
Submitted:
01 July 2025
Posted:
02 July 2025
You are already at the latest version
Abstract
The development of a vaccine against highly pathogenic avian influenza A/H5 viruses is an urgent task due to concerns about its pandemic potential. Here, we have developed an experimental mRNA vaccine mRNA-H5, encoding a modified hemagglutinin trimer of influenza virus A/turkey/Stavropol/320-01/2020 (H5N8). Immunization of BALB/c mice with the obtained mRNA-H5 vaccine using lipid nanoparticles (LNPs) and needle-free jet injection (JI) has been shown to effectively stimulate specific humoral and T-cell immune responses. Moreover, mRNA-H5 delivered by LNPs and JI provided 100% protectivity of immunized mice against lethal challenge with homologous and heterologous strains of avian influenza virus (A/Astrakhan/3212/2020 (H5N8) and A/chicken/Magadan/14-7V/2022 (H5N1), respectively). The present results indicate that needle-free jet injection can be considered as an alternative to LNPs for mRNA delivery. mRNA-H5 is a promising candidate as a vaccine against highly pathogenic avian influenza A/H5 viruses with pandemic potential.
Keywords:
mRNA vaccines
; lipid nanoparticles
; LNPs
; needle-free jet injection
; mRNA delivery
; hemagglutinin
; highly pathogenic avian influenza virus
; immune response
; H5Nx
1. Introduction
As of 5 January 2024, highly pathogenic avian influenza (HPAI) A/H5 viruses of clade 2.3.4.4b have caused the culling of more than 541 million birds to prevent the spread of the disease, causing serious economic damage to the poultry industry [1]. But this virus subtypes pose a threat not only to birds but also to human health [2,3,4]. According to the World Health Organization (WHO), as of 20 June 2025, 976 cases of A/H5N1 infection in humans have been reported, of which 470 resulted in death [5]. HPAI A/H5 viruses have also been reported to infect a variety of wild and domestic mammals, including cattle, badgers, black bears, bobcats, coyotes, ferrets, cats, foxes, leopards, opossums, pigs, raccoons, skunks, sea lions, wild otters [6]. Further circulation of HPAI A/H5 viruses among birds and mammals poses a threat of emergence of new HPAI virus capable of human-to-human transmission and raises concerns about a possible pandemic caused by these influenza virus subtypes [7]. In response to these concerns, WHO calls for increased efforts to develop a vaccine against HPAI viruses.
mRNA vaccines [8,9,10] have several properties that distinguish it from other types of influenza vaccines [11,12,13]. The mRNA vaccine platform has rapid, cost-effective and scalable production in which the target immunogen can be rapidly replaced; it is particularly relevant due to the high variability of the influenza virus. mRNA vaccines do not induce an undesirable antivector immune response and, therefore, can be administered repeatedly. An important advantage is also the effective activation of both humoral and cellular virus-specific immune responses [14,15].
Lipid nanoparticles (LNPs) effectively protect exogenous mRNA from degradation by extracellular nucleases and are currently the most widely used system for delivering mRNA into cells [16,17,18,19,20]. For researchers around the world, the standard of LNPs production is the composition developed by Moderna and Pfizer for COVID-19 vaccines [21,22]. The LNPs in these vaccines consist of four lipid components that ensure efficient encapsulation and release of mRNA, a long period of circulation in the body, and stability of the complex [23]. An additional advantage of LNPs is their adjuvant effect that enhances the immune response to the vaccine [24]. However, a problem of LNPs is the lack of their tissue specificity; untargeted delivery to liver or heart may cause undesirable adverse reactions [25,26].
Alternative delivery methods have recently attracted increasing attention. Thus, recently published in the works of Kisakov et al. [27], Abassy et al. [28] and Wang et al. [29] needle-free jet injection (JI) method is a promising alternative to LNPs for mRNA delivery. The method is based on the intradermal injection under high pressure, while the physical effects on the cell membrane provides increasing permeability to nucleic acids [30]. Additionally, the wide dispersion of the drug in the dermis facilitates the uptake of mRNA by antigen-presenting cells [31,32], which further increases its immunogenicity.
Hemagglutinin (HA) is the primary antigen for the development of influenza vaccines. Immunization with HA-based vaccines induces the production of neutralizing antibodies, which are crucial for defending against viral infection [33,34,35,36]. We have previously carried out the design of a modified HA trimer of influenza virus A/turkey/Stavropol/320-01/2020 (H5N8). The immunogen design is described in more detail in [37]. In our previous studies we obtained recombinant HA/H5 protein and pVAX-H5 DNA vaccine encoding the designed immunogen. We showed that immunization of BALB/c mice with recombinant HA/H5 protein induced the formation of neutralizing antibodies with protective properties [38]. The pVAX-H5 DNA vaccine effectively induced specific humoral and cellular immune responses in laboratory animals [37,39] and protected BALB/c mice from lethal challenge with influenza virus strains A/chicken/Khabarovsk/24-1V/2022 (H5N1) and A/turkey/Stavropol/320-01/2020 (H5N8) [40]. It is important to note that we used the JI as a delivery system for pVAX-H5 DNA vaccine.
The aim of this study was to develop mRNA vaccine encoding a modified HA trimer of influenza A/turkey/Stavropol/320-01/2020 (H5N8) virus and to analyze its immunogenic and protective properties when delivered by JI in comparison with LNPs.
2. Materials and Methods
2.1. Bacterial Strains, Viruses, Cell Cultures, Plasmids
The E. coli strain Stbl3 (Invitrogen, USA) was used to produce plasmid DNA.
In the microneutralization assay influenza virus strain A/turkey/Stavropol/320-01/2020 (H5N8) (EPI_ISL_1114749) (FBRI SRC VB «Vector» Rospotrebnadzor) and cell culture MDCK-SIAT1 (Cell culture collection of the FBRI SRC VB «Vector» Rospotrebnadzor, Russia) were used.
The vaccine strain of influenza virus A/Astrakhan/3212/2020 (H5N8) [41] and strain A/chicken/Magadan/14-7V/2022 (H5N1) (EPI_ISL_16618968) (FBRI SRC VB «Vector» Rospotrebnadzor, Russia) [42] were used for challenge study.
To obtain the DNA template, the recombinant plasmid pVAX-Cas1CC (FBRI SRC VB «Vector», Rospotrebnadzor) was used.
2.2. Preparation of DNA Template pVAX-Cas1CC-H5
The nucleotide sequence encoding the influenza virus HA was designed based on the native HA gene of the influenza virus A/turkey/Stavropol/320-01/2020 (H5N8) (EPI_ISL_1114749). The HA gene was cloned into the pVAX-Cas1CC expression cassette carrying the target gene under the control of the T7 promoter modified for efficient incorporation of the AG-Cap analog during synthesis. The cassette also contains the 5’- and 3’-untranslated regions of human α-globin, as well as a 100-nucleotide poly(A) tail. Cloning was carried out using the recognition sites of restriction endonucleases Psp124BI and BamHI (SibEnzyme, Russia). The construct was named pVAX-Cas1CC-H5. The structure of the obtained plasmid was confirmed by restriction analysis and Sanger sequencing.
The DNA template was produced and purified using the “Hipure Plasmid Mini Kit” (Magen, China) and linearized using the restriction endonuclease EcoRI (SibEnzyme, Russia).
2.3. In Vitro mRNA Synthesis
mRNA synthesis was performed using the linearized pVAX-Cas1CC-H5 template and a commercial kit (Yeasen, China). The reaction mixture included 1 μg of linearized DNA template, T7 polymerase with buffer, AG-Cap analog (m7GmAmG cap analog (Biolabmix, Russia) (6 mM), a mixture of ribonucleotide triphosphates (7 mM) with uridine replaced by N1-methylpseudouridine (Jena Bioscience, Germany), RNase inhibitor (BelBioLab, Russia) and nuclease-free water. The synthesis protocol was described previously [27]. The resulting mRNA was named mRNA-H5.
Analysis of the homogeneity and integrity of the obtained product, as well as its molecular size, was carried out by electrophoresis in 2% agarose gel.
2.4. Lipid Nanoparticles Synthesis
LNPs were synthesized by rapid microfluidic mixing of an aqueous phase containing RNA and an ethanol (organic) phase containing a lipid mixture. The aqueous phase was a solution of mRNA-H5 in 100 mM citrate buffer, pH 4. The ethanol phase consisted of a mixture of ionizable lipid:phospholipid:helper lipid:PEG lipid in a molar percentage ratio of 50:10:38.5:1.5, respectively; all lipids were dissolved in 96% ethanol. The lipids: ionizable lipid SM-102 (heptadecan-9-yl 8-((2-hydroxyethyl) (6-oxo-6-(undecyloxy)hexyl)amino)octanoate); phospholipid DSPC (1,2-distearoyl-sn-glycero-3-phosphocholine); auxiliary lipid cholesterol; PEG lipid DMG-PEG2000 (1-monomethoxypolyethyleneglycol-2,3-dimyristylglycerol with polyethyleneglycol with an average molecular weight of 2000); lipids were purchased from AVT Pharmaceutical Tech Co., China. The ratio of nitrogen in SM-102 to phosphate of mRNA was 6:1. The phases were mixed using the Automated NP System (Dolomite Microfluidics, UK). Mixing was carried out in a staggered herringbone Micromixer microfluidic chip (Dolomite Microfluidics, UK). The procedure protocol: total mixture volume - 1.6 ml; aqueous phase to organic phase ratio - 3:1; flow rate - 3000 μl/min. A chip for diluting the LNPs mixture with phosphate-buffered saline (PBS), pH 7, at a flow rate of 1000 µl/min was connected in series with the mixing chip.
Immediately after production, LNPs were diluted 20-fold with PBS, pH 7, and then concentrated using a JetSpin 30k MWCO centrifugal concentrator (Jet Biofil, China). The total volume of mRNA-H5-LNP obtained was then adjusted to obtain a final concentration corresponding to a dose of 10 or 30 μg of encapsulated mRNA-H5 in 100 μl of PBS. Before each immunization, fresh series of mRNA-H5-LNP were prepared and stored at 4°C for no more than 3 days.
For use as a control, LNPs without mRNA were obtained in a similar way, with citrate buffer as an aqueous phase.
2.5. Determination of Hydrodynamic Size and ζ-Potential of LNPs
The hydrodynamic size and charge of LNPs were determined by dynamic and electrophoretic light scattering and measured using the intensity-averaged particle size (Z-average), polydispersity index (PDI) and ζ-potential using a Zetasizer NanoZSPlus (Malvern Instruments, UK). ZEN0040 cuvettes were used for size measurements and DTS1070 cuvettes were used for ζ-potential measurements of the nanoparticle layer. All measurements were performed in triplicate at 25°C.
2.6. The Quantification of mRNA-H5-LNP Encapsulation
The encapsulation ratio and mRNA concentration were determined using the Quant-iT RiboGreen RNA reagent kit (Life Technologies, USA) according to the manufacturer's protocol, except that mRNA-H5 was the standard RNA to generate the calibration curve. Briefly, LNPs were degraded with 2% Triton X-100 in TE buffer in a 96-well plate. 1× Quant-iT RiboGreen RNA dye in TE buffer was added to the sample, and the fluorescence data of the sample (λex = 485 nm, λem = 520 nm) was measured using a Varioskan™ LUX plate reader (Thermo Fisher Scientific, USA). The total amount of nucleic acids in the sample was determined by applying the relative fluorescent units to the standard curve of nucleic acids with detergent. Simultaneously, to detect unencapsulated nucleic acid intact LNPs were measured under the same conditions but without detergent. The amount of RNA encapsulated in LNPs was determined by subtracting the amount of unencapsulated RNA from the total amount of RNA in the sample. Finally, the amount of encapsulated RNA was divided by the total amount of RNA in the sample to obtain the encapsulation ratio.
2.7. Electron Microscopy of LNPs Formulations
To determine the size and shape of mRNA-H5-LNP and LNPs without mRNA complexes, the suspension was applied to copper grids for electron microscopy coated with a carbon-stabilized film. The formulations were stained with 2% aqueous uranyl acetate. The analysis was carried out using a JEM-1400 electron microscope (Jeol, Japan). Image collection, analysis and processing were performed using a Veleta digital camera (EMSIS GmbH, Germany) and iTEM software (EMSIS GmbH, Germany).
2.8. Immunization of BALB/c Mice with mRNA-H5
The work with animals was carried out in accordance with the "Guide for the Care and Use of Laboratory Animals". The protocols were approved by the Institutional Animal Care and Use Committee at the FBRI SRC VB «Vector» Rospotrebnadzor (Bioethics Committee Protocol No. 3 dated 02/29/2024).
For immunization, female BALB/c mice weighing 16–18 g were used. The mRNA-H5-LNP formulations in 100 μl PBS were injected into the quadriceps muscle of the left hind paw using insulin syringes with a 29G needle. The mRNA-H5 preparations dissolved in 50 μl PBS were injected into the left hind paw using the needle-free jet injection method as described previously [27].
The experiment was divided into 2 stages. In the first stage, to evaluate humoral and T-cell responses, the animals were divided into five groups of 12 animals each: Group 1 was immunized with 30 μg of mRNA-H5-LNP; Group 2 was immunized with 30 μg of mRNA-H5 using JI; Group 3 was immunized with 25 μg of recombinant HA/H5 protein with aluminium hydroxide; Group 4 was immunized with LNPs without mRNA (the amount of LNPs was equivalent to the Group 1); Group 5 consisted of non-immunized animals. Immunization was performed twice with an interval of 21 days. Fourteen days after the second immunization, blood was collected from the retroorbital sinus of the eye to analyze the humoral immune response. Animals were humanely euthanized by cervical dislocation, followed by spleen collection for T-cell response analysis.
At the second stage, the animals were immunized to analyze the protective effect of the mRNA vaccine. The animals were divided into 4 groups of 20 animals each: Group 1 was immunized with 10 μg of mRNA-H5-LNP; Group 2 was immunized with 30 μg of mRNA-H5 using JI; Group 3 was immunized with 25 μg of recombinant HA/H5 protein with aluminium hydroxide; Group 4 consisted of non-immunized animals. Immunization was performed twice with an interval of 21 days. Fourteen days after the second immunization, blood was collected from the retroorbital sinus of the eye to analyze the humoral immune response.
2.9. Enzyme-Linked Immunosorbent Assay (ELISA)
ELISA was performed according to the method described in [37,43]. Recombinant HA/H5 protein of the influenza virus A/turkey/Stavropol/320-01/2020 (H5N8) was used as an antigen [38]. Sera of immune animals were used as primary antibodies (diluted 1:10). Goat antibodies against mouse IgG labeled with horseradish peroxidase (Sigma-Aldrich, USA) were used as secondary antibodies. Tetramethylbenzidine solution (IMTEK, Russia) was used as a chromogenic substrate. The reaction was stopped with 1 N hydrochloric acid solution. Optical density was measured on a Varioskan LUX device (Thermo Fisher Scientific, USA) at a wavelength of 450 nm. The final titer was determined by the last diluted sample that gave positive results for ELISA.
2.10. In Vitro Microneutralization Assay
The microneutralization assay was carried out according to the method described in [38]. First, animal sera were treated with RDE (Denka Seiken, Japan), a receptor-destroying enzyme, according to the protocol recommended by WHO [44]. Each sample was pre-diluted at a ratio of 1:10. The standard virus dose was 100 TCID50/100 μl of viral diluent. Two-fold dilutions of blood serum were prepared in 200 μl of viral diluent, then 200 μl of standardized viruses were added to each tube and incubated for 1 hour at 37°C and 5% CO2. Sera of non-immune mice were used as a negative control. Reference ferret sera (FBRI SRC VB «Vector» Rospotrebnadzor, Russia) were used as a positive control. After that, 200 μl of the suspension was transferred to the wells of a 96-well plate with a 90% monolayer of MDCK-SIAT1 cell culture; the plates were incubated for 3 days in Opti-MEMI medium with 1 μg/ml TPCK-trypsin (Sigma-Aldrich, USA) at 37°C, 5% CO2. After incubation, the cells were stained with crystal violet solution and analyzed using an Agilent BioTek Cytation 5 multi-mode cell imaging device (Thermo Fisher Scientific, USA). The serum titer in the microneutralization assay is equal to the serum dilution at which 50% of living cells remain, while in the control group no more than 5% of living cells remain.
2.11. Assessment of T-Cell Response by Intracellular Cytokine Staining (ICS)
Animal spleens were obtained two weeks after the second immunization. Splenocytes were isolated by sequential homogenization through 70 and 40 μm filters (BD Falcon, New York, NY, USA). After lysis of erythrocytes with lysis buffer (Sigma-Aldrich, USA), splenocytes were washed twice in RPMI medium and placed in 1 mL of RPMI medium with 2 mM L-glutamine and gentamicin (50 μg/mL).
Splenocytes were seeded in 96-well round-bottomed plates (Jet Biofil, China) at 7×105 cells per well; a peptides pool (20 μg/ml for each peptide), medium (as a negative control) or a mixture of mitogens (phorbol 12-myristate 13-acetate (PMA) (30 ng/ml) and ionomycin (1 μg/ml)) (as a positive control) were added.
Peptides from the hemagglutinin of the influenza A/H5N8 virus used to stimulate splenocytes: MPFHNIHPL; AGWLLGNPM; CYPGSLNDY; RVPEWSYIV; LRNSPLREKRRKRGL; YVKSNKLVL; TYNAELLVL; LYDKVRLQL; SFFRNVVWL; SPYQGAPSF; LYKNPTTYISVGTSTLNQ; VDTIMEKNVTVTHAQDILEK; SSWPNHETSLGVSAASPYQ. The peptides have a high probability of binding to MHC H-2D of BALB/c mice (determined using the NetMHCpan-4.1 service).
Cells were incubated for 3 h at 37°C in 5% CO2 and for an additional 15 h with brefeldin A (5 μg/ml, GolgiPlug BD Biosciences, USA). The next day, cells were washed and stained with anti-CD3 Alexa Fluor 700 (clone 500A2, Biolegend, USA), anti-CD4 BV785 (clone GK1.5, Biolegend, USA) and anti-CD8 FITC (clone 53-6.7, Biolegend, USA). Cells were fixed with 1% paraformaldehyde for 10 min, washed and permeabilized with 0.2% Tween-20 for 10 min. Afterwards, cytokine staining was performed with anti-IFNγ APC (clone XMG1.2, Biolegend, USA), anti-IL-2 BV 421 (clone JES6-5H4, Biolegend, USA), anti-TNFα PE (clone MP6-XT22, Biolegend, USA). After 20 min, the cells were washed, dissolved in 1% paraformaldehyde solution and incubated for 20 min, followed by analysis on a ZE5 flow cytometer (Bio-Rad, USA) using the Everest program.
2.12. Virulence
To determine the virulence of avian influenza viruses, female BALB/c mice weighing 18-20 g were infected intranasally. The mice were divided into six groups of six animals each to determine the 50% lethal dose (MLD50). The animals were lightly anesthetized with a combination of Zoletil 100 (Delpharm Tours, France) and Xyla (Interchemie, Estonia) before intranasal inoculation with 0.05 ml of the virus diluted 10-fold. The six groups were observed for 14 days to detect clinical signs and mortality. The 50% lethal dose in mice was calculated using the Reed-Munch method.
2.13. Challenge Study
All challenge studies were carried out in compliance with the requirements of SanPiN 3.3686-21 “Sanitary and epidemiological requirements for the prevention of infectious diseases” [45].
Fourteen days after the second immunization, mice were infected intranasally with 20 MLD50 of the influenza virus strain A/Astrakhan/3212/2020 (H5N8) or 20 MLD50 of the influenza virus strain A/chicken/Magadan/14-7V/2022 (H5N1). Mice were inoculated under anesthesia with a mixture of tiletamine and zolazepam, as well as xylazine chloride. The animals were observed daily after infection for 14 days; clinical signs indicating the development of the disease (ruffled fur, hypothermia, exhaustion, neurological disorders and death) were monitored and the weight of the animals was measured. In cases where mice developed severe conditions incompatible with life, such as anorexia (loss of more than 20% of the initial body weight), lethargy, the animal was humanely euthanized by cervical dislocation. All other animals were humanely euthanized by cervical dislocation at the end of the experiment.
2.14. Statistical Analysis
Statistical data processing was performed using GraphPad Prism 9.0 software (GraphPad Software, USA). Quantitative data are presented as mean with a range of values or a range from lowest to highest and analyzed using nonparametric tests. Intergroup differences were assessed using nonparametric one-way Kruskal-Wallis analysis of variance with correction for multiple comparisons and Dunn's statistical hypothesis testing. Survival function modeling was performed using the Kaplan-Meier multiplicative estimator, and survival comparison with the control group was performed using the Mantel-Cox test.
3. Results
3.1. Preparation of DNA Template and mRNA Synthesis
The gene encoding the HA of HPAI virus subtype A/H5N8, which was circulating in Russia in Stavropol in 2020 (A/turkey/Stavropol/320-01/2020) among farm poultry, was selected as an immunogen for the experimental mRNA vaccine. The amino acid sequence of the influenza virus strain A/turkey/Stavropol/320-01/2020 HA has been shown to align with the HA of the strain A/Astrakhan/3212/2020 (H5N8), which has been recommended by WHO as a vaccine strain [46]. The native sequence of the gene was modified to stabilize its conformation, and the fibritin trimerization domain of bacteriophage T4 was added to the C-terminus of the amino acid sequence [38].
The HA gene was designed and cloned into the pVAX-Cas1CC expression cassette, resulting in the DNA template pVAX-Cas1CC-H5. Synthesis of the target mRNA (Figure 1a) was then carried out from the linearized DNA template; the synthesis products were verified by electrophoresis (Figure 1b). The size of the resulting mRNA was shown to be the same as the theoretical calculation (1900 nucleotides), and the purity of the synthesis product was demonstrated.
3.2. Lipid Nanoparticles Synthesis and Characterization
The mRNA-H5 obtained was then encapsulated in LNPs for the subsequent immunization of mice. For use as a control, LNPs without mRNA were obtained in a similar way.
The obtained formulations were characterized using dynamic light scattering and electrophoretic light scattering methods. The dynamic light scattering method (Table 1) was used to analyze the samples, revealing that both the mRNA-H5-LNP complex and the LNPs without mRNA resulted in a monodisperse suspension of nanoparticles (PdI = 0.149 ± 0.01 and 0.175 ± 0.01, respectively). The mean hydrodynamic diameter of mRNA-H5-LNP was 93.5 ± 0.8 nm, while for LNPs without mRNA this parameter was higher at 131.6 ± 1.6 nm. This discrepancy may be explained by the spontaneous random self-assembly of nanoparticles in the absence of mRNA, which can result in an increase in mean size [47].
The particle distribution profile (Figure 2b) demonstrates the presence of a single peak, indicating the absence of fractions with other particle sizes and the absence of impurities.
The ζ-potential value of the mRNA-H5-LNP particles in three series of measurements was -0.02 ± 0.26 mV, in line with theoretical calculations. The almost zero charge indicates that the negatively charged mRNA molecules were almost completely packed into the LNPs. This was confirmed by Quant-iT RiboGreen RNA analysis, which also showed that an encapsulation efficiency of more than 90% was achieved. Electron microscopy (Figure 2a) confirmed that nanoparticles of the required size had formed.
3.3. A Comparison of the Immunogenicity of mRNA-H5 Administered to BALB/c Mice Using LNPs and JI
BALB/c mice were immunized to analyze the immunogenicity of mRNA-H5 and compare the efficacy of the aforementioned delivery methods.
After double immunization, the animal sera were analyzed using ELISA (Figure 3a). Specific antibody titers were detected in the experimental groups of animals which were immunized with mRNA-H5 delivered by LNPs (average titer was 1:546800), and by JI (average titer was 1:164000). Specific antibody titers were also detected in the control group of animals, which were immunized with recombinant HA/H5 protein with aluminium hydroxide (average titer was 1:182300).
Sera from immunized mice were also tested for microneutralization in MDCK cell culture. As a result, it was found that the sera of animals immunized with mRNA-H5 delivered by LNPs, mRNA-H5 delivered by JI, and recombinant HA/H5 protein are capable of neutralizing live influenza virus strain A/turkey/Stavropol/320-01/2020 (H5N8); the average 50% neutralization titer was 1:5700, 1:1700 and 1:3000 respectively.
To assess the ability of the vaccine to induce specific cellular immunity, spleens were explanted from immunized mice and examined by ICS. The response was assessed by the ability of splenocytes to respond by releasing cytokines to stimulation with peptides included in hemagglutinin.
The analysis demonstrated that immunization with mRNA-H5 resulted in the generation of both CD3+/CD8+ and CD3+/CD4+ lymphocytes, which were capable of producing IFNγ, IL-2 and TNFα after stimulation with HA-specific peptides (Figure 4).
3.4. Study of the Ability of mRNA-H5 Delivered by LNPs and JI to Protect Mice from Lethal Challenge with Avian Influenza A/H5 Viruses
After the B- and T-cell response induced by the mRNA-H5 had been analyzed, the next step was to evaluate its protective potential. The dose of mRNA-H5-LNP was reduced in accordance with the principles of humanity.
Mice were immunized twice at doses of 10 and 30 µg for LNPs and JI, respectively. The animals immunized with the recombinant HA/H5 protein and the intact animals were used as controls. The resulting titers (Figure 5a) estimated by ELISA were 1:656100 for mRNA-H5-LNP and 1:175000 for mRNA-H5 JI. The recombinant HA/H5 protein was found to stimulate a humoral immune response at a titer of 1:196800.
Then, on the 35th day after the first immunization, the mice in each group were divided into two subgroups of ten animals each. These subgroups were then infected intranasally with the influenza virus strains A/Astrakhan/3212/2020 (H5N8) [41] and A/chicken/Magadan/14-7V/2022 (H5N1) [42]. The results (Figure 5b,c) showed that the survival rate in groups of mice immunized with mRNA-H5-LNP and mRNA-H5 JI when infected with both strains was 100%. There were no signs of disease or weight loss of the animals in the experimental groups. The survival rate in the negative control group (intact animals) was 0%; as early as the fifth day after infection, neurological symptoms (e.g. decreased activity, loss of appetite) and ruffling were observed in animals, as well as significant weight loss (Figure 5d,e). The survival rate in the group of animals immunized with the recombinant HA/H5 protein was 100% in case of infection with a homologous strain, and 90% in case of infection with a heterologous strain.
4. Discussion
As mentioned earlier, mRNA vaccines have several properties that distinguish it from other types of influenza vaccines [11,12,13]. The use of mRNA platform for the development of vaccines against HPAI A/H5 viruses has been described in numerous articles [16,48,49].
In the works mentioned above, LNPs are used as a method of mRNA vaccine delivery. LNPs have demonstrated efficacy in delivering mRNA into the cytosol of target cells while protecting it from degradation within the body [50]. These particles are composed of pH-sensitive lipids; consequently, when the mRNA-LNPs complex undergoes endocytosis, the acidic environment of the late endosome initiates the degradation of the nanoparticle, resulting in the release of mRNA into the cytoplasm. However, it should be noted that certain components of LNPs have the potential to cause adverse effects. For instance, the presence of helper lipids has been observed to induce systemic inflammation through their absorption by Kupffer cells within the liver [51]. Furthermore, the interaction of certain helper lipids with phospholipases A2 has been demonstrated to result in the production of allergens, which can subsequently trigger complement activation and anaphylaxis [52,53]. It must be noted that the aforementioned list does not include all the problems reported with the use of LNPs [25,54,55,56].
An alternate approach to mRNA vaccine delivery involves JI, a method that lacks the disadvantages associated with LNPs [28,57]. In a previous study [27], it was demonstrated that the delivery of naked mRNA-RBD using a spring-loaded jet injector resulted in higher vaccine immunogenicity compared to the conventional intramuscular injection method that utilizes a needle and syringe. It was observed that naked mRNA-RBD delivered by JI has a broad distribution in tissue near the injection site but does not spread systemically, resulting in reduced vaccine reactogenicity and adverse effects.
In this study, we have developed an experimental mRNA vaccine mRNA-H5 encoding a modified hemagglutinin trimer of influenza virus A/turkey/Stavropol/320-01/2020 (H5N8). In the first stage of the study, the double immunization of BALB/c mice revealed that mRNA-H5, delivered by JI as a naked mRNA molecule or in complex with LNPs, could effectively stimulate the humoral immune response by inducing the production of H5-specific antibodies capable of neutralizing influenza virus strain A/turkey/Stavropol/320-01/2020 (H5N8) (Figure 3). The results of the study indicated that the group immunized with mRNA-H5-LNP exhibited a statistically significant increase in the level of humoral immune response in comparison with the group immunized with mRNA-H5 using JI. These data suggest that the encapsulation of mRNA in LNP may provide more efficient mRNA delivery to immune system cells. A factor that could influence the immunogenicity of mRNA delivered by JI is the partial degradation of the naked mRNA molecule by extracellular nucleases [50].
Cellular immunity also plays an important role in the prevention of influenza. The results obtained demonstrated that the percentage of CD4+ and CD8+ T cells that secrete IFN-γ, IL-2, and TNF-α was significantly higher in mice immunized with mRNA-H5 than in negative control groups (Figure 4). T-cells are incapable of providing sterilizing immunity against influenza; nevertheless, they frequently offer broader protection against various strains or subtypes of the virus [58]. This property is a significant component in the induction of comprehensive immune responses. It is also important to note that there were no statistically significant differences in IFNγ, IL-2, and TNFα production between the groups immunized with both JI and LNPs.
While the immune response in mice immunized with mRNA-H5-LNP (30 µg) was higher than in those immunized with mRNA-H5 JI, the condition of the animals in the first group was unsatisfactory. It was observed that the animals exhibited signs of distress, including coat ruffling, as well as heightened discomfort during the second immunization procedure. In consideration of the above, the protocol for evaluating the vaccine's protective efficacy was modified to reduce a dosage of mRNA-H5-LNP, set at 10 µg, in accordance with the principles of humanity. The antibody titer induced by mRNA-H5-LNP was similar at doses of 10 and 30 μg, according to the analysis of the humoral immune response (Figure 5a). A slight difference in antibody titers may be explained by a number of factors. According to the studies of several researchers, the cells have limited capacity to translate exogenous mRNA into protein. It has been determined that it is possible to achieve the maximum antigen production at a dose of 10 µg of mRNA-LNPs complex, and that an increasing of a dose does not result in a higher level of expression [59,60]. Another factor is the buffering capacity of nanoparticles. They have limited ability to effectively deliver drugs into cells due to saturable endocytosis and limited intracellular release associated with nanoparticle fusion [61,62,63]. The detection of this phenomenon indicates the necessity for further exploration of the dose-dependent effect of the mRNA-H5.
A challenge study demonstrated that immunization with both naked mRNA-H5 administered with JI and mRNA-H5 encapsulated in LNPs provided 100% protection of animals against lethal challenge of both strains of avian influenza virus (A/Astrakhan/3212/2020 (H5N8) and influenza A/chicken/Magadan/14-7V/2022 (H5N1)) (Figure 5). The vaccine's ability to provide protection against both homologous and heterologous HPAI viruses may indicate its broad-spectrum protection.
Overall, the obtained mRNA vaccine is a promising candidate for use as a vaccine against HPAI A/H5 viruses. A relevant area of further research is to evaluate the dose-dependent effect of mRNA delivered by both LNPs and JI, and estimate the protective potential of vaccine using other heterologous A/H5 viruses.
5. Conclusions
Consequently, in the current study, an experimental mRNA vaccine encoding a modified HA trimer of the influenza A/turkey/Stavropol/320-01/2020 (H5N8) virus was developed. Immunization of BALB/c mice with the obtained mRNA vaccine using LNPs and JI has been shown to effectively stimulate specific humoral and T-cell immune responses. An analysis of delivery methods demonstrated that mRNA-H5 encapsulated in LNPs resulted in a more significant stimulation of the humoral response compared to mRNA-H5 delivered by JI at the same dose. However, the level of humoral response to mRNA-H5 JI was comparable to the response to recombinant HA/H5 protein with aluminum hydroxide at a dose of 25 μg. Moreover, mRNA-vaccine mRNA-H5 delivered by LNPs and JI provided 100% protectivity of immunized mice against lethal challenge with homologous and heterologous strains of avian influenza virus (A/Astrakhan/3212/2020 (H5N8) and A/chicken/Magadan/14-7V/2022 (H5N1), respectively). The present results indicate that needle-free jet injection can be considered as an alternative to LNPs for mRNA delivery. It is necessary to study the dose-dependent effects of mRNA-H5-LNP and mRNA-H5 JI, as well as confirm the protective properties of the vaccine in other animal models, such as ferrets. Ultimately, mRNA-H5 is a promising candidate as a vaccine against HPAI A/H5 viruses with pandemic potential.
Author Contributions
Conceptualization, A.P.R., A.A.I. and L.I.K.; methodology, V.R.L., V.A.Y., N.B.R., E.V.T., D.N.K., M.B.B., S.V.S., E.V.S., L.A.K., B.N.Z., E.V.D., L.I.K., K.I.I., A.S.G. and V.Y.M.; validation, L.I.K. and A.P.R.; resources, E.V.T., S.V.S. and S.I.K.; investigation, V.R.L., V.A.Y., N.B.R., E.V.T., M.B.B., E.V.S., K.P.M., D.I.V., S.I.K., B.N.Z., E.V.D., K.I.I., A.S.G., O.N.P., N.V.D. and V.Y.M.; supervision, A.P.R., L.I.K. and A.A.I.; visualization, V.A.Y. and D.I.V.; writing—original draft preparation, V.A.Y., V.R.L. and A.P.R.; writing—review and editing, A.P.R., N.B.R., T.N.I., A.S.G., A.A.I. and L.I.K. All authors have read and agreed to the published version of the manuscript.
Funding
The study was conducted under the state assignment of FBRI SRC VB “Vector” Rospotrebnadzor.
Institutional Review Board Statement
The study was conducted according to the Declaration of Helsinki and approved by the Bioethics Committee of SRC VB Vector (No. 3 Protocol of the Bioethics Committee of 29 February 2024).
Informed Consent Statement
Not applicable.
Data Availability Statement
The data can be shared up on request.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| mRNA | Messenger RNA |
| LNPs | Lipid nanoparticles |
| JI | Needle-free jet injection |
| HPAI | Highly pathogenic avian influenza |
| WHO | World Health Organization |
| HA | Hemagglutinin |
| DNA | Deoxyribonucleic acid |
| PDI | Polydispersity index |
| ELISA | Enzyme-linked immunosorbent assay |
| ICS | Intracellular cytokine staining |
| MLD50 | 50% lethal dose |
| PMA | Phorbol 12-myristate 13-acetate |
References
- Hannoun, C. The Evolving History of Influenza Viruses and Influenza Vaccines. Expert Rev Vaccines 2013, 12, 1085–1094. [Google Scholar] [CrossRef]
- Saad, N.; Esaki, M.; Kojima, I.; Khalil, A.M.; Osuga, S.; Shahein, M.A.; Okuya, K.; Ozawa, M.; Alhatlani, B.Y. Phylogenetic Characterization of Novel Reassortant 2.3.4.4b H5N8 Highly Pathogenic Avian Influenza Viruses Isolated from Domestic Ducks in Egypt During the Winter Season 2021–2022. Viruses 2024, 16, 1655. [Google Scholar] [CrossRef]
- AbuBakar, U.; Amrani, L.; Kamarulzaman, F.A.; Karsani, S.A.; Hassandarvish, P.; Khairat, J.E. Avian Influenza Virus Tropism in Humans. Viruses 2023, 15, 833. [Google Scholar] [CrossRef]
- Purcell, R.; Giles, M.L.; Crawford, N.W.; Buttery, J. Systematic Review of Avian Influenza Virus Infection and Outcomes during Pregnancy. Emerg Infect Dis 2025, 31. [Google Scholar] [CrossRef]
- Avian Influenza Weekly Update # 1002: 20 June 2025. Available online: https://www.who.int/westernpacific/publications/m/item/avian-influenza-weekly-update---1002--20-june-2025 (accessed on 25 June 2025).
- Kim, D.-H.; Lee, S.-H.; Kim, J.; Lee, J.; Lee, J.-H.; Jeong, J.; Kim, J.; Choi, Y.-K.; Youk, S.; Song, C.-S. Newcastle Disease Virus Expressing Clade 2.3.4.4b H5 Hemagglutinin Confers Protection against Lethal H5N1 Highly Pathogenic Avian Influenza in BALB/c Mice. Front Vet Sci 2025, 12. [Google Scholar] [CrossRef]
- Ku, C.-C.; Lin, C.-Y.; Yang, C.-R.; Yang, Y.-C.; Chen, P.-L.; Lin, Y.-T.; Wang, P.-R.; Lee, M.-S.; Liang, S.-M.; Hsiao, P.-W. Vaccine Optimization for Highly Pathogenic Avian Influenza: Assessment of Antibody Responses and Protection for Virus-like Particle Vaccines in Chickens. Vaccine X 2024, 20, 100552. [Google Scholar] [CrossRef]
- Xu, S.; Yang, K.; Li, R.; Zhang, L. MRNA Vaccine Era—Mechanisms, Drug Platform and Clinical Prospection. Int J Mol Sci 2020, 21, 6582. [Google Scholar] [CrossRef]
- Sahin, U.; Karikó, K.; Türeci, Ö. MRNA-Based Therapeutics — Developing a New Class of Drugs. Nat Rev Drug Discov 2014, 13, 759–780. [Google Scholar] [CrossRef]
- Zabrodskaya, Y.A.; Gavrilova, N.V.; Elpaeva, E.A.; Lozhkov, A.A.; Vysochinskaya, V.V.; Dobrovolskaya, O.A.; Dovbysh, O.V.; Zimmerman, E.L.; Dav, P.N.; Brodskaia, A.V.; et al. MRNA Encoding Antibodies against Hemagglutinin and Nucleoprotein Prevents Influenza Virus Infection in Vitro. Biochem Biophys Res Commun 2024, 738, 150945. [Google Scholar] [CrossRef]
- Linares-Fernández, S.; Lacroix, C.; Exposito, J.-Y.; Verrier, B. Tailoring MRNA Vaccine to Balance Innate/Adaptive Immune Response. Trends Mol Med 2020, 26, 311–323. [Google Scholar] [CrossRef]
- Pardi, N.; Hogan, M.J.; Porter, F.W.; Weissman, D. MRNA Vaccines — a New Era in Vaccinology. Nat Rev Drug Discov 2018, 17, 261–279. [Google Scholar] [CrossRef]
- Scorza, F.; Pardi, N. New Kids on the Block: RNA-Based Influenza Virus Vaccines. Vaccines (Basel) 2018, 6, 20. [Google Scholar] [CrossRef]
- Cagigi, A.; Douradinha, B. Have MRNA Vaccines Sentenced DNA Vaccines to Death? Expert Rev Vaccines 2023, 22, 1154–1167. [Google Scholar] [CrossRef]
- Beasley, D.W.C. New International Guidance on Quality, Safety and Efficacy of DNA Vaccines. NPJ Vaccines 2020, 5, 53. [Google Scholar] [CrossRef]
- Furey, C.; Scher, G.; Ye, N.; Kercher, L.; DeBeauchamp, J.; Crumpton, J.C.; Jeevan, T.; Patton, C.; Franks, J.; Rubrum, A.; et al. Development of a Nucleoside-Modified MRNA Vaccine against Clade 2.3.4.4b H5 Highly Pathogenic Avian Influenza Virus. Nat Commun 2024, 15, 4350. [Google Scholar] [CrossRef]
- Li, Y.; Wang, X.; Zeng, X.; Ren, W.; Liao, P.; Zhu, B. Protective Efficacy of a Universal Influenza MRNA Vaccine against the Challenge of H1 and H5 Influenza A Viruses in Mice. mLife 2023, 2, 308–316. [Google Scholar] [CrossRef]
- Wang, Z.; Tian, C.; Zhu, J.; Wang, S.; Ao, X.; He, Y.; Chen, H.; Liao, X.; Kong, D.; Zhou, Y.; et al. Avian Influenza MRNA Vaccine Encoding Hemagglutinin Provides Complete Protection against Divergent H5N1 Viruses in Specific-Pathogen-Free Chickens. J Nanobiotechnology 2025, 23, 55. [Google Scholar] [CrossRef]
- Vysochinskaya, V.; Shishlyannikov, S.; Zabrodskaya, Y.; Shmendel, E.; Klotchenko, S.; Dobrovolskaya, O.; Gavrilova, N.; Makarova, D.; Plotnikova, M.; Elpaeva, E.; et al. Influence of Lipid Composition of Cationic Liposomes 2X3-DOPE on MRNA Delivery into Eukaryotic Cells. Pharmaceutics 2022, 15, 8. [Google Scholar] [CrossRef]
- Weng, Y.; Li, C.; Yang, T.; Hu, B.; Zhang, M.; Guo, S.; Xiao, H.; Liang, X.-J.; Huang, Y. The Challenge and Prospect of MRNA Therapeutics Landscape. Biotechnol Adv 2020, 40, 107534. [Google Scholar] [CrossRef]
- Corbett, K.S.; Flynn, B.; Foulds, K.E.; Francica, J.R.; Boyoglu-Barnum, S.; Werner, A.P.; Flach, B.; O’Connell, S.; Bock, K.W.; Minai, M.; et al. Evaluation of the MRNA-1273 Vaccine against SARS-CoV-2 in Nonhuman Primates. New England Journal of Medicine 2020, 383, 1544–1555. [Google Scholar] [CrossRef]
- Polack, F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Pérez Marc, G.; Moreira, E.D.; Zerbini, C.; et al. Safety and Efficacy of the BNT162b2 MRNA Covid-19 Vaccine. New England Journal of Medicine 2020, 383, 2603–2615. [Google Scholar] [CrossRef]
- Cheng, F.; Wang, Y.; Bai, Y.; Liang, Z.; Mao, Q.; Liu, D.; Wu, X.; Xu, M. Research Advances on the Stability of MRNA Vaccines. Viruses 2023, 15, 668. [Google Scholar] [CrossRef]
- Cui, X.; Vervaeke, P.; Gao, Y.; Opsomer, L.; Sun, Q.; Snoeck, J.; Devriendt, B.; Zhong, Z.; Sanders, N.N. Immunogenicity and Biodistribution of Lipid Nanoparticle Formulated Self-Amplifying MRNA Vaccines against H5 Avian Influenza. NPJ Vaccines 2024, 9, 138. [Google Scholar] [CrossRef]
- Lim, J.-H.; Kim, M.-S.; Kim, Y.-J.; Han, M.-H.; Jung, H.-Y.; Choi, J.-Y.; Cho, J.-H.; Kim, C.-D.; Kim, Y.-L.; Park, S.-H. New-Onset Kidney Diseases after COVID-19 Vaccination: A Case Series. Vaccines (Basel) 2022, 10, 302. [Google Scholar] [CrossRef]
- Fang, E.; Liu, X.; Li, M.; Zhang, Z.; Song, L.; Zhu, B.; Wu, X.; Liu, J.; Zhao, D.; Li, Y. Advances in COVID-19 MRNA Vaccine Development. Signal Transduct Target Ther 2022, 7, 94. [Google Scholar] [CrossRef]
- Kisakov, D.N.; Karpenko, L.I.; Kisakova, L.A.; Sharabrin, S.V.; Borgoyakova, M.B.; Starostina, E.V.; Taranov, O.S.; Ivleva, E.K.; Pyankov, O.V.; Zaykovskaya, A.V.; et al. Jet Injection of Naked MRNA Encoding the RBD of the SARS-CoV-2 Spike Protein Induces a High Level of a Specific Immune Response in Mice. Vaccines (Basel) 2025, 13, 65. [Google Scholar] [CrossRef]
- Abbasi, S.; Matsui-Masai, M.; Yasui, F.; Hayashi, A.; Tockary, T.A.; Akinaga, S.; Kohara, M.; Kataoka, K.; Uchida, S. Jet Injection Potentiates Naked MRNA SARS-CoV-2 Vaccine in Mice and Non-Human Primates by Adding Physical Stress to the Skin 2023.
- Wang, C.; Tang, X.; Jiang, C.; Zhang, Y.; Han, B.; Sun, Y.; Guo, J.; Peng, H.; Wang, Z.; Wang, Y.; et al. Intradermal Delivery of SARS-CoV-2 RBD3-Fc MRNA Vaccines via a Needle-Free Injection System Induces Robust Immune Responses in Rats. Front Immunol 2025, 16. [Google Scholar] [CrossRef]
- Miyazaki, H.; Ogura, M.; Sakaguchi, Y.; Hasegawa, T.; Atobe, S.; Terai, K. Mechanism of Jet Injector-Induced Plasmid DNA Uptake: Contribution of Shear Stress and Endocytosis. Int J Pharm 2021, 609, 121200. [Google Scholar] [CrossRef]
- Sonoda, J.; Mizoguchi, I.; Inoue, S.; Watanabe, A.; Sekine, A.; Yamagishi, M.; Miyakawa, S.; Yamaguchi, N.; Horio, E.; Katahira, Y.; et al. A Promising Needle-Free Pyro-Drive Jet Injector for Augmentation of Immunity by Intradermal Injection as a Physical Adjuvant. Int J Mol Sci 2023, 24, 9094. [Google Scholar] [CrossRef]
- Elnekave, M.; Furmanov, K.; Hovav, A.-H. Intradermal Naked Plasmid DNA Immunization: Mechanisms of Action. Expert Rev Vaccines 2011, 10, 1169–1182. [Google Scholar] [CrossRef]
- Knipe, D.; Howley, P. Fields Virology, 6th ed.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2013; pp. 374–414. [Google Scholar]
- Bhatt, S.; Holmes, E.C.; Pybus, O.G. The Genomic Rate of Molecular Adaptation of the Human Influenza A Virus. Mol Biol Evol 2011, 28, 2443–2451. [Google Scholar] [CrossRef]
- Bullard, B.L.; Weaver, E.A. Strategies Targeting Hemagglutinin as a Universal Influenza Vaccine. Vaccines (Basel) 2021, 9, 257. [Google Scholar] [CrossRef]
- Arunachalam, A.B.; Post, P.; Rudin, D. Unique Features of a Recombinant Haemagglutinin Influenza Vaccine That Influence Vaccine Performance. NPJ Vaccines 2021, 6, 144. [Google Scholar] [CrossRef]
- Litvinova, V.R.; Rudometov, A.P.; Rudometova, N.B.; Kisakov, D.N.; Borgoyakova, M.B.; Kisakova, L.A.; Starostina, E.V.; Fando, A.A.; Yakovlev, V.A.; Tigeeva, E.V.; et al. DNA Vaccine Encoding a Modified Hemagglutinin Trimer of Avian Influenza A Virus H5N8 Protects Mice from Viral Challenge. Vaccines (Basel) 2024, 12, 538. [Google Scholar] [CrossRef]
- Rudometova, N.B.; Fando, A.A.; Kisakova, L.A.; Kisakov, D.N.; Borgoyakova, M.B.; Litvinova, V.R.; Yakovlev, V.A.; Tigeeva, E.V.; Vahitov, D.I.; Sharabrin, S.V.; et al. Immunogenic and Protective Properties of Recombinant Hemagglutinin of Influenza A (H5N8) Virus. Vaccines (Basel) 2024, 12, 143. [Google Scholar] [CrossRef]
- Litvinova, V.R.; Rudometova, N.B.; Kisakova, L.A.; Kisakov, D.N.; Yakovlev, V.A.; Tigeeva, E.V.; Borgoyakova, M.B.; Ivanova, K.I.; Marchenko, V.Y.; Ilyicheva, T.N.; et al. Immunogenicity of Experimental DNA Vaccines Encoding Hemagglutinin and Hemagglutinin Stalk of Influenza A (H5N8) Virus. Сибирский научный медицинский журнал 2025, 45, 100–108. [Google Scholar] [CrossRef]
- Rudometov, A.P.; Litvinova, V.R.; Gudymo, A.S.; Ivanova, K.I.; Rudometova, N.B.; Kisakov, D.N.; Borgoyakova, M.B.; Kisakova, L.A.; Yakovlev, V.A.; Tigeeva, E.V.; et al. Dose-Dependent Effect of DNA Vaccine PVAX-H5 Encoding a Modified Hemagglutinin of Influenza A (H5N8) and Its Cross-Reactivity Against A (H5N1) Influenza Viruses of Clade 2.3.4.4b. Viruses 2025, 17, 330. [Google Scholar] [CrossRef]
- Pyankova, O.G.; Susloparov, I.M.; Moiseeva, A.A.; Kolosova, N.P.; Onkhonova, G.S.; Danilenko, A.V.; Vakalova, E.V.; Shendo, G.L.; Nekeshina, N.N.; Noskova, L.N.; et al. Isolation of Clade 2.3.4.4b A(H5N8), a Highly Pathogenic Avian Influenza Virus, from a Worker during an Outbreak on a Poultry Farm, Russia, December 2020. Eurosurveillance 2021, 26. [Google Scholar] [CrossRef]
- Marchenko, V.Y.; Svyatchenko, S.V.; Onkhonova, G.S.; Goncharova, N.I.; Ryzhikov, A.B.; Maksyutov, R.A.; Gavrilova, E.V. Review on the Epizootiological Situation on Highly Pathogenic Avian Influenza around the World and in Russia in 2022. Problems of Particularly Dangerous Infections 2023, 48–55. [Google Scholar] [CrossRef]
- Borgoyakova, M.B.; Karpenko, L.I.; Rudometov, A.P.; Volosnikova, E.A.; Merkuleva, I.A.; Starostina, E.V.; Zadorozhny, A.M.; Isaeva, A.A.; Nesmeyanova, V.S.; Shanshin, D.V.; et al. Self-Assembled Particles Combining SARS-CoV-2 RBD Protein and RBD DNA Vaccine Induce Synergistic Enhancement of the Humoral Response in Mice. Int J Mol Sci 2022, 23, 2188. [Google Scholar] [CrossRef]
- World Health Organization. Manual for the Laboratory Diagnosis and Virological Surveillance of Influenza : WHO Global Influenza Surveillance Network; WHO Press: Geneva, Switzerland, 2011; pp. 63–79. [Google Scholar]
- Decree of the Chief State Sanitary Doctor of the Russian Federation of January 28, 2021 N 4 On Approval of Sanitary Rules and Norms SanPiN 3.3686-21 “Sanitary and Epidemiological Requirements for the Prevention of Infectious Diseases”. Available online: https://Www.Rospotrebnadzor.Ru/Files/News/SP_infections_compressed.Pdf (accessed on 15 May 2025). (In Russian).
- World Health Organization. Antigenic and Genetic Characteristics of Zoonotic Influenza A Viruses and Development of Candidate Vaccine Viruses for Pandemic Preparedness. Available online: https://cdn.who.int/media/docs/default-source/influenza/whoinfluenza-recommendations/vcm-northern-hemisphere-recommendation-2022-2023/202203_zoonotic_vaccinevirusupdate.pdf.
- Mourdikoudis, S.; Pallares, R.M.; Thanh, N.T.K. Characterization Techniques for Nanoparticles: Comparison and Complementarity upon Studying Nanoparticle Properties. Nanoscale 2018, 10, 12871–12934. [Google Scholar] [CrossRef]
- Chiba, S.; Kiso, M.; Yamada, S.; Someya, K.; Onodera, Y.; Yamaguchi, A.; Matsunaga, S.; Uraki, R.; Iwatsuki-Horimoto, K.; Yamayoshi, S.; et al. An MRNA Vaccine Candidate Encoding H5HA Clade 2.3.4.4b Protects Mice from Clade 2.3.2.1a Virus Infection. NPJ Vaccines 2024, 9, 189. [Google Scholar] [CrossRef]
- Hawman, D.W.; Tipih, T.; Hodge, E.; Stone, E.T.; Warner, N.; McCarthy, N.; Granger, B.; Meade-White, K.; Leventhal, S.; Hatzakis, K.; et al. Clade 2.3.4.4b but Not Historical Clade 1 HA Replicating RNA Vaccine Protects against Bovine H5N1 Challenge in Mice. Nat Commun 2025, 16, 655. [Google Scholar] [CrossRef]
- Karikó, K.; Muramatsu, H.; Ludwig, J.; Weissman, D. Generating the Optimal MRNA for Therapy: HPLC Purification Eliminates Immune Activation and Improves Translation of Nucleoside-Modified, Protein-Encoding MRNA. Nucleic Acids Res 2011, 39, e142–e142. [Google Scholar] [CrossRef]
- Michel, T.; Luft, D.; Abraham, M.-K.; Reinhardt, S.; Salinas Medina, M.L.; Kurz, J.; Schaller, M.; Avci-Adali, M.; Schlensak, C.; Peter, K.; et al. Cationic Nanoliposomes Meet MRNA: Efficient Delivery of Modified MRNA Using Hemocompatible and Stable Vectors for Therapeutic Applications. Mol Ther Nucleic Acids 2017, 8, 459–468. [Google Scholar] [CrossRef]
- Hatziantoniou, S.; Maltezou, H.C.; Tsakris, A.; Poland, G.A.; Anastassopoulou, C. Anaphylactic Reactions to MRNA COVID-19 Vaccines: A Call for Further Study. Vaccine 2021, 39, 2605–2607. [Google Scholar] [CrossRef]
- Pniewska, E.; Pawliczak, R. The Involvement of Phospholipases A2 in Asthma and Chronic Obstructive Pulmonary Disease. Mediators Inflamm 2013, 2013, 1–12. [Google Scholar] [CrossRef]
- Banerji, A.; Wickner, P.G.; Saff, R.; Stone, C.A.; Robinson, L.B.; Long, A.A.; Wolfson, A.R.; Williams, P.; Khan, D.A.; Phillips, E.; et al. MRNA Vaccines to Prevent COVID-19 Disease and Reported Allergic Reactions: Current Evidence and Suggested Approach. J Allergy Clin Immunol Pract 2021, 9, 1423–1437. [Google Scholar] [CrossRef]
- Trougakos, I.P.; Terpos, E.; Alexopoulos, H.; Politou, M.; Paraskevis, D.; Scorilas, A.; Kastritis, E.; Andreakos, E.; Dimopoulos, M.A. Adverse Effects of COVID-19 MRNA Vaccines: The Spike Hypothesis. Trends Mol Med 2022, 28, 542–554. [Google Scholar] [CrossRef]
- Echaide, M.; Chocarro de Erauso, L.; Bocanegra, A.; Blanco, E.; Kochan, G.; Escors, D. MRNA Vaccines against SARS-CoV-2: Advantages and Caveats. Int J Mol Sci 2023, 24, 5944. [Google Scholar] [CrossRef]
- Kisakov, D.N.; Kisakova, L.A.; Sharabrin, S.V.; Yakovlev, V.A.; Tigeeva, E.V.; Borgoyakova, M.B.; Starostina, E.V.; Zaikovskaya, A.V.; Rudometov, A.P.; Rudometova, N.B.; et al. Delivery of Experimental MRNA Vaccine Encoding the RBD of SARS-CoV-2 by Jet Injection. Bull Exp Biol Med 2024, 176, 776–780. [Google Scholar] [CrossRef]
- Mooij, P.; Grødeland, G.; Koopman, G.; Andersen, T.K.; Mortier, D.; Nieuwenhuis, I.G.; Verschoor, E.J.; Fagrouch, Z.; Bogers, W.M.; Bogen, B. Needle-Free Delivery of DNA: Targeting of Hemagglutinin to MHC Class II Molecules Protects Rhesus Macaques against H1N1 Influenza. Vaccine 2019, 37, 817–826. [Google Scholar] [CrossRef]
- Mulligan, M.J.; Lyke, K.E.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Neuzil, K.; Raabe, V.; Bailey, R.; Swanson, K.A.; et al. Phase I/II Study of COVID-19 RNA Vaccine BNT162b1 in Adults. Nature 2020, 586, 589–593. [Google Scholar] [CrossRef]
- Sahin, U.; Muik, A.; Derhovanessian, E.; Vogler, I.; Kranz, L.M.; Vormehr, M.; Baum, A.; Pascal, K.; Quandt, J.; Maurus, D.; et al. COVID-19 Vaccine BNT162b1 Elicits Human Antibody and TH1 T Cell Responses. Nature 2020, 586, 594–599. [Google Scholar] [CrossRef]
- Dombu, C.Y.; Kroubi, M.; Zibouche, R.; Matran, R.; Betbeder, D. Characterization of Endocytosis and Exocytosis of Cationic Nanoparticles in Airway Epithelium Cells. Nanotechnology 2010, 21, 355102. [Google Scholar] [CrossRef]
- Palatini, P. Disposition Kinetics of Phospholipid Liposomes. In Neurobiology of Essential Fatty Acids. Advances in Experimental Medicine and Biology; Bazan, N.G., Murphy, M.G., Toffano, G., Eds.; Springer: Boston, MA, USA, 1992; Volume 318, pp. 375–391. [Google Scholar] [CrossRef]
- Heyes, J.; Palmer, L.; Bremner, K.; MacLachlan, I. Cationic Lipid Saturation Influences Intracellular Delivery of Encapsulated Nucleic Acids. Journal of Controlled Release 2005, 107, 276–287. [Google Scholar] [CrossRef]
Figure 1.
Structure and production of the mRNA-H5 vaccine. (a) Schematic representation of mRNA: mRNA has a 5'-cap, poly(A) tail, 5'- and 3'-untranslated regions of human α-globin and an open reading frame encoding the HA of HPAI A/H5N8 virus); schematic structure of the modified hemagglutinin gene; the model of the hemagglutinin trimer encoded by mRNA-H5 (the model was obtained as described in [38]). (b) Electrophoregram of mRNA-H5 synthesis products in 1% agarose gel: lane 1 – ssRNA Ladder (New England Biolabs); lane 2,3 – mRNA-H5..
Figure 1.
Structure and production of the mRNA-H5 vaccine. (a) Schematic representation of mRNA: mRNA has a 5'-cap, poly(A) tail, 5'- and 3'-untranslated regions of human α-globin and an open reading frame encoding the HA of HPAI A/H5N8 virus); schematic structure of the modified hemagglutinin gene; the model of the hemagglutinin trimer encoded by mRNA-H5 (the model was obtained as described in [38]). (b) Electrophoregram of mRNA-H5 synthesis products in 1% agarose gel: lane 1 – ssRNA Ladder (New England Biolabs); lane 2,3 – mRNA-H5..
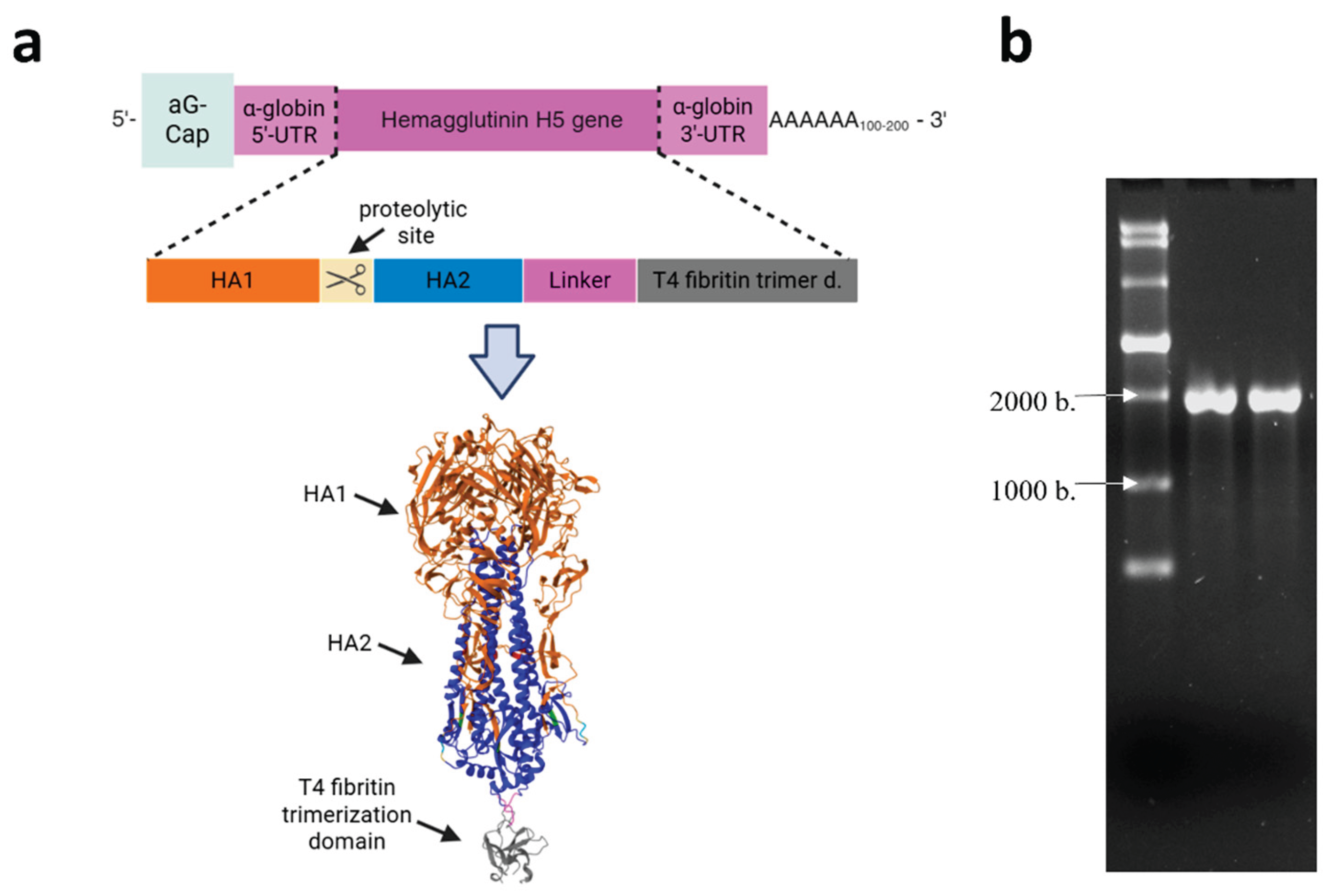
Figure 2.
Preparation and characterization of LNPs formulations. (a) Electron micrograph of complexes: (1) mRNA-H5-LNP; (2) LNPs without mRNA. (b) Dynamic light scattering: size distribution profiles of nanoparticles obtained with and without mRNA. .
Figure 2.
Preparation and characterization of LNPs formulations. (a) Electron micrograph of complexes: (1) mRNA-H5-LNP; (2) LNPs without mRNA. (b) Dynamic light scattering: size distribution profiles of nanoparticles obtained with and without mRNA. .
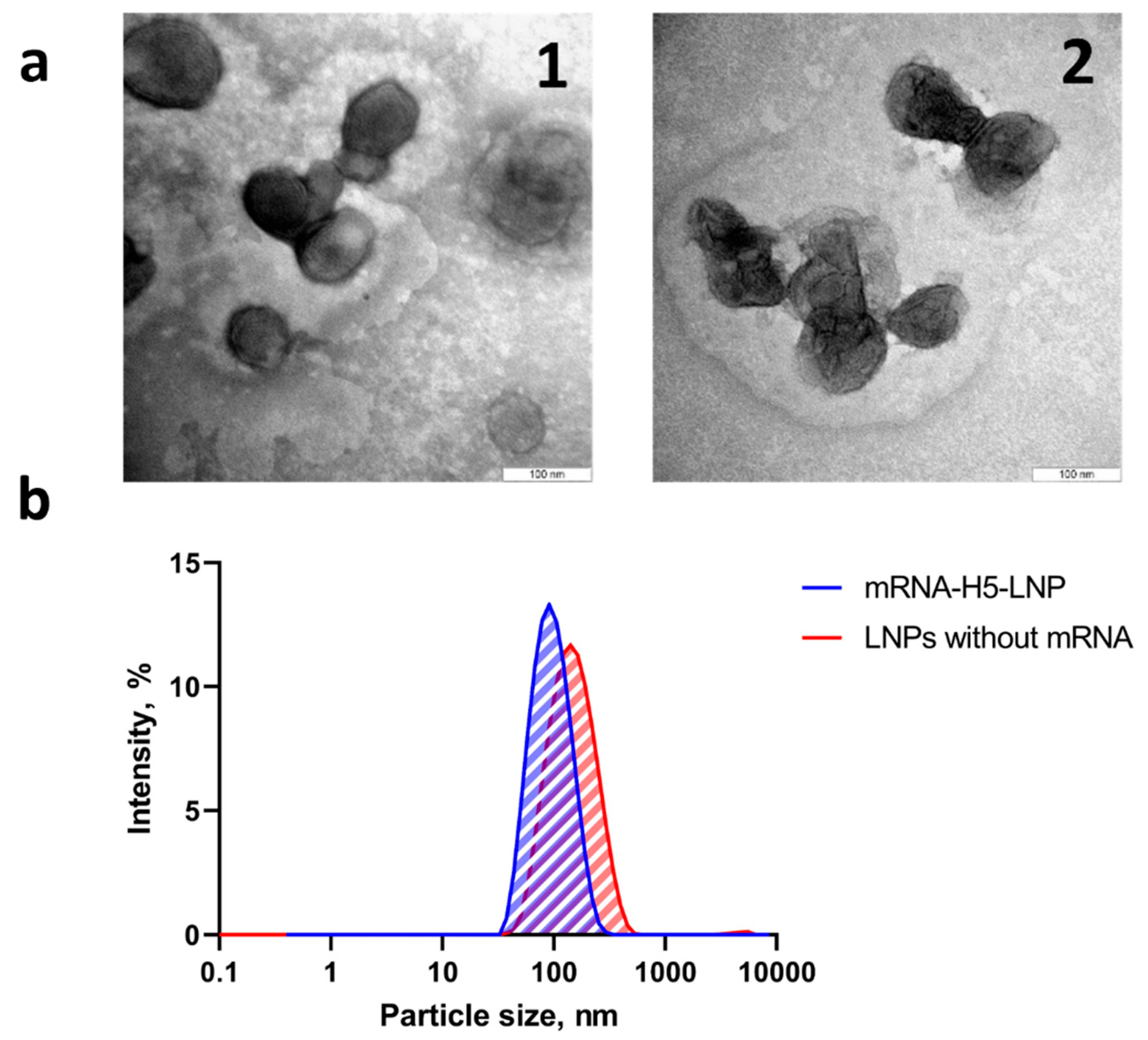
Figure 3.
Analysis of humoral immune response to mRNA-H5. (a) Reciprocal titer (dilution) of H5-specific antibodies detected in immune sera using ELISA. (b) The neutralization activity of immune sera against influenza virus A/turkey/Stavropol/320-01/2020 (H5N8). Reciprocal titer values are provided in the plot. mRNA-H5-LNP – a group of animals immunized with mRNA-H5 delivered by LNPs; mRNA-H5 JI – a group of animals immunized with mRNA-H5 delivered by JI; Recombinant protein HA/H5 – a group of animals immunized with recombinant protein HA/H5; LNPs without mRNA – a group of animals immunized with LNPs without mRNA; Intact – a group of animals that have not been subjected to any manipulation. In panels data are provided as mean with range. Significance was calculated using non-parametric one-way Kruskal–Wallis analysis of variance with correction for multiple comparisons and Dunn’s statistical hypothesis test (* p < 0.05, ** p < 0.01, **** p < 0.0001). .
Figure 3.
Analysis of humoral immune response to mRNA-H5. (a) Reciprocal titer (dilution) of H5-specific antibodies detected in immune sera using ELISA. (b) The neutralization activity of immune sera against influenza virus A/turkey/Stavropol/320-01/2020 (H5N8). Reciprocal titer values are provided in the plot. mRNA-H5-LNP – a group of animals immunized with mRNA-H5 delivered by LNPs; mRNA-H5 JI – a group of animals immunized with mRNA-H5 delivered by JI; Recombinant protein HA/H5 – a group of animals immunized with recombinant protein HA/H5; LNPs without mRNA – a group of animals immunized with LNPs without mRNA; Intact – a group of animals that have not been subjected to any manipulation. In panels data are provided as mean with range. Significance was calculated using non-parametric one-way Kruskal–Wallis analysis of variance with correction for multiple comparisons and Dunn’s statistical hypothesis test (* p < 0.05, ** p < 0.01, **** p < 0.0001). .
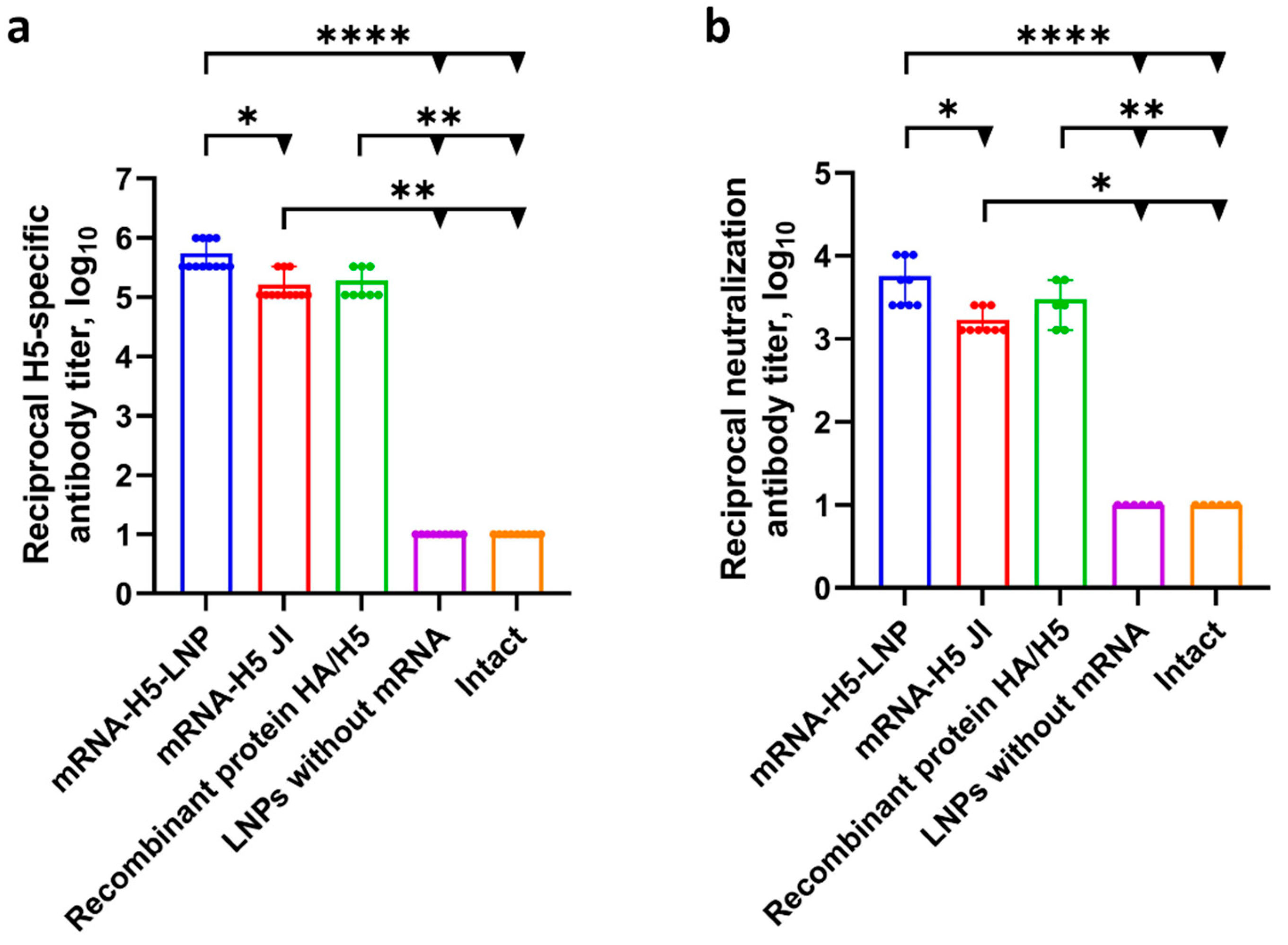
Figure 4.
Percentage of HA-specific cytokine-producing CD3+/CD4+ and CD3+/CD8+ lymphocytes analyzed using the ICS method combined with flow cytometry. mRNA-H5-LNP – a group of animals immunized with mRNA-H5 delivered by LNPs; mRNA-H5 JI – a group of animals immunized with mRNA-H5 delivered by JI; Recombinant protein HA/H5 – a group of animals immunized with recombinant protein HA/H5; LNPs without mRNA – a group of animals immunized with LNPs without mRNA; Intact – a group of animals that have not been subjected to any manipulation. Individual values are represented by dots; group medians – by lines; mean values – by pluses. p < 0.05 compared ⁰with LNPs without mRNA; *with intact mice. .
Figure 4.
Percentage of HA-specific cytokine-producing CD3+/CD4+ and CD3+/CD8+ lymphocytes analyzed using the ICS method combined with flow cytometry. mRNA-H5-LNP – a group of animals immunized with mRNA-H5 delivered by LNPs; mRNA-H5 JI – a group of animals immunized with mRNA-H5 delivered by JI; Recombinant protein HA/H5 – a group of animals immunized with recombinant protein HA/H5; LNPs without mRNA – a group of animals immunized with LNPs without mRNA; Intact – a group of animals that have not been subjected to any manipulation. Individual values are represented by dots; group medians – by lines; mean values – by pluses. p < 0.05 compared ⁰with LNPs without mRNA; *with intact mice. .
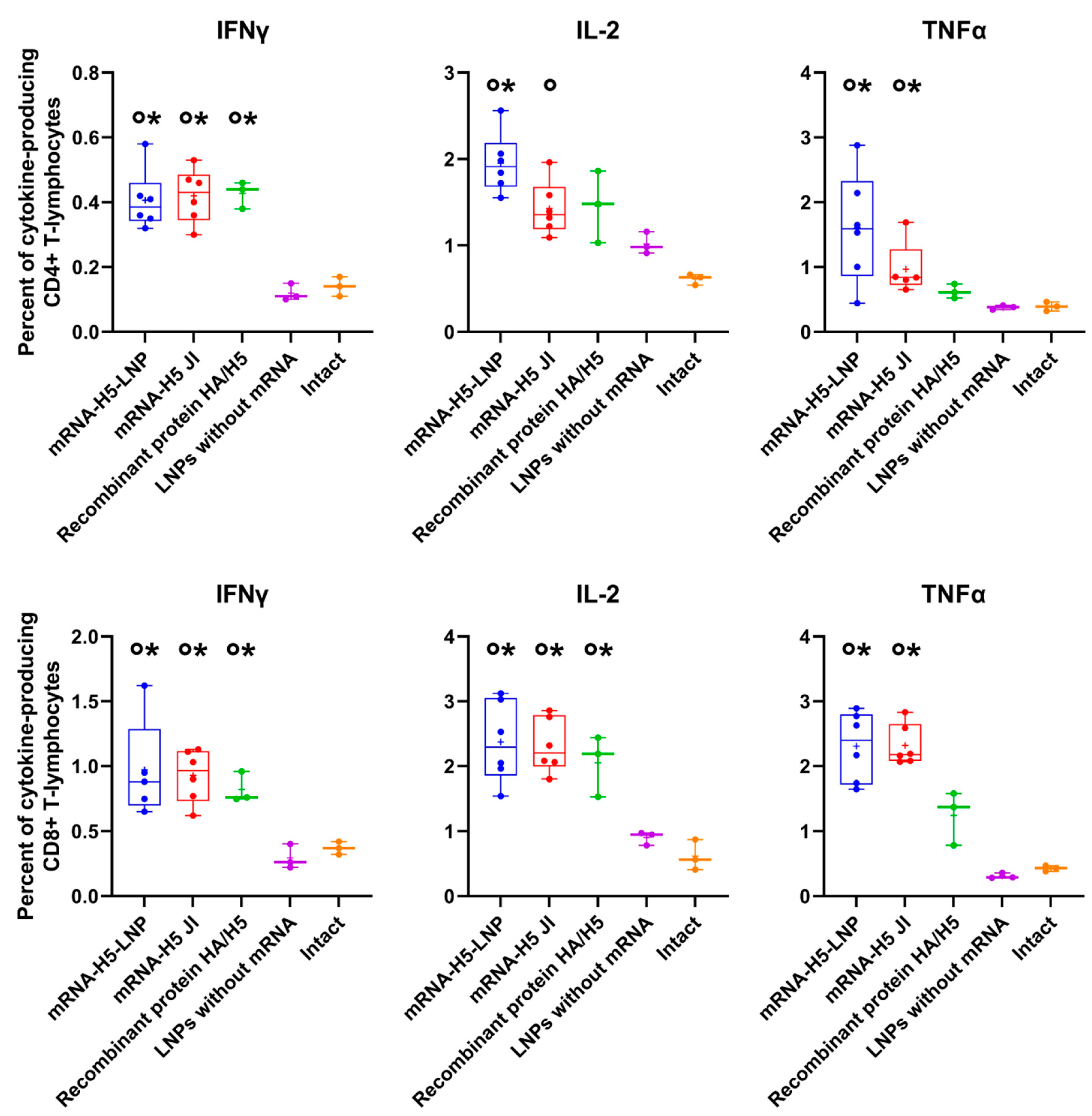
Figure 5.
Analysis of mRNA-H5 protective properties. (a) Analysis of humoral immune response before challenge. Reciprocal titer (dilution) of H5-specific antibodies detected in immune sera using ELISA. Data are provided as mean with range. (* p< 0.05; ** p< 0.01; **** p< 0.0001). Significance was calculated using non-parametric one-way Kruskal–Wallis analysis of variance with correction for multiple comparisons and Dunn’s statistical hypothesis test (** p < 0.01, *** p < 0.001). (b,c) Survival curves of immunized animals after infection with the influenza virus strain: (b) A/Astrakhan/3212/2020 (H5N8); (c) A/chicken/Magadan/14-7V/2022 (H5N1). Survival function modeling was performed using the Kaplan–Meier multiplier estimator, and comparison of survival with the control group was performed using the Mantel–Cox test. (d,e) Weight change curves of mice in a challenge study against strain: (d) A/Astrakhan/3212/2020 (H5N8); (e) A/chicken/Magadan/14-7V/2022 (H5N1). Data are presented as mean with standard error of the mean. mRNA-H5-LNP – a group of animals immunized with mRNA-H5 delivered by LNPs; mRNA-H5 JI – a group of animals immunized with mRNA-H5 delivered by JI; Recombinant protein HA/H5 – a group of animals immunized with recombinant protein HA/H5; Intact – a group of animals that have not been subjected to any manipulation.
Figure 5.
Analysis of mRNA-H5 protective properties. (a) Analysis of humoral immune response before challenge. Reciprocal titer (dilution) of H5-specific antibodies detected in immune sera using ELISA. Data are provided as mean with range. (* p< 0.05; ** p< 0.01; **** p< 0.0001). Significance was calculated using non-parametric one-way Kruskal–Wallis analysis of variance with correction for multiple comparisons and Dunn’s statistical hypothesis test (** p < 0.01, *** p < 0.001). (b,c) Survival curves of immunized animals after infection with the influenza virus strain: (b) A/Astrakhan/3212/2020 (H5N8); (c) A/chicken/Magadan/14-7V/2022 (H5N1). Survival function modeling was performed using the Kaplan–Meier multiplier estimator, and comparison of survival with the control group was performed using the Mantel–Cox test. (d,e) Weight change curves of mice in a challenge study against strain: (d) A/Astrakhan/3212/2020 (H5N8); (e) A/chicken/Magadan/14-7V/2022 (H5N1). Data are presented as mean with standard error of the mean. mRNA-H5-LNP – a group of animals immunized with mRNA-H5 delivered by LNPs; mRNA-H5 JI – a group of animals immunized with mRNA-H5 delivered by JI; Recombinant protein HA/H5 – a group of animals immunized with recombinant protein HA/H5; Intact – a group of animals that have not been subjected to any manipulation.
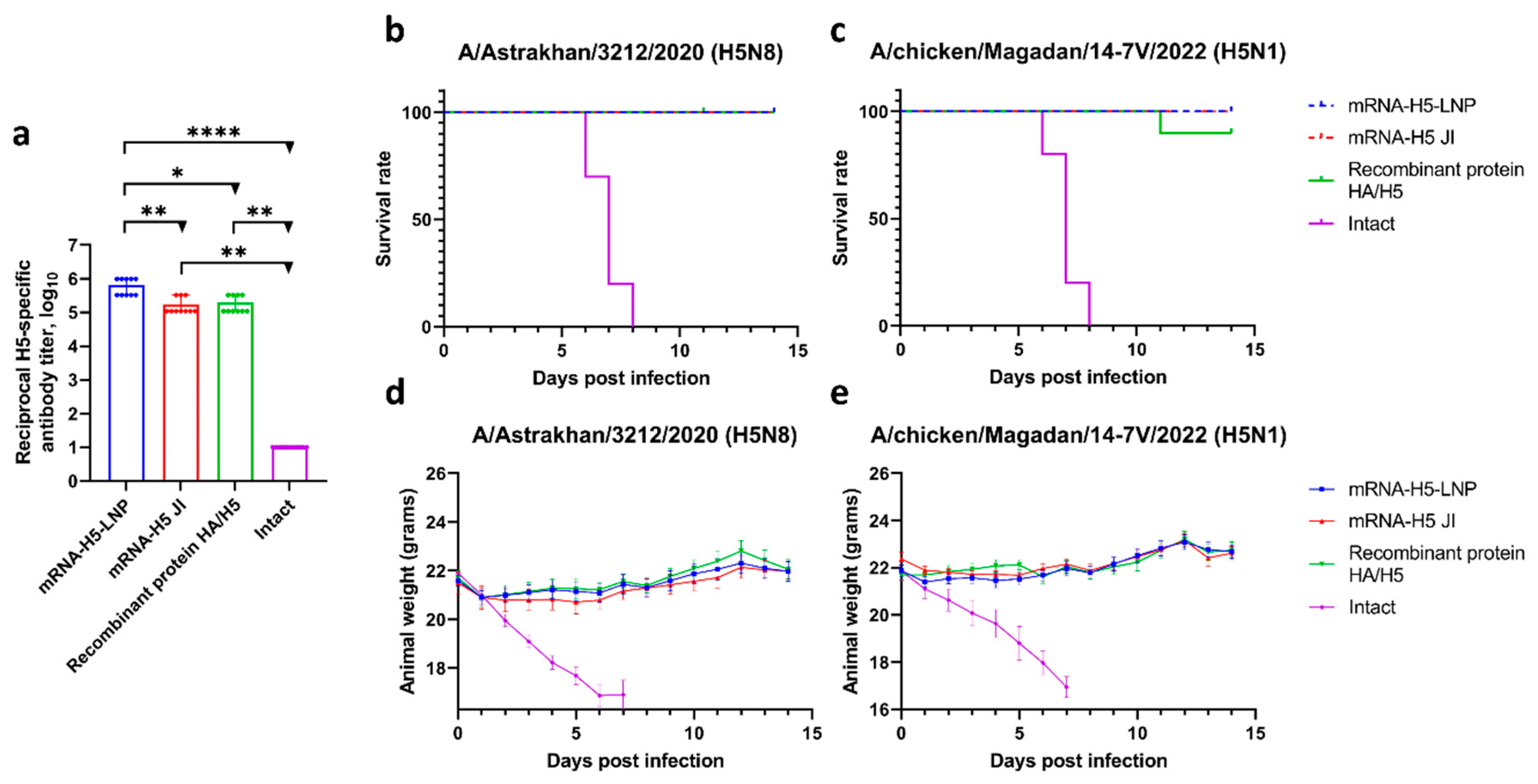

Table 1.
Physical characteristics of the obtained LNPs formulations.
| Formulations | Average particle size, nm | Polydispersity index | ζ - potential |
|---|---|---|---|
| mRNA-H5-LNP | 93.5±0.8 | 0.149 ± 0.01 | -0.02±0.26 |
| LNPs without mRNA | 131.6±1.6 | 0.175 ± 0.01 | 1.15±0.26 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



