
Submitted:
09 July 2025
Posted:
11 July 2025
You are already at the latest version
Abstract
Carbapenem-resistant Klebsiella pneumoniae (CRKP) is an urgent public health threat due to its rapid dissemination and resistance to last-line antibiotics. Cefiderocol (FDC), a novel siderophore cephalosporin, targets resistant Gram-negative pathogens by exploiting bacterial iron uptake mechanisms. However, resistance to FDC is emerging among KPC-producing K. pneumoniae strains. This study characterizes a spontaneous FDC-resistant subpopulation (IHC216) derived from a KPC-producing strain (KPNMA216) using comprehensive genomic, transcriptional, and phenotypic analyses. Given the whole-genome sequencing results, where mutations were identified in genes involved in transcriptional regulation and membrane permeability (ompC) among others, in the present work we further explore their potential implications and conduct a more detailed analysis of the IHC216 genome. qRT-PCR analysis highlighted significant downregulation of iron acquisition systems (fepA, cirA, iroN) and upregulation of alternative iron uptake pathways (iucA, fiU), reflecting a metabolic shift. A notable downregulation of blaKPC-163 correlated with restored susceptibility to carbapenems, indicating collateral susceptibility. Altered expression of pbp2 and pbp3 implicated adaptive changes in cell wall synthesis, potentially affecting FDC resistance mechanisms. Furthermore, enhanced oxidative stress responses via upregulated sodC expression and increased capsule production were observed. These findings underscore the complex interplay of genetic and transcriptional adaptations underlying FDC resistance, highlighting potential therapeutic vulnerabilities.
Keywords:
KPC
; Cefiderocol
; Klebsiella
; Carbapenem‐resistant
1. Introduction
Carbapenem-resistant K. pneumoniae (CRKP) has emerged as a significant global health concern due to its high level of resistance to multiple antibiotics, including carbapenems, which are often considered last resort treatments for severe infections. The rapid dissemination of these resistant strains poses substantial challenges in clinical settings, particularly in intensive care units (ICUs), where outbreaks can increase morbidity and mortality among critically ill patients [1]. Treatment options for infections caused by CRKP are increasingly limited, particularly as these strains develop resistance to last-line antibiotics such as ceftazidime/avibactam and colistin [2]. Studies have shown that combination therapy may be more effective than monotherapy in treating CRKP infections, with significantly more treatment failures observed in cases that received monotherapy compared to those who received combination therapy [3].
Cefiderocol (FDC) is a novel siderophore cephalosporin designed to combat infections caused by carbapenem-resistant Gram-negative bacteria, including CRKP strains. Its unique mechanism involves utilizing the bacterial iron transport system to enter the cell, thereby inhibiting cell wall synthesis [4, 5]. However, emerging evidence indicates that apart of mutation in iron uptake systems, certain NDM and KPC-producing K. pneumoniae strains are developing resistance to FDC (ref). Regarding KPC, specific variants of KPC has been linked with FDC resistance (REF). Hobson et al. revealed that KPC-31 confers cross-resistance to both ceftazidime-avibactam and FDC (REF). Additionally, a significant inoculum effect on FDC was observed, indicating that higher bacterial loads may diminish the antibiotic's efficacy [6].
Our group have recently found collateral-resistance to FDC and cefepime/zidebactam (FPZ) in eight and three of the Klebsiella isolates carrying different KPC variants, respectively (manuscript submitted). We observed that five strains exhibited the occurrence of colonies within the inhibition ellipse zones of FDC strips, suggesting the occurrence of heteroresistance or spontaneous resistant mutants. This phenomenon has been observed in gram-negative organisms and has been link to FDC resistance and to last resort antibiotics such as colistin, meropenem and ceftazidime-avibactam (CZA) [7-10].
The objective of this study is to characterize a randomly FDC resistant subpopulation (IHC216) by conducting a comprehensive genomic, transcriptional, and phenotypic analysis in comparison to its parental strain.
2. Results
2.1. Whole Genome Sequencing Analysis
As previously reported, the comparison of KPNMA216 and KPNMA216 IHC demonstrated that there are no gene content differences, however, we found 14 mutations, being 5 SNPs and 9 InDels. Remarkably, we have observed a non-synonymous difference in the dksA gene, a global transcription regulator, that it was reported to make a significant phenotype global transcription regulator, and that it was reported to make significant phenotype changes in different bacterial species. There are few differences; the main differences arere SNP/InDel level.
In detail comparative analysis of the KPNMA 216 genome with all 477 K. pneumoniae ST14 genomes available in the Genome NCBI database (Table S1) revealed 1644 conserved gene families and four unique genes in KPNMA216. Among the unique genes, we found the insertion sequence ISApu2 and the fragment of the transposase of ISKpn54, traD (conjugative system), tssK (Type VI Secretion System) genes.
A core-genome phylogenetic analysis was performed of KPNMA216 genome and 477 K. pneumoniae ST14 genomes available in Genome NCBI Database (Figure 1, Figure S1). From the phylogenetic analysis of K. pneumoniae, the KPNMA216 strain was clustered in monophyletic group that included isolates recovered from Europe (Germany, Greece, Turkey, France, United Kingdom, Italy and United Kingdom) (Figure 1). A temporal trend was observed within the KPNMA216 clade, with the most of isolates being recovered in 2014. KPNMA216 was isolated in 2022, potentially indicating a recent emergence and suggesting that the strain may have originated in Europe (Figure 1).
Fifty-two genes associated virulence phenotype was identified using VFDB database. Among the virulence genes in K. pneumoniae, we found genes involved in type VI secretion system, fimbrial biogenesis, etc.[11-13]. The capsular polysaccharide is a crucial virulence factor in Gram-negative bacteria, enabling resistance to the bactericidal activity of the complement system. K. pneumoniae KPNMA216 contains the capsular polysaccharide biosynthesis loci (KL, K locus) and LPS loci (OCL, OC locus). Comparative analysis of the KL structure in KPNMA216 showed a 99% nucleotide identity, and 100% coverage with the KL2 type. The OCL locus, responsible for O antigen synthesis, was identified as O1/02v2 (Table S2 with Virulence genes)
2.2. Collateral Susceptibility to Carbapenems in the FDC-Resistant Mutant
When the MIC for FDC in IHC216 was evaluated, it increased from 8 mg/L in KPNMA216 to 32 mg/L (Table S3, Figure S2), confirming the development of FDC resistance. This finding indicates the emergence of stable FDC-resistant colonies within the inhibition ellipse (intracolonies) in KPC-producing strains.
To investigate potential collateral susceptibility and collateral resistance, MIC values for KPNMA216 and IHC216 were compared across a panel of antimicrobial agents. IHC216 exhibited a notable decrease in resistance to carbapenems, with meropenem MIC decreasing from 32 mg/L to 0.5 mg/L and imipenem MIC decreasing from 48 mg/L to 3 mg/L (Table 1). A similar trend was observed for imipenem/relebactam (from 2 mg/L to 0.75 mg/L) and meropenem/varbobactam (from 2 mg/L to 0.064 mg/L), indicating restored susceptibility to these β-lactam/β-lactamase inhibitor combinations (Table 1).
However, susceptibility to FPZ remained unchanged (MIC = 16 mg/L). Ciprofloxacin MIC decreased slightly (0.094 mg/L to 0.064 mg/L), but no significant changes were observed for ceftolozane/tazobactam, aztreonam, amikacin, or colistin. Interestingly, tigecycline MIC increased from 0.25 mg/L to a range of 0.75-1 mg/L, suggesting a potential fitness cost associated with the observed carbapenem susceptibility restoration (Table 1).
These findings highlight differential effects of the resistance mechanism on susceptibility patterns across various antimicrobial classes and underscore the complex interplay between β-lactam resistance and collateral susceptibility in KPC-producing K. pneumoniae.
2.3. Molecular and Transcriptional Adaptations Underlying FDC Resistance in the IHC216 Mutant
To further investigate the factors contributing to FDC resistance in the emergent resistant colony, we analysed the preliminary whole-genome comparative sequence data of the IHC216 strain against the wild-type (Previous paper). The analysis identified 14 non-synonymous mutations (Paper anterior and Table S3), of which seven directly affected coding regions. Among these, several mutations occurred in genes with known functions, including dksA, ompC, licC_7, gmuC, and lacE. These mutations suggest potential alterations in transcriptional regulation, membrane permeability, and carbohydrate transport, which may contribute to the observed resistance phenotype. Notably, no mutations were detected in iron uptake systems or siderophore production, indicating that FDC resistance in IHC216 is not driven by mutations on those genes. To further assess the molecular mechanisms underlying resistance, we performed quantitative real-time PCR (qRT-PCR) to evaluate the expression of genes associated with iron uptake, antibiotic resistance, cell wall synthesis, oxidative stress response, and aromatic compound catabolism in IHC216 compared to the parental strain (Figure 2 and Table S4). The analysis of iron uptake-associated transcripts revealed a marked downregulation of fepA and cirA, which encode siderophore receptors [14-16], as well as iroN, an enterobactin receptor. Additionally, a significant decrease in entB [17] expression was observed, indicating reduced enterobactin biosynthesis. These changes suggest a reduced expression on certain iron acquisition systems or regulatory adjustments in the IHC216 strain. Conversely, there was a significant upregulation of iucA, which encodes a key enzyme in aerobactin synthesis [18], and fiU, a siderophore receptor [14], indicating a shift toward alternative iron uptake mechanisms in the IHC216 environment. In contrast, fecA expression remained unchanged, suggesting that the ferric citrate transport system is not significantly affected under these conditions (Figure 2 A, Table S4).
The expression of dksA, a key regulator influencing iron homeostasis and mutated in the IHC216, was downregulation, suggesting that the mutation may be affecting the expression of this gene, leading to the observed changes in iron metabolism (Fig. 2 B, Table S4). Given DksA’s role in bacterial stress responses and iron acquisition, its repression may impact siderophore production and iron uptake pathways, potentially influencing FDC resistance mechanisms.
Additionally, genes involved in oxidative stress responses, known to be regulated by DksA. exhibited notable changes. Specifically, sodC, which encodes a copper-zinc superoxide dismutase (Cu/Zn-SOD) involved in neutralizing reactive oxygen species, was significantly upregulated. This suggests an enhanced oxidative stress defence mechanism, likely in response to increased oxidative challenges in the presence of FDC. In addition, a metabolic shift was observed in aromatic compound catabolism, pathway control by dksA in A. baumannni [19, 20], as pcaL, which encodes a key enzyme involved in the degradation of phenolic acids, was significantly downregulated (Fig. 2 B, Table S4). Moreover, the expression of katE was found to be significantly lower in strain IHC216 compared to the parental strain. katE encodes the catalase-peroxidase enzyme, which plays a role in the bacterial oxidative stress response by breaking down hydrogen peroxide (H₂O₂) into water and oxygen. This enzyme helps protect Klebsiella from oxidative damage, especially under stress conditions such as exposure to reactive oxygen species (ROS) or during host immune responses [21]. These results suggest that strain IHC216 exhibits an impaired oxidative stress response, potentially affecting its ability to detoxify hydrogen peroxide (Figure 2 B, Table S4).
In terms of antibiotic resistance, the expression of several critical genes was significantly reduced. blaKPC-163, encoding carbapenemase enzymes responsible for resistance to carbapenems [22], showed notable downregulation, which can explain the collateral susceptibility of IHC216 towards carbapenems (Table 1). However, the outer membrane porins coding genes, ompK35 and ompK36 [23], which facilitate the influx and efflux of antibiotics, were also downregulated, potentially altering membrane permeability and reducing resistance potential supporting cefepime/zidebactam (FPZ) resistance. Furthermore, the two-component regulatory system genes baeR and baeS, associated with multidrug resistance and membrane stress responses, exhibited decreased expression. These findings suggest a diminished antibiotic resistance capability in the IHC216, possibly due to altered membrane dynamics or regulatory mechanisms. Regarding cell wall synthesis, the expression of pbp2 and pbp3, which encode penicillin-binding proteins essential for peptidoglycan synthesis and cell wall integrity, was significantly reduced. This downregulation may reflect alterations in cell wall remodelling in the IHC216 which can affects FDC activity (Figure 2 C, Table S4). This suggests a reduced utilization of aromatic compounds in the KPNMA216 IHC strains, possibly reflecting broader metabolic adaptations associated with FDC resistance.
In the IHC216 strain, quantitative real-time PCR analysis revealed significant alterations in the expression of key genes associated with biofilm formation and capsule when compared to the parental strain. Specifically, the mrkA and wzm genes exhibited upregulation, while the wbbM gene was downregulated (Fig. 2 D, Table S4). The mrkA gene encodes the major structural subunit of type 3 fimbriae, which are crucial for biofilm formation and adherence to surfaces [24]. Wzm is involved in the transport of lipopolysaccharide (LPS) components across the inner membrane, playing a role in the assembly of the bacterial outer membrane [25]. Finally, WbbM is associated with the synthesis of D-galactan I, a component of the O-antigen in LPS [26]. These expression changes in IHC216 highlight a potential shift in its pathogenic profile, with implications for its biofilm-forming ability and interaction with host defences.
2.4. Increase Capsule and Biofilm Formation Was Seen in the FDC-Resistant Mutant
A key virulence factor in K. pneumoniae is the capsule, which plays a critical role in immune evasion, resistance to antimicrobial peptides, and protection against phagocytosis by host immune cells (Ref). Capsule production has been linked to increased antibiotic resistance, pathogenicity and survival in hostile environments, including resistance to complement-mediated killing. Capsule density was evaluated showing higher capsule production in the IHC216 strains compared to its parental strain (Figure 3A).
In addition, biofilm production, known to contribute to antimicrobial resistance, was performed for the wild-type and FDC-resistant mutant (IHC216). We observed increase in biofilm formation for the IHC216 exhibited an increase in biofilm formation compared to KPNMA216 (Figure 3 B).
These results highlight potential trade-offs between capsule expression and biofilm formation in response to selective pressures in emergent of mutant population. Further investigation into the genetic mechanisms underlying these changes will provide deeper insights into the adaptation strategies of K. pneumoniae.
3. Discussion
The development of FDC resistance in KPC-producing K. pneumoniae represents a multifaceted adaptation involving genetic mutations, transcriptional reprogramming, and metabolic shifts. Previous reports have shown the FDC resistance in KPC-producing K. pneumoniae[14, 27, 28]. One mechanism involves the acquisition of plasmids carrying the ferric citrate transport (FEC) system, the presence of two co-resident plasmids—pKpQIL, which harbors variants of the blaKPC carbapenemase gene, and pKPN, containing the FEC system—can lead to decreased susceptibility to FDC in K. pneumoniae clinical isolates [14]. Additionally, mutations in the CirA receptor were reported in FDC-resistant hypervirulent K. pneumoniae [15]. Finally, the co-production of carbapenemases such as NDM and KPC has been shown to elevate the minimum inhibitory concentrations (MICs) of FDC [29]. Through whole-genome sequencing and transcriptomic analyses of the FDC-resistant subpopulation (IHC216), we revealed mutations in genes involved in transcriptional regulation, membrane permeability, iron uptake system, antibiotic resistance, and carbohydrate transport. These changes may contribute to resistance to FDC but also introduced collateral susceptibility to carbapenems, as observed in the significant reduction of meropenem (32 mg/L to 0.5 mg/L) and imipenem (48 mg/L to 3 mg/L) MICs.
Collateral susceptibility, defined as the unintended restoration of antibiotic sensitivity [30], was observed in IHC216, most likely due to the downregulation of blaKPC-163. The reduced expression of this KPC variant suggests that mutations leading to FDC resistance impose a regulatory cost on KPC expression. A similar phenomenon has been reported in KPC-31-producing strains, where a single amino acid substitution resulted in cross-resistance to both ceftazidime-avibactam and FDC [6]. Furthermore, previous studies have demonstrated that heteroresistant subpopulations of KPC-K. pneumoniae exhibit increased meropenem susceptibility under certain conditions, which aligns with our findings [31].
Interestingly, genes coding for penicillin-binding proteins pbp2 and pbp3 were also downregulated, suggesting potential cell wall remodeling in response to FDC exposure. Given that FDC inhibits peptidoglycan biosynthesis, these changes may represent an adaptive mechanism that enhances carbapenem susceptibility. In addition, the downregulation of these genes in the IHC condition may reflect a shift in bacterial physiology toward slower growth or altered division cycles. Additionally, reduced expression of pbp2 and pbp3 may serve as a protective mechanism, enhancing resistance to oxidative stress or antimicrobial agents. Similar regulatory patterns have been reported in biofilm-forming bacteria, where PBPs are differentially expressed to adapt to the biofilm matrix and reduced metabolic activity [32]. The reduced expression of ompK35 and ompK36, genes coding for outer membrane porins that facilitate β-lactam influx (REF), could further modify drug permeability, explaining why IHC216 remained resistant to cefepime/zidebactam despite restored carbapenem susceptibility.
Another important observation in IHC216 was the differential expression of iron acquisition genes. Given that FDC is a siderophore cephalosporin, bacterial iron uptake pathways play a central role in both FDC susceptibility and resistance development. The downregulation of fepA, cirA, iroN, and entB in IHC216 suggests that catecholate siderophore receptors became less relevant, potentially due to an environmental shift reducing iron competition. This finding is consistent with studies showing that bacteria downregulate siderophore receptor expression in iron-rich conditions or when alternative iron uptake systems are activated [33, 34]. Conversely, iucA and fiU were significantly upregulated, suggested a shift toward aerobactin-mediated iron acquisition, which aligns with reports demonstrating that aerobactin expression is favored in oxidative stress environments [35]. Additionally, the mutation and low expression of dksA in IHC216 suggests that this global regulator plays a central role in modulating iron metabolism. Previous work in Salmonella enterica has shown that DksA regulates iron homeostasis and oxidative stress responses, with its deletion leading to dysregulated iron uptake and increased sensitivity to reactive nitrogen species [36]. Furthermore, this effect extends to other metabolic pathways, as evidenced by the downregulation of pcaL, involved in aromatic compound metabolism, suggests a broader metabolic shift, potentially reflecting altered carbon source utilization in response to FDC exposure.
We have also observed, a significant upregulation of sodC, which encodes a copper-zinc superoxide dismutase (Cu/Zn-SOD), a major antioxidant defense enzyme in the IHC216. Increased oxidative stress resistance in IHC216 suggests that FDC exposure induces a reactive oxygen species (ROS)-rich environment, requiring an enhanced detoxification response. This agrees with previous findings that dense bacterial populations experience heightened oxidative stress due to metabolic activity and host immune responses [37, 38].
In addition to oxidative stress defenses, capsule production was significantly increased in IHC216. The capsule plays a crucial role in immune evasion and antimicrobial resistance [39, 40], and its upregulation in IHC216 may serve as a compensatory adaptation to balance FDC-induced permeability and iron acquisition shifts. Notably, capsule production has been previously associated with increased FDC resistance, suggesting that enhanced polysaccharide biosynthesis might limit drug penetration into the bacterial cell.
Lastly, biofilm formation was also increased significant in IHC216. While biofilms contribute to antimicrobial resistance, their development is often influenced by capsule production, with a potential trade-off between biofilm adherence and immune evasion. Recent studies suggest that capsule-deficient strains tend to form more robust biofilms, while hypermucoid variants rely more on capsular protection for survival.
This study highlights the complex regulatory and metabolic adaptations underlying FDC resistance in KPC-producing K. pneumoniae. This work showed that apart of recognized resistance mechanisms, IHC216 displayed collateral susceptibility to carbapenems, which could inform novel treatment strategies leveraging this vulnerability. The downregulation of blaKPC-163, alterations in pbp expression, and outer membrane modifications suggest that targeting cell wall synthesis pathways in combination with carbapenems may restore treatment efficacy. Moreover, the observed iron uptake reprogramming in IHC216 highlights potential therapeutic avenues focusing on siderophore-mediated drug delivery. Given that aerobactin synthesis was upregulated, targeting this pathway with siderophore-conjugated antimicrobials could offer a more effective strategy against FDC-resistant strains. Additionally, the induction of oxidative stress defense pathways suggests that combining FDC with ROS-enhancing agents could further sensitize resistant populations.
Recent studies have also reported heteroresistance to FDC in K. pneumoniae and Acinetobacter baumannii, where subpopulations exhibit transient resistance under host-associated conditions. Our laboratory previously demonstrated that human pleural fluid could induce cefiderocol heteroresistance in carbapenem-resistant A. baumannii, further supporting the notion that host environments drive dynamic resistance evolution. Similarly, mutations in cirA—a FDC receptor—were identified during in vivo evolution of a K. pneumoniae ST512 strain, leading to high-level FDC resistance.
In summary, the present study provides novel insights into the genetic, transcriptional, and phenotypic landscape of FDC resistance in KPC-producing K. pneumoniae. The interplay between iron homeostasis, oxidative stress defense, and membrane permeability highlights multiple vulnerabilities that could be targeted for therapeutic intervention and highlights the complexity of antibiotic resistance. The collateral susceptibility to carbapenems suggests that combination therapies exploiting this trade-off may help circumvent FDC resistance. Future investigations should focus on the clinical implications of these resistance mechanisms to develop optimized treatment strategies against multidrug-resistant K. pneumoniae and cautiously decide the optimal treatment.
4. Materials and Methods
4.1. Bacterial Strains
The KPC-producing K. pneumoniae KPNMA216 and the KPNMA216 FDC-resistant subpopulation (IHC216) strains were further analyzed in the present study (Table S3) (Hamza, paper submitted for publication). The IHC216 strain was recovered within the inhibition ellipse zones of FDC strips during the MIC determination of the KPNMA216 parental strain (Figure S2). Copies of the IHC isolates were kept at -80 °C as Luria Bertani (LB) broth containing 20% glycerol stocks that were plated on Cystine–Lactose–Electrolyte-Deficient (CLED) medium (Beckton Dickinson, Franklin Lakes, NJ, USA) and used within 24 h after overnight (16–18 h) incubation at 37 °C. The resistance phenotype stability was determined after 10 daily subcultures in CLED antibiotic-free plates.
4.2. Whole Genome Sequencing Analysis
The whole genome sequencing data of the parental strain KPNMA216 and the IHC216 generated in our previous study (Hamza, M submitted for publication), were further used for detailed resistome analysis. The fastq files, assemblies, and annotations are deposited in Zenodo (https://zenodo.org/records/14019667, accessed July 1st, 2025).
The genomes were annotated using PROKKA (version 1.14.5) [41]. The ortholog functional assignment was performed using EggNOG v2.0 (default parameter) [42]. The pangenome analysis was done using Roary software (REF). To assess core genome phylogeny, we used 477 K. pneumoniae ST14 sequences from a total of 23578 K. pneumoniae genomes available in the GenBank (Table S1). Core genome phylogeny analysis was performed using the maximum likelihood method, implemented with IQtree2 using default parameters [41].
tRNA and ncRNA predictions were conducted using tRNAscan-SE (version 1.3) and Infernal (version 1.1.5) software, respectively [43], and the Multilocus Sequence Typing (MLST) profile, OC and K locus, ICEKp associated virulence loci, colibactin (clb), salmochelin (iro), hypermucoidy (rmpA) was determined using Kleborate software [43]. Others Virulence genes were predicted using VFDB database, respectively.
4.3. RNA Extraction and Transcriptional Analysis Using Quantitative RT-qPCR
Overnight cultures of KPNMA216, and IHC216 were diluted 1:10 in LB and incubated with agitation for 18 h at 37°C. RNA was extracted from each sample using the Direct-zol RNA Kit (Zymo Research, Irvine, CA, USA) following the manufacturer’s instructions. Total RNA extractions were performed using three independent biological replicates for each condition.
The RNA samples obtained were subjected to DNase treatment (Thermo Fisher Scientific, Waltham, MA, USA) following manufacturer’s instruction, afterwards a PCR amplification of the 16S rDNA gene was performed to confirm there was no DNA contamination.
In addition, the extracted DNase-treated RNA was used to synthesize cDNA according to the iScript™ Reverse Transcription Supermix for qPCR reagents (Bio-Rad, Hercules, CA, USA) manufacturer's protocol. The cDNA concentrations were adjusted to 50 ng/μL, and qPCR was conducted using the qPCRBIO SyGreen Blue Mix Lo-ROX following the manufacturer's protocol (PCR Biosystems, Wayne, PA, USA).
Transcriptional analysis of KPNMA216, and IHC216 was performed using specific primers (Table S5). At least three independent cDNA replicates were tested in triplicate using the CFX96 Touch™ Real-Time PCR Detection System (Bio-Rad, Hercules, CA, USA). Transcriptional levels of each sample were normalized to the transcriptional level of recA. The relative quantification of gene expression was performed using the comparative threshold method 2 -ΔΔCt [44]. Differences were determined by ANOVA followed by Tukey’s multiple comparison test (P < 0.05) using GraphPad Prism (GraphPad Software, San Diego, CA, USA).
4.4. Antimicrobial Susceptibility Testing
The minimum inhibitory concentrations (MICs) for meropenem (MEM), imipenem (IMP), imipenem/relebactam (I/R), meropenem/vaborbactam (M/V), cefepime/zidebactam (FPZ), ciprofloxacin (CIP), ceftolozane/tazobactam (C/T), aztreonam (ATM), amikacin (AK), tigecycline (TGC), and colistin (CS) were determined using commercial E-strips (Liofilchem S.r.l., Roseto degli Abruzzi, Italy). The MICs for colistin was determined using the broth microdilution methods. All procedures were carried out in accordance with the manufacturer's instructions and met the standards of the Clinical and Laboratory Standards Institute (CLSI) [45] and the European Committee on Antimicrobial Susceptibility Testing (EUCAST) (https://www.eucast.org/clinical_breakpoints). The CLSI breakpoint was used for cefepime was used for FPZ categorization. Quality control strains, such as Escherichia coli ATCC 25922 and the K. pneumoniae ATCC strains were included in the experiments. Each strain was tested at least in duplicates.
4.5. Capsule and Biofilm
Capsule production assay was performed according Valcek et al. [46]. Briefly, overnight bacterial cultures (1 mL) of KPNMA216 and KPNMA216IHC were transferred to 1.5-mL microtubes and centrifuged at 7,000 × g for 2 minutes. After removing the supernatant, the pellet was resuspended in 1 mL of phosphate-buffered saline (PBS). A total of 875 µL of the PBS-resuspended bacteria was then mixed with 125 µL of Ludox LS colloidal silica (30% [wt/wt] suspension in H₂O; Merck). The mixture was centrifuged at 12,000 × g for 30 minutes, and the resulting band position was immediately photographed. The distance from the center of the band to the bottom of the microtube was measured. Each experiment was performed in triplicate, and results were statistically analyzed by calculating the standard deviation from the mean of biological replicates.
Biofilm assays were performed as previously described [47, 48]. KPNMA216 and IHC216 cells were cultured in LB broth and incubate the tubes 37 for 24-48 h without shaking (static). Experiments were performed in triplicate, with at least three technical replicates per biological replicate. Statistical analysis was performed using the test with GraphPad Prism, and a P-value <0.05 was considered statistically significant.
Supplementary Materials
Supplementary Materials associated with this article can be found, in the online version. Figure S1. Core-genome phylogenetic analysis of the KPNMA216 genome in comparison with 477 publicly available Klebsiella pneumoniae ST14 genomes retrieved from the NCBI Genome database. Figure S2. The IHC216 strain was recovered from within the inhibition ellipse zones of FDC strips during the MIC determination of the parental KPNMA216 strain. The IHC216 colony is shown marked with a red circle. Table S1. Collection of 477 publicly available Klebsiella pneumoniae ST14 genomes retrieved from the NCBI Genome database. Table S2. Virulence genes prediction in K. pneumoniae using the Blastp method. Table S3. Parental KPNMA216 and IHC216 relevant mutations associated with antibiotic resistance and MIC. Table S4. Level of expression of different genes obtained by qRT-PCR in KPNMA216 and IHC216. Table S5. qRT-PCR primers used in this study.
Author Contributions
GMT, MCR, GR, FP and MSR conceived the study and designed the experiments. VM, IL, JE, GMT, MRT, CR, GR and MSR performed the experiments and genomics and bioinformatics analyses. GMT, FP, JC, QV, RS, MRT, MCR, and MSR analyzed the data and interpreted the results. MET, RAB, GR and M.S.R. contributed reagents/materials/analysis tools. GMT, MRT, CR, MET, RAB, GR, FP and MSR wrote and revised the manuscript. All authors read and approved the final manuscript.
Funding
The authors' work was supported by NIH SC3GM125556 to MSR, 2R15 AI047115 to MET, and R01AI170889 to GGR. This study was supported in part by funds and facilities provided by the Cleveland Department of Veterans Affairs, Award Number 1I01BX001974 to RAB from the Biomedical Laboratory Research & Development Service of the VA Office of Research and Development and the Geriatric Research Education and Clinical Center VISN 10 to RAB. The content is solely the authors' responsibility and does not necessarily represent the official views of the National Institutes of Health or the Department of Veterans.
Informed Consent Statement
Not required.
Conflicts of Interest
The authors declare no conflict of interest
References
- Kerneis, S.; Lucet, J.C.; Santoro, A.; Meschiari, M. Individual and collective impact of Klebsiella pneumoniae carbapenemase (KPC)-producing K. Pneumoniae Patients Admit. ICU. J Antimicrob Chemother. 2021, 76, i19–i26. [Google Scholar]
- Li, T.; Zhu, Y.; Xiang, G.; Xu, Z.; Yang, H.; Li, M.; et al. Adaptive evolution of extensive drug resistance and persistence in epidemic ST11 KPC-producing Klebsiella pneumoniae during antimicrobial chemotherapy. Antimicrob Agents Chemother. 2025, 69, e0123524. [Google Scholar] [CrossRef]
- Lee, G.C.; Burgess, D.S. Treatment of Klebsiella pneumoniae carbapenemase (KPC) infections: a review of published case series and case reports. Ann Clin Microbiol Antimicrob. 2012, 11, 32. [Google Scholar] [CrossRef]
- Zhanel, G.G.; Golden, A.R.; Zelenitsky, S.; Wiebe, K.; Lawrence, C.K.; Adam, H.J.; et al. Cefiderocol: A Siderophore Cephalosporin with Activity Against Carbapenem-Resistant and Multidrug-Resistant Gram-Negative Bacilli. Drugs. 2019, 79, 271–89. [Google Scholar] [CrossRef]
- Bianco, G.; Boattini, M.; Cricca, M.; Diella, L.; Gatti, M.; Rossi, L.; et al. Updates on the Activity, Efficacy and Emerging Mechanisms of Resistance to Cefiderocol. Curr Issues Mol Biol. 2024, 46, 14132–53. [Google Scholar] [CrossRef] [PubMed]
- Hobson, C.A.; Cointe, A.; Jacquier, H.; Choudhury, A.; Magnan, M.; Courroux, C.; et al. Cross-resistance to cefiderocol and ceftazidime-avibactam in KPC beta-lactamase mutants and the inoculum effect. Clin Microbiol Infect. 2021, 27, 1172–e7. [Google Scholar] [CrossRef]
- Islam, M.M.; Jung, D.E.; Shin, W.S.; Oh, M.H. Colistin Resistance Mechanism and Management Strategies of Colistin-Resistant Acinetobacter baumannii Infections. Pathogens.
- Karakonstantis, S.; Rousaki, M.; Kritsotakis, E.I. Cefiderocol: Systematic Review of Mechanisms of Resistance, Heteroresistance and In Vivo Emergence of Resistance. Antibiotics (Basel).
- Bianco, G.; Boattini, M.; Iannaccone, M.; Cavallo, R.; Costa, C. Bloodstream infection by two subpopulations of Klebsiella pneumoniae ST1685 carrying KPC-33 or KPC-14 following ceftazidime/avibactam treatment: considerations regarding acquired heteroresistance and choice of carbapenemase detection assay. J Antimicrob Chemother. 2020, 75, 3075–6. [Google Scholar] [CrossRef]
- Ding, L.; Shen, S.; Chen, J.; Tian, Z.; Shi, Q.; Han, R.; et al. Klebsiella pneumoniae carbapenemase variants: the new threat to global public health. Clin Microbiol Rev. 2023, 36, e0000823. [Google Scholar] [CrossRef]
- Lin, T.L.; Lee, C.Z.; Hsieh, P.F.; Tsai, S.F.; Wang, J.T. Characterization of integrative and conjugative element ICEKp1-associated genomic heterogeneity in a Klebsiella pneumoniae strain isolated from a primary liver abscess. J Bacteriol. 2008, 190, 515–26. [Google Scholar] [CrossRef]
- Goetz, D.H.; Holmes, M.A.; Borregaard, N.; Bluhm, M.E.; Raymond, K.N.; Strong, R.K. The neutrophil lipocalin NGAL is a bacteriostatic agent that interferes with siderophore-mediated iron acquisition. Mol Cell. 2002, 10, 1033–43. [Google Scholar] [CrossRef]
- Bachman, M.A.; Miller, V.L.; Weiser, J.N. Mucosal lipocalin 2 has pro-inflammatory and iron-sequestering effects in response to bacterial enterobactin. PLoS Pathog. 2009, 5, e1000622. [Google Scholar] [CrossRef]
- Polani, R.; De Francesco, A.; Tomolillo, D.; Artuso, I.; Equestre, M.; Trirocco, R.; et al. Cefiderocol Resistance Conferred by Plasmid-Located Ferric Citrate Transport System in KPC-Producing Klebsiella pneumoniae. Emerg Infect Dis. 2025, 31, 123–4. [Google Scholar] [CrossRef]
- Lan, P.; Lu, Y.; Liao, W.; Yu, Y.; Fu, Y.; Zhou, J. Cefiderocol-resistant hypervirulent Klebsiella pneumoniae with CirA deficiency and co-production of KPC-2 and SHV-12. Clin Microbiol Infect. 2024. [CrossRef] [PubMed]
- Kumar, A.; Chakravorty, S.; Yang, T.; Russo, T.A.; Newton, S.M.; Klebba, P.E. Siderophore-mediated iron acquisition by Klebsiella pneumoniae. J Bacteriol. 2024, 206, e0002424. [Google Scholar] [CrossRef] [PubMed]
- Tsuka, T.; Kumashiro, S.; Kihara, T.; Iida, T. Correlation between Polymerase Chain Reaction Identification of Iron Acquisition Genes and an Iron-Deficient Incubation Test for Klebsiella pneumoniae Isolates from Bovine Mastitis. Microorganisms.2022;10.
- Daoud, L.; Al-Marzooq, F.; Moubareck, C.A.; Ghazawi, A.; Collyns, T. Elucidating the effect of iron acquisition systems in Klebsiella pneumoniae on susceptibility to the novel siderophore-cephalosporin cefiderocol. PLoS One. 2022, 17, e0277946. [Google Scholar] [CrossRef] [PubMed]
- Maharjan, R.P.; Sullivan, G.J.; Adams, F.G.; Shah, B.S.; Hawkey, J.; Delgado, N.; et al. DksA is a conserved master regulator of stress response in Acinetobacter baumannii. Nucleic Acids Res. 2023, 51, 6101–19. [Google Scholar] [CrossRef]
- Kim, N.; Son, J.H.; Kim, K.; Kim, H.J.; Kim, Y.J.; Shin, M.; et al. Global regulator DksA modulates virulence of Acinetobacter baumannii. Virulence. 2021, 12, 2750–63. [Google Scholar] [CrossRef]
- Switala, J.; Triggs-Raine, B.L.; Loewen, P.C. Homology among bacterial catalase genes. Can J Microbiol. 1990, 36, 728–31. [Google Scholar] [CrossRef]
- Carvalho, I.; Chenouf, N.S.; Carvalho, J.A.; Castro, A.P.; Silva, V.; Capita, R.; et al. Multidrug-resistant Klebsiella pneumoniae harboring extended spectrum beta-lactamase encoding genes isolated from human septicemias. PLoS One. 2021, 16, e0250525. [Google Scholar] [CrossRef]
- Ejaz, H. Analysis of diverse beta-lactamases presenting high-level resistance in association with OmpK35 and OmpK36 porins in ESBL-producing Klebsiella pneumoniae. Saudi J Biol Sci. 2022, 29, 3440–7. [Google Scholar] [CrossRef]
- Li, Y.; Ni, M. Regulation of biofilm formation in Klebsiella pneumoniae. Front Microbiol. 2023, 14, 1238482. [Google Scholar] [CrossRef] [PubMed]
- Shebl, R.I.; Elkhatib, W.F.; Badawy, M. Modulating the transcriptomic profile of multidrug-resistant Klebsiella pneumoniae biofilm formation by antibiotics in combination with zinc sulfate. Ann Clin Microbiol Antimicrob. 2023, 22, 84. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhang, Y.; Liu, Y.; Xu, M.; Yao, Z.; Zhang, X.; et al. Effects of chlorogenic acid on antimicrobial, antivirulence, and anti-quorum sensing of carbapenem-resistant Klebsiella pneumoniae. Front Microbiol. 2022, 13, 997310. [Google Scholar] [CrossRef] [PubMed]
- Castillo-Polo, J.A.; Hernandez-Garcia, M.; Morosini, M.I.; Perez-Viso, B.; Soriano, C.; De Pablo, R.; et al. Outbreak by KPC-62-producing ST307 Klebsiella pneumoniae isolates resistant to ceftazidime/avibactam and cefiderocol in a university hospital in Madrid, Spain. J Antimicrob Chemother. 2023, 78, 1259–64. [Google Scholar] [CrossRef]
- Findlay, J.; Bianco, G.; Boattini, M.; Nordmann, P. High-level cefiderocol and ceftazidime/avibactam resistance in KPC-producing Klebsiella pneumoniae associated with mutations in KPC and the sensor histidine kinase EnvZ. J Antimicrob Chemother. 2025, 80, 1155–7. [Google Scholar] [CrossRef]
- Yang, C.; Wang, L.; Lv, J.; Wen, Y.; Gao, Q.; Qian, F.; et al. Effects of different carbapenemase and siderophore production on cefiderocol susceptibility in Klebsiella pneumoniae. Antimicrob Agents Chemother. 2024, 68, e0101924. [Google Scholar] [CrossRef]
- Roemhild, R.; Andersson, D.I. Mechanisms and therapeutic potential of collateral sensitivity to antibiotics. PLoS Pathog. 2021, 17, e1009172. [Google Scholar] [CrossRef]
- Pournaras, S.; Kristo, I.; Vrioni, G.; Ikonomidis, A.; Poulou, A.; Petropoulou, D.; et al. Characteristics of meropenem heteroresistance in Klebsiella pneumoniae carbapenemase (KPC)-producing clinical isolates of K. pneumoniae. J Clin Microbiol. 2010, 48, 2601–4. [Google Scholar] [CrossRef]
- Stewart, P.S.; Franklin, M.J. Physiological heterogeneity in biofilms. Nat. Rev. Microbiology. 2008, 6, 199–210. [Google Scholar] [CrossRef]
- De Serrano, L.O.; Camper, A.K.; Richards, A.M. An overview of siderophores for iron acquisition in microorganisms living in the extreme. Biometals. 2016, 29, 551–71. [Google Scholar] [CrossRef]
- Page, M.G.P. The Role of Iron and Siderophores in Infection, and the Development of Siderophore Antibiotics. Clin Infect Dis. 2019, 69, S529–S37. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Pan, D.; Li, M.; Wang, Y.; Song, L.; Yu, D.; et al. Aerobactin-Mediated Iron Acquisition Enhances Biofilm Formation, Oxidative Stress Resistance, and Virulence of Yersinia pseudotuberculosis. Front Microbiol. 2021, 12, 699913. [Google Scholar] [CrossRef] [PubMed]
- Crawford, M.A.; Tapscott, T.; Fitzsimmons, L.F.; Liu, L.; Reyes, A.M.; Libby, S.J.; et al. Redox-Active Sensing by Bacterial DksA Transcription Factors Is Determined by Cysteine and Zinc Content. mBio. 2016, 7, e02161–e15. [Google Scholar] [CrossRef] [PubMed]
- Bonilla, C.Y. Generally Stressed Out Bacteria: Environmental Stress Response Mechanisms in Gram-Positive Bacteria. Integr Comp Biol. 2020, 60, 126–33. [Google Scholar] [CrossRef]
- Sun, F.; Liang, H.; Kong, X.; Xie, S.; Cho, H.; Deng, X.; et al. Quorum-sensing agr mediates bacterial oxidation response via an intramolecular disulfide redox switch in the response regulator AgrA. Proc Natl Acad Sci U S A. 2012, 109, 9095–100. [Google Scholar] [CrossRef]
- Raffatellu, M.; Chessa, D.; Wilson, R.P.; Tukel, C.; Akcelik, M.; Baumler, A.J. Capsule-mediated immune evasion: a new hypothesis explaining aspects of typhoid fever pathogenesis. Infect Immun. 2006, 74, 19–27. [Google Scholar] [CrossRef]
- Huang, X.; Li, X.; An, H.; Wang, J.; Ding, M.; Wang, L.; et al. Capsule type defines the capability of Klebsiella pneumoniae in evading Kupffer cell capture in the liver. PLoS Pathog. 2022, 18, e1010693. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: rapid prokaryotic genome annotation. Bioinformatics. 2014, 30, 2068–9. [Google Scholar] [CrossRef]
- Cantalapiedra, C.P.; Hernandez-Plaza, A.; Letunic, I.; Bork, P.; Huerta-Cepas, J. eggNOG-mapper v2: Functional Annotation, Orthology Assignments, and Domain Prediction at the Metagenomic Scale. Mol Biol Evol. 2021, 38, 5825–9. [Google Scholar] [CrossRef]
- Lam, M.M.C.; Wick, R.R.; Watts, S.C.; Cerdeira, L.T.; Wyres, K.L.; Holt, K.E. A genomic surveillance framework and genotyping tool for Klebsiella pneumoniae and its related species complex. Nat Commun. 2021, 12, 4188. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods. 2001, 25, 402–8. [Google Scholar] [CrossRef]
- (CLSI), C.L.S.I. Performance standards for antimicrobial susceptibility testing: Thirty Edition informational supplement. CLSI Document M100‐S30:2020. Clinical Lab Standards Institute, 2020. [Google Scholar]
- Valcek, A.; Philippe, C.; Whiteway, C.; Robino, E.; Nesporova, K.; Bove, M.; et al. Phenotypic Characterization and Heterogeneity among Modern Clinical Isolates of Acinetobacter baumannii. Microbiol Spectr. 2023, 11, e0306122. [Google Scholar] [CrossRef]
- Martinez, J.; Fernandez, J.S.; Liu, C.; Hoard, A.; Mendoza, A.; Nakanouchi, J.; et al. Human pleural fluid triggers global changes in the transcriptional landscape of Acinetobacter baumannii as an adaptive response to stress. Sci Rep. 2019, 9, 17251. [Google Scholar] [CrossRef]
- Mezcord, V.; Escalante, J.; Nishimura, B.; Traglia, G.M.; Sharma, R.; Valle, Q.; et al. Induced Heteroresistance in Carbapenem-Resistant Acinetobacter baumannii (CRAB) via Exposure to Human Pleural Fluid (HPF) and Its Impact on Cefiderocol Susceptibility. International journal of molecular sciences. 2023, 24. [Google Scholar] [CrossRef]
Figure 1.
A core-genome phylogenetic analysis was performed using the KPNMA216 genome and 477 Klebsiella pneumoniae ST14 genomes available in the NCBI Genome database. The KPNMA216 strain clustered within a monophyletic group located in a clade that included isolates recovered from various regions, including Germany, Greece, Turkey, France, the United Kingdom, and Italy.
Figure 1.
A core-genome phylogenetic analysis was performed using the KPNMA216 genome and 477 Klebsiella pneumoniae ST14 genomes available in the NCBI Genome database. The KPNMA216 strain clustered within a monophyletic group located in a clade that included isolates recovered from various regions, including Germany, Greece, Turkey, France, the United Kingdom, and Italy.
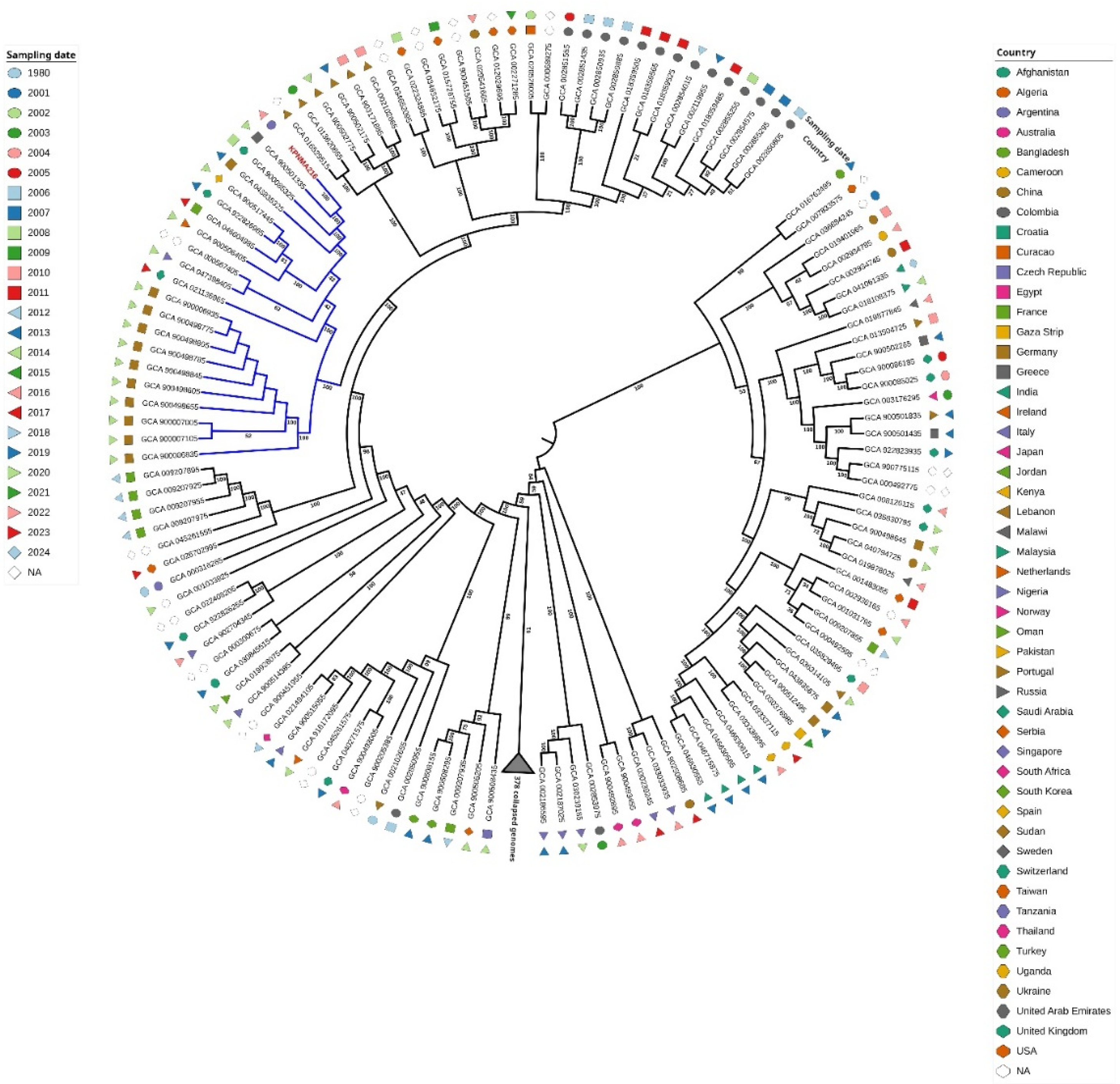
Figure 2.
Expression of genes coding for siderophores ( iucA and entB) and siderophores transporters (fepA, cirA, iroN, fiU and fecA) (A), dksA and genes regulated by DksA (sodC, pcaL and katE) (B), pbp2 and pbp3, β-lactamase blaKPC2-3, outer membrane porins ompK35 and ompK36 and the BaeRS two-component system (C), wzm, wbbM and mrkA (D) in the KPNMA216 and IHC216 strains. The data shown of qRT-PCR are mean ± SD. Fold changes were calculated using ΔΔCt analysis. At least three independent biological samples were tested using four technical replicates. Statistical significance (P < 0.05) was determined by two-way ANOVA followed by Tukey's multiple comparison test using GraphPad Prism (GraphPad software, San Diego, CA, USA). Significance was indicated by: *P < 0.05, **P < 0.01, ***P < 0.001, and **** P < 0.0001.
Figure 2.
Expression of genes coding for siderophores ( iucA and entB) and siderophores transporters (fepA, cirA, iroN, fiU and fecA) (A), dksA and genes regulated by DksA (sodC, pcaL and katE) (B), pbp2 and pbp3, β-lactamase blaKPC2-3, outer membrane porins ompK35 and ompK36 and the BaeRS two-component system (C), wzm, wbbM and mrkA (D) in the KPNMA216 and IHC216 strains. The data shown of qRT-PCR are mean ± SD. Fold changes were calculated using ΔΔCt analysis. At least three independent biological samples were tested using four technical replicates. Statistical significance (P < 0.05) was determined by two-way ANOVA followed by Tukey's multiple comparison test using GraphPad Prism (GraphPad software, San Diego, CA, USA). Significance was indicated by: *P < 0.05, **P < 0.01, ***P < 0.001, and **** P < 0.0001.
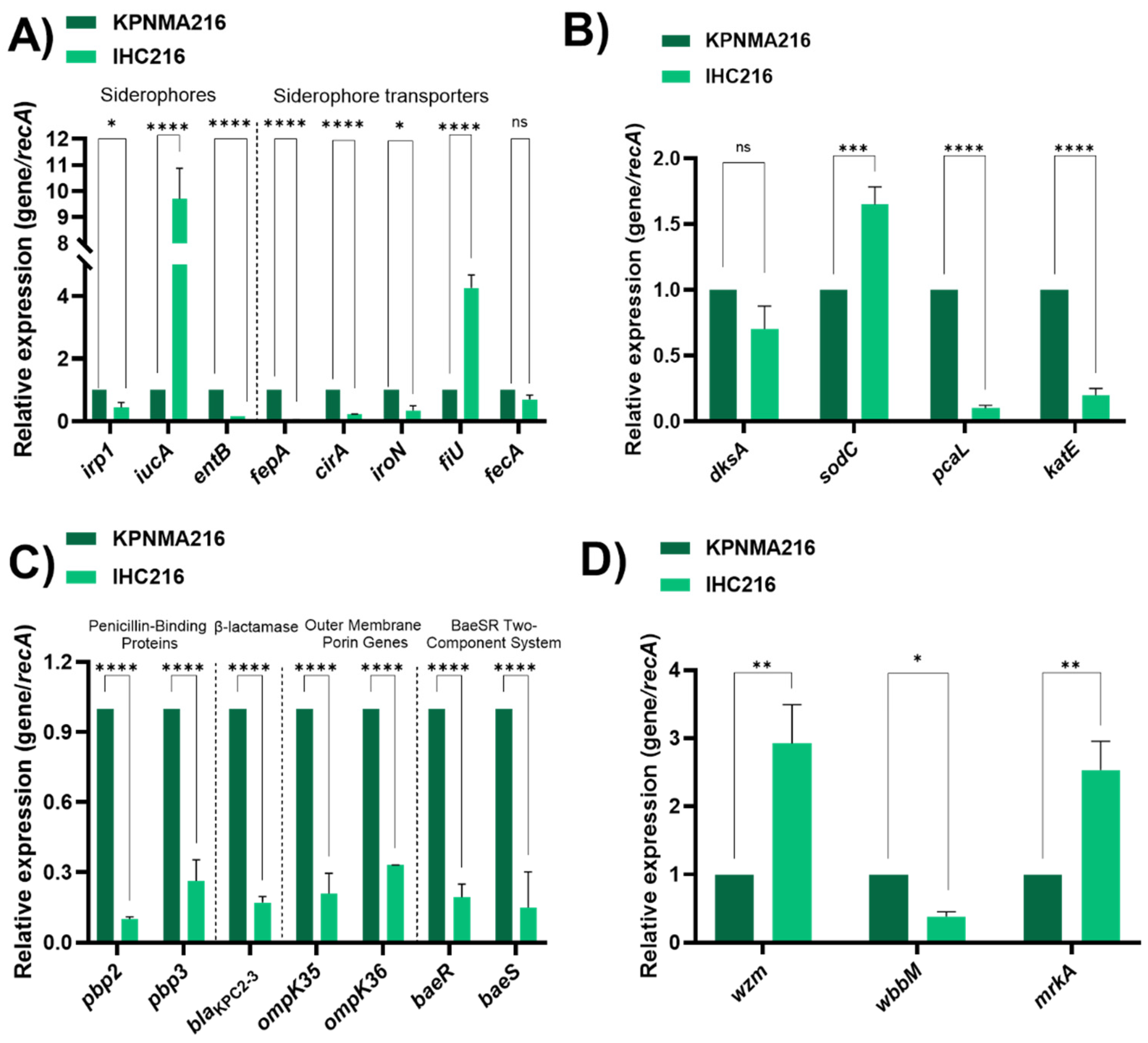
Figure 3.
(A) Capsule density in KPC216 and IHC216 strains. (B) Biofilm formation in tubes quantified by crystal violet in both strains. Statistical analysis was determined by t test (p < 0.05), using GraphPad Prism (GraphPad software, San Diego, CA, USA).
Figure 3.
(A) Capsule density in KPC216 and IHC216 strains. (B) Biofilm formation in tubes quantified by crystal violet in both strains. Statistical analysis was determined by t test (p < 0.05), using GraphPad Prism (GraphPad software, San Diego, CA, USA).
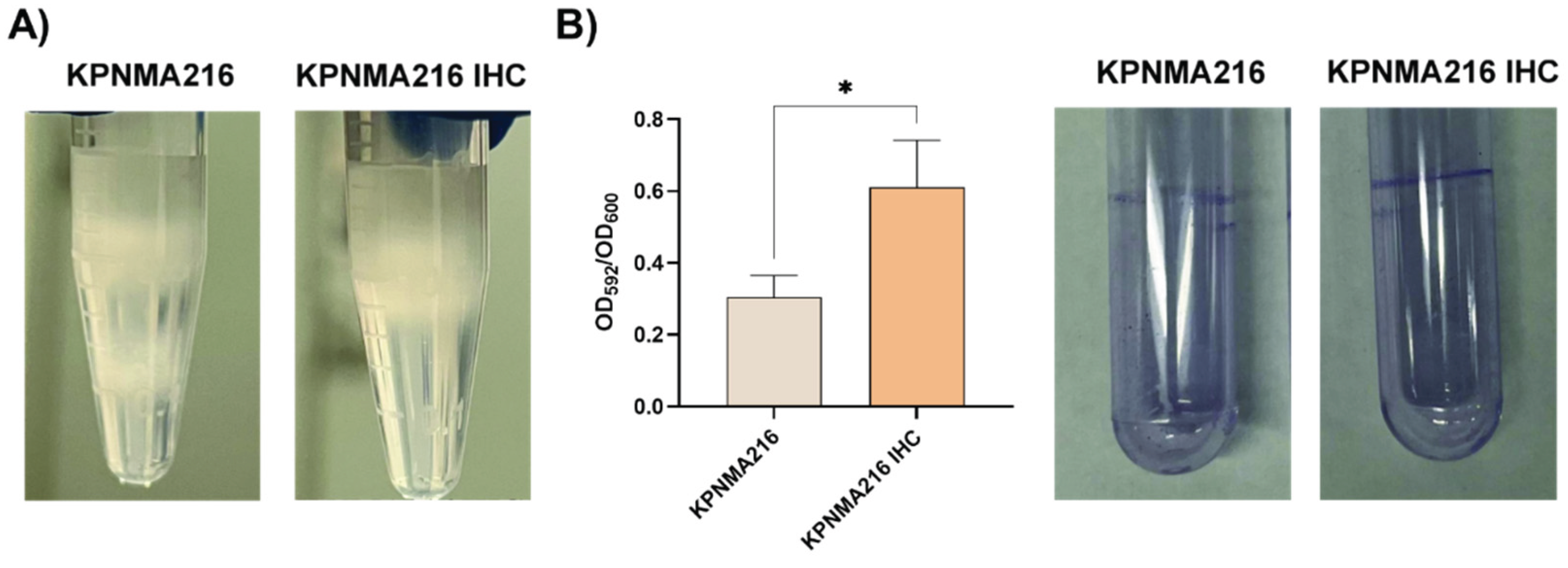

Table 1.
The minimum inhibitory concentration (MIC) for KPC216 and IHC216. The MIC was performed following manufacturer’s recommendations (Liofilchem S.r.l., Italy.
Table 1.
The minimum inhibitory concentration (MIC) for KPC216 and IHC216. The MIC was performed following manufacturer’s recommendations (Liofilchem S.r.l., Italy.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



