
Submitted:
25 March 2025
Posted:
26 March 2025
You are already at the latest version
Abstract
Background/Objectives: Proteus mirabilis is a frequent causative agent of urinary tract and wound infections in community and hospital settings. It develops re-sistance to expanded-spectrum cephalosporins (ESC) due to the production of ex-tended-spectrum β-lactamases (ESBLs) or plasmid-mediated AmpC β-lactamases (p-AmpC). During routine microbiology testing, we observed an increased rate of multi-drug-resistant (MDR) P. mirabilis isolates. Here, we report the characteristics of ESBLs and p-AmpC β-lactamases encountered among hospital and community isolates of P. mirabilis in two hospitals and the community settings in Zagreb, Croatia. Methods: Antibiotic susceptibility testing was performed by disk-diffusion and broth dilution methods. The double disk synergy test (DDST) and inhibitor-based test with clavulanic and cloxacillin were applied to screen for ESBLs and p-AmpC, respec-tively. PCR investigated the nature of ESBL, carbapenemases, and fluoroquinolone resistance determinants. Selected strains were subjected to molecular analysis of resistance traits by the Inter-array CarbaResist Kit and whole genome sequencing (WGS). Results: In total, 39 isolates were analyzed. Twenty-two isolates phenotypically tested positive for p-AmpC and seventeen for ESBLs. AmpC-producing organisms exhibited uniform resistance to amoxicillin-clavulanate, ESC, ciprofloxacin, and co-trimoxazole and uniform susceptibility to carbapenems and piperacillin-tazobactam and harbored blaCMY-16 genes. ESBL-positive isolates demonstrated resistance to amoxicillin-clavulanate, cefuroxime, cefotaxime, ceftriaxone, and ciprofloxacin, but variable susceptibility to cefepime and aminoglycosides. They possessed blaCTX-M genes that belong to cluster 1 (n=5) or 9 (n=12) with CTX-M-14 as the dominant allelic variant. Conclusions: The study's main finding is the diffusion of CTX-M ESBL and CMY-16 p-AmpC among hospital and community-acquired isolates. AmpC-producing iso-lates showed uniform resistance patterns, whereas ESBL-positive strains had varia-ble degrees of susceptibility/resistance to non-β-lactam antibiotics, resulting in more diverse susceptibility patterns. The study demonstrated diffusion resistance deter-minants among hospital and outpatient isolates, mandating improvement in de-tecting β-lactamases during routine laboratory work.
Keywords:
Proteus mirabilis
; extended-spectrum β-lactamases
; plasmid-mediated AmpC β-lactamases
; resistance
1. Introduction
Proteus mirabilis, a Gram-negative bacterium belonging to the family Morganellaceae, is an important causative agent of urinary tract (UTI) and wound infections. It is mostly isolated from catheter-associated urinary tract infections (CAUTI) associated with the production of biofilms, and can further cause bloodstream infections [1]. It develops resistance to expanded-spectrum cephalosporins (ESC) due to the production of extended-spectrum (ESBL) and plasmid-mediated AmpC β-lactamases (p-AmpC) [2].
ESBLs hydrolyze penicillins, first, second, third, and fourth-generation cephalosporins, and monobactams, but are in general inhibited by so-called suicide inhibitors such as clavulanic acid, sulbactam, and tazobactam. The first ESBLs were of TEM and SHV type, a diversification of ESBLs occurred in 2000-ties due to the emergence of the CTX-M family, concomitantly observed in Enterobacterales, most frequently in Escherichia coli and Klebsiella pneumonia [3]. CTX-M β-lactamases preferentially hydrolyze cefotaxime, have an intrinsic extended-spectrum profile, and are classified into five phylogenetic clusters: CTX-M-1 group, CTX-M-2 group, CTX-M-8 group, CTX-M-9 group and CTX-M-25 group [4,5]. They are not closely related to TEM or SHV β-lactamases but are typical members of Ambler’s class A, and are derived from a gene hosted by Kluyvera [4]. In P. mirabilis, CTX-M-ESBL variants are also the most common ESBL types today. ESBL-producing P. mirabilis has been known since 1999, when TEM and SHV enzymes were still the main responsible enzymes [6]. Plasmids encoding ESBL often carry fluoroquinolone resistance genes qnrD and qnrS [4,5].
In the late 1980-ties, cephalosporinase gene (AmpC) of chromosomal origin present in bacteria belonging to the genus Enterobacter, Serratia, Citrobacter, Pseudomonas, and Acinetobacter, were identified on plasmids spreading among Enterobacterales without chromosomal ampC gene including P. mirabilis. They are Ambler class C enzymes that possess hydrolytic activity against expanded-spectrum cephalosporins (ESC), monobactams and cephamycins but spare fourth-generation cephalosporins and carbapenems [7]. Unlike ESBLs, they are not susceptible to inhibition with clavulanic acid, sulbactam, or tazobactam but are susceptible to inhibition by cloxacillin and phenylboronic acid [7]. Genes encoding ESBLs and p-AmpC are encoded on plasmids that often contain resistance genes to non-β lactam antibiotics such as aminoglycosides, tetracyclines, sulphonamides, trimethoprim and fluoroquinolones leading to a multidrug-resistant phenotype (MDR). European studies demonstrated the spread of ESBLs and CMY AmpC β-lactamases among P. mirabilis isolates in the last decades [8,9].
Previous studies found chromosomal incorporation of genes encoding AmpC β-lactamases belonging to the CMY family and the spread of this resistant clade in Europe [9]. ESBL-positive isolates were reported as causative agents of an outbreak in the nursing home Italy [8]. Resistance to aminoglycosides in P. mirabilis is due to the acquisition of genes encoding acetylases, adenylases, and phosphorylases (aacA4, aadB, aphA6) [10].
In Croatia, the first official reports date back to 2008 when the first outbreak with ESBL-positive P. mirabilis isolates, producing TEM-52, was described in the University Hospital Split in the southern region of Croatia [11,12]. Later, in 2015, an outbreak of infections associated with a p-AmpC- positive P. mirabilis-producing CMY-16 variant from a nursing home in Zagreb was described [13]. P-AmpC-producing isolates demonstrated high-level resistance to ESC and uniform susceptibility to piperacillin-tazobactam, cefepime, and carbapenems. Recently, hospital isolates from Split were found to possess CMY-16 as well. [14]. Haemolysins, urease, motility, and biofilm formation are the virulence factors relevant to UTI [2].
In recent years, we observed an increase in ESC-resistant P. mirabilis exhibiting very similar resistance phenotypes in two tertiary hospitals in Zagreb during routine laboratory diagnostics. Herein, we analyzed the ESBLs and p-AmpC associated with resistance to ESC in a set of clinical P. mirabilis isolates obtained from two tertiary hospitals in Zagreb. Moreover, phenotypic characterization of virulence traits was done to measure the potential clinical significance exerted by these isolates. The second goal was to compare different genotypic methods for characterizing resistance genes in P. mirabilis, such as PCR, whole genome sequencing, and the Inter-array genotyping CarbaResist Kit method.
2. Results
2.1. Bacterial Isolates
Thirty-nine isolates were collected in two hospital centers: 38 from University Hospital Centre Zagreb (UHCZ) and one from University Hospital Centre Sestre Milosrdnice (UHCSM). Nine isolates were from hospitalized patients, and the rest were from the outpatient settings. In total, 19 isolates originated from urine samples, 11 from wound swabs, two from urinary catheters and sputum respectively, one from tissue specimens, and the rest from surveillance cultures. Twenty-one females and 18 males were included in the study.
2.2. Antimicrobial Susceptibility Testing and Phenotypic Tests for β-lactamases
Twenty-two isolates were phenotypically positive for p-AmpC whereas 17 tested positive for an ESBL, exhibing augmentation of the inhibition zones around ESC in the presence of clavulanic acid of 10 to 25 mm. AmpC β-lactamases were not inducible.
AmpC-producing organisms were uniformly resistant to amoxicillin alone and combined with clavulanic acid, cefuroxime, ESC (ceftazidime, cefotaxime, ceftriaxone), ciprofloxacin and sulphamethoxazole-trimethoprim and uniformly susceptible to ertapenem, meropenem and piperacillin-tazobactam as shown in Table 1. High resistance rates were observed for gentamicin (95,4%, n=21) and amikacin (90,9%, n=20). Cefepime preserved good activity, with only one strain being resistant (4,5%, n=1) and seven being intermediate susceptible or susceptible at increased exposure (32%, n=7) (Table 1). All isolates were classified as MDR. Multiple antibiotic resistance indices (MARI) ranged from 0,46 to 0,6 with mean value of 0,48.
Seventeen isolates demonstrated positive double disk-synergy test (DDST) and inhibitor-based tests with clavulanic acid, indicating the production of an ESBL. All isolates exhibited high-level resistance to amoxicillin alone and combined with clavulanic acid, cefotaxime, ceftriaxone and ciprofloxacin, with the MIC value exceeding 128 mg/L. The best activity was exerted by meropenem with MICs of all isolates in the susceptible range. There were high resistance rates to ceftazidime, cefepime, and gentamicin, with 58%, 94%, and 76% of the isolates showing resistance, respectively. The MARI ranged from 0,23 to 0,69 with mean value of 0,61. MICs and the results of phenotypic tests are shown in Supplementary Table S1.
2.3. Molecular Detection of Resistance Genes
PCR identified blaCMY genes and blaTEM-1 in all and all but two p-AmpC-positive organisms, respectively. All ESBL-producing organisms were shown to possess blaCTX-M and 10 blaTEM genes. All except five ESBL positive isolates were asigned to the phylogenetic group 9, whereas five were allocated to group 1. Eight amplicons were subjected to sequencing (Eurofin) with forward primer. Blast analysis identified CTX-M-14 encoding genes in four isolates, and CTX-M-24 and CTX-M-44, in one isolate, respectively, all belonging to the cluster CTX-M-9, and blaCTX-M-15 genes belonging to the phylogenetic cluster 1 in two isolates. blaTEM genes generated TEM-1. ISEcp element was found upstream of both CTX-M-15 and CMY encoding genes.
2.4. Inter-Array Genotyping CarbaResist Method
The AmpC-positive isolate contained the blaTEM and the blaCMY gene associated with the ISEcp insertion element. There were a plethora of aminoglycoside resistance genes encoding acetylases (aac6-Ib and aac3Ia) and adenylases (aadA1 and aadA2), as illustrated in Table 2. Two genes responsible for sulphonamide resistance were identified (sul1 and sul2) and one for trimethoprim resistance (dfrA1) (Table 2).
The ESBL-producing organism was positive for blaCTX-M-9 as the sole β-lactam resistance determinant. Similarly, as with AmpC-positive organism, there were a lot of genes associated with aminoglycoside resistance encoding acetylases (aac6-Ib and aac3-IVa) and adenylases (aadA1 and aadA2) and aph gene for aminoglycoside phosphorylase (Table 2). Identical resistance genes, as in the AmpC positive organism, were found for sulphonamide resistance (sul1 and sul2) and trimethoprim resistance, but different allelic variants of the gene for dihydrofolate reductase (dfrA5). Chloramphenicol resistance was mediated by cat genes coding for chloramphenicol acetyltransferase, as demonstrated in Table 2.
2.5. Whole Genome Sequencing (WGS)
All five tested AmpC-positive organisms harbored blaCMY-16 and all but one blaTEM-1b. There was a plethora of aminoglycoside resistance genes encoding acetylases (aac(6′)-Ib3), adenylases (aadA1, aadA2) and phosphorylases (aph(6)-Id, aph(6)-Ib, aph(6)-Ia) and 16S metylase genes (armA) conferring panaminoglycoside resistance (Table 3). Two allelic variants of sulphonamide resistance genes were found: sul1 and sul2, whereas dfrA1 and dfrA12 encode dihydrofolate reductase responsible for trimethoprim resistance. CatA1 allelic variant of the cat gene accountable for the production of chloramphenicol acetyltransferase was found in four out of five sequenced AmpC-producing organisms, as shown in Table 3.
Nine WGS analysed ESBL-producing organisms carried various blaCTX-M genes, comprising blaCTX-M-14 and blaCTX-M-65 found in three isolates respectively, and blaCTX-M—15, blaCTX-M-101 and blaCTX-M-32 in only one strain respectively, in addition to blaTEM-1d identified in two, and blaTEM-1a in one CTX-M-14 positive isolate, as illustrated in Table 3. blaOXA-1 was an additional β-lactamase gene to blaCTX-M-65. There were a lot of different aminoglycoside resistance genes present, including: aac(3)IIa, which was positive in three, and aac(3)IId, and aac(3)-IV in two isolates, respectively, encoding acetyltransferases. aadA1 was found in seven and aadA2 gene in three isolate, generating adenyltransferases, enzymes which modify aminoglycosides. The aph(3)-Ib gene encoding aminoglycoside phosphorylases was present in six and aph(6)Id in eight isolates, as shown in Table 3. Cat and cat1 genes, harbored by all but one strain, were responsible for chloramphenicol acetyltransferase, while sul1 and sul2, which mediated resistance to sulphonamides, were found simultaneously in six isolates. Two isolates harbored only sul2, while one strain resistant to sulphamethoxazole-trimetoprim was deprived of resistance genes. Dihydrofolate reductase, rendering trimethoprim inactive, was encoded by three variants of genes: dfrA1 as the most prevalent and present in six, dfrA32 in three, and dfrA17 in one isolate. Despite the uniform fluoroquinolone resistance, only one isolate positive for CTX-M-32, carried the plasmid-mediated qnrD gene (Table 3). Unlike AmpC-producing organisms, in ESBL-positive strains, the cat gene was the most frequent variant responsible for chloramphenicol resistance, found in six, followed by catA1 present in five isolates. In contrast to the wide variety of aminoglycoside resistance genes widely distributed among all isolates, there was one tetracycline resistance determinant present in all isolates- tet (J). TetC and tetH allelic variants were present in only two strains positive for the blaCTX-M-65 gene. The aac(6′)-Ib-cr gene, conferring combined high-level aminoglycoside and fluoroquinolone resistance, was identified only in two isolates harboring CTX-M-65 ESBL.
2.6. Conjugation
Conjugation experiments failed to transfer either cefotaxime or cefoxitin resistance to either of the two E. coli recipient strains.
2.7. Plasmid Analysis by PCR-Based Replicon Typing (PBRT)
No typable plasmids were demonstrated among AmpC- and ESBL- producing P. mirabilis isolates by multiplex PCR. However, WGS identified IncCol and IncQ1 plasmids in all tested AmpC producing organisms and IncQ1 in all tested ESBL - positive isolates.
2.8. Detection of Virulence Determinants
All isolates were positive for urease activity, hemolysis, and motility.
2.9. Phylogenetic Analysis
A phylogenetic tree was calculated based on WGS data and revealed that all tested AmpC- producers belonged to one closly related cluster with one strain being a singleton (PM14) as shown in Figure 1. On the contrary, the ESBL-producing organisms belonged to four different clusters. (Figure 1). The ESBL-positive isolates from 2008 (S6_320736 and S2_320735) belonged to a different lineage compared to the ESBL-positive isolates from this study (Figure 2).
3. Discussion
The study’s main finding is the diffusion of CTX-M ESBL and CMY-16 p-AmpC among hospital and community-acquired isolates in Croatia. The community setting is the major source of MDR P. mirabilis, which occurs only sporadically in the hospitals. AmpC-producing isolates showed uniform resistance patterns and resistance gene content, whereas ESBL-positive strains had a variable degree of susceptibility/resistance to non-β-lactam antibiotics, resulting in more diverse susceptibility patterns and very diverse resistance genes. The lack of inducibility is consistent with CMY variants, which are deprived of the ampR gene upstream of the blaampC gene. CMY-16 detected in p-AmpC isolates is an identical variant already described in the nursing home in Croatia [13]. Later studies demonstrated the diffusion of this important determinant in the hospital setting in the southern region of Croatia [14]. ISEcp insertion element upstream of the blaCMY genes increases the expression of the genes and the level of resistance and is probably responsible for the mobilization event. The fact that cefoxitin resistance was not transferable indicates the possibility that there was chromosomal insertion of blaCMY genes, as reported in the previous studies [9]. TEM-1 was identified as an additional broad-spectrum β-lactamase among p-AmpC and some ESBL producing organisms and attributed to increased amoxycillin resistance. This is in concordance with earlier reports from Croatia and Europe, which also found TEM-1 alongside CMY p-AmpC [9]. In the previous report from Croatia, AmpC-producing organisms were recovered predominantly from urinary catheters in the nursing home, but in this study, most of the isolates originated from mid-stream urine samples of the outpatient population.
In contrast to ESBL, Amp-C-producing organisms showed almost identical resistance phenotypes with uniform resistance to ESC, aminoglycosides, sulphonamides, and fluoroquinolones, and unform susceptibility to piperacillin-tazobactam and carbapenems, with only slight variations in MIC values and the same resistance gene content. Cefepime is not hydrolyzed by p-AmpC, but some isolates were resistant or intermediate susceptible (increased exposure). This could be attributed to other resistance mechanisms, such as porin loss or hyperexpression of efflux pumps. In a study conducted in Egypt, the coproduction of p-AmpC and ESBL belonging to the CTX-M family was identified in 60% of the isolates [15]. In their study, VIM-1 and VIM-2 metallo-β-lactamases associated with carbapenem resistance were identified as well [15]. In the present study, the isolates possessed either ESBL or p-AmpC and none of the isolates exhibited carbapenem resistance.
Regarding ESBLs, the shift from TEM-52 reported in the late 2000-ties [12,13] to the absolute dominance of CTX-M variants was observed. In Italian hospitals, TEM-52 was found to be the most prevalent variant [16].
CTX-M variants reported among P. mirabilis isolates belonged predominantly to the CTX-M-9 group, with CTX-M-14 and CTX-M-65 as the dominant variants. This is in contrast with previous studies done on E. coli and K. pneumoniae in which CTX-M-15 was produced by the vast majority of the tested isolates with only sporadic ocurrence of CTX-M-3 [17,18,19,20,21]. In the present study we detected CTX-M-15 in only three isolates. In this study, a huge number of CTX-M allelic variants were reported, in addition to a plethora of various non-β-lactam resistance genes. CTX-M-101 is the new allelic variant that has not been reported in Croatia so far but was identified among animal isolates of E. coli in South Korea [22]. Similarly as the animal isolates, our strain exhibited resistance to all cephalosporins, aminoglycosides, fluoroquinolones, sulphonamid and trimethoprim. The ISEcp insertion element was found upstream of the blaCTX-M-15 genes. It is responsible for the mobilization of the genes and increases the expression of the gene and the level of resistance [23]. IS26 was not found. European reports on ESBLs among Proteus spp. are scarce and the majority of bibliographic references originate from East Asia.
CTX-M-14 was the dominant allelic variant identified in seven isolates. It belongs to the phylogenetic cluster 9. This variant was usually reported from animal sources, but there are sporadic reports of human infections, usually from the Far East. The isolate associated with bloodstream infection in China harboured 15 antibiotic resistance genes, including cat, tet(J), blaCTX-M-14 (three copies), aph(3′)-Ia, qnrB4, blaDHA-1, qacE, sul1, armA, msr(E), mph(E), aadA1, and dfrA1. Aminoglycoside and trimethoprim resistance genes were embedded in class 2 integron [24]. The other report from China described CTX-M-140 as a new variant of CTX-M-14 with decreased hydrolytic activity against cephalosporins [25]. Our isolates exhibited very high MICs of cefotaxime, indicating the enzyme’s high hydrolytic activity. Moreover, CTX-M-14 was frequently isolated from human sources in Taiwan [26]. blaCTX-M-14 gene was a part of class 2 integron.
CTX-M-65, a derivative of CTX-M-14 and a member of cluster 9, was present in three isolates. The isolates producing CTX-M-65 were previously identified in P. mirabilis from Russia with additional blaVEB encoding Vietnam-extended-spectrum β-lactamase (VEB), aac6-Ib gene encoding aminoglycoside resistance and qnrA1 for fluoroquinolone resistance [27]. WGS revealed that the isolates belonged to two different clones. Our CTX-M-65-producing organisms harbored aac3-IIa and no plasmid-mediated fluoroquinolone resistance determinants. The same allelic variant was identified in animal P. mirabilis isolates from Hong Kong, the gene was located in Tn7-like composite transposon, and was associated with extensively drug-resistant phenotype [28]. Unlike previous studies, the ESBL-encoding genes were chromosomally encoded. Genes responsible for sulphonamide resistance sul1 and sul2, as well as chloramphenicol catB3, were located on the chromosome. CTX-M-65 was also described as an allelic variant present in pigs [29].
CTX-M-32, related to CTX-M-15 and member of CTX-M-group 1, was detected in only one isolate. This allelic variant was previously identified only in E. coli, mostly from chicken and pigs [30,31]. To our knowledge, this is the first report of this allelic variant in P. mirabilis. CTX-M-24 and CTX-M-44 were registered only sporadically each in one isolate.
The CTX-M producing organisms exhibited very variable levels of resistance to ESC which could be attributed to the differences in plasmid or gene copy numbers or mutations in the promotor region of the gene. The unique characteristic of all CTX-M producing isolates was the high level of resistance to cefotaxime, but with MICs of ceftazidime, ceftriaxone and cefepime being in the susceptible, intermediate or resistant category. Unlike other Enterobacterales (mainly K. pneumoniae and E. coli) which were analyzed in our previous studies and were harbouring CTX-M-15 associated with high level resistance to all ESC and cefepime, the blaCTX-M genes in the present study belonged predominantly to the phylogenetic cluster 9, and were linked to very variable ESC resistance phenotypes
In contrast to p-AmpC β-lactamases in which CMY-16 remained the only allelic variant for a prolonged time, a significant diversity of the encountered ESBLs was noticed.
The fact that no typable plasmids were found among AmpC-producing organisms supports the hypothesis of the chromosomal incorporation of blaCMY genes. Cefoxitin resistance was not transferable, coupled with negative PCR-based replicon typing (PBRT) could indicate chromosomal incorporation of blaCMY genes mediated by ISEcp, as reported by other authors [9]. WGS found IncCol and IncQ1 genes, but since transconjugants were not obtained, we could not prove that they carried either blaCMY or blaCTX-M genes. The fact that ESC resistance was not transferable is in line with negative PBRT which did not find any typable plasmid in the isolates. This is not surprising for p-AmpC because previous studies demonstrated chromosomal incorporation of blaCMY gene mediated by ISEcp, but it was not expected for blaCTX-M genes which are usually carried by IncFIA, IncW or IncFIB plasmids. The IncQ1 plasmid found by WGS was not previously identified to spread CTX-M encoding genes and thus its presence is not necessarily linked to blaESBL genes
ESBLs are routinely detected in microbiology laboratories by double-disk synergy test (DDST). However, there is no testing for p-AmpC in the frames of routine microbiology diagnostic, which enables the spread of isolates harboring this important resistance determinant. Failure to detect these enzymes may be responsible for the lack of appropriate infection control measures to prevent the rapid dissemination of pathogens among patients who receive inappropriate therapy. Despite that, we have not observed rapid dissemination of these isolates in the two hospital centers involved in the study. On the contrary, they were more prevalent in the outpatient setting. This is in contrast with the results from southern Croatia in which CMY-16 was described among hospital isolates [14].
The phenotypic tests proved very reliable for the detection of both p-AmpC and ESBLs and showed excellent correlation with molecular testing. However, the reporting of AmpC phenotype is underestimated in our laboratory in spite of its diffusion capacity among P. mirabilis. Inhibitor- based tests with cloxacillin are not routinely done and the results are communicated to clinicians as non-ESBL resistance, and therefore infection control measures are not recommended in contrast to ESBL phenotype. MAR indices were lower in AmpC- positive organisms, indicating a smaller number of antibiotics effective against AmpC- compared to ESBL- positive strains. Interestingly, cefepime lost activity against some CMY positive organisms, probably due to hyperproduction of AmpC enzyme. UTI and wound infections were the predominant sources of MDR P. mirabilis. The presence of ESBLs and p-AmpC complicate antibiotic therapy as the therapeutic options are very limited, particularly among oral antibiotics. Sulphamethoxazole-trimethoprim and ciprofloxacin were resistant in all, and all but one isolate. Co-resistance with non-β-lactam antibiotics, including aminoglycosides and fluoroquinolones, was detected in the majority of isolates. The plasmids encoding ESBLs and p-AmpC often possess resistance genes to non-β-lactam antibiotics, which can also exert selection pressure, enabling the horizontal spread of the plasmid. Only one isolate was shown positive for plasmid-borne qnrD gene. Therefore, fluoroquinolone resistance was probably mediated by mutations of gyrA and parC genes, as reported by other authors [32]. Still, the investigation of these resistance mechanisms was beyond this study. In Tunis, qnr6 was the most frequent fluoroquinolone resistance determinant among ESBL-producing organisms [33].
Urease activity increases the urine pH and enhances the formation of renal stones. Motility and hemolysins are important virulence factors for developing upper urinary tract infections [2].
The study found good correlation between phenotypic tests and molecular detection of resistance genes. High level β-lactam and aminoglycoside resistance was in line with a plethora of various resistance genes. However, PBRT did not find any plasmids either in AmpC- or in ESBL- producing organisms, but they were identified by WGS. This points out to the advantage of using new molecular techniques in analyzing molecular epidemiology of resistant isolates.
4. Materials and Methods
4.1. Bacterial Isolates
Bacterial isolates were collected in two largest hospital centers in Zagreb, Croatia, UHCZ and UHSM. The isolates were collected from June 2nd, 2022, until March 19th, 2024, from various clinical specimens and identified to the species level by MALDI-TOF (matrix-assisted laser desorption ionization-time of flight mass spectrometry) Biotyper (Bruker, Daltonik GmbH, Bremen, Germany).
4.2. Antimicrobial Susceptibility Testing and Phenotypic Tests for β-lactamases
The strains were tested for susceptibility to 13 antimicrobials: amoxicillin alone and combined with clavulanate, piperacillin-tazobactam, cefuroxime, expanded-spectrum cephalosporins (ESCs: ceftazidime, cefotaxime, ceftriaxone), cefepime, imipenem, meropenem, gentamicin, amikacin and ciprofloxacin by broth dilution method according to Clinical Laboratory Standard Institution CLSI [34]. A disk diffusion test according to the European Committe for Antimicrobial Susceptibility testing (EUCAST) [35] determined the susceptibility to sulphamethoxazole-trimethoprim, levofloxacin and chloramphenicol. E. coli ATCC 25922 was used as a quality control strain. The intrinsically resistant antibiotics in Proteus spp. such as colistin, nitrofurantoin, tigecycline, and tetracycline, were excluded. The isolates were classified as MDR, as described previously by Magiorakos et al. [36]. MARI were calculated by dividing the sum of antibiotics against which the strain displayed resistance by the total number of antibiotics tested [37]. ESBLs were detected by DDST [38], and confirmed by the combined disk test with ESC and cefepime with and without clavulanic acid according to CLSI [34]. Augmentation of the inhibition zones around cephalosporin disks by clavulanic acid for at least 5 mm was considered a positive result. Screening for p-AmpC was done based on reduced susceptibility to cefoxitin. P-AmpC was confirmed by cefoxitin-cloxacillin DDST (CC-DDST) based on the inhibitory effect of cloxacillin on AmpC enzyme production [39]. A disk of cefoxitin (30 µg) only and another supplemented with cloxacillin solution (20 mg/ml) were placed on Mueller-Hinton agar, inoculated with 0,5 McFarland bacterial suspension, and incubated overnight at 37° C. An augmentation of the inhibition zone by at least 5 mm by cloxacillin, compared to an unsupplemented disk, was considered a positive result. To study the inducibility of the AmpC enzyme, the cefoxitin-cefotaxime antagonist test was performed as described previously and the β-lactamase inducibility was confirmed by the presence of a blunted cefotaxime zone adjacent to cefoxitin [40].
4.3. Molecular Detection of Resistance Genes
The DNA was extracted by boiling method. Singleplex PCR was applied to detect genes encoding broad and extended-spectrum β-lactamases (blaSHV, blaTEM, blaCTX-M,, blaPER-1) [41,42,43] and plasmid-mediated fluoroquinolone resistance genes (qnrA, qnrB and qnrS) [44], was carried out on the isolates phenotypically positive for an ESBL and exhibiting reduced susceptibility to fluoroquinolones, respectively. Multiplex PCR assay was conducted for the detection of p-AmpC on isolates with reduced susceptibility to cefoxitin [45], and to determine five phylogenetic lineages of CTX-M β-lactamases (CTX-M-1, CTX-M-2, CTX-M-8, CTX-M-9 and CTX-M-25) [46], among isolates positive for blaCTX-M genes. Isolates positive for blaCTX-M and blaCMY genes were further tested for the presence of insertion sequence IS26 and ISEcp by PCR mapping using forward primer for insertion element combined with universal reverse primer for blaCTX-M genes (MA3) [47]. Eight isolates (P5, P8, P9, P10, P13, P15, P16, and P17) were subjected to sequencing for blaCTX-M genes using Eurofin Genomic service (Germany). The positive control strains producing TEM-1, TEM-2, and SHV-1 and SHV-2 were kindly provided by Prof. Adolf Bauernfeind (Max von Pettenkofer Institute, Munich, Germany); CTX-M-15 by Prof. Neil Woodford (Health Protection Agency, London, UK).
4.4. Detection of Resistance Genes by Inter-Array Kit CarbaResist
Two P. mirabilis isolates (one positive for an ESBL and the other for p-AmpC) were genotyped by an Inter-array chip according to the manufacturer’s recommendations (Inter-array fzmb GmbH, Bad Langensalza, Germany). The Inter-array genotyping Kit CarbaResist detects broad-spectrum β-lactamases, p-AmpC, ESBLs carbapenemases, and numerous other resistance genes (https://www.inter-array.com/further-genotyping-kits). RNA-free, unfragmented genomic DNA was isolated from the pure culture of the test strains, amplified, and internally labeled with biotin-dUDP according to the linear PCR amplification protocol using the antisense primer of the different targets only. Single-stranded DNA (ssDNA) reaction products were obtained. The biotin-labeled ssDNA was transferred to the ArrayWell and hybridized to DNA oligonucleotide microarrays with 230 probes for different β-lactam, aminoglycoside, fluoroquinolone, sulphonamide, trimethoprim, and colistin resistance genes. HRP-conjugated streptavidin was bound to the hybridized biotin-labeled ssDNA stains and visualized by an enzymatic reaction. The INTER-VISION Reader was used to evaluate the spots and their intensities automatically based on a digital image of the microarray. The samples obtained from the strains tested in the study were automatically analyzed for the presence or absence of specific probes, cross-checked against a database, and then information about existing resistances was output.
4.5. Whole Genome Sequencing (WGS)
Five randomly selected AmpC and nine ESBL-positive isolates were subjected to WGS. First, the strains were cultivated overnight in Tryptic Soy Broth (TSB) and Casein-Peptone Soymeal-Peptone (CASO) Broth (Merck Millipore, MA, USA) at 37 °C. Then, the genomic DNA was extracted using the QIAamp UCP Pathogen Mini Kit (Qiagen, Hilden, Germany) according to the manufacturer’s instructions. The DNA extracts were sent to the Next Generation Sequencing Facility of the Vienna Biocenter for sequencing using Illumina’s NextSeq1000 system according to the manufacturer’s instructions.
The single reads obtained were assembled with Unicycler (DOI: 10.1371/journal.pcbi.1005595) and analyzed using the webservers and services of the Center for Genomic Epidemiology (http://www.genomicepidemiology.org)[48]. A phylogenetic tree was generated using the WGS data and the REALPHY online tool (https://doi.org/10.1093/molbev/msu088). Subsequently, the calculated trees were uploaded to phylo.io (doi: 10.1093/molbev/msw080) for visalization.
4.6. Conjugation
The transferability of cefotaxime or cefoxitin resistance was determined by conjugation (broth mating method) employing E. coli A15R- strain resistant to rifampicin and E. coli J65 resistant to sodium azide [49]. Transconjugants were selected on combined plates containing cefotaxime (2 mg/L) or cefoxitin (8 mg/L) to inhibit the growth of recipient strain and rifampicin (256 mg/L) or sodium azide (100 mg/L) to suppress the donor strains.
4.7. Characterization of Plasmids
Plasmids were extracted with a Qiagen Mini kit according to the manufacturer’s instructions. PCR-based replicon typing (PBRT), according to Carattoli et al. [50] and updated version for detection of L plasmid [51]. Eighteen pairs of primers were used, including five multiplex and three simplex PCR, to assess the plasmid incompatibility group.
4.8. Detection of Virulence Determinants
Urease activity was determined by inoculating the strains in the urea-containing medium. The change of the color to pink was recorded as a positive result. The hemolytic activity was tested by culturing the strains on a 10% sheep blood plate with the addition of trimethoprim to inhibit swarming. For motility assay, one colony was stabbed with a one mcl loop into a semisolid nutrient medium. After overnight incubation at 37 °C, motility was measured as turbidity of the medium [2].
5. Conclusions
The study demonstrated the diffusion of β-lactam resistance determinants among hospital and outpatient isolates, mandating improvement in detecting β-lactamases during routine laboratory work. The studies focused on P. mirabilis are rare even though it is among the most commonly isolated causative agents of UTI, trailing only behind E. coli. P-AmpC still outnumbers ESBL-producing isolates, even though cefoxitin is not licensed in Croatia to exert selection pressure for the diffusion of blaampC genes.
Similarly, as ESBLs in K. pneumoniae and E. coli evolved from SHV-2 and SHV-5 in early 90-ties to CTX-M-15 nowadays, in P. mirabilis they evolved from TEM-52 to CTX-M-9 phylogenetic group and unlike other Enterobacterales with large number of allelic variants.MDR isolates of P. mirabilis are becoming a growing public health problem due to limited therapeutic options. The majority of the patients suffered from complicated UTIs and were not hospitalized. The options for oral therapy are very scarce, mainly because this species has intrinsic resistance to nitrofurantoin and fosfomycin. Only carbapenems exerted sustained activity against ESBL and p-AmpC-producing organisms, but parenteral antibiotics are unsuitable for outpatient settings.
The study did not find any relatedness between present isolates and those from 2008, indicating that the present isolates did not evolve from the previous ones, but developed de novo.
Our findings underscore significant challenges posed by P. mirabilis in terms of antibiotic resistance, with increasing resistance to β-lactam antibiotics due to ESBLs and p-AmpC. These results highlight the severity of P. mirabilis as a pathogen and underscore its rapid evolution and adaptability in developing resistance. This study aims to deepen our understanding of the antibiotic resistance mechanisms of P. mirabilis, to provide important insights for developing future antimicrobial drugs. It promotes effective treatment to get this pathogen under control, and to mitigate its threat to human health.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1: MICs, phenotypic tests and molecular detection of resistance gene in Proteus mirabilis.
Author Contributions
Conceptualization, B.B.; formal analysis, B.B; M.K; M.D.; J.L; G.Z; V.D; V.B; M.K; B.D.C: data curation: M.D; and B.D.C, writing—original draft preparation, B.B. and M.D. writing—review and editing B.B.; I.B; and A.G. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by a grant from the University of Zagreb (extended-spectrum β-lactamases, plasmid-mediated β-lactamases, and carbapenemases in Proteus mirabilis, grant number: 10106-24-1294).
Institutional Review Board Statement
The study was approved by the Ethical Committee of the University Hospital Centre Zagreb, class: 8.1-24/282-4, number: 02/013 AG. This is an in vitro study on bacterial isolates collected during routine laboratory diagnostic. It did not involve human or animal subjects.
Informed Consent Statement
Not applicable, as this is a retrospective in vitro study.
Data Availability Statement
The original contributions presented in the study are included in the article, further inquiries can be directed to the corresponding author.
Acknowledgments
We thank Marina Nađ for technical assistance.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
ESBL-extended-spectrum β-lactamases, p-AmpC-plasmid-mediated AmpC β-lactamases, ESC: expanded-spectrum cephalosporinsMIC—minimum inhibitory concentration; WGS—whole genome sequencing, PFGE—pulsed-field gel electrophoresis; BSI—bloodstream infection; CLSI—Clinical Laboratory Standard Institution; EUCAST—European Committee for Antimicrobial Susceptibility Testing; MHA—Mueller-Hinton agar; CAZ—ceftazidime; FEP—cefepime; CTX—cefotaxime; CRO—ceftriaxone; AMT—aztreonam; IMI—imipenem, MEM—meropenem; ERT—ertapenem; MIC—minimum-inhibitory concentration.
References
- Sanches, M.S.; Silva, L.C.; Silva, C.R.D.; Montini, V.H.; Oliva, B.H.D.; Guidone, G.H.M.; Nogueira, M.C.L.; Menck-Costa, M.F.; Kobayashi, R.K.T.; Vespero, E.C.; Rocha, S.P.D. Prevalence of Antimicrobial Resistance and Clonal Relationship in ESBL/AmpC-Producing Proteus mirabilis Isolated from Meat Products and Community-Acquired Urinary Tract Infection (UTI-CA) in Southern Brazil. Antibiotics (Basel). 2023, 2023 10, 370. [Google Scholar] [CrossRef]
- Li, Y.; Yin, M.; Fang, C.; Fu, Y.; Dai, X.; Zeng, W.; Zhang, L. Genetic analysis of resistance and virulence characteristics of clinical multidrug-resistant Proteus mirabilis isolates. Front. Cell. Infect. Microbiol. 2023, 11, 1229194. [Google Scholar] [CrossRef] [PubMed]
- Paterson, D.L.; Bonomo, R.A. Extended-spectrum β-lactamases: A clinical update. Clin. Microbiol. Rev. 2005, 18, 657–686. [Google Scholar] [CrossRef] [PubMed]
- Cantón, R.; Coque, T.M. The CTX-M β-lactamase pandemic. Curr. Opin. Microbiol. 2006, 9, 466–475. [Google Scholar] [CrossRef]
- Coque, T.M.; Baquero, F.; Canton, R. Increasing prevalence of ESBL-producing Enterobacteriaceae in Europe. Eurosurveillance 2008, 13, 19044. [Google Scholar] [PubMed]
- de Champs, C.; Bonnet, R.; Sirot, D.; Chanal, C.; Sirot, J. Clinical relevance of Proteus mirabilis in hospital patients: a two year survey. J Antimicrob Chemother. 2000, 45, 537–9. [Google Scholar] [CrossRef]
- Jacoby, G.A. AmpC β-lactamases. J. Clin. Microbiol. 2009, 22, 161–182. [Google Scholar] [CrossRef]
- Migliavacca, R.; Migliavacca, A.; Nucleo, E.; Ciaponi, A.; Spalla, M.; De Luca, C.; Pagani, L. Molecular epidemiology of ESBL producing P. mirabilis strains from a long-term care and rehabilitation facility in Italy. New Microbiol. 2007, 30, 362–6. [Google Scholar]
- D’Andrea, M.M.; Literacka, E.; Zioga, A.; Giani, T.; Baraniak, A.; Fiett, J.; Sadowy, E.; Tassios, P.T.; Rossolini, G.M.; Gniadkowski, M.; Miriagou, V. Evolution and spread of a multidrug-resistant Proteus mirabilis clone with chromosomal AmpC-type cephalosporinases in Europe. Antimicrob Agents Chemother. 2011, 55, 2735–42. [Google Scholar] [CrossRef]
- Girlich, D.; Bonnin, R.A.; Dortet, L.; Naas, T. Genetics of Acquired Antibiotic Resistance Genes in Proteus spp. Front Microbiol. 2020, 21, 256. [Google Scholar] [CrossRef]
- Sardelić, S.; Bedenić, B.; Sijak, D.; Colinon, C.; Kalenić, S. Emergence of Proteus mirabilis isolates producing TEM-52 extended-spectrum β-lactamases in Croatia. Chemotherapy. 2010, 56, 208–13. [Google Scholar] [CrossRef] [PubMed]
- Tonkić, M.; Mohar, B.; Šiško-Kraljević, K.; Meško-Meglič, K.; Goić-Barišić, I.; Novak, A.; Kovačić, A.; Punda-Polić, V. High prevalence and molecular characterization of extended-spectrum β-lactamase-producing Proteus mirabilis strains in southern Croatia. J Med Microbiol. 2010, 59, 1185–1190. [Google Scholar] [CrossRef]
- Bedenić, B.; Firis, N.; Elveđi-Gašparović, V.; Krilanović, M.; Matanović, K.; Štimac, I.; Luxner, J.; Vraneš, J.; Meštrović, T.; Zarfel, G.; Grisold, A. Emergence of multidrug-resistant Proteus mirabilis in a long-term care facility in Croatia. Wien Klin Wochenschr. 2016, 128, 404–13. [Google Scholar] [CrossRef] [PubMed]
- Rubić, Z.; Soprek, S.; Jelić, M.; Novak, A.; Goić-Barisić, I.; Radić, M.; Tambić-Andrasević, A.; Tonkić, M. Molecular Characterization of β-Lactam Resistance and Antimicrobial Susceptibility to Possible Therapeutic Options of AmpC-Producing Multidrug-Resistant Proteus mirabilis in a University Hospital of Split, Croatia. Microb Drug Resist. 2021, 27, 162–169. [Google Scholar] [CrossRef]
- Shaaban, M.; Elshaer, S.L.; Abd El-Rahman, O.A. Prevalence of extended-spectrum β-lactamases, AmpC, and carbapenemases in Proteus mirabilis clinical isolates. BMC Microbiol. 2022, 11, 247. [Google Scholar] [CrossRef] [PubMed]
- Perillli, M.; Segatore, B.; Mugnaioli, C.; Celenza, G.; Rossolini, G.M. , Stefani, S.; Luzzaro, F.; Pini, B.; Amicosante, G. Persistence of TEM-52/TEM-92 and SHV-12 extended-spectrum β-lactamases in clinical isolates of Enterobacteriaceae in Italy. Microb Drug Resist. 2011, 17, 521–524. [Google Scholar] [CrossRef]
- Literacka, E.; Bedenic, B.; Baraniak, A.; Fiett, J.; Tonkic, M.; Jajic-Bencic, I.; Gniadkowski, M. blaCTX-M genes in Escherichia coli strains from Croatian Hospitals are located in new (blaCTX-M-3a) and widely spread (blaCTX-M-3a and blaCTX-M-15) genetic structures. Antimicrob Agents Chemother. 2009, 53, 1630–5. [Google Scholar] [CrossRef]
- Krilanović, M.; Tomić-Paradžik, M.; Meštrović, T.; Beader, N.; Herljević, Z.; Conzemius, R.; Barišić, I.; Vraneš, J.; Elveđi-Gašparović, V.; Bedenić, B. Extended-spectrum beta-lactamases and plasmid diversity in urinary isolates of Escherichia coli in Croatia: a nation-wide, multicentric, retrospective study. Folia Microbiol (Praha). 2020, 65, 649–667. [Google Scholar] [CrossRef]
- Car, H.; Dobrić, M.; Pospišil, M.; Nađ, M.; Luxner, J.; Zarfel, G.; Grisold, A.; Nikić-Hecer, A.; Vraneš, J.; Bedenić, B. Comparison of Carbapenemases and Extended-Spectrum β-Lactamases and Resistance Phenotypes in Hospital- and Community-Acquired Isolates of Klebsiella pneumoniae from Croatia. Microorganisms. 2024, 2, 2224. [Google Scholar] [CrossRef]
- Bandić Pavlović, D.; Pospišil, M.; Nađ, M.; Vrbanović Mijatović, V.; Luxner, J.; Zarfel, G.; Grisold, A.; Tonković, D.; Dobrić, M.; Bedenić, B. Multidrug-Resistant Bacteria in Surgical Intensive Care Units: Antibiotic Susceptibility and β-Lactamase Characterization. Pathogens. 2024, 15, 11. [Google Scholar] [CrossRef]
- Bedenić, B.; Pešorda, L.; Krilanović, M.; Beader, N.; Veir, Z.; Schoenthaler, S.; Bandić-Pavlović, D.; Frančula-Zaninović, S.; Barišić, I. Evolution of Beta-Lactamases in Urinary Klebsiella pneumoniae Isolates from Croatia; from Extended-Spectrum Beta-Lactamases to Carbapenemases and Colistin Resistance. Curr Microbiol. 2022, 15, 355. [Google Scholar] [CrossRef]
- Seo, K.W.; Do, K.H.; Lee, W.K. Comparative Genetic Characterization of CTX-M-Producing Escherichia coli Isolated from Humans and Pigs with Diarrhea in Korea Using Next-Generation Sequencing. Microorganisms. 2023, 28, 1922. [Google Scholar] [CrossRef]
- Mahrouki, S.; Belhadj, O.; Chihi, H.; Mohamed, B.M.; Celenza, G.; Amicosante, G.; Perilli, M. Chromosomal blaCTX-M-₁₅ associated with ISEcp1 in Proteus mirabilis and Morganella morganii isolated at the Military Hospital of Tunis, Tunisia. J Med Microbiol. 2012, 61, 1286–1289. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Shan, G.; Yu, G.; Wei, J.; Zhang, Q.; Su, W.; Lin, Q.; Zheng, Z.; Wu, G.; Li, G.; Chang, Q.; Yuan, H.; He, Y.; Chen, Y.; Zhang, Y.; Huang, H.; Hu, W.; Song, R.; Weng, Y.; Li, X.; Liu, S. Whole genome sequencing of multidrug-resistant Proteus mirabilis strain PM1162 recovered from a urinary tract infection in China. J Glob Antimicrob Resist. 2023, 33, 44–50. [Google Scholar] [CrossRef]
- Tian, G.B.; Jiang, Y.Q.; Huang, Y.M.; Qin, Y.; Feng, L.Q.; Zhang, X.F.; Li, H.Y.; Zhong, L.L.; Zeng, K.J.; Patil, S.; Xing, Y.; Huang, X. Characterization of CTX-M-140, a Variant of CTX-M-14 Extended-Spectrum β-Lactamase with Decreased Cephalosporin Hydrolytic Activity, from Cephalosporin-Resistant Proteus mirabilis. Antimicrob Agents Chemother. 2016, 23, 6121–6. [Google Scholar] [CrossRef]
- Chao, C.M.; Lai, C.C.; Yu, W.L. Epidemiology of extended-spectrum β-lactamases in Enterobacterales in Taiwan for over two decades. Front Microbiol. 2023, 23, 1060050. [Google Scholar] [CrossRef]
- Karpenko, A.; Shelenkov, A.; Petrova, L.; Gusarov, V.; Zamyatin, M.; Mikhaylova, Y.; Akimkin, V. Two multidrug-resistant Proteus mirabilis clones carrying extended spectrum beta-lactamases revealed in a single hospital department by whole genome sequencing. Heliyon. 2024, 2, e40821. [Google Scholar] [CrossRef]
- Biggel, M.; Boss, S.; Uea-Anuwong, T.; Lugsomya, K.; Magouras, I.; Stephan, R. Complete Genome Sequence of the Extensively Drug-Resistant Extended-Spectrum β-Lactamase-Producing Proteus mirabilis Isolate HK294, Obtained from Poultry Feces in Hong Kong. Microbiol. Resour. Announc. 2023, 20, e0022523. [Google Scholar] [CrossRef]
- Martínez-Álvarez, S.; Châtre, P.; François, P.; Zarazaga, M.; Madec, J.Y.; Haenni, M.; Torres, C. Comparative phylogenomics of extended-spectrum β-lactamase-producing Escherichia coli revealed a wide diversity of clones and plasmids in Spanish chicken meat. Int. J. Food Microbiol 2025, 2, 110900. [Google Scholar] [CrossRef]
- Lemlem, M.; Aklilu, E.; Mohamed, M.; Kamaruzzaman, N.F.; Devan, S.S.; Lawal, H.; Kanamma, A.A. Prevalence and molecular characterization of ESBL-producing Escherichia coli isolated from broiler chicken and their respective farms environment in Malaysia. BMC. Microbiol. 2024, 26, 499. [Google Scholar] [CrossRef]
- Martínez-Álvarez, S.; Châtre, P.; François, P.; Abdullahi, I.N.; Simón, C.; Zarazaga, M.; Madec, J.Y.; Haenni, M.; Torres, C. Unexpected role of pig nostrils in the clonal and plasmidic dissemination of extended-spectrum β-lactamase-producing Escherichia coli at farm level. Ecotoxicol. Environ. Saf. 2024, 15, 116145. [Google Scholar] [CrossRef]
- Nakano, R.; Nakano, A.; Abe, M.; Nagano, N.; Asahara, M.; Furukawa, T.; Ono, Y.; Yano, H.; Okamoto, R. Prevalence and mechanism of fluoroquinolone resistance in clinical isolates of Proteus mirabilis in Japan. Heliyon. 2019, 1, e01291. [Google Scholar] [CrossRef]
- Mahrouki, S.; Perilli, M.; Bourouis, A.; Chihi, H.; Ferjani, M.; Ben Moussa, M.; Amicosante, G.; Belhadj, O. Prevalence of quinolone resistance determinant qnrA6 among broad- and extended-spectrum beta-lactam-resistant Proteus mirabilis and Morganella morganii clinical isolates with sul1-type class 1 integron association in a Tunisian Hospital. Scand J Infect Dis. 2013, 45, 600–5. [Google Scholar] [CrossRef] [PubMed]
- Clinical Laboratory Standard Institution. Performance Standards for Antimicrobial Susceptibility Testing, 28th ed.; Approved Standard M100-S22; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018. [Google Scholar]
- European Committee for Antimicrobial Susceptibility Testing. Breakpoint Tables for Interpretation of MICs and Zone Diameters. Version 12. 2022. Available online: http://www.eucast.org (accessed on 1st October 2023).
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G. ; Harbarth, S; Hindler, J.F; Kahlmeter, G; Olsson-Liljequist, B; Paterson, D.L; Rice, L.B; Stelling J.; Struelens, M.J.; Vatopoulos, A.; Weber, J.T.; Monnet, D. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect 2002, 18, 268–281. [Google Scholar] [CrossRef]
- Davis, R.; Brown, P.D. Multiple Antibiotic Resistance Index, Fitness and Virulence Potential in 408 Respiratory Pseudomonas aeruginosa from Jamaica. J. Med. Microbiol. 2016, 65, 261–271. [Google Scholar] [PubMed]
- Jarlier, V.; Nicolas, M.H.; Fournier, G.; Philippon, A. Extended broad-spectrum beta-lactamases conferring transferable resistance to newer beta-lactam agents in Enterobacteriaceae: hospital prevalence and susceptibility patterns. Rev Infect Dis 1988, 10, 867–878. [Google Scholar] [CrossRef]
- Polsfuss, S.; Bloemberg, G.V.; Giger, J.; Meyer, V.; Böttger, E.C.; Hombach, M. Practical approach for reliable detection of AmpC β-lactamase-producing Enterobacteriaceae. J. Clin. Microbiol. 2011, 49, 2798–803. [Google Scholar] [CrossRef]
- Sanders, C.C.; Sanders, W.E. Jr; Goering, R.V. In vitro antagonism of beta-lactam antibiotics by cefoxitin. Antimicrob. Agents. Chemother. 1982, 21, 968–75. [Google Scholar] [CrossRef]
- Arlet, G.; Brami, G.; Decre, D.; Flippo, A.; Gaillot, O.; Lagrange, P.H.; Philippon, A. Molecular characterization by PCR restriction fragment polymorphism of TEM β-lactamases. FEMS Microbiol. Lett. 1995, 134, 203–208. [Google Scholar] [CrossRef]
- Nüesch-Inderbinen, M.T.; Hächler, H.; Kayser, F.H. Detection of genes coding for extended-spectrum SHV β-lactamases in clinical isolates by a molecular genetic method, and comparison with the E test. Eur. J. Clin. Microbiol. Infect. Dis. 1996, 15, 398–402. [Google Scholar] [CrossRef]
- Woodford, N.; Ward, M.E.; Kaufmann, M.E.; Turton, J.; Fagan, E.J.; James, D.; Johnson, A.P.; Pike, R.; Warner, M.; Cheasty, T.; Pearson, A.; Harry, S.; Leach, J.B; Loughrey, A.; Lowes, J.A.; Warren, R.E.; Livermore, D.M. Community and hospital spread of Escherichia coli producing CTX-M extended-spectrum β-lactamases in the UK. J. Antimicrob. Chemother. 2004, 54, 735–743. [Google Scholar] [CrossRef]
- Robicsek, A.; Jacoby, G.A.; Hooper, D.C. The worldwide emergence of plasmid-mediated quinolone resistance. Lancet Infect. Dis. 2006, 6, 629–640. [Google Scholar] [CrossRef] [PubMed]
- Perez-Perez, F.J.; Hanson, N.D. Detection of plasmid-mediated AmpC β-lactamase genes in clinical isolates by using multiplex PCR. J. Clin. Microbiol. 2002, 40, 2153–2162. [Google Scholar] [CrossRef] [PubMed]
- Woodford, N.; Fagan, E.J.; Ellington, M.J. Multiplex PCR for rapid detection of genes encoding CTX-M extended-spectrum β-lactamases. J. Antimicrob. Chemother. 2006, 57, 154–155. [Google Scholar] [CrossRef] [PubMed]
- Saladin, M.; Cao, V.T.B.; Lambert, T.; Donay, J.L.; Hermann, J.; Ould-Hocine, L. Diversity of CTX-M β-lactamases and Their Promoter Regions from Enterobacteriaceae Isolated in Three Parisian Hospitals. FEMS Microbiol. Lett. 2002, 209, 161–168. [Google Scholar] [CrossRef]
- Zankari, E.; Hasman, H.; Cosentino, S.; Vestergaard, M.; Rasmussen, S.; Lund, O.; Aarestrup, F.M.; Larsen, M.V. Identification of acquired antimicrobial resistance genes. J Antimicrob Chemother. 2012, 67, 2640–4. [Google Scholar] [CrossRef]
- Elwell, L.P.; Falkow, S. The characterization of R plasmids and the detection of plasmid-specified genes. In Antibiotics in Laboratory Medicine, 2nd ed.; Lorian, V., Ed.; Williams and Wilkins: Baltimore, MD, USA, 1986; pp. 683–721. [Google Scholar]
- Carattoli, A.; Bertini, A.; Villa, L.; Falbo, V.; Hopkins, K.L.; Threfall, E.J. Identification of plasmids by PCR-based replicon typing. J Microbiol Methods. 2005, 63, 219–228. [Google Scholar] [CrossRef]
- Carattoli, A.; Seiffert, S.N.; Schwendener, S.; Perreten, V.; Endimiani, A. Differentiation of IncL and IncM Plasmids Associated with the Spread of Clinically Relevant Antimicrobial Resistance. PLoS ONE 2015, 10, e0123063. [Google Scholar] [CrossRef]
Figure 1.
Phylogenetic tree showing relatedness of the ESBL and the AmpC- producing organisms analysed in this study.
Figure 1.
Phylogenetic tree showing relatedness of the ESBL and the AmpC- producing organisms analysed in this study.
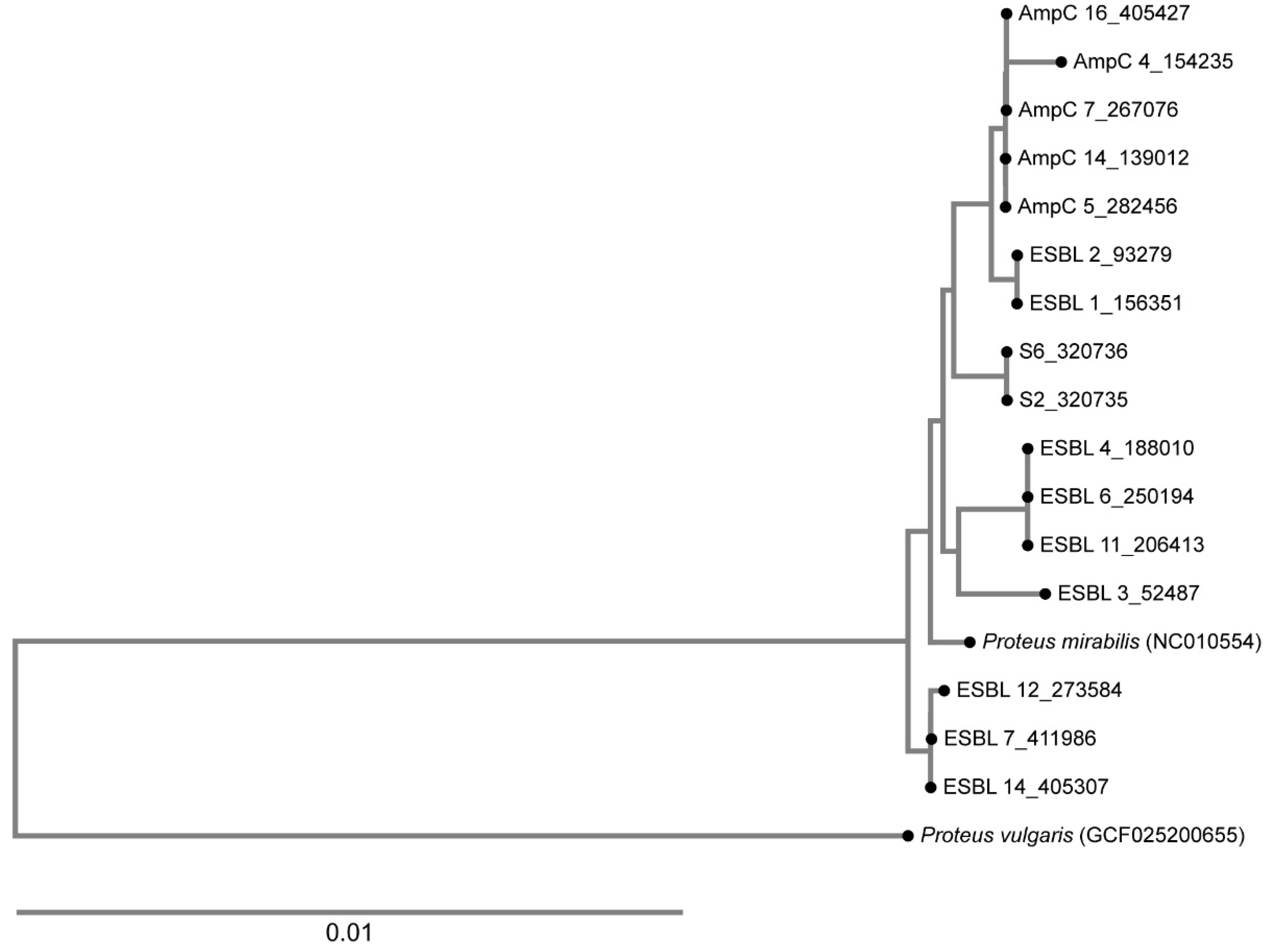
Figure 2.
Phylogenetic tree showing relatedness of the present isolates with those from 2008 which are designated as S6 and S7.
Figure 2.
Phylogenetic tree showing relatedness of the present isolates with those from 2008 which are designated as S6 and S7.


Table 1.
Antibiotic susceptibility of ESBL and p-AmpC-producing P. mirabilis isolates. MIC values are expressed as mg/L.
Table 1.
Antibiotic susceptibility of ESBL and p-AmpC-producing P. mirabilis isolates. MIC values are expressed as mg/L.
| ESBL | AmpC | |||||||
| MIC range | MIC50 | MIC90 | Number and % of resistant isolates | MIC range | MIC50 | MIC90 | Number and % of resistant isolates | |
| amoxycillin-clavulanate | >128->128 | ≥128 | ≥128 | 17/17 (100%) | >128->128 | ≥128 | ≥128 | 22/22 (100%) |
| cefuroxime | >128->128 | ≥128 | ≥128 | 17/17 (100%) | >128->128 | ≥128 | ≥128 | 22/22 (100%) |
| piperacillin-tazobactam | 4-32 | 16 | 32 | 0/17 (0%) | 2-64 | 16 | 64 | 0/22 (0%) |
| ceftazidime | 2->128 | 16 | ≥128 | 10/17(58,8%) | 16->128 | >128 | >128 | 22/22 (95) |
| cefotaxime | 32->128 | ≥128 | ≥128 | 17/17 (100%) | >128->128 | ≥128 | ≥128 | 22/22 (100%) |
| ceftriaxone | 8->128 | 64 | ≥128 | 17/17 (100%) | 32->128 | ≥128 | ≥128 | 17/17 (100%) |
| cefepime | 4-64 | 32 | 64 | 16/17 (94%) | 4-32 | 8 | 32 | 6/22 (27%) |
| imipenem | 0,5-4 | 1 | 2 | 0/17 (0%) | 0,25-1 | 0,5 | 1 | 0/22 (0%) |
| meropenem | 0,06-0,25 | 0,06 | 0,12 | 0/17 (0%) | 0,06-0,25 | 0,12 | 0,25 | 0/22 (0%) |
| gentamicin | 0,25->128 | 32 | >128 | 13/17(76,4%) | 0,25->128 | 64 | >128 | 19/22 (86,3%) |
| amikacin | 8->128 | 32 | 128 | 6/17 (35,2%) | 32->128 | 128 | >128 | 20/22 (91%) |
| ciprofloxacin | 1->128 | 128 | >128 | 17/17 (100%) | 16_>128 | 128 | >128 | 22/22 (100%) |
Table 2.
Inter-array chip CarbaResist Kit results of two representative P. mirabilis isolates.
| Isolate and Protocol Number | β-Lactam | AG | SUL | THR | |
| AmpC 1 (284989) |
blaCMY-16 blaTEM |
aac(6′) aac(3″)-Ia aac(6′)-Ib-cr aadA1 aadA2 |
Sul1 Sul2 |
dfrA1 | |
| ESBL 5 (156351) |
blaCTX-M-9 |
aac(6′) aac(6′)-Ib aac(3″)-IVa aadA1 aadA2 aphA |
Sul1 Sul2 |
dfrA5 |
Abbreviations: AG, aminoglycosides; SUL, sulphonamides; THR, trimethoprim.
Table 3.
Whole genome sequencing of representative isolates (accession number are provided in parenthesis).
Table 3.
Whole genome sequencing of representative isolates (accession number are provided in parenthesis).
|
Isolate and protocol number |
β-Lactam | Aminoglycosides | Sulphonamide | Trimethoprim | Chloramphenicol | Tetracycline | Fluoroquinolones | Plasmid Inc group |
| AmpC 4 (154235) |
blaCMY-16 (AJ781421) |
dfrA1 (X00926) |
||||||
| AmpC 5 (282456) |
blaCMY-16 AJ781421 blaTEM-1b AY458016 |
aac(6′)-Ib3 X60321 aadA1 JX185132 aph(6)-Id M28829 aph(3)-Ia X62115 |
Sul1 U12338 Sul2 HQ840942 |
dfrA1 X00926 |
CatA1 V00622 |
Tet (A) (AJ517790) |
||
| AmpC 7 (267076) |
blaCMY-16 (AJ781421) |
aadA1 (789/789) |
dfrA1 (X00926) |
CatA1 V00622 |
||||
| AmpC 14 (139012) |
blaCMY-16 (AJ781421) blaTEM-1b (AY458016) |
aac(6′)-Ib3 X60321 aadA1 (JX185132) aadA2 (JQ364967) aph(6)-Id (M28829) aph(3)-Ia (X62115) |
Sul1 U12338 Sul2 HQ840942 |
dfrA1 X00926 |
CatA1 V00622 |
Tet (J) (ACLE01000065) |
||
| AmpC 16 (405427) |
blaCMY-16 (AJ781421) blaTEM-1b (AY458016) |
aph(6)-Id (M28829) aph(3)-Ia X(62115) aadA1 (JX185132) aadA2 (JQ364967) aac(6′)-Ib-cr (DQ303918) aph(3″)-Ib (AF321551) armA (AY220558) |
Sul1 U12338 Sul2 HQ840942 |
dfrA1 (X00926) dfrA12 (AM040708) |
CatA1 (V00622) |
Tet (J) (ACLE01000065) Tet (A) (AJ517790) |
||
| ESBL 1 (156351) |
blaCTX-M-101 (HQ398214 |
aph(6)-Id (M28829) aac(3)-IId (EU022314) aadA1 (JX185132) aadA5 (AF137361) aph(3″)-Ia (X62115) aph(3″)-Ib (AF321551) |
Sul1 U12338 Sul2 (HQ840942) |
dfrA1 (X00926) |
cat (M11587) CatA1 (V00622) |
Tet (J) (ACLE01000065) |
||
| ESBL 2 (93279) |
blaCTX-M-15 (HQ398214) blaTEM-1d (AF188200) |
aph(6)-Id (M28829) aac(3)-IId (EU022314) aadA1 (JX185132) aadA5 (AF137361) aph(3)-Ia (X62115) aph(3″)-Ib (AF321551) aac(3)-IV (DQ241380) |
Sul1 U12338 Sul2 HQ840942 |
dfrA1 (X00926) dfrA17 (FJ460238) |
cat (M11587) CatA1 V00622 |
Tet (J) (ACLE01000065) |
||
| ESBL 3 (52487) |
blaCTX-M-32 (AJ557142) |
aph(6)-Id (M28829) aph(3)-Ia (X62115) aph(4)-Ia (V01499) aadA1 (JX185132) aph(3″)-Ib AF024602 |
Sul1 U12338 Sul2 HQ840942 |
dfrA1 (X00926) |
cat (M11587) |
Tet (J) (ACLE01000065) |
qnrD1 FJ228229 |
|
| ESBL 4 (188010) |
blaCTX-M-14b (DQ359215) |
aph(6)-Id (M28829 aac(3)-IIa (X51534) aadA1 (JX185132) aph(3″)-Ib (AF321551) |
dfrA1 (X00926) |
cat (M11587) |
Tet (J) (ACLE01000065) |
|||
| ESBL 6 (250194) |
blaCTX-M-14b (DQ359215) blaTEM-1d (AF188200) |
aac(3)-IIa (X51534) aadA1 (JX185132) aph(3″)-Ib (AF321551) |
Sul2 HQ840942 |
dfrA1 (X00926 |
cat (M11587) |
Tet (J) (ACLE01000065 |
||
| ESBL 7 (411986) |
blaCTX-M-65 (EF418608) blaOXA-1 (HQ170510) |
aph(6)-Id (M28829) aph(3′)-Ia (X62115) aph(4)-Ia (V01499) aac(6′)-Ib-cr (DQ303918) aph(3″)-Ib (AF321551) aac(3)-IV (DQ241380) aadA2b (D43625) |
Sul1 (U12338) Sul2 (HQ840942) |
dfrA32 (GU067642) |
catB3 (U13880) |
Tet (J) (ACLE01000065 tet(C) (AF055345) |
||
| ESBL 11 (206414) |
blaCTX-M-14b (DQ359215)¸ blaTEM-1d (AF188200 |
aph(6)-Id (M28829) aac(3)-IIa (X51534) aadA1 (JX185132) aph(3″)-Ib (AF321551) |
Sul2 (HQ840942) |
dfrA1 (X00926) |
cat (M11587) |
Tet (J) (ACLE01000065) |
||
| ESBL 12 (273584) |
blaCTX-M-65 (EF418608) blaOXA-1 (HQ170510) blaTEM-1A (HM749966) |
aph(6)-Id (M28829) aph(3′)-Ia (X62115) aph(4)-Ia (V01499) aph(3″)-Ib (AF321551) aac(6′)-Ib-cr (DQ303918) aac(3)-IV (DQ241380) aadA2b (D43625) |
Sul1 (U12338) Sul2 (HQ840942) |
dfrA32 (GU067642) |
catB3 (U13880) |
tet(C) (AF055345) tet(H) (Y15510) Tet (J) (ACLE01000065) |
||
| ESBL 14 (405307) |
blaCTX-M-65 (DQ359215) blaTEM-1d (AF188200) blaOXA-1 (HQ170510 |
aph(6)-Id aph(3′)-Ia (X62115) (M28829) aph(3″)-Ib (AF321551 aac(3)-IIa (X51534) aac(6′)-Ib-cr (DQ303918 aadA1 (JX185132) aadA2 (JQ364967) |
Sul1 (U12338) Sul2 (HQ840942 |
dfrA32 (GU067642) |
cat (M11587) catB3 (U13880) |
Tet (J) (ACLE01000065 tet(C) (AF055345) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



