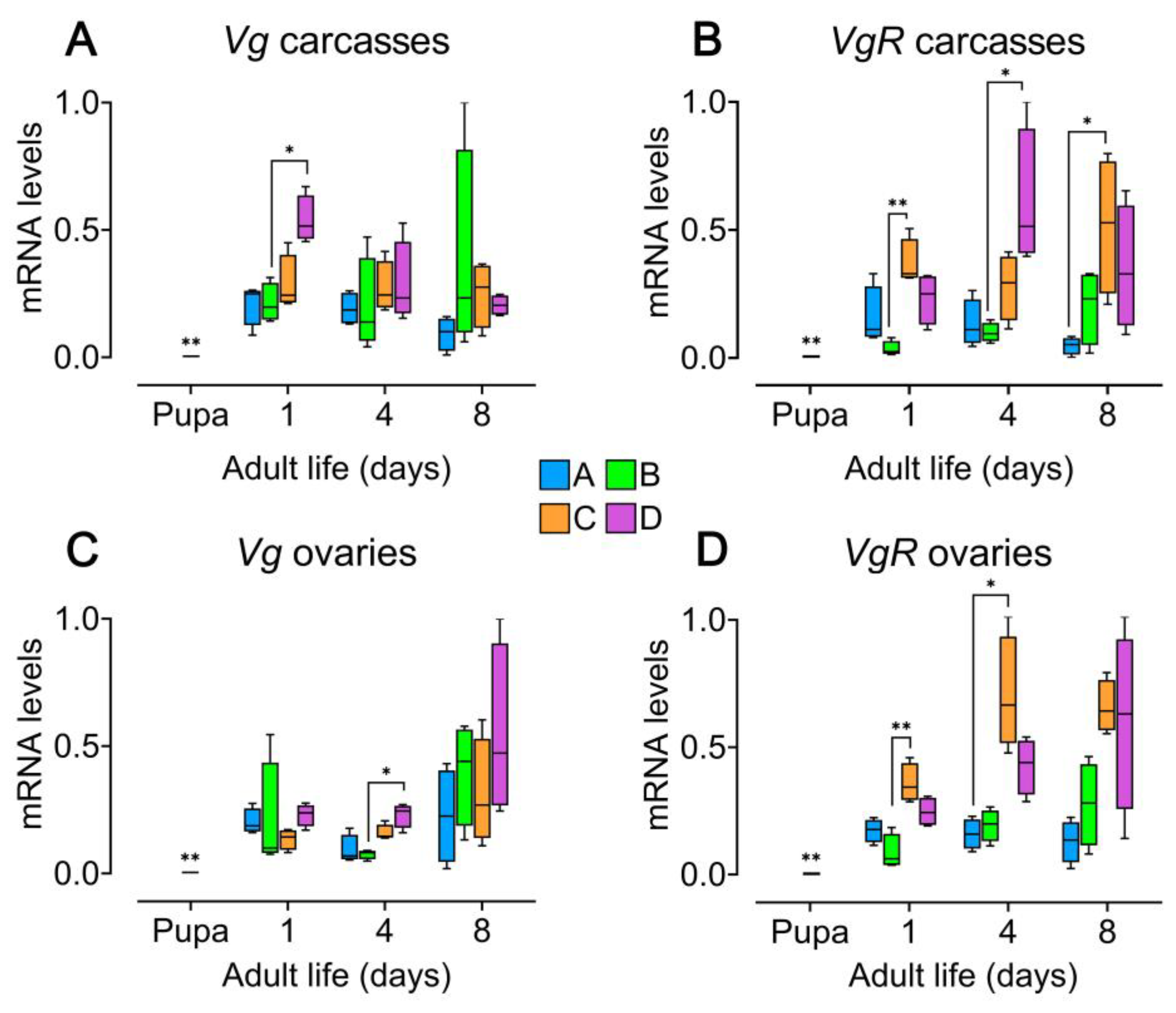
Figure 5.
Proposed working model of oviposition regulation in the bean weevil Zabrotes subfasciatus. The proposed increase in juvenile hormone (JH) synthesis following female exposure to host seeds, as well as the chemical nature of the sex pheromone involved, require experimental validation. CA= corpora allata; FB= fat body; Ov= ovaries; JH= juvenile hormone; Vg= vitellogenin; VgR= vitellogenin receptor.
Figure 5.
Proposed working model of oviposition regulation in the bean weevil Zabrotes subfasciatus. The proposed increase in juvenile hormone (JH) synthesis following female exposure to host seeds, as well as the chemical nature of the sex pheromone involved, require experimental validation. CA= corpora allata; FB= fat body; Ov= ovaries; JH= juvenile hormone; Vg= vitellogenin; VgR= vitellogenin receptor.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



