
Submitted:
16 July 2025
Posted:
17 July 2025
You are already at the latest version
Abstract
Lead (Pb) is a non-essential, toxic heavy metal with no known biological function that has caused widespread environmental contamination throughout human history. Pb toxicity represents one of the most persistent environmental health challenges, with no safe exposure threshold identified. The metal demonstrates remarkable persistence in biological systems, with approximately 90% of it stored in bone tissue for decades, mimicking calcium due to its similar ionic properties. Contemporary contamination primarily stems from mining activities, battery manufacturing, electronic waste recycling, and deteriorating infrastructure. Pb enters organisms through multiple pathways and causes severe health impacts across all biological systems, with particularly devastating neurodevelopmental and bone effects in children and cardiovascular, and reproductive consequences in adults. On a molecular level, Pb disrupts cellular processes through ion mimicry, replacing essential metals in enzymes and proteins, leading to mitochondrial dysfunction, oxidative stress, DNA damage, and epigenetic modifications. This review examines the sources of Pb pollution and its toxicological impacts on bacteria, fungi, plants, animals, and humans. It explores the molecular mechanisms underlying these effects, including neuroinflammation, genotoxicity, and cell death pathways. The paper considers current approaches for Pb removal from contaminated environments and therapeutic interventions for Pb poisoning.
Keywords:
Pb pollution
; Pb toxicity
; bioaccumulation
; neurotoxicity
; oxidative stress
; ion mimicry
; environmental remediation
; molecular mechanisms
1. Introduction
Lead (Pb, atomic number: 82, atomic mass: 207.2, period number: 6, Group 14, electron configuration: [Xe]6s²4f¹⁴5d¹⁰6p², isotopes: 4 stable) is one of the earliest metals used by humans. The name "lead" originates from the Anglo-Saxon word of unknown origin, while its chemical symbol, Pb, is derived from the Latin word plumbum [1]. Historical records indicate that lead was widely used in ancient societies for pipes, cookware, cosmetics, and even wine storage, which significantly contributed to environmental contamination [2]. In Table 1, some of Pb’s physical and chemical properties are described.
During the Industrial Revolution and especially throughout the 20th century, anthropogenic activities such as the use of tetraethyl lead in gasoline dramatically increased global lead emissions. Although the use of leaded gasoline has been banned in most countries since the 1980s, lead remains persistent in soils, buildings, and older infrastructure [3]. Today, the primary sources of lead exposure include mining, battery manufacturing, smelting, electronic waste recycling, and the deterioration of lead-based paints, as well as aging plumbing systems that continue to contaminate drinking water [2,3].
The WHO classifies Pb as one of the ten most hazardous chemical elements for humans. In 2016, data from the Institute for Health Metrics and Evaluation (IHME) indicated that Pb poisoning resulted in 540.000 deaths globally. Most of these fatalities occurred in low-income countries. Pb contributes to 64% of primary idiopathic intellectual disabilities, 3% of ischemic heart disease cases, and approximately 3.1% of heart attacks worldwide [4]. Pb has long been a part of human life; for centuries, its compounds were used to sweeten sour wines, and it is still produced today in Pb-Zn processing plants, as well as in paint factories, photocopiers, and in automotive and jet fuels to boost octane levels. According to the WHO, Pb emissions into the atmosphere come from various recycling activities. Pb is also utilized in the manufacture of batteries, rivets, ammunition, ceramic glazes, jewelry, toys, and traditional medicine. A significant portion of Pb is used in car batteries [4]. The International Lead Association (ILA) reports that global annual primary Pb production is around 5 million tons, valued at approximately $15 billion, which has doubled over the past decade. When released into outdoor air, Pb becomes part of street dust, potentially causing allergic reactions in the population. The US Environmental Protection Agency (EPA) recommends that Pb levels in city air should not exceed 9 µg/m³. In the USA, Pb is one of six required air quality parameters, alongside CO, NO2, O3, SO2, and PM2.5 and PM10 particulate matter. In freshwater, Pb concentrations typically range from 1 to 10 µg/L [4]. Figure 1 depicts the concentration of Pb in agricultural soils and pasture lands in Europe.
Lead is a non-essential, highly toxic metal with no known biological function. Once absorbed—mainly through inhalation or ingestion—it is distributed to the brain, liver, kidneys, and bones. Lead mimics essential metals such as calcium and zinc, disrupting cellular signaling and enzyme activity. It induces oxidative stress, inflammation, and interferes with gene expression and DNA repair mechanisms [6,2].
Due to its long biological half-life and poor excretion, lead bioaccumulates in soft and hard tissues over time. Chronic exposure affects multiple organ systems, particularly the nervous, renal, cardiovascular, hematopoietic, and reproductive systems. Children are especially vulnerable due to greater absorption rates and ongoing neurodevelopment, with even low blood lead levels associated with cognitive and behavioral impairments [6,2]. According to the Global Burden of Disease Study, lead exposure contributed to approximately 900,000 deaths globally in 2019, reinforcing its classification as a probable human carcinogen [2]. Pb and inorgarnic compounds are classified in Group 2A of probable carcinogens according to IARC (International Agency of Reasearch on Cancer - https://monographs.iarc.who.int/list-of-classifications Accessed on 09.07.2025), while organic compounds are classified in Group 3.
2. The Origin of Pb Pollution
Lead is a toxic heavy metal that has been extensively utilized throughout human history, resulting in its widespread environmental distribution. Despite natural sources, such as volcanic emissions and erosion, most of the environmental Pb burden is of anthropogenic origin (Figure 2) [2]. The industrial use of lead peaked during the 20th century with its incorporation in gasoline additives (e.g., tetraethyl lead), batteries, paints, ceramics, and plumbing systems [7]. Even after regulatory efforts to eliminate leaded gasoline, Pb emissions from smelting, battery recycling, and mining continue to be prominent pollution sources worldwide [8].
Historically, the use of lead in urban infrastructure—such as water pipes, roofing, and household paint—resulted in significant accumulation in soil and dust, especially in densely populated or industrialized regions. In the United States, legacy contamination from such uses has led to persistent soil Pb levels exceeding standard thresholds, particularly in urban neighborhoods with older housing stock [10]. Likewise, regions in Poland, such as Upper Silesia, experienced prolonged air pollution due to nearby heavy industry, which contributed to substantial lead deposition in forest and park soils [11].
In many low- and middle-income countries, lead exposure remains an urgent health concern. Informal recycling of lead-acid batteries, illegal smelting, and adulteration of consumer products (e.g., spices, cosmetics) are major contributors to environmental and human contamination [8,12]. Sustained environmental exposure, even at low levels, persists due to Pb's long environmental half-life and strong binding to soil and sediment matrices [13].
The persistence of lead in soil is influenced by factors such as pH, organic matter content, and soil texture. For instance, acidic soils or those rich in organic carbon tend to enhance Pb mobility and bioavailability, posing risks to soil microflora and indirectly affecting plant health and nutrient cycling [11]. In Polish mountain landscapes affected by industrial emissions, these soil conditions amplified Pb toxicity and microbial stress, particularly in the upper soil layers [11].
Lead contamination of water systems is another concern, primarily through runoff from polluted soil or the corrosion of plumbing components containing lead. Even minimal levels of Pb in drinking water or food can cumulatively harm biological systems due to Pb's poor excretion and high affinity for biological molecules [13,2]. These properties make Pb a persistent threat to environmental and public health, particularly in developing regions where monitoring and remediation infrastructure is lacking.
2.1. Natural Occurrence of Pb
2.1.1. Soil
Pb naturally occurs in trace concentrations within the Earth’s crust, constituting approximately 0.002% of its composition [14]. Background concentrations of lead in soil primarily arise from the weathering of bedrock containing lead-bearing minerals and other geogenic processes, such as sea spray and volcanic activity, particularly in regions with significant natural mineral deposits [14,15].
Over time, soils may become enriched with lead through the slow breakdown of parent materials such as galena (PbS), anglesite (PbSO₄), and cerussite (PbCO₃). These minerals release lead ions into the soil matrix during prolonged physical and chemical weathering processes [16]. The mobility and chemical behavior of naturally sourced lead in soil are strongly influenced by local biogeochemical conditions, particularly soil pH and redox potential. In acidic soils (pH < 4.0), lead is more likely to exist in a soluble ionic form (Pb²⁺), increasing its bioavailability and potential for plant uptake [16]. Conversely, in alkaline conditions (pH > 7.0), lead tends to precipitate as insoluble compounds such as Pb(OH)₂ or PbCO₃, reducing both its mobility and toxicity [16]. In areas rich in iron (Fe) and manganese (Mn) oxides, naturally occurring lead is often adsorbed onto these mineral surfaces, significantly limiting its movement and uptake by plants [15].
Additionally, lead released through natural weathering may form long-lived complexes with clay minerals and organic matter, further influencing its persistence and behavior in the soil environment [15]. High-molecular-weight organic compounds can bind Pb ions, forming stable complexes that reduce lead solubility and limit biological interactions. However, low-molecular-weight organics may increase lead mobility by forming soluble chelates [16].
Microbial activity is another critical factor affecting the fate of naturally present lead. Soil microbes can influence pH and redox conditions through metabolic processes, thereby altering lead speciation and its ecological risk profile [16]. Some bacteria, particularly under anaerobic conditions, can convert lead into less mobile sulfide forms (e.g., PbS), effectively immobilizing the metal [16].
2.1.2. Water
Based on the analyzed studies, lead (Pb) concentrations in marine and freshwater environments show significant variation and contamination patterns. In marine sediments from Mallorca's southwest coast, background Pb levels averaged 10.1 mg/kg. Still, concentrations near wastewater discharge points reached heavily polluted levels, with three samples at Baluard station exceeding probable effect levels (PEL) according to NOAA standards. In contrast, six additional samples fell between the threshold and PEL [17]. The Red Sea coastal sediments showed elevated contamination with mean Pb concentrations of 57.8 mg/kg in Egypt, 39.7 mg/kg in Saudi Arabia, and 96.67 mg/kg in Jordan (Table 2) – all significantly above the upper continental crust reference level of 17.0 mg/kg. One extreme case in Egypt's Hurghada region reached 865 mg/kg, indicating serious non-carcinogenic health risks [18].
Freshwater systems demonstrate comparatively lower but still concerning levels of Pb. In China's Fenghe River Basin, water Pb concentrations ranged from 0.70 to 1.51 μg/L, remaining below the WHO standard (10 μg/L). In contrast, sediment concentrations averaged 30.2 mg/kg, exceeding the regional background value of 21.4 mg/kg at all sampling points [20]. African water bodies show alarming contamination levels, with WHO drinking water standards (0.01 mg/L) exceeded by ratios of 82-669 times in various locations, including Lake Victoria, boreholes, and rivers, highlighting the urgent need for improved water treatment and monitoring systems across different aquatic environments [21].
Lead contamination in water arises primarily from anthropogenic sources, including industrial discharge, urban runoff, aging infrastructure, and untreated domestic wastewater [3]. In urban environments, rain and snowmelt mobilize historical lead deposits from roads, rooftops, and construction surfaces, allowing them to enter surface water bodies [3]. Although lead concentrations in the water column may diminish through dilution or sedimentation, sediments located near urban and industrial areas retain significant amounts of lead, serving as persistent secondary sources of contamination [3]. Furthermore, domestic waste from urban centers — especially where wastewater treatment is insufficient—introduces various heavy metals, including lead, into river systems [22]. In remote regions, such as the Arctic and subarctic, exposure remains a concern due to reliance on untreated water sources and older infrastructure, particularly among Indigenous populations [23].
Industrial processes constitute a major contributor to lead input in aquatic environments. Heavy industries such as smelting, coal processing, tanning, and chemical manufacturing discharge significant volumes of effluents, often containing lead and other toxic metals, into surface and groundwater systems [22]. When inadequately treated or released directly, these discharges severely compromise water quality and accumulate in sediments [22]. Alongside this, agriculture also acts as a widespread non-point source of heavy metal pollution. Fertilizers, pesticides, and other chemical inputs applied on farmlands can contain lead and related metals, which are then mobilized by rainwater runoff into nearby aquatic systems [22]. Continuous input from agricultural activities has led to notable long-term contamination in regions of intensive farming [22].
Improper waste management practices also play a significant role in lead contamination. Landfills and open dumping sites lacking containment systems allow leachates enriched with heavy metals, including lead, to infiltrate surrounding soils and water bodies [22]. Municipal solid waste, which often contains a mix of industrial, household, and healthcare refuse, contributes to this problem when it is dumped without segregation [22,3]. Over time, toxic substances from decomposing waste migrate into surface and groundwater via percolation or runoff [22,3]. Additionally, natural processes, such as the weathering of rocks and minerals, can release trace amounts of lead into water systems; however, these sources are generally less significant compared to anthropogenic inputs [22].
2.1.3. Air
Lead enters the atmosphere and binds to particulate matter that can be transported over long distances and subsequently deposited onto soil and dust surfaces, creating a significant exposure pathway for humans through both inhalation and ingestion [24]. Regulatory standards for workplace air exposure vary significantly, with OSHA establishing limits of 0.5 mg/m³ for elemental lead in general industry and 50 μg/m³ in construction environments, while WHO maintains an air quality standard of 0.5 μg/m³ [24,25].
Atmospheric monitoring studies have documented varying lead concentrations across different urban and industrial environments. In the industrial areas of Coimbatore city, suspended particulate matter contained lead concentrations ranging from 5 to 7 μg/m³, with lead ranking fourth among the heavy metals detected (Fe > Zn > Cu > Pb > Ni > Cr) [26]. Dust fall particles in Zarand, Iran, showed lead concentrations of 1.01 mg/kg with an enrichment factor of 194.2, indicating predominantly anthropogenic origins from vehicle and industrial emissions [19]. These atmospheric lead particulates pose significant health risks through inhalation, particularly affecting the cardiovascular and respiratory systems, and contribute to soil and household dust contamination [24,26]. Table 3 summarizes the maximal permissible limits of Pb in food, drinking water, soil, and air.
2.2. Anthropogenic Pb Sources
Industrial and agricultural activities synergistically contribute to environmental lead contamination through multiple pathways. Rapid industrialization in developing nations has intensified lead release through manufacturing processes, waste incineration, and improper disposal of effluents. At the same time, agricultural practices involving contaminated fertilizers, pesticides, and wastewater irrigation perpetuate soil contamination [31].
Atmospheric deposition represents the predominant contributor to agricultural lead contamination, particularly in industrialized regions. In China, atmospheric deposition accounted for 80-94% of total lead inputs to agrarian soils between 2006 and 2015, compared to 43-85% between 1999 and 2005. Coal combustion contributed 22% and vehicle emissions 78% to lead in atmospheric dust [32]. Industrial activities, including fossil fuel combustion, manufacturing processes, and mining operations, constitute primary anthropogenic sources. Phosphate fertilizers contain average lead concentrations of 2.9 mg/kg¹ across 12 European countries [32].
Metal cookware emerges as a significant and underrecognized source of lead exposure due to manufacturing practices that utilize contaminated scrap materials. Aluminum and brass cookware frequently exceed 100 ppm lead content (0,01%), with some brass components reaching 10,000 ppm (1%), particularly in handles [33]. Traditional Afghan pressure cookers have been found to contain lead levels of up to 69,000 ppm in the pressure relief vent pipes. At the same time, Hindalium cookware leached lead amounts exceeding childhood dietary limits by 1400-fold after 24 hours of contact with acidic solutions [33].
Occupational exposure is the most prevalent anthropogenic source of lead in the Middle Eastern and North African regions, accounting for 26.5% of documented cases. Workers in petrol stations, battery manufacturing, smelting operations, and automotive repair face elevated exposure risks, with secondary contamination affecting families through contaminated clothing [12]. Cosmetic products, particularly traditional kohl, represent the second most common source, accounting for 23.5% of cases. In contrast, residential factors, including lead-based paint in buildings older than 20 years, account for 20.6% of exposure sources [12].
3. Pb Toxicity on Living Organisms
3.1. Pb Accumulation and Toxicity on Bacteria
Lead toxicity varies significantly among different strains, with some showing remarkable tolerance while others demonstrate high sensitivity. Gram-positive bacteria, such as Bacillus species, Lactobacillus strains (L. acidophilus, L. rhamnosus, and L. plantarum), and Micrococcus species, exhibit considerable lead resistance and removal capabilities through biosorption and bioaccumulation mechanisms [34]. Similarly, Gram-negative bacteria, including Pseudomonas aeruginosa, Enterobacter cloacae, and Stenotrophomonas maltophilia, demonstrate effective lead tolerance and detoxification abilities [34]. Examples of Pb-resistant bacteria isolated from contaminated sites include also Acinetobacter junii Pb1, Bacillus subtilis X3, Delftia tsuruhatensis, Halomonas sp., and Pseudomonas aeruginosa N6P6 [35]. However, particular species, such as Desulfovibrio desulfuricans G20, show extreme sensitivity to lead, with complete growth inhibition at just 3 mg/L Pb(II) chloride, whereas Thermus thermophilus strain Samu-SA1 experiences reduced protein content and enzyme activity at lead concentrations of 100-300 mg/L [34] (Figure 3).
3.1.1. Resistance Mechanisms of Bacteria
Bacteria have evolved sophisticated resistance mechanisms to counteract lead toxicity, employing multiple strategies for survival in contaminated environments. Efflux systems play a central role in microbial defense against lead toxicity in Pb/Zn tailings, with membrane-bound enzymes called P-type ATPases such as ZntA actively exporting Pb²⁺ ions from the cytoplasm to the external environment through ATP hydrolysis, thereby maintaining cellular homeostasis under metal stress [37,38]. Under high lead exposure, the pbrTRABCD gene cluster becomes specifically activated, with elevated expression of genes such as pbrB, pbrT, and pbrD, which encode proteins involved in lead transport and sequestration. In parallel, other efflux systems — particularly RND family transporters and ABC-type pumps — utilize ATP hydrolysis or proton gradients to extrude toxic Pb²⁺ ions, further contributing to microbial resilience in contaminated environments [35, 39].
Metal sequestration mechanisms play crucial roles in lead resistance through intracellular binding and precipitation strategies. Cysteine-rich metallothioneins effectively trap toxic heavy metals, including Pb, allowing bacteria to tolerate high concentrations by forming non-toxic complexes within the cytoplasm [35]. Glutathione acts as an alternative chelator, scavenging and detoxifying lead through its thiol group, while metallochaperones assist in transporting metal ions within cells to minimize toxic impact [35]. Some bacterial species, including Bacillus, Staphylococcus aureus, Shewanella, Providencia, and Vibrio harveyi, employ precipitation mechanisms by sequestering lead as phosphate salts, forming intracellular precipitates that reduce bioavailability [35,38].
Lead-resistant bacterial communities demonstrate specific evolutionary adaptations and regulatory responses to metal stress. Proteobacteria are noted to carry most heavy metal resistance genes, with metal-resistant taxa including Afipia, Bradyrhizobium, Sphingomonas, and Miltoncostaea showing particular adaptations to lead-rich environments [37]. Pseudomonas S8A employs comprehensive strategies including morphological differentiation, surfactant production, and specific protein expression for lead resistance [37].
3.1.2. Bacteria in Pb-Contaminated Soil
Bacterial adaptation mechanisms in Pb-contaminated environments involve biosorption, where Pb ions directly adsorb onto functional groups on bacterial cell surfaces, including hydroxyl, amide, carboxyl, and phosphate groups, and intracellular sequestration as a detoxification strategy [40]. The efficiency of microbial-mediated immobilization depends on bacterial taxa and ambient environmental conditions, including pH, oxygen status, and metal concentrations [40]. Microorganisms employ key mechanisms, including immobilization of lead into insoluble precipitates through phosphate solubilization and microbially induced carbonate precipitation (MICP) [40]. Phosphate Solubilizing Bacteria (PSB), widely distributed in soils including the rhizosphere, can convert insoluble phosphorus compounds into reactive phosphate that binds with Pb to form insoluble Pb-phosphate precipitates like pyromorphite [Pb5(PO4)3X] [40]. Examples of PSB involved in Pb immobilization include Bacillus subtilis, Pantoea sp., Enterobacter sp., Acinetobacter pittii, Serratia marcescens, Leclercia adecarboxylata, and Pseudomonas putida [40]. MICP processes involve ureolytic organisms that produce carbonate (CO3²⁻) through urea hydrolysis, which then binds with Pb²⁺ to form insoluble lead carbonate (PbCO₃) precipitates, with species including Sporosarcina spp., Viridibacillus arenosi, Enterobacter cloacae, Lysinibacillus sphaericus, Kocuria flava, and Staphylococcus epidermis [40].
3.2. Pb Accumulation and Toxicity on Fungi
Lead exposure induces significant morphological and physiological changes in fungal cells, including wrinkling, deformation, and the formation of crystalline precipitates such as lead hydroxyl apatite [41,42]. Lead toxic effects trigger oxidative stress responses and activate antioxidant defense systems, while simultaneously disrupting cellular membranes, DNA structure, and enzymatic functions [36]. Despite these adverse effects, metal-tolerant fungi maintain their bioremediation capacity through mechanisms including bioaccumulation, biosorption, and biotransformation, making them promising candidates for environmental cleanup applications [36,43].
The mechanisms of lead removal vary significantly with metal concentration and fungal species. At lower concentrations (25-50 mg/L), fungi primarily utilize intracellular accumulation, while at higher concentrations (75-300 mg/L), extracellular biosorption becomes dominant [44]. The biosorption process involves multiple functional groups including O-H, C-H, N-H, C=O, and C-O groups, with peak biosorption capacity typically occurring on day 5 of incubation, reaching values up to 97.82 mg/g in A. niger [42]. Sarocladium strain Pb-9 showed maximum adsorption rates of 37.75% at 2000 mg/L concentration [41].
Multiple fungal species demonstrate remarkable tolerance to high lead concentrations, with Aspergillus awamori and Aspergillus niger strains showing outstanding tolerance at concentrations up to 10,000 mg/L [42]. Verticillium insectorum J3 survived and grew in medium containing 500 mg/L Pb(II), achieving approximately 90% removal rates at concentrations between 75-250 mg/L [44]. White rot fungi also demonstrate exceptional performance, with Phanerochaete chrysosporium removing over 90% of lead from 50 mg/L solutions and achieving up to 99% efficiency in industrial wastewater treatment [43].
3.3. Pb Accumulation and Toxicity on Edible Mushrooms
Wild edible mushrooms demonstrate significant capacity for lead bioaccumulation, with concentrations varying considerably among species and collection sites. Lead content in mushrooms typically ranges from 0.004 to 37.1 mg/kg dry weight, with Umbilicaria esculenta showing the highest accumulation potential at 12.3 ± 7.0 mg/kg d.w. [45]. Among commonly consumed species, Suillus luteus accumulated 2.21 ± 0.48 mg/kg d.w., while Cantharellus cibarius contained 0.424-1.01 mg/kg d.w. [46,47]. Transfer factors from soil to mushroom tissues range from 1.528% to 35.2%, with Armillaria mellea demonstrating particularly high bioaccumulation efficiency at 18.6% relative to underlying soils [48].
Most wild mushroom samples exceed the established regulatory limits for lead content, with mean concentrations surpassing the EU maximum level of 0.800 mg/kg dry weight (d.w.) for wild fungi [48]. However, when compared to the 3.0 mg/kg dry weight standard, naturally collected mushrooms from uncontaminated areas generally remain within acceptable limits [46]. The estimated daily intake of lead from mushroom consumption (2.51x10-3 to 3.12x10-3 mg/day) typically does not exceed provisional tolerable weekly intake levels, with health risk indices remaining below 1 for non-carcinogenic effects [49,45]. Despite relatively low daily intake levels, lead exposure from mushroom consumption presents concerning carcinogenic risks that exceed acceptable thresholds (10⁻⁶ to 10⁻⁴), with carcinogenic risk values ranging from 5.67 × 10⁻⁴ to 1.68 × 10⁻³ for adults [48].
3.4. Pb Accumulation and Toxicity on Plants
Plants absorb Pb primarily through root systems via passive diffusion, facilitated transport, and active uptake mechanisms involving Zinc-Iron Permease (ZIP) and Natural Resistance-Associated Macrophage Protein (NRAMP) families (Figure 4) [50]. Soil physicochemical properties significantly influence uptake, with lower pH increasing Pb solubility and bioavailability [51,50]. The Casparian strip in root endodermis acts as a critical barrier, restricting Pb translocation from roots to shoots, resulting in more than 95% of absorbed Pb accumulating in root tissues [51]. This distribution pattern follows the sequence: roots > leaves > stem > inflorescence > seeds, with root vegetables typically showing the highest Pb concentrations [52]. Pb accumulation varies significantly among plant species, with hyperaccumulators like Arabis paniculata, Isachne globosa, and Pogonatherum crinitum capable of accumulating up to 1000 mg kg⁻¹ Pb in tissue [53].
The World Health Organization has established a permissible limit of 2 mg/kg for Pb concentration in plants, while soil tolerance reaches 85 mg/kg [55]. However, Pb contamination varies significantly among agricultural systems, as demonstrated by lettuce grown in bioslurry-treated soils containing 16.7 mg/kg Pb, considerably exceeding the WHO/FAO permissible limit of 0.3 mg/kg for vegetables [56]. Fiber crops demonstrate variable accumulation patterns: Kenaf can accumulate up to 867.55 mg/kg Pb in shoots compared to 329.66 mg/kg in roots, while Hemp shows preferential root accumulation with concentrations reaching 38.2 mg/kg in roots versus 16.5 mg/kg in leaves [57].
Lead toxicity severely impairs plant growth and development, causing a 42% reduction in root growth and a 14-30% decrease in rice seed germination at 1 mM concentrations [52]. In maize cultivation, Pb exposure at 3000 mg/kg soil concentration caused a 20% reduction in plant height, a 15% decrease in fresh weight, and a 25% reduction in dry weight, alongside increased electrical conductivity (15.22%) and decreased leaf relative water content (62.32%) [58]. Lead exposure significantly affects stomatal structure and function, inducing stomatal closure in multiple species, including tobacco, white leadtree, black gram, and soybean, which contributes to reduced stomatal conductance and decreased transpiration rates [59]. Paradoxically, Pb can increase stomatal number and density in species such as sunflower and plantain, while causing ultrastructural changes in guard cells, including a decrease in guard cell diameter and the accumulation of starch grains and plastid globules [59].
Pb exposure disrupts essential physiological processes, including photosynthesis, chlorophyll biosynthesis (particularly chlorophyll B), and ATP synthesis [52,50]. The metal interferes with nutrient uptake of essential cations (Mn²⁺, Zn²⁺, Fe²⁺, Ca²⁺, Mg²⁺) and induces oxidative stress through excessive production of reactive oxygen species (ROS), including H₂O₂, O₂²⁻, and OH⁻ radicals [51,50]. Plants have evolved sophisticated defense mechanisms to counteract Pb toxicity, including activation of antioxidant defense systems comprising enzymatic antioxidants superoxide dismutase (SOD), catalase (CAT), peroxidase (POD) glutathione peroxidase (GPx) and non-enzymatic compounds (ascorbic acid, glutathione, carotenoids) [50,51]. Metal sequestration strategies involve the synthesis of phytochelatins and metallothioneins for Pb chelation, followed by vacuolar compartmentalization [60,61]. Cell wall binding through pectin-Pb complexes provides additional physical barriers, while specific genes, such as JcMT2a, JcPAL, and NtCBP4, have been identified as contributing to Pb tolerance in various species [60,61]. Advanced OMICS approaches have revealed complex molecular responses to Pb stress, including differential expression of 36 genes in maize roots and 181 proteins in Rhoeo discolor leaves exposed to Pb toxicity [62]. Epigenetic modifications play a crucial role in Pb tolerance, with DNA hypomethylation of transporter gene promoters (TaABCCs and TaHMA2) enhancing the metal sequestration capacity in wheat [63]. Genome-wide association studies have identified 4 quantitative trait loci (QTL) and 9 candidate genes related to Pb resistance in Brassica napus, while 21 QTLs were linked to Pb toxicity in rice grain [62].
3.5. Pb Accumulation and Toxicity on Animals
Pb bioaccumulation patterns vary significantly across animal species and tissues, with preferential accumulation in metabolically active organs. In invertebrates, lead accumulates in body tissues, nests (in social species), Malpighian tubules, and exoskeletons, where it may be temporarily stored as part of detoxification before molting [64]. Terrestrial invertebrates demonstrate heightened sensitivity to Pb exposure, experiencing detrimental effects at concentrations below current international permissible limits, including impaired fertility, developmental defects, and behavioral disturbances [64]. In marine fish, Pb preferentially binds to calcium-rich tissues such as bones and gills, while also accumulating in liver and kidney tissues through detoxification processes [65]. Unlike mercury, Pb generally does not biomagnify strongly through marine food webs and often exhibits trophic dilution, where concentrations decrease at higher trophic levels due to lower bioavailability and faster excretion rates [65]. Terrestrial food chains show distinct patterns of lead accumulation; for example, European raptors — especially obligate scavengers like Griffon Vultures — exhibit the highest mean Pb concentrations in liver and kidney tissues due to consumption of lead-contaminated prey [66]. Moreover, bone Pb concentrations exceeding 20 mg/kg dry weight are linked to clinical symptoms, with raptors and scavengers being particularly vulnerable to Pb-induced mortality, and hunting ammunition identified as a significant source of exposure [66,67].
The mechanism of lead toxicity in animals involves the production of free radicals that cause oxidative stress and damage to essential biological molecules, including enzymes, proteins, lipids, and DNA [68]. In cattle, chronic Pb exposure induces nephrotoxicity through increased lipid peroxidation and decreased antioxidant enzyme activity, with renal dysfunction occurring at concentrations as low as 10 g/dL [69]. Studies on wood mice from contaminated mining areas revealed that 93% of individuals at mine sites and 76% at private properties exceeded bone Pb thresholds (25 mg/kg dry weight) associated with kidney damage [70].
The ecological and public health implications of Pb contamination extend beyond individual animal toxicity to encompass population-level effects and concerns regarding food safety. Contaminated forage crops can lead to significant lead accumulation in grazing animals. For instance, cows grazing on polluted pastures showed mean Pb concentrations of 15 ± 2.6 µg/kg in milk, representing 75% of the maximum limit suggested by Codex Alimentarius [71]. Studies on farm ruminants showed that buffaloes consistently displayed the highest Pb levels in blood (up to 2.47 mg/l). At the same time, sheep exhibited the highest concentrations in wool hair samples (ranging from 1.05 to 2.35 mg/kg) [72]. In poultry, bioaccumulation primarily occurs in the kidneys, liver, reproductive organs, and lungs, with studies in various countries reporting varying tissue-specific concentrations [73]. The transfer of Pb through food chains poses significant human health risks, with studies in Middle Eastern countries frequently reporting Pb levels in red meat exceeding FAO and WHO permissible limits [74], emphasizing the critical need for comprehensive monitoring and mitigation strategies across agricultural and wildlife management systems.
3.6. Pb Accumulation and Toxicity on Human
Humans are primarily exposed to lead through the inhalation of polluted air and dust particles, ingestion of contaminated water or food, and, to a lesser extent, through dermal absorption [75]. Once absorbed, lead accumulates in the human body through bioaccumulation and biomagnification processes, with approximately 90% of absorbed lead stored in mineralized tissues, particularly bones and teeth, where it can persist for decades by displacing calcium in hydroxyapatite crystals [2,75]. The respiratory tract efficiently absorbs micron lead particles at approximately 69% efficiency, while gastrointestinal absorption occurs at 10-15% in adults and at higher rates in children, primarily in the duodenum [2]. Children are particularly vulnerable due to higher absorption rates and greater susceptibility to lead's toxic effects, as lead is not absorbed by their bones as efficiently as in adults, leaving more circulating in soft tissues [6].
Lead toxicity manifests systemically, affecting virtually every organ system including the nervous system (causing neurotoxicity, cognitive impairment, and behavioral disorders), cardiovascular system (hypertension, increased risk of stroke and coronary heart disease), renal system (nephrotoxicity and chronic kidney disease), reproductive system (reduced fertility, sperm abnormalities, pregnancy complications), and hematopoietic system (anemia and altered blood cell parameters) [2,6,76] (Figure 5). The World Health Organization has established that no level of lead exposure is considered safe for human health, with the international level of concern set at 10 μg/dL in blood. Centers for Disease Control and Prevention USA has further restricted the permissible level of Pb in blood in children from 20 to 5 μg/dL. However, adverse effects can occur at even lower concentrations [76]. Lead's persistence in biological systems, combined with its ability to cross critical physiological barriers, accumulate in various organs, and perturb protein function, makes it particularly hazardous to human health [77,78].
3.6.1. Pb Distribution in the Human Body
Understanding the distribution patterns of lead within the human body following different routes of exposure (Figure 6) is crucial for comprehensive risk assessment and the development of effective prevention strategies.
3.6.1.1. Inhalation
Lead exposure through inhalation represents a significant pathway for human contamination, particularly in occupational and urban environments where anthropogenic sources such as industrial processes, particularly in sectors like welding and alloy smelting processes, fossil fuel combustion, and waste incineration release these toxic elements into the atmosphere [79,80]. Once inhaled, lead is primarily absorbed from the respiratory system, although it can also be absorbed from the digestive system when larger particles are swallowed, making inhalation a dual-route exposure mechanism [75,76]. In the blood, lead is primarily transported into erythrocytes by an anion exchanger and interacts with intracellular proteins like aminolevulinic acid dehydratase (ALAD), which serves as a major lead-binding site in red blood cells [75].
Lead concentrations in ambient air vary dramatically by location and source proximity, ranging from 0.05-0.1 µg/m³ in rural Mexican cities to 0.3-1.0 µg/m³ in major developing country cities [2]. Educational and office environments typically show low exposures of 0.02-0.274 µg/m³, while tobacco cafés demonstrate elevated levels of 0.7-0.9 µg/m³ [80]. Urban-industrial disparities are evident in Polish cities, where Pb particles smaller than 1 µm averaged 7.1 ± 0.46 µg/m³ in Warsaw versus 19 ± 1.4 µg/m³ in industrial Zabrze [79]. The highest concentrations occur near anthropogenic sources, with foundry areas exceeding 10 µg/m³ and Chinese lead-zinc smelters reporting annual averages of 8.05 μg/m³ Pb in air with winter peaks of 34.6 μg/m³ [2]. These concentrations are regulated by WHO annual limits of only 0.5 µg/m³, EPA three-month limits of 0.15 µg/m³, and OSHA occupational limits of 50 µg/m³, with successful interventions demonstrated by Korea's reduction from 0.34 μg/m³ in 1991 to <0.03 μg/m³ by 2018 following leaded gasoline prohibition [2].
3.6.1.2. Ingestion
Once ingested, Pb is absorbed through the gastrointestinal tract, with absorption rates varying significantly based on the chemical form and population demographics [81]. The gastrointestinal tract serves as the critical entry point for lead absorption from contaminated foods, where the metal undergoes oxidation and forms stable bonds to enzymes and protein molecules [81]. Following absorption, Pb is transported systemically via the bloodstream, primarily bound to erythrocytes, and subsequently distributes to various tissues and organs throughout the body [77]. Organic lead demonstrates substantially higher absorption rates at approximately 90%, accumulated in bone tissue, where it replaces calcium ions, leading to reduced bone mineral density and prolonged retention, while inorganic lead absorption ranges from 40-50% in children, who exhibit markedly higher intestinal absorption capabilities compared to adults [81]. This enhanced absorption in pediatric populations is particularly concerning, given that children consume three times as much food relative to their body mass compared to adults [82,77].
International regulatory standards have established maximum permissible limits for lead in various food categories: 0.01 mg/kg in infant products and natural mineral waters, 0.02 mg/kg in milk and dairy products, and 0.1 mg/kg in fruits, vegetables, meat products, and wine [81]. However, monitoring studies have documented concerning exceedances of these limits, with apple samples from Ukraine and Kosovo, lettuce and red potatoes from Romania, milk and dairy products from Egypt, and various edible oils (olive, rapeseed, sesame, sunflower, corn, cottonseed, and soybean) from multiple countries all showing lead concentrations above regulatory thresholds (Table 4) [81].
Dietary fiber intake can modulate lead absorption by binding to Pb within the digestive tract, facilitating its excretion through feces and reducing systemic bioavailability [77]. Elimination of absorbed inorganic lead occurs primarily through renal excretion (approximately 75%) and fecal elimination (about 16%), though the metal's slow excretion rate and prolonged retention time, especially in bone tissue, result in chronic bioaccumulation over extended periods [81].
3.6.1.3. Permeation
Dermal penetration of lead represents a significant yet under-studied exposure route that can contribute substantially to systemic lead burden [83]. While the outermost protective layer of the skin, the stratum corneum, generally restricts large-scale penetration, traces of lead present in various sources can reach the circulatory system through skin appendages or via transcellular and intracellular pathways [84]. The vast majority of studies (92% of 24 study summaries) have reported detectable levels of dermal absorption of inorganic lead compounds, including both water-soluble and water-insoluble forms, with average calculated diffusion rates for animal and human skin data ranging from 10⁻⁷ to 10⁻⁴ mg cm⁻² h⁻¹ and permeability coefficient (Kp) values ranging from 10⁻⁷ to 10⁻⁵ cm h⁻¹ [83]. These dermal exposures demonstrate significant potential impact, as they could potentially elevate blood lead levels by over 6 µg dl⁻¹, representing more than 100% of the 5 µg dl⁻¹ blood lead level associated with adverse health effects in adults [83]. Occupational exposure studies have documented measurable dermal lead concentrations, with the highest detected concentration of 6.91 ng/cm² found in workers involved in stamping processes, corresponding to an Average Daily Dose of 301.59 ng/kg-day [85].
The bioaccumulation potential is particularly concerning given that some data suggest a "reservoir effect", where lead may accumulate in skin layers and then act as a source for extended systemic exposure, highlighting the importance of comprehensive risk assessment that considers dermal penetration alongside traditional inhalation and ingestion pathways [83].
3.6.2. Harmful Effects on Human Organs and Systems
Heavy metals exert toxic effects on various human organs and biological systems. Upon exposure, heavy metals can cause pulmonary, cardiovascular, hepatic, gastrointestinal, dermatological, and nervous system disorders. Central to their toxicity is the generation of ROS, which induces oxidative stress, leading to DNA damage, protein misfolding, lipid peroxidation, and membrane disruption. These molecular alterations contribute to neurotoxicity, carcinogenesis, impaired DNA repair, loss of cellular functions, and overall cell damage (Figure 7).
3.6.2.1. Effects on the Blood and Circulatory System
Chronic lead exposure is recognized as a significant risk factor for cardiovascular diseases (CVD), contributing substantially to global CVD deaths and disability-adjusted life-years (DALYs), with an estimated burden of 5.5 million deaths per year in 2019 [86]. Lead exposure affects the blood and circulatory system through multiple interconnected mechanisms, with approximately 99% of blood lead concentration bound to red blood cells [87]. The most extensively studied cardiovascular effect is elevated blood pressure,
where a meta-analysis demonstrated that a 10 µg/g increase in bone lead was associated with a 0.26 mmHg increase in systolic blood pressure, while population studies have shown that declines in blood lead levels greater than 0.1 μg/dL result in marked reductions in systolic blood pressure [88,89]. Even low-level lead exposure around 2 μg/dL increases the risk of hypertension, peripheral arterial disease, and renal dysfunction, with estimated mortality relative risk from CVD being higher for subjects with blood lead levels above 5–9 μg/dL [87]. Participants experiencing the highest tertile of blood lead decline (>0.91 μg/dL) compared to the lowest tertile (<0.27 μg/dL) demonstrated a mean difference of -7.08 mm Hg in systolic blood pressure change [89]. Lead exposure results in increased blood pressure through oxidative stress, where chronic exposure intensifies the deactivation of nitric oxide by reactive oxygen species, leading to functional nitric oxide deficiency and compensatory upregulation of nitric oxide synthase [88].
Atherosclerosis serves as the major mediator of CVD from lead exposure, with studies showing that a 10-μg/L increase in blood lead is associated with a 7% increase in the risk of high coronary artery calcium score (≥100) in men, indicating significant coronary artery calcification [86]. Lead exposure contributes to ischemic heart disease, which accounts for the largest proportion of lead exposure-attributable CVD deaths and DALYs, and stroke, representing the second most significant cause of lead-related cardiovascular mortality [88]. The metal damages the blood vessel inner lining, promoting inflammation and plaque formation that narrows blood vessels, while also affecting cardiac structure through associations with decreased interventricular septum thickness [88,89].
At the cellular level, lead exposure interferes with cardiac conduction and contractility by reducing Na+/K+ ATPase pump activity, affecting myofibril phosphorylation, and disrupting signaling pathways, while also promoting lipid metabolism alterations, including increased triglycerides, lipid peroxidation, and decreased HDL cholesterol [87]. Lead exposure is also associated with anemia risk at high exposure levels, and the measurement accuracy of blood lead can be improved by correcting for hemoglobin or hematocrit levels, as lower erythrocyte volume fraction results in underestimated blood lead values [86].
3.6.2.2. Effects on the Reproductive System
Lead exposure during critical developmental periods, particularly puberty, causes lasting reproductive consequences in both males and females, including delayed sexual maturation, altered hormone profiles, and reduced reproductive capability [90]. Lead disrupts the hypothalamic-pituitary-gonadal axis in both males and females, interfering with essential hormone synthesis and release pathways that regulate reproductive function [91,92]. Lead exposure caused significant changes in serum sex hormone levels, with progesterone decreasing by 80.2% and testosterone by 49.9%, while estradiol increased by 69.8% in the highest dose group [90]. Furthermore, a meta-analysis found a statistically significant association between paternal lead exposure and congenital anomalies with a pooled odds ratio of 2.09 (95% confidence intervals: 2.09-3.35; P < .01) [92].
In males, lead exposure causes impaired spermatogenesis and disrupts the release of testosterone and luteinizing hormone, essential for sperm production and maturation [92]. Blood lead levels exceeding 40 µg/dL of Pb have been associated with significant impairments in male reproductive health, including reduced sperm count and concentration, decreased semen volume, diminished motility and vitality, and abnormal morphology, as demonstrated by both individual studies and a systematic review and meta-analysis [91,93]. Studies have shown a dose-response relationship between blood and seminal lead levels and worsening semen parameters, with even relatively low blood levels affecting male fertility [93].
In females, lead exposure adversely affects reproductive function by disrupting ovarian physiology, including impairments in follicle formation, oocyte maturation, ovulation, and corpus luteum development. These disruptions contribute to reduced fertility, extended time to conception, and adverse pregnancy outcomes such as spontaneous abortion, stillbirths, and developmental abnormalities [91]. Additionally, epidemiological evidence links lead exposure to broader reproductive issues, including infertility, premature birth, and early menopause [90]. At the cellular level, lead induces oxidative stress in ovarian tissue by generating ROS, which damages DNA, lipids, and proteins, potentially causing genomic instability. This oxidative damage, combined with lead-induced inflammatory responses, may further contribute to pregnancy complications [91,92]. The molecular mechanisms underlying lead's reproductive toxicity include the activation of endoplasmic reticulum stress-related signaling pathways, particularly the IRE1α-JNK signaling pathway, leading to increased follicular atresia and decreased ovarian reserves through apoptosis [90,91].
3.6.2.3. Effects on the Respiratory System
Adults are mainly exposed by breathing lead-contaminated fine particulate matters and fumes, with respirable particles capable of deep lung penetration and deposition in the alveoli, readily passing the air-blood barrier for systemic distribution [94]. Prenatal and postnatal lead exposure has been associated with adverse respiratory health outcomes in children, including increased risk for atopic sensitization to common aeroallergens at 5 years of age and lung function deficits [95].
Several studies indicate that environmental lead pollution and lead exposure negatively impact the respiratory system in humans. In a study of 200 children aged 5–14 with bronchial asthma, higher blood lead levels were associated with increased asthma severity, as well as higher frequencies of eosinophilia and elevated IgE levels. Asthmatic children with elevated blood lead showed significantly higher rates of eosinophilia (66.7%) and increased total IgE (83.3%), along with more severe asthma compared to children with normal blood lead levels. Another cross-sectional study, involving 1,788 children from the National Health and Nutrition Examination Survey (2005–2006), also found an association between blood lead levels and serum IgE levels, eosinophil counts, and asthma prevalence. Additionally, higher frequency of asthma and respiratory symptoms were observed in industrial workers in the United Arab Emirates exposed to lead, compared to non-industrial workers, and a correlation between blood lead levels and asthma progression was reported among Caucasian Americans [96].
3.6.2.4. Effects on the Kidney System
Lead exposure poses significant nephrotoxic risks across multiple pathways, with the kidney serving as a primary target organ due to its role in lead excretion [97]. Chronic lead exposure results in accumulation in renal tissue, leading to continuous insults that manifest as proximal tubular dysfunction, chronic interstitial nephritis, and progressive chronic kidney disease (CKD), potentially culminating in end-stage renal failure [98]. Blood lead levels even below 5 µg/dL negatively affect kidney function, with the lowest level associated with longitudinal changes in estimated glomerular filtration rate (eGFR) documented at 2.2 µg/dL [99]. Elevated blood lead levels ≥1.5 µg/dL are significantly associated with increased CKD risk, maintaining significance after adjustment for confounders [100]. Meta-analysis reveals that chronic lead-exposed groups exhibit significantly higher blood lead levels averaging 25.6 µg/dL compared to unexposed populations, with elevated biomarkers including urinary N-acetyl-β-D-glucosaminidase (NAG), α1-microglobulin, β2-microglobulin, serum creatinine, and kidney injury molecule-1 (KIM-1) [98]. Pathological manifestations include glomerulosclerosis, tubular atrophy, interstitial fibrosis, and renal arteriosclerosis [97]. Lead exposure exhibits a dose-response relationship with kidney stone formation, where a 1-unit increase in urinary lead levels is associated with a 7% increased risk, and participants in the highest quartile have a 64% higher risk of kidney stones compared to those in the lowest quartile [97]. Among patients with advanced CKD (eGFR <43 mL/min/1.73 m²), each 1 µg/g increase in tibial lead concentration correlates with 0.12 g/dL lower hemoglobin concentration and 23% higher odds of anemia, indicating lead's role in CKD-related myelosuppression [101]. Systematically, lead enters proximal tubular cells through endocytosis, inhibits mitochondrial respiration, generates reactive oxidants, depletes intracellular glutathione, and triggers programmed cell death while binding to calcium-sensitive receptors and inducing hypercalciuria [99]. The nephrotoxic effects are mediated through oxidative stress, inflammation, activation of NF-κB, renin-angiotensin system activation, and macrophage recruitment, culminating in tubulointerstitial damage and progressive renal dysfunction [99,102].
3.6.2.5. Effects on the Bones
In bones, lead’s biological half-life lasts up to 30 years [103]. The metal exerts detrimental effects on bone health through multiple mechanisms, including its ability to replace calcium in hydroxyapatite crystals due to higher affinity for bone sialoprotein than calcium [104], inhibition of vitamin D activation and dietary calcium absorption [105], and cytotoxic effects on osteoblasts, osteoclasts, and chondrocytes [103]. Studies demonstrate significant negative correlations between lead exposure and bone mineral density (BMD), with urinary lead levels showing correlation coefficients of -0.015 for total bone density and -0.019 for lumbar spine bone density in adults [104]. In children and adolescents, every 1 mg/dl increase in blood lead levels corresponds to BMD decreases of 0.011 g/cm² at the total spine, 0.008 g/cm² at the total femur, and 0.006 g/cm² at the femur neck [105]. Lead exposure demonstrates site-specific effects, with lumbar spine BMD showing consistently negative associations while femoral BMD effects vary by study population [106]. Age and gender significantly influence lead's impact on bone health, with stronger correlations observed in individuals ≥39 years and in women, particularly postmenopausal women where lead may induce BMD decreases through estrogen level reduction [104]. Direct tissue analysis reveals significantly elevated lead levels in osteoporotic patients, with median bone tissue concentrations of 1.67 mg/kg in osteoporotic subjects compared to 0.57 mg/kg in healthy controls (p = 0.002), and corresponding negative correlations between bone lead levels and BMD of the total femur and femoral neck (rho = -0.33, p = 0.03) [103]. These findings demonstrate that lead exposure represents a significant risk factor for osteoporosis development, with effects detectable at low exposure levels, for children – even within current reference ranges [105].
3.6.2.6. Effects on the Liver and Intestines
The liver serves as a significant reservoir of lead among soft tissues, holding approximately 33% of the lead content, and chronic exposure results in statistically significant alterations in serum liver enzymes including increased levels of lactate dehydrogenase (LDH), aspartate aminotransferase (AST), anaplastic lymphoma kinase (ALK), alanine aminotransferase (ALT), and bilirubin, alongside decreased levels of total serum protein, albumin, and globulin [107]. Lead toxicity operates through multiple mechanisms including oxidative stress generation, lipid peroxidation, inflammatory cytokine enhancement (IL-1β, IL-6, TNF-α), and interference with essential cellular processes, with even very low-dose exposure (median blood lead level of 0.9 μg/dl) potentially causing organ injury [108,109]. Studies demonstrate that the proportion of advanced liver fibrosis significantly increases with blood lead level, with higher concentrations found in fibrotic groups (1.2 μg/dl vs 0.9 μg/dl, p < 0.001), and blood lead levels remain an independent risk factor for advanced liver fibrosis even after controlling for confounding factors (OR = 1.249; 95% CI, 1.048–1.489; p = 0.013) [108]. The duration of lead exposure correlates significantly with the degree of liver fibrosis, with 26.7% of exposed individuals showing liver stiffness values above the significant fibrosis cut-off level [109]. Table 5 outlines selected hepatotoxic effects of lead exposure as observed in various in vivo studies utilizing animal models.
Regarding intestinal effects, lead causes gut dysbacteriosis by altering gut microbiota composition at phylum, family, and genus levels, destroys gut physiological homeostasis through local oxidative stress and inflammation. Pb also increases gut permeability by reducing tight junction proteins expression including peripheral membrane protein Zonula occludens-1 (ZO-1), Zonula occludens-2 (ZO-2), transmembrane proteins claudin-1 and occludin [111]. Lead exposure impairs intestinal absorption of essential nutrients, reducing glucose, glycine, lysine, and phenylalanine absorption, while disrupting the transport of divalent mineral elements like iron, calcium, and zinc by hijacking transporters such as Divalent metal transport 1 (DMT1) [111]. The gut-liver axis becomes compromised as dietary lead enters the portal vein through enterohepatic circulation, leading to disruption of hepatic lipid metabolism and dose-dependent increases in mRNA levels of genes related to fatty acid transport, β-oxidation, and triglyceride production [111].
3.6.2.7. Effects on the Central Nervous System
The neurotoxic mechanisms underlying lead's deleterious effects involve complex neurochemical, molecular, and morphological changes that converge to influence neuronal activity and functional neural circuits [112]. Lead acts as a potent non-competitive antagonist of N-methyl-D-aspartate (NMDA) receptors, this inhibition leads to reduced brain-derived neurotrophic factor release, negatively impacting synaptogenesis and neuronal networks, ultimately resulting in impaired learning and memory abilities [113,114]. Lead's ability to substitute for calcium ions facilitates its passage across the blood-brain barrier and disrupts calcium-dependent processes throughout the brain [115]. Additional mechanisms include oxidative stress generation through ROS production, mitochondrial dysfunction leading to decreased ATP levels, interference with neurotransmitter systems particularly affecting GABA and glutamate release, and disruption of myelin formation [112]. Chronic exposure results in neuronal loss, reduced cortical thickness, deficient dendritic development, and decreased neurogenesis in the adult hippocampus [112]. Meta-analysis evidence indicates significantly increased risk for malignant brain cancers associated with occupational lead exposure (pooled OR = 1.13, 95% CI: 1.04-1.24) [116].
Developmental neurotoxicity constitutes the most critical health effect, as prenatal and early postnatal exposure triggers irreversible alterations in CNS structure and function [112]. The developing brain demonstrates heightened vulnerability due to its rapid growth, immature blood-brain barrier, and high rates of cellular development [112]. Even low-level exposure below 10 μg/dL blood lead concentration produces significant cognitive impairments, with no evidence for a safe threshold identified [115]. Studies demonstrate that an increase in blood lead from <1 to 30 μg/dL associates with approximately 9 IQ point deficits, with the largest portion (~6 IQ points) occurring below 10 μg/dL [114]. An integrated analysis revealed a 6.9 IQ point decrement associated with increased concurrent blood lead levels from 2.4 to 30 μg/dL [115]. Lead exposure during elementary school age demonstrates maximal detrimental effects on IQ, resulting in deficits ranging from 5-7 points to up to 15 points in cognitive assessments [112].
3.6.2.8. Effects on the Immune System
The immune system demonstrates particular vulnerability to lead toxicity, with blood lead concentrations as low as 3.5 µg/dL causing significant immunological alterations in children [117]. Occupational exposure studies reveal dramatic effects, with shipyard workers showing median blood lead levels of 37.07 µg/dL — 8.7-fold higher than controls at 4.3 µg/dL — and experiencing an 8.4% reduction in phagocytic activity, a 34% decrease in cytotoxic T cells (CD3+CD8+), and a 2.7-fold increase in regulatory T cells, alongside a 2-fold elevation in the IL-4/IFN-γ ratio indicating a shift toward humoral immunity [118]. At the cellular level, neutrophils accumulate the highest lead levels among immune cells and suffer vesicular damage through glutathione/multidrug resistance-associated protein (GSH/MRP) mediated sequestration, while CD4+ T cells, characterized as thiol-poor, experience severe ribosomal damage with reduced 18S rRNA levels [119]. Immunological perturbations involve oxidative stress mechanisms, inflammatory pathway activation including Mitogen-Activated Protein Kinase (MAPK), which is a type of serine/threonine-specific protein kinase involved in directing cellular responses, and NF-κB signaling (crucial intracellular pathway that regulates the expression of a wide range of genes), and result in immune dysregulation that increases susceptibility to infections, autoimmune diseases, and cancer [118,120].
4. Molecular Mechanisms of Pb Toxicity
4.1. Ion Mimicry and Cellular Disruption
Lead toxicity is driven by ion mimicry, wherein Pb²⁺ imitates essential divalent cations such as Ca²⁺, Mg²⁺, Zn²⁺, and Fe²⁺ due to similar biophysicochemical properties, allowing it to compete with or displace them at critical binding sites in proteins and enzymes, thereby disrupting their normal function [121,78]. Unlike these essential metals, however, Pb²⁺ cannot fulfill their physiological roles, making its substitution particularly harmful to mammalian cells. Pb²⁺ binds to functional sites normally occupied by calcium, magnesium, or zinc in key proteins such as calmodulin (calcium-binding protein), protein kinase C (play a crucial role in signal transduction pathways), troponin C (regulatory protein of striated muscle contraction,), and various synaptic proteins, with some Ca²⁺-binding proteins exhibiting even greater affinity for lead than for calcium [121]. In neurons, Pb²⁺ interferes with voltage-gated and receptor-operated calcium channels, impairing synaptic development and downstream signaling pathways, with potentially lasting neurotoxic effects [78].
Pb²⁺ competes with Zn²⁺ binding sites of several Zn²⁺ finger proteins (presence of zinc ions) and the NMDA receptor channel [121]. The metal also targets displacement of Fe²⁺, particularly in the DMT1, which may facilitate Pb²⁺ transport and uptake. Pb²⁺ exposure increases Fe²⁺ content in rat brain while decreasing ferroportin 1 (FP1), an Fe²⁺ efflux protein [121].
4.2. Mitochondrial Dysfunction and Energy Metabolism
Mitochondria represent primary targets of lead toxicity, with Pb²⁺ accumulating in the mitochondrial matrix and promoting imbalance in mitochondrial homeostasis [121]. Lead exposure remarkably reduces activities of mitochondrial respiratory chain enzyme complexes, especially complex II and complex III, impacting oxidative phosphorylation and decreasing ATP synthesis [122]. Studies have demonstrated that Pb exposure damages mitochondrial membrane potential, leading to decreased ATP production in rat proximal tubular cells, while chronic exposure causes mitochondrial edema in mice hepatocytes [123].
Lead disturbs intracellular calcium homeostasis through competition with Ca²⁺ for plasma membrane transport systems, including Ca²⁺ channels and pumps [122]. This leads to subcellular calcium redistribution, increasing Ca²⁺ levels in cytoplasm and mitochondria while decreasing endoplasmic reticulum (ER) Ca²⁺ [122]. Pb exposure increases expression of IP3R-1 and IP3R-2, enhancing ER Ca²⁺ release and subsequently increasing mitochondrial Ca²⁺ concentration [122].
The pathological opening of the mitochondrial permeability transition pore (MPTP) represents a critical mechanism of lead-induced cellular damage (Figure 8) [122]. This abnormal MPTP opening results in a significant decrease in mitochondrial membrane potential, ultimately leading to cell death through apoptosis [122]. Lead exposure alters expression of MPTP components, including voltage-dependent anion channel (VDAC), adenine nucleoside translocator protein (ANT), and cyclophilin-D (Cyp-D), while significantly increasing expression of pro-apoptotic protein Bax and decreasing anti-apoptotic proteins Bcl-2 and Bcl-xL [122].
4.3. Oxidative Stress and Antioxidant Depletion
Lead exposure generates ROS and reactive nitrogen species (RNS) either directly or indirectly within mitochondria [122]. The brain demonstrates particular susceptibility due to high oxygen consumption, low antioxidant defenses, abundance of polyunsaturated fatty acids, and presence of redox-active metals like Fe²⁺ [121].
Lead significantly inhibits the mitochondrial antioxidant system, including activities of GPx, SOD, CAT, and glutathione (GSH) [122]. Pb demonstrates high affinity for reactive -SH groups of GSH, effectively decreasing GSH levels [76]. The metal inhibits antioxidant enzymes by competing for essential metal cofactors like Zn²⁺ and Cu²⁺ [121].
Lead contributes to free radical generation through pro-oxidative effects of δ-aminolaevulinic acid (δ-ALA), which accumulates due to Pb²⁺-induced inhibition of
the enzyme ALA dehydratase (an enzyme known also as porphobilinogen synthase that plays a crucial role in the biosynthesis of heme, chlorophyll, and vitamin B12) [121]. This leads to increased levels of oxidative stress markers including malondialdehyde (MDA) and H₂O₂ concentrations [76]. Lipid peroxidation and protein carbonylation serve as significant indicators of oxidative stress under Pb exposure [122].
4.4. Neuroinflammation and Immune Response
Lead exposure stimulates neuroinflammation through activation of microglia and astroglia with subsequent release of inflammatory mediators [121]. Pb-induced mitochondrial damage promotes inflammation potentially via TLR4-stimulated signaling cascades [122]. In general, TLRs are an important family of pattern recognition receptors (PRRs) that recognize exogenous and endogenous pathogens, heat shock proteins and different components of the extracellular matrix. MPTP opening releases Mitochondrial Damage-Associated Molecular Patterns (MTDs), including the circular mtDNA, ATP, cardiolipin, and formyl-peptides, which act as DAMPs and activate pattern recognition receptors like TLR4 and TLR9 [122]. Cardiolipin is 1,3-bis(sn-3’-phosphatidyl)-sn-glycerol is an important component of the inner mitochondrial membrane responivble for cytochrome-C-Oxidase quaternary structure while formyl-peptides, also located in mitochondria play a role in the initiation of inflammation by activating the formyl-peptide receptor (FPR). Astroglial and microglial activation also associates with elevated levels of NADPH oxidase (NOx), the main non-mitochondrial ROS source activated by Pb²⁺ [121]. Lead exposure triggers assembly and activation of NLRP3 inflammasomes (multiprotein complexes that plays a pivotal role in regulating the innate immune system and inflammatory signaling) leading to pyroptosis through pro-caspase-1 cleavage and maturation of IL-1β and IL-18 [122]. Pyroptosis a highly inflammatory-driven, caspase-1 dependent programmed lytic cell death that is often caused by the invasion of a intracellular pathogen.
The TLR4-MyD88-NF-κB axis represents a significant pathway in Pb-induced inflammation [122]. Lead increases expression of TLR4 and MyD88 (Myeloid differentiation 88 protein, with important role in the immune system), activating downstream molecules including IRAK-4, IRAK-1 (IRAK - interleukin-1 receptor-associated kinase), involved in innate immune responses), and TRAF6, ultimately leading to NF-κB (a key modulator of innate and adaptive immune responses) activation [122]. Activated NF-κB and P38 MAPK (Mitogen-Activated Protein Kinase) pathways translocate to the nucleus, upregulating expression of cytokines including TNF-α, IL-1, IL-6, IL-8, IL-10, and IL-18 and provoking chronic inflammation [122,76].
4.5. DNA Damage and Genotoxicity
Lead induces genotoxic effects primarily through oxidative stress and inhibition of DNA repair mechanisms [125]. The accumulation of ROS impairs energy metabolism and causes various DNA alterations, including fragmentation, rearrangements, deletions, and point mutations [78]. Additionally, lead can directly interact with nuclear proteins and DNA, leading to site-specific damage and further ROS production [78]. One contributing factor is the accumulation of δ-ALA, which promotes ROS generation and oxidative DNA damage, such as the formation of 8-hydroxy-2′-deoxyguanosine (8-OHdG) — a widely recognized biomarker of oxidative stress that shows a positive correlation with lead exposure [125].
Lead interferes with DNA processing and repair enzymes by substituting essential metals like calcium and zinc [125]. Studies demonstrate altered expression of DNA repair genes due to Pb toxicity, with significant decreases in expression of genes involved in base excision repair (BER), nucleotide excision repair (NER), and double-strand break repair [125]. Specific genes including OGG1 (encodes the enzyme 8-oxoguanine DNA glycosylase responsible for the excision of 8-oxoguanine, a mutagenic base byproduct which occurs as a result of exposure to reactive oxygen), APE1(encodes multifunctional protein involved in DNA repair), and XRCC1 (BER pathway), XPD (xeroderma pigmentosum group D (XPD) protein with a function in nucleotide excision repair of DNA damage caused by UV radiation) and POLD1 (NER pathway), and BRCA1 (homologous recombination) show down-regulation following Pb exposure [125].
4.6. Epigenetic Modifications
Lead acts as an epigenetic toxicant by altering DNA methylation, histone modifications, and the expression of non-coding RNAs [126]. DNA methylation dysregulation involves changes in 5-methylcytosine (5mC) levels and abnormal methylation of genes involved in neurodevelopment, cognitive function, and cell cycle regulation [126]. Lead exposure alters expression levels of DNA methyltransferases (DNMTs) including Dnmt1, Dnmt3a, and Dnmt3b, along with demethylases and methyl-binding proteins like MeCP2 [126].
Histone modifications include methylation and acetylation of N-terminal tails of histones H3 and H4 [126]. Lead exposure causes significant differences in levels of post-translational histones’ modifications H3K9/14ac and H3K9me3 (acetylation of specific lysine residues), with decreased H3K27me3 levels observed in hippocampus of lead-exposed rodents [126]. Lead treatment results in HDAC2 (Histone Deacetylase 2) upregulation and Ac-H3K9 (modification of Histone H3, indicating the acetylation at the 9th lysine) downregulation, with HDAC2 considered essential for regulating lead-induced neurotoxicity [126].
MicroRNA alterations include upregulation of miR-520c-3p, miR-211, miR-148a, and miR-572 in occupationally exposed Chinese workers [127]. miR-520c-3p, miR-211, miR-148a, and miR-572 are closely related with cancer miR-520c-3p recently known as a novel supressor of lung aadenocarcinoma, miR-211 was upregulated in varous tumors including gliomas), miR-148a acts as a tumor suppressorand showsagreat potential in crenal cancer therapy, and miR-572 (both squamous cell carcinom of the head and neck, and ovarian ncancer). Studies show significant upregulation of miR-155 and miR-221, with functional analysis predicting targets involved in cellular pathways including cell differentiation, development, and apoptosis [127].
Studies show significant upregulation of miR-155 and miR-221, with functional analysis predicting targets involved in cellular pathways including cell differentiation, development, and apoptosis [127].
4.7. Autophagy and Cell Death Pathways
Lead triggers autophagy, particularly mitophagy (selective degradation of damaged mitochondria), via mitochondrial pathways [122]. Pb affects mitochondrial dynamics by altering expression of proteins including MFF (Mitochondrial fission factor controlling the division of mitochondria), Drp1 (dynamin-related protein 1, tightly regulated to clear the damaged mitochondria), MFN1 (mitofusin, functions as a tumor suppressor gene, activating the fusion process to depress the metastasis and invasion in vitro and in vivo), MFN2, and OPA1 (crucial regulators of the mitochondrial dynamics), increasing fission factors while decreasing fusion factors [122]. Lead exposure accelerates mitophagy via the PINK1/Parkin pathway (an autophagy machinery cleaning defected mitochondria), causing mitochondrial depolarization and accumulation of uncleaved PINK1 (a kinase which functions as a mitochondrial damage sensor) on outer mitochondrial membrane [122]. Lead exposure triggers non-apoptotic cell death pathways, including necroptosis — via upregulation of RIPK3 (lipid metabolism regulator contributing to inflammation and carcinogenesis in non-alcoholic fatty liver disease) and MLKL (Mixed lineage kinase domain like pseudokinase, an executioner of necroptosis) in olfactory cells. Besides, lead induce ferroptosis, by increasing free iron levels, downregulating GPX4 (Glutathione peroxidase, a selenonzyme that plays a critical role in maintaining oxidative homeostasis), and altering SLC7A11 (L-cystine/ L-glutamate transporter across the cell membrane) expression in the most commonly used in neuroscience research, especially for the Parkinson, cell line - PC12 [123].
5. Pb Resistance Mechanisms: Summary
Lead is lethal to bacterial cells even at small concentrations [34], at high concentrations Pb inhibits cell division, disrupts cellular respiration, and alters bacterial cell morphology and membrane integrity [38]. In response to this toxicity, organisms have developed sophisticated defense mechanisms (Figure 9) mediated by heavy metal resistance genes (HMRGs) [35]. Resistance is typically determined by measuring the minimum inhibitory concentration (MIC), with strains showing high Pb resistance often having an MIC greater than 1 mM [40].
5.1. Efflux and Active Transport Systems
Active transport of metal ions outside the cell represents a primary resistance mechanism across different organisms. PIB-type ATPases mediate efflux of lead, as observed in Pseudomonas species, which actively extrude metal ions to withstand heavy metal stress [34]. The zntA gene, encoding a P-type ATPase, is particularly significant and highly expressed in both rhizosphere and non-rhizosphere soils of milltailings regions [37]. Bacteria also employ RND efflux systems for exporting various metal ions [38].
Plants utilize active efflux mechanisms through ATP-dependent transporters at root level to limit heavy metal uptake, creating competition with other nutrients for absorption sites [60]. Once heavy metals enter the plant cytosol, they can be transported out of cells or restricted to vacuoles through sequestration involving transporter families like ABC (transport of sugars, amino acids, and metals), CDF (transport of divalent cadmium, zinc, and cobalt), HMA (heavy-metal-associated domain), and NRAMP (Natural Resistance-associated Macrophage Protein), transport of divalent iron and manganese) [60].
5.2. Metal Chelation and Sequestration
Metallothioneins are cysteine-rich proteins that sequester heavy metals through thiol groups, forming non-toxic complexes in bacterial, yeast, plant and animal, includng human, cells (Figure 10). For example, the genetically engineered Pseudomonas putida strain 15420352 overexpressing metallothioneins has shown enhanced capability to capture and immobilize Pb and Hg in wastewater, proving effective in pilot-scale bioreactors for treating industrial wastewater from mining and battery production [38]. Under lead stress, microorganisms adjust the expression of metallothionein genes by utilizing metal-sensing proteins, metal-responsive transcription factors, metal-binding proteins, and phosphorylation mechanisms to cope with fluctuating metal ion concentrations [37].
Siderophores, produced by bacteria such as Alcaligenes eutrophus and Pseudomonas aeruginosa, can reduce Pb toxicity. P. aeruginosa produces explicitly pyoverdine and pyochelin, which can block Pb absorption [38]. Enhanced siderophore production serves as a lead-resistant mechanism, typically produced to promote plant growth in iron-deficient environments while influencing heavy metal bioavailability [34].
Plants employ phytochelatins (PCs) as specialized metal-binding peptides under heavy metal stress. PCs are high-affinity ligands that bind to metal cations including Pb²⁺ to immobilize them, preventing interference with essential biochemical pathways and cell signaling [60]. Plants also synthesize organic acids to bind free metal ions, thereby minimizing toxic effects [60].
5.3. Biosorption and Surface Binding
In bacteria, biosorption involves the binding of metal ions to bacterial cell walls and their entrapment in extracellular capsules or exopolysaccharides (EPS), with functional groups such as hydroxyl, amino, amide, and phosphate playing key roles in heavy metal ion adsorption [34,37]. Bacillus megaterium plays a crucial role in detoxifying Cd and Pb through production of EPS [38]. Biofilms, composed of EPS, create protective niches for bacteria and bind heavy metals, thereby lowering their toxicity and bioavailability [38]. Binding of Pb²⁺ has been noted at the interface of Burkholderia cepacia biofilms and hematite (Fe³⁺ oxide) [35].
In fungi, cell surface binding and extracellular biosorption serve as the primary mechanisms for removing Pb(II) at higher concentrations, with extracellular polymeric substances or metabolic products coating the cell surface and facilitating lead removal through mechanisms such as surface binding, ion exchange, or precipitation [44]. Fourier transform infrared spectroscopy analysis identified various functional groups involved in biosorption, including O-H and N-H bonds from proteins, C-H stretching vibrations from hydrophilic lipid molecules, and protein components like Amide I (C=O stretching) and Amide II (NH₂ deformation) peaks [41].
Plant cell walls act as physical barriers where bivalent metal cations such as Pb²⁺ bind to carboxyl groups in pectin, restricting heavy metals to cell walls and preventing cellular entry [60]. Lead accumulates in cell walls of epidermal cells in certain plants [60].
The sorption capacity of certain microorganisms is presented in Table 6.
5.4. Bioaccumulation and Intracellular Sequestration
Bioaccumulation refers to the accumulation of metal ions inside cells in less toxic states or as intracellular precipitation, often being metabolic-activity dependent [34]. Intracellular sequestration involves complexation and storage of metal ions within the bacterial cytoplasm [38].
In fungi, intracellular accumulation plays an important role in Pb(II) removal particularly at lower concentrations (25-50 mg L⁻¹). Verticillium insectorum J3 can take up and accumulate Pb(II) ions inside fungal cells [44]. Scanning Electron Microscope Observation of Sarocladium Pb-9 indicated that Pb element was contained within the mycelium after treatment with Pb(NO₃)₂ [41].
5.5. Precipitation and Biotransformation
Lead precipitation represents an effective detoxification strategy across multiple organism types. Sulfate-reducing bacteria can precipitate Pb, and precipitation can be catalyzed by enzymes such as phosphatase enzyme (PbrB) [34]. The mechanism of MICP precipitates Pb in its carbonate form, serving as a resistance mechanism by sequestering Pb²⁺ and other toxic metals, thereby protecting microbial cells [40].
5.6. Morphological Adaptations
Morphological alterations play a critical role in microbial resistance to heavy metal stress. Under lead (Pb) exposure, bacteria often exhibit changes in cell shape, size, and membrane integrity as part of their adaptive response [38]. Pseudomonas species, in particular, demonstrate advanced survival strategies, including morphological differentiation, biosurfactant production, and the expression of stress-responsive proteins [37].
5.7. Genetic Regulation and Molecular Mechanisms
Bacterial tolerance involves genetic mutations targeting genes responsible for regulating metal ion absorption, efflux, and storage. Horizontal Gene Transfer (HGT) significantly contributes to metal tolerance by facilitating rapid spread of resistance genes across bacterial species through mobile genetic elements like plasmids and transposons [38].
Heavy metals can directly trigger the transcription of resistance genes, thereby initiating detoxification and tolerance responses [37]. Inorganic ion transport and metabolism play a crucial role in maintaining bacterial homeostasis and mediating heavy metal transport by regulating intracellular ion levels and metal metabolism [37].
6. Methods for Pb Removal
6.1. From the Environment
6.1.1. Physical and Chemical Methods
Traditional physicochemical approaches for lead removal include ion exchange, adsorption, membrane processing, reverse osmosis, filtration, electro-coagulation, chemical precipitation, nanofiltration, and coagulation-flocculation [130, 131]. However, these methods often prove ineffective or expensive, particularly at very low heavy metal concentrations below 100 mg/L, and physical separation methods cannot remove heavy metal salts dissolved in water [130,36].
6.1.1.1. Limestone-Based Adsorption
Limestone mining waste and its derived CaO (calcined limestone) demonstrate exceptional effectiveness for lead removal through adsorption mechanisms. Calcined limestone, prepared by grinding limestone and thermal decomposition at 1100°C for 2 hours, exhibits significantly higher surface area (14.3 m²/g) compared to pristine limestone (4.2 m²/g) [131]. Under optimized conditions, limestone achieved 98.9% Pb²⁺ removal efficiency at 2 g/L dosage, while calcined limestone achieved 100% removal at only 0.4 g/L dosage [131]. The optimal pH for both materials was 5, with maximum contact time of 30 minutes yielding 95.4% removal for limestone and 99.5% for calcined limestone [131]. The primary mechanism involves ion exchange, where reactions with calcium carbonate and calcium oxide result in precipitation of poorly soluble lead carbonates and hydroxides [131].
6.1.1.2. Nanotechnology Applications
Nanotechnology has emerged as a cost-effective solution for environmental remediation. Polyvinylpyrrolidone-coated magnetic nanoparticles (PVP–Fe₃O₄ NPs) demonstrate exceptional efficiency for lead removal, achieving nearly 100% removal at 0.1 mg L⁻¹ initial concentration and more than 80% at 1 mg L⁻¹ in both soft water and seawater using 167 mg L⁻¹ of nanoparticles [132]. The adsorption capacity reached 59.6–61.6 mg g⁻¹ in soft water, with lead binding occurring immediately and completely within the first 1.5 hours of mixing [132]. Lead exhibited higher adsorption affinity than other metals due to its larger ionic radius, and the kinetic data fitted well to the pseudo-second-order model, indicating covalent bonding through electron sharing or exchange [132].
6.1.2. Biological Remediation Methods
6.1.2.1. Microbial Bioremediation
Microbial bioremediation employs various mechanisms including biosorption, biotransformation, bioleaching, and bacterial immobilization [36]. Specific microbial species demonstrate high efficiency, with Penicillium janthinillum GXCR achieving 85–99% efficiency, Aspergillus flavus reaching 46–97%, Sporosarcina pastaurii attaining 94–98%, Variovorax boronicumulans showing 73–95%, and Stenotrophomonas rhizophila demonstrating 63–96% removal rates [36].
Three key microbial-mediated processes facilitate lead immobilization: PSB, SRM and MICP. Phosphate Solubilizing Bacteria (PSB) convert inorganic or organic phosphorus into reactive phosphate that binds lead, forming highly recalcitrant lead phosphate precipitates such as pyromorphite [40]. Sulfate Reducing Microorganisms (SRM) operate under anoxic conditions, using sulfate as terminal electron acceptor to produce hydrogen sulfide, which complexes with Pb²⁺ to form stable lead sulfide (PbS) [40]. Microbial Induced Carbonate Precipitation (MICP) is facilitated by ureolytic organisms with urease activity, converting urea into ammonium and carbonate under alkaline conditions, allowing Pb²⁺ to bind with carbonate forming insoluble lead carbonate (PbCO₃) [40].
A novel biological template utilizing extracellular bacterial protein (~30 kDa) isolated from hot spring bacterium MDH1 provides an eco-efficient route for lead removal. When this purified protein is added to lead nitrate [Pb(NO₃)₂] solutions, insoluble lead oxide nanoplates with thickness of 20-25 nm are formed, which can be easily separated by centrifugation and filtration [130]. Toxicity studies confirmed complete lead removal, as the filtered supernatant showed no zone of inhibition on E. coli growth [130].
6.1.2.2. Phytoremediation
Phytoremediation techniques utilizing hyperaccumulator plants (transfer factor >1) offer promising solutions for Pb-contaminated environments through phytoextraction, phytostabilization, and rhizofiltration approaches [52,51]. Jute demonstrates high accumulation in both roots (367.83 mg/kg) and shoots (370.43 mg/kg), while Flax shows 68.6% average Pb removal efficiency from soil [57]. Complementary strategies include biochar application for Pb immobilization, nanotechnology approaches, and plant growth regulators to enhance tolerance mechanisms [51]. Phytoremediation efficiency can be enhanced using chemical amendments such as EDTA chelators, which increase metal solubility and facilitate translocation to shoots [57].
Multi-species plant–microbe consortia have demonstrated superior effectiveness, with combinations of Rhizobium leguminosarum, Sinorhizobium meliloti, Pseudomonas fluorescens, Luteibacter sp., Variovorax sp., Bacillus simplex, and B. megaterium resulting in significant increases in Pb absorption by Lathyrus sativus [38].
Despite significant advances in understanding Pb-plant interactions, further research is needed to elucidate specific molecular mechanisms, develop transgenic plants with enhanced remediation capabilities using CRISPR/Cas9-mediated genetic modifications, and establish sustainable management practices for Pb-contaminated agricultural systems to ensure food safety and sustainable crop production [60,51,62].
6.1.3. Soil Amendment Strategies
Agricultural soil remediation employs various amendment strategies to immobilize lead and reduce bioavailability. Lime application using CaO, CaCO₃, or Ca(OH)₂ increases soil pH, decreasing bioavailability of cationic metals like lead through enhanced competition with calcium [32]. Phosphate minerals immobilize lead through pH increase, electrostatic interaction, ion exchange, surface complexation, and precipitation [32]. Clay minerals including sepiolite, attapulgite, bentonite, and kaolinite with negatively charged surfaces adsorb lead through complexation, lattice diffusion, and isomorphic substitution [32].
Biochar affects lead mobility through cation-π interactions, electrostatic attraction, ion exchange, complexation, and precipitation [32]. Combined zero-valent iron-biochar treatment significantly decreased lead content in rice plants [32]. Water management strategies significantly impact lead bioavailability, where flooding conditions decrease availability of cationic metals, while alternate wetting and drying effectively reduces lead accumulation compared to continuous ponding [32]. Sprinkler irrigation minimizes lead contents up to 50% in rice cultivars [32].
6.1.4. Advanced Technologies
Advanced microbial technologies encompass Microbial Fuel Cells (MFCs) for lead removal and recovery from wastewater, biofilm technology utilizing extracellular polymeric substances for metal sequestration, and nanomaterials like carbon nanotubes with exceptional adsorption capabilities [36]. Biotechnological tools such as genetic engineering and recombinant DNA technology can enhance microbial strains for improved lead bioremediation [36].
6.2. From the Human Body
Lead elimination occurs primarily through natural excretion in feces and urine [16], though once lead enters the body, it cannot be totally removed [6]. The most effective treatment is chelation therapy using CaNa2EDTA or Succimer (DMSA), with Succimer dosed at 350 mg/m² three times daily for five days, then twice daily for 14 days, though limited to severe cases due to side effects [6]. Dietary interventions with calcium, zinc, and iron supplementation help lower lead levels by counteracting lead's calcium analog behavior, while vitamins and flavonoids scavenge ROS generated by lead-induced oxidative stress [6]. Environmental regulations have reduced average blood lead concentrations in developed nations to 1.5 μg/dl, with studies showing 32% decreases over 5.2 years following reduced exposure [94].
7. Conclusion
Lead contamination represents one of the most pervasive and persistent environmental health challenges, affecting virtually every biological system through complex molecular mechanisms involving ion mimicry, mitochondrial dysfunction, oxidative stress, and DNA damage. With an estimated 900,000 deaths annually and no safe exposure threshold, lead poses irreversible neurodevelopmental risks in children and cardiovascular consequences in adults, persisting in bone tissue for decades after exposure. While organisms have evolved resistance mechanisms, including efflux systems, metal chelation, and biosorption, and promising remediation technologies such as nanotechnology applications, microbial bioremediation, and phytoremediation continue to advance, prevention through source control remains the most effective approach. The ultimate goal must be the elimination of lead exposure to the greatest extent possible, requiring sustained research, technological innovation, policy development, and international cooperation to address this preventable global health crisis and protect current and future generations from one of humanity's oldest and most persistent environmental toxins.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Acknowledgments
This work was conducted at UPIZ Educational and Research Laboratory of Biology-MF-NBU, New Bulgarian University, Sofia, Bulgaria.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| OSHA | Occupational Safety and Health Administration |
| RND | Resistance-Nodulation-Division (family) |
| MICP | Microbially Induced Carbonate Precipitation |
| ZIP | ZRT-IRT-like Protein (family) |
| NRAMP | Natural Resistance-Associated Macrophage Protein |
| ROS | Reactive Oxygen Species |
| FAO | Food and Agriculture Organization (of the United Nations) |
| GSH/MRP | Glutathione / Multidrug Resistance-associated Protein |
References
- Lead | Pb (Element) – PubChem. Available online: https://pubchem.ncbi.nlm.nih.gov/element/Lead (accessed on 11 June 2025).
- Gonzalez-Villalva, A.; Marcela, R.-L.; Nelly, L.-V.; Patricia, B.-N.; Guadalupe, M.-R.; Brenda, C.-T.; Maria Eugenia, C.-V.; Martha, U.-C.; Isabel, G.-P.; Fortoul, T.I. Lead Systemic Toxicity: A Persistent Problem for Health. Toxicology 2025, 515, 154163. [Google Scholar] [CrossRef]
- Levin, R.; Zilli Vieira, C.L.; Rosenbaum, M.H.; Bischoff, K.; Mordarski, D.C.; Brown, M.J. The Urban Lead (Pb) Burden in Humans, Animals and the Natural Environment. Environmental Research 2021, 193, 110377. [Google Scholar] [CrossRef]
- Satchanska, G. Toxicity of heavy metals and pesticides on the living world 2025, A Monograph, 103 pages.
- Soil contamination by heavy metals. Europa.eu. https://www.eea.europa.eu/en/analysis/maps-and-charts/soil-contamination-by-heavy-metals (accessed 2025-07-08).
- Collin, M.S.; Venkatraman, S.K.; Vijayakumar, N.; Kanimozhi, V.; Arbaaz, S.M.; Stacey, R.G.S.; Anusha, J.; Choudhary, R.; Lvov, V.; Tovar, G.I.; et al. Bioaccumulation of Lead (Pb) and Its Effects on Human: A Review. Journal of Hazardous Materials Advances 2022, 7, 100094. [Google Scholar] [CrossRef]
- Charkiewicz, A.E.; Backstrand, J.R. Lead Toxicity and Pollution in Poland. IJERPH 2020, 17, 4385. [Google Scholar] [CrossRef] [PubMed]
- Ericson, B.; Hu, H.; Nash, E.; Ferraro, G.; Sinitsky, J.; Taylor, M.P. Blood Lead Levels in Low-Income and Middle-Income Countries: A Systematic Review. The Lancet Planetary Health 2021, 5, e145–e153. [Google Scholar] [CrossRef]
- Bouida, L.; Rafatullah, M.; Kerrouche, A.; Qutob, M.; Alosaimi, A.M.; Alorfi, H.S.; Hussein, M.A. A Review on Cadmium and Lead Contamination: Sources, Fate, Mechanism, Health Effects and Remediation Methods. Water 2022, 14, 3432. [Google Scholar] [CrossRef]
- O’Shea, M.J.; Toupal, J.; Caballero-Gómez, H.; McKeon, T.P.; Howarth, M.V.; Pepino, R.; Gieré, R. Lead Pollution, Demographics, and Environmental Health Risks: The Case of Philadelphia, USA. IJERPH 2021, 18, 9055. [Google Scholar] [CrossRef] [PubMed]
- Borgulat, J.; Łukasik, W.; Borgulat, A.; Nadgórska-Socha, A.; Kandziora-Ciupa, M. Influence of Lead on the Activity of Soil Microorganisms in Two Beskidy Landscape Parks. Environ Monit Assess 2021, 193, 839. [Google Scholar] [CrossRef]
- Sukaiti, W.S.A.; Al Shuhoumi, M.A.; Balushi, H.I.A.; Faifi, M.A.; Kazzi, Z. Lead Poisoning Epidemiology, Challenges and Opportunities: First Systematic Review and Expert Consensuses of the MENA Region. Environmental Advances 2023, 12, 100387. [Google Scholar] [CrossRef]
- Wu, D.; Hu, Y.; Cheng, H.; Ye, X. Detection Techniques for Lead Ions in Water: A Review. Molecules 2023, 28, 3601. [Google Scholar] [CrossRef]
- Raj, K.; Das, A.P. Lead Pollution: Impact on Environment and Human Health and Approach for a Sustainable Solution. Environmental Chemistry and Ecotoxicology 2023, 5, 79–85. [Google Scholar] [CrossRef]
- Kumar, A.; Kumar, A.; M.M.S., C.-P.; Chaturvedi, A.K.; Shabnam, A.A.; Subrahmanyam, G.; Mondal, R.; Gupta, D.K.; Malyan, S.K.; Kumar, S.S.; et al. Lead Toxicity: Health Hazards, Influence on Food Chain, and Sustainable Remediation Approaches. IJERPH 2020, 17, 2179. [CrossRef]
- Huang, H.; Guan, H.; Tian, Z.-Q.; Chen, M.-M.; Tian, K.-K.; Zhao, F.-J.; Wang, P. Exposure Sources, Intake Pathways and Accumulation of Lead in Human Blood. Soil Security 2024, 15, 100150. [Google Scholar] [CrossRef]
- Ardila, P.A.R.; Alonso, R.Á.; Valsero, J.J.D.; García, R.M.; Cabrera, F.Á.; Cosío, E.L.; Laforet, S.D. Assessment of Heavy Metal Pollution in Marine Sediments from Southwest of Mallorca Island, Spain. Environ Sci Pollut Res 2023, 30, 16852–16866. [Google Scholar] [CrossRef]
- Al-Mutairi, K.A.; Yap, C.K. A Review of Heavy Metals in Coastal Surface Sediments from the Red Sea: Health-Ecological Risk Assessments. IJERPH 2021, 18, 2798. [Google Scholar] [CrossRef]
- Javid, A.; Nasiri, A.; Mahdizadeh, H.; Momtaz, S.M.; Azizian, M.; Javid, N. Determination and Risk Assessment of Heavy Metals in Air Dust Fall Particles. Environ Health Eng Manag 2021, 8, 319–327. [Google Scholar] [CrossRef]
- Luo, P.; Xu, C.; Kang, S.; Huo, A.; Lyu, J.; Zhou, M.; Nover, D. Heavy Metals in Water and Surface Sediments of the Fenghe River Basin, China: Assessment and Source Analysis. Water Science and Technology 2021, 84, 3072–3090. [Google Scholar] [CrossRef]
- Okpara, E.C.; Fayemi, O.E.; Wojuola, O.B.; Onwudiwe, D.C.; Ebenso, E.E. Electrochemical Detection of Selected Heavy Metals in Water: A Case Study of African Experiences. RSC Adv. 2022, 12, 26319–26361. [Google Scholar] [CrossRef]
- Zhang, P.; Yang, M.; Lan, J.; Huang, Y.; Zhang, J.; Huang, S.; Yang, Y.; Ru, J. Water Quality Degradation Due to Heavy Metal Contamination: Health Impacts and Eco-Friendly Approaches for Heavy Metal Remediation. Toxics 2023, 11, 828. [Google Scholar] [CrossRef]
- Stalwick, J.A.; Ratelle, M.; Gurney, K.E.B.; Drysdale, M.; Lazarescu, C.; Comte, J.; Laird, B.; Skinner, K. Sources of Exposure to Lead in Arctic and Subarctic Regions: A Scoping Review. International Journal of Circumpolar Health 2023, 82, 2208810. [Google Scholar] [CrossRef]
- Moyebi, O.D.; Lebbie, T.; Carpenter, D.O. Standards for Levels of Lead in Soil and Dust around the World. Reviews on Environmental Health 2025, 40, 185–196. [Google Scholar] [CrossRef] [PubMed]
- Briffa, J.; Sinagra, E.; Blundell, R. Heavy Metal Pollution in the Environment and Their Toxicological Effects on Humans. Heliyon 2020, 6, e04691. [Google Scholar] [CrossRef] [PubMed]
- Sadheesh, S.; Malathi, R.; Arundhathi; Rino, M.A.; Swetha, R. Monitoring and Analyzing the Effect of Heavy Metals in Air Based on Ecological and Health Risks in Coimbatore. IOP Conf. Ser.: Earth Environ. Sci. 2022, 1125, 012002. [CrossRef]
- EUR-Lex - 32023R0915 - EN - EUR-Lex. Europa.eu. https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX%3A32023R0915. (accessed on 07 July 2025).
- EUR-Lex - 32020L2184 - EN - EUR-Lex. Europa.eu. https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=celex%3A32020L2184. (accessed on 07 July 2025).
- Vieira, D.C.S.; Yunta, F.; Baragaño, D.; Evrard, O.; Reiff, T.; Silva, V.; de la Torre, A.; Zhang, C.; Panagos, P.; Jones, A.; Wojda, P. Soil Pollution in the European Union – an Outlook. Environmental Science & Policy 2024, 161, 103876. [Google Scholar] [CrossRef]
- European Commission. EU air quality standards. environment.ec.europa.eu. https://environment.ec.europa.eu/topics/air/air-quality/eu-air-quality-standards_en. (accessed on 07 July 2025).
- Angon, P.B.; Islam, Md.S.; Kc, S.; Das, A.; Anjum, N.; Poudel, A.; Suchi, S.A. Sources, Effects and Present Perspectives of Heavy Metals Contamination: Soil, Plants and Human Food Chain. Heliyon 2024, 10, e28357. [Google Scholar] [CrossRef]
- Wan, Y.; Liu, J.; Zhuang, Z.; Wang, Q.; Li, H. Heavy Metals in Agricultural Soils: Sources, Influencing Factors, and Remediation Strategies. Toxics 2024, 12, 63. [Google Scholar] [CrossRef]
- Fellows, K.M.; Samy, S.; Whittaker, S.G. Evaluating Metal Cookware as a Source of Lead Exposure. J Expo Sci Environ Epidemiol 2025, 35, 342–350. [Google Scholar] [CrossRef]
- Ashkan, M.F. Lead: Natural Occurrence, Toxicity to Organisms and Bioremediation by Lead-Degrading Bacteria: A Comprehensive Review. J. Pure Appl. Microbiol. 2023, 17, 1298–1319. [Google Scholar] [CrossRef]
- Balta, I.; Lemon, J.; Gadaj, A.; Cretescu, I.; Stef, D.; Pet, I.; Stef, L.; McCleery, D.; Douglas, A.; Corcionivoschi, N. The Interplay between Antimicrobial Resistance, Heavy Metal Pollution, and the Role of Microplastics. Front. Microbiol. 2025, 16, 1550587. [Google Scholar] [CrossRef]
- Abd Elnabi, M.K.; Elkaliny, N.E.; Elyazied, M.M.; Azab, S.H.; Elkhalifa, S.A.; Elmasry, S.; Mouhamed, M.S.; Shalamesh, E.M.; Alhorieny, N.A.; Abd Elaty, A.E.; et al. Toxicity of Heavy Metals and Recent Advances in Their Removal: A Review. Toxics 2023, 11, 580. [Google Scholar] [CrossRef]
- Zhu, S.; Mao, H.; Yang, X.; Zhao, W.; Sheng, L.; Sun, S.; Du, X. Resilience Mechanisms of Rhizosphere Microorganisms in Lead-Zinc Tailings: Metagenomic Insights into Heavy Metal Resistance. Ecotoxicology and Environmental Safety 2025, 292, 117956. [Google Scholar] [CrossRef] [PubMed]
- Nnaji, N.D.; Anyanwu, C.U.; Miri, T.; Onyeaka, H. Mechanisms of Heavy Metal Tolerance in Bacteria: A Review. Sustainability 2024, 16, 11124. [Google Scholar] [CrossRef]
- Davidova, S.; Milushev, V.; Satchanska, G. The Mechanisms of Cadmium Toxicity in Living Organisms. Toxics 2024, 12, 875. [Google Scholar] [CrossRef] [PubMed]
- Elizabeth George, S.; Wan, Y. Microbial Functionalities and Immobilization of Environmental Lead: Biogeochemical and Molecular Mechanisms and Implications for Bioremediation. Journal of Hazardous Materials 2023, 457, 131738. [Google Scholar] [CrossRef]
- Li, W.; Zhao, L.; Liu, C.; Bai, X.; Xu, C.; An, F.; Sun, F. Isolating and Identifying One Strain with Lead-Tolerant Fungus and Preliminary Study on Its Capability of Biosorption to Pb2+. Biology 2024, 13, 1053. [Google Scholar] [CrossRef]
- Titilawo, M.A.; Ajani, T.F.; Adedapo, S.A.; Akinleye, G.O.; Ogunlana, O.E.; Aderibigbe, D. Evaluation of Lead Tolerance and Biosorption Characteristics of Fungi from Dumpsite Soils. Discov Environ 2023, 1, 24. [Google Scholar] [CrossRef]
- Singh, V.K.; Singh, R. Role of White Rot Fungi in Sustainable Remediation of Heavy Metals from the Contaminated Environment.
- Feng, C.; Li, J.; Li, X.; Li, K.; Luo, K.; Liao, X.; Liu, T. Characterization and Mechanism of Lead and Zinc Biosorption by Growing Verticillium Insectorum J3. PLoS ONE 2018, 13, e0203859. [Google Scholar] [CrossRef]
- An, J.-M.; Gu, S.-Y.; Kim, D.-J.; Shin, H.-C.; Hong, K.-S.; Kim, Y.-K. Arsenic, Cadmium, Lead, and Mercury Contents of Mushroom Species in Korea and Associated Health Risk. International Journal of Food Properties 2020, 23, 992–998. [Google Scholar] [CrossRef]
- Muszyńska, B.; Rojowski, J.; Łazarz, M.; Kała, K.; Dobosz, K.; Opoka, W. The Accumulation and Release of Cd and Pb from Edible Mushrooms and Their Biomass. Pol. J. Environ. Stud. 2018, 27, 223–230. [Google Scholar] [CrossRef]
- Gałgowska, M.; Pietrzak-Fiećko, R. Cadmium and Lead Content in Selected Fungi from Poland and Their Edible Safety Assessment. Molecules 2021, 26, 7289. [Google Scholar] [CrossRef]
- Bucurica, I.A.; Dulama, I.D.; Radulescu, C.; Banica, A.L.; Stanescu, S.G. Heavy Metals and Associated Risks of Wild Edible Mushrooms Consumption: Transfer Factor, Carcinogenic Risk, and Health Risk Index. JoF 2024, 10, 844. [Google Scholar] [CrossRef]
- Orywal, K.; Socha, K.; Nowakowski, P.; Zoń, W.; Kaczyński, P.; Mroczko, B.; Łozowicka, B.; Perkowski, M. Health Risk Assessment of Exposure to Toxic Elements Resulting from Consumption of Dried Wild-Grown Mushrooms Available for Sale. PLoS ONE 2021, 16, e0252834. [Google Scholar] [CrossRef]
- Gupta, M.; Dwivedi, V.; Kumar, S.; Patel, A.; Niazi, P.; Yadav, V.K. Lead Toxicity in Plants: Mechanistic Insights into Toxicity, Physiological Responses of Plants and Mitigation Strategies. Plant Signaling & Behavior 2024, 19, 2365576. [Google Scholar] [CrossRef] [PubMed]
- Ur Rahman, S.; Qin, A.; Zain, M.; Mushtaq, Z.; Mehmood, F.; Riaz, L.; Naveed, S.; Ansari, M.J.; Saeed, M.; Ahmad, I.; et al. Pb Uptake, Accumulation, and Translocation in Plants: Plant Physiological, Biochemical, and Molecular Response: A Review. Heliyon 2024, 10, e27724. [Google Scholar] [CrossRef] [PubMed]
- Collin, S.; Baskar, A.; Geevarghese, D.M.; Ali, M.N.V.S.; Bahubali, P.; Choudhary, R.; Lvov, V.; Tovar, G.I.; Senatov, F.; Koppala, S.; et al. Bioaccumulation of Lead (Pb) and Its Effects in Plants: A Review. Journal of Hazardous Materials Letters 2022, 3, 100064. [Google Scholar] [CrossRef]
- He, S.; Niu, Y.; Xing, L.; Liang, Z.; Song, X.; Ding, M.; Huang, W. Research Progress of the Detection and Analysis Methods of Heavy Metals in Plants. Front. Plant Sci. 2024, 15, 1310328. [Google Scholar] [CrossRef]
- Angulo-Bejarano, P.I.; Puente-Rivera, J.; Cruz-Ortega, R. Metal and Metalloid Toxicity in Plants: An Overview on Molecular Aspects. Plants 2021, 10, 635. [Google Scholar] [CrossRef]
- Alengebawy, A.; Abdelkhalek, S.T.; Qureshi, S.R.; Wang, M.-Q. Heavy Metals and Pesticides Toxicity in Agricultural Soil and Plants: Ecological Risks and Human Health Implications. Toxics 2021, 9, 42. [Google Scholar] [CrossRef]
- Meskelu, T.; Senbeta, A.F.; Keneni, Y.G.; Sime, G. Heavy Metal Accumulation and Food Safety of Lettuce ( Lactuca Sativa L.) Amended by Bioslurry and Chemical Fertilizer Application. Food Science & Nutrition 2024, 12, 7449–7460. [Google Scholar] [CrossRef]
- Cleophas, F.N.; Zahari, N.Z.; Murugayah, P.; Rahim, S.A.; Mohd Yatim, A.N. Phytoremediation: A Novel Approach of Bast Fiber Plants (Hemp, Kenaf, Jute and Flax) for Heavy Metals Decontamination in Soil—Review. Toxics 2022, 11, 5. [Google Scholar] [CrossRef]
- Elik, Ü.; Gül, Z. Accumulation Potential of Lead and Cadmium Metals in Maize (Zea Mays L.) and Effects on Physiological-Morphological Characteristics. Life 2025, 15, 310. [Google Scholar] [CrossRef]
- Guo, Z.; Gao, Y.; Yuan, X.; Yuan, M.; Huang, L.; Wang, S.; Liu, C.; Duan, C. Effects of Heavy Metals on Stomata in Plants: A Review. IJMS 2023, 24, 9302. [Google Scholar] [CrossRef]
- Ejaz, U.; Khan, S.M.; Khalid, N.; Ahmad, Z.; Jehangir, S.; Fatima Rizvi, Z.; Lho, L.H.; Han, H.; Raposo, A. Detoxifying the Heavy Metals: A Multipronged Study of Tolerance Strategies against Heavy Metals Toxicity in Plants. Front. Plant Sci. 2023, 14, 1154571. [Google Scholar] [CrossRef]
- El-Sappah, A.H.; Zhu, Y.; Huang, Q.; Chen, B.; Soaud, S.A.; Abd Elhamid, M.A.; Yan, K.; Li, J.; El-Tarabily, K.A. Plants’ Molecular Behavior to Heavy Metals: From Criticality to Toxicity. Front. Plant Sci. 2024, 15, 1423625. [Google Scholar] [CrossRef]
- Rahman, S.U.; Nawaz, M.F.; Gul, S.; Yasin, G.; Hussain, B.; Li, Y.; Cheng, H. State-of-the-Art OMICS Strategies against Toxic Effects of Heavy Metals in Plants: A Review. Ecotoxicology and Environmental Safety 2022, 242, 113952. [Google Scholar] [CrossRef] [PubMed]
- Fasani, E.; Giannelli, G.; Varotto, S.; Visioli, G.; Bellin, D.; Furini, A.; DalCorso, G. Epigenetic Control of Plant Response to Heavy Metals. Plants 2023, 12, 3195. [Google Scholar] [CrossRef] [PubMed]
- Monchanin, C.; Devaud, J.-M.; Barron, A.B.; Lihoreau, M. Current Permissible Levels of Metal Pollutants Harm Terrestrial Invertebrates. Science of The Total Environment 2021, 779, 146398. [Google Scholar] [CrossRef]
- Oros, A. Bioaccumulation and Trophic Transfer of Heavy Metals in Marine Fish: Ecological and Ecosystem-Level Impacts. JoX 2025, 15, 59. [Google Scholar] [CrossRef]
- Monclús, L.; Shore, R.F.; Krone, O. Lead Contamination in Raptors in Europe: A Systematic Review and Meta-Analysis. Science of The Total Environment 2020, 748, 141437. [Google Scholar] [CrossRef]
- Lazarova, I.; Balieva, G. Effects of Lead Exposure in Wild Birds as Causes for Incidents and Fatal Injuries. Diversity 2025, 17, 387. [Google Scholar] [CrossRef]
- Vajargah, M.F.; Azar, H. Investigating the Effects of Accumulation of Lead and Cadmium Metals in Fish and Its Impact on Human Health. JAMB 2023, 12, 209–213. [Google Scholar] [CrossRef]
- Tahir, I.; Alkheraije, K.A. A Review of Important Heavy Metals Toxicity with Special Emphasis on Nephrotoxicity and Its Management in Cattle. Front. Vet. Sci. 2023, 10, 1149720. [Google Scholar] [CrossRef] [PubMed]
- Sartorius, A.; Johnson, M.F.; Young, S.; Bennett, M.; Baiker, K.; Edwards, P.; Yon, L. Trace Metal Accumulation through the Environment and Wildlife at Two Derelict Lead Mines in Wales. Heliyon 2024, 10, e34265. [Google Scholar] [CrossRef] [PubMed]
- Chirinos-Peinado, D.; Castro-Bedriñana, J.; Ríos-Ríos, E.; Mamani-Gamarra, G.; Quijada-Caro, E.; Huacho-Jurado, A.; Nuñez-Rojas, W. Lead and Cadmium Bioaccumulation in Fresh Cow’s Milk in an Intermediate Area of the Central Andes of Peru and Risk to Human Health. Toxics 2022, 10, 317. [Google Scholar] [CrossRef]
- Hussain, M.I.; Khan, Z.I.; Ahmad, K.; Naeem, M.; Ali, M.A.; Elshikh, M.S.; Zaman, Q.U.; Iqbal, K.; Muscolo, A.; Yang, H.-H. Toxicity and Bioassimilation of Lead and Nickel in Farm Ruminants Fed on Diversified Forage Crops Grown on Contaminated Soil. Ecotoxicology and Environmental Safety 2024, 283, 116812. [Google Scholar] [CrossRef]
- Aljohani, A.S.M. Heavy Metal Toxicity in Poultry: A Comprehensive Review. Front. Vet. Sci. 2023, 10, 1161354. [Google Scholar] [CrossRef]
- Hassan Emami, M.; Saberi, F.; Mohammadzadeh, S.; Fahim, A.; Abdolvand, M.; Ali Ehsan Dehkordi, S.; Mohammadzadeh, S.; Maghool, F. A Review of Heavy Metals Accumulation in Red Meat and Meat Products in the Middle East. Journal of Food Protection 2023, 86, 100048. [Google Scholar] [CrossRef]
- Jomova, K.; Alomar, S.Y.; Nepovimova, E.; Kuca, K.; Valko, M. Heavy Metals: Toxicity and Human Health Effects. Arch Toxicol 2025, 99, 153–209. [Google Scholar] [CrossRef]
- Balali-Mood, M.; Naseri, K.; Tahergorabi, Z.; Khazdair, M.R.; Sadeghi, M. Toxic Mechanisms of Five Heavy Metals: Mercury, Lead, Chromium, Cadmium, and Arsenic. Front. Pharmacol. 2021, 12, 643972. [Google Scholar] [CrossRef]
- Mititelu, M.; Neacșu, S.M.; Busnatu, Ștefan, S.; Scafa-Udriște, A.; Andronic, O.; Lăcraru, A.-E.; Ioniță-Mîndrican, C.-B.; Lupuliasa, D.; Negrei, C.; Olteanu, G. Assessing Heavy Metal Contamination in Food: Implications for Human Health and Environmental Safety. Toxics 2025, 13, 333. [CrossRef]
- Witkowska, D.; Słowik, J.; Chilicka, K. Heavy Metals and Human Health: Possible Exposure Pathways and the Competition for Protein Binding Sites. Molecules 2021, 26, 6060. [Google Scholar] [CrossRef]
- Majewski, G.; Klik, B.K.; Rogula-Kozłowska, W.; Rogula-Kopiec, P.; Rybak, J.; Radziemska, M.; Liniauskienė, E. Assessment of Heavy Metal Inhalation Risks in Urban Environments in Poland: A Case Study. J. Ecol. Eng. 2023, 24, 330–340. [Google Scholar] [CrossRef] [PubMed]
- Khoshakhlagh, A.H.; Ghobakhloo, S.; Gruszecka-Kosowska, A. Inhalational Exposure to Heavy Metals: Carcinogenic and Non-Carcinogenic Risk Assessment. Journal of Hazardous Materials Advances 2024, 16, 100485. [Google Scholar] [CrossRef]
- Scutarașu, E.C.; Trincă, L.C. Heavy Metals in Foods and Beverages: Global Situation, Health Risks and Reduction Methods. Foods 2023, 12, 3340. [Google Scholar] [CrossRef] [PubMed]
- Bair, E.C. A Narrative Review of Toxic Heavy Metal Content of Infant and Toddler Foods and Evaluation of United States Policy. Front. Nutr. 2022, 9, 919913. [Google Scholar] [CrossRef]
- Niemeier, R.T.; Maier, A.; Reichard, J.F. Rapid Review of Dermal Penetration and Absorption of Inorganic Lead Compounds for Occupational Risk Assessment. Annals of Work Exposures and Health 2022, 66, 291–311. [Google Scholar] [CrossRef]
- Arshad, H.; Mehmood, M.Z.; Shah, M.H.; Abbasi, A.M. Evaluation of Heavy Metals in Cosmetic Products and Their Health Risk Assessment. Saudi Pharmaceutical Journal 2020, 28, 779–790. [Google Scholar] [CrossRef]
- Oginawati, K.; Suharyanto; Susetyo, S.H.; Sulung, G.; Muhayatun; Chazanah, N.; Dewi Kusumah, S.W.; Fahimah, N. Investigation of Dermal Exposure to Heavy Metals (Cu, Zn, Ni, Al, Fe and Pb) in Traditional Batik Industry Workers. Heliyon 2022, 8, e08914. [CrossRef]
- Rosengren, E.; Barregard, L.; Sallsten, G.; Fagerberg, B.; Engström, G.; Fagman, E.; Forsgard, N.; Lundh, T.; Bergström, G.; Harari, F. Exposure to Lead and Coronary Artery Atherosclerosis: A Swedish Cross-Sectional Population-Based Study. JAHA 2025, 14, e037633. [Google Scholar] [CrossRef]
- Nucera, S.; Serra, M.; Caminiti, R.; Ruga, S.; Passacatini, L.C.; Macrì, R.; Scarano, F.; Maiuolo, J.; Bulotta, R.; Mollace, R.; et al. Non-Essential Heavy Metal Effects in Cardiovascular Diseases: An Overview of Systematic Reviews. Front. Cardiovasc. Med. 2024, 11, 1332339. [Google Scholar] [CrossRef]
- Dang, P.; Tang, M.; Fan, H.; Hao, J. Chronic Lead Exposure and Burden of Cardiovascular Disease during 1990–2019: A Systematic Analysis of the Global Burden of Disease Study. Front. Cardiovasc. Med. 2024, 11, 1367681. [Google Scholar] [CrossRef]
- Lieberman-Cribbin, W.; Li, Z.; Lewin, M.; Ruiz, P.; Jarrett, J.M.; Cole, S.A.; Kupsco, A.; O’Leary, M.; Pichler, G.; Shimbo, D.; et al. The Contribution of Declines in Blood Lead Levels to Reductions in Blood Pressure Levels: Longitudinal Evidence in the Strong Heart Family Study. JAHA 2024, 13, e031256. [Google Scholar] [CrossRef] [PubMed]
- Shan, D.; Wen, X.; Guan, X.; Fang, H.; Liu, Y.; Qin, M.; Wang, H.; Xu, J.; Lv, J.; Zhao, J.; et al. Pubertal Lead Exposure Affects Ovary Development, Folliculogenesis and Steroidogenesis by Activation of IRE1α-JNK Signaling Pathway in Rat. Ecotoxicology and Environmental Safety 2023, 257, 114919. [Google Scholar] [CrossRef] [PubMed]
- Osowski, A.; Fedoniuk, L.; Bilyk, Y.; Fedchyshyn, O.; Sas, M.; Kramar, S.; Lomakina, Y.; Fik, V.; Chorniy, S.; Wojtkiewicz, J. Lead Exposure Assessment and Its Impact on the Structural Organization and Morphological Peculiarities of Rat Ovaries. Toxics 2023, 11, 769. [Google Scholar] [CrossRef]
- Aliche, K.A.; Umeoguaju, F.U.; Ikewuchi, C.; Diorgu, F.C.; Ajao, O.; Frazzoli, C.; Orisakwe, O.E. Paternal Lead Exposure and Pregnancy Outcomes: A Systematic Review and Meta-Analysis. Environmental Health Insights 2025, 19. [Google Scholar] [CrossRef] [PubMed]
- Giulioni, C.; Maurizi, V.; De Stefano, V.; Polisini, G.; Teoh, J.Y.-C.; Milanese, G.; Galosi, A.B.; Castellani, D. The Influence of Lead Exposure on Male Semen Parameters: A Systematic Review and Meta-Analysis. Reproductive Toxicology 2023, 118, 108387. [Google Scholar] [CrossRef]
- Yu, Y.-L.; Yang, W.-Y.; Hara, A.; Asayama, K.; Roels, H.A.; Nawrot, T.S.; Staessen, J.A. Public and Occupational Health Risks Related to Lead Exposure Updated According to Present-Day Blood Lead Levels. Hypertens Res 2023, 46, 395–407. [Google Scholar] [CrossRef]
- Alcala, C.S.; Lane, J.M.; Midya, V.; Eggers, S.; Wright, R.O.; Rosa, M.J. Exploring the Link between the Pediatric Exposome, Respiratory Health, and Executive Function in Children: A Narrative Review. Front. Public Health 2024, 12, 1383851. [Google Scholar] [CrossRef]
- Boskabady, M.; Marefati, N.; Farkhondeh, T.; Shakeri, F.; Farshbaf, A.; Boskabady, M.H. The Effect of Environmental Lead Exposure on Human Health and the Contribution of Inflammatory Mechanisms, a Review. Environment International 2018, 120, 404–420. [Google Scholar] [CrossRef]
- Liang, D.; Liu, C.; Yang, M. The Association between Urinary Lead Concentration and the Likelihood of Kidney Stones in US Adults: A Population-Based Study. Sci Rep 2025, 15, 1653. [Google Scholar] [CrossRef]
- Upadhyay, K.; Viramgami, A.; Bagepally, B.S.; Balachandar, R. Association between Chronic Lead Exposure and Markers of Kidney Injury: A Systematic Review and Meta-Analysis. Toxicology Reports 2024, 13, 101837. [Google Scholar] [CrossRef]
- Li, L.; Zhao, J. Association of Urinary and Blood Lead Concentrations with All-Cause Mortality in US Adults with Chronic Kidney Disease: A Prospective Cohort Study. Sci Rep 2024, 14, 23230. [Google Scholar] [CrossRef]
- Kuo, P.-F.; Huang, Y.-T.; Chuang, M.-H.; Jiang, M.-Y. Association of Low-Level Heavy Metal Exposure with Risk of Chronic Kidney Disease and Long-Term Mortality. PLoS ONE 2024, 19, e0315688. [Google Scholar] [CrossRef] [PubMed]
- Paudel, S.; Geetha, P.; Kyriazis, P.; Specht, A.; Hu, H.; Danziger, J. Association of Environmental Lead Toxicity and Hematologic Outcomes in Patients with Advanced Kidney Disease. Nephrology Dialysis Transplantation 2023, 38, 1337–1339. [Google Scholar] [CrossRef] [PubMed]
- Qiao, L.; Li, M.; Wen, X.; Deng, F.; Hou, X.; Zou, Q.; Liu, H.; Fan, X.; Han, J. Epidemiological Trends in the Burden of Chronic Kidney Disease Due to Lead Exposure in China from 1990 to 2021 Compared to Global, and Projections until 2050. International Journal of Environmental Health Research 2025, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Battistini, B.; Greggi, C.; Visconti, V.V.; Albanese, M.; Messina, A.; De Filippis, P.; Gasperini, B.; Falvino, A.; Piscitelli, P.; Palombi, L.; et al. Metals Accumulation Affects Bone and Muscle in Osteoporotic Patients: A Pilot Study. Environmental Research 2024, 250, 118514. [Google Scholar] [CrossRef]
- Wang, S.; Zhao, X.; Zhou, R.; Jin, Y.; Wang, X.; Ma, X.; Lu, X. The Influence of Adult Urine Lead Exposure on Bone Mineral Densit: NHANES 2015-2018. Front. Endocrinol. 2024, 15, 1412872. [Google Scholar] [CrossRef]
- Cui, A.; Xiao, P.; Hu, B.; Ma, Y.; Fan, Z.; Wang, H.; Zhou, F.; Zhuang, Y. Blood Lead Level Is Negatively Associated With Bone Mineral Density in U.S. Children and Adolescents Aged 8-19 Years. Front. Endocrinol. 2022, 13, 928752. [Google Scholar] [CrossRef]
- Zabihi, A.; Mehrpour, O.; Nakhaee, S.; Atabati, E. Lead Poisoning and Its Effects on Bone Density. Sci Rep 2025, 15, 15619. [Google Scholar] [CrossRef]
- Firoozichahak, A.; Rahimnejad, S.; Rahmani, A.; Parvizimehr, A.; Aghaei, A.; Rahimpoor, R. Effect of Occupational Exposure to Lead on Serum Levels of Lipid Profile and Liver Enzymes: An Occupational Cohort Study. Toxicology Reports 2022, 9, 269–275. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, Z.; Lin, H.; Zeng, Z.; Huang, J.; Li, D. Lead Exposure Was Associated with Liver Fibrosis in Subjects without Known Chronic Liver Disease: An Analysis of NHANES 2017–2020. Front. Environ. Sci. 2022, 10, 995795. [Google Scholar] [CrossRef]
- Teerasarntipan, T.; Chaiteerakij, R.; Prueksapanich, P.; Werawatganon, D. Changes in Inflammatory Cytokines, Antioxidants and Liver Stiffness after Chelation Therapy in Individuals with Chronic Lead Poisoning. BMC Gastroenterol 2020, 20, 263. [Google Scholar] [CrossRef]
- Renu, K.; Chakraborty, R.; Myakala, H.; Koti, R.; Famurewa, A.C.; Madhyastha, H.; Vellingiri, B.; George, A.; Valsala Gopalakrishnan, A. Molecular Mechanism of Heavy Metals (Lead, Chromium, Arsenic, Mercury, Nickel and Cadmium) - Induced Hepatotoxicity – A Review. Chemosphere 2021, 271, 129735. [Google Scholar] [CrossRef]
- Yu, L.; Yu, Y.; Xiao, Y.; Tian, F.; Narbad, A.; Zhai, Q.; Chen, W. Lead-Induced Gut Injuries and the Dietary Protective Strategies: A Review. Journal of Functional Foods 2021, 83, 104528. [Google Scholar] [CrossRef]
- Ramírez Ortega, D.; González Esquivel, D.F.; Blanco Ayala, T.; Pineda, B.; Gómez Manzo, S.; Marcial Quino, J.; Carrillo Mora, P.; Pérez De La Cruz, V. Cognitive Impairment Induced by Lead Exposure during Lifespan: Mechanisms of Lead Neurotoxicity. Toxics 2021, 9, 23. [Google Scholar] [CrossRef] [PubMed]
- Alva, S.; Parithathvi, A.; Harshitha, P.; Dsouza, H.S. Influence of Lead on cAMP-Response Element Binding Protein (CREB) and Its Implications in Neurodegenerative Disorders. Toxicology Letters 2024, 400, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Gundacker, C.; Forsthuber, M.; Szigeti, T.; Kakucs, R.; Mustieles, V.; Fernandez, M.F.; Bengtsen, E.; Vogel, U.; Hougaard, K.S.; Saber, A.T. Lead (Pb) and Neurodevelopment: A Review on Exposure and Biomarkers of Effect (BDNF, HDL) and Susceptibility. International Journal of Hygiene and Environmental Health 2021, 238, 113855. [Google Scholar] [CrossRef]
- Sanders, T.; Liu, Y.; Buchner, V.; Tchounwou, P.B. Neurotoxic Effects and Biomarkers of Lead Exposure: A Review. Reviews on Environmental Health 2009, 24. [Google Scholar] [CrossRef]
- Ahn, J.; Park, M.Y.; Kang, M.-Y.; Shin, I.-S.; An, S.; Kim, H.-R. Occupational Lead Exposure and Brain Tumors: Systematic Review and Meta-Analysis. IJERPH 2020, 17, 3975. [Google Scholar] [CrossRef]
- Zhang, X.; Sun, L.; Dapar, M.L.G.; Zhang, Q. Novel Plasma Cytokines Identified and Validated in Children during Lead Exposure According to the New Updated BLRV. Sci Rep 2024, 14, 30323. [Google Scholar] [CrossRef]
- Pukanha, K.; Yimthiang, S.; Kwanhian, W. The Immunotoxicity of Chronic Exposure to High Levels of Lead: An Ex Vivo Investigation. Toxics 2020, 8, 56. [Google Scholar] [CrossRef]
- Hong, Y.; Ye, T.; Jiang, H.; Wang, A.; Wang, B.; Li, Y.; Xie, H.; Meng, H.; Shen, C.; Ding, X. Panoramic Lead-Immune System Interactome Reveals Diversified Mechanisms of Immunotoxicity upon Chronic Lead Exposure. Cell Biol Toxicol 2025, 41, 81. [Google Scholar] [CrossRef] [PubMed]
- Harshitha, P.; Bose, K.; Dsouza, H.S. Influence of Lead-Induced Toxicity on the Inflammatory Cytokines. Toxicology 2024, 503, 153771. [Google Scholar] [CrossRef] [PubMed]
- Virgolini, M.B.; Aschner, M. Molecular Mechanisms of Lead Neurotoxicity. In Advances in Neurotoxicology; Elsevier, 2021; Vol. 5, pp. 159–213 ISBN 978-0-12-823775-5.
- Han, Q.; Zhang, W.; Guo, J.; Zhu, Q.; Chen, H.; Xia, Y.; Zhu, G. Mitochondrion: A Sensitive Target for Pb Exposure. J. Toxicol. Sci. 2021, 46, 345–358. [Google Scholar] [CrossRef] [PubMed]
- Koyama, H.; Kamogashira, T.; Yamasoba, T. Heavy Metal Exposure: Molecular Pathways, Clinical Implications, and Protective Strategies. Antioxidants 2024, 13, 76. [Google Scholar] [CrossRef]
- Chlubek, M.; Baranowska-Bosiacka, I. Selected Functions and Disorders of Mitochondrial Metabolism under Lead Exposure. Cells 2024, 13, 1182. [Google Scholar] [CrossRef]
- Hemmaphan, S.; Bordeerat, N.K. Genotoxic Effects of Lead and Their Impact on the Expression of DNA Repair Genes. IJERPH 2022, 19, 4307. [Google Scholar] [CrossRef]
- Mei, Z.; Liu, G.; Zhao, B.; He, Z.; Gu, S. Emerging Roles of Epigenetics in Lead-Induced Neurotoxicity. Environment International 2023, 181, 108253. [Google Scholar] [CrossRef]
- Mitra, P.; Misra, S.; Sharma, P. Epigenetics in Lead Toxicity: New Avenues for Future Research. Ind J Clin Biochem 2021, 36, 129–130. [Google Scholar] [CrossRef]
- Yang, R.; Roshani, D.; Gao, B.; Li, P.; Shang, N. Metallothionein: A Comprehensive Review of Its Classification, Structure, Biological Functions, and Applications. Antioxidants 2024, 13, 825. [Google Scholar] [CrossRef]
- Zulfiqar, U.; Farooq, M.; Hussain, S.; Maqsood, M.; Hussain, M.; Ishfaq, M.; Ahmad, M.; Anjum, M.Z. Lead Toxicity in Plants: Impacts and Remediation. Journal of Environmental Management 2019, 250, 109557. [Google Scholar] [CrossRef]
- Ghosal, C.; Alam, N.; Tamang, A.; Chattopadhyay, B. Removal of Lead Contamination through the Formation of Lead-Nanoplates by a Hot Spring Microbial Protein. AiM 2021, 11, 681–693. [Google Scholar] [CrossRef]
- Fathy, A.T.; Moneim, M.A.; Ahmed, E.A.; El-Ayaat, A.M.; Dardir, F.M. Effective Removal of Heavy Metal Ions (Pb, Cu, and Cd) from Contaminated Water by Limestone Mine Wastes. Sci Rep 2025, 15, 1680. [Google Scholar] [CrossRef]
- Hong, J.; Xie, J.; Mirshahghassemi, S.; Lead, J. Metal (Cd, Cr, Ni, Pb) Removal from Environmentally Relevant Waters Using Polyvinylpyrrolidone-Coated Magnetite Nanoparticles. RSC Adv. 2020, 10, 3266–3276. [Google Scholar] [CrossRef]
Figure 1.
Concentration of Pb in topsoils in Europe (0-25cm) [5].
Figure 1.
Concentration of Pb in topsoils in Europe (0-25cm) [5].
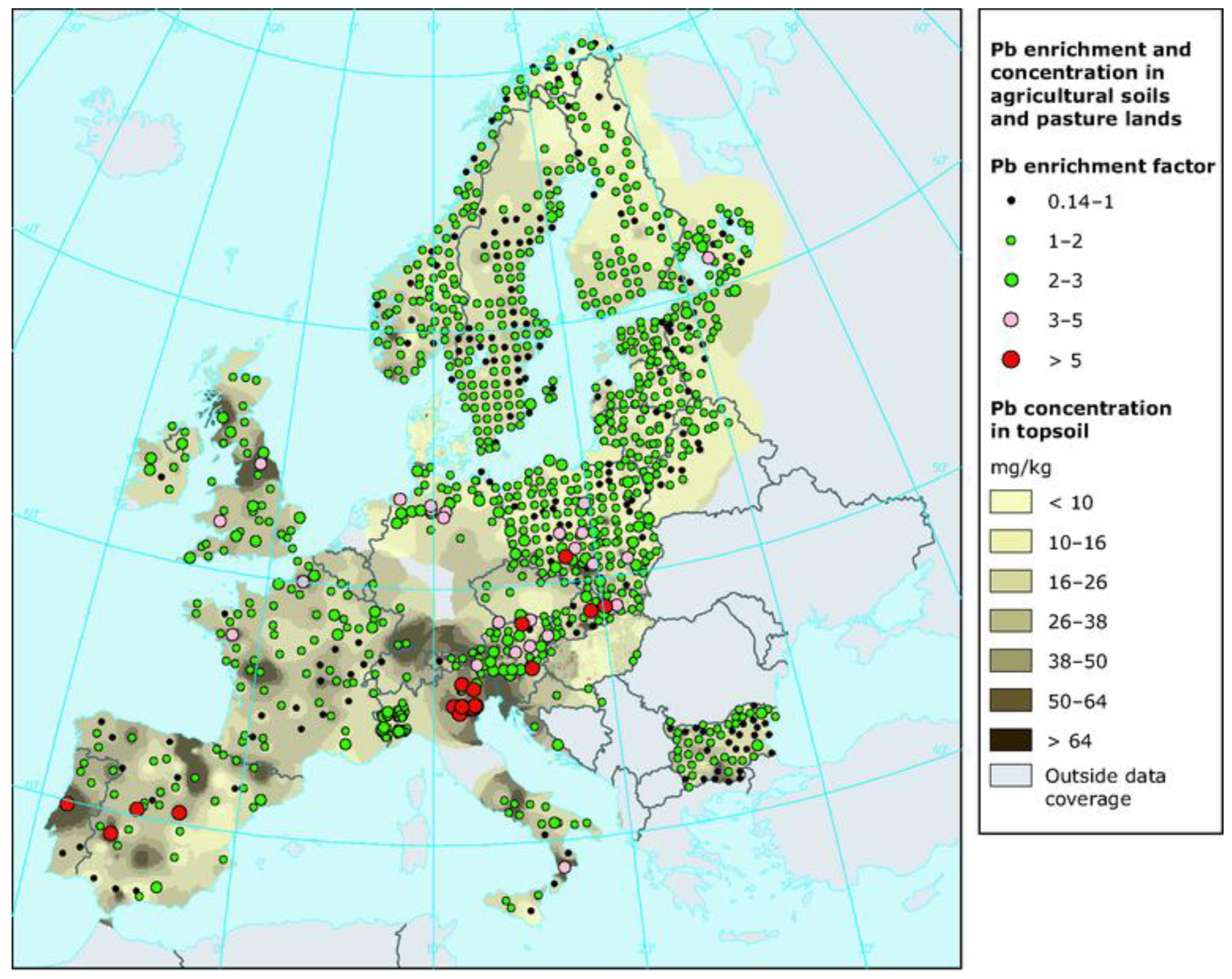
Figure 2.
Principal origins of lead contamination in the soil-plant system [9].
Figure 2.
Principal origins of lead contamination in the soil-plant system [9].
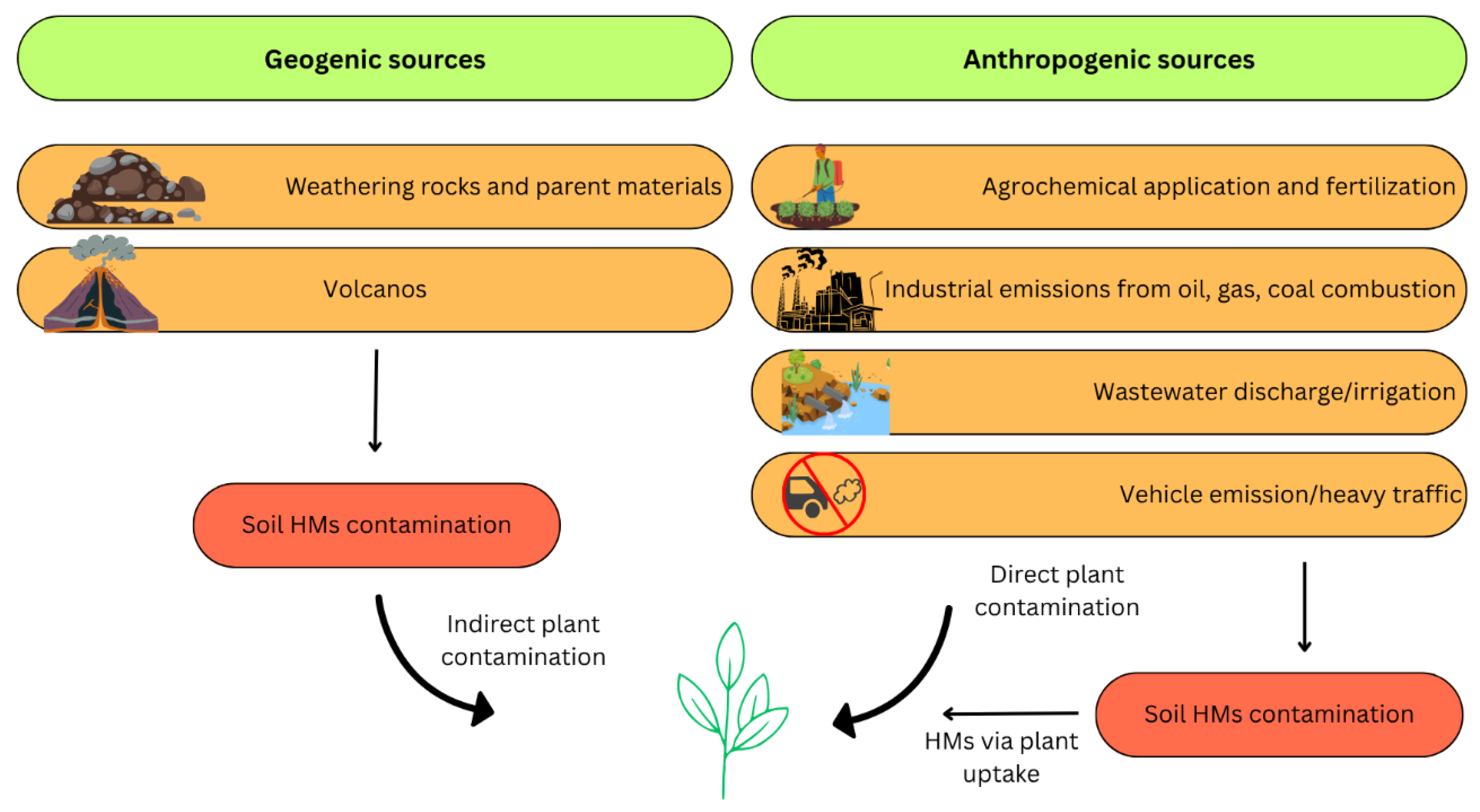
Figure 3.
Toxicity of heavy metals on microorganisms [36].
Figure 3.
Toxicity of heavy metals on microorganisms [36].
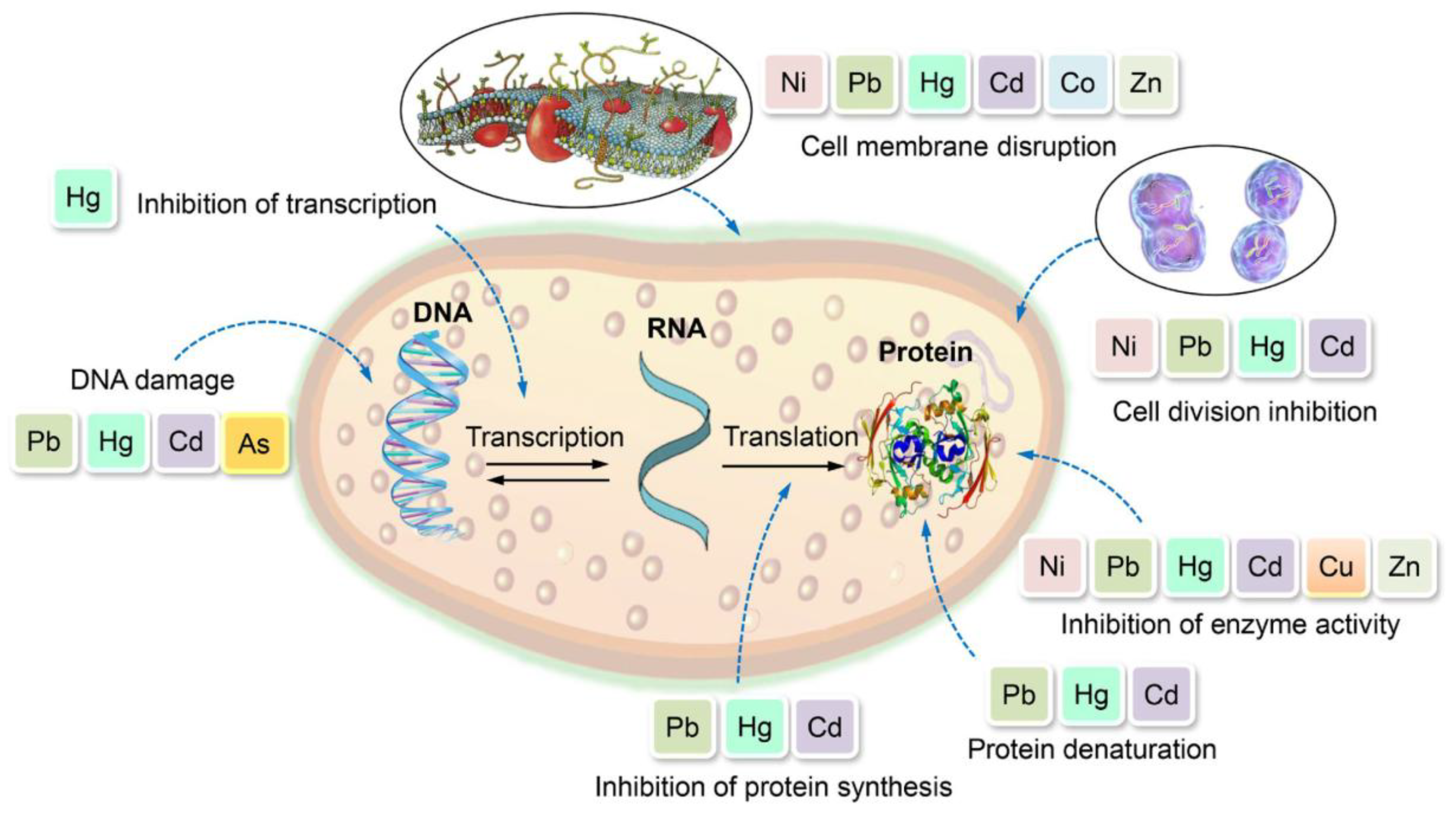
Figure 4.
Visualization of lead uptake through the root system of plants, highlighting the roles of passive diffusion, facilitated transport, and active uptake. The image shows Pb entering root cells (a, b) via transporters including the ZIP (e.g., ZIP4/9) and NRAMP families, as labeled in sections (b) and (d). These transporters mediate the internalization of Pb ions into the cytosol and vacuole [54].
Figure 4.
Visualization of lead uptake through the root system of plants, highlighting the roles of passive diffusion, facilitated transport, and active uptake. The image shows Pb entering root cells (a, b) via transporters including the ZIP (e.g., ZIP4/9) and NRAMP families, as labeled in sections (b) and (d). These transporters mediate the internalization of Pb ions into the cytosol and vacuole [54].
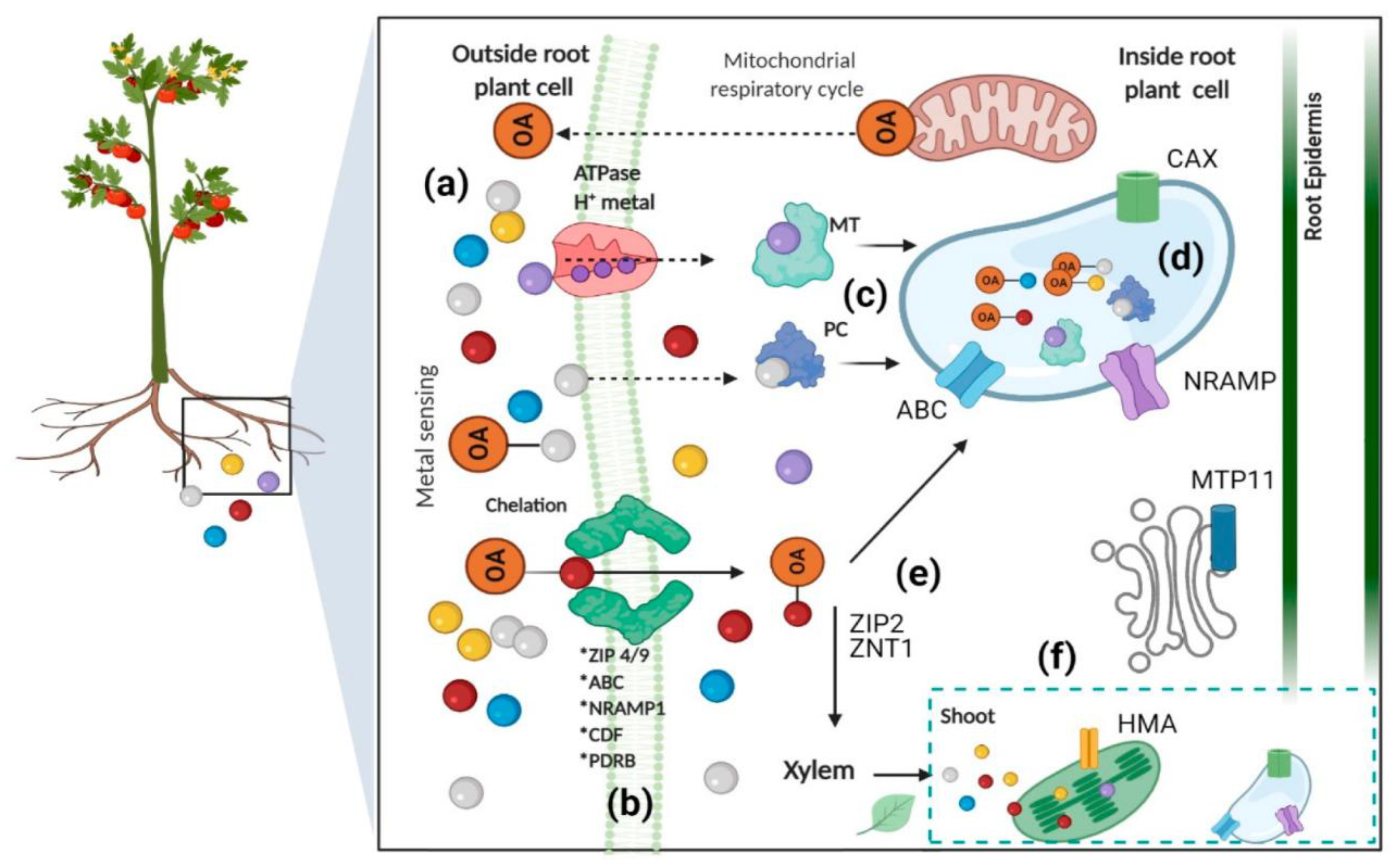
Figure 5.
Lead (Pb) exposure induces toxic systemic effects [2].
Figure 5.
Lead (Pb) exposure induces toxic systemic effects [2].
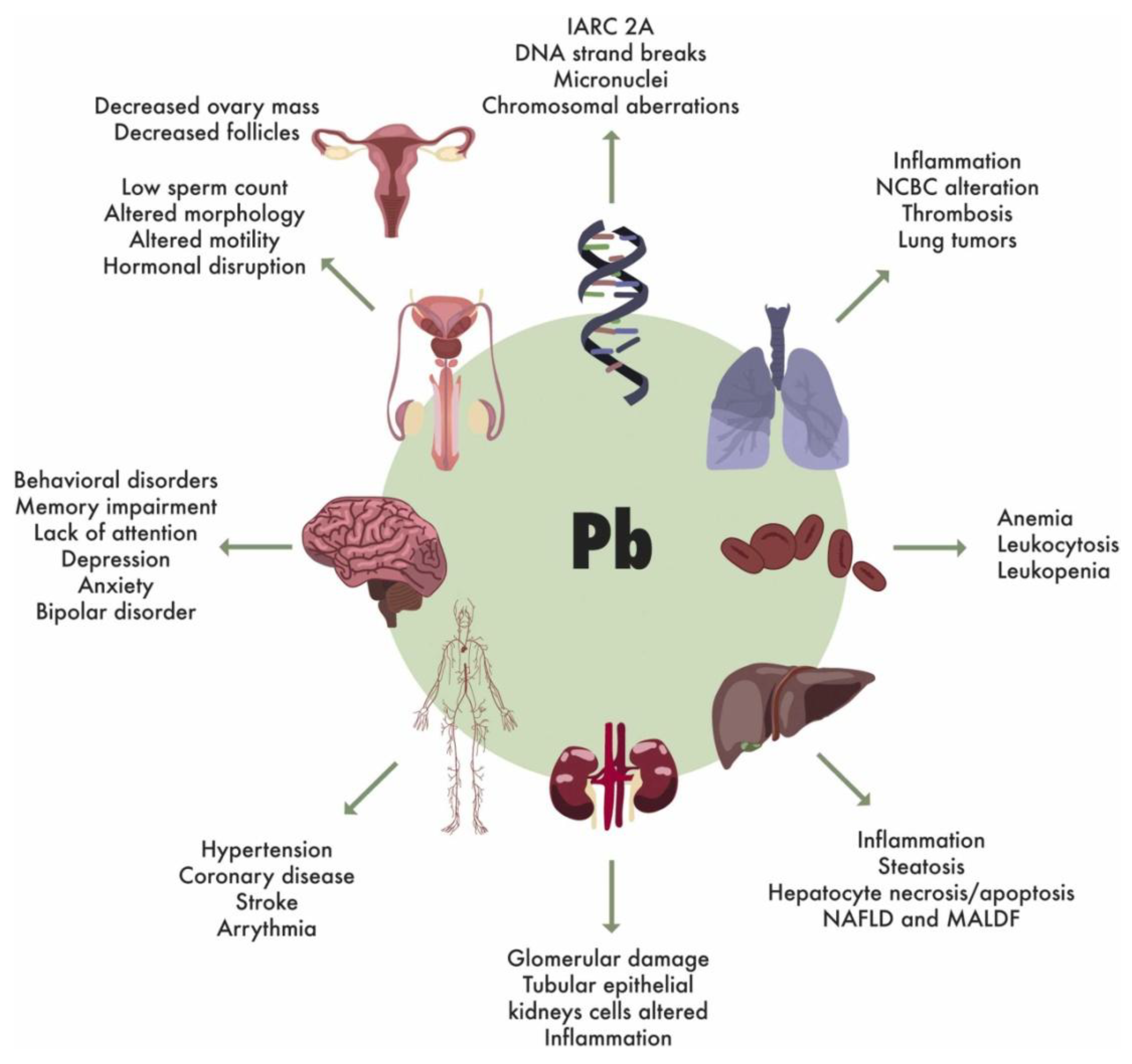
Figure 6.
Pathways of Lead Accumulation and Its Effects on Human Health [7].
Figure 6.
Pathways of Lead Accumulation and Its Effects on Human Health [7].
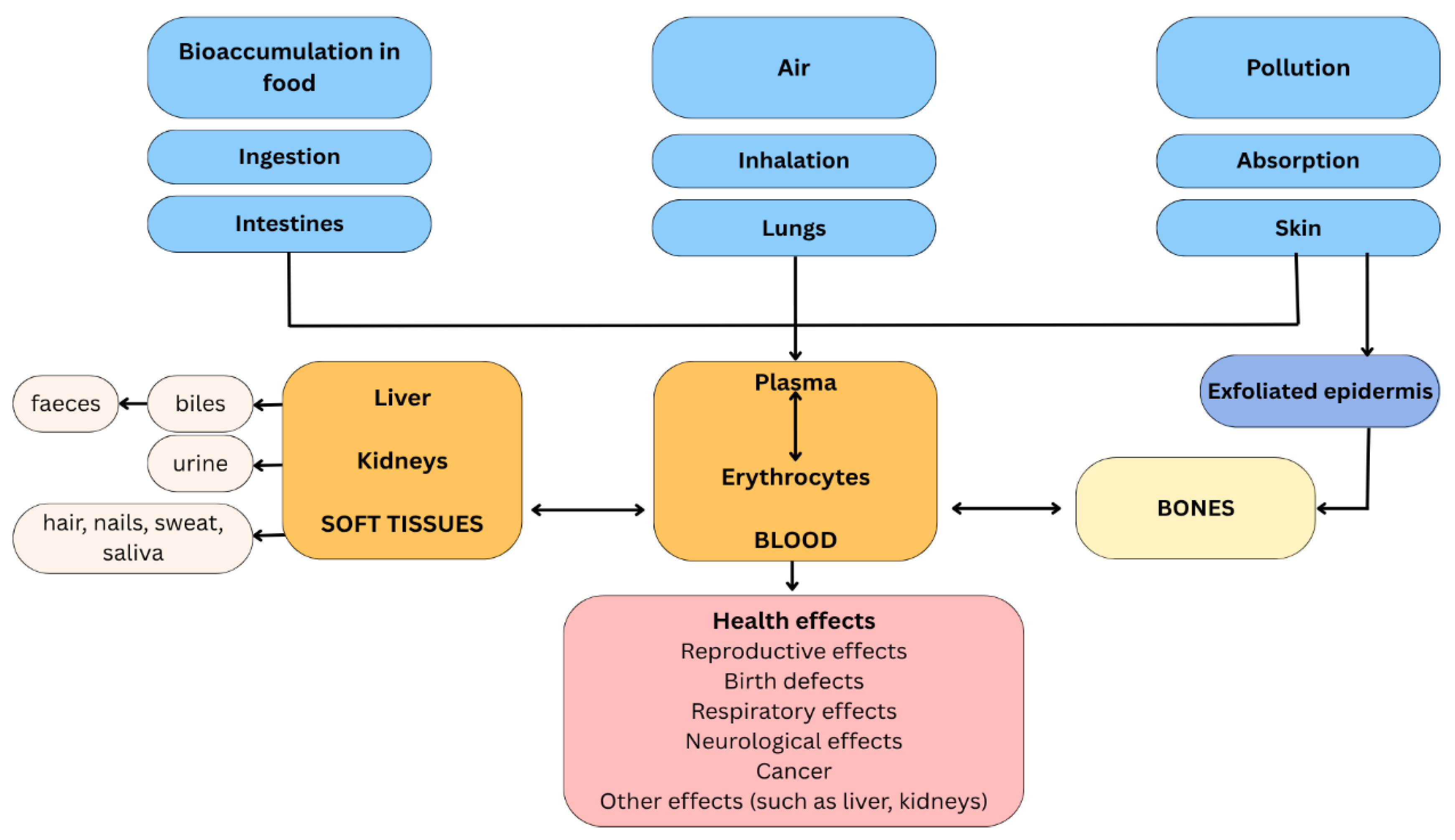
Figure 7.
Oxidative stress and human organ toxicity following exposure to HMs [36].
Figure 7.
Oxidative stress and human organ toxicity following exposure to HMs [36].
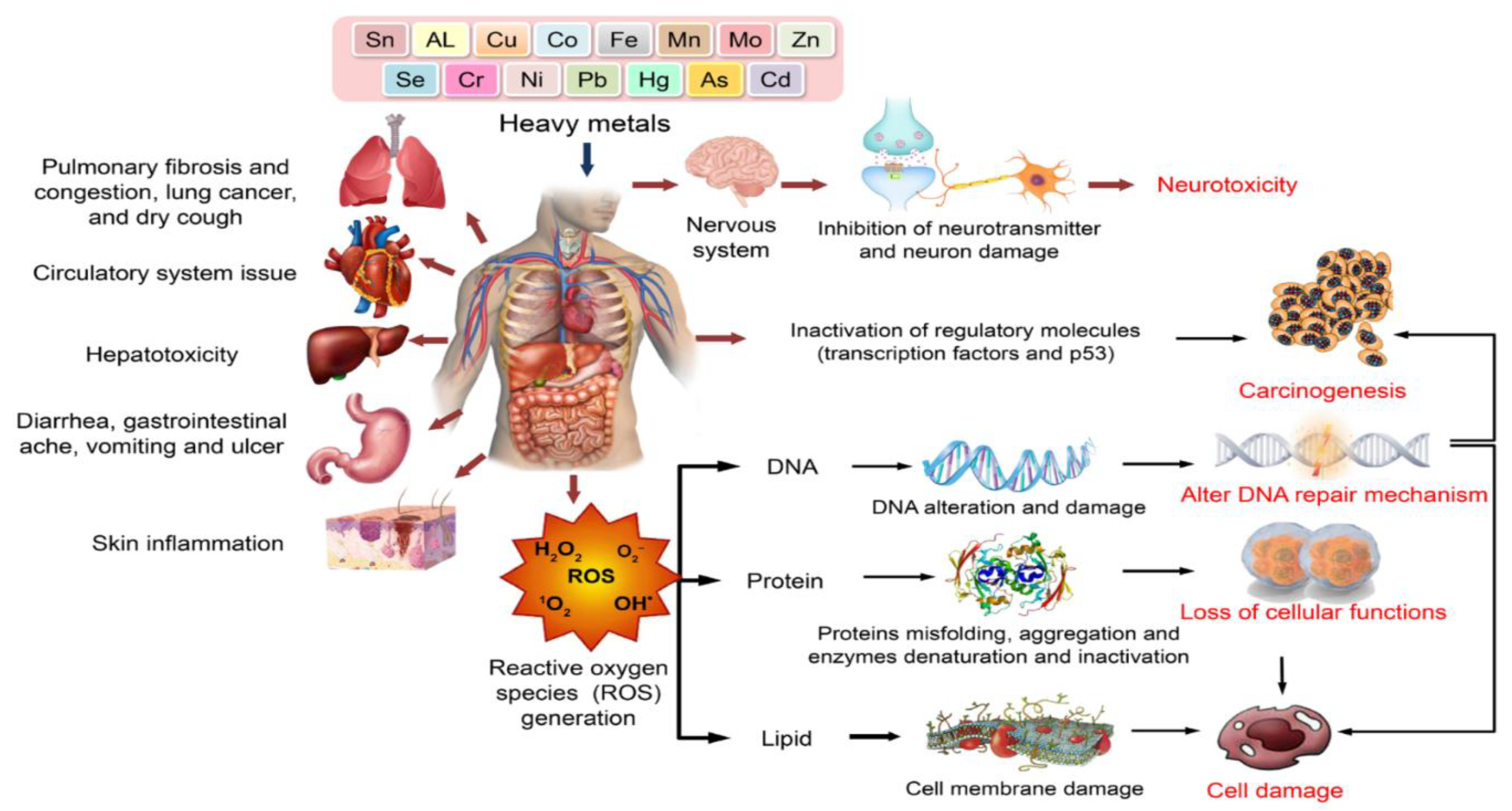
Figure 8.
Lead and mitochondrial permeability transition pore involvement in late apoptosis and necrosis. Extensive metabolic changes in a cell exposed to lead result in the opening of the mitochondrial permeability transition pores. The dramatically increased permeability of the outer mitochondrial membrane is accompanied by mitochondrial swelling (A) and a non-selective release of apoptosis-inducing factors from the intermembrane space into the cytoplasm (B) [124].
Figure 8.
Lead and mitochondrial permeability transition pore involvement in late apoptosis and necrosis. Extensive metabolic changes in a cell exposed to lead result in the opening of the mitochondrial permeability transition pores. The dramatically increased permeability of the outer mitochondrial membrane is accompanied by mitochondrial swelling (A) and a non-selective release of apoptosis-inducing factors from the intermembrane space into the cytoplasm (B) [124].
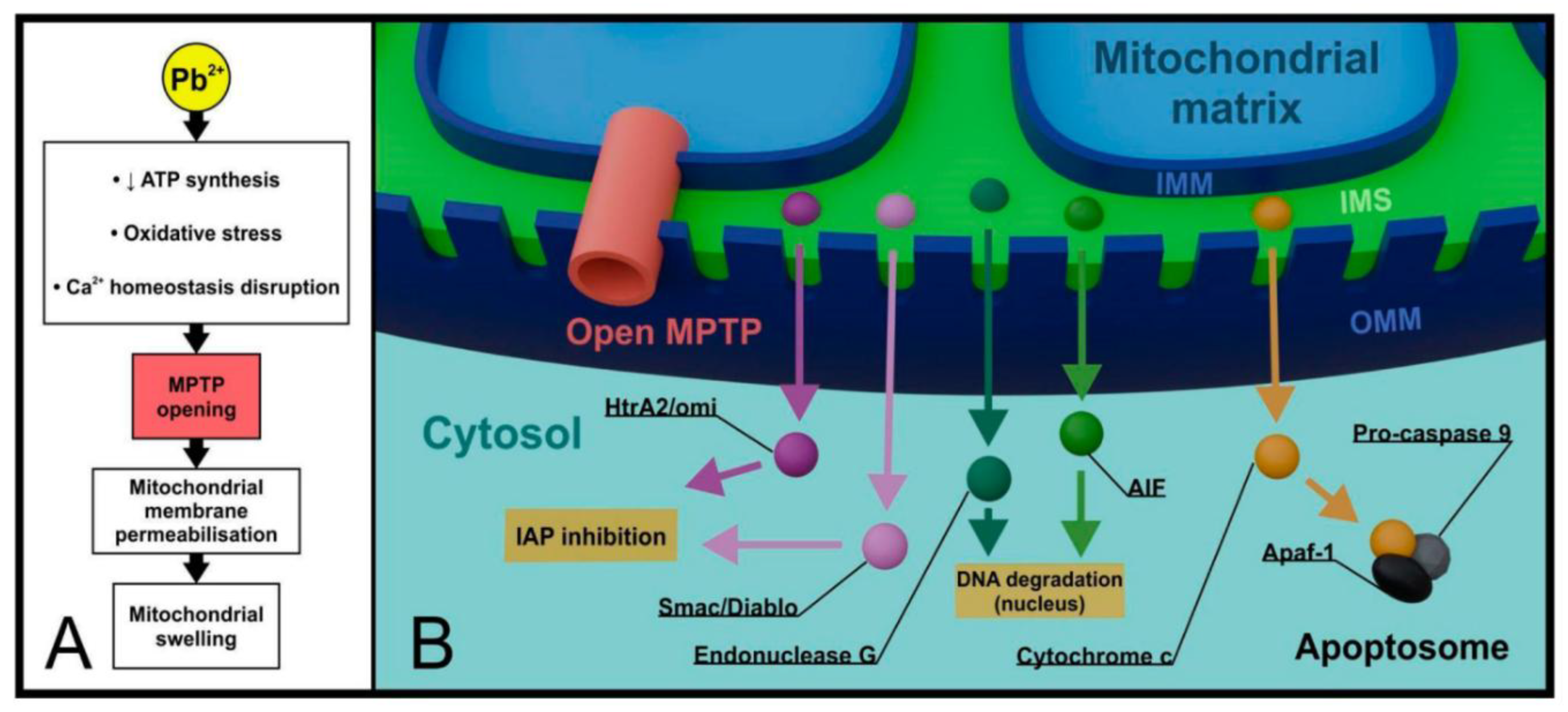
Figure 9.
Microbial remediation mechanisms against heavy metals, including Pb [36].
Figure 9.
Microbial remediation mechanisms against heavy metals, including Pb [36].
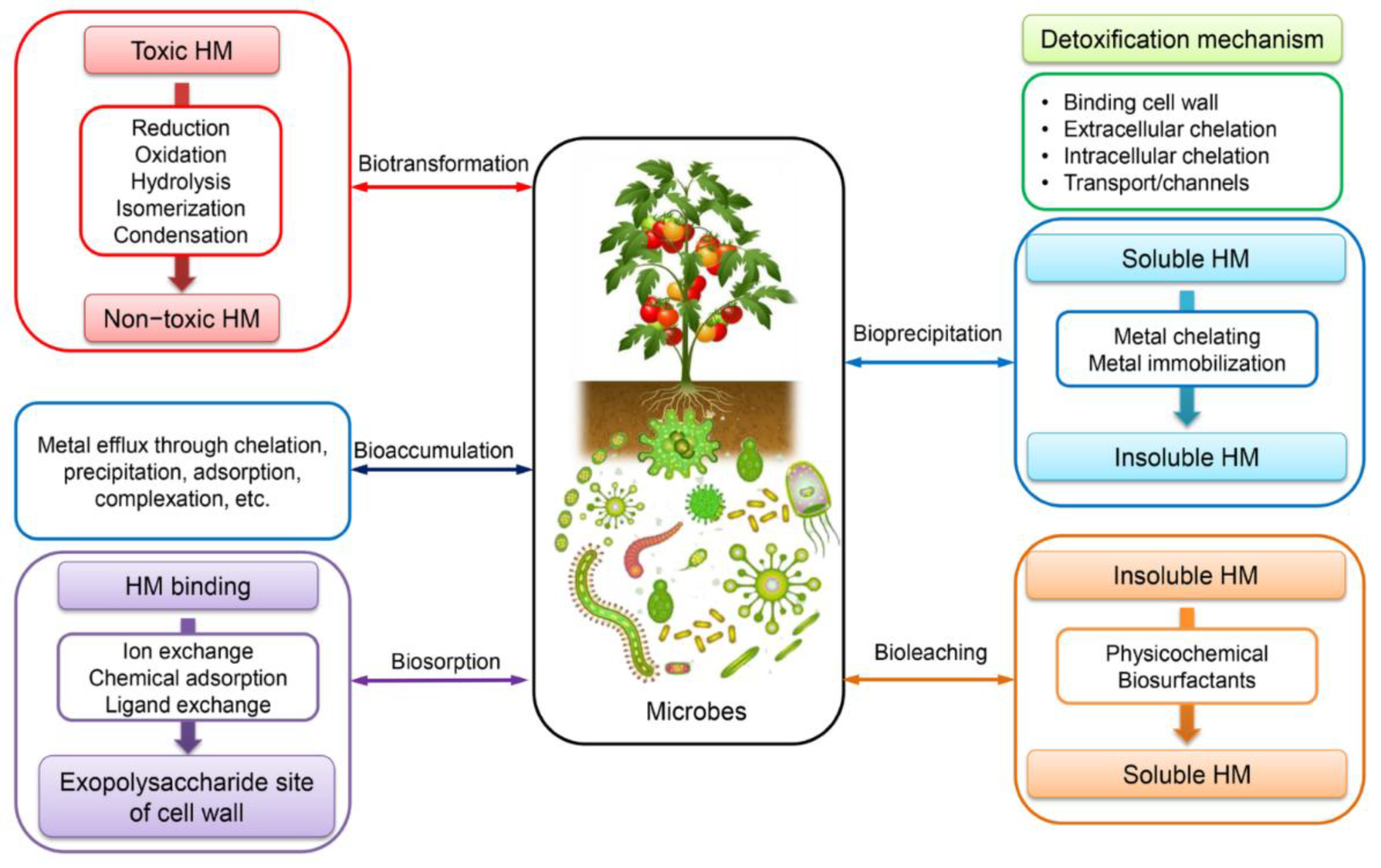
Figure 10.
The NMR structure of the α-domain and β-domain of rat MT-2. The α-domain has four metal ion binding sites, and the β-domain has three metal ion binding sites [128].
Figure 10.
The NMR structure of the α-domain and β-domain of rat MT-2. The α-domain has four metal ion binding sites, and the β-domain has three metal ion binding sites [128].

Table 1.
Physical and chemical properties of Pb [1].
Table 1.
Physical and chemical properties of Pb [1].
| Property | Value |
|---|---|
| Atomic number Atomic weight Atomic radius Electronic configuration Melting point Boiling point Density at 20 °C Reduction potential Pb²⁺ + 2e⁻ → Pb(s) Heat of fusion Heat of vaporization Electronegativity (Pauling scale) First ionization energy Second ionization energy |
82 207.2 u 180 pm (Empirical) [Xe]6s²4f¹⁴5d¹⁰6p² 327.46 °C 1749 °C 11.342 g/cm³ −0.126 V 4.77 kJ/mol 179.5 kJ/mol 2.33 7.417 eV 15.03 eV |
Table 2.
Lead Concentration (mg/kg) in Soil, Dust, and Marine Samples from Selected Global Regions. Table is optimized after [18,19,9].
| Sample | Lead Concentration Range |
|---|---|
| Urban soil, Klang district, Selangor | 52.7 |
| Soil from the mango plantation area, Perlis | 0.4 |
| Surface soils from iron ore mining sites, Kuala Lipis, Pahang | 63.5–72.5 |
| Grassland, arable land, forest, wasteland, Malopolska, Poland | 3-586 |
| Average concentration of heavy metals in the Earth's crust | 12.5 |
| Dust fall particles, Zarand, Iran | 1.01 |
| Road dust, Delhi city, India | 128.7 |
| Red Sea (North), Gulf of Aqaba | 96.67 |
| Red Sea (North), Hurghada City | 53 |
Table 3.
Maximal permissible limits of Pb.
| Metric | Pb | Ref. |
|---|---|---|
| Food | 0.01-3 mg/kg | [27] |
| Drinking water | 5 µg/L | [28] |
| Soil | 50–300 mg/kg | [29] |
| Air | 0.5 µg/m3 | [30] |
Table 4.
Measured Lead Levels in Selected Products. Table is optimized after [81].
Table 4.
Measured Lead Levels in Selected Products. Table is optimized after [81].
| Food Type | Country | Concentration (μg/kg or μg/L) | Year | |
|---|---|---|---|---|
| FRUITS | Grape white varieties | Croatia | 0.001–0.021 | 2008 |
| Grape red varieties | Croatia | 0.002–0.039 | 2008 | |
| Banana | Bangladesh | 3 | 2016 | |
| Mango | Bangladesh | 642 | 2016 | |
| Apples | Kosovo | 1490–2170 | 2019 | |
| Apples | Ukraine | 1347–3886 | 2021 | |
| VEGETABLES | Lettuce | Romania | 820–2220 | 2021 |
| Tomato | Romania | 0.7–0.8 | 2023 | |
| Potato | China | 67 | 2009 | |
| Potato | Bangladesh | 7 | 2016 | |
| White potato | Romania | 300–400 | 2021 | |
| Red potato | Romania | 370–1030 | 2021 | |
| Onion | Romania | 160–180 | 2021 | |
| Carrot | Romania | 540–940 | 2021 | |
| Beans | Romania | 80–520 | 2021 | |
| Soybean | Monte Carlo | 33–70 | 2022 | |
| Grain, maize | China | 20–13 | 2015 | |
| MEATS | Pork meat products | Italy | 220–380 | 2020 |
| Pork | Italy | 0.024 | 2020 | |
| Bacon | Romania | 580 | 2014 | |
| Ham | Romania | 650 | 2014 | |
| Salami | Romania | 210 | 2014 | |
| Sausages | Romania | 820 | 2014 | |
| Red meat | Asia | 605–1435 | 2023 | |
| Red meat | Africa | 840–1094 | 2023 | |
| Beef | Italy | 0.019 | 2020 | |
| Mutton meat | China (Beijing) | 128 | 2019 | |
| DAIRY | Milk | Monte Carlo | 550 | 2023 |
| Milk | Turkey | 0.85 | 2023 | |
| Milk | Tanzania | 263 | 2023 | |
| Raw cow milk | Turkey | 16.7 | 2012 | |
| Raw cow milk | Egypt | 101.6 | 2023 | |
| Sheep and goat milk | Italy | 0.002 | 2020 | |
| Milk and dairy products | Egypt | 0.044–0.751 | 2014 | |
| Full-fat UHT milk | Cyprus | 2.66 | 2021 | |
| Full-fat yogurt | Cyprus | 3 | 2021 | |
| Halloumi cheese | Cyprus | 35.3 | 2021 | |
| OILS | Corn oil | Iran | 99 | 2020 |
| Olive oil | London | 143 | 2022 | |
| Olive oil | Pakistan | 4285 | 2022 | |
| Rapeseed oil | China | 1960 | 2016 | |
| Rapeseed oil | Poland | 56 | 2017 | |
| Coconut oil | London | 158 | 2022 | |
| Sesame oil | Pakistan | 4005 | 2022 | |
| Sesame oil | Korea | 36 | 2019 | |
| Sunflower oil | London | 274 | 2022 | |
| Sunflower oil | Iran | 99 | 2020 | |
| Flaxseed oil | Korea | 25.7 | 2019 | |
| DRINKS | Beer | Ethiopia | 6 | 2022 |
| Beer | Brazil | 13–33 | 2005 | |
| Muscat Ottonel | Romania | 2.5–632 | 2017 | |
| low-alcoholic Muscat Ottonel | Romania | 67–575 | 2017 | |
| White wine | Croatia | 30 | 2008 |
Table 5.
Effect of Lead on liver. Table is optimized after [110].
Table 5.
Effect of Lead on liver. Table is optimized after [110].
| Effect | Concentration | Exposure time | Biological models | Mode of action | The outcome of treatment |
|---|---|---|---|---|---|
| Oxidative stress | Lead acetate (Pb 0.2%) | 5 weeks | Rat | Upregulating the transcription process of the cyclooxygenase-2 gene | Oxidative stress, lipid peroxidation |
| Ultrastructural changes | 0.13% lead acetate | 4–8 weeks | Adult albino rats | Megalocytosis complex III of the respiratory chain affected | Nuclear pyknosis, juxtanuclear inclusion bodies |
| Cholesterol functions of the liver | Lead acetate (500 mg Pb/L) | 10-11 weeks | Male Wistar rats | Inhibition of the activity of HMGR and decrease in the expression of cholesterol 7 alpha-hydroxylase (CYP7A1) genes | reduction of metabolism of cholesterol, increase in plasma cholesterol levels |
| Metabolic functions | Lead acetate or lead nitrate (20 mg/kg) | 4 weeks | Swiss albino male mice | Reduced enzymatic activity of glucose-6-phosphatase (G6PASE) | pyruvic acid content was increased, disruption in glycogen related mechanisms |
| Hepatic hyperplasia | Lead acetate trihydrate | 4-52 weeks | Wistar Albino Rats | Increase in the activity of DNA polymerase-β, Protein kinase C alpha (PKC-α) overexpression, suppression of the mRNA of the CYP1A2 gene, increased production of TNF-α | hyperplasia of Kupffer cells, oxidative stress of the hepatocytes |
| Cell death | Lead acetate, 1 mg/ml | 1 week | Female mice | Overexpression of apoptotic markers like Bax, Caspase 8, Caspase 3 | Apoptosis, Oxidative stress |
Table 6.
Biosorption by different microbes. Table is optimized after [129].
Table 6.
Biosorption by different microbes. Table is optimized after [129].
| Microbial Biosorbent | pH | Temperature (°C) | Time (h) | Initial Metal Ion Concentration (mg/L) | Sorption Capacity (mg/g) |
|---|---|---|---|---|---|
| Enterobacter cloacae | 8 | 40 | 72 | 400 | 172 |
| Pseudomonas aeruginosa | 7.5 | 40 | 24 | 50 | 40 |
| Micrococcus luteus | - | 27 | 48 | 272 | 1965 |
| Aspergillus niger | 4.5 | 30 | 72 | 100 | 34.4 |
| Aspergillus fumigatus | 4 | 30 | 48 | 100 | 35 |
| Saccharomyces cerevisiae | 8 | 60 | 6 | 98.25 | 80 |
| Phanerochaete chrysosporium | 6 | 20 | 1 | 100 | 88.16 |
| Botrytis cinerea | 4 | 25 | 1.5 | 350 | 107.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



