
Submitted:
01 July 2025
Posted:
03 July 2025
You are already at the latest version
Abstract
The Atlantic Forest coastal tablelands is home to a wide diversity of small-sized freshwater fish sharing a biogeographically congruent distributional pattern. The annual killifish Xenurolebias are among these remarkable inhabitants in the riparian wetlands with four species in the area. Xenurolebias pataxo between the Jucuruçu and Mucuri rivers, geographically replaced by Xenurolebias myersi at the Riacho Doce and the Rio Itaúnas, and further south Xenurolebias cricarensis in the floodplains of the Rio São Mateus and Xenurolebias izecksohni, living in temporary environments of the Rio Barra Seca, north of the Rio Doce. Due to their restricted habitat at lowlands, these annual killifish could be all signed as target species, providing benefits in the conservation of riparian wetlands. However, less is known about their habits and real distribution. Otherwise, the accelerated habitat loss is worrisome. Two of these species are threatened with extinction on Red Lists; one is Near Threatened and one Data Deficient. After ten years since the last species description, our investigations revealed a fifth species, the first recorded south of Rio Doce. The present contribution aims to highlight the habitat occupied by each species within the temporary environments, and present an updated distributional data on these fishes. Additionally, a new Xenurolebias is described, the first one recorded on south of Rio Doce.
Keywords:
Atlantic Forest
; Rio Riacho
; swamp
; cloud fish
; endemism
1. Introduction
Among the Rivulidae fish family, the Cynolebiinae contains annual species, or cloud fish, which complete their life cycle regulated by the rainy season. Their lives are short, quickly reaching sexual maturity and dying during the dry season, when the pools that are their habitat dry up. The eggs remain in a state of diapause in the damp mud at the bottom of the puddle and hatch during the rains [1].
The Atlantic Forest in Eastern Brazil have certain stretches of mild relief, comprising a phyto physiognomy called Coastal Tablelands, between Southern Bahia and Northern Espírito Santo. This area is full of small independent streams, lakes, swamps and temporary flooded environments, and these habitats are home to a wide diversity of small-sized fish, often endemic [2,3,4,5].
The annual killifish Xenurolebias are among these remarkable inhabitants in the riparian wetlands with four species restricted to the Coastal Tablelands. These fish appear only in brief periods of the year (5 Costa, 2002). They live in pools of dark, reddish and acidic waters, in the restinga, or on the edges of forests, in the coastal plains of the north of Espírito Santo to the north of the Doce River [[6[]. The habitats of these endemic species are sometimes ephemeral wetlands that require special conservation efforts. The distribution of each taxon is linked to the lowlands of a certain river basin on coastal tablelands bioregion (5 Vieira-Guimarães et al., 2024). In extreme southern Bahia, Xenurolebias pataxo Costa, 2014, between the Jucuruçu and Mucuri rivers, geographically replaced by Xenurolebias myersi [[7[] between the Riacho Doce and the Itaúnas river, in the extreme north of Espírito Santo, and further south Xenurolebias cricarensis Costa, 2014, and in the floodplain of the Rio São Mateus and Xenurolebias izecksohni (Da Cruz, 1983), which occurs in temporary environments of the Rio Barra Seca, north of the Doce River. Due to their restricted habitat at lowlands, the annual killifish of the Xenurolebias could be signed as target species, providing benefits in the conservation of riparian wetlands in the Atlantic Forest. However, less is known about them, their habits, real distribution and human current impacts on its populations.
Two of these species are threatened with extinction on Red Lists; one is Near Threatened and one Data Deficient [8]. After ten years since the last species description, our investigations revealed a fifth species, the first recorded south of the Rio Doce. These fishes inhabiting swamp areas of the Rio Riacho, are distinct from its geographically closest congener on the Rio Barra Seca swamps, and revealed to be a new taxon.
The present study aims to update distributional data on these fishes, evaluate their habitat and describe a new species at lowlands south of Rio Doce.
2. Materials and Methods
2.1. Study Area
The phytophysiognomy of the region is composed of coastal tablelands forest, a formation of the Atlantic Forest with mild relief, sandy soils covered by dense forest and pioneer vegetation, in mostly flat to wavy terrains [9,10,11]. These tablelands include coastal ecosystem formations of vegetation such as restinga, muçununga, and swamps. Water accumulates in flooded areas that, due to the concentration of large quantities of leaves and tannin, display a tea-like or amber coloration. The climate in the region is Am (tropical monsoon), according to the climatic classification system (Köppen – Geiger classification). The annual average temperature is 25 °C, with averages of 28 °C in summer and 21.4 °C in winter. The average annual rainfall is 1,354 mm, with rainy season between October and April.
Coastal floodplains along river basins of Mucuri, Riacho Doce, Rio Itaúnas, Rio São Mateus, Rio Doce and Rio Riacho are temporal subject to floods, generating temporary pools.
2.2. Sample Design and Data Collection
In search of fish populations, expeditions and field campaigns were carried out between December 2021 and March 2025, with visits to the Xenurolebias species type localities, as well as the exploration of additional environments.
Each sampling point was located by GPS (Global Positioning System), and plotted in a map using the GPS Trackmaker Professional program 4.9 [8]. The results of distance and cartographic area were calculated on base maps constructed under the same program.
Collection points were photographed and characterized for environmental conditions, as well as its abiotic parameters. Fish sampling was carried out with seines and sieves. At each point, abiotic parameters were recorded.
The material was fixed in 10% formalin and taken to be deposited in the fish collection to be cataloged. Voucher specimens of the new taxon species were collected and deposited in the ichthyological collections of the Museu Nacional (MNRJ) and the Museu de Biologia Mello Leitão (MBML). All captured individuals were first euthanized before fixation and field methods were approved by the Ethics Committee for Animal Use of Federal University of Espírito Santo (CEUA-UFES; permit number: 008/23).
Abiotic and biotic data were collected and systematized through a field protocol. In addition, the characteristics of the environments and specimens were recorded through photographs. The pH and Salinity measurements were carried out in the laboratory from water samples collected. The other parameters were measured in the field.
Morphometric and meristic data were taken according to [12]; measurements are presented as percent of standard length (SL), except for those related to head morphology, which are expressed as percent of head length (HL). Fin-ray counts include all elements.
The Extension of Occupancy (EOO) for the habitat occupied were estimated with the area of minimum convex polygon [19]. The calculated distributional area, in squares kilometers, is used in the process of assessing the risk of extinction of a certain taxon according to IUCN Red List guidelines. The EOO is one of the key metrics adopted to determine a species conservation status, and the same methodology is adopted and padronized by the Brazilian Ministério do Meio Ambiente in the evaluation of priority areas for conservation of annual fish of the Rivulidae family [22].
3. Results
3.1. Xenurolebias Tupinikin sp. nov. (Figure 1, Figure 2, Figure 3, Figure 4, Figure 5, Figure 6 and Figure 7- Table 1 and Table 2)
Holotype. MNRJ 56004, 23.4 mm SL, Brazil, Espírito Santo State. Aracruz, Rio Riacho basin. Swamp area on Sertão do Riacho Stream, -19,73992S -40,04357W, 26 Jan 2035, B. Pinheiro.
Paratypes. MNRJ 56005, 9, 16.4–23.4 mm SL, Brazil, Espírito Santo State. Aracruz, Rio Riacho basin, Swamp area on Sertão do Riacho Stream, -19,73992S -40,04357W, 26 Jan 2025, B. Pinheiro, collected with the holotype.
Diagnosis. Xenurolebias tupinikin is distinguished from remaining congeners by adult males presenting in life a dark blotch overlapping the median portion of the last stripe on caudal peduncle (Figure 2). Further differs from congeners, except X. myersi, by presenting the dorsal and anal fin borders with black outline. Additionally distinct from X. myersi by body depth 25.9- 27.9 in SL (vs. 29.0- 31.4 in SL).
Differs from X. izecksohni and X. cricarensis by the caudal fin in males with 5-6 bars (vs. 7- 14 bars) and by male head depth 64.5-76.9% of HL (vs. 81.2- 85.6% of head length in males). Further differs from X. pataxo and X. myersi by no yellow spots on distal half of dorsal fin in males (vs. yellow spot present).
Description. Morphometric data available in Table 1. Maximum adult size 23.9 mm SL. Body slender, sub-cylindrical anteriorly, slightly deeper than wide, to compressed posteriorly; greatest body depth at level of pelvic-fin base. Dorsal and ventral profiles gently convex from snout to end of dorsal and anal-fin bases, nearly straight on caudal peduncle. Jaws short, snout blunt. Eye small, positioned on dorsal portion of head side. Extremity of dorsal fin pointed and long in males, rounded to slightly pointed in females; tip of anal fin pointed in males, rounded in females; in males, one or two filamentous rays on tip of dorsal and anal fins reaching vertical between base and middle of caudal fin. Caudal fin rounded or sub-lanceolate in shape. Pectoral fin elliptical, posterior margin reaching vertical between base of third and sixth anal-fin rays in males, reaching urogenital papilla in females. Pelvic fin small, tip reaching base of second or third anal-fin ray in males, reaching base of third anal-fin ray in females. Pelvic-fin bases medially united. Dorsal-fin origin on vertical through base of eighth or nineth anal-fin ray in both sexes. Dorsal-fin rays 17 in males, 12-13 in females; anal-fin rays 21 in males, 21-23 in females; caudal-fin rays 21; pectoral-fin rays 13; pelvic-fin rays 6. Scales small, cycloid. Body and head entirely scaled, except ventral surface of head. Body squamation extending over anterior 25% of caudal-fin base; few scales slightly extending on middle of anal-fin base; no scales on dorsal-fin base. Scales arranged in regular transverse pattern. Two small supraorbital scales. Longitudinal series of scales 27; transverse series of scales 9; scale rows around caudal peduncle 12. Contact organ on each scale of ventral portion of flank in male. Minute papillate contact organs on 2 dorsal-most rays of pectoral fin in males. Single neuromast on each scale of lateral line; 2 neuromasts on caudal-fin base. Cephalic neuromasts: supraorbital 10-12, parietal 1, anterior rostral 1, posterior rostral 1, infraorbital 2 + 16-18, preorbital 3, otic 1, post-otic 2, supratemporal 1, median opercular 1, ventral opercular 2, pre-opercular plus mandibular 23-24, lateral mandibular 5-7, paramandibular 1.

Table 1.
Morphometric data of Xenurolebias tupinikin, sp. nov.
| Xenurolebias tupinikin | |||
| Holotype male |
Male (n=5) |
Female (n=10) | |
| Standard length (mm) | 23.3 | 20.3-23.3 | 16.5-23.9 |
| Percents of standard length | |||
| Body depth | 26.9 | 25.9-27.9 | 19.0-32.9 |
| Caudal peduncle depth | 14,3 | 11.4-14.4 | 6.6-14.7 |
| Predorsal length | 59.0 | 56.6-64.1 | 48.8-73.9 |
| Prepelvic length | 44.0 | 42.7-49.4 | 44.1-56.2 |
| Length of dorsal-fin base | 24.2 | 24.2-29.2 | 09.1-16.8 |
| Length of anal-fin base | 51.0 | 47.6-56.1 | 17.8-26.1 |
| Caudal-fin length | 40.7 | 37.3-40.7 | 29.2-39.5 |
| Pectoral-fin length | 32.9 | 27.8-33.2 | 15.8-29.7 |
| Head length | 29,9 | 29.2-31.5 | 28.5-33.8 |
| Percents of head length | |||
| Head depth | 76.8 | 64.4-76.8 | 61.27-85.5 |
| Lower jaw length | 25.5 | 18.4-25.5 | 15.1-21.6 |
| Eye diameter | 32.2 | 31.5-46.1 | 10.8-34.8 |
Live coloration. Male (Figure 3a). Side of body brownish yellow, with 12-13 dark brown bars; caudal peduncle with a dark spot united to posterior-most bar. Dorsum pale brown. Venter pale pink. Opercular and infraorbital region yellowish, with two difuse dark bars. Iris light yellow with dark reddish brown bar on middle part. Dorsal fin dark brown with scattered irregular marks and fin margin becoming dark towards its tip. Anal fin dark brown to dark red, with irregular marks. Caudal fin pale yellow, with 5-6 dark irregular bars. Pectoral fin hyaline with a browinsh border. Pelvic fin light brown.
Female (Figure 3b). Side of body light brown, with 10-11 dark grey bars; one or two black blotches on anterocentral part of flank. Dorsum pale brown. Venter light pink. Opercular region yellowish. Iris light yellow, with dark brownish bar. Unpaired fins hyaline with small dark brownish spots. Paired fins hyaline.
Distribution. The Riacho River corresponds to the first independent coastal river basin south of the Rio Doce. Its tributaries are mostly small watercourses, with predominantly sandy substrate and banks covered by riparian vegetation in different states of conservation. The largest tributary of Rio Riacho corresponds to the Rio dos Comboios, which runs in parallel to coast line until meeting the River Riacho close to its mouth.
The new species is known from three localities, on two tributaries of the rio Riacho, central north of Espírito Santo, Brazil (Figure 4- P1 to P3). The Sertão do Riacho stream and also found in neighbor stream, the Caipora stream, all contributors to the right margin of the Rio Riacho, a coastal river drainage south of the Rio Doce, Espirito Santo (Figure 4).
Ecological notes. The Sertão do Riacho isolated pool, identified as an extensive swamp area, with approximately 2 km². Completely covered by dense emerging vegetation, formed mainly by grasses and ferns. The substrate is formed by emerging vegetation, decomposing plant material and sand (Figure 5). Abiotic information. Dark tea colored waters in a lentic environment. Water depth at pool between 40 cm to 60 cm, Humidity: 60% / Water temperature: 27.1ºC / pH: 4.3-6.2 / Salinity (ppm): 0 (Table 2).
The swampy area at type locality (Figure 4- blue X) is about 100 m from point 1 (Figure 4- yellow circle). It features dense emergent vegetation, marginal vegetation, and floating macrophytes. The swamp showed an abundance of Xenurolebias tupinikin, adults and juveniles, scattered throughout the swampy area, found mainly under vegetation.
Sampling was carried out in various parts of the pool, between edges and central part, with some limitation due to the dense vegetation that emerged from the bottom of the pool. Only adult individuals were captured, three females and one male for record. Additional specimens were immediately returned to the environment. As for the accompanying fauna, aquatic insects in a wide variety and a specimen of Callichthys callichthys was captured.
Etymology. The specific name is a reference to the Tupiniquim indigenous people, inhabitants of lowlands at central north of Espírito Santo. On left margin of the Rio Comboios is located the indigenous land Comboios (Figure 4- green stripes near coast), which correspond to the nearest human occupation relative to the environments inhabited by these fishes. A noun in apposition.
Conservation concerns. The estimated Extent of occupancy (EOO) of the population found in Riacho river basin was defined as 0.096 Km2 (two locations)(Figure 6). From May until September, the pools where the rivulids were captured become dry.The Sertão do Riacho pool is near the road to the village of Riacho, and quite anthropized. At the site, garbage dumping points (solid waste) and debris (civil construction waste) were identified, in addition to sand extraction. The distance from the pool to the sea is approximately 4 km in straight line (Figure 6).
3.2. Xenurolebias izeckshohni (Figure 8, Figure 9 and Figure 10)
Xenurolebias izecksohni was described for the Barra Seca river basin, at the locality of Canto Grande, Farias, within the Vale Natural Reserve (RNV), a private preserved area [13,14]. Until now, the distribution of the species was limited to the Barra Seca river basin. Between 2023 and 2025, field campaigns were carried out in search of populations. New populations were found at the Doce River basin, considerably expanding its area of occurrence to include two river basins- Doce and Barra Seca (see map on Figure 7). The collections were carried out in different periods, covering both the dry and rainy seasons, which allowed the measurement of important data hitherto unknown for the species. It was observed that the life cycle of the species is configured as biannual, and not annual, as expected. That is, throughout the year, the puddles dry up and reestablish themselves twice, usually between November and March the first cycle, and between May and August the second, and there may be variations according to the climatic conditions of each year. In addition, it was found that puddles completely exposed to the sun dry out more quickly, while those with vegetation cover tend to persist for longer, and may even remain flooded throughout the year, which maintain populations alive in these environments.
Most of the new populations of X. izecksohni registered are distributed in private areas, including rural properties and lands of the Linhares Agroindustrial Plant - LASA, an ethanol producing company. In addition to places around the RNV, in the vicinities of the type locality. The number of individuals per pond varied between a few specimens (1 to 5) and concentrations of more than fifty individuals. The morphometric analysis revealed a considerable variation in the standard length (SL), with values higher than those recorded in the literature for the species (30–43 mm)[14]. Males with up to 54.24 mm and females with up to 41.36 mm were measured, evidencing the occurrence of specimens larger than those previously described. Behaviors observed in the field indicate a preferential occupation of the edges of the puddles by Xenurolebias izecksohni, especially under vegetation cover or between submerged leaves. The color of the individuals varied according to the shade of the water, which, in many cases, presented a dark color due to the presence of tannins from the decomposition of organic matter. Under these conditions, the fish showed equally darkened coloration, which may indicate phenotypic plasticity and adaptation to the environment, possibly related to camouflage mechanisms and protection against predators.
Native Vegetation and Landscape. In view of the new records, it was possible to verify that the habitats occupied by Xenurolebias izecksohni present considerable variation in their structural aspects. Individuals have been found in different types of temporary aquatic environments, including puddles, ponds, and even intermittent streams. Such environments varied in terms of vegetation cover, with conditions of total exposure, partial shading or complete vegetation cover. In addition, the composition of the vegetation was also diverse, with records of marginal, floating and emerging plants. The puddles, specifically, showed great variability in terms of morphology (circular, elongated and amoeboid shapes), substrate (sandy, leafy or mixed), depth and color of the water, revealing a considerable ecological plasticity of the species. Puddles in open areas exposed to the sun showed clear waters and emergent herbaceous or aquatic vegetation, while those in areas shaded by shrub or forest vegetation exhibited darker waters and substrate rich in organic matter. Abiotic information. Dark tea colored waters in a lentic environment. Water depth at pool between 20 cm to 70 cm, Humidity: 60%-88% / Water temperature: 22ºC- 37ºC / pH: 3.5 – 5.6 / Salinity (ppm): 0 – 2 (Table 2).
Similar to what occurs with many species of rivulids (15-16), Xenurolebias izecksohni occupies ephemeral habitats, with very peculiar characteristics, becoming inhospitable to most fish species. This condition was observed throughout the expeditions, in which records of coexistence of X. izecksohni with other species of ichthyofauna were rare. In less than 10% of the sampled points, specimens of other species, such as Astyanax spp., were collected. Hoplias malabaricus, Hoplerythrinus unitaeniatus and Callichthys callichthys, usually in larger, newly formed puddles, and close to springs or perennial watercourses. In these cases of coexistence with other fishes, the density of X. izecksohni was considerably reduced, suggesting a possible negative impact of the presence of predators or competition for resources. In contrast, in the smaller, more isolated pools, in which X. izecksohni was often the only species recorded, the population density was significantly higher, indicating a possible adaptive specialization to environments with low biotic pressure.
In newly formed puddles after rain events, newly hatched fish were recorded, which indicates recent reproductive episodes and reinforces the importance of these temporary environments ass nursery for the reproductive success of the species. In addition, an unusual observation was the simultaneous capture of individuals at different stages of development, including fry and adults living in the same environment. Such coexistence is uncommon among rivulids, whose life cycle usually involves synchronization between hatching and periods of temporary flooding, with no overlap of generations [17,18]. These data may indicate local phenological variations or plastic responses to changes in environmental conditions.
Conservation concerns. The estimated Extent of occupancy (EOO) of the population found in both Doce and Barra Seca river basins was defined as 173.6 Km2 (more than 5 locations)(Figure 10). The pools where the rivulids occurs were full of water at least twice a year. The current estimated distribution area is approximately 385 km², with a sampling perimeter of 95.6 km, demonstrating a significant expansion of the previously known distribution. Xenurolebias izecksohni is listed as "Vulnerable" by the [19]. Otherwise, the new information obtained indicates that its situation may be still delicate for its environments than previously estimated, especially in the face of intense anthropogenic pressure on unprotected private areas, where the species was widely recorded during this study.
3.3. Xenurolebias cricarensis (Costa, 2014) (Figure 11, Figure 12 and Figure 13)
Xenurolebias cricarensis is known to occur in its type locality only, a temporary swamp located inside a farm, near road from BR-101 to Conceição da Barra city, north of Espírito Santo.
In 2024, two expeditions were carried out in search of the species. During dry season, on June, an expedition visited the type locality, no specimens were found, since the environment was completely dry. Other points nearby were visited, but were unsuccessful. A second expedition was carried out on wet season, between November 23 and 29, 2024, in the same region. This time the pond was flooded, and populations were recorded, the first findings since the species description.
During the wet season field campaign, 14 specimens of X. cricarensis, were collected, along with the accompanying fauna composed of aquatic insects, tadpoles and mollusks. Specimens of Oreochromis niloticus were also captured. Additional areas, with potential occurrence were visited, but the only with populations is theat single puddle in the sub-basin of the São Domingos stream.
Considering that X. cricarensis is dependent on temporary pools, its persistence is directly related to climatic conditions and the integrity of these environments. Degradation can lead to the local extinction of the species, which highlights the need for urgent actions, such as the creation of conservation units and the awareness of landowners about the ecological importance of these systems.
Native Vegetation in the Landscape. The environment in São Domingos is a large temporary swamp, with approximately 3 km². with dark colored acidic waters, clay soil, and a substrate with decomposing leaves and branches. These characteristics refer to the initial portion of the puddle, which is easier to access, located next to the road. The bottom portion was not accessible and remains to be investigated. Abiotic information. Dark tea colored waters in a lentic environment. Water depth at pool between 50 cm to 60 cm, Humidity: 68% / Water temperature: 24.6ºC / pH: 5.6 (Table 2).
Conservation concerns. The estimated Extent of occupancy (EOO) of the population found in São Mateus river basin corresponds to a single location (Figure 13). Only one pool have populations. From May until the beginning of the rainy season, the pool was completely dry. Categorized as "Data Deficient" (DD) by the [8], the recent data obtained reinforces that Xenurolebias cricarensis needs to be categorized as threatened with extinction. The species has only been found in a single puddle in the sub-basin of the São Domingos Stream with approximately 85 thousand square meters, totalling an approximate occupation area of 0.15 km².
This small area is largely anthropized, with the detection of the practice of cattle grazing in the wetlands. Additionally, such puddle is in the vicinities of the city, and subject to urban pressure. These evidences reinforces the urgent need to create protected areas around the local wetlands habitats. Coexistence with exotic species, such as tilapia, may also compromises population viability due to competition and habitat degradation.
3.4. Xenurolebias myersi (Figure 14, Figure 15 and Figure 16)
The Itaúnas cloud fish inhabits the sub-basins of the Velha Antônia stream, Moças stream, in the Itaúnas river basin and additionally in the Limo stream, a tributary of Doce creek basin [20,21]. These bright colored fishes are sexually dimorphic. Lives in peculiar environments, where water is transparent, translucent, in a yellowish or dark orange tone, without turbidity.
Native Vegetation in the Landscape. The species inhabits seasonal swamps in open areas of taboal (Typha spp.) or in restinga forests. They are found in temporary freshwater floods, of varying size, from small defined puddles with tens of meters to very extensive swamp areas, with a few square kilometers. Such environments are in the floodplain of the rivers, some very close to the sea, just over a hundred meters from the beach. The puddles are shallow, between 20 and 70 cm deep. The bed of the marsh is composed of a triple layer of substract: leaf litter, red mud, which occupies about 20 centimeters or more deep, and then sand underneath. Abiotic information. Dark tea colored waters in a lentic environment. Water depth at pool between 40 cm to 110 cm, Humidity: 60% / Water temperature: 27.1ºC / pH: 3.5-5.6/ Salinity (ppm): 0 (Table 2).
Conservation concerns. The estimated Extent of occupancy (EOO) of the population found in both Itaúnas and Riacho Doce river basins was defined as 12.879 Km2 (more than 5 locations)(Figure 16). The pools where the rivulids occurs were full of water at least twice a year. Xenurolebias myersi is endemic to coastal lowland swamps and restinga environments at Itaúnas village and the Riacho Doce locality. Categorized as "Endangered" (EN), in both on the national red list [8] and on the State red list of Espírito Santo [18]. The species is also included in the Action Plan to conservation of Rivulidae fishes [22].
Its noticeably that few populations were recorded inside the Itaúnas State Park, and mostly of them are found in environments outside the limits of the park, are susceptible to impacts, running imminent risks of extirpation. Attention should be given to the village of Itaúnas, that recently undergone several modifications, driven by tourism and immobiliary pression, urban expansion as well as the pressures of monocultural Eucalyptus plantations in its surroundings.
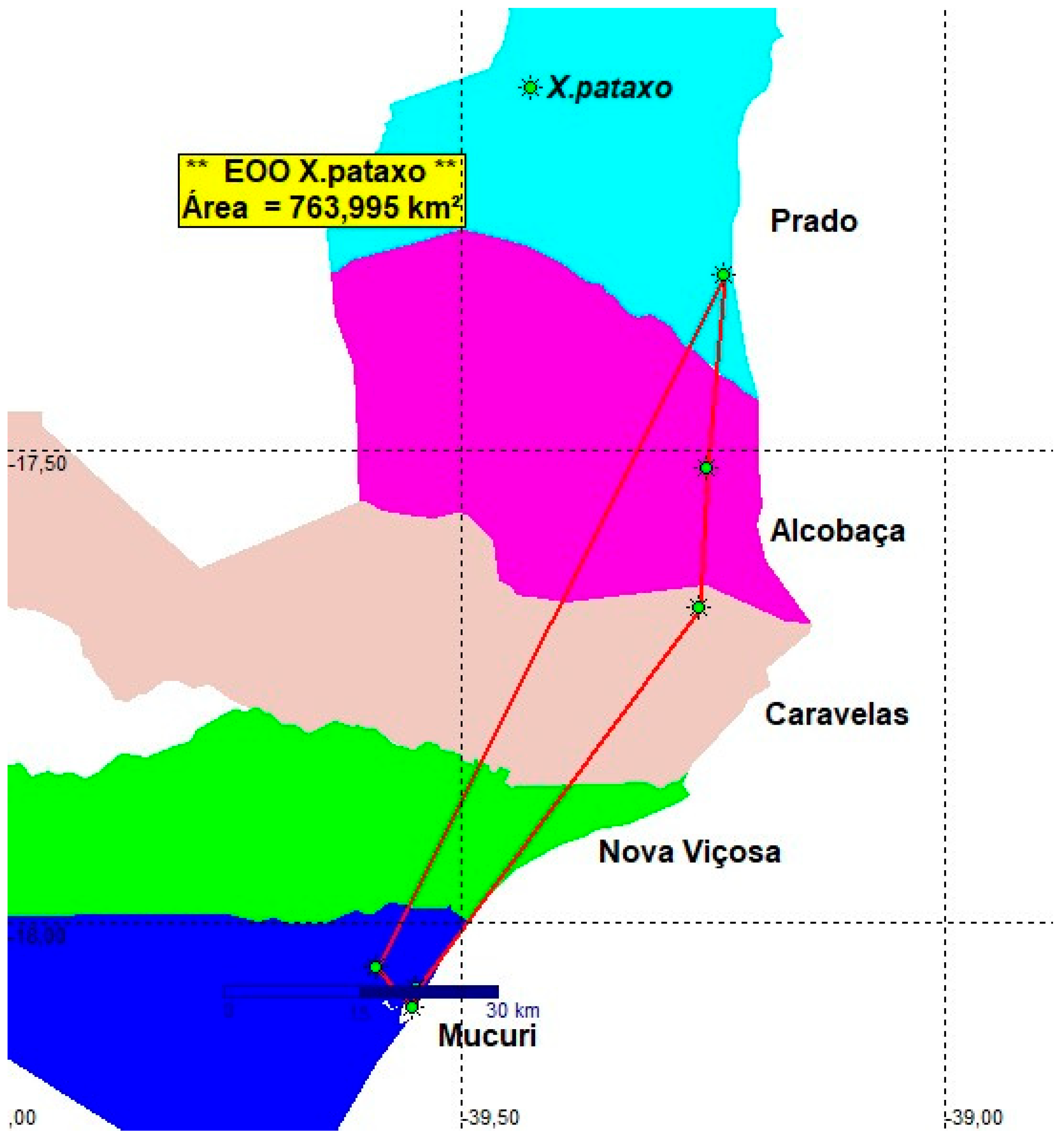
3.5. Xenurolebias pataxo (Figure 17, Figure 18 and Figure 19- Table 2)
Xenurolebias pataxo is recorded from a broad coastal plain area along coastal tabletands formation at southern Bahia between the Jucuruçu river basin and the Mucuri river basin, about 95 km in a straight line [1][]. The area is situated within a touristic zone, which extends to the town of Porto Seguro, one of the most popular touristic destinations in South America. As a consequence, the natural habitats have undergone a great decline since the 1980s, with remarkable habitat loss in the last two decades following intensive urbanization. Populations recorded between 1989 and 1998, situated in the Jucuruçu, Itanhém and Peruípe river basins, as well as some populations previously found around the village of Mucuri close to the Mucuri river mouth, have disappeared. Collecting efforts between Prado and Peruípe were not successful in finding these fishes. A population was detected on Itanhém river lowlands in 2007 (Figure 14).
Conservation concerns. The estimated Extent of occupancy (EOO) of the population found between Mucuri and Jucuruçu river basins was defined as 763.995 Km2 (5 locations)(Figure 19). All records were sampled more than ten years ago. Nowadays the species is found at a single location only, the type locality. Although the original area of occupancy was about 200 km², today X. pataxo is found in a small area about 10 km². The abrupt and extensive environmental decline indicates that X. pataxo is not at least Vulnerable. The whole area is in need of additional investigation.
Table 2.
Abiotic data for Xenurolebias species.
| Environmental features | Values per species | ||||
| X. myersi | X. izecksohni | X. pataxo | X. cricarensis | X. tupinikin | |
| Occupation Area (m²) | 60 | 385 | 10 | 3 | 1.58 |
| EOO (Km²) | 12.879 | 173.6 | 763.995 | NA* | 0.096 |
| Depth (cm) | 40-110 | 20-70 | 30-50 | 50-60 | 30-60 |
| Temperature (ºC) | 24-28 | 22-37 | 25 | 24.6 | 26.5-29.2 |
| pH | 3.5-5.6 | 3.5-5.6 | 5.2 | 5.6 | 4.3-6.2 |
| humidity (%) | 60-80 | 60-88 | NE* | 68 | 54-60 |
| Salinity | 0 | 0-2 | 0 | NE* | 0 |
*NA. Not Applicable (single location). *NE. Not Evaluated.
4. Discussion
The Xenurolebias tupinikin is recognized as a member of genus [14] by possessing sexual dimorphism in number of dorsal and anal-fin rays and numerous supraorbital neuromasts (9 or more); and the 3 synapomorphies uniquely shared by species of Xenurolebias: presence of filamentous rays on the posterior margin of the anal fin in females, caudal fin sub-lanceolate to lanceolate and asymmetric due to an expansion of the ventral portion of the fin in males, and a dark reddish bar on the posterior portion of the caudal peduncle posteriorly extending over the dorsal and ventral margins of the caudal fin in males [23].
The Rivulidae family is considered one of the most endangered among freshwater fishes [25], an even more critical condition for species with an annual life cycle, which depend directly on the rainfall regime to complete their development. These fish inhabit temporary environments that are highly susceptible to fragmentation and degradation, which makes their populations extremely vulnerable. Such factors should be carefully considered in the conservation status assessments of the species.
All Xenurolebias species are endemic to coastal tablelands domain in Atlantic Forest. Two of these species are threatened with extinction on Red Lists (X. myersi- EN and X. pataxo- VU); one is Near Threatened (X. izecksohni- NT) and one Data Deficient (X. cricarensis- DD). Most of these fishes figure out on brazilian redlists [25] state list of Espírito Santo [26] and state list of Bahia [27].
The National Action Plan for the conservation of endangered rivulid fish - PAN Rivulids [23] highlights in its specific objective 3 the action of popularization of the species, through "disseminating knowledge about the focal species of rivulids, sensitizing society about the importance of wetlands for their conservation" [25]. Additionally, in Espírito Santo state it is a species benefited in the Capixaba-Gerais Territorial Action Plan – PAT Capixaba- Gerais [24].
5. Conclusions
In last decades, species of Xenurolebias have been the focus of taxonomic review and molecular studies [14,29]. Despite of these, knowledge about these annual fish populations and their environment conditions preferences are still poorly known. Data on geographic distribution is often limited to that appearing in their original descriptions. Citizen science consists of the partnership between the residents of the place and the researchers in the discovery of novelties and collection of data for scientific research. The dialogue of knowledge, in which the community already knows about the place these fish lives, and the protagonism of local actors, are sources of knowledge that were explored in Xenurolebias myersi [22,23,24]. Associating traditional knowledge with the technical-scientific nature can also enhance conservation actions for other species, through social participation in the construction of knowledge. We hope present contribution may inspire future works on habitat and people interactions towards the survival of these species.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: Records of fish specimens listed for this study.
Author Contributions
All authors designed the study; BPG, LMSS, RFMP participated on fieldwork. RFMP built the species vochers list and checked species distribution ranges. LMSS performed data curation and taxonomic validation. Statistical analysis was performed by GRL. All authors wrote the manuscript.
Funding
Fieldwork was in part funded by GEF Pró Espécies, process no. .
Institutional Review Board Statement
The animal study protocol was approved by the Ethics Committee for Animal Use of Federal University of Espírito Santo (CEUA-UFES); permit number: 008/23 for studies involving field methods.
Data Availability Statement
Data is available in Supplementary Material Table S1.
Acknowledgments
We extend our gratitude to Rodrigo Damásio, Mateus Nogueira Gama Filho, Vinicio Martins, Juca Damásio and Aurikson Correa for their help with field activities. We are grateful to C. Moreira, M.R. Britto, P.A. Buckup (MNRJ) and J. P. da Silva Novelli (MBML) for providing collection records and for the courtesy extended during the visit to their institutions. Thanks to the SAPI - Sociedade de Amigos por Itaúnas team for partnership. To the team of Parque Estadual de Itaúnas, in special to Juliana, Gustavo and Savana at IEMA-Insituto Estadual do meio Ambiente for logistic support during fieldwork. To Izabel Correa Boock de Garcia for the exchange of ideas within the scope of PAN Rivulídeos. This work received laboratory support from the Instituto Nossos Riachos (INR). To ICMBio for the license for field activities. Our gratitude to the population of the village of Itaúnas for the partnership and cordiality.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results”.
References
- Furness, A.I. The evolution of an annual life cycle in killifish: adaptation to ephemeral aquatic environments through embryonic diapause. Biol Rev 2015, 1, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Bizerril, C.R.S.F. Análise taxonômica e biogeográfica da ictiofauna de água doce do leste brasileiro. Acta Biol. Leopoldensia 1994, 16, 51–80. [Google Scholar]
- Menezes, N.A.; Weitzman, S.H.; Oyakawa, O.T.; Lima, F.C.; Castro, R.M.C.; Weitzman, M.J. Peixes de água doce da Mata Atlântica. Neotrópica: São Paulo, Brazil, 2007; 407 p.
- Sarmento-Soares, L. M.; Martins-Pinheiro, R. F. A fauna de peixes nas bacias do norte do Espírito Santo, Brasil. Sitientibus Cien. Biol 2012, 12, 27–52. [Google Scholar] [CrossRef]
- Vieira-Guimarães, F.; Sarmento-Soares, L. M.; Martins-Pinheiro, R. F. Freshwater fishes of the Northeastern Mata Atlântica ecoregion, Brazil: an updated checklist with distributional patterns of a highly endemic ichthyofauna. Zootaxa 2024, 5475 1, 1–72. [Google Scholar] [CrossRef]
- Costa, W.J.E.M. Peixes anuais brasileiros: diversidade e conservação. Editora, Universidade Federal do Paraná, Curitiba, Brazil, 2002, 180 p.
- Carvalho, A.L. Um novo peixe anual do estado do Espírito Santo (Pisces, Cyprinodontidae, Rivulinae). Revta Brasil Biol 1971, 31, 401–404. [Google Scholar]
- Ferreira JO. GPS TrackMakerPRO Version 4.9.610 GeoStudio Technology. 2022.
- Sarmento-Soares, L. M.; Martins-Pinheiro, R. F. A fauna de peixes na REBIO Córrego Grande e seu entorno direto, Espírito Santo, Brasil. Bol Mus Biol Mello Leitão 2013, 31, 25–57. [Google Scholar]
- Sarmento-Soares, L. M.; Martins-Pinheiro, R. F. A fauna de peixes na bacia do rio Barra Seca e REBIO de Sooretama, Espírito Santo, Brasil. Bol Mus Biol Mello Leitão 2014, 35, 49–104. [Google Scholar]
- Vieira-Guimarães, F.; Sarmento-Soares, L. M.; Martins-Pinheiro, R. F.; Duboc, L.F. Assessment of Stream Environmental Condition Using Fishbased Metrics In A Protected Area And Its Disturbed Buffer Zone, Northeastern Atlantic Rainforest. Oecol. Aust. 2022, 26, 461–475. [Google Scholar] [CrossRef]
- Costa, W.J.E.M. 1995. Pearl killifishes, the Cynolebiatinae: systematics and biogeography of the neotropical annual fish subfamily (Cyprinodontiformes: Rivulidae). TFH, Neptune City, NJ, USA.
- Cruz, C.A.G. Uma nova espécie de Cynolebias do estado do Espirito Santo, Brazil (Pisces, Cyprinodontidae). Pap Avul Zoologia, São Paulo 1983, 35, 73–77. [Google Scholar] [CrossRef]
- Costa, W.J.E.M.; Amorim, P.F. Integrative taxonomy and conservation of seasonal killifishes, Xenurolebias (Teleostei: Rivulidae), and the Brazilian Atlantic Forest. Syst Biodiv 2014, 12, 350–365. [Google Scholar] [CrossRef]
- Costa, W.J.E.M. Peixes anuais brasileiros: diversidade e conservação. Editora, Universidade Federal do Paraná, Curitiba, Brasil. 2002.
- Volcan, M.V.; Gonçalves, A.C.; Lanés, L.E.K.; Guadagnin, D.L. Annual Fishes (Rivulidae) from Southern Brazil: A Broad-Scale Assessment of Their Diversity and Conservation. In: Berois N, García G, de Sá RO, editors. Annual Fishes: life history strategy, diversity, and evolution. Boca Ratón: CRC Press; 2016. p.185-203.
- Guedes, G.H.S. The recapture of Leptopanchax opalescens (Aplocheiloidei: Rivulidae), a critically endangered seasonal killifish: habitat and aspects of population structure. Zoologia Curitiba 2020, 37, e54982. [Google Scholar] [CrossRef]
- Severo-Neto, F.; Volcan MV, Population dynamics of Melanorivulus rossoi, a restricted geographic distribution killifish species. Environ Biol Fish. [CrossRef]
- IUCN. (2025). The IUCN Red List of Threatened Species. Version 2024-1. https://www.iucnredlist.org accessed June 21, 2025. IUCN.IUCN SSC Red List Technical Working Group, 2021.
- Sarmento-Soares, L.M.; Castro, R.D.R.; Martins-Pinheiro, R.F.; Ledermann, M.R.; Silva, J.P. O peixinho das nuvens Xenurolebias myersi (Carvalho, 1971) (Cyprinodontiformes: Rivulidae) reaparece em Itaúnas, Conceição da Barra, Espírito Santo, Brasil. Bol Soc Brasil Ictiol 2022, 140, 17–29. [Google Scholar]
- Sarmento-Soares, L.M., Castro, R.D.R., Martins-Pinheiro, R.F. ; Garcia, I.B. (2024). Ways to Protect the Environments of the Itaúnas Cloud Fish - Xenurolebias myersi (Carvalho, 1971) - Inhabitant of the Restinga Swamps, Conceição Da Barra, Northern Espírito Santo, Southeastern Brazil. Qeios CC-BY 4.0: 1-6. [CrossRef]
- MMA-Ministério do Meio Ambiente. Portaria MMA Nº 148, de 7 de Junho de 2022. Alteração dos Anexos da Portaria Nº 445, referentes à atualização da Lista Nacional de Espécies Ameaçadas de Extinção. Anexo III. MMA, Brasília.[unknown pagination].
- Costa, W.J.E.M. Descriptive morphology and phylogenetic relationships among species of the Neotropical annual killifish genera Nematolebias and Simpsonichthys (Cyprinodontiformes: Aplocheiloidei: Rivulidae). Neotrop Ichthyol 2006, 4, 1–26. [Google Scholar] [CrossRef]
- IEMA - Instituto Estadual De Meio Ambiente E Recursos Hídricos. (2021). Plano de Ação Territorial para Conservação de Espécies Ameaçadas de Extinção do Território Capixaba-Gerais: Sumário Executivo. Cariacica: IEMA.
- MMA - Ministério Do Meio Ambiente. Livro Vermelho da Fauna Brasileira Ameaçada de Extinção: Volume VI - Peixes. Brasília: MMA/ICMBio 2018.
- Hostim-Silva, M.; Duboc, L.F.; Pimentel, C.R.; Vilar, C.C.; Machado, D.F.; Dario, F.D., Guimarães, F.V., Pinheiro, I.E.G., Adelir-Alves, J., Musiello- Fernandes, J., Santander-Neto, J., Nunes, J.A.C.C., Silva, J.P., Ingenito, L.F.S., Sarmento- Soares, L.M., Britto, M.R., Lopes, M.M., Freitas, M.O., Buckup, P.A., Martins-Pinheiro, R.F., Silva, T.G., Volpi, T.A.; Giglio, V.J. (2019). Peixes ameaçados de extinção no estado do Espírito Santo, p. 230-255. In: Fraga, C.N., Formigoni, M.H.; Chaves, F.G. (Orgs). Fauna e flora ameaçadas de extinção no estado do Espírito Santo. Santa Teresa: Instituto Nacional da Mata Atlântica.
- Silva, A.T., Chagas, R.J., Santos, A.C.A., Zanata, A.M., Rodrigues, B.K., Polaz, C.N.M., Alves, C.B.M., Vieira, C.S., Souza, F.B., Vieira, F., Sampaio, F.A.C., Ferreira, H., Alves, H.R.A., Sarmento-Soares, L.M., Pinho, M., Martins-Pinheiro, R.F., Lima, S.M.Q., Campiolo, S., Camelier, P. Freshwater fishes of the Bahia State, Northeastern Brazil. Biota Neotropica 2020, 20, e20200969. [CrossRef]
- ICMBio- Instituto Chico Mendes de Conservação da Biodiversidade. Sumário Executivo do Plano de Ação Nacional de Conservação de Peixes Rivulídeos Ameaçados de Extinção 2013; www.icmbio.gov.br/portal/images/stories/docs-plano-de-acao/pan-rivulideos/sumario-executivo-rivulideos.pdf [accessed Jun 21 2025].
- Costa, W.J.E.M. Taxonomic revision of the seasonal South American killifish genus Simpsonichthys (Teleostei: Cyprinodontiformes: Aplocheiloidei: Rivulidae). Zootaxa 2007, 1669, 1–134. [Google Scholar] [CrossRef]
Figure 1.
Xenurolebias tupinikin, A. MNRJ 56004, male, holotype, 23.4 mm SL; B. MNRJ 56005, female, paratype, 35.4 mm SL. Pool at flooded area on Sertão do Riacho stream, Aracruz, Espírito Santo, Brazil.
Figure 1.
Xenurolebias tupinikin, A. MNRJ 56004, male, holotype, 23.4 mm SL; B. MNRJ 56005, female, paratype, 35.4 mm SL. Pool at flooded area on Sertão do Riacho stream, Aracruz, Espírito Santo, Brazil.

Figure 2.
Diagnostic features for Xenurolebias tupinikin. Dark blotch overlapping the median portion of the last stripe on caudal peduncle.
Figure 2.
Diagnostic features for Xenurolebias tupinikin. Dark blotch overlapping the median portion of the last stripe on caudal peduncle.

Figure 3.
Xenurolebias tupinikin. A. Male. B. Famale.

Figure 4.
Sampling localities for Xenurolebias tupinikin. P1- Sertão do riacho stream. P2- Type locality (blue X). P3- Temporaty pool on wetlands of Caipora stream. Green striped area between left margin of Rio dos Comboios and ocean corresponds to indigenous land Comboios of Tupiniquim people (green tree).
Figure 4.
Sampling localities for Xenurolebias tupinikin. P1- Sertão do riacho stream. P2- Type locality (blue X). P3- Temporaty pool on wetlands of Caipora stream. Green striped area between left margin of Rio dos Comboios and ocean corresponds to indigenous land Comboios of Tupiniquim people (green tree).
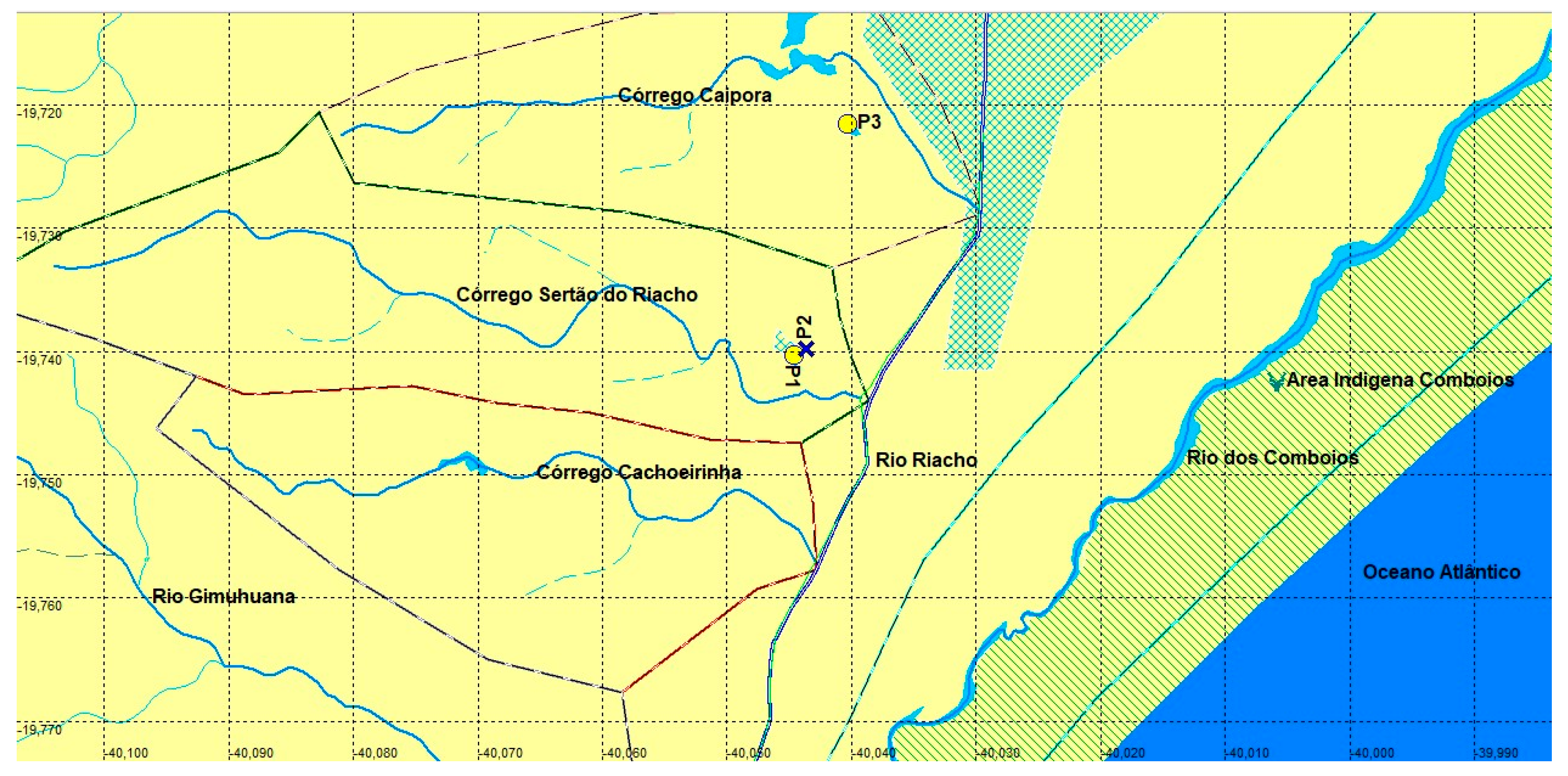
Figure 5.
Sertão do Riacho stream. Type locality of Xenurolebias tupinikin.

Figure 6.
Estimates of EOO, cartographic area and the number of locations for Xenurolebias tupinikin.
Figure 6.
Estimates of EOO, cartographic area and the number of locations for Xenurolebias tupinikin.
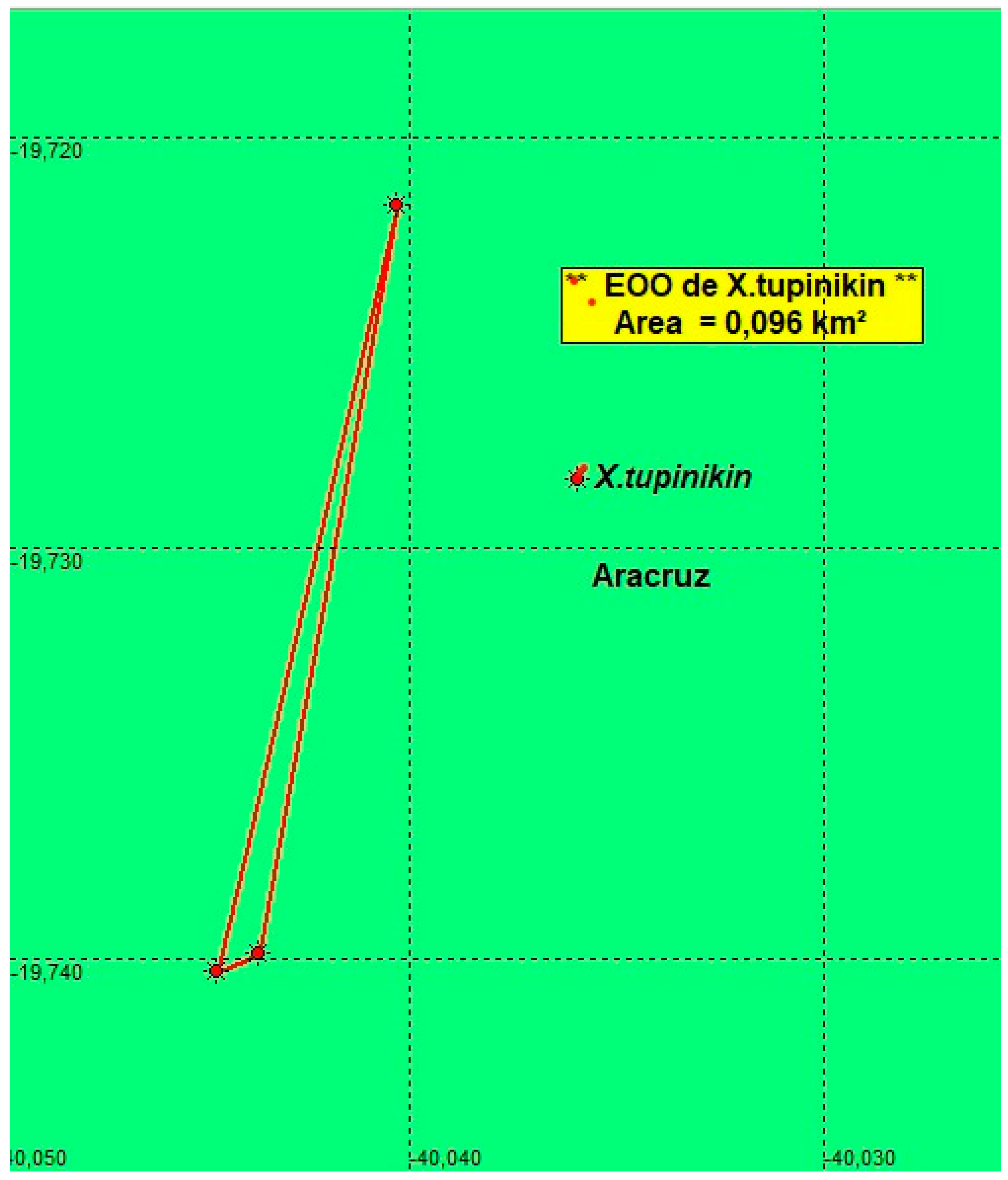
Figure 7.
Study area location within the coastal tablelands between Bahia and Espírito Santo, Brazil, with records of Xenurolebias species. Xenurolebias tupinikin (red); Xenurolebias izecksohni (white); Xenurolebias cricarensis (rose); Xenurolebias myersi (yellow) and Xenurolebias pataxo (green). Colored contours correspond to each municipality, from south to north: Aracruz (light green); Linhares (purple); São Mateus (dark green); Conceição da Barra (dark blue); Mucuri (light blue); Nova Viçosa (brown); Caravelas (marine blue); Alcobaça (rose) and Prado (blue). Scale bar 60 Km.
Figure 7.
Study area location within the coastal tablelands between Bahia and Espírito Santo, Brazil, with records of Xenurolebias species. Xenurolebias tupinikin (red); Xenurolebias izecksohni (white); Xenurolebias cricarensis (rose); Xenurolebias myersi (yellow) and Xenurolebias pataxo (green). Colored contours correspond to each municipality, from south to north: Aracruz (light green); Linhares (purple); São Mateus (dark green); Conceição da Barra (dark blue); Mucuri (light blue); Nova Viçosa (brown); Caravelas (marine blue); Alcobaça (rose) and Prado (blue). Scale bar 60 Km.
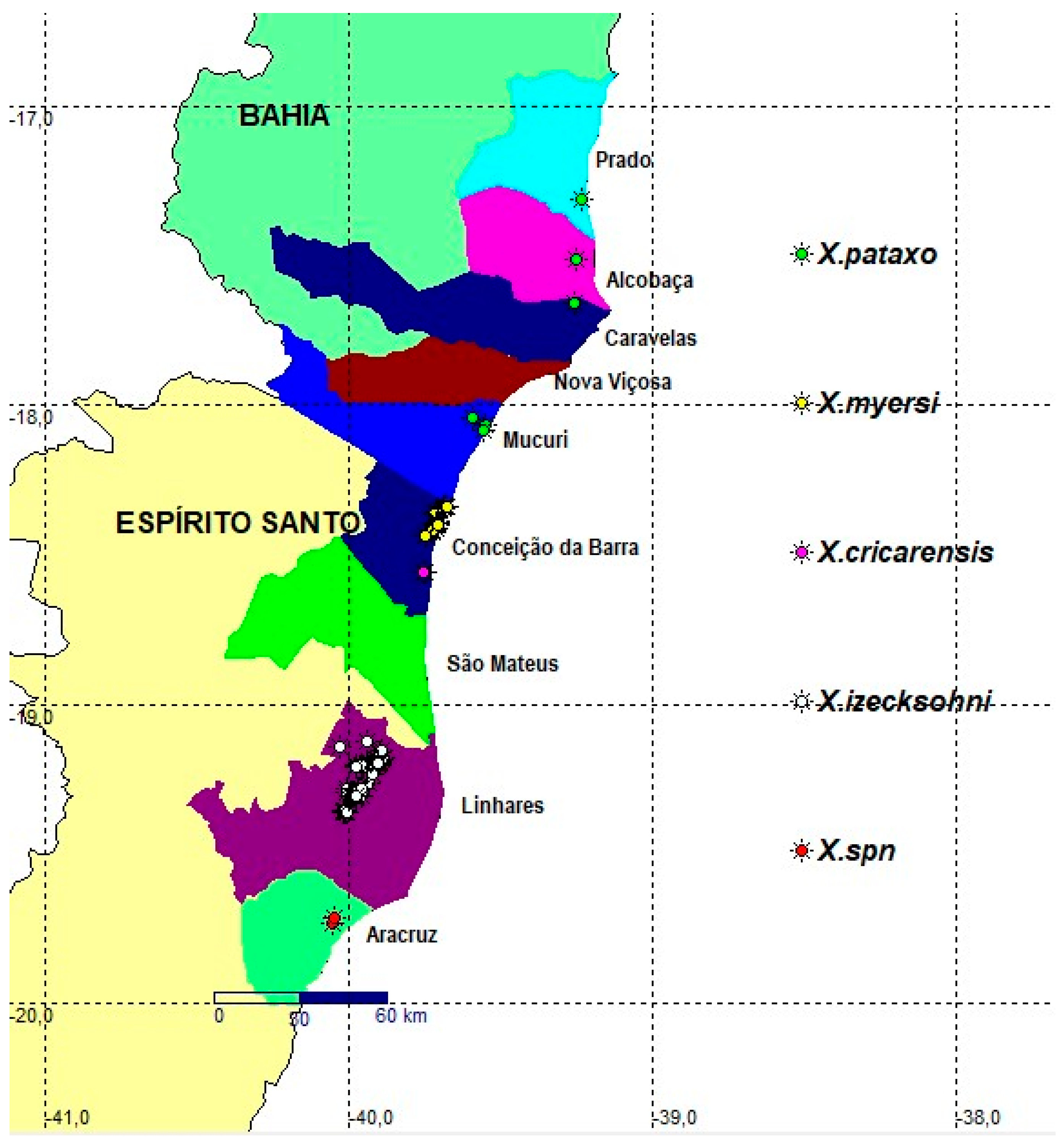
Figure 8.
Xenurolebias izecksohni, A. MBML 14205, male, 54.2 mm SL; B. MBML 14208, female, 29.6 mm SL. Isolated Pool at flooded area on Barra Seca, Linhares, Espírito Santo, Brazil. Images: Bruno Pinheiro.
Figure 8.
Xenurolebias izecksohni, A. MBML 14205, male, 54.2 mm SL; B. MBML 14208, female, 29.6 mm SL. Isolated Pool at flooded area on Barra Seca, Linhares, Espírito Santo, Brazil. Images: Bruno Pinheiro.

Figure 9.
Representation of the local riparian (right).

Figure 10.
Estimates of EOO, cartographic area and the number of locations for Xenurolebias izecksohni.
Figure 10.
Estimates of EOO, cartographic area and the number of locations for Xenurolebias izecksohni.
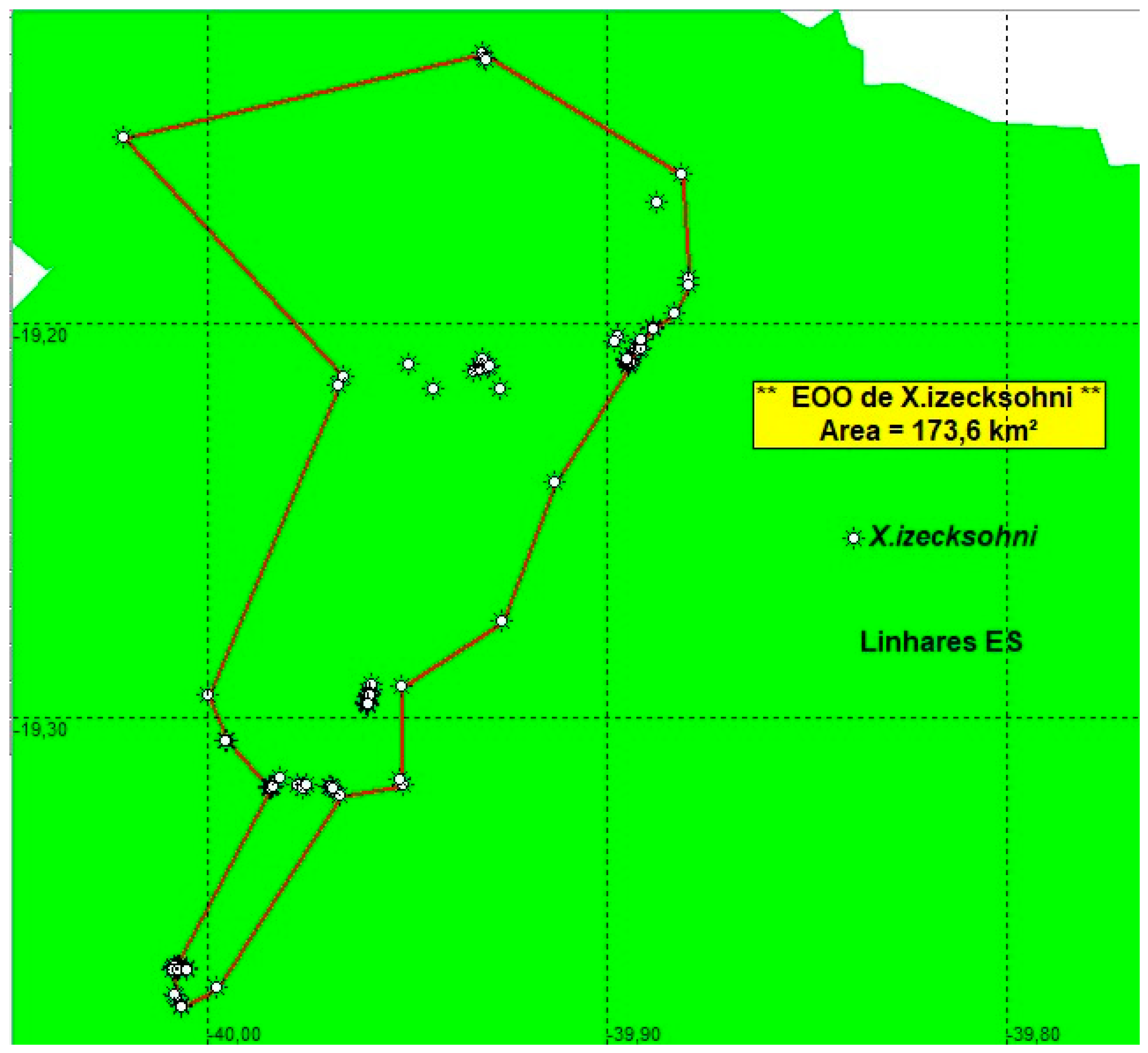
Figure 11.
Couple of Xenurolebias cricarensis in field aquarium. Female in front, male behind. Image: Bruno Pinheiro.
Figure 11.
Couple of Xenurolebias cricarensis in field aquarium. Female in front, male behind. Image: Bruno Pinheiro.

Figure 12.
Flooded area in the sub-basin of the São Domingos Stream at Km 16 of the ES-421 Adolfo Barbosa Serra Highway on the left in the direction of Conceição da Barra (Type Locality of Xenurolebias cricarensis). Image: Ronaldo Pinheiro.
Figure 12.
Flooded area in the sub-basin of the São Domingos Stream at Km 16 of the ES-421 Adolfo Barbosa Serra Highway on the left in the direction of Conceição da Barra (Type Locality of Xenurolebias cricarensis). Image: Ronaldo Pinheiro.

Figure 13.
Estimates of EOO, cartographic area and the number of locations for Xenurolebias cricarensis.
Figure 13.
Estimates of EOO, cartographic area and the number of locations for Xenurolebias cricarensis.
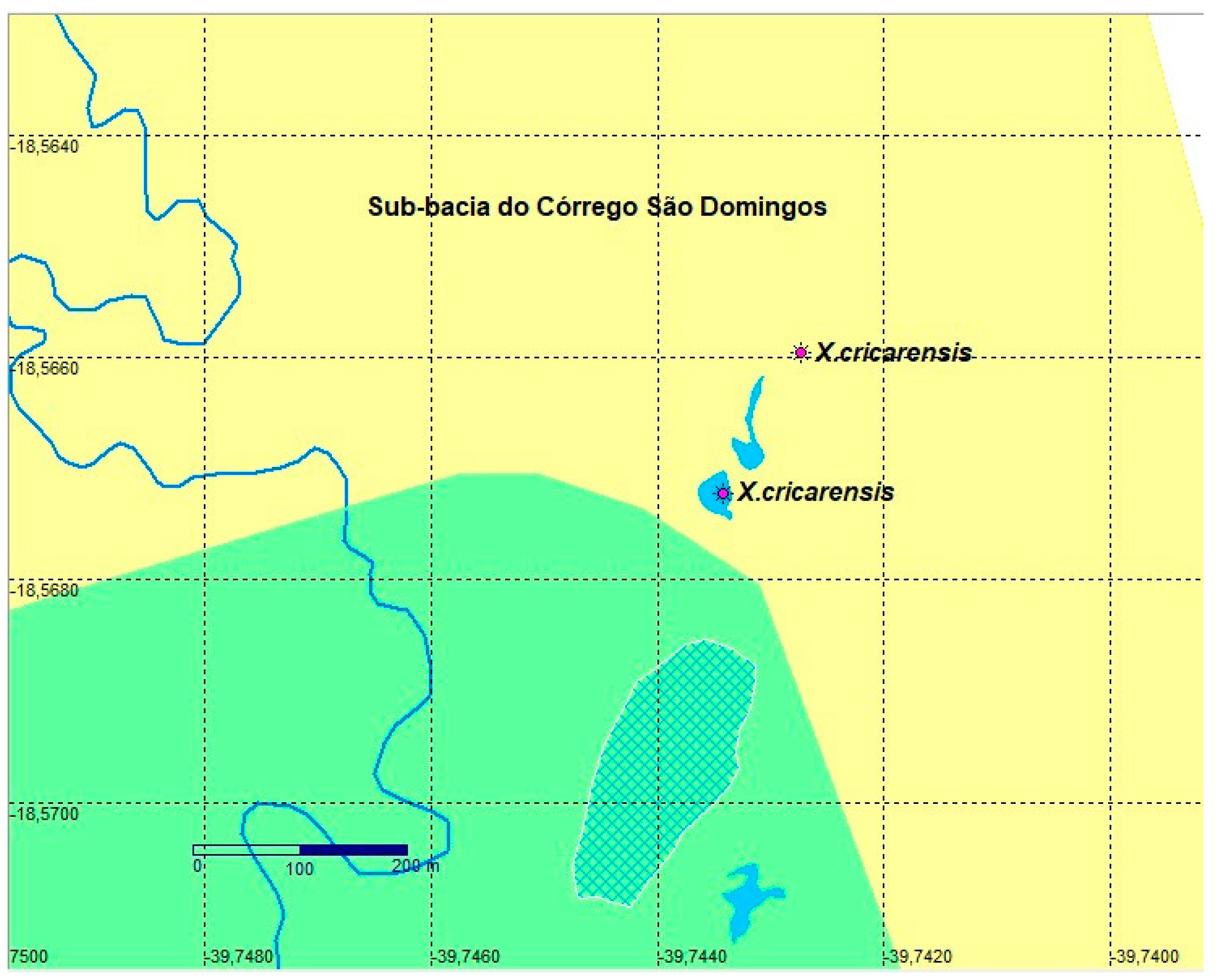
Figure 14.
Xenurolebias myersi, adult male with lanceolate dorsal and anal fins and contrasting vertical bars on flanks. Image: Bruno Pinheiro. Xenurolebias myersi, adult female with translucent fins and dark dots on flanks. Image: Frederico Pereira. Images in field aquarium. Specimens not preserved.
Figure 14.
Xenurolebias myersi, adult male with lanceolate dorsal and anal fins and contrasting vertical bars on flanks. Image: Bruno Pinheiro. Xenurolebias myersi, adult female with translucent fins and dark dots on flanks. Image: Frederico Pereira. Images in field aquarium. Specimens not preserved.

Figure 15.
Velha Antônia stream, near village of Itaúnas. Type locality of Xenurolebias myersi. Image: Luisa Sarmento.
Figure 15.
Velha Antônia stream, near village of Itaúnas. Type locality of Xenurolebias myersi. Image: Luisa Sarmento.

Figure 16.
Estimates of EOO, cartographic area and the number of locations for Xenurolebias myersi.
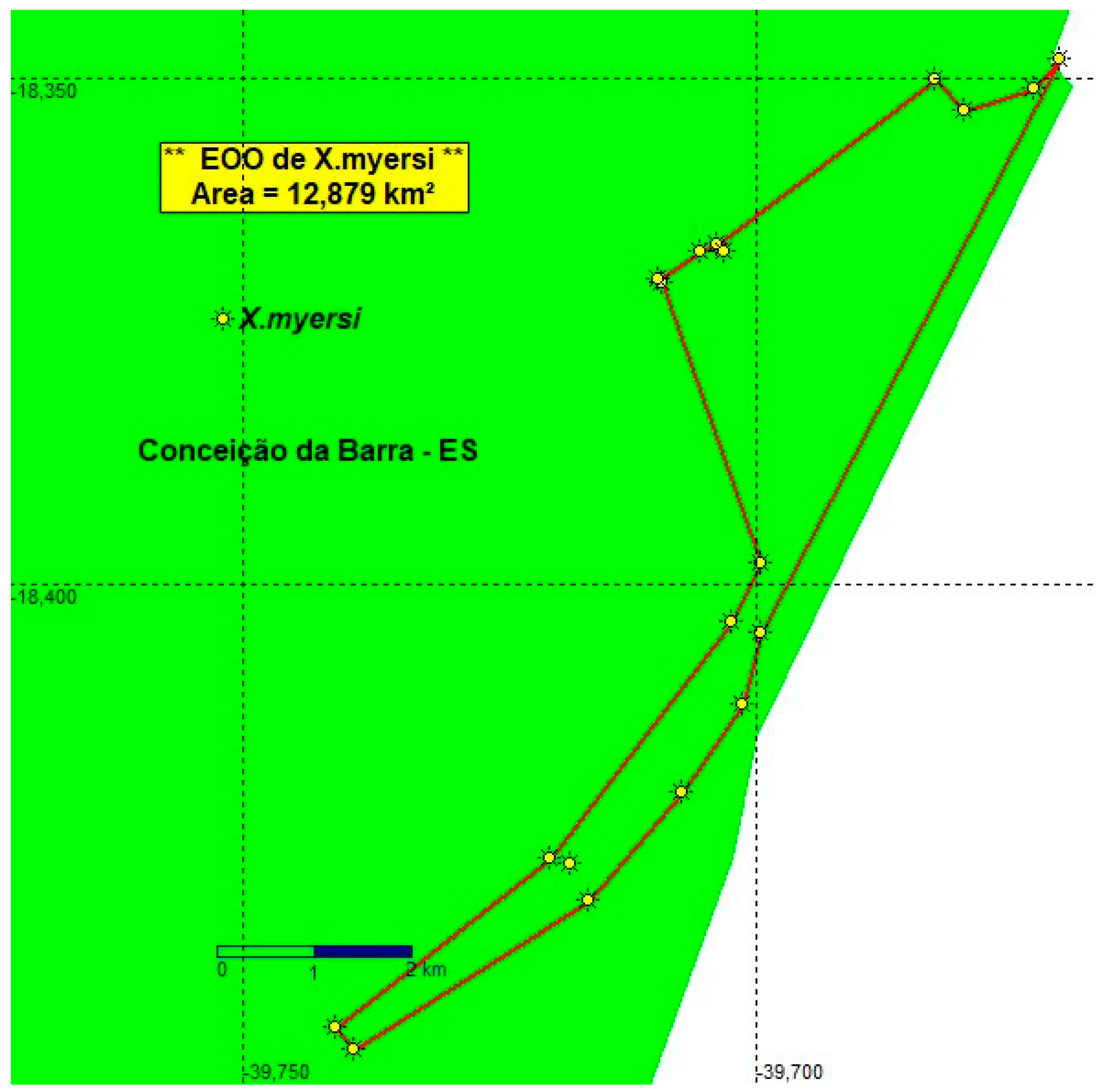
Figure 13.
Xenurolebias pataxo. Image: Luisa Sarmento. Images in field aquarium. Specimen not preserved.
Figure 13.
Xenurolebias pataxo. Image: Luisa Sarmento. Images in field aquarium. Specimen not preserved.

Figure 14.
Flooded area adjacent to Itanhém river, Alcobaça municipality, Bahia. Image: Luisa Sarmento.
Figure 14.
Flooded area adjacent to Itanhém river, Alcobaça municipality, Bahia. Image: Luisa Sarmento.

Figure 19.
Estimates of EOO, cartographic area and the number of locations for Xenurolebias pataxo.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



