Submitted:
19 November 2024
Posted:
21 November 2024
You are already at the latest version
Abstract
Stress is recognized as an adaptive response to potentially harmful environmental stimuli. The primary physiological adaptation to stress is an increase in circulating cortisol levels, which, in excess, can be transferred and incorporated into the oocytes of maturing females, affecting the embryonic developmental program. Additionally, maternal energy availability is an essential environmental factor that modulates this program. Based on this background, we investigated the effects of maternal cortisol on the development of the somatotropic axis in the zebrafish offspring and juveniles. Zebrafish mothers were divided into two groups based on diet: Group 1 received a cortisol-enriched diet, to mimic maternal stress, while Group 2 (control) received a standard diet, for five days, followed by breeding on day three. Offspring were assessed at 0, 24, 48, 72-, 96-, 120, and 144 -hours post-fertilization (hpf). Morphological analyses were performed during embryonic development, including survival rate, body length, presence of pericardial edema, and heartbeat. We examined the gene expression of key somatotropic axis components, including mtor, foxO3a, mafbx, murf1, mstna, gh, igf1, igf2a, igf2b, 11hsdb2, and fkbp5. Our finding offers critical insights for aquaculture by elucidating the complex interaction between maternal factors and somatotropic axis development in offspring. The study demonstrated that cortisol-treated females significantly affected offspring survival, morphology, and cardiac structures, in addition to altering gene expression related to protein synthesis, and showed signs of accelerated development. At 30 days post-fertilization, juveniles exhibited a notable increase in muscle bundle size and cross-sectional diameter. This evidence underscores the importance of aquaculture and animal research regarding how maternal environments influence somatotropic axis development in fish, providing a foundation for improved breeding protocols and management practices.
Keywords:
cortisol
; maternal stress
; stress response
; zebrafish
; somatotropic axis
1. Introduction
Developmental programming is a concept originating from maternal environment stimuli or early environmental conditions, which can lead to significant consequences in embryonic development and result in physiological changes that persist throughout life. Epigenetic mechanisms are central in mediating the effects of adverse environmental factors during early life, influencing long-term metabolic health by inducing enduring alterations in gene expression and organ function [1]. The exploration of early programmed changes dates back several decades. Notably, research conducted in 1957 on rats revealed modifications in the Hypothalamic-Pituitary-Adrenal (HPA) axis response to parental stress [2]. This study demonstrated that early life experiences could alter adults' nervous system reactivity to stress. This subject is being extensively explored across different species, including humans [3]. Exposure to a stressful environment during fetal development has been associated with behavioral dysfunction and disruptions in the HPA axis [4,5,6], along with metabolic [7], neurological [8,9,10], immunological [11,12], cardiovascular [13], and aging-related dysfunctions [14]. Studies on reptiles and birds have shown that maternal stress can lead to abnormal corticosterone deposition in embryos, the primary hormone involved in stress response in vertebrates. This abnormal corticosterone deposition has been found to cause developmental alterations after hatching [15,16]. In fish, cortisol manipulations in embryos or parents have been shown to result in significant alterations in the development of the somatotropic axis [17,18].
The environmental conditions that fish are cultured can subject them to a diverse array of stressors, both physical and chemical. Consequently, understanding the physiological alterations induced by stress is significant in aquaculture [19]. Stress is an adaptative response that triggers the activation of the central nervous system and endocrine pathways to restore homeostasis [11]. The primary response involves the release of catecholamines and corticosteroids via the Hypothalamic-Pituitary-Interrenal (HPI) axis. This is followed by secondary responses, which include the mobilization of energy substrates, enhancement of oxygen transport capacity, and modifications to the immune system. Finally, tertiary effects manifest as behavior, reproduction, development, and growth changes. Thus, stress responses extend beyond simple glucose mobilization, reflecting a complex metabolic adaptation that impacts multiple systems within the organism [20,21].
The HPI axis plays a central role in the primary stress response and releases glucocorticoids in fish. The HPI axis is functionally like the HPA axis in mammals, sharing key similarities. Upon exposure to stress, the hypothalamus releases corticotropin-releasing hormone (CRH), which in turn stimulates the synthesis of adrenocorticotropic hormone (ACTH). ACTH then targets the interrenal tissue, which is comparable to the adrenal gland in mammals, leading to the release of cortisol. Cortisol is the major glucocorticoid in the circulatory system of humans and teleost fish, mediating the physiological stress response [17].
In zebrafish, cortisol during the embryonic phase originates from the mother, as de novo synthesis of this steroid occurs only post-hatching [22]. Early cortisol is crucial for modulating developmental programming, including the activating of the HPI axis [23]. During maternal stress, cortisol is transferred into developing oocytes and subsequently deposited into fertilized embryos. In the embryos, cortisol binds to glucocorticoid receptors (GRs). These GRs when bound to cortisol act as transcription factors that regulate the expression of various target genes in the embryo. Maternal cortisol is deposited in the developing oocyte and is present in decreasing quantities throughout embryogenesis. The enzyme 11β-hydroxysteroid dehydrogenase (11β-HSD) converts cortisol into its inactive form, cortisone, thereby protecting oocytes and embryos from excessive cortisol during maternal stress [17,23]. The interplay between maternal stress and cortisol in oocytes is significant and complex. Therefore, research into the effects of maternal cortisol during early embryonic development is essential for understanding how these effects influence early developmental processes and contribute to phenotypic changes in offspring.
Furthermore, the development of muscle tissue or myogenesis, which initiates in the early stages of fish development can be affected by stress and cortisol. Embryonic muscle formation originates from cellular compartments known as somites, which are derived from mesodermal tissues. The dorsal region of the somites gives rise to myogenic precursor cells (MPCs) that express transcription factors Pax3 and Pax7, which are specific to muscle fiber types. This region also produces a population of reserve progenitor cells, referred to as satellite cells, which are crucial for adult muscle growth [24]. Muscle tissue growth needs a balance between self-renewal and differentiation of precursor stem cells. This balance is regulated by different factors that influence the cell cycle of precursor stem cells, including Meox1, a critical factor for their activity. Zebrafish's muscular system maintains a continuous population of self-renewing precursor cells, facilitating secondary myogenesis throughout life. Secondary myogenesis occurs through hyperplasia and hypertrophy [25]. The activation of precursor stem cells, proliferation of myoblasts, differentiation, formation of myotubules, and maturation of myofibrils are regulated by myogenic regulatory factors (MRFs) such as MyoD, Myf5, myogenin, and MRF4, each with specific roles [26].
In teleost fish and other vertebrates, skeletal muscle represents a significant portion of body mass, and its growth is regulated by a balance between anabolic and catabolic processes influenced by stress and cortisol. The absence of glucocorticoid signaling has been shown to disrupt the expression of myogenic regulatory factors and muscle-specific genes, as evidenced by studies on zebrafish lacking glucocorticoid receptors (GRs) [13,27]. Additionally, stress and cortisol can affect the somatotropic axis. The somatotropic axis comprises the hypothalamus, the pituitary gland, which releases growth hormone, GH-, and the liver, the primary source of insulin-like growth factors (IGFs). Studies in salmonids have demonstrated that acute stress and intraperitoneal cortisol administration lead to significant GH levels decrease in plasma [20]. Therefore, it is plausible that stress and cortisol also influence IGF1 production, as IGF1 synthesis in the liver is stimulated by GH. IGF1 is a key hormone in promoting the proliferation, differentiation, and hypertrophy of muscle precursor cells.
The importance of the somatotropic axis in the growth of fish has been elucidated. On the other hand, perturbations in the growth axis involving environmental factors, including energy availability, stress, photoperiod, temperature, and others, can influence the development of this axis. Recent studies suggest that excessive maternal cortisol can impact embryonic developmental programming. However, the molecular mechanisms underlying these processes remain unclear. This study aimed to explore how the maternal environment affects the development of the somatotropic axis in offspring, using zebrafish as a model organism. Zebrafish were chosen due to their rapid development, ease of manipulation, well-characterized genome, and the transparency of embryos and larvae, which facilitate structural and morphological observations. Their genomic similarity to humans also makes zebrafish a valuable model. The knowledge generated from this study will contribute to research in mammals, including humans, and optimize aquaculture techniques to ensure proper growth and development of offspring.
2. Materials and Methods
2.1. Animal husbandry
Sexually mature males and females (outbred) (4-5 months old) were maintained in the aquarium facility of the Department of Structural and Functional Biology, Institute of Biosciences, Botucatu, São Paulo State University (UNESP) in 6-L tanks on a recirculating system with a 14:10 light: dark cycle, and constant temperature conditions (28 °C). The following water parameters were monitored in all tanks every other day: pH, dissolved oxygen, and ammonia concentration. Males were fed twice a day with commercial food (Pentair Aquatic Habitats, Florida, USA), while females were fed twice a day with a diet prepared with hydrocortisone to assess the effects of maternal stress-associated cortisol on offspring development, as described below. Handling and experimentation were performed according to the Brazilian legislation regulated by the National Council for the Control of Animal Experimental (CONCEA) and Ethical Principles in Animal Research (Protocol n. 8520250320 -CEUA).
2.2. Maternal stress-associated cortisol
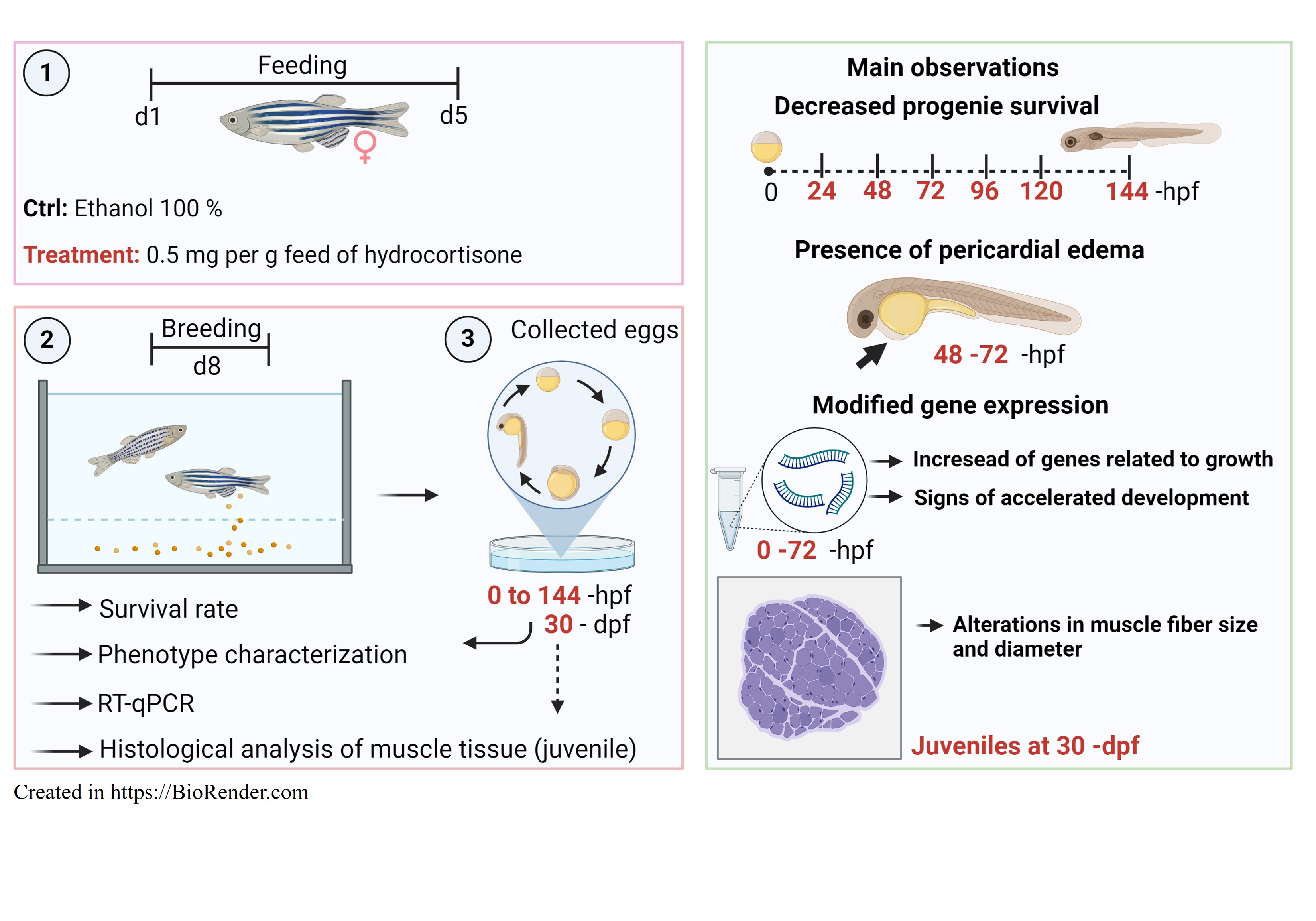
Ten sexually mature female zebrafish were assigned to each of the two 6-L tanks. One group (n = 5) was fed a control zebrafish diet, while the other group (n = 5) was fed a diet laced with cortisol, according to methods previously described [17]. For that, the zebrafish diet was prepared by soaking food pellets in 100 % ethanol either alone (control group) or with 0.5 mg per g feed (hydrocortisone group) (Sigma-Aldrich, St Louis, MO, USA) and allowing ethanol to evaporate [17,28]. This concentration was chosen based on previous studies with maternal cortisol [17]. Zebrafish mothers received thereabout of 25 µg cortisol/ g body mass. Fish were fed twice per day for 5 days. On day 3 post-treatment, five females from each group were transferred to breeding tanks (Figure 1).
2.3. Breeding
On day 3, female fish from each experimental group (control and cortisol) (n = 5/group) were placed for reproduction. The females were divided among five tanks (n = 1 per tank) along with unexposed males in each tank, following the usual methods for zebrafish reproduction [29]. The daily breeding set-up at 18 h, fish were transferred into breeding traps in the same tank, and eggs were collected within the first light (at 9 h) after mating the following morning. Fertilized eggs were collected in Petri dishes containing embryo medium (Hank’s medium, https://zfin.org/zf_info/zfbook/chapt10.html#wptohtml16). The eggs, embryos, and larvae from each group were collected and evaluated from 0-, 24-, 48- 72-, 96-, 120-, and 144 -hours post-fertilization (hpf). In the period from 0 to 72 -hpf, the eggs, embryos, and early larvae were counted and pools of 20 (n = 5 pools of 20 individuals for each pool) were snap-frozen on liquid nitrogen and stored at – 80 °C for gene expression analysis. Furthermore, eggs, embryos, and larvae from 0 to 144 -hpf were sampled for phenotype characterization and larval morphology. Juveniles at 30 -days post fertilization (dpf) were also collected for muscle fiber analysis in both groups, as described above (Figure 1).
2.4. Transcript levels of the somatotropic axis by quantitative real-time PCR (qPCR)
mRNA abundance of key genes involved in the somatotropic axis was measured by qPCR in embryos from 0 to 72 -hpf. Embryos were pooled into 20 (n = 5 pools of 20 individuals for each pool) per experimental group (control and cortisol treatment). Total RNA was extracted from the pools using a commercial kit (PureLink TM RNA mini kit, Ambion, Life Technologies, Carlsbad, CA, United States). After RNA extraction, the usual downstream methods were followed according to the manufacturer’s instructions, and quality and purity were checked with a NanoDropTM One Spectrophotometer (Thermo Scientific, Madison, WI, United States). cDNA synthesis was performed as described by Nóbrega et al. (2010) [30]. qPCR reactions were conducted using 5 μl of 2X SYBR-Green Universal Master Mix, 1 μl of forward primer (1.125 nM), 1 μl of reverse primer (1.125 nM), 0.5 μl of DEPC water, and 2.5 μl of cDNA. The relative mRNA levels of intracellular genes related to growth regulation through syntheses, such as mtor (mammalian target of rapamycin), and protein degradation, including foxo3a, mafbx, and murf1 (forkhead box O3, muscle atrophy F-box, and muscle ring-finger protein-1, respectively) were quantified. Additionally, genes involved in the systemic or hormonal control of anabolic pathways, such as gh, igf1, igf2a, and igf2b (growth hormone, insulin-like growth factor 1, insulin-like growth factor 2a and 2b, respectively), as well as mstna (myostatin), which is related to the regulation of catabolism, were also measured. Furthermore, the relative mRNA levels of hsd11b2 (hydroxysteroid 11-beta dehydrogenase 2) and fkbp5 (glucocorticoid receptor chaperone protein) were evaluated to denote changes in the Hypothalamic-Pituitary-Interrenal (HPI) axis [17,31]. mRNA levels of the targets (Cts) were normalized by the transcript levels of β-actin and expressed as relative values of the control group (as fold induction), according to the 2−(∆∆CT) methods. Primers were designed according to zebrafish sequences (Table 1). Each primer was validated using a melting curve (Figure S1).
2.5. Embryo and larvae phenotypes
Embryos and larvae at 48 to 72 -hpf from both control and cortisol-treated groups were collected from spawning tanks and evaluated for morphological changes. The selected period corresponds to the post-hatching stage, during which larvae are presumed to experience the most significant influence of maternal cortisol deposition, as de novo synthesis of this steroid begins only after hatching. A total of 30 larvae were randomly chosen from each group, and 30 bright-field images from each specimen were captured using an Olympus MVX10 MacroView stereomicroscope. These images were subsequently utilized to measure morphometric parameters in developing larvae with annotation performed using the Olympus cellSens dimensions software (https:www.olympuslifescience.com/pt/software/cellsens/), and analysis conducted via ImageJ software (available at http://imagej.nih.gov/ij/index.html). The developmental differences between the control and treatment groups were assessed through morphometric evaluations. The total length of larvae was measured, and abnormal developments were identified, including pericardial and yolk sac edema, inflated swim bladder, and frequency of body deformation and/or abnormal curvature. The survival rate of zebrafish embryos and larvae in the range of 0 to 144 -hpf was evaluated for each group using the Kaplan-Meier method. Survival was assessed based on the percentage of viable larvae relative to the total number of specimens collected post-fertilization (0 hpf). Embryos and larvae found at the bottom of the Petri dish with opaque appearance and abnormal coloration, along with hyporesponsiveness to mechanical stress were considered dead.
2.6. Heart rate measurement in embryos at 48 -hpf
Videos from zebrafish embryos at 48 -hpf from both maternal control and cortisol-treated groups (n = 30/condition) were recorded to calculate heart rate per minute. The non-anesthetized embryos were immobilized using excavated slide blanks containing sufficient embryonic medium to ensure survival during filming. The setup consists of an Olympus MVX10 MacroView stereomicroscope linkage to Olympus cellSens dimensions software (https://www.olympuslifescience.com/pt/software/cellsens/) for video acquisition. The video files (.AVI) were obtained at 30 frames per second (fps) totaling 60 seconds of recording, with a resolution of 4080 x 3072 pixels. Heart rate quantification was performed using a Python script named CardiOT2® software, which employs the Fast Fourier (FFT) methodology to calculate heart rate contraction cycles.
2.7. Histological analysis of muscle tissue
Five fish at 30 days post-fertilization (dpf) were collected from each experimental group (control and cortisol) for cross-sectional analysis, while three larvae per group were used for longitudinal analysis. The specimens were anesthetized with 0.1% benzocaine and sacrificed for skeletal muscle tissue collection. Samples were immediately fixed in Karnovsky solution (glutaraldehyde 2.5 %, paraformaldehyde 2 %, 0.1 M phosphate buffer, pH 7.2) and preserved in 70 % alcohol. Samples were dehydrated using graded ethanol concentrations (80, 95, and 95 %) and embedded in Technovit 7100 resin (Heraeus Kulzer, Wehrheim, Germany), according to the manufacturer’s protocol. Longitudinal and cross-section histological areas (4 µm) of muscle fibers were prepared through a microtome equipped with a glass knife. Sections were stained with 0.1 % toluidine blue in 1 % sodium borate. Fiber diameter and frequency were determined using a Leica DMI6000 microscope (Leica, Heidelberg, Germany) coupled with an image analyzer. Muscle fibers were classified based on the criteria previously described [32], with fibers smaller than 9 µm categorized as thin fibers, and those larger than 9 µm as thick fibers.
2.8. Statistical analysis
The data were subjected to the mean ± SEM (Standard Error of Mean). Significant differences between the two groups were identified using Student’s t-tests non-parametric (unpaired) to compare two groups, the treated group versus the control group. The significance level (p) was considered ≤ 0.05 in both cases. The normal distribution of the samples was assessed using the Kolmogorov-Smirnov test, which was performed for all genes used in this study. All analysis was carried out using Graph Pad Prism software 8.0.1 (Graph Pad Software, Inc., San Diego, CA, United States).
3. Results
3.1. Maternal cortisol influences the mRNA expression levels of genes associated with the somatotropic and HPI axes in zebrafish offspring
Samples were collected at 0, 24, 48, and 72 -hpf of both the control and cortisol-treated groups for gene expression analysis by qPCR (Figure 1). The expression levels of essential components involved in the somatotropic and HPI axes, such as mtor, foxo3a, mafbx, murf1, mstna, gh, igf1, igf2a, igf2b, fkbp5, and 11hsdb2, were significantly altered in the zebrafish progeny from females fed with cortisol-enriched diet when compared to the control (Figure 2). Our analysis revealed that maternal cortisol treatment up-regulated mtor mRNA levels in embryos at 24 -hpf (Figure 2A). We also evaluated the genes involved in muscle protein degradation. Embryos and larvae from the maternal cortisol-treated group exhibited increased levels of foxo3a at 24, 48, and 72 -hpf compared to the control (Figure 2B). The mRNA expression levels of mafbx e murf1, which are genes involved in the catabolic pathway, were significantly altered in the treated group. While mafbx levels were significantly elevated at 48 and 72- hpf (Figure 2C), murf1 increased expression at 0 and 48 -hpf (Figure 2D). In this regard, the expression of mstna, a protein that inhibits myogenesis, a significant decrease in its expression was observed in the treated group at 0 -hpf, followed by a significant increase at 24 -hpf (Figure 2E). Maternal cortisol treatment also significantly increased the expression of gh in embryos at 0 -hpf (Figure 2F). However, a substantial decrease in primary gh transcripts was observed in early larvae at 72 -hpf from females fed with a cortisol-treated diet (Figure 2F). The expression levels of igf1, igf2a, and igf2b were measured. igf1 was up-regulated from 0 to 48 -hpf in the cortisol-treated group (Figure 2G), while igf2b expression increased only at 0 -hpf (Figure 2I). On the other hand, the mRNA levels of igf2a remained unchanged across the experimental groups at the sampled time points (Figure 2H). Finally, we evaluated the expression of genes responsive to the HPI axis. A significant decrease in fkbp5 expression levels was observed exclusively at 0 -hpf (Figure 2J). However, the mRNA levels of the enzyme 11β-HSD2 (11hsdb2), responsible for converting cortisol into its inactive form, cortisone, showed no significant differences between the experimental groups (Figure 2K).
3.2. Maternal cortisol treatment affected different stages of offspring development in different parameters of evaluation
We investigated the impact of maternal cortisol on embryos and larvae survival and phenotype. Embryos and larvae were collected at intervals from 0 to 144 -hpf in both the control and cortisol-treated groups for survival analysis. The survival rate was conducted using the Kaplan-Meier method. The results showed a higher mortality rate among offspring from females fed a cortisol-enriched diet compared to the control group (Figure 3A). The Log-rank test (Mantel-Cox) indicated a significant relative risk of death between the groups during the study period (chi-square = 64; p < 0.0001). The control group maintained a survival rate of 90.5 % at 24 -hpf, with no decline observed up to 144 -hpf. However, the offspring from the cortisol group showed survival rates of 79.6 %, 78.6 %, 73.5 %, and 70.9 % at 24, 48, 72, and 144 -hpf, respectively.
Body length measurements of larvae at 48 and 72 -hpf were measured. Maternal cortisol exposure significantly increased larval length at 48 -hpf compared to the control group (Figure 3B). The morphological evaluation indicated that offspring from females fed a cortisol-enriched diet exhibited moderate cardiac edema, as classified previously [13]. Specifically, 63.3 % and 36.6 % of larvae displayed moderate pericardial edema at 48 and 72 -hpf, respectively (Figure 3C-D). No morphological abnormalities were noted in the control group (Figure 3E-F).
Additionally, heart rate measurements at 48 -hpf showed no significant difference between offspring from cortisol-treated and control females (Figure 4).
At 30 -dpf, histological analysis of muscle fibers in the progeny of cortisol-treated females revealed distinct changes. Longitudinal sections revealed that juveniles from the maternal cortisol-treated group had significantly longer muscle bundles compared to the control group (mean 387.6 ± 6.71 µm, and 287.6 ± 24.81 µm, respectively) (Figure 5A-C).
Furthermore, cross-section analysis indicated that the total muscle diameter was larger in juveniles from mothers treated with cortisol (254 ± 9.27 µm) compared to those from control females (187.7 ± 6.44 µm) (Figure 6A, B, E). Furthermore, cross-sections showed a higher percentage of thick muscle fiber (> 9 µm) in the juvenile from maternal cortisol-treated, averaging 72.5 %, compared to 35.7 % in the control group. However, the progeny from the control group exhibited a higher proportion of thin muscle fibers (< 9 µm), averaging 63.7 % compared to 27.5 % in the progeny from the maternal cortisol group (Figure 6C, D, F).
4. Discussion
Zebrafish (Danio rerio) is an ideal model to investigate the early developmental effects of cortisol signaling, as de novo synthesis of cortisol begins only after hatching (48 – 72 hours post-fertilization, hpf). Therefore, maternal cortisol is a major influence on early developmental programming [22]. Previous research has highlighted the critical role of maternal cortisol in embryogenesis [13,27]. Studies employing morpholino oligonucleotides to reduce glucocorticoid receptor expression in zebrafish embryos showed mesodermal malformations, including bent tails and deformed somites, as well as reduced growth and decreased survival rates [13,27].
On the other hand, to investigate the influence of increased maternal cortisol deposition in embryos, several experiments have been conducted in fish to assess phenotypic changes in offspring, particularly concerning the somatotropic axis [33]. In zebrafish, Nesan and Vijayan (2016) [23] demonstrated that microinjection of 32 pg pf cortisol into newly fertilized embryos – a technique used to mimic maternal cortisol transfer and deposition – resulted in increased embryo size at 48 and 72 -hpf. Similarly, our study found that adding cortisol to the maternal diet resulted in maternal transfer of cortisol, which increased offspring size at 48 -hpf. Interestingly, the same researchers [23] demonstrated that injection of cortisol-specific antibodies resulted in decreased embryo length and abnormal morphologies, including moderate tail twist and deformed body curvature. These findings demonstrated the crucial role of cortisol in mesoderm formation and muscle development [13], prompting further investigation into whether a positive adjustment of cortisol levels could be beneficial for embryogenesis and myogenesis.
Another critical aspect to consider is the cardiac structure. Offspring from females fed a cortisol-enriched diet exhibit pericardial edema. This finding is particularly relevant to our study, as the heart is composed of muscle tissue and originates from the mesodermal layer during embryogenesis [34]. Interestingly, Nesan and Vijayan (2012) [13] demonstrated that increasing cortisol levels in newly fertilized zygotes via microinjection also disrupted cardiogenesis, leading to pericardial edema, and cardiac chamber malformations. Similar results support the hypothesis that maternal cortisol programs embryogenesis, though elevated levels of this steroid in the embryo can have adverse effects. Moreover, the morphometric data from our study align with the literature confirming the effectiveness of using a cortisol-enriched diet to simulate maternal stress and subsequent cortisol transfer to the offspring [17]. Nesan and Vijayan (2012) [13] also observed a reduction in heart rate in 72 -hpf larvae that were exposed to elevated cortisol levels during pre-hatch. On the other hand, our study showed that maternal cortisol treatment did not alter the heart rate of larvae at 48 -hpf, despite observed pericardial edema. We hypothesize that the discrepancy between the results may be attributed to differences in the treatment methods – such as cortisol microinjection into embryos versus maternal cortisol diet supplementation – or to a specific time when heart rate was measured.
To gain a deeper understanding of how cortisol influences zebrafish embryonic development, we focused on gene expression analysis. We examined key developmental genes involved in the somatotropic axis, including gh, igf1, igf2a, and igf2b. In vertebrates, GH functions directly by binding to receptors on the muscle sarcolemma and indirectly by stimulating the production and release of insulin-like growth factors (Igf) in the liver and peripheral tissues [35]. In zebrafish, the Igf system includes igf1, igf2, and igf3, which igf3 being specific to gonadal tissue [36]. igf2 and igf3 are involved in midline development. The particular relevance to our study is igf1, which acts through the PI3K-Akt-TOR anabolic pathways by binding to its receptor, thereby being essential for normal embryonic development and myogenesis [24]. Indeed, reduced igf1 signaling in zebrafish has been associated with decreased synthesis of major muscle proteins, leading to a reduction in muscle fiber diameter [32]. The gene expression data reveal a significant increase in gh mRNA levels after fertilization, suggesting that this transcript is maternally inherited and positively regulated by maternal cortisol. As development progresses, relative gh levels normalize at 24 and 48 -hpf. However, there was a marked decline of gh levels at 72 -hpf in the offspring of cortisol-treated females. Consistent with this, igf1 mRNA levels increased at 0, 24, and 48 -hpf in embryos and larvae from cortisol-treated females. Notably, the increase in larval size at 48 -hpf brings into line with the elevated levels of the growth factor. In our study, the elevated transfer of maternal cortisol resulted in a coincidence increase in gh and igf1 levels in the first hour post-fertilization, followed by a subsequent downregulation during embryogenesis. This suggests a direct relationship between the HPI and somatotropic axes. Our findings are consistent with the literature, which emphasizes the crucial role of cortisol in the proper morphological development of zebrafish [21,37].
Additionally, we analyzed the mRNA levels of genes related to skeletal muscle development, including mtor, foxo3a, and murf1. The regulation of muscle fiber volume and quantity involves a complex interplay of anabolic and catabolic proteins, which respectively promote protein synthesis and degradation [24]. The IGF-Akt pathway plays a pivotal role in this process, functioning through two key mechanisms in fish. In the anabolic process, mtor protein is phosphorylated and activated, playing a central role in cell growth and proliferation, which drives hypertrophy and hyperplasia. On the other hand, muscle atrophy is prevented by the phosphorylation and inactivation of the foxo3a protein, resulting in the downregulation of the ubiquitin ligases murf1 and mafbx, which mark proteins for degradation by the proteasome complex [38]. In this study, we observed an upregulation of mtor and foxo3a expression in embryos at 24 -hpf in the maternal cortisol-treated group, suggesting an increase in muscle protein turnover. This phenomenon may explain the increased embryo length observed in larvae at 48 -hpf in the treated group. Subsequently, we observed the upregulation in the mRNA expression of protein degradation markers, such as foxo3a, murf1, and mafbx in embryos at 48 and 72 -hpf along with stabilization of mtor expression, which may have prevented the exacerbated growth post-fertilization. Consequently, there was no significant difference in embryo length at 72 -hpf between maternal cortisol-treated and control groups.
On the other hand, myostatin, a member of the tumor growth factor-β (TGF-β) superfamily, is a well-established negative regulator of myogenesis in vertebrates. In zebrafish, inhibition of myostatin expression leads to a significant increase in muscle mass [39,40]. Furthermore, two myostatin gene variants, mstna and mstnb, can be found in zebrafish. mstna mRNA being maternally inherited and highly expressed in embryos at 48 -hpf [41,42,43]. Our study demonstrated that mstna mRNA levels were inhibited in embryos after fertilization in the maternal cortisol-treated group. This suggests that the increase in larval length at 48 -hpf is related to the inhibition of mstna levels, leading to enhanced muscle growth, as indicated by elevated mtor expression. However, this interpretation was complex when we observed the suppression of mRNA transcripts of genes involved in protein degradation, murf1 and mafbx. Additionally, there was a significant increase in mstna expression in embryos at 24 -hpf corresponding to the treated group, which could be directly related to the elevated expression of foxo3a, murf1, and mafbx in the period of 48 and 72 -hpf. These findings suggest that this process's regulatory mechanisms are still poorly understood, particularly during the post-fertilization period.
Finally, we also evaluated the expression of cortisol-responsive genes: hsd11b2 e fkbp5 [17,31,44]. FK506-binding protein 5 (Fkbp5) is a glucocorticoid receptor chaperone protein that is responsive to elevated glucocorticoid levels in zebrafish. This observation was confirmed in Fdx1b-deficient zebrafish males, in which its expression was decreased in response to decreased plasma cortisol levels [45]. In the present study, fkbp5 expression was reduced in 0 -hpf embryos from cortisol-treated females, indicating either that maternal cortisol transfer did not occur as expected, or that there are counter-regulatory mechanisms in response to excess glucocorticoids in the offspring. The low response to cortisol receptors activating during the early stages of embryogenesis could have influenced the increased larval size observed at 48 -hpf in the treated group, which indicates the crosstalk between the HPI and somatotropic axes.
The literature suggests that ovarian follicles, eggs, and embryos are not passive recipients of maternal steroids [33]; they possess mechanisms to either transport cortisol out of the embryo or metabolize it into its inactive form. Consequently, the transfer of maternal cortisol to the offspring and its subsequent effects involves passive and counter-regulatory mechanisms. Faught et al. (2016) [17] demonstrated an increase in the hsd11b2 transcript in zebrafish ovarian follicles following ex vivo cortisol incubation, indicating that maternal stress may lead to the activating of ovarian enzymes that mitigate cortisol levels in oocytes. However, our findings suggest that cortisol deposition is transient due to the asynchronous type of zebrafish reproductive, in which vitellogenic oocytes might be susceptible to excessive steroid uptake during a specific window before the upregulation of hsd11b2 activity in ovarian follicles in response to increased cortisol levels. Notably, no significant differences in hsd11b2 expression were observed between experimental groups up to 72 -hpf, supporting our hypothesis. Although hsd11b2 mRNA levels were unchanged, the expression of fkbp5 was reduced post-fertilization. Therefore, the mechanism behind the increased cortisol levels in embryos remains unclear, and further research is required to better understand the mechanism of maternal cortisol transfer in fish. We propose that maternal cortisol transfer occurred without a counter-regulatory effect on fkbp5 mRNA levels, as previously demonstrated by Tovo-Neto et al. (2020) [31], who showed that high cortisol concentrations increase fkbp5 expression. The results obtained from morphological and histological analysis in this study are consistent. We believe that feeding females with cortisol-enriched feed may lead to the transfer of cortisol to their offspring.
Histological analysis of muscle fiber in larvae at 30 -dpf from a mother-fed cortisol-enriched diet revealed a significantly larger muscle bundle size compared to the larvae from the control group. Moreover, larvae from the treated group exhibited an increased cross-section diameter. These findings are consistent with previous studies. Wilson et al. (2016) [44] showed that treating fertilized embryos with the corticosteroid dexamethasone for 5 days resulted in increased larval size at 84 -dpf. This suggests that changes in corticosteroid levels in ovarian follicles or directly in the embryos can induce long-term morphological changes. The same study [44] investigated the acute and chronic effects of the synthetic glucocorticoid receptor (GR) agonist dexamethasone. At 120 -hpf, larvae treated exhibited reduced cortisol levels compared to control larvae. Additionally, Nesan and Vijayan (2016) [23] demonstrated that alterations in maternal cortisol availability affect the cortisol response in zebrafish embryos post-hatching. Specifically, reducing cortisol levels in embryos via microinjection of specific antibodies resulted in an enhanced cortisol response. In contrast, larvae from embryos microinjected with cortisol did not show a cortisol response following an acute physical stressor, indicating potential disruption of the HPI axis activity. In vertebrates, muscle growth occurs through a combination of two main processes: hypertrophy, which is the increase in the size and length of existing muscle fibers, and hyperplasia, which involves the formation of new muscle fibers from the differentiation of muscle stem cells. Research in zebrafish has demonstrated that the cross-section area (CSA) of muscle fibers correlates directly with the fish's age [46]. Additionally, newly formed muscle fibers through hyperplasia in older zebrafish exhibit increased CSA and length compared to those in younger fish [47]. Our results showed that juveniles at 30 -dpf from cortisol-fed females had a higher percentage of thick fibers compared to the control group. These findings suggest that maternal cortisol treatment may have accelerated the metabolic rate and developmental progression of embryos, as evidenced by changes in length during larval development. Another hypothesis is that the treatment induced greater hypertrophy than muscle hyperplasia.
5. Conclusions
In this study, we showed that maternal stress linked to cortisol deposition significantly impacts the morphological and cardiac structures of offspring development. Embryos from cortisol-treated females exhibited modified expression of genes associated with protein synthesis and showed signs of accelerated development. In juveniles, alterations in muscle fiber size and diameter were observed. Overall, our study suggests that parental treatment and environmental conditions can influence the function of the hypothalamic-pituitary-interrenal (HPI) axis in offspring, thereby impacting the growth and viability of offspring. This highlights the importance of standardizing breeding and reproductive methods in the aquaculture system.
Supplementary Materials
The following supporting information can be downloaded at: www.mdpi.com/xxx/s1, Figure S1: (A) Primer specificity. The primer specificity was verified according to the melt curve temperature (dissociation temperature). The dissociation temperature in all qPCR reactions is demonstrated for each primer. Each dot represents the melt curve temperature for one sample. (B) A scatter plot was generated to assess the stability of β-actin as a reference gene in samples of embryo/early larvae collected at 0 to 72 -hpf from females fed only with commercial food (control group) and fed with a diet laced with 0.5 mg hydrocortisone per gram of feed. No significant differences (p>0.05; ANOVA following Tukey’s multiple comparison test) were observed in β-actin Ct values between control and treated samples across the experimental groups. Table S1: Primers used for gene expression studies (qPCR) (FW = Forward; RV = Reverse).
Author Contributions
Conceptualization, R.H.N., M.S.R. and V.P.B.T.; methodology, V.P.B.T. and M.S.R.; formal analysis, V.P.B.T. and M.S.R.; investigation, V.P.B.T. and M.S.R.; data curation, V.P.B.T. and M.S.R.; writing—original draft preparation, M.S.R.; writing—review and editing, M.S.R. and R.H.N.; visualization, M.S.R. and R.H.N.; supervision, M.S.R. and R.H.N. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP) (2019/22997-3 granted to M.S.R.; 2019/22702-3 granted to V.P.B.T.; 2021/06742-5 granted to R.H.N.).
Institutional Review Board Statement
The current research was conducted following the Ethical Principles on Animal Experimentation adopted by the National Council for the Control of Animal Experimentation (CONCEA/Brazil). All procedures used in this study were approved by the CEUA (Committee on Ethics in the Uses of Animals) of São Paulo State University (UNESP), protocol number 8520250320-CEUA.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation, to any qualified researcher. The data are available from the author, M.S.R., upon request.
Acknowledgments
The authors gratefully acknowledge the Reproductive and Molecular Biology Research Group for their support in the development of this study.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Tobi, E. W.; Slieker, R. C.; Luijk, R.; Dekkers, K. F.; Stein, A. D.; Xu, K. M. DNA methylation as a mediator of the association between prenatal adversity and risk factors for metabolic disease in adulthood. Sci Adv. 2018, 4, eaao4364, 1-10.
- Levine, S. Infantile experience and resistance to physiological stress. Science. 1957, 126 (3270), 405-405.
- Joseph, D.; Whirledge, S. Stress and the HPA axis: Balancing homeostasis and fertility. Int J Mol Sci. 2017, 18, 2224.
- Brunton, P. J.; Russell, J. A. Prenatal social stress in the rat programs neuroendocrine and behavioral responses to stress in the adult offspring: Sex-specific effects. J Neuroendocrinol. 2010, 22, 258–271.
- Dreiling, M.; Schiffner, R.; Bischoff, S.; Rupprecht, S.; Kroegel, N.; Schubert, H.; Rakers, F. Impact of chronic maternal stress during early gestation on maternal-fetal stress transfer and fetal stress sensitivity in sheep. Stress. 2017, 21, 1–10.
- D’Agostino, S.; Testa, M.; Aliperti, V.; Venditti, M.; Minucci, S.; Aniello, F.; Donizetti, A. Expression pattern dysregulation of stress- and neuronal activity-related genes in response to prenatal stress paradigm in zebrafish larvae. Cell Stress Chaperones. 2019, 24, 1005–1012.
- Entringer, S. Impact of stress and stress physiology during pregnancy on child metabolic function and obesity risk. Curr Opin Clin Nutr Metab Care. 2013, 16, 320–327.
- Coulon, M.; Wellman, C. L.; Marjara, I. S.; Janczak, A. M.; Zanella, A. J. Early adverse experience alters dendritic spine density and gene expression in prefrontal cortex and hippocampus in lambs. Psychoneuroendocrinology.2013, 38, 1112-1121.
- Babenko, O.; Kovalchuk, I.; Metz, G. A. S. Stress-induced perinatal and transgenerational epigenetic programming of brain development and mental health. Neurosci Biobehav Rev. 2015, 48, 70–91.
- Hirst, J. J.; Cumberland, A. L.; Shaw, J. C.; Bennett, G. A.; Kelleher, M. A.; Walker, D. W.; Palliser, H. K. Loss of neurosteroid-mediated protection following stress during fetal life. J Steroid Biochem Mol Biol. 2016, 160, 181–188.
- Veru, F.; Laplante, D. P.; Luheshi, G., King, S. Prenatal maternal stress exposure and immune function in the offspring. Stress. 2014, 17, 133–148.
- Marques, A. H.; Bjørke-Monsen, A.-L.; Teixeira, A. L.; Silverman, M. N. Maternal stress, nutrition and physical activity: Impact on immune function, CNS development, and psychopathology. Brain Res. 2015, 1617, 28–46.
- Nesan, D.; Vijayan, M.M. Embryo exposure to elevated cortisol level leads to cardiac performance dysfunction in zebrafish. Mol Cell Endocrinol. 2012, 363, 85–91.
- Haussmann, M. F.; Longenecker, A. S.; Marchetto, N. M.; Juliano, S. A.; Bowden, R. M. Embryonic exposure to corticosterone modifies the juvenile stress response, oxidative stress and telomere length. Proc Biol Sci. 2011, 279, 1447–1456.
- De Fraipont, M.; Clobert, J.; John-Alder, H.; Meylan, S. Increased pre-natal maternal corticosterone promotes philopatry of offspring in common lizards Lacerta vivipara. Journal of Animal Ecology. 2000, 69, 404–4013.
- Tissier, M. L.; Williams, T. D.; Criscuolo, F. Maternal effects underlie ageing costs of growth in the zebra finch (Taeniopygia guttata). PloS ONE. 2014, 9, e97705.
- Faught, E.; Best, C.; Vijayan, M. M. Maternal stress-associated cortisol stimulation may protect embryos from cortisol excess in zebrafish. R Soc Open Sci. 2016, 24, 160032.
- Faught, E.; Vijayan, M. M. The mineralocorticoid receptor is essential for stress axis regulation in zebrafish larvae. Sci Rep. 2018, 8, 18081.
- Yoshioka, E. T.O.; Mariano, W.S.; Santos, L.R.B. Estresse em peixes cultivados: Agravantes e atenuantes para o manejo rentável. In Manejo e sanidade de peixes em cultivo, 1st ed.; Tavares-Dias, M; Macapá: Embrapa Amapá, Brasil, 2009; pp. 226–247.
- Leatherland, J. F.; Li, M.; BARKATAKI, S. Stressors, glucocorticoids and ovarian function in teleosts. J of Fish Biol. 2010, 76, 86–111.
- Nesan, D.; Vijayan, M. M. Role of glucocorticoid in developmental programming: Evidence from zebrafish. Gen Comp Endocrinol. 2013, 181, 35–44.
- Alsop, D.; Vijayan, M. M. Development of the corticosteroid stress axis and receptor expression in zebrafish. Am J Physiol Regul Integr Comp Physiol. 2008, 294, R711–R719.
- Nesan, D.; Vijayan, M. M. Maternal cortisol mediates hypothalamus-pituitary-interrenal axis development in zebrafish. Sci Rep. 2016, 6, 22582.
- Johnston, I. A.; Bower, N. I.; Macqueen, D. J. Growth and the regulation of myotome muscle mass in teleost fish. J Exp Biol. 2011, 214, 1617–1628.
- Keenan, S.; Currie, P. The developmental phases of zebrafish myogenesis. J Dev Biol. 2019, 7, 12.
- Hinits, Y.; Osborn, D. P. S.; Hughes, S. M. Differential requirements for myogenic regulatory factors distinguish medial and lateral somitic, cranial and fin muscle fibre populations. Development. 2009, 136, 403–414.
- Pikulkaew, S.; Benato, F.; Celeghin, A.; Zucal, C.; Skobo, T.; Colombo, L.; Valle, L. D. The knockdown of maternal glucocorticoid receptor mRNA alters embryo development in zebrafish. Dev Dyn. 2011, 240, 874–889.
- Alderman, S.L.; Mcguire, A.; Bernier, N.J.; Vijayan, M.M. Central and peripheral glucocorticoid receptors are involved in the plasma cortisol response to an acute stressor in rainbow trout. Gen Comp Endocrinol. 2012, 176, 79–85.
- Tsang, B.; Zahid, H.; Ansari, R.; Lee, R.C. Partap, A.; Gerlai, R. Breeding zebrafish: A review of different methods and a discussion on standardization. Zebrafish. 2017, 14, 561–573.
- Nóbrega R.H.; Greebe C.D.; Van De Kant H.; Bogerd J.; França L.R.; Schulz R.W. Spermatogonial stem cell niche and spermatogonial stem cell transplantation in zebrafish. PLoS One. 2010, 5, e12808.
- Tovo-Neto, A.; Martinez, Emanuel R. M.; Melo, A.G.; Doretto, L. B.; Butzge, A. J.; Rodrigues, M. S.; Nakajima, R.T.; Habibi, H. R.; Nóbrega, R.H. Cortisol directly stimulates spermatogonial differentiation, meiosis, and spermiogenesis in zebrafish (Danio rerio) testicular explants. Biomolecules. 2020, 10, 429.
- Figueiredo, M.A.; Mareco, E.A.; Silva, M.D.P.; Marins, L.F. Muscle-specific growth hormone receptor (GHR) overexpression induces hyperplasia but not hypertrophy in transgenic zebrafish. Transgenic Res. 2012, 21, 457–469.
- Sopinka, N. M.; Capelle, P. M.; Semeniuk, C. A. D.; Love, O. P. Glucocorticoids in fish eggs: variation, interactions with the environment, and the potential to shape offspring fitness. Physiol Biochem Zool. 2017, 90, 15–33.
- Glickman, N.; Yelon, D. Cardiac development in zebrafish: coordination of form and function. Semin Cell Dev Biol. 2002, 13, 507–513.
- Wood, A. W.; Duan, C.; Bern, H.A. Insulin-like growth factor signaling in fish. Int Rev Cytol. 2005, 243, 215–285.
- Nóbrega, R.H.; Morais, R.D.V.S.; Crespo, D.; De Waal, P.P.; França, L.R.; Schulz, R.W.; Bogerd, J. Fsh stimulates spermatogonial proliferation and differentiation in zebrafish via Igf3. Endocrinology. 2015, 156, 3804–3817.
- Wilson, K. S.; Matrone, G.; Livingstone, D. E. W.; Al-Dujaili, E. A. S.; Mullins, J. J.; Tucker, C.S.; Hadoke, P. W. F.; Kenyon, C. J.; Denvir, M. A. Physiological roles of glucocorticoids during early embryonic development of the zebrafish (Danio rerio). J Physiol. 2013, 591, 6209–6220.
- Cleveland, B.M.; Weber, G.M.; Blemings, K.P.; Silverstein, J.T. Insulin-like growth factor-I and genetic effects on indexes of protein degradation in response to feed deprivation in rainbow trout (Oncorhynchus mykiss). Am J Physiol Regul Integr Comp Physiol. 2009, 297, R1332–R1342.
- Acosta J.; Carpio Y.; Borroto I.; Gonzálezo.; Estradam.P. Myostatin gene silenced by RNAi show a zebrafish giant phenotype. J Biotechnol. 2005, 119, 324–331.
- Lee, C.; Hu, S.; Gong, H.; Chen, M.; Lu, J.; Wu, J. Suppression of myostatin with vector-based RNA interference causes a double-muscle effect in transgenic zebrafish. Biochem Biophys Res Commun. 2009, 387, 766–771.
- Maccatrozzo, L.; Bargelloni, L. ; Cardazzo, B.; Rizzo, G.; Patarnello, T. A novel second myostatin gene is present in teleost fish. FEBS Lett. 2001, 509, 36–40.
- Amali, A.A.; Lin, C. J.-F.; Chen, Y.-H.; Wang, W.-L.; Gong, H.-Y.; Lee, C.-Y.; Wu, J.-L. Up-regulation of muscle-specific transcription factors during embryonic somitogenesis of zebrafish (Danio rerio) by knock-down of myostatin-1. Dev Dyn. 2004, 229, 847–856.
- Gao, Y.; Dai, Z.; Shi, C.; Zhai, G.; Jin, X.; He, J.; Lou, Q.; Yin Z. Depletion of myostatin b promotes somatic growth and lipid metabolism in zebrafish. Front Endocrinol. 2016, 10, 332.
- Wilson, K. S.; Matrone, G.; Livingstone, D. E. W.; Al-Dujaili, E. A. S.; Mullins, J. J.; Tucker, C.S.; Hadoke, P. W. F.; Kenyon, C. J.; Denvir, M. A. Physiological roles of glucocorticoids during early embryonic development of the zebrafish (Danio rerio). J Physiol. 2013, 591, 6209–6220.
- Oakes, J.A.; Li, N.; Wistow, B.R.C.; Griffin, A.; Barnard, L.; Storbeck, K.-H.; Cunliffe, V.T.; Krone, N.P. Ferredoxin 1b deficiency leads to testis disorganization, impaired spermatogenesis, and feminization in zebrafish. Endocrinology. 2019, 160, 2401–2416.
- Keenan, S.; Currie, P. The developmental phases of zebrafish myogenesis. J Dev Biol. 2019, 7, 12.
- Nguyen, P.D.; Gurevich, D.B.; Sonntag, C.; Hersey, L.; Alaei, S.; Nim, H.T.; Siegel, A.; Hall, T.E.; Rossello, F.J.; Boyd, S.E. Muscle stem cells undergo extensive clonal drift during tissue growth via meox1-mediated induction of G2 cell-cycle arrest. Cell Stem Cell. 2017, 21, 107–119.
Figure 1.
xperimental design. (1) Two groups of female zebrafish (n = 30) were assigned into three replicates per experimental group. One group was fed commercial food, while the other one was fed commercial food containing 0.5 mg per feed of hydrocortisone. Zebrafish mothers received thereabout of 25 µg cortisol/ g body mass. Fish were fed twice per day for 5 days. (2) On day 3 post-treatment, five females from each group were transferred to breeding tanks with males untreated. (3) Fertilized eggs were collected in Petri dishes containing embryo medium. (4) Eggs, embryos, and larvae were collected at 0 to 144 -hpf for survival rate, and phenotype characterization such as length, and pericardial edema. The eggs, embryos, and early larvae from 0 to 72 -hpf were counted and pools were collected for gene expression analysis. Embryos at 48 -hpf from the control and cortisol groups were taken to calculate the heartbeat rate per minute. (5) Juveniles at 30 -dpf were also collected for histological analysis of muscle fiber in both groups. Illustration created in https://BioRender.com.
Figure 1.
xperimental design. (1) Two groups of female zebrafish (n = 30) were assigned into three replicates per experimental group. One group was fed commercial food, while the other one was fed commercial food containing 0.5 mg per feed of hydrocortisone. Zebrafish mothers received thereabout of 25 µg cortisol/ g body mass. Fish were fed twice per day for 5 days. (2) On day 3 post-treatment, five females from each group were transferred to breeding tanks with males untreated. (3) Fertilized eggs were collected in Petri dishes containing embryo medium. (4) Eggs, embryos, and larvae were collected at 0 to 144 -hpf for survival rate, and phenotype characterization such as length, and pericardial edema. The eggs, embryos, and early larvae from 0 to 72 -hpf were counted and pools were collected for gene expression analysis. Embryos at 48 -hpf from the control and cortisol groups were taken to calculate the heartbeat rate per minute. (5) Juveniles at 30 -dpf were also collected for histological analysis of muscle fiber in both groups. Illustration created in https://BioRender.com.
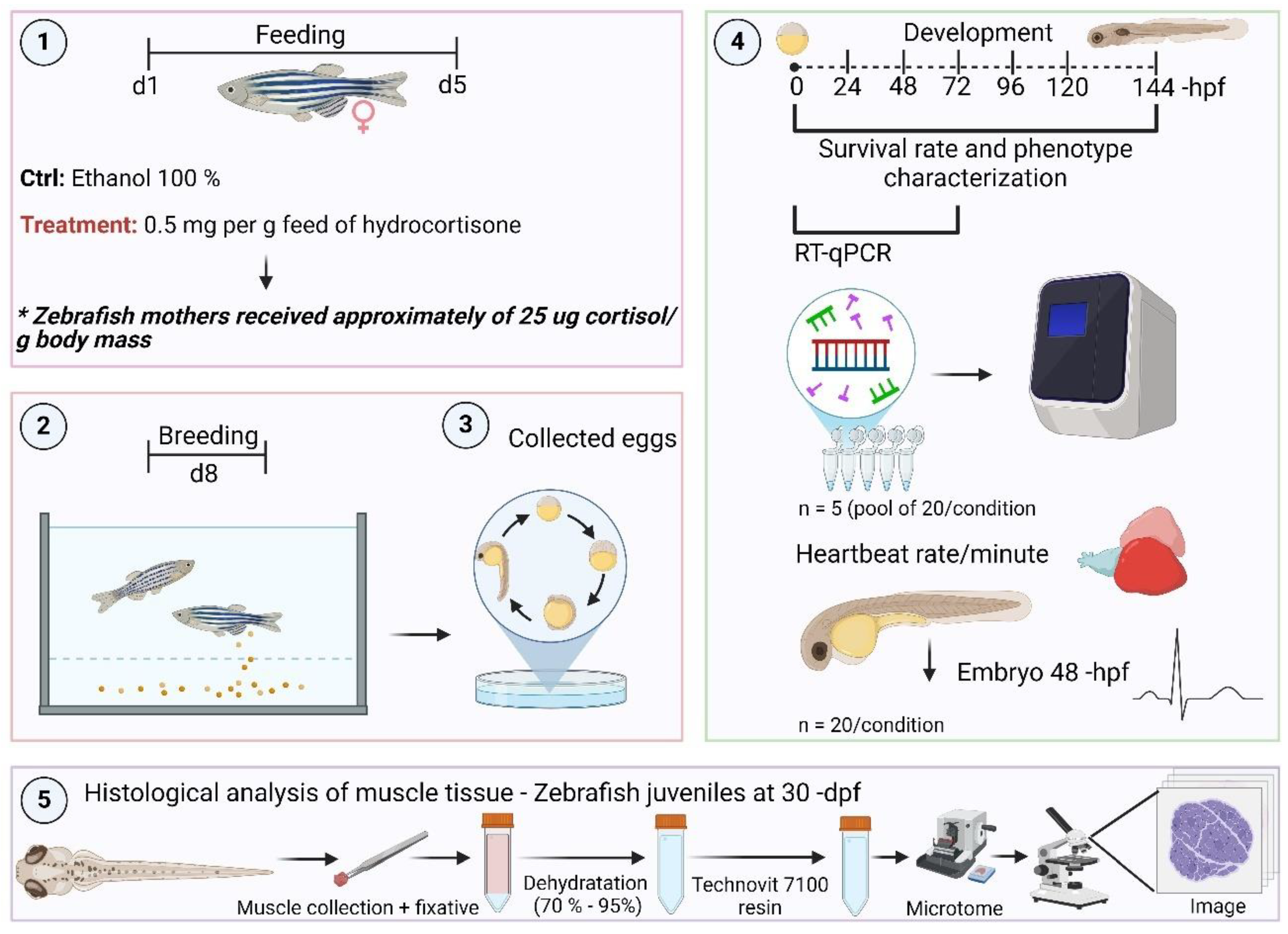
Figure 2.
Somatotropic and HPI-related genes in embryos and early larvae at 0 to 72 -hpf from mothers fed with hydrocortisone enriched in the diet (0.5 mg per g feed) compared to the untreated group (control) (dotted black line set at 1). The selected genes mtor (mammalian target of rapamycin) (A); foxo3a (forkhead box O3) (B); mafbx (muscle atrophy F-box) (C); murf1 (muscle ring-finger protein-1) (D); mstna (myostatin) (E); gh (growth hormone) (F); igf1 (insulin-like growth factor 1) (G); igf2a (insulin-like growth factor 2a) (H); igf2b (insulin-like growth factor 2b) (I); fkbp5 (glucocorticoid receptor chaperone protein) (J); and 11hsdb2 (hydroxysteroid 11-beta dehydrogenase 2) (K) were evaluated. Ct values were normalized with β-actin and expressed as relative values of control (no treated fish) levels of expression. Bars represent the mean ± SEM fold change (n = 5 pools of 20 individuals for each pool) relative to the control. Student unpaired t-test, *p < 0.05, **p < 0.01, ***p < 0.001. NS not significant.
Figure 2.
Somatotropic and HPI-related genes in embryos and early larvae at 0 to 72 -hpf from mothers fed with hydrocortisone enriched in the diet (0.5 mg per g feed) compared to the untreated group (control) (dotted black line set at 1). The selected genes mtor (mammalian target of rapamycin) (A); foxo3a (forkhead box O3) (B); mafbx (muscle atrophy F-box) (C); murf1 (muscle ring-finger protein-1) (D); mstna (myostatin) (E); gh (growth hormone) (F); igf1 (insulin-like growth factor 1) (G); igf2a (insulin-like growth factor 2a) (H); igf2b (insulin-like growth factor 2b) (I); fkbp5 (glucocorticoid receptor chaperone protein) (J); and 11hsdb2 (hydroxysteroid 11-beta dehydrogenase 2) (K) were evaluated. Ct values were normalized with β-actin and expressed as relative values of control (no treated fish) levels of expression. Bars represent the mean ± SEM fold change (n = 5 pools of 20 individuals for each pool) relative to the control. Student unpaired t-test, *p < 0.05, **p < 0.01, ***p < 0.001. NS not significant.
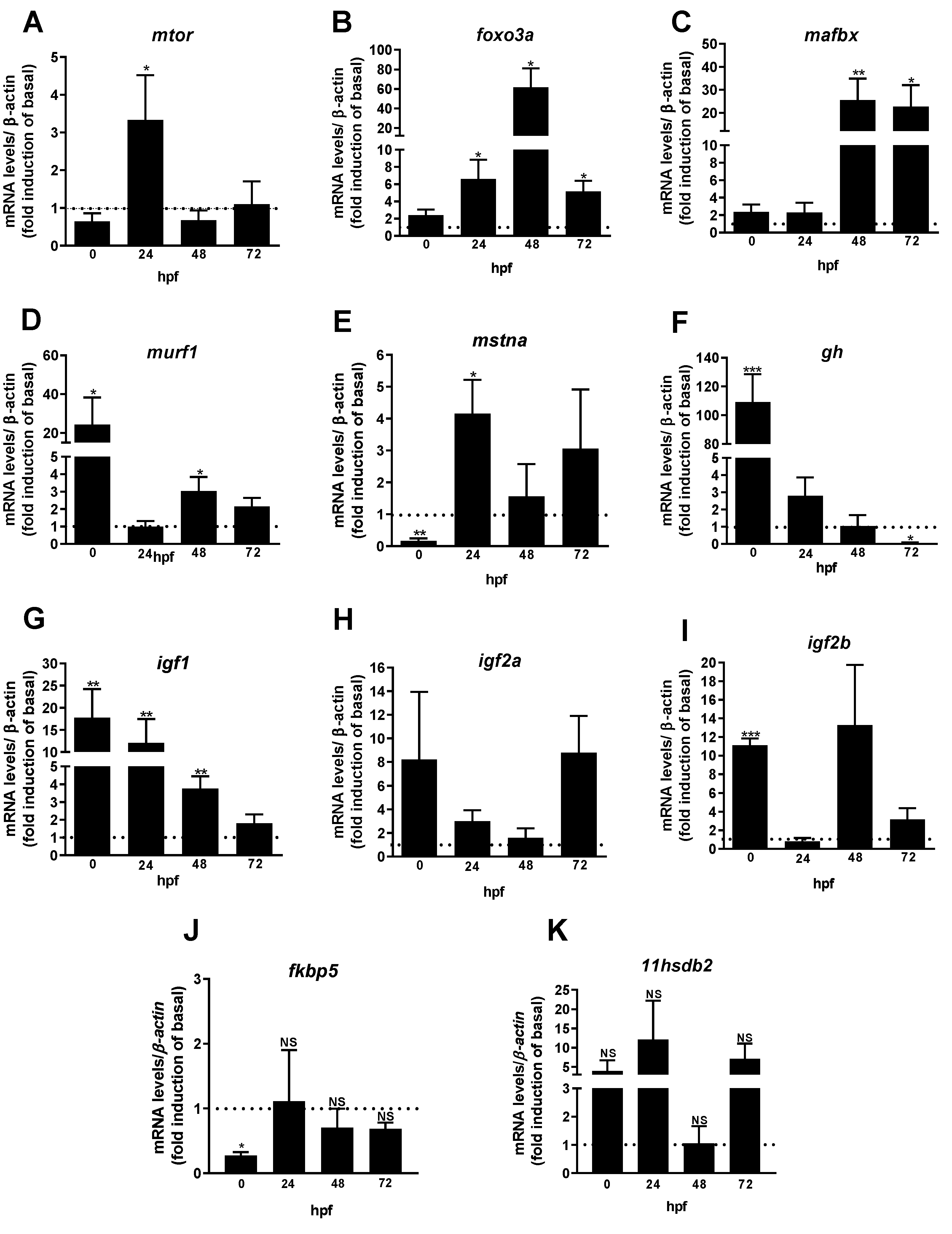
Figure 3.
The survival rate of embryos and larvae at 0 to 144 -hpf (A) was demonstrated in both control and treated animals. Embryos and larvae length at 48 to 72 -hpf (B) were measured (Bars represent the mean ± SEM; ANOVA followed by Tukey’s multiple comparison tests or Student unpaired t-test. Distinct letters denote significant differences; p ≤ 0.05). The phenotype of embryos and early larvae from control females (C-D) compared with those of progeny impacted by maternal transfer of corticosteroid (E-F). Control animals showed normal morphology of pericardium (C-D). However, embryos from maternal cortisol-treated were affected. Arrows highlight the presence of pericardial edema (E-F). Scale bar = 500 µm (n = 30, basal and treatment).
Figure 3.
The survival rate of embryos and larvae at 0 to 144 -hpf (A) was demonstrated in both control and treated animals. Embryos and larvae length at 48 to 72 -hpf (B) were measured (Bars represent the mean ± SEM; ANOVA followed by Tukey’s multiple comparison tests or Student unpaired t-test. Distinct letters denote significant differences; p ≤ 0.05). The phenotype of embryos and early larvae from control females (C-D) compared with those of progeny impacted by maternal transfer of corticosteroid (E-F). Control animals showed normal morphology of pericardium (C-D). However, embryos from maternal cortisol-treated were affected. Arrows highlight the presence of pericardial edema (E-F). Scale bar = 500 µm (n = 30, basal and treatment).
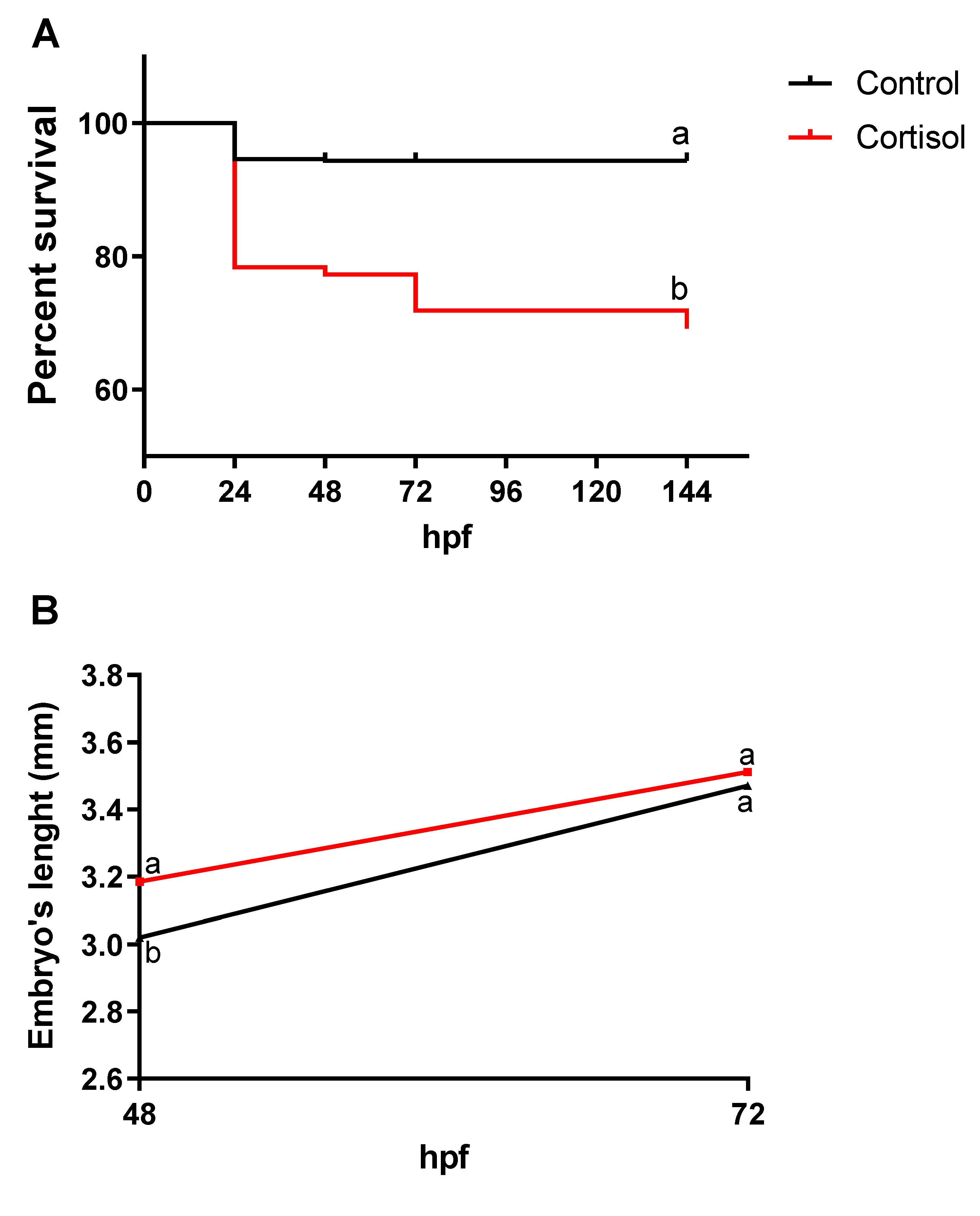
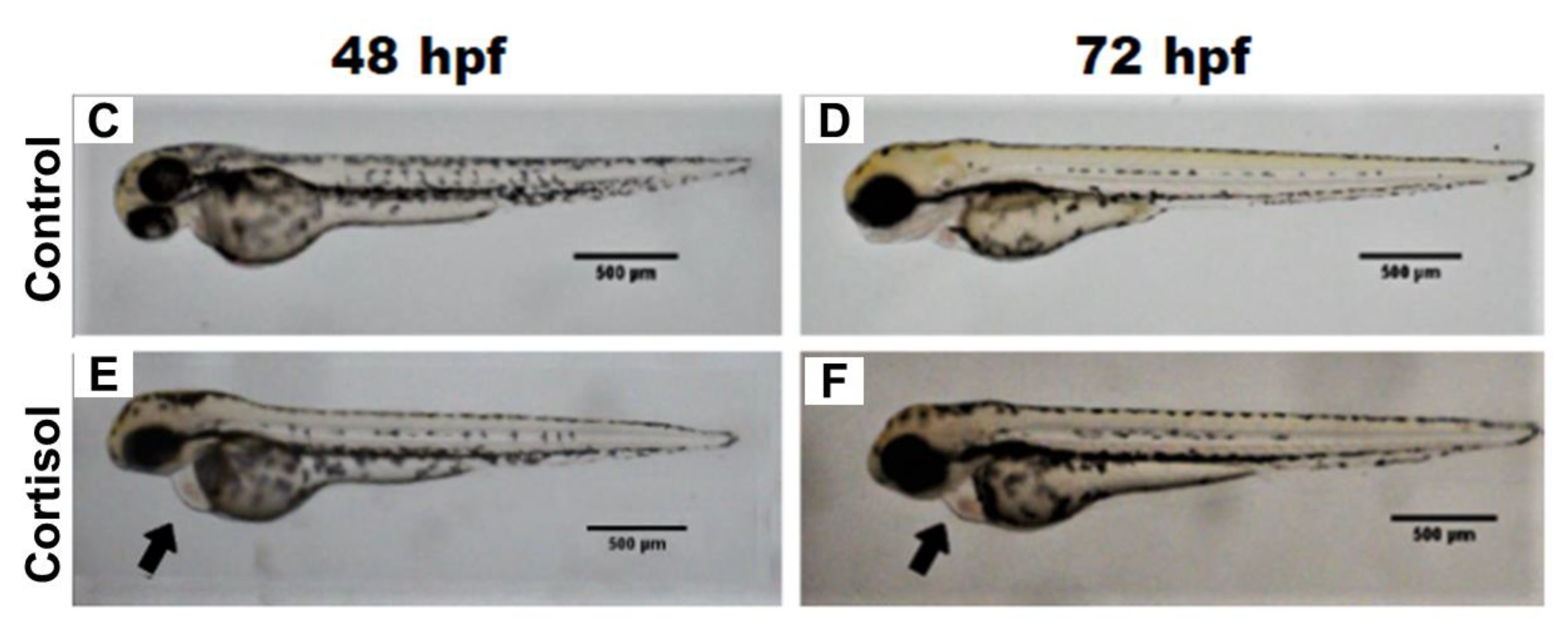
Figure 4.
The heartbeat of embryos with 48 -hpf from control (red circle) and maternal cortisol treatment (blue circle) was demonstrated. No significant heartbeat difference was found between control and treatment embryos. Student unpaired t-test, p = 0.1459.
Figure 4.
The heartbeat of embryos with 48 -hpf from control (red circle) and maternal cortisol treatment (blue circle) was demonstrated. No significant heartbeat difference was found between control and treatment embryos. Student unpaired t-test, p = 0.1459.
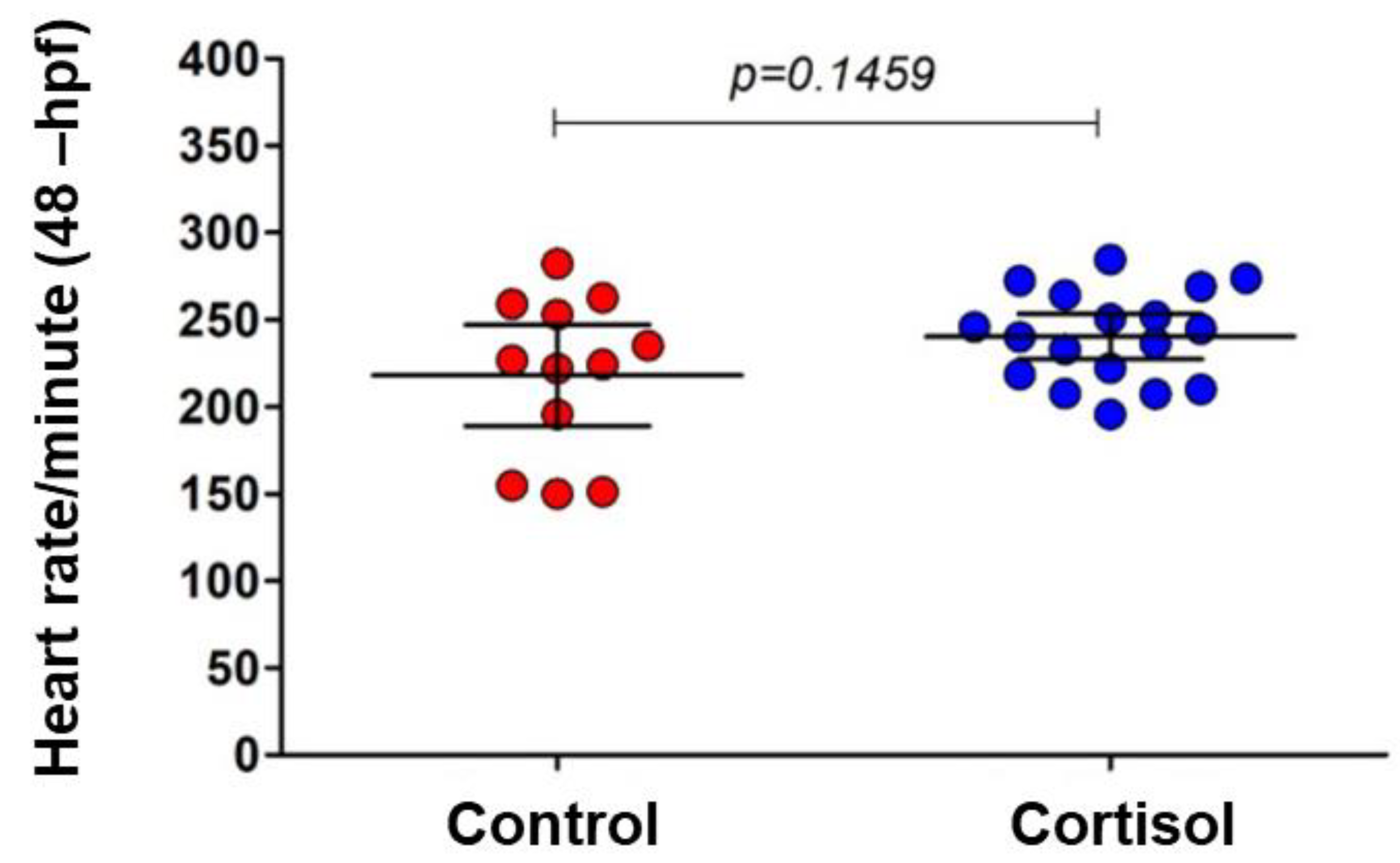
Figure 5.
Longitudinal histological section of muscle from 30 -dpf zebrafish juvenile from females fed with a commercial diet (A) or a diet supplemented with cortisol (B). Muscle bundle length in 30 -dpf from juvenile from control (black) and cortisol-treated (red) females (C). (Bars sampled in each group represent the mean ± SEM; Student unpaired t-test, *p < 0.05). Scale bar = 100 µm. Staining: Toluidine blue (n = 3, basal and treatment).
Figure 5.
Longitudinal histological section of muscle from 30 -dpf zebrafish juvenile from females fed with a commercial diet (A) or a diet supplemented with cortisol (B). Muscle bundle length in 30 -dpf from juvenile from control (black) and cortisol-treated (red) females (C). (Bars sampled in each group represent the mean ± SEM; Student unpaired t-test, *p < 0.05). Scale bar = 100 µm. Staining: Toluidine blue (n = 3, basal and treatment).

Figure 6.
Histological cross-section of zebrafish muscle juvenile at 30 -dpf from females fed a commercial diet (control) (A, C) or a diet supplemented with cortisol (B, D). The cross-sectional diameter of muscle fiber in both progeny from non-treated and cortisol-treated females (E). The proportion of thin (< 9 µm) and thick (> 9 µm) muscle fiber in zebrafish juveniles at 30 -dpf from control and cortisol-treated groups. (Symbols or bars represent the mean ± SEM; Student unpaired t-test, *p < 0.05; ***p < 0.001). Scale bar = 25 µm (A-B); 10 µm (C-D). Staining: Toluidine blue (n = 5; 75 muscle fiber per animal, basal and treatment).
Figure 6.
Histological cross-section of zebrafish muscle juvenile at 30 -dpf from females fed a commercial diet (control) (A, C) or a diet supplemented with cortisol (B, D). The cross-sectional diameter of muscle fiber in both progeny from non-treated and cortisol-treated females (E). The proportion of thin (< 9 µm) and thick (> 9 µm) muscle fiber in zebrafish juveniles at 30 -dpf from control and cortisol-treated groups. (Symbols or bars represent the mean ± SEM; Student unpaired t-test, *p < 0.05; ***p < 0.001). Scale bar = 25 µm (A-B); 10 µm (C-D). Staining: Toluidine blue (n = 5; 75 muscle fiber per animal, basal and treatment).

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



