Submitted:
30 June 2025
Posted:
01 July 2025
You are already at the latest version
Abstract
Nucleoside modifications to the template of mRNA vaccines were important in achieving the desired level of antigen expression and immune response for the vaccine platform to function successfully during the COVID-19 pandemic. The innovation solved a crucial issue and resulted in the awarding of the 2023 Nobel Prize in Medicine to Katalin Karikó and Drew Weissman. Despite the spotlight on nucleoside modifications, several important aspects of its behavior in living systems were only discovered recently, particularly context specific translational accuracy, and the stimulation of +1 ribosomal frameshifting at specific “slippery sequences”. The latter has been observed in living subjects (mouse and human) to produce both off-target antigens and an off-target immune response. While adverse effects cannot currently be attributed to poor translational fidelity, this may spell issues for future iterations of mRNA therapeutics if not addressed. Fortunately, in most cases, the transcript sequence can be modified to allay these concerns and produce faithful protein products, by avoiding sequence contexts resulting in either incorrect amino acid incorporation or +1 frameshifting. The modularity of the mRNA platform makes these fixes easy to incorporate, as long as this is accounted for in future mRNA therapeutics design.
Keywords:
n1-methyl-pseudouridine
; mRNA vaccines
; Pfizer/BioNTech BNT162b2
; Moderna mRNA-1273
; ribosomal frameshifting
Introduction
Messenger RNA (mRNA) vaccines were well poised to respond to the challenge of the COVID-19 pandemic. While SARS-CoV-2 was spreading rapidly, the teams at Pfizer/BioNTech and Moderna produced prototype vaccines merely from the sequence released on January 10, 2020[1]. While previous generations of vaccines required laborious culturing from infectious material, Pfizer/BioNTech and Moderna were able to leverage the digital revolution in biology, and develop the vaccine sequence based on the publicly available genome of SARS-CoV-2 on GenBank [1].
At the core of the vaccine is the genetic code. All of the major COVID-19 vaccines, mRNA and otherwise, express the spike (S) protein from SARS-CoV-2, with some modifications. Importantly, for the vaccines to function, the genetic code needs to remain intact long enough to enter a cell, to find a ribosome, and be translated by the ribosome to produce the antigen, in this case the modified SARS-CoV-2 S protein.
Early mRNA vaccines used naked mRNA, composed of the same bases as mRNA in all organisms, cytosine (C), guanine (G), adenine (A) and uridine (U), which takes the place of the DNA base thymine (T) [2]. The development of mRNA vaccines demonstrated promising results in generating targeted immune responses and validating the platform's core concept [3]. However, initial formulations faced significant challenges due to rapid degradation of unprotected mRNA strands by host ribonucleases and unintended activation of innate immune pathways [4]. These biological defense mechanisms, which normally break down cellular transcripts or detect viral/bacterial RNA, often intercepted therapeutic mRNA before it could trigger the desired adaptive immunity [3]. Researchers subsequently addressed these issues through innovations in nucleotide modification and lipid nanoparticle delivery systems [5].In 2005, Karikó and Weissman demonstrated that by modifying the uridines to pseudouridines (Ψ), the immune response was lessened, and the modified mRNA was able to remain intact and express the target antigen in greater amounts than with naked mRNA [6](Figure 1). A further upgrade came in 2015, when experiments demonstrated that n1-methyl-pseudouridine (m1Ψ) modifications increased the translational efficiency further still [7]. However, although this was not an active concern at the time, Ψ-modified RNA was later found to exhibit reduced translational fidelity compared to naked RNA, which may result in the production of unintended proteins [8].. In 2019, m1Ψ-RNA demonstrated high translational fidelity compared to Ψ-RNA, at a similar level to naked mRNA, the standard for translational fidelity [9]. Other developments critical for the development of mRNA vaccines are shown in Figure 1, adapted from the events in [5]. These are broken down by events related to the development of mRNA vaccines [5,10,11,12,13,14,15], events related to pseudouridine [6,16,17,18], and events related to n1-methylpseudouridine [8,19,20,21,22,23,24,25,26,27].
The importance of these modifications to the effectiveness of mRNA vaccines is illustrated by a counterexample. CureVac, founded in 2000 to deliver RNA therapeutics, uses unmodified mRNA with codon-optimized sequences selected to minimize the use of uridine [28], as GU sequences can stimulate toll like receptors 7 and 8 to provoke an immune response [29,30]. However, CureVac’s prospective vaccine performed comparatively poorly compared to the modified nucleoside mRNA vaccines, with the CureVac’s vaccine achieving a mere 48% efficacy [31], significantly lower than the modified nucleoside mRNA vaccines [14,15]. CureVac has changed to using modified nucleosides in its recent vaccine candidates [32].
In short, nucleoside modifications are important for mRNA vaccines, as they allow the genetic code to enter the cytoplasm of the cell and be expressed [33]. Chemical modification of RNA allows them to escape host immune response [6,34,35] and also independently increase host ribosomal density [36]. Both of these factors increase the number of copies of the antigen produced. Karikó and Weissman's discovery enabled mRNA vaccines to express enough of the antigen for recognition by the host immune system [37]. Compared to unmodified RNA, pseudouridinylated RNA is translated roughly 9x more [18].
The discovery solved the crucial challenge of mRNA vaccines, allowing the lipid nanoparticle-encapsulated mRNA to produce amounts of the target antigen sufficient to stimulate a robust immune response. In 2020, their decades of work came to fruition when clinical trials of the Pfizer and Moderna messenger RNA vaccines, both using n1-methyl-pseudouridine in place of uridine, demonstrated efficacies of 95% (95% Confidence interval (CI), 90.3,97.6%) [14] and 94.1% (95% CI, 89.3 to 96.8%) [15], respectively. Katalin Karikó and Drew Weissman were awarded the 2023 Nobel Prize in Medicine for their breakthrough development of the chemical modifications of mRNA [38], which solved a crucial challenge in mRNA vaccines, and are likely responsible for millions of lives saved from COVID-19 [39].
The Biology of Ψ and m1Ψ
The nucleobases of RNA were elucidated during the first half of the 20th century [40]. Pseudouridine, the first RNA modification to be discovered, and the most ubiquitous, was first discovered in 1951 in ribonucleic acid extracted from calf liver [16]. Following its discovery, over 150 additional nucleic acid modifications have been discovered [41], with wide ranging effects on gene expression [42]. Pseudouridine is ubiquitous throughout diverse organisms [43], including humans, where 0.2 to 0.6% of human RNA uridines are converted to pseudouridines by endogenous enzymes [44].
N1-methyl-pseudouridine was first discovered in 1976 in Streptomyces platensis, a bacterium [19]. Later discoveries of m1Ψ in 1982 in an archaea (Halococcus morrhuae) transfer RNA [21] and again in 1984 in Halobacterium volcanii [22]. Additional archaea species containing m1 Ψ have since been discovered, shown in Figure 2, including Methanococcoides burtonii [45], Methanosarcina barkeri [46], Archaeoglobus fulgidus [47], Methanococcus voltae [48], Methanococcus jannaschii [49], Methanomicrobium mobile [50], Halococcus morrhuae [51], Haloferax volcanii [52], Halobacterium salinarum [53], Pyrobaculum islandicum [54], Thermoproteus neutrophilus [55], Methanococcus igneus [56], Methanothermus fervidus [57] and Methanococcus vannielii [58].
In archaea, m1Ψ has only been found at position 54 of tRNA (Figure 3A). In Saccharomyces cerevisiae, the enzyme Nep1 catalyzes the conversion of the uridine at position 1191 of 18S ribosomal RNA (rRNA) into m1Ψ (Figure 3B). While, to our knowledge, m1Ψ has not been directly observed in other eukaryotic cells, it is plausible that m1Ψ may be present in the 18S rRNA of other eukaryotic species sharing Nep1 or an ortholog [59]. m1Ψ is thus rarely observed in nature, and its ubiquitous use as artificially incorporated in mRNA COVID-19 vaccines is without precedent.
Important Differences from Uridine
Little is known about the role of m1 Ψ in nature, though recent experiments have elucidated how it behaves in biological systems. As m1Ψ has a different chemical structure than both U and Ψ, there are subtle differences in base pairing energies [8,60]. This may result in context-specific translational fidelity, which has been observed for m1Ψ [8]. Earlier work on the translational fidelity of m1Ψ had characterized tRNA misincorporation levels for a subset of possible codons, finding accuracy similar to uridine-containing mRNA codons [9]. The signal of altered translational fidelity may have been missed in these initial experiments. The error rate for m1Ψ-incorporated transcripts, while lesser than that for Ψ-incorporated transcripts, was 25% higher (p=0.006) than for U-containing transcripts (Figure 4A) [61].
Another challenge to translational fidelity is m1Ψ-rich sequences inducing +1 translational frameshifting and producing off-target proteins [27] (Figure 4B). This appears to impact the immune response, as antibodies for the frameshifted proteins were observed in both mice and humans injected with BNT162b2 (Pfizer/BioNTech vaccine) [27]. Fortunately, there have not been adverse events attributed to these off-target proteins. However, given the panoply of possible mRNA templates in future products, a space which expands considerably when personalized mRNA transcripts are used for cancer vaccines or gene therapy, it may be difficult to characterize the biological effects of all possible off-target proteins.
Resolution: Moving Forward with mRNA
Fortunately, introducing synonymous mutations to the mRNA sequence to avoid “slippery sequences”, the specific short sequences which induce +1 ribosomal frameshifting, can limit this issue [27,62]. Fortunately, the slippery sequences appear to be quite specific, and may be avoided without changing the desirable properties of m1Ψ, namely, its decreased immunogenecity, and increased translational efficiency, compared to both unmodified RNA and Ψ-RNA [24]. In the Pfizer/BioNTech BNT162b2 sequence, for example, there are six predicted +1 ribosomal frameshift sites [27]; introduction of synonymous mutations at these six locations may be sufficient to reduce the production of off-target proteins to the same level as in unmodified RNA [27,62].
With dozens of mRNA therapeutic products in development, approval and marketing pipelines [63,64], it is timely that these design challenges were spotted. The strength of mRNA vaccines is their ability to go from sequence to prototype quickly [65], which makes the changes to ensure faithful translation easy to apply to future iterations of the technology. Further studies should be undertaken to elucidate additional potential biological impacts and dose effects of m1Ψ in mRNA therapeutics, considering their extreme rare presentation in nature, particularly in eukaryotic cells.
References
- Severe acute respiratory syndrome coronavirus 2 isolate Wuhan-Hu-1, complete genome [Internet]. 2020 [cited 2025 May 3]. Available from: http://www.ncbi.nlm.nih.gov/nuccore/NC_045512.2.
- Pardi N, Hogan MJ, Porter FW, Weissman D. mRNA vaccines — a new era in vaccinology. Nat Rev Drug Discov. 2018, 17, 261–79. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary N, Weissman D, Whitehead KA. mRNA vaccines for infectious diseases: principles, delivery and clinical translation. Nat Rev Drug Discov. 2021, 20, 817–38. [Google Scholar] [CrossRef] [PubMed]
- Cheng F, Wang Y, Bai Y, Liang Z, Mao Q, Liu D, et al. Research Advances on the Stability of mRNA Vaccines. Viruses 2023, 15, 668. [Google Scholar] [CrossRef] [PubMed]
- Dolgin E. The tangled history of mRNA vaccines. Nature. 2021 Sep 14;597(7876):318–24.
- Karikó K, Buckstein M, Ni H, Weissman D. Suppression of RNA Recognition by Toll-like Receptors: The Impact of Nucleoside Modification and the Evolutionary Origin of RNA. Immunity 2005, 23, 165–75. [Google Scholar] [CrossRef] [PubMed]
- Pardi N, Tuyishime S, Muramatsu H, Kariko K, Mui BL, Tam YK, et al. Expression kinetics of nucleoside-modified mRNA delivered in lipid nanoparticles to mice by various routes. J Control Release Off J Control Release Soc. 2015 Nov 10;217:345–51.
- Monroe J, Eyler DE, Mitchell L, Deb I, Bojanowski A, Srinivas P, et al. N1-Methylpseudouridine and pseudouridine modifications modulate mRNA decoding during translation. Nat Commun. 2024, 15, 8119. [Google Scholar] [CrossRef] [PubMed]
- Kim KQ, Burgute BD, Tzeng SC, Jing C, Jungers C, Zhang J, et al. N1-methylpseudouridine found within COVID-19 mRNA vaccines produces faithful protein products. Cell Rep [Internet]. 2022 Aug 30 [cited 2022 Oct 5];40(9). Available from: https://www.cell.com/cell-reports/abstract/S2211-1247(22)01120-2.
- Melton DA, Krieg PA, Rebagliati MR, Maniatis T, Zinn K, Green MR. Efficient in vitro synthesis of biologically active RNA and RNA hybridization probes from plasmids containing a bacteriophage SP6 promoter. Nucleic Acids Res. 1984, 12, 7035–56. [Google Scholar] [CrossRef] [PubMed]
- Malone RW, Felgner PL, Verma IM. Cationic liposome-mediated RNA transfection. Proc Natl Acad Sci. 1989 Aug;86(16):6077–81.
- Felgner PL, Wolff JA, Rhodes GH, Malone RW, Carson DA. Induction of a protective immune response in a mammal by injecting a DNA sequence. Biotechnol Adv. 1997;15(3–4):663.
- Alberer M, Gnad-Vogt U, Hong HS, Mehr KT, Backert L, Finak G, et al. Safety and immunogenicity of a mRNA rabies vaccine in healthy adults: an open-label, non-randomised, prospective, first-in-human phase 1 clinical trial. Lancet Lond Engl. 2017 Sep 23;390(10101):1511–20.
- Polack FP, Thomas SJ, Kitchin N, Absalon J, Gurtman A, Lockhart S, et al. Safety and Efficacy of the BNT162b2 mRNA Covid-19 Vaccine. N Engl J Med. 2020 Dec 31;383(27):2603–15.
- Baden LR, El Sahly HM, Essink B, Kotloff K, Frey S, Novak R, et al. Efficacy and Safety of the mRNA-1273 SARS-CoV-2 Vaccine. N Engl J Med. 2021 Feb 4;384(5):403–16.
- Cohn WE, Volkin E. Nucleoside-5′-Phosphates from Ribonucleic Acid. Nature. 1951 Mar;167(4247):483–4.
- Cortese R, Kammen HO, Spengler SJ, Ames BN. Biosynthesis of Pseudouridine in Transfer Ribonucleic Acid. J Biol Chem. 1974 Feb 25;249(4):1103–8.
- Karikó K, Muramatsu H, Welsh FA, Ludwig J, Kato H, Akira S, et al. Incorporation of Pseudouridine Into mRNA Yields Superior Nonimmunogenic Vector With Increased Translational Capacity and Biological Stability. Mol Ther. 2008 Nov 1;16(11):1833–40.
- Argoudelis AD, Mizsak SA. 1-methylpseudouridine, a metabolite of Streptomyces platensis. J Antibiot (Tokyo). 1976 Aug;29(8):818–23.
- Earl RA, Townsend LB. A chemical synthesis of the nucleoside 1-methylpseudouridine. J Heterocycl Chem. 1977;14(4):699–700.
- Pang H, Ihara M, Kuchino Y, Nishimura S, Gupta R, Woese CR, et al. Structure of a modified nucleoside in archaebacterial tRNA which replaces ribosylthymine. 1-Methylpseudouridine. J Biol Chem. 1982 Apr 10;257(7):3589–92.
- Gupta R. Halobacterium volcanii tRNAs. Identification of 41 tRNAs covering all amino acids, and the sequences of 33 class I tRNAs. J Biol Chem. 1984 Aug 10;259(15):9461–71.
- Wurm JP, Griese M, Bahr U, Held M, Heckel A, Karas M, et al. Identification of the enzyme responsible for N1-methylation of pseudouridine 54 in archaeal tRNAs. RNA. 2012 Mar;18(3):412–20.
- Andries O, Mc Cafferty S, De Smedt SC, Weiss R, Sanders NN, Kitada T. N1-methylpseudouridine-incorporated mRNA outperforms pseudouridine-incorporated mRNA by providing enhanced protein expression and reduced immunogenicity in mammalian cell lines and mice. J Controlled Release. 2015 Nov 10;217:337–44.
- Nance KD, Meier JL. Modifications in an Emergency: The Role of N1-Methylpseudouridine in COVID-19 Vaccines. ACS Cent Sci. 2021 May 26;7(5):748–56.
- Bansal A. From rejection to the Nobel Prize: Karikó and Weissman’s pioneering work on mRNA vaccines, and the need for diversity and inclusion in translational immunology. Front Immunol [Internet]. 2023 Nov 8 [cited 2025 May 5];14. Available from: https://www.frontiersin.orghttps://www.frontiersin.org/journals/immunology/articles/10.3389/fimmu.2023.1306025/full.
- Mulroney TE, Pöyry T, Yam-Puc JC, Rust M, Harvey RF, Kalmar L, et al. N1-methylpseudouridylation of mRNA causes +1 ribosomal frameshifting. Nature. 2024 Jan;625(7993):189–94.
- Thess A, Grund S, Mui BL, Hope MJ, Baumhof P, Fotin-Mleczek M, et al. Sequence-engineered mRNA Without Chemical Nucleoside Modifications Enables an Effective Protein Therapy in Large Animals. Mol Ther J Am Soc Gene Ther. 2015 Sep;23(9):1456–64.
- Buschmann MD, Carrasco MJ, Alishetty S, Paige M, Alameh MG, Weissman D. Nanomaterial Delivery Systems for mRNA Vaccines. Vaccines. 2021 Jan;9(1):65.
- Heil F, Hemmi H, Hochrein H, Ampenberger F, Kirschning C, Akira S, et al. Species-Specific Recognition of Single-Stranded RNA via Toll-like Receptor 7 and 8. Science. 2004 Mar 5;303(5663):1526–9.
- Kremsner PG, Guerrero RAA, Arana E, Aroca Martinez GJ, Bonten MJ, Chandler R, et al. Efficacy and safety of the CVnCoV SARS-CoV-2 mRNA vaccine candidate: results from herald, a phase 2b/3, randomised, observer-blinded, placebo-controlled clinical trial in ten countries in Europe and Latin America.
- CureVac Comes Around [Internet]. [cited 2025 May 27]. Available from: https://www.science.org/content/blog-post/curevac-comes-around.
- Liu A, Wang X. The Pivotal Role of Chemical Modifications in mRNA Therapeutics. Front Cell Dev Biol [Internet]. 2022 Jul 13 [cited 2024 Oct 5];10. Available from: https://www.frontiersin.org/journals/cell-and-developmental-biology/articles/10.3389/fcell.2022.901510/full.
- Diebold SS, Massacrier C, Akira S, Paturel C, Morel Y, Reis e Sousa C. Nucleic acid agonists for Toll-like receptor 7 are defined by the presence of uridine ribonucleotides. Eur J Immunol. 2006;36(12):3256–67.
- Rubio-Casillas A, Cowley D, Raszek M, Uversky VN, Redwan EM. Review: N1-methyl-pseudouridine (m1Ψ): Friend or foe of cancer? Int J Biol Macromol. 2024 May 1;267:131427.
- Svitkin YV, Cheng YM, Chakraborty T, Presnyak V, John M, Sonenberg N. N1-methyl-pseudouridine in mRNA enhances translation through eIF2α-dependent and independent mechanisms by increasing ribosome density. Nucleic Acids Res. 2017 Jun 2;45(10):6023–36.
- Uchida S, Kataoka K, Itaka K. Screening of mRNA Chemical Modification to Maximize Protein Expression with Reduced Immunogenicity. Pharmaceutics. 2015 Sep;7(3):137–51.
- Krammer F, Palese P. Profile of Katalin Karikó and Drew Weissman: 2023 Nobel laureates in Physiology or Medicine. Proc Natl Acad Sci. 2024 Feb 27;121(9):e2400423121.
- Watson OJ, Barnsley G, Toor J, Hogan AB, Winskill P, Ghani AC. Global impact of the first year of COVID-19 vaccination: a mathematical modelling study. Lancet Infect Dis. 2022 Sep 1;22(9):1293–302.
- Allen, FW. The Biochemistry of the Nucleic Acids, Purines, and Pyrimidines. Annu Rev Biochem. 1941 Jul 1;10(Volume 10, 1941):221–44.
- Ontiveros RJ, Stoute J, Liu KF. The chemical diversity of RNA modifications. Biochem J. 2019 Apr 26;476(8):1227–45.
- Frye M, Harada BT, Behm M, He C. RNA modifications modulate gene expression during development. Science. 2018 Sep 28;361(6409):1346–9.
- Gilbert, WV. Recent developments, opportunities, and challenges in the study of mRNA pseudouridylation. RNA. 2024 May;30(5):530–6.
- Borchardt EK, Martinez NM, Gilbert WV. Regulation and Function of RNA Pseudouridylation in Human Cells. Annu Rev Genet. 2020 Nov 23;54:309–36.
- Franzmann PD, Springer N, Ludwig W, Conway De Macario E, Rohde M. A Methanogenic Archaeon from Ace Lake, Antarctica: Methanococcoides burtonii sp. nov. Syst Appl Microbiol. 1992 Dec 1;15(4):573–81.
- Fu L, Zhou T, Wang J, You L, Lu Y, Yu L, et al. NanoFe3O4 as Solid Electron Shuttles to Accelerate Acetotrophic Methanogenesis by Methanosarcina barkeri. Front Microbiol [Internet]. 2019 Mar 5 [cited 2024 Oct 12];10. Available from: https://www.frontiersin.org/journals/microbiology/articles/10.3389/fmicb.2019.00388/full.
- Toso DB, Javed MM, Czornyj E, Gunsalus RP, Zhou ZH. Discovery and Characterization of Iron Sulfide and Polyphosphate Bodies Coexisting in Archaeoglobus fulgidus Cells. Archaea. 2016;2016(1):4706532.
- Ossmer R, Mund T, Hartzell PL, Konheiser U, Kohring GW, Klein A, et al. Immunocytochemical localization of component C of the methylreductase system in Methanococcus voltae and Methanobacterium thermoautotrophicum. Proc Natl Acad Sci. 1986 Aug;83(16):5789–92.
- Mukhopadhyay B, Johnson EF, Wolfe RS. A novel pH2 control on the expression of flagella in the hyperthermophilic strictly hydrogenotrophic methanarchaeaon Methanococcus jannaschii. Proc Natl Acad Sci. 2000 Oct 10;97(21):11522–7.
- Mochimaru H, Tamaki H, Katayama T, Imachi H, Sakata S, Kamagata Y. Methanomicrobium antiquum sp. nov., a hydrogenotrophic methanogen isolated from deep sedimentary aquifers in a natural gas field. Int J Syst Evol Microbiol. 2016;66(11):4873–7.
- Legat A, Gruber C, Zangger K, Wanner G, Stan-Lotter H. Identification of polyhydroxyalkanoates in Halococcus and other haloarchaeal species. Appl Microbiol Biotechnol. 2010 Jul 1;87(3):1119–27.
- de Silva RT, Abdul-Halim MF, Pittrich DA, Brown HJ, Pohlschroder M, Duggin IG. Improved growth and morphological plasticity of Haloferax volcanii. Microbiology. 2021;167(2):001012.
- Stan-Lotter H, Fendrihan S. Halophilic Archaea: Life with Desiccation, Radiation and Oligotrophy over Geological Times. Life. 2015 Sep;5(3):1487–96.
- Kashefi K, Moskowitz BM, Lovley DR. Characterization of extracellular minerals produced during dissimilatory Fe(III) and U(VI) reduction at 100 °C by Pyrobaculum islandicum. Geobiology. 2008;6(2):147–54.
- Thermoproteus neutrophilus - microbewiki [Internet]. [cited 2024 Oct 6]. Available from: https://microbewiki.kenyon.edu/index.php/Thermoproteus_neutrophilus.
- Burggraf S, Fricke H, Neuner A, Kristjansson J, Rouvier P, Mandelco L, et al. Methanococcus igneus sp. nov., a Novel Hyperthermophilic Methanogen from a Shallow Submarine Hydrothermal System. Syst Appl Microbiol. 1990 Aug 1;13(3):263–9.
- Stetter KO, Thomm M, Winter J, Wildgruber G, Huber H, Zillig W, et al. Methanothermus fervidus, sp. nov., a novel extremely thermophilic methanogen isolated from an Icelandic hot spring. Zentralblatt Für Bakteriol Mikrobiol Hyg Abt Orig C Allg Angew Ökol Mikrobiol. 1981 Aug 1;2(2):166–78.
- Jones JB, Bowers B, Stadtman TC. Methanococcus vannielii: ultrastructure and sensitivity to detergents and antibiotics. J Bacteriol. 1977 Jun;130(3):1357–63.
- Chatterjee K, Blaby IK, Thiaville PC, Majumder M, Grosjean H, Yuan YA, et al. The archaeal COG1901/DUF358 SPOUT-methyltransferase members, together with pseudouridine synthase Pus10, catalyze the formation of 1-methylpseudouridine at position 54 of tRNA. RNA. 2012 Mar;18(3):421–33.
- Dutta N, Deb I, Sarzynska J, Lahiri A. Structural and Thermodynamic Consequences of Base Pairs Containing Pseudouridine and N1-methylpseudouridine in RNA Duplexes. ChemistrySelect. 2025;10(4):e202400006.
- Chen TH, Potapov V, Dai N, Ong JL, Roy B. N1-methyl-pseudouridine is incorporated with higher fidelity than pseudouridine in synthetic RNAs. Sci Rep. 2022 Jul 29;12(1):13017.
- Song, Y. Tweaks in vaccine. Nat Chem Biol. 2024 Feb;20(2):131–131.
- Dong Y, Anderson DG. Opportunities and Challenges in mRNA Therapeutics. Acc Chem Res. 2022 Jan 4;55(1):1–1.
- Rohner E, Yang R, Foo KS, Goedel A, Chien KR. Unlocking the promise of mRNA therapeutics. Nat Biotechnol. 2022 Nov;40(11):1586–600.
- Ball, P. The lightning-fast quest for COVID vaccines — and what it means for other diseases. Nature. 2020 Dec 18;589(7840):16–8.
Figure 1.
Timeline of events in the use of modified nucleosides in mRNA vaccines. Adapted from [5]. Timeline sources: mRNA vaccines events [5,10,11,12,13,14,15], pseudouridine [6,16,17,18], n1-methylpseudouridine [8,19,20,21,22,23,24,25,26,27].
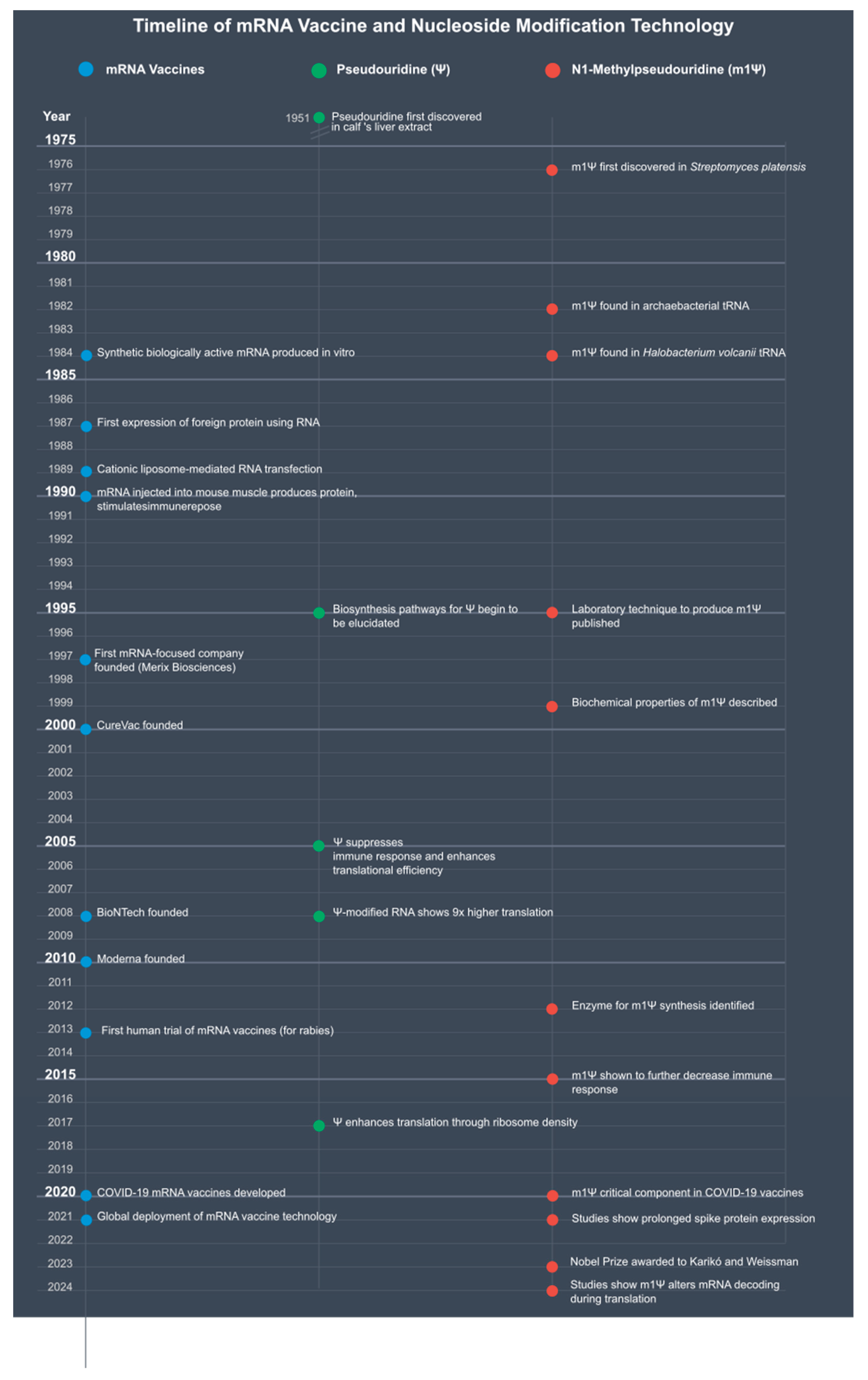
Figure 2.
Archaeal species with naturally occurring m1Ψ, and their habitats. Adapted from [59]. Images for Methanosarcina barkeri [46], Archaeoglobus fulgidus [47], Methanococcus voltae [48], Methanococcus jannaschii [49], Methanomicrobium mobile [50], Halococcus morrhuae [51], Haloferax volcanii [52], Halobacterium salinarum [53], Pyrobaculum islandicum [54] are reproduced under Creative Commons licenses, from their respective references. The image for Thermoproteus neutrophilus [55] is reproduced with permission from Prof. Lennon. Image for Methanococcoides burtonii is reprinted from A Methanogenic Archaeon from Ace Lake, Antarctica: Methanococcoides burtonii sp. nov.," P.D. Franzmann, N. Springer, W. Ludwig, E. Conway De Macario, M. Rohde, Systematic and Applied Microbiology, Vol. 15, Issue 4, pp. 573–581, 1992. [45], with permission from Elsevier. Image for Methanococcus igneus is reprinted from Systematic and Applied Microbiology, Vol. 13, Siegfried Burggraf, Hans Fricke, Annemarie Neuner, Jakob Kristjansson, Pierre Rouvier, Linda Mandelco, Carl R. Woese, Karl O. Stetter, Methanococcus igneus sp. nov., a Novel Hyperthermophilic Methanogen from a Shallow Submarine Hydrothermal System, pp. 263-269. Reference [56], Copyright (1990), with permission from Elsevier. The images of Methanothermus fervidus [57] and Methanococcus vannielii [58] are reproduced from their associated references, with permission from Elsevier.
Figure 2.
Archaeal species with naturally occurring m1Ψ, and their habitats. Adapted from [59]. Images for Methanosarcina barkeri [46], Archaeoglobus fulgidus [47], Methanococcus voltae [48], Methanococcus jannaschii [49], Methanomicrobium mobile [50], Halococcus morrhuae [51], Haloferax volcanii [52], Halobacterium salinarum [53], Pyrobaculum islandicum [54] are reproduced under Creative Commons licenses, from their respective references. The image for Thermoproteus neutrophilus [55] is reproduced with permission from Prof. Lennon. Image for Methanococcoides burtonii is reprinted from A Methanogenic Archaeon from Ace Lake, Antarctica: Methanococcoides burtonii sp. nov.," P.D. Franzmann, N. Springer, W. Ludwig, E. Conway De Macario, M. Rohde, Systematic and Applied Microbiology, Vol. 15, Issue 4, pp. 573–581, 1992. [45], with permission from Elsevier. Image for Methanococcus igneus is reprinted from Systematic and Applied Microbiology, Vol. 13, Siegfried Burggraf, Hans Fricke, Annemarie Neuner, Jakob Kristjansson, Pierre Rouvier, Linda Mandelco, Carl R. Woese, Karl O. Stetter, Methanococcus igneus sp. nov., a Novel Hyperthermophilic Methanogen from a Shallow Submarine Hydrothermal System, pp. 263-269. Reference [56], Copyright (1990), with permission from Elsevier. The images of Methanothermus fervidus [57] and Methanococcus vannielii [58] are reproduced from their associated references, with permission from Elsevier.
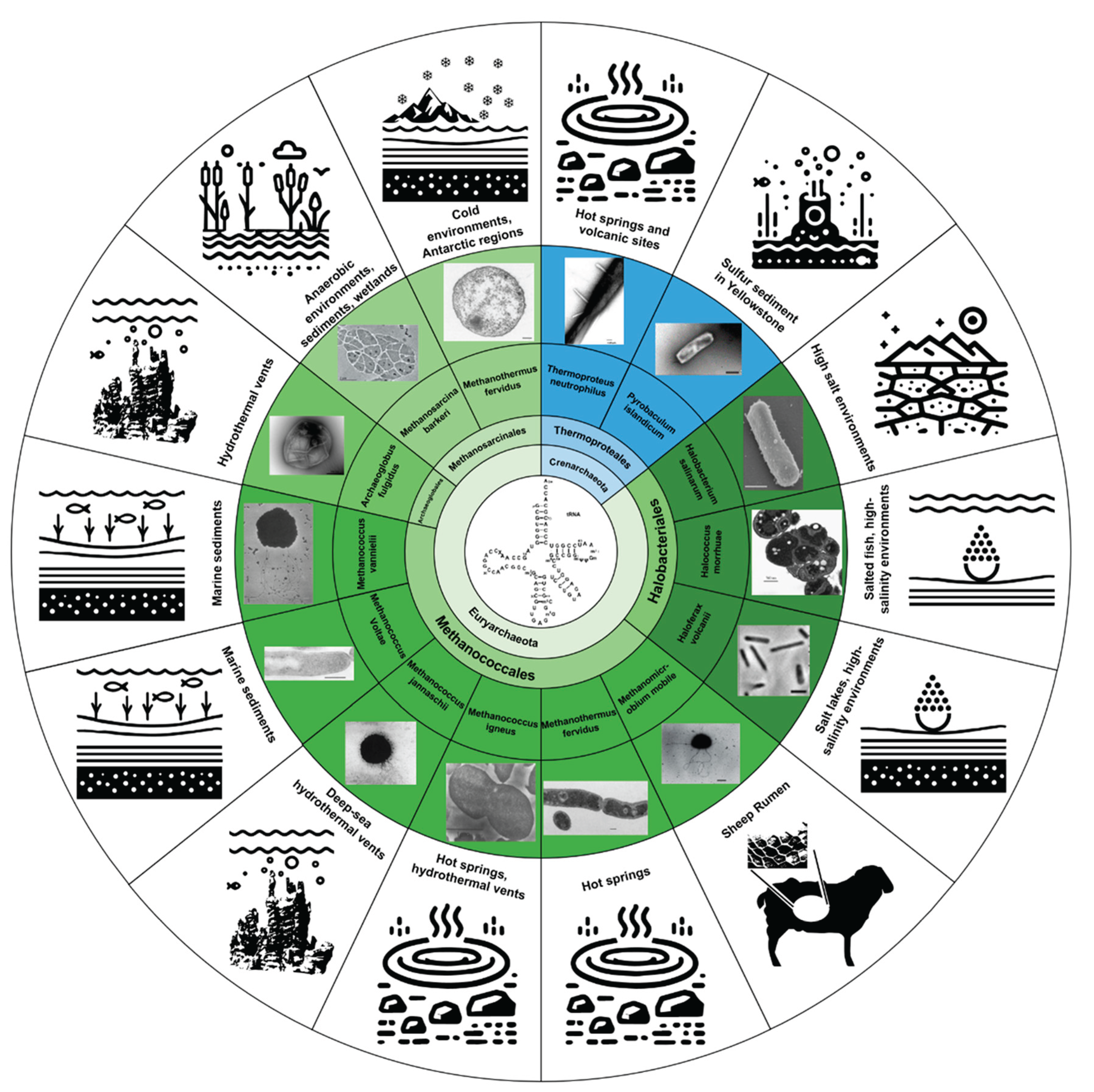
Figure 3.
Two biological pathways where n1-methyl-pseudouridine is present. Panel A. tRNA modifications at position 54 in the conserved TΨ-loop of tRNA. The modification pathway varies by organism type, in most archaea, U54 is first converted to pseudouridine (Ψ54) by the enzyme aPus10, then can be further methylated to m1Ψ54 by the methyltransferase TrmY1. In Thermococcales, Bacteria, and Eukarya, U54 is directly methylated to m5U54 by different methyltransferases (aTrmU54, bTrmA, and eTrm2p respectively). Panel B. Modifications in the 16S rRNA helix 35 region. In Archaea U914 is converted to pseudouridine by an unidentified archaeal enzyme system. In Eukarya the corresponding U1191 in 18S rRNA is modified by a snoRNP complex containing the eCbf5 pseudouridine synthase and snR35 guide RNA. The pseudouridine can be methylated to m1Ψ by the Nep1 methyltransferase, and in some eukaryotes, further modified to acp3m1Ψ. The right side of the figure displays the chemical structures of these various uridine derivatives, with dashed lines indicating the axis of rotation during isomerization and asterisks marking atoms involved in reverse Hoogsteen base pairing.Reproduced from [59] under a Creative Commons License (Attribution-NonCommercial 4.0 International License).
Figure 3.
Two biological pathways where n1-methyl-pseudouridine is present. Panel A. tRNA modifications at position 54 in the conserved TΨ-loop of tRNA. The modification pathway varies by organism type, in most archaea, U54 is first converted to pseudouridine (Ψ54) by the enzyme aPus10, then can be further methylated to m1Ψ54 by the methyltransferase TrmY1. In Thermococcales, Bacteria, and Eukarya, U54 is directly methylated to m5U54 by different methyltransferases (aTrmU54, bTrmA, and eTrm2p respectively). Panel B. Modifications in the 16S rRNA helix 35 region. In Archaea U914 is converted to pseudouridine by an unidentified archaeal enzyme system. In Eukarya the corresponding U1191 in 18S rRNA is modified by a snoRNP complex containing the eCbf5 pseudouridine synthase and snR35 guide RNA. The pseudouridine can be methylated to m1Ψ by the Nep1 methyltransferase, and in some eukaryotes, further modified to acp3m1Ψ. The right side of the figure displays the chemical structures of these various uridine derivatives, with dashed lines indicating the axis of rotation during isomerization and asterisks marking atoms involved in reverse Hoogsteen base pairing.Reproduced from [59] under a Creative Commons License (Attribution-NonCommercial 4.0 International License).
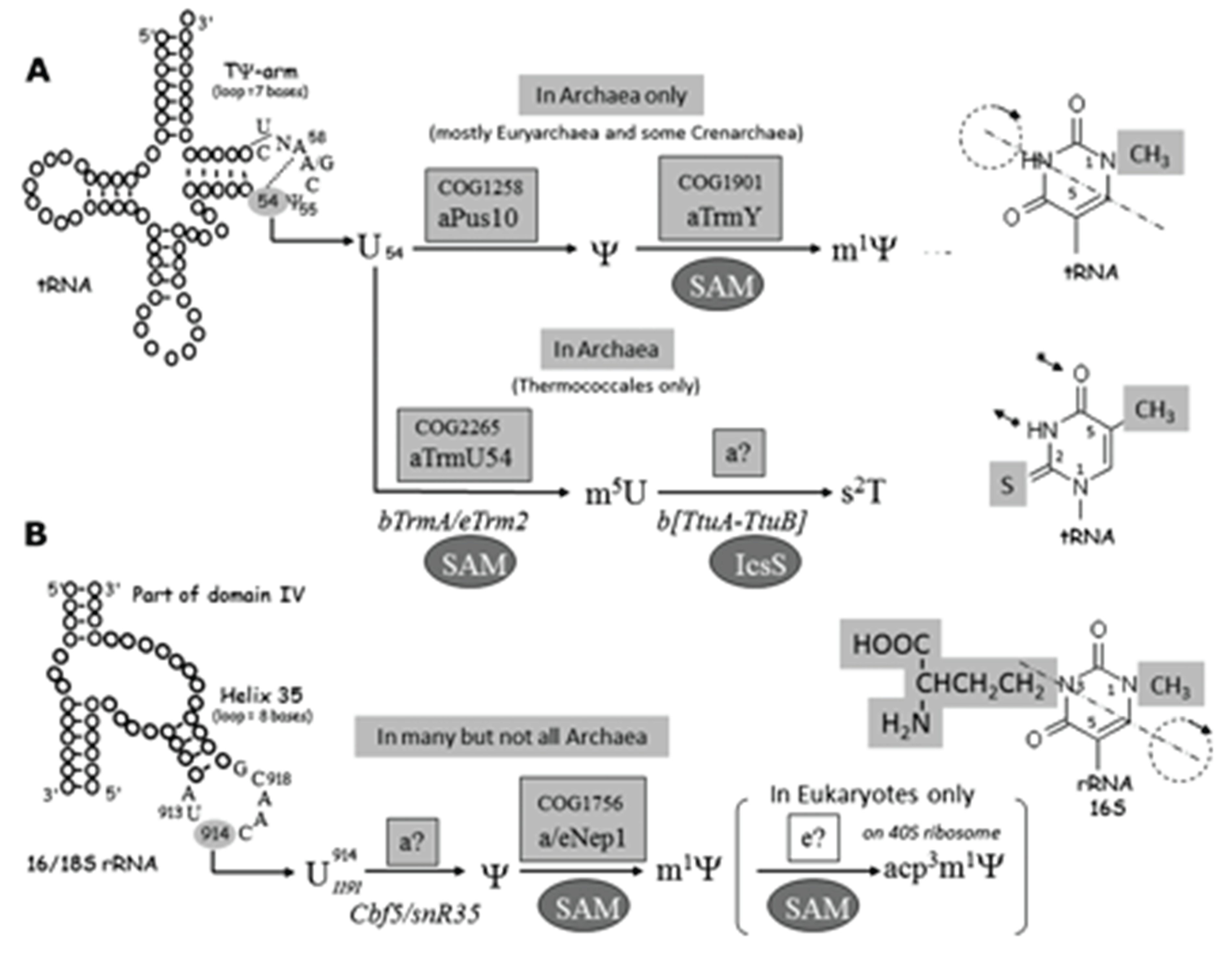
Figure 4.
Translational challenges with modified nucleosides. Panel A. Translational error rates (misuse of tRNAs) of modified uridine-containing transcripts. Reproduced from [61] under a Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/). Panel B. Out of frame expression of a firefly luciferase reporter by nucleoside type. m1Ψ, N1-methyl-pseudouridine; m5C, 5-methylcytidine; m1Ψ + m5C, combination of N1-methyl-pseudouridine and 5-methylcytidine; mo5U, 5-methoxyuridine; mo5U + m5C, combination of 5-methoxyuridine and 5-methylcytidine. Reproduced from (23) under a Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/).
Figure 4.
Translational challenges with modified nucleosides. Panel A. Translational error rates (misuse of tRNAs) of modified uridine-containing transcripts. Reproduced from [61] under a Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/). Panel B. Out of frame expression of a firefly luciferase reporter by nucleoside type. m1Ψ, N1-methyl-pseudouridine; m5C, 5-methylcytidine; m1Ψ + m5C, combination of N1-methyl-pseudouridine and 5-methylcytidine; mo5U, 5-methoxyuridine; mo5U + m5C, combination of 5-methoxyuridine and 5-methylcytidine. Reproduced from (23) under a Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



