Submitted:
18 July 2025
Posted:
18 July 2025
You are already at the latest version
Abstract
In this study, the invasive orange spiny whitefly (“OSW”; Aleurocanthus spiniferus) and a species of Encarsia parasitising its puparia were studied in three different areas of the province of Modena (Emilia-Romagna, northern Italy): a pear orchard in Bomporto, an organic pear orchard in Carpi, and the semi-natural botanical garden "La Pica" in San Felice sul Panaro. The material of both species was collected for taxonomic and molecular studies. The abundance of OSW and parasitoid activity was surveyed in whitefly puparia, with a focus on the botanical garden. A total of 1,800 leaves of Malus domestica, Pyrus communis and Vitis vinifera were sampled to assess whitefly infestation. The results showed a significant variation in the abundance of A. spiniferus puparia, with the highest infestation observed in V. vinifera. Morphological and molecular analyses, including mitochondrial COI and rRNA 16S for whiteflies and COI and 28S for parasitoids, confirmed the identity of A. spiniferus and identified the parasitoid as Encarsia nipponica. Molecular data also revealed the presence of three haplotypes of A. spiniferus, including a haplotype from China new to Europe. The parasitisation rate by E. nipponica was low (0.015%). Our results update the distribution of A. spiniferus and E. nipponica in Italy, providing insights into their ecological interactions and potential implications for pest management strategies.
Keywords:
haplotype diversity
; host–parasitoid interactions
; biological control potential
; Aleyrodidae
; Aphelinidae
; grapevine agroecosystems
1. Introduction
Aleurocanthus Quaintance & Baker, 1914 (Hemiptera: Aleyrodidae) includes 93 species, predominantly found in tropical and subtropical regions [1] Due to their widespread distribution and considerable potential threat to crops, all species within this genus are classified as quarantine pests by the European Union [2].
Among these species, Aleurocanthus spiniferus (Quaintance, 1903), commonly known as the orange spiny whitefly (OSW), is one of the most damaging worldwide and is particularly notable for its polyphagous habits. Native to Asia, OSW has spread to most tropical and subtropical regions, including Africa, Australia, and the Pacific Islands (Hawaii)[1,3,4,5,6]. More recently it has entered Europe, first reported in Italy in 2008 [7], followed by reports in Croatia (2012) [8], Montenegro (2013) [9], Greece (2016) [10], Albania (2018) [11], and France (2023) [12]. In Italy, the species was initially detected in the Lecce district of the Apulia region [7] and has since been reported from eleven regions: Basilicata, Calabria, Campania, Emilia-Romagna, Lazio, Liguria, Lombardy, Marche, Tuscany and Sicily [6,7,8,9,10,11,12,13].
In 2018, the Emilia-Romagna phytosanitary service, a prominent fruit producing region in northern Italy, especially for the cultivation of pear and grapevines, detected a new pest for this region, initially identified as a species of Aleurocanthus [13] and subsequently ascribed to A. spiniferus. In the following years, more outbreaks were reported in different parts of the region. Accurate species identification within this genus is complicated by polyphenism, in which morphological traits vary depending on the host plant, requiring both taxonomic and molecular approaches for precise identification [1,2,3,4,5,6,7,8,9,10,11,12,13,14,15,16].
Aleurocanthus spiniferus infests approximately 90 host plant species, including several economically significant crops, with a notable preference for Citrus spp. and grapes (Vitis vinifera L.) [14,15,16,17]. Infestations can weaken plants by direct sap loss and indirect damage caused by honeydew production, which promotes sooty mould and alters photosynthesis [11]. As a quarantine pest, OSW also impacts international trade by hindering the export of fruits and plants, which poses a significant threat to European agriculture.
As it is a quarantine pest, the phytosanitary practices that can be applied for the containment of OSW outbreaks vary according to the areas affected (nurseries, private gardens, public green areas). Integrated pest management (IPM) strategies for the management of OSW in agricultural crops include the use of soft soaps, wetting agents, and approved synthetic insecticides, depending on regional authorisations. However, these methods often provide incomplete control and can negatively affect biocontrol agents in the agroecosystem concerned, leading to secondary pest outbreaks [18]. Several parasitoids of A. spiniferus have been documented worldwide [19,20], primarily within the chalcidoid family Aphelinidae. Biological control efforts have been successful globally, using parasitoids such as Encarsia, including E. perplexa Huang & Polaszek [as E. opulenta (Silvestri)], E. smithi (Silvestri), and Amitus. [21,22,23], with some cases reaching mean parasitism rates of around 80% [24]. However, due to the European "Habitats Directive" (Council Directive 92/43/EEC, 1992), the use of exotic biocontrol agents is restricted in European countries.
Recent surveys in several central and southern Italian regions (Apulia, Campania, Latium, Marche and Sicily) identify Eretmocerus iulii Laudonia and Melone as parasitoid of A. spiniferus [15,16,17,18,19,20,21,22,23,24,25]. Eretmocerus iulii showed parasitisation rates ranging from 4.07% to 71.43% [15]. Given the threat posed by OSW, more research is needed in northern Italy to assess the impact of the pest and the role of biocontrol agents.
This study aims to confirm the identity of the Aleurocanthus species that occur in the Emilia-Romagna region by morphological and molecular analysis, evaluate its level of infestation in common crop species in Northern Italy, and assess the presence and role of naturally occurring parasitoids, identifying them at the species level using both morphological and molecular methods.
2. Materials and Methods
2.1. Sampled Areas
The collection of samples was carried out at three different sites in the province of Modena (Emilia-Romagna region, Italy), respectively represented by the edge of a pear orchard located in the municipal area of Bomporto (44.73164076133763, 11.039053971141435), by an organic pear orchard in the municipality of Carpi (44.727952774732834, 10.874274022562624) and by the Botanical Garden 'La Pica' in San Felice sul Panaro (44.87540090470623, 11.097329719787881).). The latter site is a seminatural area with the presence of agricultural and ornamental plants, covering an area of approximately 21,000 m2 and characterised by the presence of over 1,000 plant species from all over the world.
Puparia (4th instar nymphs) of OSW were collected from all three sites (Table 1) for taxonomic studies. In the botanical garden “La Pica”, a whitefly puparia survey was also carried out in different host plant species and specimens were collected to assess levels of parasitisation. For the latter study, both Aleurocanthus sp. and its parasitoid samples were preserved in 100% ethanol after collection, prior to morphological and molecular identification.
2.2. Morphological Study of Aleurocanthus sp.
As is the accepted standard for morphological studies on whiteflies, observations were carried out on puparia, which are considered to have better documented characters than adults. To this end, puparia of Aleurocanthus sp. collected from all three sampling areas were treated with cold 10% KOH until they turned light brown; they were then rinsed in 70% ethanol for 5 minutes, transferred to glacial acetic acid for 5 minutes, clove oil for 15 minutes and mounted in Canada balsam. The prepared slide-mounted specimens were observed using differential interference contrast (DIC) on a compound microscope.
2.3. Field Survey
The survey mentioned above in the botanical garden 'La Pica' was carried out in July 2022 on 20 plants selected for each of the following species: Malus domestica (Suckow) Borkh., Pyrus communis L. and Vitis vinifera L. In each of the 60 selected plants, 30 randomly selected leaves were visually sampled, making a total of 600 leaves for each plant species, to quantify the level of infestation by Aleurocanthus sp. puparia on the three different species.
The evaluation of the parasitisation rate was carried out exclusively on 10 pear plants of the Abate fetél variety (the most common pear variety in the Emilia Romagna region), from which a total of 80 leaves were collected. Each leaf was kept in a 9 cm diameter Petri dish and all the material was stored outdoors in the facilities of the botanical garden, protected from sunlight, under the porch of the visitor centre. Each sample / Petri dish was checked every two days for possible parasitoid emergence.
2.4. Data Analysis
Parasitisation impact on host populations [26], which describes the efficacy of a parasitoid in reducing a pest (parasitism level), was calculated as the number of parasitoids emerged / number of OSW pupae collected * 100.
To analyse the effect of tree species on Aleurocanthus sp. abundance (measured as the number of whitefly puparia per leaf), a Generalised Linear Mixed Model (GLMM) was fitted using the glmm TMB package in R [27]. The model was specified with a negative binomial distribution to account for overdispersion in the count data. The response variable in the model was the number of Aleurocanthus sp. puparia per leaf, and the fixed effect included in the model was the tree species (species). The structure of the random effect included random intercepts for each plant (plant) and each leaf nested within the plant (leaf), to account for the hierarchical structure of the data, where multiple leaves were sampled from the same plant and multiple plants were sampled within each species. Before fitting the GLMM model, summary statistics were calculated for the number of Aleurocanthus sp. puparia observed on the leaves of different tree species. All statistical analyses were performed with R 4.4.1[28]. For each test, α = 0.05.
2.5. Morphological Study of Parasitoids
Specimens were mounted on microscope slides after Noyes (1982) [29], but without maceration in 10% KOH after DNA extraction. However, this is not a general recommendation, as in many cases proteinase K /ATL buffer extraction leaves behind rhodopsin or other eye pigments, which require further maceration in KOH. The samples were then washed in distilled water for 1 hour and dehydrated for 5 minutes in graded ethanol of the following concentrations: 35%, 70%, 85%, 100%. After being cleared in clove oil and allowing alcohol evaporation, samples were dissected in Canada balsam. The wings, antennae, head, and remaining body parts were mounted separately on a single slide. In total, 22 females and no males, from 18 samples / petri dishes, were examined. Specimens examined (Figure 3), different structures were scanned with an Olympus BX63 microscope with DIC. The image sections were stacked and combined using Helicon Focus software, and the final images were edited with Adobe Photoshop CC®.
2.6. Molecular Analysis
To confirm the identification of the species according to morphological characteristics, some of the collected specimens of the whitefly and its parasitoid were stored at -20 ° C until they were used for molecular analysis. Genomic DNA was extracted using QuickExtractTM DNA Extraction Solution (Lucigen, Middleton, WI, USA), according to the manufacturer’s protocol. As this is a non-destructive method, the samples were immediately washed with 100 μL of distilled water at the end of the extraction procedure, then replaced with 100 μL of 100% ethanol and stored as vouchers at -20 ° C at UNIMORE. A negative extraction control was also performed with all the kit solutions but without insect tissue to check for contamination.
The genetic diversity of the examined species was analysed using mitochondrial genes cytochrome oxidase subunit I (COI) and rRNA 16S (16S) for whitefly samples and using the COI and the nuclear 28S gene for parasitoids. The markers were chosen due to their high variability and the large number of previously scored samples (CIT). In whiteflies, PCR amplification of a portion of the COI gene was carried out using primers AsFmik (5’-GTG TCC CAT TTA ATT AGT AGA GA-3’) and AsRmik (5’-GAG CCA TAA TAA AAG ACT CCA TC-3’; [30], with the protocol described in Uesugi et al. (2016) [31]. A portion of the 16S gene was also amplified for whiteflies with the primer pair 16Sbr (5’-CCG GTC TGA ACT CAG ATC ACG T-3’) and 16 Sar (5’-CGC CTG TTT AAC AAA AAC AT-3’[32]; with the following protocol: 35 cycles with 30 s at 94 °C, 30 s at 48 °C, and 30 s at 72 °C, with a final elongation step at 72 °C for 7 min. For parasitoids, a portion of the COI gene was amplified using primers LCO-1490 (5′-GGT CAA CAA ATC ATA AAG ATA TTG G-3′; Folmer et al. 1994) and C1-N-2191 (5’-CCC GGT AAA ATT AAA ATA TAA ACT TC-3’; [32], with a step-up procedure, with the following protocol: the initial five cycles were performed with 1 min at 94 °C, 1.5 min at 42 °C and 1.5 min at 72 °C, and they were followed by 35 cycles with 1 min at 94 °C, 1.5 min at 50 °C and 1 min at 72 °C, with a final elongation step at 72 °C for 7 min. The 28S rDNA D2 and D3 expansion regions were also amplified in parasitoids using the primer pair. D23F (5′-GAG AGT TCA AGA GTA CGT G-3′; [33] and 28Sb (a.k.a. D3B; 5′-TCGGAAGGAACC AGC TAC TA-3′, [34], with the protocol described Polaszek et al [35].
The amplified products were gel purified using the Wizard Gel and PCR cleaning kit (Promega) kit. Then both strands were subjected to sequencing reactions using the Big Dye Terminator 1.1 kit (Applied Biosystems, Foster City, CA, USA) and sequenced using an ABI Prism 3100 sequencer (Applied Biosystems). Nucleotide sequences of the newly analysed samples were submitted to GenBank (accession numbers: ZZ000000-ZZ000000 for COI; KK000000-KK000000 for 16S; and WW000000-WW000000 for 28S). Chromatograms were checked for the presence of ambiguous bases and COI sequences were translated to amino acids using the invertebrate mitochondrial code implemented in MEGA11 [36] to check for the presence of stop codons and, therefore, pseudogenes. The nucleotide sequences were aligned with the Clustal algorithm implemented in MEGA11 (pairwise and multiple alignment parameters: Gap opening penalty: 15, Gap extend penalty: 6.66) and checked by visual inspection.
Both newly analysed and reference sequences of COI and 16S for whiteflies and COI and 28S for GenBank parassitoid samples (see Supplementary Material Table S1) were included in the analysis, to provide better qualitative information for possible invasion pathways. Uncorrected p distances between scored haplotypes were determined using MEGA11, after checking that scored sequences were not subject to saturation using the programme DAMBE [37]. The relationships among haplotypes were estimated using a parsimony network by applying the method described by Templeton et al. (1992) [38], as implemented in TCS 1.21 [39] and visualised using tcsBU [40].
2.7. Phylogenetic Analysis
Six sequences of 28S D2 ribosomal DNA captured from Encarsia nipponica were combined with previously published sequence data from 75 outgroup taxa (Table X). Outgroups were comprised of a broad sampling of species-groups of Encarsia, Coccophagus Westwood, Coccophagoides Girault, Pteropterix Westwood, Aphytis Howard. Sequences were aligned using the E-INS-I algorithm in MAFFT v7.490 [41]. Alignments were trimmed using spruceup with cut-off values of 0.99, 0.97, 0.95 and 0.9 [42]. Phylogenetic reconstruction was performed using maximum likelihood in IQ-TREE v. 2.2.2.6, implementing a General Time Reversible model with invariant sites and gamma-distributed rate variation (-m GTR + I + G) [43]. Ten independent iterations of maximum likelihood were run with each trimmed data set (--runs 10). Bootstrap support was estimated from 1000 bootstrap trees constructed using ultrafast bootstrapping (-b 1000) [44].
3. Results
3.1. Morphological Study of Aleurocanthus sp.
Puparia collected in Emilia Romagna belong to the genus Aleurocanthus on the basis of the presence of many stout dorsal spines, of a white marginal waxy fringe and of earlier instar exuviae maintained in a stack on the dorsum. For this genus, a globally valid key for species identification based on morphological characters is currently not available. Thus, the samples collected in the present study have been identified using multiple valid keys for various local faunas, such as those provided by Martin (1999)[45], Dubey and Sundararaj (2004)[46], Dubey and Ko (2012)[47], Jansen and Porcelli (2018)[48], as well as by comparison with material stored in the CR collection, at the University of Catania, Italy.
The samples studied were ascribed to Aleurocanthus spiniferus, based on their overall shape and the width of their wax secretion (usually broad, ranging from almost 20 to 30% of puparial width) (Figure 1: a, b), their dorsal spine characteristics (with the third posterior-most pair of submarginal spines always single) (Figure 1: b), the overall shape of the posterior part of the abdomen (including the aspect, position and size of the vasiform orifice), with the VIII abdominal tergite around 70 μm (Figure 1: c), and the shape and dimensions of marginal teeth (never less than 6 per 0.1 mm and usually exceeding 200 units in their total number) (Figure 1: d).
3.2. Field Survey
On the 1,800 leaves collected (600 leaves per species), 2,701 A. spiniferus puparia were counted, from which 40 parasitoid samples emerged, all belonging to a single species and with a parasitoid impact on the host population of 0.015%.
The median number [IQR] of A. spiniferus individuals is reported for each species (Table 2, Figure 2). Data indicate considerable variation in the abundance of A. spiniferus between the three host plant species. Vitis vinifera had the highest median number of individuals of A. spiniferus per leaf (16 [8,27]), followed by Pyrus communis (4 [1,8]), and Malus domestica (1 [0,2]).
The results of the GLMM model indicate a significant effect of the host tree species on the abundance of A. spiniferus (Table 2, Figure 2). Specifically, the Pyrus communis and Vitis vinifera species had significantly higher numbers of A. spiniferus puparia compared to the reference species (Table 2). The random effects of the model showed negligible variance at the leaf level (variance=2.77e-09; Std. Dev. = 0.00005), suggesting that most of the variability in A. spiniferus counts was due to differences between plants (variance=0.129; Std. Dev. = 0.36) rather than between individual leaves.
3.3. Morphological Study of Parasitoids
Using the key to Chinese Encarsia by Huang and Polaszek (1998)[48], and by comparison with specimens in the collection of the Natural History Museum, London, slide-mounted specimens were identified as Encarsia nipponica Silvestri, 1927. The species can be diagnosed as follows: antennal formula 1,1,4,2 (Figure 3c); tarsal formula 5,5,5 (Fig 3a); fore wing with asetose area around stigmal vein and around the lower distal area; wing infuscate below marginal vein (Figure 3d); colour pattern distinctive, with the terminal antennal segment conspicuously darker than the rest of the antenna (Figure 3c); a broad dark band above the mouth opening, reaching to the lower eyes (Figure 3b); the following are dark in contrast to the rest of the body: pronotum, anterior mesoscutum, T1-T3, T5, and T7+8 of the metasoma (Figure 3a).
In Viggiani’s (1988)[49] key to Italian Encarsia, E. nipponica keys out as either E. partenopea (Masi) (currently a synonym of E. inaron (Walker)), or as E. pergandiella Howard, depending on the interpretation of the antenna as either having a 2-merous clava or an indistinct clava.
3.4. Phylogenetic Analysis
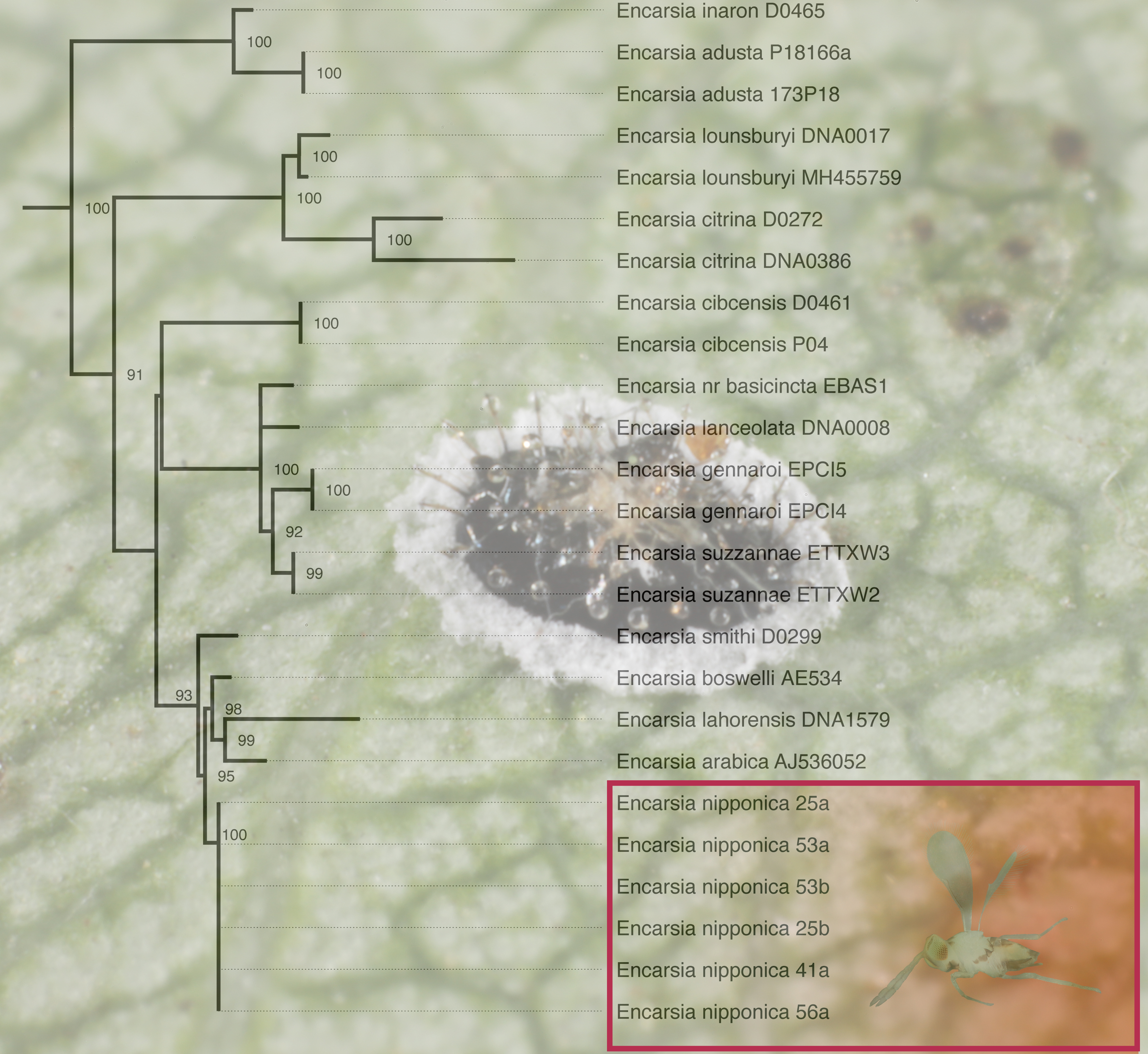
Five maximum likelihood trees were constructed based on the trimmed 28S D2 rDNA alignments produced from spruceup, with the untrimmed data set yielding the most likely overall phylogeny. While Encarsia was recovered as a strongly monophyletic group in all analyses, the backbone relationships therein are largely unsupported (Supplementary Material Table S2). Encarsia nipponica was always recovered in a strongly supported clade containing the boswelli, citrina, inaron, lahorensis, longifasciata, parvella, perflava, and smithi species-groups (Figure 4).
3.5. Molecular Analysis
Uncorrected p-distances of both genes assigned Emilia-Romagna specimens to Aleurocanthus spiniferus, thus confirming the morphological identification, with p-distances ranging from 0-21.1% for COI and from 0-15.7% for 16S (Supplementary Material Tables S3-S4). The haplotype network analysis (Figure 5) computed on the COI gene shows that the Emilia-Romagna specimens harbour three different haplotypes. Two of them had already been discovered in other Italian regions, in the Balkan region and in China, while a third one (red arrow, Figure 5), previously described for Chinese specimens, was discovered for the first time in Europe.
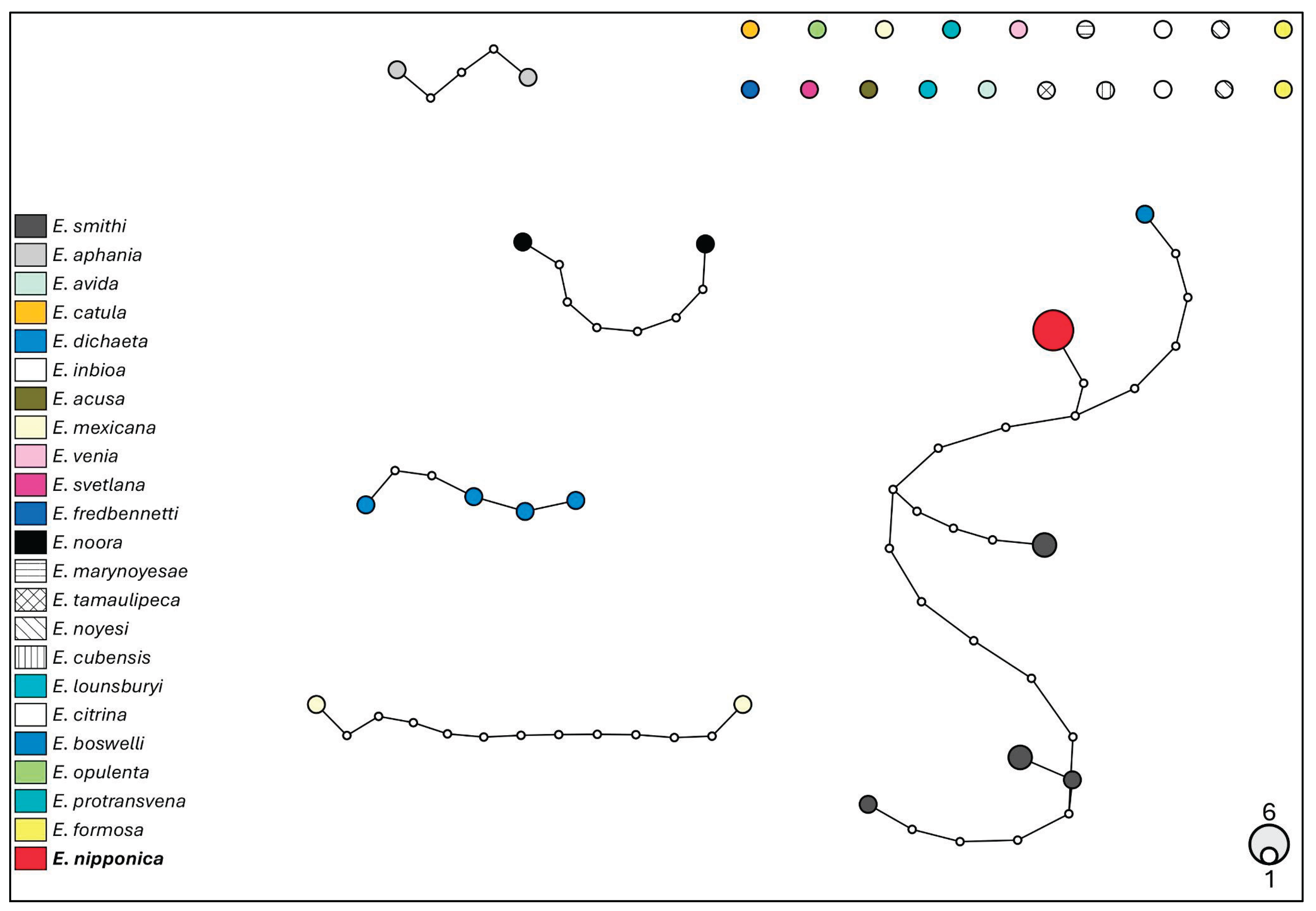
Molecular data indicate that parasitoids emerging from A. spiniferus in Emilia-Romagna belong to a distinct species of Encarsia, as shown by the COI gene analysis (Figure 7), with a p-distance not lower than 8.3% (Supplementary Material Table S5). The 28S gene is a less variable marker and shows that this species is more closely related to E. smithi (Silvestri, 1926) and E. boswelli (Girault, 1915) (Figue 6; p-distances ranging 1.4-2.5%; Supplementary Material Table S6). These findings strengthen the conclusion reported above to assign the species to E. nipponica based on morphological analysis.
Figure 5.
Haplotype network analysis computed on 657 bp of the COI gene of Aleurocanthus spiniferus. The circles denote the haplotypes, while the circle area represents the haplotype frequency. Small white circles indicate putative/missing haplotypes. Networks that fall below the value of the 95% connection limit are disconnected. The red arrow shows the new haplotype for Europe, found in Emilia-Romagna.
Figure 5.
Haplotype network analysis computed on 657 bp of the COI gene of Aleurocanthus spiniferus. The circles denote the haplotypes, while the circle area represents the haplotype frequency. Small white circles indicate putative/missing haplotypes. Networks that fall below the value of the 95% connection limit are disconnected. The red arrow shows the new haplotype for Europe, found in Emilia-Romagna.
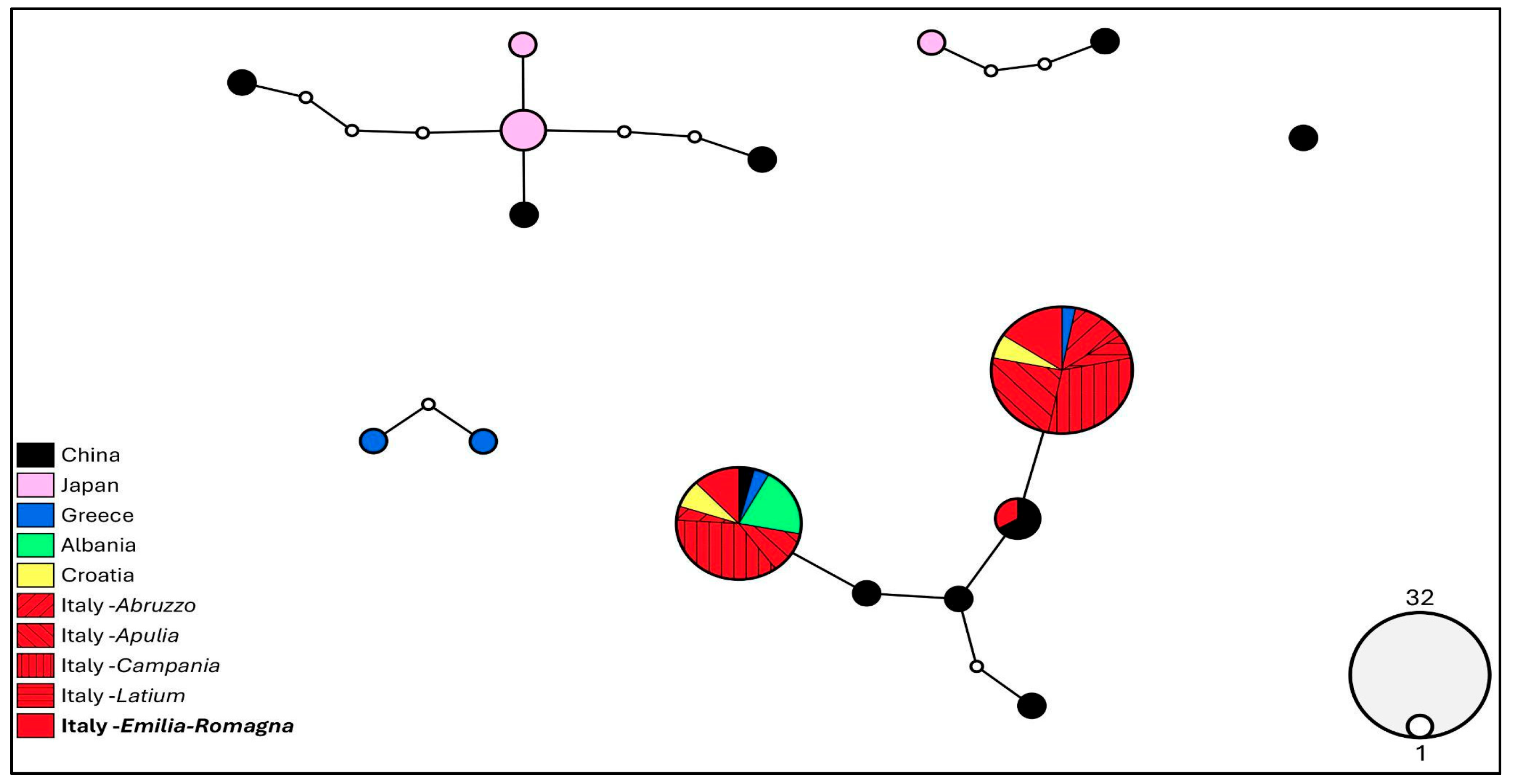
Figure 6.
Haplotype network analysis on 672 bp of the COI gene of Encarsia. Circles denote haplotypes, while circle area represents haplotype frequency. Small white circles indicate putative/missing haplotypes. Networks that fall below the value of the 95% connection limit are disconnected. The red arrow shows the haplotype found in Emilia-Romagna specimens.
Figure 6.
Haplotype network analysis on 672 bp of the COI gene of Encarsia. Circles denote haplotypes, while circle area represents haplotype frequency. Small white circles indicate putative/missing haplotypes. Networks that fall below the value of the 95% connection limit are disconnected. The red arrow shows the haplotype found in Emilia-Romagna specimens.
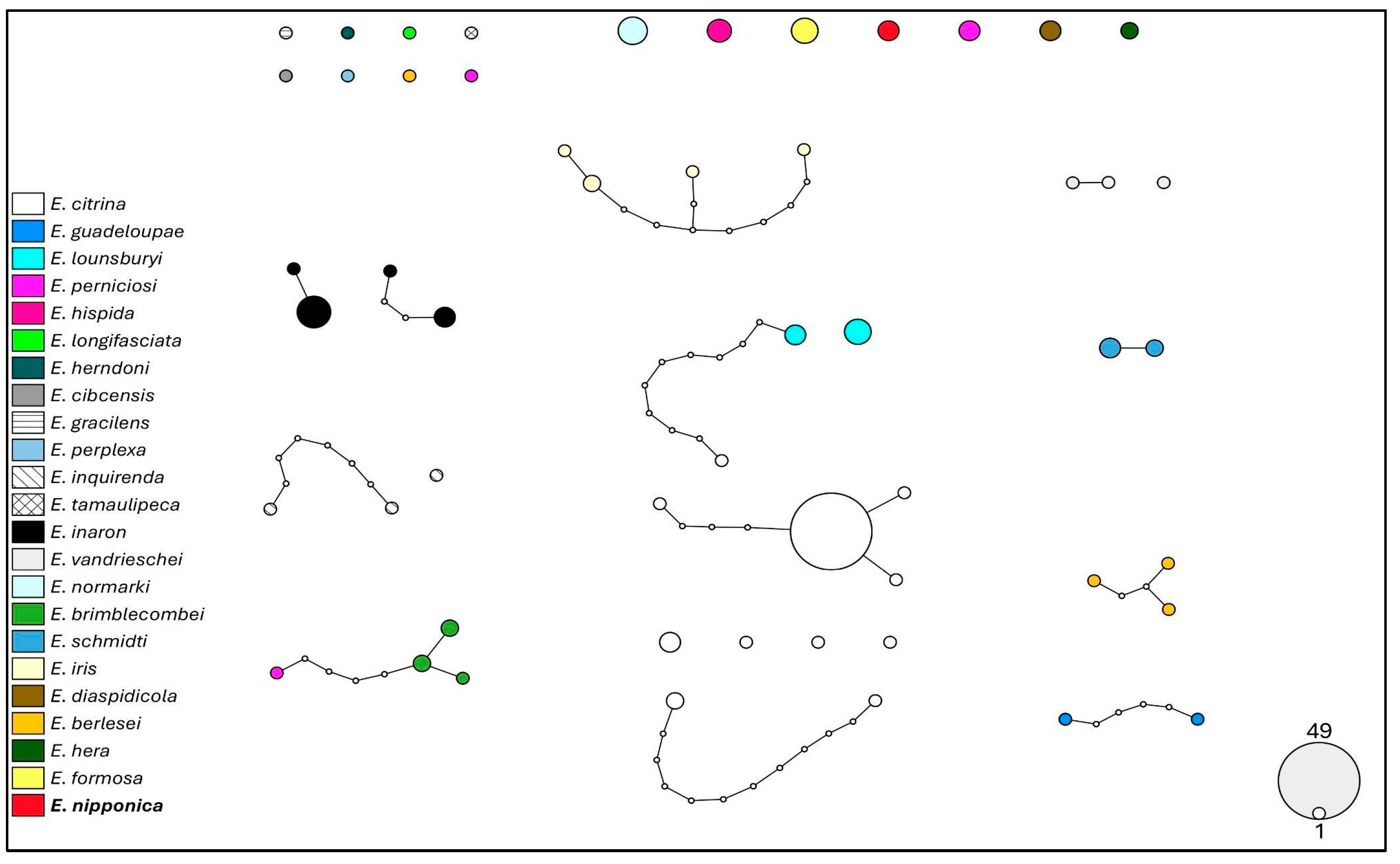
Figure 7.
Haplotype network analysis on 858 bp of the 28S gene of Encarsia. Circles denote haplotypes, while the circle area represents the frequency of the haplotype. Small white circles show putative/missing haplotypes. Networks that fall below the value of the 95% connection limit are disconnected. The red arrow shows the haplotype found in the Emilia-Romagna specimens.
Figure 7.
Haplotype network analysis on 858 bp of the 28S gene of Encarsia. Circles denote haplotypes, while the circle area represents the frequency of the haplotype. Small white circles show putative/missing haplotypes. Networks that fall below the value of the 95% connection limit are disconnected. The red arrow shows the haplotype found in the Emilia-Romagna specimens.
4. Discussion
The recent identification of Aleurocanthus spiniferus in the Emilia-Romagna region of Italy marks a significant development in the understanding of the distribution of this invasive species in Europe. The initial detection was made through careful morphological analysis, where the distinctive spiny pupal cases and the dark adult colour with white waxy secretions were key identifiers. These characteristics are essential to distinguish A. spiniferus from other species of whiteflies commonly found in the region. However, given the economic importance of the agricultural sector in Emilia-Romagna, which includes extensive viticulture and fruit production, morphological identification alone was deemed insufficient.
Molecular techniques were employed to ensure accurate identification, focussing on the mitochondrial cytochrome c oxidase I (COI) gene. The sequences obtained were compared with existing GenBank entries, confirming a high similarity with A. spiniferus populations from other Italian regions [11]. Furthermore, a new haplotype was also found, defined here as H5, raising concerns about the potential impact of the pest on local agriculture. H5 was previously found in Chinese specimens from Yichang, Hubei province [31]. The presence of A. spiniferus in Emilia-Romagna could be due to two non-mutually exclusive scenarios: a) natural expansion of the populations already present in other Italian regions and/or in Albania, Croatia and Greece (as indicated by the presence of haplotypes H1 and H2); b) newly introduced specimens from China (haplotype H5).
The impact of A. spiniferus on different host plants has been well documented in other regions, showing a wide host range that includes citrus, tea, and various ornamental plants. In Asia, where the pest is endemic, significant infestations of citrus crops have been reported, leading to substantial economic losses (USDA 1982; [10]. Similarly, in the newly invaded regions of southern Italy, including Calabria and Puglia, A. spiniferus has been found on citrus plants, grapevines and even ornamental plants, demonstrating its adaptability and potential to cause widespread damage across different plant species [11].
In Emilia-Romagna, A. spiniferus infestation varied significantly among the different host plants in the study area. In particular, the highest infestation rates were observed in grapevines and pear, while apple exhibited lower infestation levels. These findings are consistent with previous studies indicating a preference for A. spiniferus for certain host plants. For instance, Kapantaidaki et al. (2019)[10] noted a similar preference in Greece, where A. spiniferus was more prevalent in grapevine and citrus compared to other fruit trees. Nugnes et al .(2020)[11] also reported higher infestation rates on citrus and grapevine in southern Italy, suggesting that these plants may provide more favourable conditions for the pest's development. The reasons for this preference for the host plant may be multifactorial, involving aspects such as plant physiology, leaf surface characteristics, and chemical cues that influence the oviposition behaviour of A. spiniferus. Grapevine and pear trees can provide a more suitable environment for the development of the pest. However, the pest's ability to thrive on a wide variety of host plants suggests that its spread could have far-reaching consequences for different agricultural sectors. This adaptability highlights the importance of early detection and rapid response to prevent the establishment and spread of A. spiniferus in new areas. Therefore, the presence of A. spiniferus in Emilia-Romagna not only represents a new challenge for local farmers but also provides a case study in the spread and management of this invasive pest. The findings from this region will contribute to a broader understanding of A. spiniferus dynamics and inform future pest management strategies in Europe.
In addition, the present study marks the first documented occurrence of Encarsia nipponica in Europe, specifically as a parasitoid of A. spiniferus in the Emilia-Romagna region of Italy. This is the first detection of this species in Europe and the second discovery of a previously unrecorded species of Aphelinidae associated with A. spiniferus in the country, following the discovery of Eretmocerus iulii (serius group) earlier this year (Melone et al. 2024). This finding significantly expands the known geographic range of E. nipponica, which was previously reported mainly in Asian regions, including Japan and Taiwan [48]. This discovery is particularly important given the increasing incidence of A. spiniferus in Europe, especially in fruit producing regions [13,14].
Encarsia nipponica has historically been placed within the parvella species-group (Huang and Polaszek 1998)[48], however based on our assessment it appears to be better placed within the perflava species-group. The main characteristics of the E. perflava group are as follows: Fore wing with an asetose area around the stigma and in the lower distal area (Figure 3d); antenna with F1 and F2 much shorter than the remaining antennomeres, and without longitudinal sensilla (Figure 3c). The wing characters are shared by the closely related parvella and longifasciata groups.
Our phylogenetic analysis suggests that E. nipponica is aligned with taxa currently placed in the perflava, boswelli, lahorensis, smithi, and longifasciata species-groups. The relationship between these species-groups is poorly understood and inconclusively supported in our phylogenetic hypothesis. Ongoing investigations in the taxonomy and phylogenetics of the Encarsia species groups aim to better understand the limits between these species-groups.
The parasitisation rate observed in this study was relatively low, with a parasitism impact of 0.015% on the A. spiniferus population. This is substantially lower than the parasitism rates reported for other Encarsia species used in biological control programmes. For instance, Encarsia smithi has achieved parasitism rates of up to 80% in controlling A. spiniferus in tropical regions [24]. The low parasitisation rate observed here could be attributed to several factors, including the novelty of the host-parasitoid association in this geographical area, environmental conditions that may not yet favour E. nipponica, or competition with other parasitoid species.
Finally, more studies are needed to assess the long-term establishment and efficacy of E. nipponica in European ecosystems, as well as its potential interactions with native parasitoid species.
In conclusion, the first record of E. nipponica in Europe provides new insights into the biocontrol potential of A. spiniferus. Although the observed parasitisation rate was low, the establishment of this parasitoid could represent an important step forward in the integrated management of this invasive pest. Ongoing monitoring and further research will be crucial in understanding the dynamics of this new parasitoid-host interaction and to optimise its use in European agroecosystems.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Supplementary Table S1: GenBank sequences utilized for comparison. (NA: geographic origin not available); Supplementary Material Table S2: GenBank sequences used for comparison; Supplementary Material Table S3: Genetic distance values (p-distance) for the COI gene among Aleurocanthus species. The analysis was carried out on a dataset of 657 bp. Newly analyzed specimens are in bold; Supplementary Material Table S4: Genetic distance values (p-distance) for the 16S gene among Aleurocanthus species. The analysis was carried out on a dataset of 488 bp. Newly analyzed specimens are in bold; Supplementary Material Table S5: Genetic distance values (p-distance) for the COI gene among Encarsia species. The analysis was carried out on a dataset of 672 bp. Newly analyzed specimens are in bold; Supplementary Material Table S6: Genetic distance values (p-distance) for the 28S gene among Encarsia species. The analysis was carried out on a dataset of 858 bp. Newly analyzed specimens are in bold.
Author Contributions
Conceptualization, E.C., D.G. and M.C.; methodology, E.C., D.G., M.C., C.R., A.P. and R.L.K.; validation, C.R., A.P. and R.L.K.; formal analysis, E.C., M.C. and R.L.K.; investigation, E.C., D.G., M.C., C.R., A.P. and R.L.K.; resources, L.M.; data curation, E.C., D.G. and M.C.; writing—original draft preparation, E.C.; writing—review and editing, E.C., D.G., M.C., C.R., A.P., R.L.K. and L.M.; visualization, E.C., D.G. and M.C.; supervision, L.M.; project administration, L.M.; funding acquisition, L.M. All authors have read and agreed to the published version of the manuscript.
Funding
This work was partially funded by the National Recovery and Resilience Plan (NRRP), Mission 4 Component 2 Investment 1.4 – Call for tender No. 3138 of 16 December 2021, rectified by Decree n. 3175 of 18 December 2021 of the Italian Ministry of University and Research funded by the European Union – NextGenerationEU. Project Code [CN_00000033], Concession Decree No. 1034 of 17 June 2022 adopted by the Italian Ministry of University and Research, CUP E93C22001090001, Project title “National Biodiversity Future Center – NBFC”. EC was funded by the EU, FSE-REACT-EU, PON Research and Innovation 2014-2020 DM1062/2021.
Data Availability Statement
The original contributions presented in this study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
Acknowledgments
We thank Niccolò Patelli for collecting A. spiniferus and Elena Monari for performing DNA extractions. We are also grateful to Patrizia Casari, Adriano Cazzuoli and Enrico Gabrielli for providing the facilities of the La Pica Botanical Garden, and cooperation in sampling. The sixth author is a participant of the Oak Ridge Institute for Science and Education (ORISE) Agricultural Research Service (ARS) Research Participation Program, supported by the USDA-ARS Systematic Entomology Lab in Washington D.C., USA. The mention of trade names or commercial products in this article is solely for the purpose of providing specific information and does not imply recommendation or endorsement by the USDA. USDA is an equal opportunity provider and employer. This article was written without the use of artificial intelligence.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Martin, J.H. An identification guide to common whitefly pest species of the world (Homoptera: Aleyrodidae). Trop. Pest Manag. 1987, 33, 298–322. [Google Scholar] [CrossRef]
- EFSA Panel on Plant Health. Evaluation of a paper by Guarnaccia et al. (2017) on the first report of Phyllosticta citricarpa in Europe. EFSA J. 2018, 16, 1–48. [Google Scholar] [CrossRef]
- Van den Berg, M.A.; De Beer, M.S. Natural enemies of the spiny blackfly, Aleurocanthus spiniferus (Hem. : Aleyrodidae), in Mpumalanga, South Africa. In Proceedings of the International Society of Citriculture, Sun City Resort, South Africa, 12–17 May 1996; pp. 667–669. [Google Scholar]
- Nguyen, R.; Fasulo, T.R. A Citrus blackfly parasitoid, Encarsia perplexa Huang & Polaszek (Insecta: Hymenoptera: Aphelinidae). EDIS 2010. [Google Scholar] [CrossRef]
- Gillespie, P.S. A review of the whitefly genus Aleurocanthus Quaintance & Baker (Hemiptera: Aleyrodidae) in Australia. Zootaxa 2012, 3252, 1–61. [Google Scholar] [CrossRef]
- EPPO. Aleurocanthus spiniferus. Available online: https://gd.eppo.int/taxon/ALEUSI (accessed on 15 July 2024).
- Porcelli, F. First record of Aleurocanthus spiniferus (Homoptera: Aleyrodidae) in Apulia, Southern Italy. EPPO Bull. 2008, 38, 516–518. [Google Scholar] [CrossRef]
- Šimala, M.; Pintar, M.; Milek, T.M.; Markotić, V. Results of a two year survey (2015-2016) of quarantine whitefly species from genus Aleurocanthus Quaintance & Baker 1914 on citrus in Croatia. In Proceedings of the 13th Slovenian Conference on Plant Protection, Rimske Toplice, Slovenia, 7–8 March 2017. [Google Scholar]
- Radonjić, S.; Hrnčić, S.; Malumphy, C. First record of Aleurocanthus spiniferus (Quaintance) (Hemiptera Aleyrodidae) in Montenegro. Redia 2014, 97, 141–145. [Google Scholar]
- Kapantaidaki, D.E.; Antonatos, S.; Kontodimas, D.; Milonas, P.; Papachristos, D.P. Presence of the invasive whitefly Aleurocanthus spiniferus (Hemiptera: Aleyrodidae) in Greece. EPPO Bull. 2019, 49, 127–131. [Google Scholar] [CrossRef]
- Nugnes, F.; Laudonia, S.; Jesu, G.; Jansen, M.G.M.; Bernardo, U.; Porcelli, F. Aleurocanthus spiniferus (Hemiptera: Aleyrodidae) in some European countries: Diffusion, hosts, molecular characterization, and natural enemies. Insects 2020, 11, 42. [Google Scholar] [CrossRef]
- Streito, J.C.; Mendes, E.; Sanquer, E.; Strugarek, M.; Ouvrad, D.; Robin-Havret, V.; Poncet, L.; Lannou, C.; Rossi, J.P. Incursion Preparedness, Citizen Science and Early Detection of Invasive Insects: The Case of Aleurocanthus spiniferus (Hemiptera, Aleyrodidae) in France. Insects 2023, 14, 916. [Google Scholar] [CrossRef]
- Bariselli, M.; Bortolotti, P.P.; Nannini, R. Scheda 27 - Aleurocanthus spiniferus. In Schede tecniche per il riconoscimento degli organismi nocivi da quarantena; Regione Emilia-Romagna, Assessorato Agricoltura, Caccia e Pesca, Servizio fitosanitario Emilia-Romagna: Bologna, Italy, 2019; p. 4. [Google Scholar]
- Rapisarda, C.; Longo, S. First report from Sicily (Italy) of the orange spiny whitefly, Aleurocanthus spiniferus (Quaintance) (Hemiptera: Aleyrodidae), and its potential risk for the Italian citrus industry. EPPO Bull. 2021, 51, 329–332. [Google Scholar] [CrossRef]
- Melone, G.; Ascolese, R.; Nugnes, F.; Porcelli, F.; Rapisarda, C.; Farina, A.; Picciotti, U.; Garganese, F.; Laudonia, S. An Eretmocerus species, parasitoid of Aleurocanthus spiniferus, was found in Europe: The secret savior of threatened plants. Sustainability 2024, 16, 2970. [Google Scholar] [CrossRef]
- Gill, R.J. The morphology of whiteflies. In Whiteflies: Their bionomics, pest status and management; Gerling, D., Ed.; Intercept: Andover, Hants, UK, 1990; pp. 13–46. [Google Scholar]
- Mound, L.A.; Halsey, S.H. Whitefly of the World; British Museum (Natural History) and John Wiley & Sons: Chichester, UK, 1978; p. 340. [Google Scholar]
- Cioffi, M.; Cornara, D.; Corrado, I.; Jansen, M.G.M.; Porcelli, F. The status of Aleurocanthus spiniferus from its unwanted introduction in Italy to date. Bull. Insectology 2013, 66, 273–281. [Google Scholar]
- Evans, G. The whiteflies (Hemiptera: Aleyrodidae) of the world and their host plants and natural enemies; USDA/Animal Plant Health Inspection Service (APHIS): Riverdale, MD, USA, 2008. [Google Scholar]
- Silvestri, F. Contribuzione alla conoscenza degli Aleurodidae (Insecta: Hemiptera) viventi su Citrus in Estremo Oriente e dei loro parassiti. II. Descrizione e notizie biologiche dei parassiti di Aleurodidi viventi su Citrus. Boll. Lab. Zool. Gen. Agrar. R. Ist. Super. Agrar. Portici 1927, 21, 20–60. [Google Scholar]
- Van den Berg, M.A.; Hoppner, G.; Greenland, J. An economic study of the biological control of the spiny blackfly, Aleurocanthus spiniferus (Hemiptera: Aleyrodidae), in a citrus orchard in Swaziland. Biocontrol Sci. Techn. 2000, 10, 27–32. [Google Scholar] [CrossRef]
- Muniappan, R.; Purea, M.; Sengebau, F.; Reddy, G.V.P. Orange spiny whitefly, Aleurocanthus spiniferus (Quaintance) (Homoptera: Aleyrodidae), and its parasitoids in the Republic of Palau. Proc. Hawaiian Entomol. Soc. 2006, 38, 21–25. [Google Scholar]
- Gyeltshen, J.; Hodges, A.; Hodges, G.S. Orange Spiny Whitefly, Aleurocanthus spiniferus Quaintance (Insecta: Hemiptera: Aleyrodidae); University of Florida IFAS Extension: Gainesville, FL, USA, 2019. [Google Scholar]
- Laudonia, S.; Melone, G.; Ascolese, R.; Nugnes, F. Eretmocerus iulii Laudonia et Melone sp.n.: parasitoid associated with Aleurocanthus spiniferus. Bull. Insectology 2024, 77, 263–269. [Google Scholar]
- Colazza, S.; Bin, F. Efficiency of Trissolcus basalis (Hymenoptera: Scelionidae) as an egg parasitoid of Nezara viridula (Heteroptera: Pentatomidae) in Central Italy. Environ. Entomol. 1995, 24, 1703–1707. [Google Scholar] [CrossRef]
- Brooks, M.E.; Kristensen, K.; van Benthem, K.J.; Magnusson, A.; Berg, C.W.; Nielsen, A.; Skaug, H.J.; Mächler, M.; Bolker, B.M. glmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. The R J. 2017, 9, 378–400. [Google Scholar] [CrossRef]
- R Core Team. R: A language and environment for statistical computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Noyes, J.S. Collecting and preserving chalcid wasps (Hymenoptera: Chalcidoidea). J. Nat. Hist. 1982, 16, 315–334. [Google Scholar] [CrossRef]
- Nugnes, F.; Russo, E.; Viggiani, G.; Bernardo, U. First record of an invasive fruit fly belonging to Bactrocera dorsalis complex (Diptera: Tephritidae) in Europe. Insects 2018, 9, 182. [Google Scholar] [CrossRef]
- Uesugi, R.; Sato, Y.; Han, B.; Huang, Z.D.; Yara, K.; Furuhashi, K. Molecular evidence for multiple phylogenetic groups within two species of invasive spiny whiteflies and their parasitoid wasp. Bull. Entomol. Res. 2016, 106, 328–340. [Google Scholar] [CrossRef]
- Simon, C.; Frati, F.; Beckenbach, A.; Crespi, B.; Liu, H.; Flook, P. Evolution, weighting, and phylogenetic utility of mitochondrial gene sequences and a compilation of conserved Polymerase Chain Reaction primers. Ann. Entomol. Soc. Am. 1994, 87, 651–701. [Google Scholar] [CrossRef]
- Park, J.K.; O’Foighil, D. Sphaeriid and corbiculid clams represent separate heterodont bivalve radiations into freshwater environments. Mol. Phylogenetics Evol. 2000, 14, 75–88. [Google Scholar] [CrossRef] [PubMed]
- Nunn, G.B.; Theisen, B.F.; Christensen, B.; Arctander, P. Simplicity correlated size growth of the nuclear 28S ribosomal RNA D3 expansion segment in the crustacean order Isopoda. J. Mol. Evol. 1997, 42, 211–223. [Google Scholar] [CrossRef] [PubMed]
- Polaszek, A.; Ayshford, T.; Yahya, B.E.; Fusu, L. Wallaceaphytis: an unusual new genus of parasitoid wasp (Hymenoptera: Aphelinidae) from Borneo. J. Nat. Hist. 2014, 48, 1111–1123. [Google Scholar] [CrossRef]
- Kumar, D.; Singh, M.; Kushwaha, M.; Makarana, G.; Yadav, M.R. Integrated use of organic and inorganic nutrient sources influences the nutrient content, uptake and nutrient use efficiencies of fodder oats (Avena sativa). Indian J. Agron. 2021, 66, 466–473. [Google Scholar] [CrossRef]
- Xia, X.; Xie, Z. DAMBE: Software package for data analysis in molecular biology and evolution. J. Hered. 2001, 92, 371–373. [Google Scholar] [CrossRef]
- Templeton, A.R.; Crandall, K.A.; Sing, C.F. A cladistic analysis of phenotypic associations with haplotypes inferred from restriction endonuclease mapping and DNA sequence data. III. Cladogram estimation. Genetics 1992, 132, 619–633. [Google Scholar] [CrossRef]
- Clement, M.; Posada, D.; Crandall, K.A. TCS: a computer program to estimate gene genealogies. Mol. Ecol. 2000, 9, 1657–1659. [Google Scholar] [CrossRef]
- Múrias dos Santos, A.; Cabezas, M.P.; Tavares, A.I.; Xavier, R.; Branco, M. tcsBU: a tool to extend TCS network layout and visualization. Bioinformatics 2016, 32, 627–628. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Borowiec, M.L. Spruceup: fast and flexible identification, visualization, and removal of outliers from large multiple sequence alignments. J. Open-Source Softw. 2019, 4, 1635. [Google Scholar] [CrossRef]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [PubMed]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the Ultrafast Bootstrap Approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef]
- Martin, J.H. The whitefly fauna of Australia (Sternorrhyncha: Aleyrodidae). A taxonomic account and identification guide; CSIRO Entomology Technical Paper 38; CSIRO Entomology: Canberra, Australia, 1999; p. 197. [Google Scholar]
- Dubey, A.K.; Sundararaj, R. Whitefly species of the genus Aleurocanthus Quaintance & Baker (Hemiptera: Aleyrodidae) from India, with descriptions of six new species. Orient. Insects 2005, 39, 295–321. [Google Scholar] [CrossRef]
- Dubey, A.K.; Ko, C.C. Sexual dimorphism among species of Aleurocanthus Quaintance & Baker (Hemiptera: Aleyrodidae) in Taiwan, with one new species and an identification key. Zootaxa 2012, 3177, 1–23. [Google Scholar] [CrossRef]
- Huang, J.; Polaszek, A. A revision of the Chinese species of Encarsia Förster (Hymenoptera: Aphelinidae): parasitoids of whiteflies, scale insects and aphids (Hemiptera: Aleyrodidae, Diaspididae, Aphidoidea). J. Nat. Hist. 1998, 32, 1825–1966. [Google Scholar] [CrossRef]
- Viggiani, G. Le specie italiane del genere Encarsia Foerster (Hymenoptera: Aphelinidae). Boll. Lab. Entomol. Agrar. Filippo Silvestri Portici 1987, 44, 121–179. [Google Scholar]
Figure 1.
Puparia of Aleurocanthus spiniferus with the characteristic wax secretion (A), the characteristic of the dorsal spines (B), the posterior part of the abdomen (with the vasiform orifice) (C), and the marginal teeth (D). Scale bar: 200 µm.
Figure 1.
Puparia of Aleurocanthus spiniferus with the characteristic wax secretion (A), the characteristic of the dorsal spines (B), the posterior part of the abdomen (with the vasiform orifice) (C), and the marginal teeth (D). Scale bar: 200 µm.

Figure 2.
Violin plots showing the distribution of Aleurocanthus spiniferus puparia per leaf in host tree species: Malus domestica, Pyrus communis, and Vitis vinifera. Each violin plot is overlaid with a box plot and individual data points to illustrate the central tendency and spread of the data. The thick horizontal line within each box represents the median, while the box boundaries indicate the interquartile range (IQR).
Figure 2.
Violin plots showing the distribution of Aleurocanthus spiniferus puparia per leaf in host tree species: Malus domestica, Pyrus communis, and Vitis vinifera. Each violin plot is overlaid with a box plot and individual data points to illustrate the central tendency and spread of the data. The thick horizontal line within each box represents the median, while the box boundaries indicate the interquartile range (IQR).
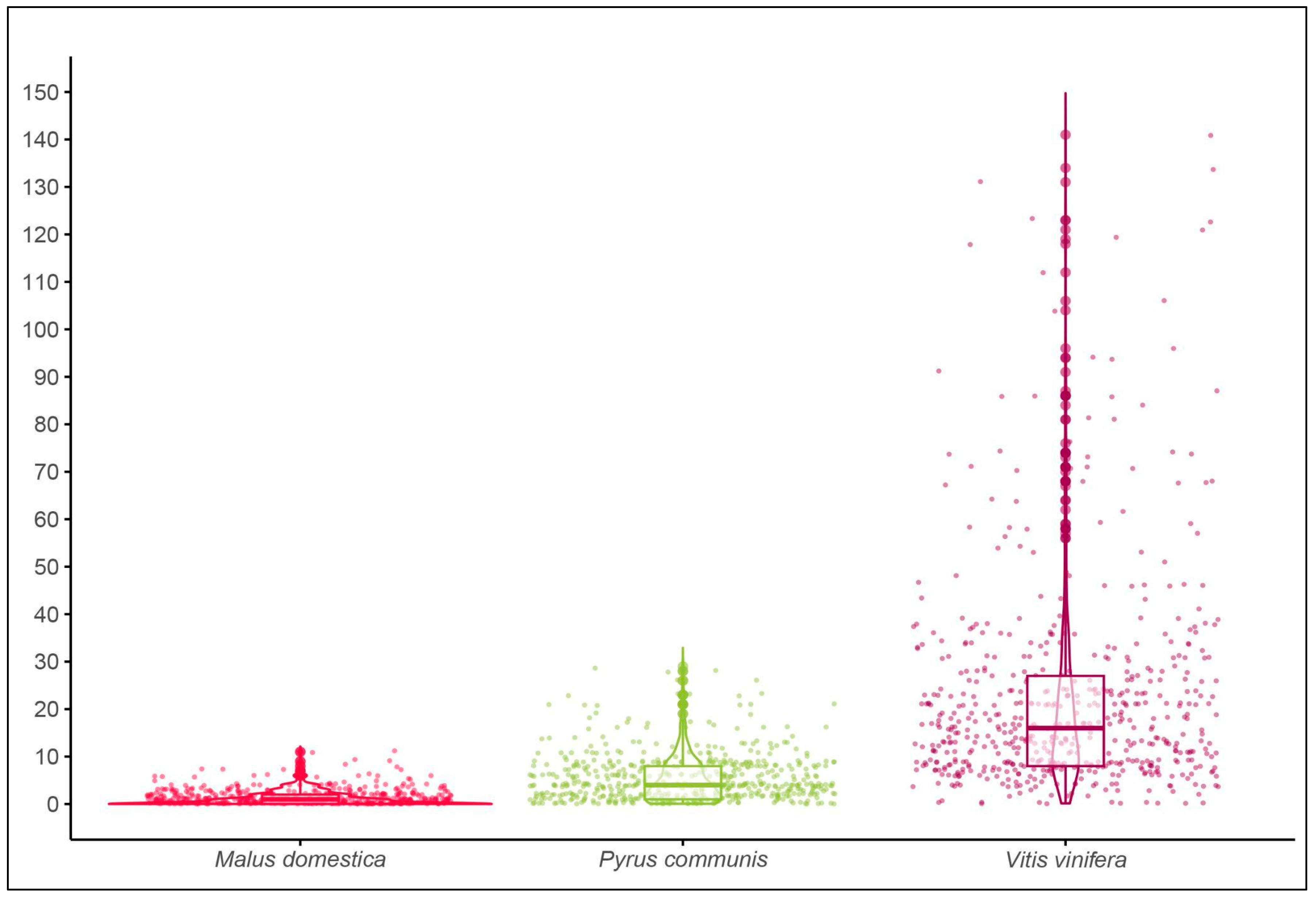
Figure 3.
A) Dorsal meso-metasoma and legs; B) Face; C) antenna, and D) fore wing of Encarsia nipponica. Photos were taken with an Olympus BX63 microscope.
Figure 3.
A) Dorsal meso-metasoma and legs; B) Face; C) antenna, and D) fore wing of Encarsia nipponica. Photos were taken with an Olympus BX63 microscope.

Figure 4.
Subtree from maximum likelihood tree (IQ-TREE 2) based on 28s D2 ribosomal DNA (535 bp) from 81 taxa (6 ingroup, 75 outgroup); support values from 1000 ultrafast bootstrap replicates, shown if above 90.
Figure 4.
Subtree from maximum likelihood tree (IQ-TREE 2) based on 28s D2 ribosomal DNA (535 bp) from 81 taxa (6 ingroup, 75 outgroup); support values from 1000 ultrafast bootstrap replicates, shown if above 90.
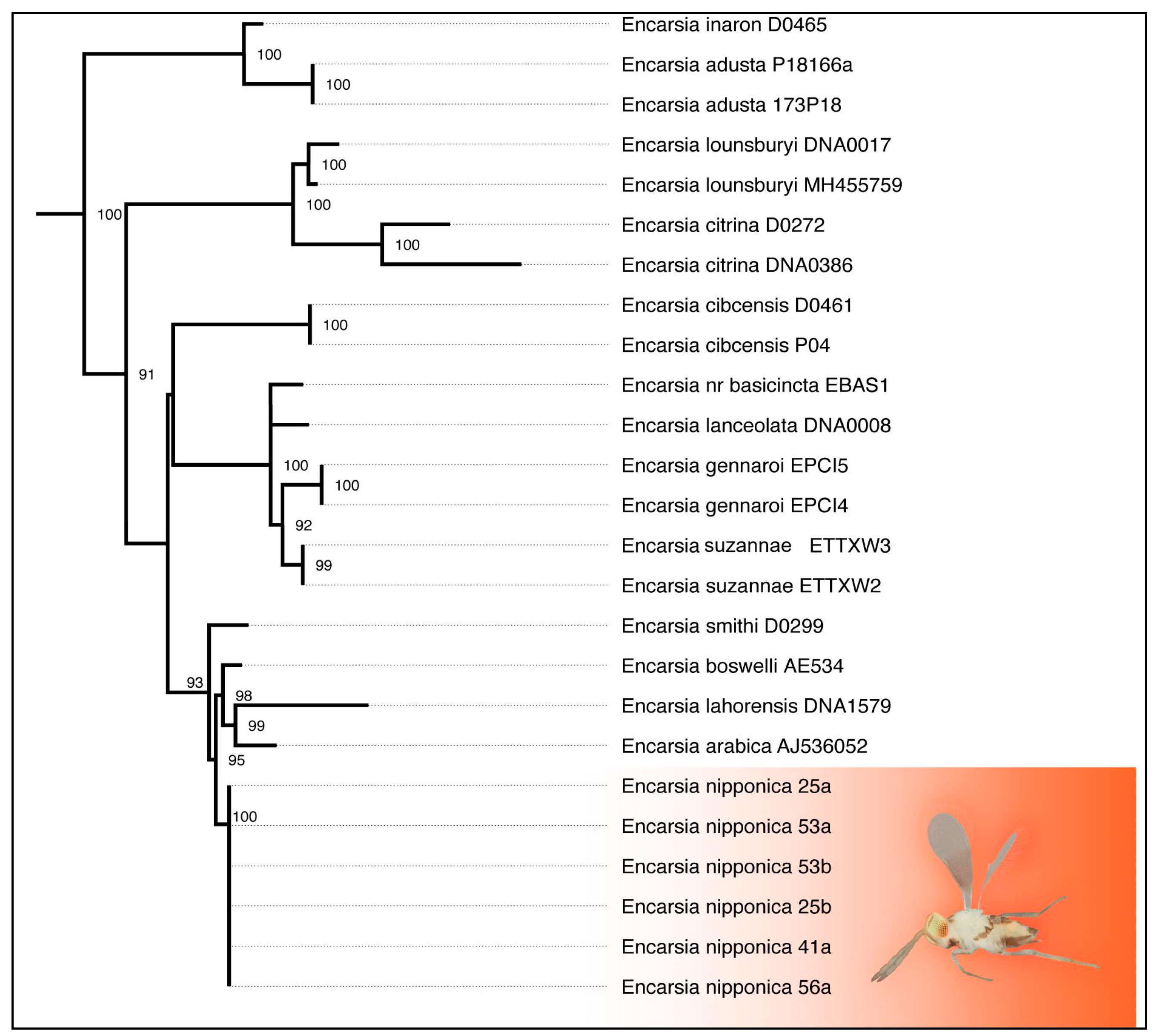

Table 1.
Information on species, sampling year, sites, and host plants for specimens utilised for molecular analyses in the present study.
Table 1.
Information on species, sampling year, sites, and host plants for specimens utilised for molecular analyses in the present study.
| Specimen Code | Preliminary Identification | Year | Sampling site | Host Plant/Host |
|---|---|---|---|---|
| As GBP V1 | Aleurocanthus sp. | 2023 | Giardino Botanico La Pica San Felice sul Panaro |
Vitis vinifera |
| As GBP V2 | Aleurocanthus sp. | 2023 | Giardino Botanico La Pica San Felice sul Panaro |
Vitis vinifera |
| As GBP V3 | Aleurocanthus sp. | 2023 | Giardino Botanico La Pica San Felice sul Panaro |
Vitis vinifera |
| As GBP V4 | Aleurocanthus sp. | 2023 | Giardino Botanico La Pica San Felice sul Panaro |
Vitis vinifera |
| As GBP M1 | Aleurocanthus sp. | 2023 | Giardino Botanico La Pica San Felice sul Panaro |
Pyrus communis |
| As GBP M2 | Aleurocanthus sp. | 2023 | Giardino Botanico La Pica San Felice sul Panaro |
Pyrus communis |
| As GBP M3 | Aleurocanthus sp. | 2023 | Giardino Botanico La Pica San Felice sul Panaro |
Pyrus communis |
| As GBP M4 | Aleurocanthus sp. | 2023 | Giardino Botanico La Pica San Felice sul Panaro |
Pyrus communis |
| As M226 1 | Aleurocanthus sp. | 2023 | Azienda Magnanini Carpi |
Pyrus communis |
| As M226 2 | Aleurocanthus sp. | 2023 | Azienda Magnanini Carpi |
Pyrus communis |
| As M226 3 | Aleurocanthus sp. | 2023 | Azienda Magnanini Carpi |
Pyrus communis |
| As bom 5 | Aleurocanthus sp. | 2018 | Bomporto | Orchard hedge |
| 25a | Encarsia sp. | 2022 | Giardino Botanico La Pica S. Felice sul Panaro |
Aleurocanthus sp. |
| 25b | Encarsia sp. | 2022 | Giardino Botanico La Pica S. Felice sul Panaro |
Aleurocanthus sp. |
| 41° | Encarsia sp. | 2022 | Giardino Botanico La Pica S. Felice sul Panaro |
Aleurocanthus sp. |
| 53° | Encarsia sp. | 2022 | Giardino Botanico La Pica S. Felice sul Panaro |
Aleurocanthus sp. |
| 53b | Encarsia sp. | 2022 | Giardino Botanico La Pica S. Felice sul Panaro |
Aleurocanthus sp. |
| 56a | Encarsia sp. | 2022 | Giardino Botanico La Pica S. Felice sul Panaro |
Aleurocanthus sp. |
Table 2.
Results from a generalised linear mixed model (GLMM) with a negative binomial distribution. The table shows the median count with interquartile range (IQR) in square brackets for each tree species (Malus domestica; Pyrus communis and Vitis vinifera), along with the estimated effects of each species relative to M. domestica. The z-value and P-value indicate the statistical significance of the difference in the number of Aleurocanthus spiniferus puparia compared to M. domestica.
Table 2.
Results from a generalised linear mixed model (GLMM) with a negative binomial distribution. The table shows the median count with interquartile range (IQR) in square brackets for each tree species (Malus domestica; Pyrus communis and Vitis vinifera), along with the estimated effects of each species relative to M. domestica. The z-value and P-value indicate the statistical significance of the difference in the number of Aleurocanthus spiniferus puparia compared to M. domestica.
| median [IQR] | estimate | z value | P value | |
|---|---|---|---|---|
| Malus domestica | 1[0,2] | - | - | - |
| Pyrus communis | 4[1,8] | 1.45 | 22.21 | <0.001 *** |
| Vitis vinifera | 16[8,27] | 2.891 | 45.88 | <0.001 *** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



