
Submitted:
03 July 2025
Posted:
04 July 2025
You are already at the latest version
Abstract
Background/objectives: Proteus mirabilis is a frequent causative agent of urinary and wound in-fections in both community and hospital setting. It develops resistance to expanded-spectrum cephalosporins (ESC) due to the production of extended-spectrum β-lactamases (ESBLs) or plasmid-mediated AmpC β-lactamases (p-AmpC). Recently, carbapenem resistant isolates of P. mirabilis emerged due to production of carbapenemases, mostly belonging to Ambler class B and D. Here we report an outbreak of infections due to carbapenem-resistant P. mirabilis ob-served in a psychiatric hospital in Zagreb, Croatia. The characteristics of ESBL and car-bapenemase produced by P. mirabilis associated with an outbreak were analyzed. Material and methods: The antibiotic susceptibility testing was performed by disk-diffusion and broth dilution method. Double disk synergy test (DDST) and inhibitor- based test with clavulanic and phenylboronic acid were applied to screen for ESBLs and p-AmpC, respectively. Carbapenemases were screened by modified Hodge test (MHT) while carbapenem hydrolysis was investigated by CIM and eCIM test. The nature of ESBL, carbapenemases, and fluoroquin-olone resistance determinants was investigated by PCR. Transferability of cefotaxime and imipenem resistance was determined by conjugation. Plasmids were characterized by PCR-based replicon typing. Selected isolates were subjected to molecular characterization of re-sistome by the Interarray Carba Resist Kit and whole genome sequencing (WGS) Results: In total, 20 isolates were collected and analyzed. All isolates exhibited resistance to amoxicillin alone and combined with clavulanic acid, cefuroxime, ceftazidime, cefotaxime, ceftriaxone, cefepime, imipenem, ceftazidime-avibactam, ceftolozane-tazobactam, gentamicin, amikacin and ciprofloxacin. There was uniform susceptibility to ertapenem, meropenem and cefiderocol. DDST and combined disk test with clavulanic acid tested positive indicating pro-duction of an ESBL. MHT was negative in all isolates while CIM showed moderate sensitivity but only with imipenem as indicator disk. E-CIM tested positive in all CIM positive isolates, consistent with an MBL. PCR and sequencing of selected amplicons identified VIM-1 and VIM-4. The Interarray Carba Resist Kit and whole genome sequencing (WGS) identified β-lactam re-sistance genes: blaVIM, blaCTX-M-15, and blaTEM genes, aminoglycoside resistance genes: aac(3)-IId, aph(6)-Id, aph(3'')-Ib, aadA1, armA, and aac(6')-IIc, and resistance genes for sulphonamides sul1 and sul2, trimethoprim dfr1, chloramfenicol cat and tetracycline tetJ. Conclusions: The study revealed epidemic spread of carbapenemase producing P. mirabilis in two wards in a psychiatric hospital. This is the first report of VIM producing P. mirabilis in Croatia.
Keywords:
Proteus mirabilis
; multidrug resistance
; VIM
; CTX-M-15
; epidemic spread
1. Introduction
Proteus mirabilis is an opportunistic pathogen belonging to the family Enterobacterales, widely distributed in the environment and as a normal microbiota in the bowel of human and animals [1]. Along with other Enteroacterales, it is considered to be an important causative agent of urinary tract infections (UTI), particularly catheter associated urinary tract infections (CAUTI) due to its ability to form biofilm and wound infections [2]. P. mirabilis is intrinsically resistant to tigecycline, polymixins, tetracyclines and nitrofurantoin [3], but does not have any intrinsic β-lactamase.
Resistance to β-lactams in P. mirabilis is mediated by β-lactamases, enzymes which cleave β-lactam antibiotics, modification of PBR receptors, upregulation of efflux pumps and porin alteration or loss. The β-lactamases encountered in Proteus species are broad spectrum penicillinases (TEM-1, TEM-2, SHV-1, CARB, OXA-1-4), extended-spectrum β-lactamases (ESBLs), plasmid-mediated AmpC β-lactamase (p-AmpC) and recently carbapenemases [3,4,5,6]. ESBLs hydrolyze penicillins, all cephalosporins and monobactams, whereas p-Amp inactivate expanded-spectrum cephalosporins (ceftazidime, cefoaxime, ceftriaxone) and cephamycins, but spare carbapenems and cefepime, Carbapenems are potent β-lactam antibiotics and the last resort treatment for infections due ESBL and AmpC producing organisms [3]. Carbapenemases encountered in P. mirablis belong to class B or metallo-β-lactamases (MBLs) (VIM, IMP, NDM), class D or carbapenem hydrolyzing oxacillinases or CHDL (OXA-48, OXA-181, OXA-23, OXA-58) designated as CHDL and rarely A (KPC) [3,7]. The resistance to aminoglycosides in P. mirabilis is most frequently caused by enzymes which modify aminoglycosides and render them inactive. Adenyltransferases are encoded by aadA1 and aad2, acetyltransferases by aac(6'')-Ib and aacA4 genes and phosphorylases by aph(6)-Id, aph(6)-Ib and aph(6)-Ia) genes [3]. Panaminoglycoside resistance associated with 16S rRNA methylase was also reported in P. mirabilis [3]. P. mirabilis develops resistance to fluoroqinolones by mutation of the genes encoding DNA gyrase (gyrA) and topoisomerase (parC) or acquisition of plasmid-mediated qnr genes encoding qnr proteins which protect topoisomerase IV. Sul1 and sul2 genes usually confer resistance to sulphonamides while dfr genes encode dihydropholate reductase antagonizing the effect of trimethoprim [3].
Bibliographical data on β-lactam resistance mechanisms in P. mirabilis are mostly focused on ESBLs and p-AmpC. ESBLs belonged most frequently to TEM family in early 90-ties, but a shift to CTX-M was reported at the beginning of 2000-ties [3]. CMY is almost the only p-AmpC family reported in P. mirabilis [3]. In Croatia, an outbreak of infections with TEM-52 β-lactamase producing P. mirabilis was reported in Split in 2008 [8,9]. Later, an outbreak of infections with CMY-16 was described in a nursing home in Zagreb [10]. The same allelic variant was much later identified in a hospital in Split [11]. Intensive use of carbapenems in the treatment of infections with ESBL and p-AmpC producing organisms lead to proliferation of carbapenemases in P. mirabilis. The majority of studies detected carbapenemases belonging to VIM and NDM family and OXA-48 [3]. However, there are no reports addressing carbapenemases in this species from Croatia.
Here we report extensively drug-resistant carbapenemase producing P. mirabilis associated with an outbreak in a psychiatric hospital in Zagreb. We aimed to characterize β-lactamases and other resistance mechanisms carried by these isolates and their molecular epidemiology.
2. Materials and Methods
2.1. Patients and Bacterial Isolates
The bacterial isolates with reduced susceptibility to at least one carbapenem (imipenem, meropenem, ertapenem) were collected from 2023 to 2024 from a psychiatric hospital in Zagreb. The isolates were identified to a species level by MALDI-TOF MS (matrix-assisted laser desorption ionization–time of flight mass spectrometry) (Vitek MS, bioMerieux, France). The initial antibiotic susceptibility was tested according to EUCAST guidelines [12] in the Dr. Andrija Štampar Teaching Institute of Public Health which provides microbiological service for Psychiatric Hospital “Sveti Ivan”. Isolates which demonstrated reduced susceptibility to at least one carbapenem were sent to the Clinical Department for Clinical Microbiology and Infection Control and Prevention of the University Hospital Centre Zagreb for further analysis.
2.2. Antimicrobial Susceptibility Testing
Broth dilution method was applied to determine the minimum-inhibitory concentrations (MICs) of 13 antibiotic: amoxycillin alone and with clavulanic acid, piperacillin-tazobactam, cefuroxime, ceftazidime, cefotaxime, ceftriaxone, cefepime, imipenem, meropenem, gentamicin, amikacin and ciprofloxacin. The results were interpreted according to CLSI guidelines [13]. The susceptibility to ceftazidime-avibactam, sulphametoxazole-trimethoprim and chloramphenicol was determined only by disk-diffusion test. The antibiotic containing disks were provided by Oxoid (Basingstoke, UK). The isolates were classified as multidrug-resistant (MDR), extensively-drug resistant (XDR) or pandrug-resistant (PDR) as described previously by Magiorakos et al [14]. MDR is recognized as being non-susceptible to at least one antimicrobial agent in three antimicrobial classes. XDR is recognized as being resistant to at least one antimicrobial agent in all antimicrobial classes and susceptible to only two or less. PDR isolates are resistant to all antibiotics available for the specific species. Escherichia coli ATCC 25922 and Klebsiella pneumoniae 700603 served as quality control strains for MIC determination.
2.3. Phenotypic Detection of β-Lactamases
ESBLs were detected by a double disk- synergy test [15] and combined disk test with cephalosporins and clavulanic acid [13]. Plasmid-mediated AmpC β-lactamases were detected among cefoxitin resistant isolates by combined disk test using cephalosporin disks combined with cloxacllin [6]. Initial screening for carbapenemases was carried out for the purpose of routine microbiology testing by immunochromatographic test [16]. A modified Hodge test (MHT) with imipenem disk was used to confirm the production of carbapenemases [17]. Since identification of carbapenemases in P.mirabilis poses an identification challenge due to low level of resistance in this species, additional tests were done. Carbapenem inactivation method (CIM) was done to assess hydrolysis of carbapenems with disks of imipenem and meropenem [18]. A suspension of the test strain was adjusted to McFarland 0.5 (108 CFU/mL) and a meropenem disk was placed in the suspension. The suspension was incubated for 2 h at 37 °C. E. coli ATCC 25922 was inoculated on Mueller–Hinton agar (MH) and the disk was placed in the middle of the plate. The plates were incubated overnight and the lack of inhibition zone, decreased inhibition zone (<15 mm) or colonies within the inhibition zone indicated carbapenem hydrolysis. The isolates positive in CIM test were subjected to eCIM for detection of MBLs. The eCIM test was performed in the same way, but with one tube containing 0.5 mM EDTA to inhibit MBLs. The test indicated MBL positivity if there was ≥5 mm increase in zone diameter in eCIM experiment compared to the control sample without EDTA [18]. The strains from own collection, known to be positive for KPC, VIM, NDM and OXA-48, were used as positive and negative controls [18]. Since there was a high rate of negativity in eCIM among isolates positive in immunochromatographic method, they were additionally tested by combined disk tests with imipenem and meropenem alone and combined with PBA, 0.1 M EDTA, or both to detect for KPC, MBLs, or simultaneous production of KPC and MBL, respectively [17].
2.4. Molecular Detection of Resistance Genes
DNA was extracted by thermal lysis at 100 C for 10 minutes. Cellular debris was removed by centrifugation and the supernatant was used as DNA template. Antimicrobial resistance genes comprising β-lactam (blaTEM, blaSHV, blaCTX-M) [19,20,21] and fluoroquinolones (qnrA, qnrB, qnrS) [22], were determined by singleplex PCR as described previously. Carbapenemases coding genes belonging to class A (blaKPC), B or MBLs (blaIMP, blaVIM, blaNDM) and class D (blaOXA-48), [23], blaOXA-23, blaOXA-24/40, blaOXA-51, blaOXA-58) [24] blaampC [25] and the genes of five clonal groups of CTX-M β-lactamases were amplified via multiplex PCR according to the protocols reported previously [26]. The randomly selected amplicons (11 CTX-M and 8 VIM) obtained by singleplex PCR were sequenced with forward and reverse using Eurofin Genomic service Eurofins Genomics (https://eurofingenomics.eu) and the sequences were compared with those from the GenBank nucleotide database at http://www.ncbi.nlm. nih.gov/blast (accessed on 1st February 2025. The flanking regions of blaCTX-M genes were investigated by PCR mapping, using forward primer for ISEcp combined with universal reverse primer for CTX-M encoding genes (MA3) [27].
2.5. CarbaResist Inter-Array Genotyping Kit
Genotyping of six randomly selected isolates was conducted using the microarray-based CarbaResist Genotyping Kit, according to the manufacturer’s instructions, version 1012012100004 (INTER-ARRAY, fzmb GmbH, Bad Langensalza, Germany). In short, genomic DNA was isolated from monoclonal overnight cultures using the Qiagen DNeasy Blood and Tissue Kit, according to the manual. The unfragmented DNA was linear amplified using one primer for each target sequence (antisense) and internally labeled with biotin dUTP. The obtained ssDNA (single-stranded) products were transferred into the ArrayWells for hybridization. These wells contain 230 probes corresponding to distinct genes for the most relevant carbapenemases, ESBL and AmpC, as well as genes associated with β-lactam-, aminoglycoside-, fluoroquinolone-, sulphonamide-, trimethoprim- and colistin-resistance. After washing steps to remove any unbound DNA, horseradish peroxidase (HRP)-conjugated streptavidin was bounded to all hybridized sections, resulting in dark spots on the chip due to an enzymatic reaction. The detection of these spots and data acquisition was performed automatically using the INTER-VISION Reader.
2.6. Whole Genome Sequencing (WGS)
Four representative strains were subjected to WGS. First, the strains were cultivated in Tryptic Soy Broth (Merck Millipore, MA, USA) at 37 °C overnight. Then, the genomic DNA was extracted using the QIAamp DNA Mini Kit (Qiagen, Hilden, Germany) according to the manufacturer’s instructions. Subsequently, the extracts were barcoded via the Rapid Barcoding Kit 96 V14 and sequenced using the minION (Oxford Nanopore Technologies). For the assembly of the single reads Flye (https://www.pnas.org/doi/full/10.1073/pnas.1604560113, accessed on 20 June 2025) was utilized. Subsequently, the data was analyzed using the webservers and services of the Center for Genomic Epidemiology (http://www.genomicepidemiology.org, accessed on 20 June 2025) [48]. The phylogenetic tree was generated with REALPHY online tool (https://doi.org/10.1093/molbev/msu088, accessed on 20 June, 2025) and visualized using phylo.io (https://doi.org/10.1093/molbev/msw080, accessed on 20 June 2025).
2.7. Characterization of Plasmids
PCR-based replicon typing (PBRT) was according to Carattoli et al. [29]. Since it was observed previously that PBRT can be inefficient in identifying L/M plasmid incompatibility type, an updated method designated to identify and distinguish between IncL and IncM plasmids was applied [30]. Positive control strains were kindly provided by Dr. Alessandra Carattoli, Instituto Superiore di Sanita, Rome, Italy.
2.8. Detection of Virulence Determinants
Urease activity was determined by inoculating the strains in the urea-containing medium. The change of the color to pink was recorded as a positive result. The haemolytic activity was tested by culturing the strains on a 10% sheep blood plate with the addition of trimethoprim to inhibit swarming. For the motility assay, one colony was stabbed with a one-microliter loop into a semisolid nutrient medium. After overnight incubation at 37 °C, motility was measured as turbidity of the medium [2].
2.9. Genotyping
Phylogenetic tree was constracted using WGS.
3. Results
3.1. Patients and Bacterial Isolates
In total 20 isolates with reduced susceptibility to carbapenems were collected from November 3rd 2023 until 24th July 2024. All isolates originated from urinary tract and all but two from urinary catheters (Supplementary Material Table S1). All patients were females with age ranging from 49-90 years (median-79)
3.2. Antibiotic Susceptibility
Antimicrobial susceptibility results for the 20 P. mirabilis isolates are listed in Table 1. They all exhibited identical resistance profile with MICs of amoxycillin alone and combined with clavulanic acid, cefuroxime, cefotaxime, ceftriaxone, cefepime, gentamicin, amikacin and ciprofloxacin exceeding 128 mg/L in the majority of isolates. Regarding carbapenems, isolates displayed reduced susceptiblity towards imipenem with variable MIC values ranging from to 4 to 128 mg/L, but susceptiblity to meropenem and ertapenem. High resistance rate of 90% (18/20) was observed for ceftazidime. (Table 1) Piperacillin-tazobactam retained activity with all isolates being susceptible according to CLSI with MIC ≤ 16 mg/L . However, if EUCAST criteria were applied with lower resistance breakpoint (16 mg/L), 35% (n=7) of the isolates would be resistant. On the other hand, according to EUCAST there would be only four isolates resistant to imipenem (20%). All but one isolate showed susceptibility to cefiderocol. All isolates were classified as XDR. Meropenem, ertapenem and cefiderocol exerted high activity with MICs of meropenem below 0,5 mg/L.
3.3. Phenotypic Detection of β-Lactamases
Based on DDST using clavulanic acid and combined disk-test all isolates were ESBL positive. All isolates were susceptibile to cefoxitin, indicating the lack of p-AmpC. Regarding carbapenemases, Hodge test was negative in all but one isolate (95%) as no distortion of the inhibition zone was seen. Furthermore, CIM and e IM were positive in 80% (n=16), indicating production of an MBL, but only with imipenem disk. However, inhibitor based disk test with EDTA yielded positive result in 70% (n=14) again only with imipenem disk, rasing suspicion of an MBL.
3.4. Molecular Detection of Resistance Genes
PCR revealed blaCTX-M cluster 1 and blaVIM in all tested isolates. Sequencing of eight randomly selected VIM amplicons identified one blaVIM-1, blaVIM-4, and blaVIM-78 genes, respectively and five blaVIM-78 genes.All 11 CTX-M amplicons revealed CTX-M-15.
3.5. Genotpying by Interarray CarbaResist Kit
The genomes of all tested isolates possessed the identical resistance gene content including blaTEM, blaCTX-M-9 gene associated with the ISEcp insertion element and blaVIM providing β-lactam resistance, aac(6)-Iic, aadA1 and aadA2 and armA coding enyzmes for aminoglycoside modification, sul1 and sul2 for sulphonamide resistance and dfrA1 for dihydrofolate reductase, rendering trimethoprim resistant. One isolate harboured dfrA15 allelic variant. All isolates harboured integrase genes for class 1 and class 2 integrons.

Table 2.
Resistome of P. mirabilis isolates by Interarray CarbaResist Kit.
| Isolate and Protocol Number | Res phenotype | β-Lactam | Aminoglycoside | sulphonami | trimethoprim | Integrase genes |
| PM 2 | AMX, AMC, TZP, CAZ,CTX, CRO, FEP, GM,AMI CIP |
ISEcpblaCTX-M-15 blaTEM blaVIM |
aac(6′’)IIc aadA1 aadA2 armA |
sul1 |
dfrA1 |
Intl2 |
| PM 3 | AMX, AMC, TZP, CAZ,CTX, CRO, FEP, GM,AMI CIP |
ISEcpblaCTX-M-15 blaCMY blaTEM blaVIM |
aac(6′’)IIc aadA1 aadA2 armA |
sul1 sul2 |
dfrA1 |
Intl1 Intl2 |
|
PM4 |
AMX, AMC, TZP, CAZ,CTX, CRO, FEP, GM,AMI CIP |
ISEcpblaCTX-M-15 blaTEM blaVIM |
aac(6′’)IIc aadA1 aadA2 armA |
sul1 sul2 |
dfrA1 dfrA15 |
Intl1 Intl2 |
| PM11 | AMX, AMC, TZP, CAZ,CTX, CRO, FEP, GM,AMI CIP |
ISEcpblaCTX-M-15 blaTEM blaVIM |
aac(6′’)IIc aadA1 aadA2 armA |
sul1 sul2 |
dfrA1 |
Intl1 Intl2 |
| PM14 | AMX, AMC, TZP, CAZ,CTX, CRO, FEP, GM,AMI CIP |
ISEcpblaCTX-M-15 blaTEM blaVIM |
aac(6′’)IIc aadA1 aadA2 armA |
sul1 sul2 |
dfrA1 |
Intl1 Intl2 |
| PM 19 | AMX, AMC, TZP, CAZ,CTX, CRO, FEP, GM,AMI CIP |
ISEcpblaCTX-M-15 blaTEM blaVIM |
aac(6′’)IIc aadA1 aadA2 armA |
sul1 sul2 |
dfrA1 |
Intl1 Intl2 |
| PM 20 | AMX, AMC, TZP, CAZ,CTX, CRO, FEP, GM,AMI CIP |
ISEcpblaCTX-M-15 blaTEM blaVIM |
aac(6′’)IIc aadA1 aadA2 armA |
sul1 sul2 |
dfrA1 |
Intl1 Intl2 |
3.6. WGS
All four tested isolates presented identical antimicrobial resistance genes (AMR): blaCTX-M-202 blaTEM-2, blaTEM-1A, blaVIM-4 for β-lactam resistance, aac(3)-IId, aph(6)-Id, aph(3'')-Ib, aadA1, armA, aac(6')-IIc for aminoglycoside resistance, sul1 and sul2 for sulphonamide resistance, dfrA1 encoding dihydropholate reductase, cat for chloraphenicol acetyltransferase and tet(J) responsible for tetracycline resistance as shown in Table 3. Fluoroquinolone resistance genes were not detected.
3.7. Plasmid Analysis
There we not typable plasmids found by multiplex PCR but WGS detected IncQ1 plasmid in representative isolates.
3.8. Detection of Virulence Determinants
All isolates were positive for urease activity, hemolysis, and motility.
3.9. Genotyping
Genotyping demonstrated that four representative isolates were clonally related but distinct from ESBL and p-AmpC positive isolates from previous study as shown in Figure 1.
4. Discussion
The study described an outbreak of infections with VIM-1 positive P. mirabilis. We characterized resistance determinants and molecular epidemiology of the isolates. All isolates had identical resistance patterns and genes. The study showed development of resistance of this important hospital pathogen and its ability to accumulate β-lactam resistance determinants. The bibliographical references on MBLs in P. mirabilis are scarce and mostly reported from Far East [31]. In Europe, Greece and Bulgaria are focal points of VIM producing P. mirabilis [32,33,34,35]. Sporadic occurrence was recorded in Germany [36]. Our isolates were resistant or intermediate susceptible to imipenem, but susceptible to ertapenem and meropenem and thus represent a hidden reservoir of VIM-MBL in Enterobacterales.The use of molecular methods and immunochromatographic tests could help to uncover the underestimated carriage of VIM producing P. mirabilis since the isolates remain susceptible to two out of three carbapenems available in Croatia. VIM-4 and VIM-75 allelic variants are reported for the first time in Croatia. They were previously found among XDR P. mirabilis isolates from Germany [37]. Similarly, as in Germany two of our isolates harbored duplicate VIM carbapenemases. In contrast to the wide variety of VIM allelic variants all CTX-M amplicons encoded CTX-M-15, the most widespread allelic variant of CTX-M enyzmes. All isolates were categorized as XDR, as they were susceptible only to meropenem, ertapenem and cefiderocol. However, the resistance phenotype is dependent on which criteria were applied. According to CLSI all isolates were susceptible to piperacillin-tazobactam and resistant to imipenem, but if MICs were interpreted using EUCAST recommendations, all isolates would be resistant to piperacillin-tazobactam and the majority intermediately susceptible to imipenem (susceptible at increased exposure). The isolates exhibited variable level of resistance to expanded-spectrum cephalosporins and carbapenems with MICs of imipenem ranging from 4 to >128 mg/L. This could be due to variable expression of blaVIM genes.
MBL encoding genes were probably embedded in class 1 or class 2 integrons which can be transferred and integrated in the plasmids or chromosomes and are responsible for the spread of gene cassetes with β-lactam, aminoglycosides and sulphonamide resistance genes, similarly as those from Germany [37]. CHDL previously identified only in Acinetobacter baumannii, are dominant in P. mirabilis in France, Belgium and Germany and similarly as MBLs also pose a diagnostic challenge due to high susceptibility to carbapenems [37,38,39]. In the past, Pseudomonas aeruginosa was a typical host for VIM carbapenemases, but later they were frequently detected among carbapenem-resistant Enterobacter cloacae isolates [40]. In contrast to other Enterobacterales, VIM positivity affected only imipenem susceptibility and rendered it resistant. Meropenem and ertapenem remained susceptible. Among Enterobacterales, K. pneumoniae and E. coli are typical hospital pathogens, bearing blaCARB genes, associated with hospital outbreaks. However, P. mirabilis is gaining importance as nosocomial pathogen and outbreaks due to ESBL, p-AmpC and carbapenemase producing isolates occur all over the world. In Croatia, two hospital outbreaks were reported: one with TEM-52 producing P. mirabilis and the other with CMY-16 producing P. mirabilis, both from Split [8,9,11].
CTX-M-15 ESBL was detected as additional β-lactamase to VIM. It was conferring on the producing isolates high-level resistance to third and fourth generation cephalosporins and amoxicillin-clavulanate. This allelic variant has achieved worldwide spread and was described in E. coli and K. pneumoniae from Croatia [41,42]. This is in contrast with the recent study on ESBLs in P. mirabilis from Croatia which identified CTX-M-9 cluster to be dominant with CTX-M-14 as the most frequent variant [43]. ISEcp upstream of blaCTX-M gene acts as promotor and increased the expression of the gene. In spite of very high MICs of aminoglycosides, there were no plasmid-born resistance genes and thus the resistance is most likely due the mutations of gyrA and parCgenes. High-level resistance to aminoglycosides is in line with a plethora of resistance genes encoding acetylases, adenylases and phosphorilases which modify aminoglycosides and render them inactive. The presence of sul, tet and cat genes is in concordance with sulphonamide, tetracycline and chloramphenicol resistance, respectively. Intrestingly, there was only one allelic variant of dfr gene coding for dihydropteroat syntethase, unlike previous studies on ESBLs in P. mirabilis which revealed several allelic variants [43].
Unfortunately, phenotypic tests based on carbapenem hydrolysis proved unreliable. MHT did not detect any of the isolates as carbapenemase producers and CIM exhibited positive results only with imipenem disk. This is due to low expression of blaVIM genes and weak carbapenem hydrolysis. Phenotypic detection of MBLs is mostly based on carbapenem-EDTA synergy test. In the present study we performed two forms of tests: combined-disk test with EDTA and eCIM, but both exerted moderate sensitivity, visible only with imipenem disk. On the contrary, immunochromatographic test showed high concordance with PCR, WGS and Interarry CarbaResist Kit. All four methods detected VIM resistant determinant in all tested isolates. A similar problem was observed with OXA-23 and OXA-58 producing isolates in France and Belgium [38,39]. Apparently, expression of the carbapenemase encoding genes is very weak among Proteus spp. isolates, regardless of the carbapenemase type. Some researchers recommended the use of dipicolinate as metal chelator for Proteus spp. which provides better sensitivity compared to EDTA [43]. However, the compound is not available in the most routine laboratories.
Therapeutic options are very limited also due to the intrinsic resistance to tigecycline and colistin. Fortunately, cefiderocol susceptibility tested positive in all except one strain. There was no resistance recorded to meropenem and ertapenem. However, in case of carbapenem administration, there is a risk of development of mutants hyperproducing VIM β-lactamase under selection pressure.
The fact that all isolates possessed identical resistance genes points towards the common source. The described MDR clone successfully spread within two women's wards of a psychiatric hospital, causing urinary infections in catheterized patients. The hospital epidemic was stopped after the introduction of measures to prevent the spread of the VIM-positive P. mirabilis clone, but after that, urinary infections caused by this clone were detected in two nursing homes in Zagreb, because the patients were transferred from the hospital. Thanks to the implemented prevention measures, the clone did not spread in those two homes.
5. Conclusions
Analysis of the local antimicrobial resistance patterns is necessary in order to avoid hospital outbreaks with difficult to treat isolates. Hospital hygiene measures should be implemented to limit the spread of MDR P. mirabilis.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
B.B.: laboratory analysis of the isolates, manuscript preparation, conceptualization: M.A., V.T.. and J.V.: data collection. M.N.: statistical analysis. V.D., A.G, I.B. J.L, V.D,. and G.Z.: laboratory analysis of the isolates, A.G., J.V.: critical reviewing of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
Grant from the University of Zagreb School of Medicine (Carbapenemases in hospitals, nursing homes and environment, grant number: 380-59-10106-16-2983).
Institutional Review Board Statement
The permission for the study was obtained from the Ethical Commission of the Dr Andrija Štampar Teaching Institute of Public Health, Zagreb, Croatia (Permission No. 641-01/25-01/1).
Informed Consent Statement
Was not required.
Data Availability Statement
The data presented in this study are available on request from the corresponding author. Resistance genes sequences were deposited in the NCBI Gene bank with the following accession numbers: aac(3)-IId- EU022314, aph(6)-Id- M28829, aph(3'')-Ib- AF321551, aadA1- JQ480156, armA- AY220558; aac(6')-IIc- NC_012555; blaCTX-M-202- MF195067; blaTEM-1A- HM749966; blaVIM-4- EU581706; blaVIM-1- Y18050, blaTEM-2- X54606, cat- M11587; sul1- U12338; sul2- FN995456; tet(J)- ACLE01000065; dfrA1- X00926.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Sanches, M.S.; Silva, L.C.; Silva, C.R.D.; Montini, V.H.; Oliva, B.H.D.; Guidone, G.H.M.; Nogueira, M.C.L.; Menck-Costa, M.F.; Kobayashi, R.K.T.; Vespero, E.C.; Rocha, S.P.D. Prevalence of Antimicrobial Resistance and Clonal Relationship in ESBL/AmpC-Producing Proteus mirabilis Isolated from Meat Products and Community-Acquired Urinary Tract Infection (UTI-CA) in Southern Brazil. Antibiotics (Basel). 2023, 2023 10, 370. [Google Scholar] [CrossRef]
- Li, Y.; Yin, M.; Fang, C.; Fu, Y.; Dai, X.; Zeng, W.; Zhang, L. Genetic analysis of resistance and virulence characteristics of clinical multidrug-resistant Proteus mirabilis isolates. Front. Cell. Infect. Microbiol. 2023, 11, 1229194. [Google Scholar] [CrossRef] [PubMed]
- Girlich, D.; Bonnin, R.A.; Dortet, L.; Naas, T. Genetics of Acquired Antibiotic Resistance Genes in Proteus spp. Front Microbiol. Front Microbiol. 2020, 21, 256. [Google Scholar] [CrossRef]
- Paterson, D.L.; Bonomo, R.A. Extended-spectrum β-lactamases: A clinical update. Clin. Microbiol. Rev. 2005, 18, 657–686. [Google Scholar] [CrossRef]
- Cantón, R.; Coque, T.M. The CTX-M β-lactamase pandemic. Curr. Opin. Microbiol. 2006, 9, 466–475. [Google Scholar] [CrossRef] [PubMed]
- Jacoby, G.A. AmpC β-lactamases. J. Clin. Microbiol. 2009, 22, 161–182. [Google Scholar] [CrossRef]
- Potter RF, D'Souza AW, Dantas G. The rapid spread of carbapenem-resistant Enterobacteriaceae. Drug Resist Updat. 2016, 29, 30–46. [CrossRef] [PubMed] [PubMed Central]
- Sardelić, S.; Bedenić, B.; Sijak, D.; Colinon, C.; Kalenić, S. Emergence of Proteus mirabilis isolates producing TEM-52 extended-spectrum β-lactamases in Croatia. Chemotherapy. 2010, 56, 208–13. [Google Scholar] [CrossRef]
- Tonkić, M.; Mohar, B.; Šiško-Kraljević, K.; Meško-Meglič, K.; Goić-Barišić, I.; Novak, A.; Kovačić, A.; Punda-Polić, V. High prevalence and molecular characterization of extended-spectrum β-lactamase-producing Proteus mirabilis strains in southern Croatia. J Med Microbiol. 2010, 59, 1185–1190. [Google Scholar] [CrossRef]
- Bedenić, B.; Firis, N.; Elveđi-Gašparović, V.; Krilanović, M.; Matanović, K.; Štimac, I.; Luxner, J.; Vraneš, J.; Meštrović, T.; Zarfel, G.; Grisold, A. Emergence of multidrug-resistant Proteus mirabilis in a long-term care facility in Croatia. Wien Klin Wochenschr. 2016, 128, 404–13. [Google Scholar] [CrossRef]
- Rubić, Z.; Soprek, S.; Jelić, M.; Novak, A.; Goić-Barisić, I.; Radić, M.; Tambić-Andrasević, A.; Tonkić, M. Molecular Characterization of β-Lactam Resistance and Antimicrobial Susceptibility to Possible Therapeutic Options of AmpC-Producing Multidrug-Resistant Proteus mirabilis in a University Hospital of Split, Croatia. Microb Drug Resist. 2021, 27, 162–169. [Google Scholar] [CrossRef] [PubMed]
- European Committee for Antimicrobial Susceptibility Testing. Breakpoint Tables for Interpretation of MICs and Zone Diameters. Version 12. 2022. Available online: http://www.eucast.org (accessed on 1st October 2023).
- Clinical Laboratory Standard Institution. Performance Standards for Antimicrobial Susceptibility Testing, 28th ed.; Approved Standard M100-S22; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018. [Google Scholar]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S; Hindler, J.F; Kahlmeter, G; Olsson-Liljequist, B; Paterson, D.L; Rice, L.B; Stelling J.; Struelens, M.J.; Vatopoulos, A.; Weber, J.T.; Monnet, D. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2002, 18, 268–281. [CrossRef]
- Jarlier, V.; Nicolas, M.H.; Fournier, G.; Philippon, A. Extended broad-spectrum beta-lactamases conferring transferable resistance to newer beta-lactam agents in Enterobacteriaceae: hospital prevalence and susceptibility patterns. Rev Infect Dis. 1988, 10, 867–78. [Google Scholar] [CrossRef]
- MacDonald, J.W.; Chibabhai, V. Evaluation of the RESIST-4 O.K.N.V Immunochromatographic Lateral Flow Assay for the Rapid Detection of OXA-48, KPC, NDM and VIM Carbapenemases from Cultured Isolates. Access Microbiol. 2019, 1, e000031. [Google Scholar] [CrossRef]
- Lee, K.; Lim, Y.S.; Yong, D.; Yum, J.H.; Chong, Y. Evaluation of the Hodge Test and the Imipenem-EDTA-double-disk Synergy Test for Differentiating Metallo-β-lactamase-producing Isolates of Pseudomonas spp. and Acinetobacter spp. J. Clin. Microbiol. 2003, 41, 4623–4629. [Google Scholar] [CrossRef]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing, M100, 31st ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2021. [Google Scholar]
- Arlet, G.; Brami, G.; Decre, D.; Flippo, A.; Gaillot, O.; Lagrange, P.H.; Philippon, A. Molecular characterization by PCR restriction fragment polymorphism of TEM β-lactamases. FEMS Microbiol. Lett. 1995, 134, 203–208. [Google Scholar] [CrossRef]
- Nüesch-Inderbinen, M.T.; Hächler, H.; Kayser, F.H. Detection of genes coding for extended-spectrum SHV β-lactamases in clinical isolates by a molecular genetic method, and comparison with the E test. Eur. J. Clin. Microbiol. Infect. Dis. 1996, 15, 398–402. [Google Scholar] [CrossRef] [PubMed]
- Woodford, N.; Ward, M.E.; Kaufmann, M.E.; Turton, J.; Fagan, E.J.; James, D.; Johnson, A.P.; Pike, R.; Warner, M.; Cheasty, T.; Pearson, A.; Harry, S.; Leach, J.B; Loughrey, A.; Lowes, J.A.; Warren, R.E.; Livermore, D.M. Community and hospital spread of Escherichia coli producing CTX-M extended-spectrum β-lactamases in the UK. J. Antimicrob. Chemother. 2004, 54, 735–743. [Google Scholar] [CrossRef] [PubMed]
- Robicsek, A.; Jacoby, G.A.; Hooper, D.C. The worldwide emergence of plasmid-mediated quinolone resistance. Lancet Infect. Dis. 2006, 6, 629–640. [Google Scholar] [CrossRef]
- Poirel, L.; Walsh, T.R.; Cuveiller, V.; Nordman, P. Multiplex PCR for Detection of Acquired Carbapenemases Genes. Diagn. Microbiol. Infect. Dis. 2011, 70, 119–123. [Google Scholar] [CrossRef]
- Woodford, N. , Ellington, M. J; Coelho, J; Turton, J; Ward, M.E.; Brown, S; Amyes S.G; Livermore, D.M.. Multiplex PCR for genes encoding prevalent OXA carbapenemases. Int. J. Antimicrob. Agents. 2006, 27, 351–353. [Google Scholar] [PubMed]
- Perez-Perez, F.J.; Hanson, N.D. Detection of plasmid-mediated AmpC β-lactamase genes in clinical isolates by using multiplex PCR. J. Clin. Microbiol. 2002, 40, 2153–2162. [Google Scholar] [CrossRef]
- Woodford, N.; Fagan, E.J.; Ellington, M.J. Multiplex PCR for rapid detection of genes encoding CTX-M extended-spectrum β-lactamases. J. Antimicrob. Chemother. 2006, 57, 154–155. [Google Scholar] [CrossRef]
- Saladin, M.; Cao, V.T.B.; Lambert, T.; Donay, J.L.; Hermann, J.; Ould-Hocine, L. Diversity of CTX-M β-lactamases and Their Promoter Regions from Enterobacteriaceae Isolated in Three Parisian Hospitals. FEMS Microbiol. Lett. 2002, 209, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Zankari, E.; Hasman, H.; Cosentino, S.; Vestergaard, M.; Rasmussen, S.; Lund, O.; Aarestrup, F.M.; Larsen, M.V. Identification of acquired antimicrobial resistance genes. J Antimicrob Chemother. 2012, 67, 2640–4. [Google Scholar] [CrossRef]
- Carattoli, A.; Bertini, A.; Villa, L.; Falbo, V.; Hopkins, K.L.; Threfall, E.J. Identification of plasmids by PCR-based replicon typing. J Microbiol Methods. 2005, 63, 219–228. [Google Scholar] [CrossRef] [PubMed]
- Carattoli, A.; Seiffert, S.N.; Schwendener, S.; Perreten, V.; Endimiani, A. Differentiation of IncL and IncM Plasmids Associated with the Spread of Clinically Relevant Antimicrobial Resistance. PLoS ONE 2015, 10, e0123063. [Google Scholar] [CrossRef]
- Shaaban, M.; Elshaer, S.L.; Abd El-Rahman, O.A. Prevalence of extended-spectrum β-lactamases, AmpC, and carbapenemases in Proteus mirabilis clinical isolates. BMC Microbiol. 2022, 11, 247. [Google Scholar] [CrossRef]
- Miriagou, V; Papagiannitsis, C. C; Tzelepi, E; Casals, J.B; Legakis, N.J; Tzouvelekis, L.S. Detecting VIM-1 production in Proteus mirabilis by an imipenem-dipicolinic acid double disk synergy test. J. Clin. Microbiol. 2010, 48, 667–8. [CrossRef]
- Papagiannitsis, C. C; Miriagou, V; Kotsakis, S. D; Tzelepi, E; Vatopoulos, A.C; Petinaki, E; Tzouvelekis,.LS. Characterization of a transmissible plasmid encoding VEB-1 and VIM-1 in Proteus mirabilis. Antimicrob. Agents. Chemother. 2012, 56, 4024–5. [Google Scholar] [CrossRef]
- Protonotariou E, Poulou A, Politi L, Meletis G, Chatzopoulou F, Malousi A, Metallidis S, Tsakris A, Skoura L. Clonal outbreak caused by VIM-4-producing Proteus mirabilis in a Greek tertiary-care hospital. Int J Antimicrob Agents. Int J Antimicrob Agents. 2020, 56, 106060. [CrossRef] [PubMed]
- Markovska R, Schneider I, Keuleyan E, Ivanova D, Lesseva M, Stoeva T, Sredkova M, Bauernfeind A, Mitov I. Dissemination of a Multidrug-Resistant VIM-1- and CMY-99-Producing Proteus mirabilis Clone in Bulgaria. Microb Drug Resist. 2017, 23, 345–350. [CrossRef]
- Fritzenwanker, M; Falgenhauer, J; Hain, T; Imirzalioglu, C; Chakraborty T, Yao, Y. The Detection of Extensively Drug-Resistant Proteus mirabilis Strains Harboring Both VIM-; and VIM-75 Metallo-β-Lactamases from Patients in Germany. Microorganisms. 2025 25, 266. [CrossRef]
- Sattler, J.; Noster, J.; Stelzer, Y.; Spille, M.; Schäfer, S.; Xanthopoulou, K.; Sommer, J.; Jantsch, J.; Peter, S.; Göttig, S.; et al. OXA-48-like carbapenemases in Proteus mirabilis—Novel genetic environments and a challenge for detection. Emerg. Microbes. Infect. 2024, 13, 2353310. [Google Scholar] [CrossRef] [PubMed]
- Potron, A.; Hocquet, D.; Triponney, P.; Plésiat, P.; Bertrand, X.; Valot, B. Carbapenem-Susceptible OXA-23-Producing Proteus mirabilis in the French Community. Antimicrob. Agents Chemother. 2019, 63, e00191–e19. [Google Scholar] [CrossRef]
- Bonnin, R.A.; Girlich, D.; Jousset, A.B.; Gauthier, L.; Cuzon, G.; Bogaerts, P. A single Proteus mirabilis lineage from human and animal sources: A hidden reservoir of OXA-23 or OXA-58 carbapenemases in Enterobacterales. Sci Rep. 2020, 8, 9160. [Google Scholar] [CrossRef] [PubMed]
- Bedenić, B; Sardelić, S; Luxner, J; Bošnjak, Z; Varda-Brkić, D; Lukić-Grlić, A; Mareković, I; Frančula-Zaninović, S; Krilanović, M; Šijak, D; Grisold, A; Zarfel, G. Molecular characterization of clas B carbapenemases in advanced stage of dissemination and emergence of class D carbapenemases in Enterobacteriaceae from Croatia. Infect Genetic Evol 2016,. 43,74-82. Doi10.1016.
- Literacka, E.; Bedenic, B.; Baraniak, A. ; Fiett, J; Tonkic, M. ; Jajic-Bencic, I.; Gniadkowski, M. blaCTX-M genes in Escherichia coli strains from Croatian Hospitals are located in new (blaCTX-M-3a) and widely spread (blaCTX-M-3a and blaCTX-M-15) genetic structures. Antimicrob. Agents Chemother. 2009, 53, 1630–1635. [Google Scholar] [CrossRef]
- Car, H.; Dobrić, M.; Pospišil, M.; Nađ, M.; Luxner, J.; Zarfel, G.; Grisold, A.; Nikić-Hecer, A.; Vraneš, J.; Bedenić, B. Comparison of Carbapenemases and Extended-Spectrum β-Lactamases and Resistance Phenotypes in Hospital- and Community-Acquired Isolates of Klebsiella pneumoniae from Croatia. Microorganisms 2024, 2, 2224. [Google Scholar] [CrossRef]
- Bedenić, B; Luxner, J; Zarfel, G; Grisold, A; Dobrić, M; Đuras-Cuculić, B; Kasalo, M; Bratić, V; Dobretzberger, V; Barišić, I. First Report of CTX-M-32 and CTX-M-101 in Proteus mirabilis from Zagreb, Croatia. Antibiotic, 2025, 30, 462. [CrossRef]
- Miriagou, V; Papagiannitsis, C. C; Tzelepi, E; Casals, J.B; Legakis, N.J, Tzouvelekis, L.S. Detecting VIM-1 production in Proteus mirabilis by an imipenem-dipicolinic acid double disk synergy test. J. Clin. Microbiol. 2010, 48, 667–8. [CrossRef]
Figure 1.
Phylogenetic tree showing genetic relatedness between isolates. Four ioslates from the present study showed high level of genetic relatedness but not similarity with ESBL and p-AmpC producing organisms from the previous study.PM3-36810, PM5-5990, PM6-41149, PM8-11646.
Figure 1.
Phylogenetic tree showing genetic relatedness between isolates. Four ioslates from the present study showed high level of genetic relatedness but not similarity with ESBL and p-AmpC producing organisms from the previous study.PM3-36810, PM5-5990, PM6-41149, PM8-11646.
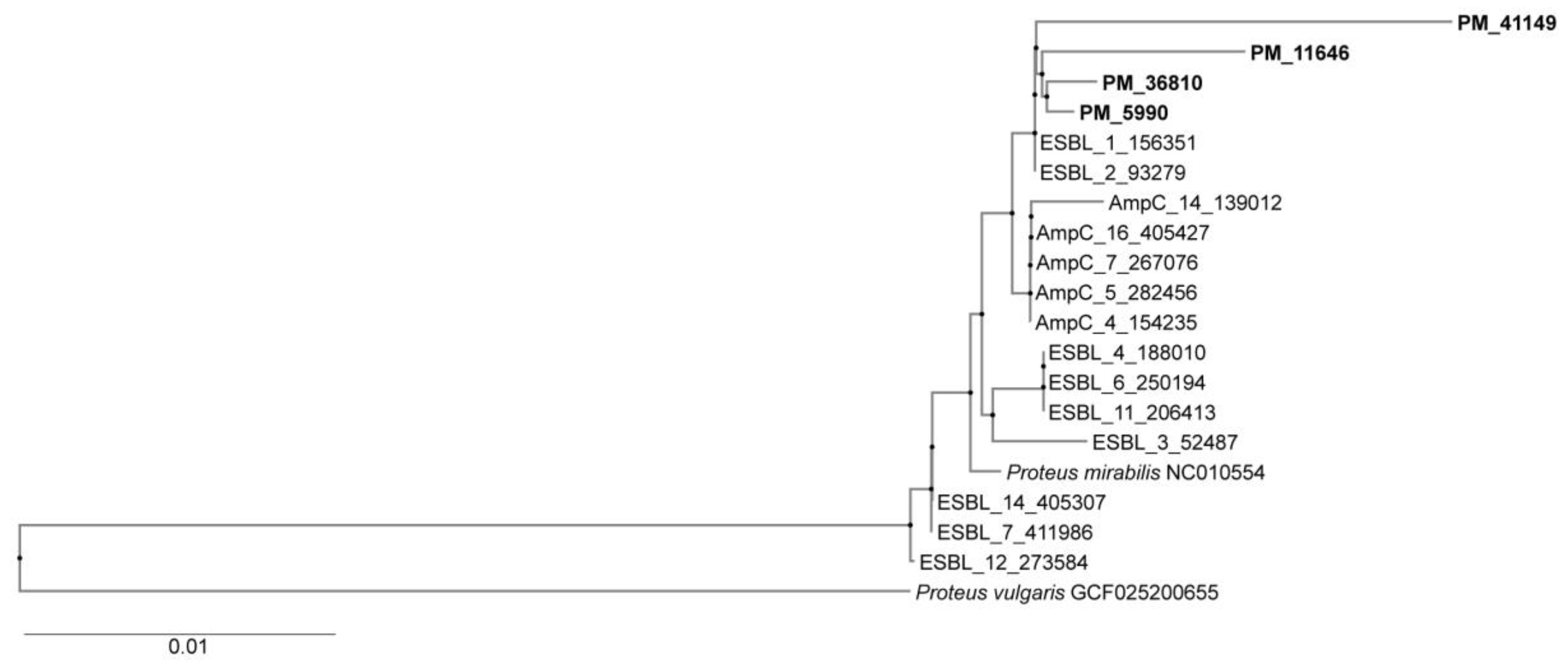
Table 1.
Antibiotic susceptibility profiles of VIM bearing P. mirabilis strains to different antibiotics.
Table 1.
Antibiotic susceptibility profiles of VIM bearing P. mirabilis strains to different antibiotics.
| AMX 32 |
AMC 32/16 |
TZP 128/4 |
CXM 32 |
CAZ 16 |
CTX 4 |
CRO 4 |
FEP 16 |
IMI 4 |
MEM 4 |
GM 16 |
CIP 0,25 |
|
| 1 | >128 (R) | 32/16(R) | 8/4 (S) | >128(R) | 8 (R) | >128(R) | >128(R) | 16(R) | 128 (R) | 0,25(S) | >128(R) | >128(R) |
| 2 | >128 (R) | 64/32(R) | 8/4 (S) | >128(R) | 16(R) | >128(R) | >128(R) | >128(R) |
128 R) | 0,25(S) | >128(R) | >128(R) |
| 3 | >128 (R) | 64/32(R) | 8/4(S) | >128(R) | 8 (R) | >128(R) | >128(R) | >128(R) | 128 (R) | 0,25(S) | >128(R) | >128(R) |
| 4 | >128 (R) | 128/64(R) | 8/4(S) | >128(R) | 2 (I) | >128(R) | >128(R) | >128(R) | 128 (R) | 0,25(S) | >128(R) | >128(R) |
| 5 | >128 (R) | 32/16(R) | 16/4 (S) | >128(R) | 2 (I) | >128(R) | >128(R) | >128(R) | 4 (R) | 0,12 S) | >128(R) | >128(R) |
| 6 | >128 (R) | 32/16(R) | 16/4(S) | >128(R) | 8(R) | >128(R) | >128(R) | >128(R) | 4 (R) | 0,25(S) | >128(R) | >128(R) |
| 7 | >128 (R) | 128/64(R) | 8/4 (S) | >128(R) | 8(R) | >128(R) | >128(R) | >128(R) | 4 (R) | 0,5 (S) | >128(R) | >128(R) |
| 8 | >128 (R) | 128/64(R) | 8/4 (S) | >128(R) | 16(R) | >128(R) | >128(R) | >128(R) | 8 (R) | 0,25(S) | >128(R) | >128(R) |
| 9 | >128 (R) | 128/64(R) | 8/4 (S) | >128(R) | 16(R) | >128(R) | >128(R) | >128(R) | 4 (R) | 0,25(S) | >128(R) | >128(R) |
| 10 | >128 (R) | 128/64(R) | 8/4 (S) | >128(R) | 32(R) | >128(R) | 32(R) | >128(R) | 4 (R) | 0,12 S) | >128(R) | >128(R) |
| 11 | >128 (R) | 128/64(R) | 8/4 (S) | >128(R) | 128(R) | >128(R) | 32(R) | >128(R) | 4 (R) | 0,25(S) | >128(R) | >128(R) |
| 12 | >128 (R) | 128/64(R) | 16/4(S) | >128(R) | 2(I) | >128(R) | >128(R) | >128(R) | 8 (R) | 0,25(S) | >128(R) | >128(R) |
| 13 | >128 (R) | 128/64(R) | 8/4 (S) | >128(R) | 32(R) | >128(R) | 32(R) | >128(R) | 8 (R) | 0,5 (S) | >128(R) | >128(R) |
| 14 | >128 (R) | 128/64(R) | 16/4(S) | >128(R) | 128(R) | >128(R) | 16((R) | >128(R) | 4 (R) | 0,5 (S) | >128(R) | >128(R) |
| 15 | >128 (R) | 128/64(R) | 8/4 (S) | >128(R) | 32(R | >128(R) | 16(R) | >128(R) | 4 (R) | 0,25(S) | >128(R) | >128(R) |
| 16 | >128 (R) | 128/64(R) | 16/4(S) | >128(R) | 128(R) | >128(R) | 128(R) | >128(R) | 4 (R) | 0,12 S) | >128(R) | >128(R) |
| 17 | >128 (R) | 64/32(R) | 16/4(S) | >128(R) | 16(R) | >128(R) | 32(R) | >128(R) | 8 (R) | 0,06 (S) | >128(R) | >128(R) |
| 18 | >128 (R) | 128/64(R) | 8/4 (S) | >128(R) | 64(R) | >128(R) | 64(R) | >128(R) | 4 (R) | 0,25(S) | >128(R) | >128(R) |
| 19 | >128 (R) | 128/64(R) | 8/4 (S) | >128(R) | 64(R) | >128(R) | 32(R) | >128(R) | 4 (R) | 0,25(S) | >128(R) | >128(R) |
| 20 | >128 (R) | 64/32(R) | 16/4(S) | >128(R) | 64(R) | >128(R) | 128(R) | >128(R) | 4 (R) | 0,5 (S) | >128(R) | >128(R) |
Abbreviations: AMX-amoxycillin; AMC—amoxycillin/clavulanic acid; TZP—piperacillin–tazobactam; CXM-cefuroxime; CAZ—ceftazidime; CTX-cefotaxime CRO—ceftriaxone; FEP—cefepime; IMI—imipenem; MEM—meropenem; GM—gentamicin; AMI—amikacin; CIP—ciprofloxacin; R-resistant, I-Intermediate susceptible (susceptible at increased exposure), S-susceptible. Resistance breakpoint is provided.
Table 3.
Whole-genome sequencing of representative isolates. Accession numbers are provided in a separate section at the end of the manuscript.
Table 3.
Whole-genome sequencing of representative isolates. Accession numbers are provided in a separate section at the end of the manuscript.
|
Isolate and Protocol Number |
β-Lactam | Aminoglycosides | Sulphonamide | Trimethoprim | Chloramphenicol | Tetracycline |
| PM 3 | blaCTX-M-202, blaTEM-156, blaTEM-1A, blaTEM-2, blaVIM-4, blaVIM-1, | aac(3)-IId, aph(6)-Id, aph(3'')-Ib, aadA1, armA, aac(6')-IIc, |
sul1 sul2 |
dfrA1 | cat | tet(J) |
| PM 5 | blaCTX-M-202 blaTEM-2, blaTEM-1A, blaVIM-4 | aac(3)-IId, aph(6)-Id, aph(3'')-Ib, aadA1, armA, aac(6')-IIc, |
sul1 sul2 |
dfrA1 | cat | tet(J) |
| PM 6 | blaCTX-M-202 blaTEM-2, , blaVIM--4, | aph(6)-Id, aph(3'')-Ib, armA, aac(6')-IIc, aac(3)-IId, aac(6')-IIc, aadA1 |
sul1 sul2 |
dfrA1 | cat | tet(J) |
| PM 8 | blaCTX-M-202, blaTEM-2, blaTEM-1A, blaVIM-1, blaVIM-4, | aac(3)-IId, aph(6)-Id, aph(3'')-Ib, aadA1, armA, aac(6')-IIc, |
sul1 sul2 |
dfrA1 |
cat | tet(J) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



