
Submitted:
07 July 2025
Posted:
08 July 2025
You are already at the latest version
Abstract
The feminization of the protandric teleost Centropomus undecimalis was evaluated under contrasting salinity conditions using 17β-estradiol (E2) administered orally to juveniles and via implants to adults. Two factorial experiments were designed to assess the hormonal effect on histological, hormonal, molecular, and zootechnical indicators. A 100% feminization rate was achieved in both life stages, confirmed by histological analysis and the expression of female sex- and reproduction-related genes (foxl2a, cyp19a1a, zar1l, ddx5, and H2A), particularly in marine environments. Hormonal assays showed increased plasma vitellogenin and decreased 11-ketotestosterone in treated groups, indicating suppression of male endocrine pathways. Zootechnical performance was not significantly affected in adults, but juveniles reared in freshwater exhibited better feed conversion and protein efficiency, likely due to lower osmoregulatory demands. These findings demonstrate that E2-induced feminization is highly effective in C. undecimalis across different salinities, with improved physiological outcomes in freshwater conditions. This study provides valuable insight into the reproductive manipulation of a commercially important species and supporting its sustainable aquaculture.
Keywords:
Centropomidae
; euryhaline fish
; reproduction
; androgens
; estrogens
; aquaculture
; gene expression
; protandric hermaphrodite
1. Introduction
Sex manipulation in fish represents a key biotechnological tool for optimizing aquaculture production, particularly in hermaphroditic species, where the induction of monosex populations allows for the maximization of productive, economic, and reproductive performance [1,2]. Induced feminization through hormonal treatments—typically using synthetic estrogens such as 17β-estradiol (E2)—has proven effective in numerous teleost species, altering the sex of individuals without compromising their viability [3,4]. This strategy not only addresses reproductive limitations and facilitates domestication under captive conditions but also offers an alternative to reduce pressure on vulnerable natural populations [5,6].
Centropomus undecimalis, commonly known as common snook, is a commercially important euryhaline species exhibiting sequential protandric hermaphroditism and inhabiting estuarine and coastal ecosystems in the Caribbean and other tropical and subtropical regions [7,8]. Although this reproductive strategy is advantageous in the wild, it poses a barrier to aquaculture development, especially when a supply of functional females is required for captive reproduction. In Colombia, the species is classified as vulnerable due to overfishing and habitat degradation [9].
Several studies have shown that E2 can successfully induce complete feminization in both juvenile and adult C. undecimalis without adverse effects on survival or growth [4,10,11]. This process is associated with the upregulation of key genes such as foxl2, cyp19a1a, zar1l, and ddx5, along with hormonal changes including increased vitellogenin (Vtg) and reduced 11-ketotestosterone (11-KT), indicating suppression of the male endocrine axis [12,13,14]. However, changes in environmental conditions (freshwater vs. marine) may interfere with the efficacy of feminization by affecting processes such as osmoregulation, steroidogenesis, and the absorption of exogenous hormones [15,16,17].
In euryhaline fish, the osmotic environment can directly influence gene expression and metabolic rates associated with sexual differentiation [15]. Studies on barramundi (Lates calcarifer) and other Centropomus species have shown that saline environments promote a faster response to hormonal treatments, likely due to more efficient E2 uptake and energy redistribution toward vitellogenesis [18,19,20].
Despite these advances, the combined effect of salinity and hormonal treatment on feminization and reproductive physiology in C. undecimalis remains poorly explored, representing a critical gap in the reproductive biotechnology of this species. This study aims to evaluate the synergistic potential of E2 and culture environment (freshwater vs. seawater) on the feminization of wild juveniles and adults of C. undecimalis, using histological, molecular, hormonal, somatic, and zootechnical indicators. The findings will support the optimization of rearing protocols for this high-value hermaphroditic species, with implications for the sustainability and reproductive efficiency of aquaculture systems.
2. Materials and Methods
To evaluate the reproductive physiology and the synergistic effect of 17β-estradiol (E2) on the feminization of Centropomus undecimalis, two experiments were conducted, distinguished by the route of hormone administration: enriched feed for juveniles and intramuscular implants for adults.
2.1. Study Area and Specimen Collection
The study was conducted at the Aquaculture Station and the Aquaculture and Fisheries Development Center of the University of Magdalena in Taganga (Santa Marta, Colombia). A total of 300 juveniles of C. undecimalis were collected from the Ciénaga Grande of Santa Marta, Colombian Caribbean, using cast nets with 8–15 mm mesh size. Fish were transported in capture-site water (8 ppt salinity) with supplemental oxygen (240 mg/L), following standardized transport protocols [21].
2.2. Acclimation and Feeding
Fish received prophylactic salt baths and were acclimated in 2000 L plastic tanks. Salinity was gradually adjusted: for freshwater conditions, it was reduced by 5 ppt/day to 0 ppt; for marine conditions, it was increased to 35 ppt. During adaptation, fish were fed live prey (Oreochromis spp.) and then transitioned to a formulated diet comprising fishmeal (Opisthonema oglinum), shrimp meal, commercial feed with 45% crude protein (CP), and vitamin supplements. The final feed composition was 55.7% CP, 16.2% moisture, 9.4% fat, and 13.8% ash, verified by NIRS™ DA1650.
2.3. Experimental Design
The experimental design was performed using a linear mixed-effects model (LMM), which was appropriate for the 2 × 2 × 3 factorial design employed. Fixed effects included hormonal treatment or H (with E2 and control), culture environment or E (marine and freshwater), and sampling time or T (at three distinct sampling times). The experimental unit was defined as the tank (n = 12), as treatments were applied at the group level. Each combination of hormonal treatment and environment included three independent tank replicates, and within each tank, two fish were sacrificed per time point as subsamples. To account for the hierarchical structure of the experiment and to avoid pseudoreplication, tank was included as a random effect. The fitted model for the dependent variable (DVar) was as follows:
DVar₍ᵢⱼₖₗ₎ = μ + Hᵢ + Eⱼ + Tₖ + (H × E)ᵢⱼ + (H × T)ᵢₖ + (E× T)ⱼₖ + (H × E × T)ᵢⱼₖ + (1 | Tankₗ) + εᵢⱼₖₗ
Where:
- μ is the overall mean,
- Hᵢ, Eⱼ, and Tₖ represent the fixed effects of hormone, environment, and time, respectively,
- (1 | Tankₗ) is the random effect of the tank (nested within the H × E combination),
- εᵢⱼₖₗ is the residual error term associated with within-tank variability.
Plasma hormonal analyses were conducted evaluating 11-ketotestosterone (11-KT) and vitellogenin (Vtg) concentrations as dependent variables. Experiment 1 involved juveniles and Experiment 2 included adults. Tissue, molecular, and somatic samples were collected from sacrificed individuals, maintaining equal sample sizes across time points. All fish were tagged with a PIT tag (Avid®) for individual identification. Zootechnical parameters were evaluated as dependent variables at the end of each experiment using a one-way ANOVA.
2.4. Experiment 1: Juveniles
Juveniles (13.47 ± 0.14 cm; 18.09 ± 0.60 g) were assigned to four treatments: T1 (E2 feed, seawater), T2 (E2 feed, freshwater), T3 (non-E2 feed, seawater), and T4 (non-E2 feed, freshwater), each with three replicates (n = 3), with 25 fish per 2000 L tank. Freshwater was treated with carbon and sand filters, while seawater underwent sequential filtration (25–1 µm), UV treatment, and supplemental oxygen. Feminization was induced by feeding a diet containing 100 mg·kg⁻¹ of E2 (55.7% CP), provided twice daily at 5% biomass for 45 days. Control groups received the same feed without hormones. Fish were cultured for 365 days post-treatment, with monthly biometric evaluations. Feed preparation followed the method described by Carvalho et al. [3].
2.5. Experiment 2: Adults
Adults (40.56 ± 5.2 cm; 582.65 ± 74.80 g) underwent the same four treatments, with hormones delivered via intramuscular implants: T1 (E2 implant, seawater), T2 (E2 implant, freshwater), T3 (no implant, seawater), T4 (no implant, freshwater). Each treatment had two replicates (n = 2), with 10 fish per 25 m³ tank. Implants were prepared following modified protocols for snook [22], blending 475 mg of cholesterol (95%) and 25 mg of cellulose (5%) with 30 mg of E2 in 80% ethanol. After drying (35–40 °C), they were combined with coconut oil (13 mg), molded (2 mm × 7 mm), and refrigerated at 5 °C. A dose of 2 mg E2 was administered per fish. Fish were fed once daily at a rate of 3% biomass for 180 days. Filtration and monitoring systems matched those used in juveniles. Six fish were sampled per treatment at each time point.
2.6. Gonadal Histological Evaluation
Sampling occurred at day 0 and on post-treatment days 46, 180, and 365 (Experiment 1); and on days 46, 90, and 180 (Experiment 2). Six fish were randomly selected per treatment from juveniles and adults. Fish were euthanized with 400 mg·L⁻¹ eugenol per AVMA guidelines [23]. Gonads were fixed in 4% buffered formalin, paraffin-embedded, sectioned (7 μm), and stained with hematoxylin-eosin [24]. Gonadal stages were classified per Brown-Peterson et al. [25] for males and Young et al. [26] for females.
2.7. Plasma 11-KT and Vtg Concentrations
11-KT and Vtg concentrations were measured in the same individuals used for histological and molecular analysis. Blood (0.5 mL) was drawn via caudal puncture using heparinized syringes. Plasma was obtained by centrifugation (15,000 rpm, 15 min, 4 °C), stored at –80 °C, and analyzed using competitive ELISA kits (MBS7253710 for 11-KT and MBS8807715 for Vtg; MyBioSource, USA). Absorbance was read at 450 nm using a Varioskan™ LUX reader; accuracy was confirmed through spike recovery and parallelism [27].
2.8. Gene Expression Analysis in Juveniles
At 180 days post-treatment in Experiment 1, six gonadal samples per treatment were collected, preserved in RNAlater (–80 °C), and processed by Novogene (USA). Total RNA was extracted with TRIzol® (Invitrogen), and libraries prepared using TruSeq Stranded mRNA (Illumina). Quality was verified via agarose gels and Bioanalyzer 2100. Libraries were sequenced (Illumina NovaSeq 6000, 150 bp paired-end), filtered with Fastp [28] (Q < 30), and aligned to the Lates calcarifer reference genome using HISAT2 [29]. Gene expression was quantified with featureCounts [30]. StringTie v1.3.3b [31] was used for transcript assembly, enabling discovery of novel genes, new exons, and accurate transcript annotation. Differential expression was analyzed with DESeq2 [32] (log2FC ≥ 1, padj ≤ 0.05), with enrichment assessed using clusterProfiler [33] and the GO/KEGG databases.
2.9. Zootechnical Performance and Water Quality
Parameters such as absolute growth rate (AGR), specific growth rate (SGR), absolute length gain (ALG), feed conversion ratio (FCR), feed efficiency ratio (FER) and protein efficiency ratio (PER) were estimated as follows [34]:
AGR (g/day) = [Wf (g) – Wo (g)] / t (day)
SGR (%/day) = [(Ln Wo − Ln Wf)/t] × 100
Where Wo and Wf represent initial body weight and final body weight, and t is the duration of the experiment (days).
ALG = [Ltf (cm) – Lto (cm)] / t (day)
Where Lto and Ltf represent initial total length and final, and t is the duration of the experiment (days).
FCR = Feed intake (g)/biomass gain (g) x 100
FER = Wet weight gain (g) /dry feed fed (g)
PER = Wet weight gain (g) /dry protein fed (g)
With the weights of the gonads and liver, the visceral fat indices (VFI), the hepatosomatic index (HSI) and the gonadosomatic index (GSI) were estimated using the following equations [34,35].
VFI (%) = [(visceral fat weight (g)/body weight (g)] x 100
HSI (%) = [(liver weight (g)/body weight (g)] x 100
GSI (%) = [(gonad weight (g)/body weight (g)] x 100
2.10. Statistical Analysis
All statistical analyses were performed using R software (R Core Team, 2024). Prior to conducting inferential tests, assumptions of normality and homogeneity of variances were assessed using the Shapiro–Wilk and Levene’s tests, respectively. These diagnostics were applied to the residuals of each model using the functions shapiro.test(residuals(model)) and leveneTest(Dvar ~ group, data = data), where Dvar represents the dependent variable and group the independent factor under evaluation.
Given that the assumptions of normality and homoscedasticity were satisfied, a linear mixed-effects model (LMM) was used to evaluate the effects of hormonal treatment, environment, and sampling time, accounting for the hierarchical structure of the data. Tanks were specified as a random effect to reflect the experimental design. The model was fitted using the lmer() function from the lme4 package as follows:
model <- lmer(Dvar ~ Hormone * Environment * Time + (1 | Tank), data = data)
For zootechnical parameters measured at the conclusion of each trial, a one-way analysis of variance (ANOVA) was conducted using the aov() function, followed by Tukey’s Honest Significant Difference (HSD) test for post hoc pairwise comparisons using the TukeyHSD() function. The R syntax applied was:
model <- aov(Dvar ~ Group, data = data)
TukeyHSD(model)
3. Results
3.1. Experiment 1: Juveniles
3.1.1. Histological Evaluation of Gonadal Development
On day 0 (before the experiment began) and at 46 days, all treatments showed 100% male gonadal development. However, at 180 and 365 days, treatment T1 showed complete feminization, with 100% females, indicating the effectiveness of the hormonal treatment applied to this group. On day 180, in treatment T2, 83% of females and 17% of individuals at the transitional stage were recorded. By day 365, 100% of females had achieved feminization in this treatment.
In contrast, no signs of sex change were observed in treatments T3 and T4 during the experimental period in either environment. All males analyzed exhibited small, translucent, filiform testes with the presence of primary spermatogonia (Sg1) but no visible lumen in the lobules (Figure 1A). In transitional individuals, intersex testes of reduced size were observed, containing both Sg1 and scattered primary oocytes, with a mean diameter of 5.09 ± 1.02 µm (Figure 1B).
Females from treatments T1 and T2 were in the chromatin-nucleolar stage, with primary growing oocytes (PO), well-organized ovarian lamellae, and thin ovarian walls. The mean diameter of oocyte was 13.8 ± 2.88 µm and 12.7 ± 1.41 µm for treatments T1 and T2, respectively (Figure 1C). At the end of the experiment, both macroscopic observations and histological evaluations of gonads in fish treated with estradiol confirmed the effectiveness of the hormonal treatment in both environments, with a final proportion of 100% females in groups T1 and T2.
3.1.2. Gene Expression Associated with Sexual Processes and Gonadal Development in Feminized Juveniles of Centropomus undecimalis
Transcriptomic analysis revealed significant differential gene expression between treatments and environments. In the marine environment, the comparison between T1 (E2) and T3 (control) identified 1,947 differentially expressed genes (DEGs), with 861 upregulated and 1,086 downregulated genes (Figure 2A). In contrast, in the freshwater environment, the comparison between T2 (E2) and T4 (control) revealed only 136 differentially expressed genes (DEGs), with 126 upregulated and 10 downregulated genes (Figure 2B).
Among the DEGs in T1 vs T3 (marine), four key genes related to female sex determination were identified: zar1l, foxl2a, ddx5, and H2A. In T2 vs T4 (freshwater), two genes were associated with female gonadal development: ddx5 and H2A. Notably, ddx5 and H2A were shared between both environments, suggesting a consistent role in the feminization process induced by E2 (Table 3).
Subsequent complementary analyses sought the presence of key genes in the transcriptome. In both environments and E2-treated groups, the genes Cyp19a1, GADPH, and foxl2 were expressed, with transcript sizes ranging from 507 to 686 base pairs. These sequences were specific to Centropomus undecimalis, although no sequences for this species are currently listed in public databases (GenBank or BOLD). Therefore, similar percentages were not established, although BLAST results indicated 98% identity for each gene.
3.1.3. Plasma 11-KT and VTG Levels in Juvenile Centropomus undecimalis
In juvenile C. undecimalis, plasma vitellogenin (Vtg) levels were significantly influenced by hormonal treatment (F₁,₈ = 239.23, p < 0.001) and sampling time (F₂,₅₃ = 9.57, p < 0.001), with a significant treatment × time interaction (F₂,₅₃ = 4.53, p = 0.015). The environment had a marginal effect on Vtg levels (F₁,₈ = 4.44, p = 0.068), whereas no significant interactions were observed for environment × treatment, environment × time, or the three-way interaction. For 11-ketotestosterone (11-KT), the hormonal treatment (F₁,₈ = 156.13, p < 0.001), time (F₂,₅₂ = 9.08, p < 0.001), and treatment × time interaction (F₂,₅₂ = 12.98, p < 0.001) were highly significant. No significant effects were observed for environment or any of its interactions. The inclusion of tank as a random effect effectively accounted for variability among replicate units and avoided pseudoreplication due to subsampling structure.
At 46 days post-feminization, the highest plasma vitellogenin (Vtg) concentrations were recorded in treatments T1 (marine E2) and T2 (freshwater E2), with peaks of 1.76 and 1.91 pg/mL, respectively. These levels decreased slightly by 180 days (1.61 pg/mL in T1 and 1.82 pg/mL in T2), and at 365 days, the values were 1.48 pg/mL in T1 and 1.66 pg/mL in T2. In contrast, hormone-free treatments (T3 and T4) showed significantly lower Vtg concentrations, at 0.59 and 0.55 pg/mL, respectively (p < 0.05) (Figure 3A). In contrast, 11-KT levels showed the opposite trend. At 365 days, E2-treated groups (T1 and T2) had concentrations of 0.33 and 0.28 pg/mL, respectively, while the highest values were recorded in the control groups: 1.33 pg/mL in T3 and 1.04 pg/mL in T4 (Figure 3B).
3.1.4. Visceral Fat, Gonadosomatic, and Hepatosomatic Indices in Juvenile Centropomus undecimalis
In juvenile C. undecimalis, all three somatic indices exhibited significant temporal variation. The gonadosomatic index (GSI) showed significant main effects for hormonal treatment (F₁,₈ = 8.26, p = 0.021) and time (F₂,₅₂ = 113.43, p < 0.001), along with a significant environment × time interaction (F₂,₅₂ = 7.11, p = 0.0019), indicating that temporal changes in GSI differed between environments. For the hepatosomatic index (HSI), time had a strong effect (F₂,₅₂ = 56.89, p < 0.001), while treatment showed a trend toward significance (F₁,₈ = 4.60, p = 0.064), and significant interaction effects were observed for environment × time (F₂,₅₂ = 6.11, p = 0.0041) and treatment × time (F₂,₅₂ = 10.76, p < 0.001). The visceral fat index (VFI) was significantly affected only by time (F₂,₅₂ = 13.40, p < 0.001), with no significant main effects of treatment or environment, nor any significant interactions. These findings suggest that developmental timing plays a predominant role in modulating somatic energy allocation in juveniles, with hormone and environmental influences acting in a time-dependent manner.
The highest VFI value was recorded in T4 (freshwater control) at 46 days post-treatment (2.44%), decreasing steadily to 0.76% by day 365 (Figure 4A). The highest GSI value occurred in T2 (freshwater E2) at 46 days (0.69%), dropping to 0.08% by day 365 (Figure 4B). Similarly, HSI followed a decreasing pattern, with T2 also showing the highest initial value (4.04%), which declined to 0.72% by the end of the experiment (Figure 4C).
3.1.5. Growth Performance Indicators in Juvenile Centropomus undecimalis
Treatments carried out in freshwater showed superior zootechnical indicators compared to those in seawater. Specifically, significantly higher values were observed in specific growth rate (SGR), protein efficiency ratio (PER), feed efficiency ratio (FER), and a lower feed conversion ratio (FCR), indicating better feed utilization under freshwater conditions.
SGR was highest in treatments T2 (freshwater E2) and T4 (freshwater control), with values of 0.64 and 0.73, respectively. Similarly, FER reached 18.94 in T2 and 21.43 in T4, with significant differences between treatments (p < 0.05). PER was also higher in freshwater (0.32 in T2 and 0.37 in T4), while FCR was consistently lower in these treatments, reinforcing the improved feed efficiency in this environment. Absolute growth rate (AGR) and daily length gain (DLG) were highest in T4, with values of 0.52 g/day and 0.04 cm/day, respectively, highlighting this treatment as having the greatest growth. No significant differences in survival were observed among treatments or environments (Table 4).
3.2. Experiment 2: Adults
3.2.1. Histological Observation of Gonads in Centropomus undecimalis
At the beginning of the experiment (time 0), all the sampled fish corresponded to immature male individuals (100%). characterized by a low number of Sertoli cells in the connective tissue, the absence of cyst formation, and classified as stage I of development (Figure 5A and B). At 46 days post-treatment, fish implanted with 17β-estradiol (T1 and T2) exhibited 100% of individuals in sexual transition from male to female. Previtelogenic oocytes with multiple nucleoli (n), large nuclei (N), and basophilic cytoplasm (C) were observed, with mean diameters of 16.31 ± 0.41 µm in T1 and 14.24 ± 5.03 µm in T2 (Figure 5C and D).
In contrast, the control treatments (T3 and T4) showed no signs of sex change at 180 days. The gonads were classified as immature testes at stage II, characterized by the presence of connective tissue (CT), primary spermatocytes (PS), and secondary spermatocytes (SS) as well as light pink-colored testes (Figure 5E and F). Unlike by day 180, treatments T1 and T2 exhibited complete feminization, with 100% of individuals developing stage II ovaries. Oocytes displayed well-defined nuclei and cytoplasm. minimal connective tissue, and mean diameters of 41.01 ± 7.43 µm (T1) and 37.63 ± 2.15 µm (T2), confirming the effectiveness of the hormonal treatment (Figure 5G and H).
3.2.2. Plasma Levels of Vitellogenin (Vtg) and 11-Ketotestosterone (11-KT) in Adult Centropomus undecimalis
In adult specimens, Vtg concentrations were significantly affected by hormonal treatment (F₁,₈ = 10.97, p = 0.011), while neither the environment nor time had significant main effects. The treatment × time interaction approached significance (F₂,₅₂ = 3.13, p = 0.052), suggesting a potential temporal modulation of hormonal effects. For 11-KT, a significant main effect of time was detected (F₂,₅₂ = 5.35, p = 0.007), along with significant environment × time (F₂,₅₂ = 4.00, p = 0.024), treatment × time (F₂,₅₂ = 4.08, p = 0.023), and environment × treatment × time interactions (F₂,₅₂ = 3.52, p = 0.037). These findings indicate that the endocrine response in adults is modulated by complex interactions between hormonal treatment, environmental conditions, and exposure time. The tank-level random effect captured variation attributable to experimental units, maintaining the integrity of the hierarchical structure.
In treatment with 17β-estradiol implants (T2), plasma levels of vitellogenin (Vtg) progressively increased throughout the experiment. At 90 days, significant differences were observed between the hormonally treated groups (T1 and T2) and the control groups without implants (T3 and T4). This trend became more pronounced at 180 days, when both T1 and T2 showed significantly higher concentrations than the controls. Notably, T2 reached the highest value with 2.291 pg/mL. In contrast, treatments T3 and T4 showed the lowest Vtg levels at 180 days, with 0.839 and 0.912 pg/mL, respectively (Figure 6A).
Regarding 11-ketotestosterone (11-KT), significant differences were observed both across sampling times and among treatments. At 90 days, levels in T1 and T2 were lower compared to the control groups, while T4 showed a higher concentration (0.202 pg/mL) (Figure 6B). This trend continued and became more pronounced by 180 days, when T4 reached the highest concentration recorded during the experiment (0.603 pg/mL), indicating a hormonal response opposite to that induced by E2.
3.2.3. Visceral Fat, Gonadosomatic, and Hepatosomatic Indices in Adult Centropomus undecimalis
In adult specimens, somatic indices were primarily influenced by hormonal treatment and time. GSI was significantly affected by treatment (F₁,₈ = 7.28, p = 0.027) and time (F₁,₃₂ = 12.53, p = 0.0012), but no significant interactions were detected. HSI responded significantly to treatment (F₁,₈ = 9.30, p = 0.0158), whereas no other main effects or interactions were statistically significant. For VFI, a significant effect of time was observed (F₁,₃₂ = 7.03, p = 0.0124), with no significant contributions from treatment, environment, or their interactions. Collectively, these results indicate that in adults, hormonal treatment had a consistent effect on gonadal and hepatic indices, while VFI remained predominantly influenced by temporal dynamics, suggesting physiological adjustments in lipid metabolism associated with reproductive status over time.
In this experiment, the Visceral Fat Index (VFI) showed a decreasing trend over time. The highest value was recorded in treatment T4 (control in freshwater) at 2.445%, while the lowest was observed in T1 (E2 in seawater) at 0.800% by day 180 (Figure 7A). As for the Gonadosomatic Index (GSI), an increasing trend was evident in the E2 treatments (T1 and T2), with the highest value recorded in T1 at 180 days (0.396%). The lowest GSI value was recorded in T3 (control in seawater) at 46 days (0.110%) (Figure 7B). The Hepatosomatic Index (HSI), in turn, exhibited an overall decreasing pattern. The highest value was observed in T2 (E2 in freshwater) at 46 days (1.210%), while the lowest value was recorded in T4 (control in freshwater) at 0.320% (Figure 7C).
These results suggest that E2 implantation in adults induces an initial reduction in VFI and HSI, with higher values observed at the first sampling (46 days) and a decreasing trend by day 180. In contrast, the GSI increased over time, particularly in the E2-treated groups, indicating a progressive stimulation of female gonadal development induced by exogenous estrogen.
3.2.4. Growth Performance Indicators in Adult Centropomus undecimalis
In adult fish implanted with 17β-estradiol (E2), no significant differences were observed between environments (marine vs. freshwater) or between hormonal treatments. Regarding growth, the highest values for absolute growth rate (AGR) and specific growth rate (SGR) were recorded in T3 (control in seawater), at 1.186 g/day and 0.193 %/day, respectively. On the other hand, treatment T1 showed the best performance in terms of feed efficiency ratio (FER: 0.342) and protein efficiency ratio (PER: 0.624), along with a low feed conversion ratio (FCR: 3.100) (Table 6).
4. Discussion
Induced feminization of Centropomus undecimalis through the administration of 17β-estradiol (E2) proved highly effective in both juvenile and adult stages [10,26]. The efficacy of the treatment was evidenced by histological analyses and supported by well-established hormonal mechanisms described for protandric teleosts [38]. The ability to reverse the male phenotype into functional ovaries is consistent with findings in other species subjected to similar hormonal protocols, e.g., Oreochromis niloticus [39] and C. viridis [11]. In this study, a 100% feminization rate was achieved in both juveniles fed hormone-enriched diets and adults receiving implants, surpassing the variable results reported by Vidal-López et al. [10], who observed age- and protocol-dependent outcomes.
In adults, intramuscular implantation proved to an effective tool for inducing gonadal change, with functional oocytes observed at 180 days post-treatment. This result is particularly relevant since many previous studies have focused on the juvenile stage as the critical window for sexual differentiation [26,38,40]. The current results reinforce the idea that gonadal tissue in common snook retains plasticity beyond the juvenile phase, as suggested in studies on Lates calcarifer, where adults underwent sex reversal after prolonged treatments [18]. C. poeyi [41] and C. undecimalis [42] where adults underwent sex reversal after prolonged treatments.
At the molecular level, transcriptomic analyses at 180 days post-treatment in juveniles revealed a high number of differentially expressed genes (DEGs), especially under marine conditions. Over 1900 DEGs were identified between the E2-treated and control groups, including key female sex-determining genes such as foxl2a, cyp19a1a, zar1l, ddx5, and H2A. The upregulation of these genes has been recognized as a reliable molecular marker of ovarian commitment in fish [12,13,14,43]. In previous studies with barramundi, foxl2 and cyp19a1a were also upregulated in response to estrogen treatments, acting as essential regulators of androgen aromatization and gonadal epithelium differentiation toward a female phenotype [18]. The genes ddx5 and H2A were commonly expressed in both environments, suggesting their role in stabilizing the feminization process regardless of osmotic conditions. In euryhaline fish such as C. undecimalis, the environment can modulate reproductive gene expression, influencing processes like vitellogenesis and hypothalamic-pituitary-gonadal axis plasticity [44]. This effect was confirmed by the greater number of DEGs observed in marine fish, potentially due to heightened estrogen sensitivity or stronger activation of reproductive metabolism under seawater conditions, as previously documented in Dicentrarchus labrax and Sparus aurata [45,46].
Likewise, plasma vitellogenin (Vtg) levels confirmed the estrogenic activity induced by E2, with both juvenile and adult treatments showing significant increases compared to controls. This finding aligns with multiple studies that recognize Vtg as a sensitive and specific biomarker of estrogen exposure in fish [47,48]. In juveniles, Vtg levels peaked within the first 180 days and then stabilized, while in adults, values rose progressively, reaching maximum levels at day 180. This temporal difference may be attributed to the administration route (oral for juveniles, implant for adults), which strongly influences the hormone's action kinetics and its effectiveness in triggering specific physiological processes in gonadal tissue [49,50].
Conversely, plasma 11-ketotestosterone (11-KT) concentrations—the dominant androgen in teleosts—were consistently lower in E2-treated fish of both life stages. This reflects suppression of the male endocrine axis, likely due to negative feedback from estradiol on gonadotropin release [51]. The most pronounced suppression was observed in implanted groups, supporting the efficacy of sustained delivery in achieving prolonged endocrine inhibition of the male phenotype. Interestingly, E2 may also increase 11-KT and testosterone levels in C. undecimalis, suggesting the involvement of endocrine feedback mechanisms in regulating sexual differentiation [52]. These results highlight the importance of precise dose management to avoid adverse effects on hormonal homeostasis. Thus, sustained 11-KT suppression in E2-treated fish indicates disrupted spermatogenesis and a redirection of metabolism toward vitellogenesis. In this context, the 11-KT to E2 ratio may serve as a robust indicator of sexual status, providing an additional diagnostic tool to monitor the progress of feminization [53]. Nevertheless, hormonal response variability may be influenced by environmental factors such as temperature and photoperiod, which modulate gene expression linked to gonadal differentiation [54]. These findings corroborate E2's capacity to modulate the hypothalamic–pituitary–gonadal axis, induce differential expression of genes such as cyp19a, and promote ovarian development [13].
E2 administration in C. undecimalis triggered differential physiological responses in juveniles and adults, as reflected in somatic indices. In juveniles, the initial increase in gonadosomatic index (GSI) and hepatosomatic index (HSI), followed by a decline in HSI, suggests a transient hormonal effect consistent with the activation of hepatic metabolism for Vtg synthesis [11]. In adults, increased GSI and a slight reduction in late HSI reduction suggest a prolonged metabolic adjustment associated with E2 implantation. This dynamic aligns with studies indicating E2's role in gonadal development stimulation [3], while others warn that high HSI may indicate metabolic overload from Vtg synthesis in protandric fish [55]. In contrast, the visceral fat index (VFI) did not vary significantly, suggesting that E2 does not directly affect lipid energy reserves [56].
Growth parameters showed specific effects: a temporary growth rate reduction during treatment was observed, but no significant differences emerged between E2-treated fish and controls by the end of the experiment. These findings are consistent with studies on Centropomus species, which report neutral growth effects after E2 exposure [10]. In freshwater conditions, juveniles exhibited better zootechnical performance (SGR, FER, PER, FCR) in treatments T2 and T4, consistent with authors who found improved growth under less osmotically demanding environments [57,58].
The better feed conversion ratio (FCR) observed in freshwater-reared juveniles supports the hypothesis that osmoregulation requires less energy under low salinity. In C. undecimalis, osmotic pressure decreases at salinities below 25 ppt [59], resulting in reduced metabolic costs in freshwater systems [16]. In adults, no significant differences in growth parameters were observed, reflecting stage-specific physiological responses to treatment [17].
In light of these results, we conclude that feminization induced with 17β-estradiol in C. undecimalis is an effective strategy for both juveniles and adults under contrasting salinity conditions. Histological, molecular, endocrine, and physiological evidence confirms that E2 promotes female sexual differentiation and functional gonadal development without significantly compromising productive traits. This finding is especially relevant for designing monosex breeding programs in snook, a species of high commercial and biological value.
Future research should focus on a deeper transcriptomic and epigenetic characterization of sex-reversal mechanisms as well as the development of gradual dosage protocols and encapsulated formulations for controlled, localized E2 release. Additionally, sustainable reproductive alternatives—such as bioactive plant extracts or gene-editing techniques targeting key endocrine pathways—should be comparatively assessed. Lastly, efforts should be made to establish a regulatory framework that ensures the safe application of sex-control technologies in aquaculture, striking a balance between productivity with environmental and health responsibility.
5. Conclusions
This study provides robust evidence that feminization of Centropomus undecimalis through 17β-estradiol (E2) is a highly effective strategy in both juvenile and adult stages across contrasting salinity conditions. The use of histological, hormonal, molecular, and physiological indicators allowed for a comprehensive assessment of sex reversal, revealing a consistent upregulation of ovarian markers and suppression of androgenic pathways in treated fish. The linear mixed-effects model confirmed significant main effects and interactions for treatment, environment, and time, particularly influencing Vtg and 11-KT levels and somatic indices such as GSI and HSI. These findings underscore the biological plasticity of this protandric species and the feasibility of sex control protocols tailored to life stage and environmental context. Moreover, the results support the development of monosex broodstock as a biotechnological tool for enhancing reproductive efficiency and promoting
6. Patents
This section is not mandatory but may be added if there are patents resulting from the work reported in this manuscript.
Author Contributions
Conceptualization, S.G.-I., W.C.-S. and N.V.-V.; methodology, S.G.-I. and S.C.-B.; investigation, S.G.-I. and S.C.-B.; supervision, W.C.-S. and N.V.-V.; data curation and writing—original draft preparation, S.G.-I.; writing—review and editing, W.C.-S., N.V.-V. and V.A.-G. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Sistema General de Regalías (SGR), project “Implementation of productive systems in marine snook aquaculture to promote its production in the department of Magdalena.” SG-I acknowledges the “Excellence Doctoral Scholarship Program of the Bicentennial I (MinCiencias)” for supporting her doctoral studies in Marine Sciences.
Institutional Review Board Statement
This study is approved by the Institutional Ethics Committee for Research (CEI) of the Universidad del Magdalena (date: 23 May 2019) and the National Authority of Environmental Licenses (ANLA) (code: Res.001264; date: 26 June 2024).
Data Availability Statement
The data supporting the findings of this study are available upon request from the corresponding author.
Acknowledgments
The authors thank the entire technical and scientific team of the Aquaculture Hangar project from the Research Group on Biodiversity and Applied Ecology – GIBEA at the University of Magdalena.
Conflicts of Interest
The authors declare no conflict of interest related to this manuscript.
References
- Piferrer, F. Endocrine sex control strategies for the feminization of teleost fish. Aquaculture 2001, 197, 229–281. [Google Scholar] [CrossRef]
- Contreras-García, M.; Contreras-Sánchez, W.M.; Mendoza-Carranza, M.; Mcdonal-Vera, A.; Cruz-Rosado, L. Induced Sex Reversal in Adult Males of the Protandric Hermaphrodite Centropomus undecimalis Using 17 β-Estradiol. Aquac. J. 2023, 3, 196–208. [Google Scholar] [CrossRef]
- Carvalho, C.; Passini, G.; Melo-Costa, W.; Nunes-Vieira, B.; Ronzani-Cerqueira, V. Effect of estradiol-17β on the sex ratio, growth and survival of juvenile common snook (Centropomus undecimalis). Acta Sci. Anim. Sci. 2014, 36, 239–245. [Google Scholar] [CrossRef]
- Passini, G.; Sterzelecki, F.C.; De Carvalho, C.V.A.; Cerqueira, V.R. Induction of sex inversion in common snook (Centropomus undecimalis) males using 17-β estradiol implants. Aquac. Res. 2016, 47, 1090–1099. [Google Scholar] [CrossRef]
- García, C.B.; Duarte, L.O.; Altamar, J.; Manjarrés, L.M. Demersal fish density in the upwelling ecosystem off Colombia, Caribbean Sea: Historic outlook. Fish. Res. 2007, 85, 68–73. [Google Scholar] [CrossRef]
- Gaitán-Ibarra, S.; Villamizar-Villamizar, N. Piscicultura marina del róbalo (Centropomus undecimalis); Editorial Unimagdalena: Santa Marta, Colombia, 2024; 128p. [Google Scholar]
- Caballero-Chávez, J. Revisión de aspectos biológicos y pesqueros del róbalo (Centropomus undecimalis). Rev. Colomb. Cienc. Pecu. 2011, 24, 161–176. [Google Scholar]
- Navarro-Flores, J.; Ibarra-Castro, L.; Martinez-Brown, J.M.; Zavala-Leal, O.I. Hermaphroditism in teleost fishes and their implications in commercial aquaculture. Rev. Biol. Mar. Oceanogr. 2019, 54, 1–10. [Google Scholar] [CrossRef]
- Grijalba-Bendeck, M.; Leal-Flórez, J.; Bolaños-Cubillos, N.; Acero, A. Centropomus undecimalis. In Libro Rojo de Peces Marinos de Colombia; INVEMAR: Santa Marta, Colombia, 2017; pp. 224–227. [Google Scholar]
- Vidal-López JM, Álvarez-González CA, Sánchez WM, Patiño R, Hernández-Franyutti AA, Hernández-Vidal U, García RM. Feminización de juveniles del Robalo Blanco Centropomus undecimalis (Bloch 1792) usando 17β-estradiol. Revista Ciencias Marinas y Costeras. 2012, 4:83-93. https://www.redalyc.org/articulo.oa? 6337.
- Navarro-Flores, J.; Manuel, M.B.J.; Iram, Z.L.; Humberto, R.C.A.; Leonardo, I.C. Assessing the feasibility of exogenous 17β-estradiol for inducing sex change in white snook, C. viridis: From growth, resting and maturation studies. Aquac. Rep. 2023, 33, 101767. [Google Scholar] [CrossRef]
- Baron, D.; Cocquet, J.; Xia, X.; Fellous, M.; Guiguen, Y.; Veitia, R.A. An evolutionary and functional analysis of FoxL2 in rainbow trout gonad differentiation. J. Mol. Endocrinol. 2004, 33, 705–715. [Google Scholar] [CrossRef]
- Cui H, Zhu H, Ban W, Li Y, Chen R, Li L, Zhang X, Chen K, Xu H. Characterization of Two Gonadal Genes, zar1 and wt1b, in Hermaphroditic Fish Asian Seabass (Lates calcarifer). Animals 2024, 14, 508. [Google Scholar] [CrossRef]
- Sone, R.; Taimatsu, K.; Ohga, R.; et al. Critical roles of the ddx5 gene in zebrafish sex differentiation and oocyte maturation. Sci. Rep. 2020, 10, 14157. [Google Scholar] [CrossRef]
- Hernández-Vidal, U.; Chiappa-Carrara, X.; Contreras-Sánchez, W.M. Reproductive variability of the common snook, Centropomus undecimalis, in environments of contrasting salinities. Cienc. Mar. 2012, 40, 173–185. [Google Scholar] [CrossRef]
- Mylonas, C.C.; et al. Growth performance and osmoregulation in the shi drum (Umbrina cirrosa) adapted to different environmental salinities. Aquaculture 2009, 287, 203–210. [Google Scholar] [CrossRef]
- Logan, C.A.; Buckley, B.A. Transcriptomic responses to environmental temperature in eurythermal and stenothermal fishes. J. Exp. Biol. 2015, 218, 1915–1924. [Google Scholar] [CrossRef]
- Banh, Q.Q.T.; Domingos, J.A.; Pinto, R.C.C.; et al. Dietary 17β-estradiol and 17α-ethinyloestradiol alter gonadal morphology and gene expression in juvenile barramundi (Lates calcarifer). Aquac. Res. 2020, 51, 3415–3425. [Google Scholar] [CrossRef]
- Wu, N.; Yue, H.M.; Chen, B.; Gui, J.F. Histone H2A Has a Novel Variant in Fish Oocytes. Biol. Reprod. 2009, 81, 275–283. [Google Scholar] [CrossRef]
- Yan, H.; Shen, X.; Jiang, J.; et al. Gene expression of Takifugu rubripes gonads during AI-or MT-induced masculinization and E2-induced feminization. Endocrinology. 2021, 162, bqab068. [Google Scholar] [CrossRef]
- Polonía-Rivera, C.; Gaitán, S.; Chaparro-Muñoz, N.; Villamizar, N. Captura, transporte y aclimatación de juveniles y adultos de róbalo Centropomus undecimalis. Rev. Intropica. 2017, 12, 61–64. [Google Scholar] [CrossRef]
- Contreras-Sánchez, W.M. Manual para la producción de róbalo blanco: Centropomus undecimalis en cautiverio; UJAT: Villahermosa, México, 2012. [Google Scholar]
- AVMA guidelines for the euthanasia of animals. American Veterinary Medical Association. Recuperado el 3 de abril de 2024, de https://www.avma.
- Luna, H. Manual of Histologic Staining Methods of the Armed Forces Institute of Pathology; 3rd, *!!! REPLACE !!!* (Eds.) ; American Registry of Pathology: New York, NY, USA, 1968.
- Brown-Peterson, N.J.; Wyanski, D.M.; Saborido-Rey, F.; et al. A standardized terminology for describing reproductive development in fishes. Mar. Coast. Fish. 2011, 3, 52–70. [Google Scholar] [CrossRef]
- Young, J.; Yeiser, B.G.; Whittington, J.A.; Dutka-Gianelli, J. Maturation of female common snook Centropomus undecimalis: Implications for managing protandrous fishes. J. Fish Biol. 2020, 97, 1317–1331. [Google Scholar] [CrossRef]
- Scott, A.P.; Ellis, T. Measurement of fish steroids in water: A review. Gen. Comp. Endocrinol. 2007, 153, 392–400. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Kim, D.; et al. HISAT2: Graph-based alignment of next-generation sequencing reads. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef]
- Pertea M, Pertea GM, Antonescu CM, Chang TC, Mendell JT, Salzberg SL. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nature biotechnology. 2015, 33(3):290-5.
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Yu, G. , Wang, LG., Han, Y., He, QY. clusterProfiler: an R package for comparing biological themes among gene clusters. Omics: a journal of integrative biology. 2012, 1;16(5):284-7. [CrossRef]
- Zheng J, Zhang W, Dan Z, Zhuang Y, Liu Y, Mai K and Ai Q (2022), Replacement of dietary fish meal with Clostridium autoethanogenum meal on growth performance, intestinal amino acids transporters, protein metabolism and hepatic lipid metabolism of juvenile turbot (Scophthalmus maximus L.). Front. Physiol. 13:981750. [CrossRef]
- Sadekarpawar, S. , Parikh, P. ( 8(1), 110–118. [CrossRef]
- Sokal, RR. The principles and practice of statistics in biological research. Biometry. 1995, 451–554. [Google Scholar]
- Zar, JH. Dichotomous variables. Biostatistical Analysis 5th ed. Upper Saddle River, NJ: Prentice-Hall. 2010, 557-8.
- Baroiller, J.F.; Guiguen, Y.; Fostier, A. Endocrine and environmental aspects of sex differentiation in fish. Cell. Mol. Life Sci. 1999, 55, 910–931. [Google Scholar] [CrossRef]
- Pandian, T.J.; Sheela, S.G. Hormonal induction of sex reversal in fish. Aquaculture 1995, 138, 1–22. [Google Scholar] [CrossRef]
- Devlin, R.H.; Nagahama, Y. Sex determination and sex differentiation in fish: An overview of genetic, physiological, and environmental influences. Aquaculture 2002, 208, 191–364. [Google Scholar] [CrossRef]
- Vidal-López, J. M. , Contreras-Sánchez, W. M., Hernández-Franyutti, A., Contreras-García, M. D. J., & Uribe-Aranzábal, M. D. C. (2019). Functional feminization of the Mexican snook (Centropomus poeyi) using 17β-estradiol in the diet. Latin american journal of aquatic research, 47(2), 240-250.
- Contreras-García, M. J. , Contreras-Sánchez, W. M., Mendoza-Carranza, M., De La Cruz-Hernández, E. N., Mcdonal-Vera, A., & Hernández-Vidal, U. (2025). Gene expression, gonadal morphology, and steroid hormone profiles during induced sex-change in the protandric hermaphrodite Centropomus undecimalis. Aquaculture, 604, 742429.
- Vizziano-Cantonnet, D.; Baron, D.; Randuineau, G.; Guiguen, Y. Characterization of early molecular sex differentiation in rainbow trout, Oncorhynchus mykiss. Dev. Dyn. 2007, 236, 2198–2206. [Google Scholar] [CrossRef]
- Mylonas, C.C.; Fostier, A.; Zanuy, S. Broodstock management and hormonal manipulations of fish reproduction. Gen. Comp. Endocrinol. 2010, 165, 516–534. [Google Scholar] [CrossRef]
- Blázquez, M.; Zanuy, S.; Carillo, M.; Piferrer, F. Effects of rearing temperature on the sex differentiation of the European sea bass (Dicentrarchus labrax L.). J. Exp. Zool. 1998, 281, 207–216. [Google Scholar] [CrossRef]
- Giménez, I.; Estévez, A.; Zanuy, S.; et al. Environmental sex determination in the sea bream (Sparus aurata L.): Relationship between growth and sex. Aquaculture 2007, 263, 1–4. [Google Scholar] [CrossRef]
- Tyler, C.R.; van der Eerden, B.; Jobling, S.; Panter, G.; Sumpter, J.P. Measurement of vitellogenin in male fathead minnows (Pimephales promelas) as a biomarker for estrogenic exposure. Environ. Toxicol. Chem. 1999, 18, 2291–2296. [Google Scholar] [CrossRef]
- Arukwe, A.; Goksøyr, A. Eggshell and egg yolk proteins in fish: Hepatic expression, synthetic control and use as biomarkers of estrogenic compounds. Mar. Environ. Res. 2003, 55, 331–334. [Google Scholar] [CrossRef]
- Santos, E.M.; Kille, P.; Tyler, C.R. Gene expression and molecular responses in fish: Integration with ecotoxicology and endocrine disruption. Gen. Comp. Endocrinol. 2007, 152, 1–4. [Google Scholar] [CrossRef]
- Hara, A.; Hiramatsu, N.; Fujita, T. Vitellogenesis and choriogenesis in fishes. Fish Physiol. Biochem. 2016, 42, 141–150. [Google Scholar] [CrossRef]
- Filby, A.L.; Thorpe, K.L.; Tyler, C.R. Multiple molecular effect pathways of an environmental oestrogen in fish. J. Mol. Endocrinol. 2006, 37, 121–134. [Google Scholar] [CrossRef] [PubMed]
- Passini, G.; Sterzelecki, F.C.; De Carvalho, C.V.A.; Baloi, M.F.; Naide, V.; Cerqueira, V.R. 17α-Methyltestosterone implants accelerate spermatogenesis in common snook, Centropomus undecimalis, during first sexual maturation. Theriogenology 2018, 106, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Kohn, Y.Y.; Lokman, P.M.; Damsteegt, E.L.; Closs, G.P.; Young, G. Sex identification in captive hapuku (Polyprion oxygeneios) using plasma levels of vitellogenin and sex steroids. Aquaculture. [CrossRef]
- Nieto-Vera, M.T.; Rodríguez-Pulido, J.A.; Góngora-Orjuela, A. ¿Qué sabemos de los esteroides sexuales y las gonadotropinas en la reproducción de teleósteos neotropicales? Orinoquia 2020, 24, 52–63. [Google Scholar] [CrossRef]
- Young, J.; Yeiser, B.G.; Whittington, J.A.; Dutka-Gianelli, J. Maturation of female common snook Centropomus undecimalis: Implications for managing protandrous fishes. J. Fish Biol. 2020, 97, 1317–1331. [Google Scholar] [CrossRef]
- De Carvalho, C.V.A.; Passini, G.; De Melo Costa, W.; Cerqueira, V.R. Feminization and growth of juvenile fat snook Centropomus parallelus fed diets with different concentrations of estradiol-17β. Aquac. Int. 2014, 22, 1499–1512. [Google Scholar] [CrossRef]
- Alvarez-Lajonchère, L.; Hernández-Molejón, O.G. Producción de juveniles de peces estuarinos para un Centro en América Latina y el Caribe: diseño, operación y tecnologías. World Aquaculture Society, 2001. [Google Scholar]
- Álvarez-Lajonchère, L.; Tsuzuki, M.Y. A review of methods for Centropomus spp. (snooks) aquaculture and recommendations for the establishment of their culture in Latin America. Aquac. Res. 2008, 39, 684–700. [Google Scholar] [CrossRef]
- Gracia-López, V.; Rosas-Vázquez, C.; Brito-Pérez, R. Effects of salinity on physiological conditions in juvenile common snook Centropomus undecimalis. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2006, 145, 340–345. [Google Scholar] [CrossRef]
Figure 1.
Histological characterization of gonads in juvenile Centropomus undecimalis treated with 17β-estradiol. A) Immature testis (Stage I). B) Transitional gonad with intersex features. C) Ovary at chromatin-nucleolar stage (Stage I). Sg: primary spermatogonia, WT: testicular wall, CT: connective tissue, C: cysts, PO: primary growing oocytes, A: atretic oocyte, N: nucleus, OL: ovarian lamellae. Hematoxylin-eosin staining, 40X magnification.
Figure 1.
Histological characterization of gonads in juvenile Centropomus undecimalis treated with 17β-estradiol. A) Immature testis (Stage I). B) Transitional gonad with intersex features. C) Ovary at chromatin-nucleolar stage (Stage I). Sg: primary spermatogonia, WT: testicular wall, CT: connective tissue, C: cysts, PO: primary growing oocytes, A: atretic oocyte, N: nucleus, OL: ovarian lamellae. Hematoxylin-eosin staining, 40X magnification.
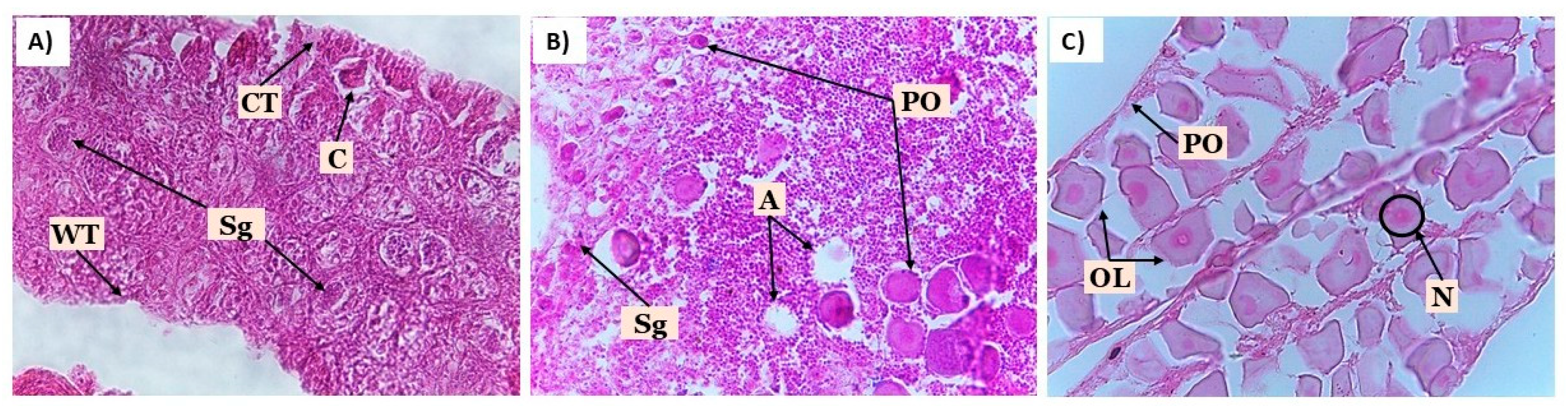
Figure 2.
Comparative analysis of differentially expressed genes (DEGs) in juvenile Centropomus undecimalis treated with 17β-estradiol (E2). Volcano plots for A) T1 vs T3 (marine environment) and B) T2 vs T4 (freshwater environment). The dashed line indicates the threshold for DEG selection criteria.
Figure 2.
Comparative analysis of differentially expressed genes (DEGs) in juvenile Centropomus undecimalis treated with 17β-estradiol (E2). Volcano plots for A) T1 vs T3 (marine environment) and B) T2 vs T4 (freshwater environment). The dashed line indicates the threshold for DEG selection criteria.
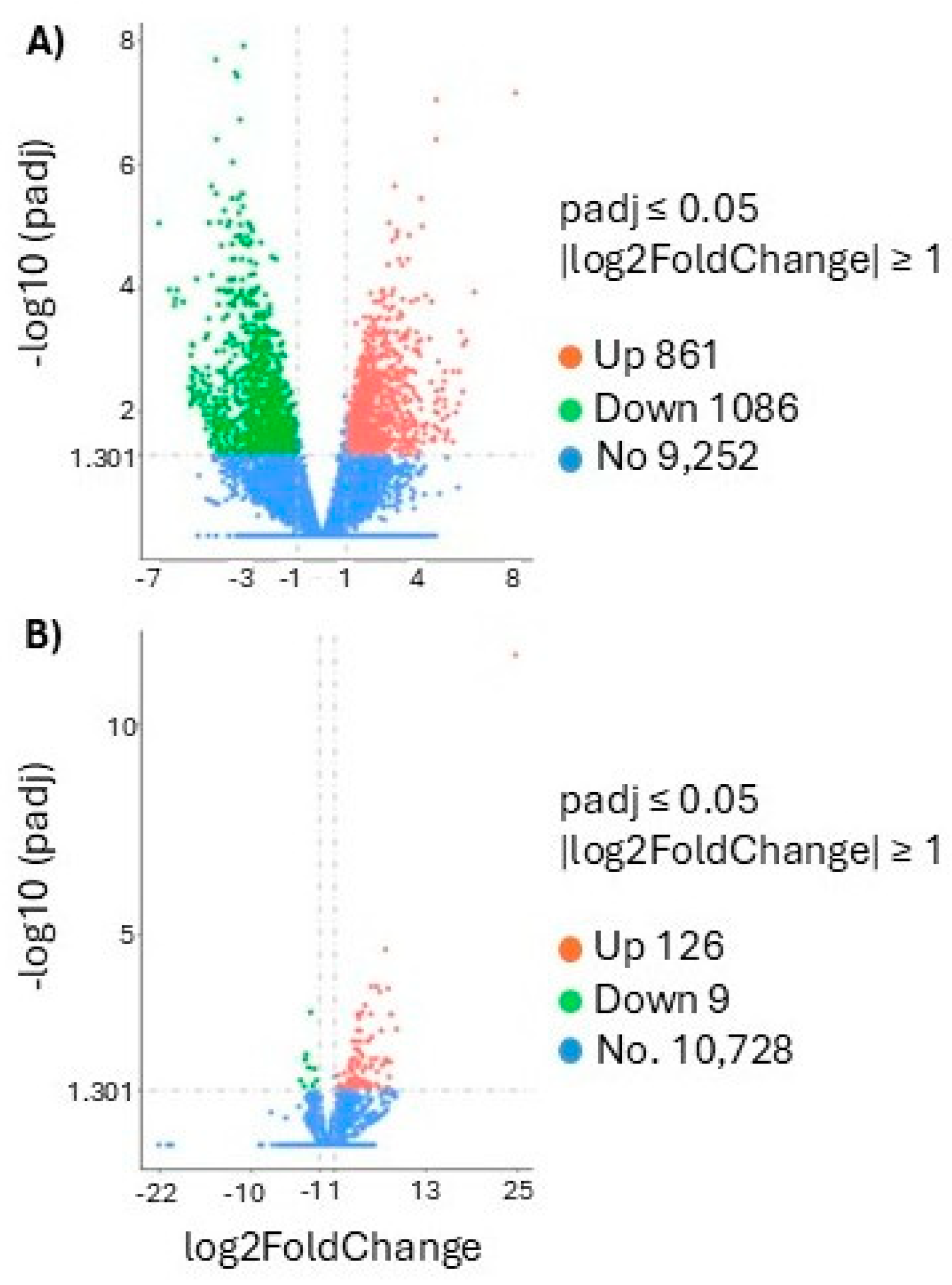
Figure 3.
Plasma concentrations of A) vitellogenin (Vtg) and B) 11-ketotestosterone (11-KT) in juvenile Centropomus undecimalis at 46, 180, and 365 days post-feminization under different treatments. Values are presented as means with 95% confidence intervals derived from Tukey’s HSD test (p < 0.05).
Figure 3.
Plasma concentrations of A) vitellogenin (Vtg) and B) 11-ketotestosterone (11-KT) in juvenile Centropomus undecimalis at 46, 180, and 365 days post-feminization under different treatments. Values are presented as means with 95% confidence intervals derived from Tukey’s HSD test (p < 0.05).
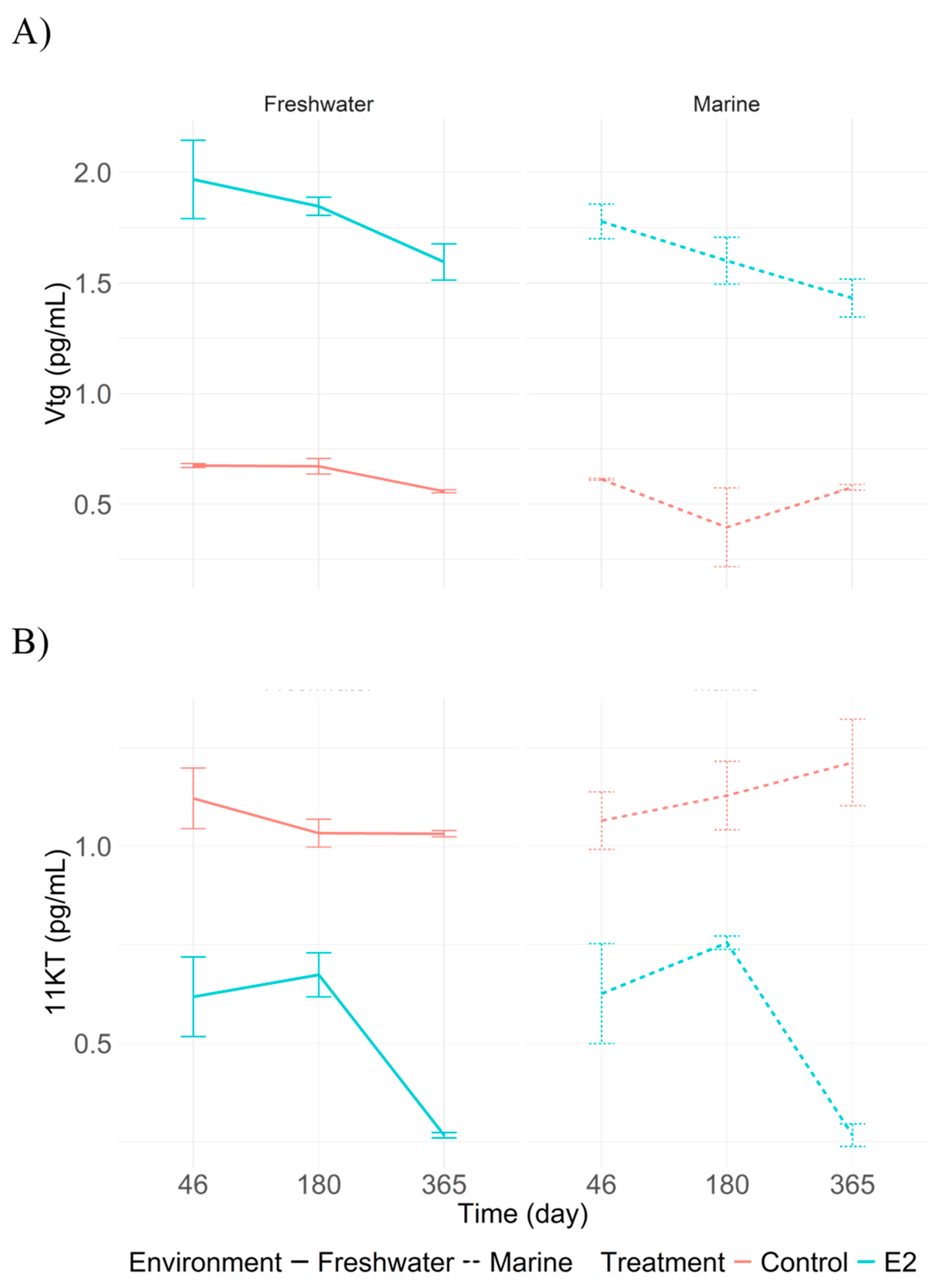
Figure 4.
Comparison of physiological indices in juvenile Centropomus undecimalis exposed to 17β-estradiol (E2) and control conditions in marine and freshwater environments. A) Visceral Fat Index (VFI); B) Gonadosomatic Index (GSI); C) Hepatosomatic Index (HSI). Values are shown as means with 95% confidence intervals. Differences between sampling times and treatments were evaluated using Tukey’s HSD test (p < 0.05).
Figure 4.
Comparison of physiological indices in juvenile Centropomus undecimalis exposed to 17β-estradiol (E2) and control conditions in marine and freshwater environments. A) Visceral Fat Index (VFI); B) Gonadosomatic Index (GSI); C) Hepatosomatic Index (HSI). Values are shown as means with 95% confidence intervals. Differences between sampling times and treatments were evaluated using Tukey’s HSD test (p < 0.05).
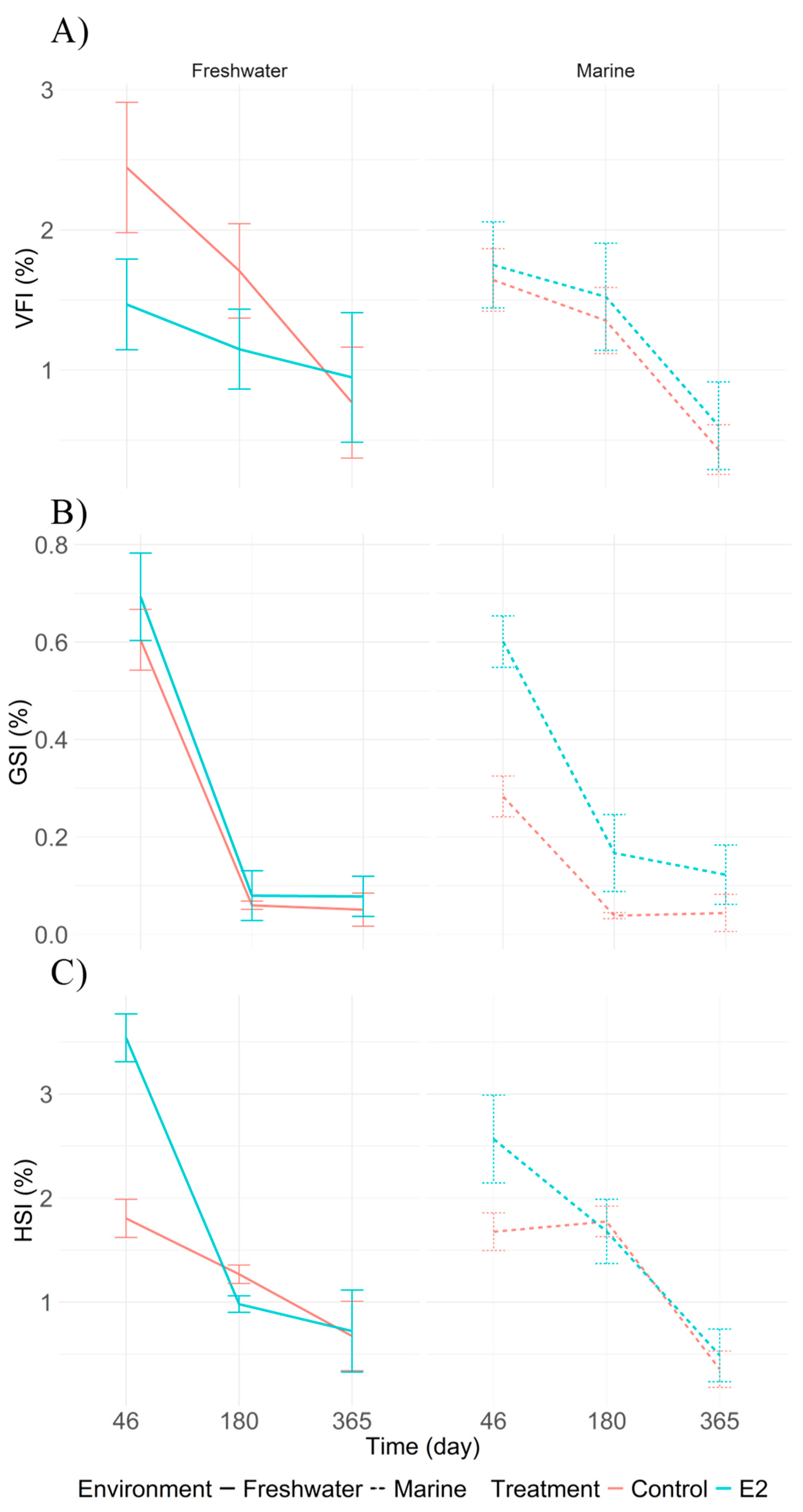
Figure 5.
Gonads from adult Centropomus undecimalis treated with 17β-estradiol were observed at 0. 46. and 180 days. A-B) Immature testis (Stage I); C-D) Transitional ovary (Stage I); E-F) Immature testis (Stage II); G-H) Developing ovary (Stage II). Staining: hematoxylin-eosin. Magnification: 40X. Observed structures: CT: connective tissue; PS: primary spermatocyte; SS: secondary spermatocyte; Sg: spermatogonia; TW: testicular wall; Pv: previtellogenic oocytes; N: nucleus; n: nucleoli; Cy: cytoplasm.
Figure 5.
Gonads from adult Centropomus undecimalis treated with 17β-estradiol were observed at 0. 46. and 180 days. A-B) Immature testis (Stage I); C-D) Transitional ovary (Stage I); E-F) Immature testis (Stage II); G-H) Developing ovary (Stage II). Staining: hematoxylin-eosin. Magnification: 40X. Observed structures: CT: connective tissue; PS: primary spermatocyte; SS: secondary spermatocyte; Sg: spermatogonia; TW: testicular wall; Pv: previtellogenic oocytes; N: nucleus; n: nucleoli; Cy: cytoplasm.
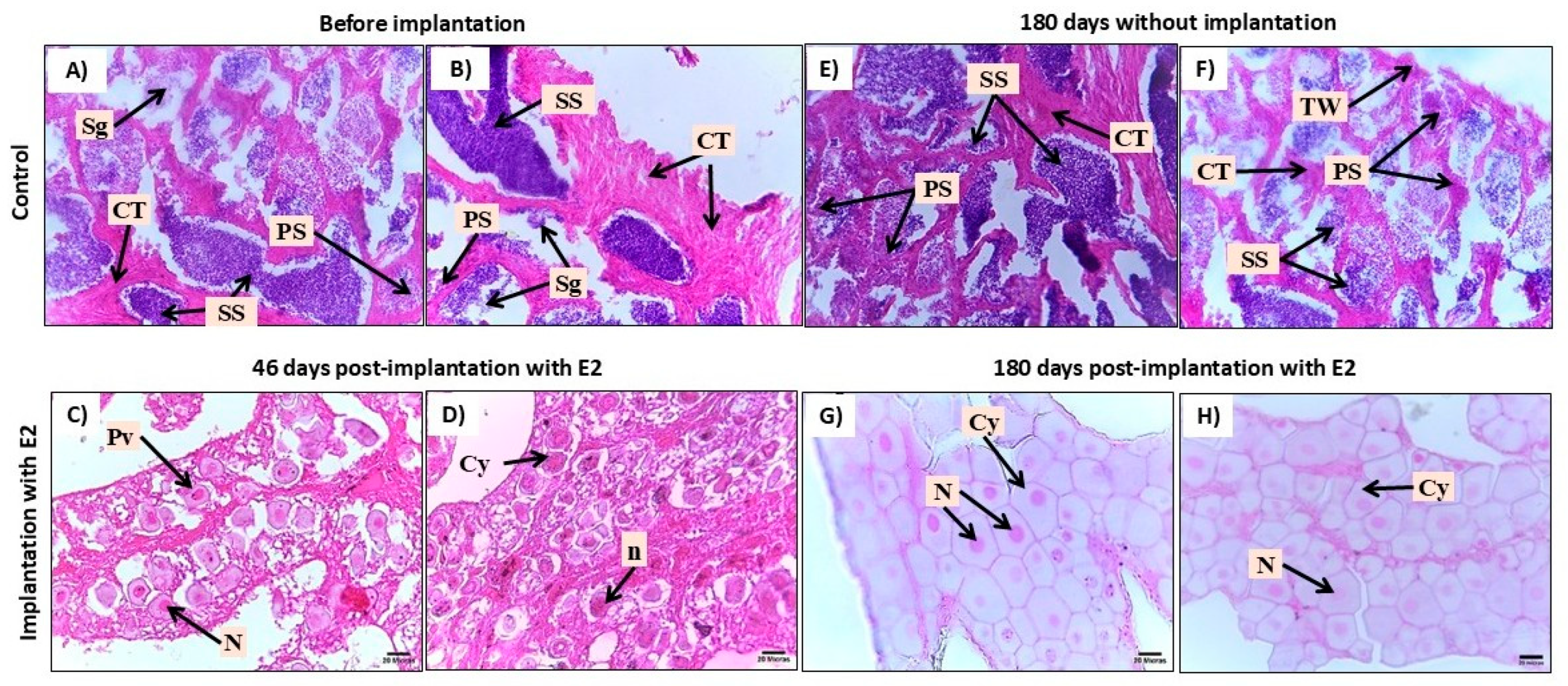
Figure 6.
Plasma concentrations of (A) vitellogenin (Vtg) and (B) 11-ketotestosterone (11-KT) in adult Centropomus undecimalis at 46, 180, and 365 days post-feminization under different treatments. Values are presented as means with 95% confidence intervals derived from Tukey’s HSD test (p < 0.05).
Figure 6.
Plasma concentrations of (A) vitellogenin (Vtg) and (B) 11-ketotestosterone (11-KT) in adult Centropomus undecimalis at 46, 180, and 365 days post-feminization under different treatments. Values are presented as means with 95% confidence intervals derived from Tukey’s HSD test (p < 0.05).
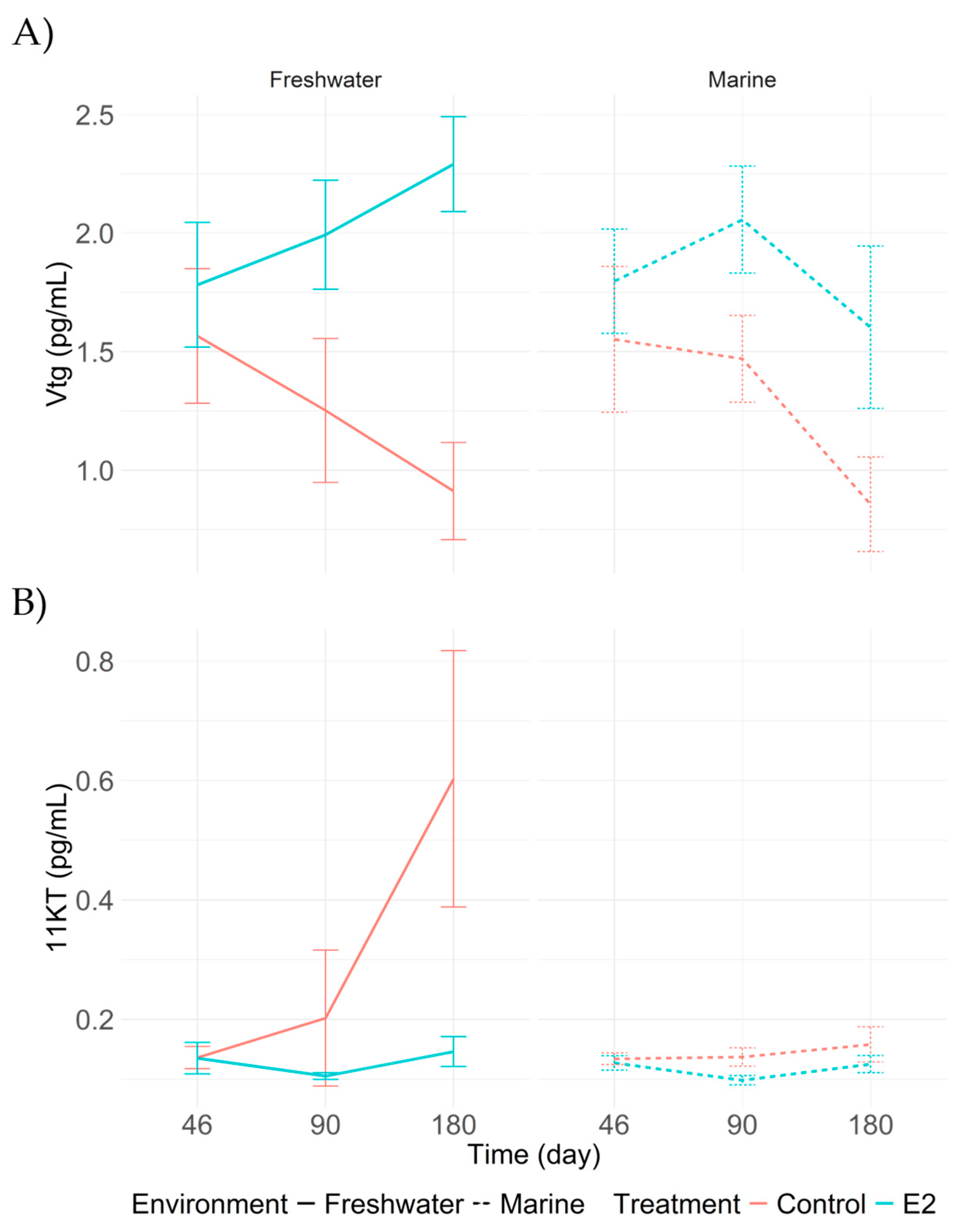
Figure 7.
Comparison of physiological indices in adults Centropomus undecimalis exposed to 17β-estradiol (E2) and control conditions in marine and freshwater environments. A) Visceral Fat Index (VFI); B) Gonadosomatic Index (GSI); C) Hepatosomatic Index (HSI). Values are shown as means with 95% confidence intervals. Differences between sampling times and treatments were evaluated using Tukey’s HSD test (p < 0.05).
Figure 7.
Comparison of physiological indices in adults Centropomus undecimalis exposed to 17β-estradiol (E2) and control conditions in marine and freshwater environments. A) Visceral Fat Index (VFI); B) Gonadosomatic Index (GSI); C) Hepatosomatic Index (HSI). Values are shown as means with 95% confidence intervals. Differences between sampling times and treatments were evaluated using Tukey’s HSD test (p < 0.05).
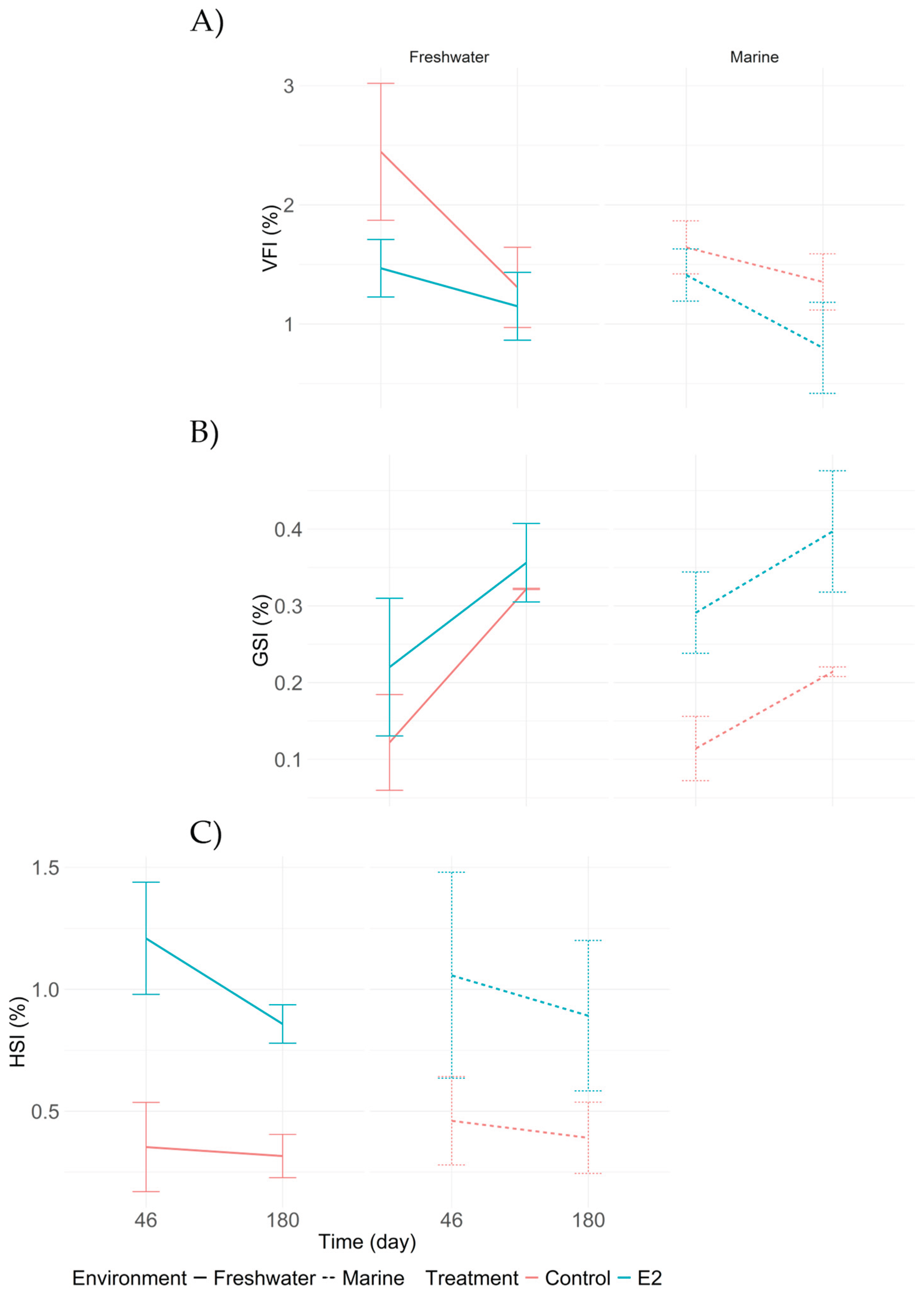

Table 3.
Differentially expressed genes (DEGs) in juvenile Centropomus undecimalis treated with 17β-estradiol (E2) under marine (T1 vs T3) and freshwater (T2 vs T4) conditions, associated with female sex determination and gonadal development.
Table 3.
Differentially expressed genes (DEGs) in juvenile Centropomus undecimalis treated with 17β-estradiol (E2) under marine (T1 vs T3) and freshwater (T2 vs T4) conditions, associated with female sex determination and gonadal development.
| Gene | p-Value | log2FoldChange | ||
| T1 vs T3 | T2 vs T4 | |||
| zar1l | 0.001 | 246829734073046 | ||
| foxl2a | 0.001 | 651651063065463 | ||
| ddx5 | 0.001 | 401534593566732 | ||
| H2A | 0.009 | 306331524924077 | ||
| ddx5 | 0.001 | 307831256725416 | ||
| H2A | 0.001 | 261544452401545 | ||
Table 4.
Zootechnical parameters (mean ± standard error) for juvenile Centropomus undecimalis treated with and without 17β-estradiol (E2) in marine and freshwater environments. Different letters indicate significant differences between treatments for a given variable (p < 0.05).
Table 4.
Zootechnical parameters (mean ± standard error) for juvenile Centropomus undecimalis treated with and without 17β-estradiol (E2) in marine and freshwater environments. Different letters indicate significant differences between treatments for a given variable (p < 0.05).
| Parameters | Treatments | |||
|
T1 (E2-marine) |
T2 (E2-freshwater) |
T3 (marine) |
T4 (freshwater) |
|
| AGR (g/day) | 0.25±0.02 | 0.36±0.06 | 0.24±0.02 | 0.52±0.08 |
| ALG. (cm/day) | 0.03±0.01 | 0.03±0.00 | 0.02±0.00 | 0.04±0.01 |
| SGR (%/day) | 0.39±0.04a | 0.64±0.05ab | 0.38±0.04a | 0.734±0.051b |
| FCR | 8.85±0.69a | 5.68±0.56bc | 8.47±0.62ab | 4.90±0.47bc |
| FER | 12.19±1.98a | 18.94±2.56ab | 12.53±2.14a | 21.43±2.73b |
| PER | 0.205±0.02a | 0.320±0.01bc | 0.215±0.02ab | 0.370±0.05c |
| S (%) | 71 | 67 | 68 | 77 |
AGR, absolute growth rate; ALG, absolute length gain; SGR, specific growth rate; FCR, feed conversion ratio, FER feed efficiency ratio, PER, protein efficiency ratio; S, survival rate.
Table 6.
Zootechnical parameters (mean ± standard error) in adult Centropomus undecimalis treated with 17β-estradiol (E2) via implants in marine and freshwater environments. Different letters indicate significant differences between treatments (p < 0.05).
Table 6.
Zootechnical parameters (mean ± standard error) in adult Centropomus undecimalis treated with 17β-estradiol (E2) via implants in marine and freshwater environments. Different letters indicate significant differences between treatments (p < 0.05).
| Parameters | Treatments | |||
|
T1 (E2-Marine) |
T2 (E2-Freshwater) |
T3 (Marine) |
T4 (Freshwater) |
|
| AGR (g/day) | 1.031±0.33 | 0.615±0.31 | 1.186±0.14 | 0.980±0.39 |
| ALG (cm/day) | 0.016±0.00 | 0.013±0.00 | 0.029±0.01 | 0.024±0.00 |
| SGR (%/day) | 0.163±0.06 | 0.081±0.04 | 0.193±0.02 | 0.153±0.06 |
| FCR | 3.100±0.51 | 3.933±0.63 | 3.080±0.42 | 3.624±0.82 |
| FER | 0.342±0.05 | 0.269±0.04 | 0.336±0.04 | 0.317±0.09 |
| PER | 0.624±0.10 | 0.492±0.08 | 0.615±0.07 | 0.579±0.16 |
| S (%) | 68a | 95b | 73a | 98b |
AGR, absolute growth rate; ALG, absolute length gain; SGR, specific growth rate; FCR, feed conversion ratio, FER feed efficiency ratio, PER, protein efficiency ratio; S, survival rate.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



