
Submitted:
27 June 2025
Posted:
01 July 2025
You are already at the latest version
Abstract
Oxidative stress is a biological imbalance that contributes to cellular damage and is a major driver of aging and age-related disorders, prompting the search for natural antioxidant agents. Our study is a phytochemical, electrochemical and biological characterization of antioxidant potential of aqueous extracts from aerial parts of A. occidentale—leaves, bark, fruit, and cashew nuts- traditionally used in folklore medicine. Extracts were analyzed using FT-IR spectroscopy, GC×GC-TOFMS, polyphenol quantification and antioxidant capacity assays (ABTS, FRAP, DPPH). Biological activity was tested in different mice and human cell lines (SH-SY5Y, MEF, ARPE-19 and HLEC). Aqueous extracts from the leaves and bark of A. occidentale exhibited significantly higher antioxidant activity compared to those from the fruit and cashew nut. These extracts showed elevated polyphenol content and strong performance in antioxidant capacity assays. In vitro, leaf and bark extracts enhanced cell viability under H₂O₂-induced oxidative stress, preserved mitochondrial membrane potential and upregulated cytoprotective genes (HMOX1, NQO1, GCLC, and GCLM) in multiple cell lines. In contrast, fruit and nut extracts showed minimal antioxidant activity and no significant gene modulation. Our findings underscore the therapeutic potential of A. occidentale leaf and bark extracts as effective natural antioxidants and support their further development as candidates for phytotherapeutic interventions.
Keywords:
phytocompounds
; Anacardium
; oxidative stress
; antioxidants
; NRF2
1. Introduction
Oxidative stress plays a pivotal role in human pathology, being critical in the onset and progression of multiple diseases, including ocular and neurodegenerative diseases. High levels of oxidative stress are particularly relevant during aging when a progressive age-related functional decline of the antioxidant shielding network is believed to occur, leading to the aberrant accumulation of reactive oxygen species (ROS) [1,2,3]. This is in alignment with the association of oxidative stress to the etiology and progression of multiple age-related diseases, including Alzheimer’s disease, Parkinson’s disease, age-related macular degeneration, cataracts or glaucoma among others [4,5]. Nowadays, the need to identify therapeutic strategies to mitigate the effects of oxidative stress in aging and age-related disorders represents one of the major challenges the scientific community faces.
Oxidative stress refers to the imbalance between the production of ROS and the ability of an organism to detoxify them through antioxidant cellular capacities. The production of free radicals in an organism is a naturally occurring phenomenon due to cellular metabolism and external factors such as xenobiotics, pollutants, UV radiation, and heavy metals can stimulate the production of harmful oxidative molecules [6]. Optimal redox status is essential for the maintenance of proper cellular function and, when the production of ROS overwhelms the antioxidant defense, gradual damage is accumulated in cells and tissues. A better understanding of the modulation of the redox status imbalance is imperative for developing antioxidant-based therapies aimed at mitigating disease progression and improving patient outcomes.
Our body possesses both enzyme-based and non-enzymatic mechanisms that contribute to its total antioxidant capacity, which counteract the toxic effects of free radicals. In the antioxidant defense, the nuclear factor erythroid 2-related factor 2 (NRF2) is the best-established master regulator of oxidative stress response, a transcriptional factor modulating the expression level of genes encoding antioxidant and detoxifying enzymes to neutralize the oxidative stress-derived cellular damage [7,8,9]. Some of these cytoprotective genes are involved in glutathione metabolism, such as glutamate-cysteine ligase catalytic subunit (GCLC) and glutamate-cysteine ligase modifier subunit (GCLM), in heme metabolism, such as heme oxygenase 1 (HMOX1), or in detoxifying routes such as NAD(P)H: quinone oxidoreductase-1 (NQO1). Of note, loss of NRF2 activity is considered a pivotal driving force behind the imbalance of redox status and age-related proteotoxicity associated with multiple ageing-related diseases [10].
In recent years, there has been growing interest in identifying compounds derived from plants -also known as phytocompounds- that can exert antioxidant properties, acting as either ROS scavengers or boosting the transcriptional upregulation of NRF2-target genes. These phytocompounds are often secondary metabolites, non-essential molecules produced by plants that confer the ability to respond to stimuli, adapt to stress, and provide protection against oxidative stress. Glycosides, alkaloids, terpenoids and polyphenols are some of these metabolites, and the latter are now being deeply investigated for their promising pharmacological properties [11,12,13].
In this regard, ethnobotany becomes especially important for gathering all information related to the properties, therapeutic potential, and use of plants by societies and communities. Medicinal plants constitute one of the most widely utilized forms of traditional medicine. For many communities, herbal remedies often serve as a primary—or even sole—means of accessing healthcare. The therapeutic properties of many plants have not been thoroughly researched and analyzed from a scientific perspective. The combination of ethnobotanical knowledge and omics technologies has already been proposed as a promising method to identify new phytocompounds or evaluate the potential of different plant extracts [14].
Growing evidence suggests that the family Anacardiaceae, which comprises approximately 81 genera and around 800 species, might be a promising source of bioactive phytochemicals and nutraceuticals [15]. In most cases, these are tropical plant species, typically exhibiting arboreal or shrubby growth forms, and different members of the family have garnered the scientific community's attention, such as Pistacia vera, Mangifera indica, or Anacardium occidentale. The latter is a tree about 5 to 7 meters tall, native to South America, specifically the northwest of Brazil. However, due to the agri-food interest in the cashew nut, A. occidentale is now cultivated in other tropical regions such as Africa, Asia, and India. Apart from the cashew nuts, a product well-known worldwide as a food snack, the A. occidentale fruit, known as the cajú apple, is often consumed as juice, jam, or simply as a fresh fruit when mature. Additionally, leaves and bark are used in folklore medicine to treat different ailments, and some studies suggest that these two aerial parts seem to possess certain antidiabetic and antihyperglycemic activities [16,17].
Despite the pharmaceutical potential of A. occidentale being explored, there is fragmented knowledge about the antioxidant potential of the different aerial parts. This work develops a multidisciplinary approach to analyze the antioxidant properties of the aqueous extract of four aerial parts of the plant (leaves, bark, fruit, and cashew nuts) and their protective effects against H2O2-derived toxicity.
2. Materials and Methods
2.1. Preparation of Plant Extracts
The aerial parts of A. occidentale (leaves, bark, fruit and cashew nut) were harvested from a local crop field (16°07'29.1"N 61°34'53.9"W) in Goyave (Guadeloupe (FWI)). 374 g of dried leaves were introduced into a microwave extractor (ETHOS X advanced extraction system (Milestone®, Bergamo, Italy)) chamber containing 1.5 L of distilled water. After the extraction, the residue was lyophilized using a freeze dryer model Alpha 1-4 LD plus (Martin Christ®, Germany). After lyophilization, 11.98 g of dried leaves extract was obtained, corresponding to a 3.2% yield. For the bark extract, 1009.2 g of A. occidentale bark was introduced into a microwave extractor chamber containing 1.5 L of distilled water. After the extraction, the residue was lyophilized. After the lyophilization, 41.29g of dried bark extract were obtained, corresponding to a 4.09% yield. The starting material for the obtention of the aqueous A. occidentale extract was 125.21g of lyophilized fruits. The material was smashed and introduced into a microwave extractor chamber containing 1.5 L of distilled water. Analogous to the bark aqueous extract, a microwave-assisted extraction was performed. The remaining material underwent lyophilization. After the lyophilization process, 61.33g of dried fruit extract were obtained, corresponding to a 48.98% yield. 1 Kg of dried A. occidentale nuts were introduced into a microwave extractor chamber containing 1.5 L of distilled water. A microwave-assisted aqueous extraction was performed. After the extraction, the residue was lyophilized. After the lyophilization, 94.86g of dried nuts extract were obtained, corresponding to a 9.5% yield.
2.2. Metabolomic Profile
2.2.1. Fourier Transform Infrared Spectroscopy (FT-IR)
Fourier Transform Infrared Spectroscopy (FT-IR) was used for preliminary phytochemical characterization of plant extracts. This technique identifies functional groups and molecular structures based on vibrational transitions, providing insights into the chemical composition of complex mixtures. Samples for FT-IR analysis were prepared by dissolving four distinct plant extracts in distilled water to achieve a uniform concentration of 1 mg/mL, ensuring optimal signal intensity while minimizing saturation effects. Spectral data acquisition was performed with the Spectrum Two FT-IR spectrometer (Perkin Elmer, Madrid, Spain), equipped with an Attenuated Total Reflection (ATR) accessory for analyzing liquid and semi-solid samples, allowing direct measurement without extensive preparation. Each extract was analyzed separately, and spectra were recorded over the mid-infrared range (4000–400 cm⁻¹). After acquisition, raw spectra underwent baseline correction and normalization using Spectrum 10™ software (Perkin Elmer), which is essential for minimizing noise and enhancing peak identification accuracy. Characteristic water absorption bands were computationally removed to prevent interference with phytochemical signal interpretation. Key absorption bands were identified and assigned to specific functional groups like hydroxyl (-OH), carbonyl (C=O), alkene (C=C) and amine (N-H) groups, indicative of phytochemical classes such as flavonoids, alkaloids, terpenoids, and phenolic compounds.
2.2.2. Determination of the Total Polyphenol Content (TPC)
The total polyphenol content (TPC) of the four extracts was determined by the Folin-Ciocalteu method. Briefly, 1mg/mL solutions of each extract were prepared by dissolving the dried extract in distilled water. The solutions were filtered to avoid the presence of undesired particles. The reaction was carried out by mixing 100µL of each 1mg/mL aqueous extract, 600µL of distilled water, 600µL of sodium carbonate 7.5%, and 200µL of the Folin-Ciocalteu reagent. The mixture was incubated for 10 minutes at 50 °C. The absorbance was measured at a wavelength of 760 nm using the spectrophotometer Genesys 20 (Thermo Scientific). For the analysis, a standard curve using gallic acid 2.5 mM was used, and the results were expressed as gallic acid equivalents (GAE). Three experiments were conducted in triplicate to ensure the reliability and reproducibility of the results.
2.2.3. GC×GC-TOFMS Analysis
2.2.3.1. Derivatization Analysis
Samples were prepared and analyzed as previously described [18] with minor modifications. Analyte extracted in 1 mL 50:50 methanol (Optima Grade, Fisher Scientific, Hampton, NH, USA) and chloroform (HPLC-grade, Fisher Scientific) in a 2 mL microcentrifuge tube (VWR, Radnor, PA, USA). The extraction was vortexed for 5 min, then centrifuged at 10.000 rpm for 10 min (MIKRO 185, Hettich Zentrifugen, Westphalia, Germany). The supernatant was then removed and diluted by a factor of 40, then 750 μL of the diluent was aliquoted into a 2 mL clear glass GC vial (Chromatographic Specialties) and capped. The extract was then dried under nitrogen at 40 °C. A 100 μL aliquot of toluene dried with Na₂SO₄ was added to the sample and dried under nitrogen at 50 °C. A 50 μL aliquot of 20 mg/mL of CH3ONH2 · HCl (Fisher Scientific) in pyridine (HPLC Grade, Fisher Scientific) was added to the vial and incubated in the heating block for 1 h at 60 °C. Following this, a 75 μL aliquot of MSTFA + 1% TMCS (Fisher Scientific) was added to the vial, and the sample was again incubated for 1 h at 40 °C. The sample was then transferred into a 300 μL glass insert autosampler vial (Chromatographic Specialties) and capped.
2.2.3.2. Untargeted GC×GC-TOFMS
Untargeted GC×GC-TOFMS metabolomics was performed using a method reported previously [19]. A total volume of 1 µL of each sample was injected using a MultiPurpose Sampler (MPS; Gerstel, Linthicum Heights, MD, USA) into a CIS-4 inlet (Gerstel) operated in splitless inlet mode at 250 °C. Separation and analysis were performed with a LECO BT 4D GC×GC-TOFMS (LECO, St. Joseph, MI, USA). The first-dimension chromatographic column was a 60 m × 0.25 mm × 0.25 μm Rxi-5SilMS (Chromatographic Specialities, Brockville, ON, Canada), and the second-dimension column was a 1.4 m × 0.25 mm × 0.25 μm Rtx-200MS (Chromatographic Specialities). Ultra-pure helium gas (Linde Canada Inc., Edmonton, AB, Canada) was used as the carrier gas at a constant flow rate of 2.0 mL/min. The initial oven temperature was held at 80 °C for 4 min before ramping up to a final temperature of 315 °C at a rate of 3.5 ℃/min, where it was held for 10 min. The secondary oven and modulator temperature offsets were set at +10 °C relative to the GC oven and +15 °C relative to the secondary oven, respectively. The modulation period was 2.5 s. Data acquisition was conducted at a rate of 200 Hz over a mass range between 40 and 800 m/z. A detector voltage was applied, and an electron impact energy of −70 eV was set for ionization. The mass spectrometer transfer line and ion source temperatures were set to 250 °C and 200 °C, respectively. The total chromatographic analysis time was 81.1 min for each run. A reagent blank and instrument blanks were performed for quality control purposes.
2.2.3.3. Data Processing
The GC×GC-TOFMS data were processed using LECO ChromaTOF® for BT (v5.58.05; Leco). Peak finding was performed with an S/N above 1000 and a minimum stick count of 3. Subpeaks were combined with a mass spectral match factor over 500. Peaks eluting within 0.8 s to 1.1 s in the second dimension were removed, as they elute in the column bleed. Retention indices for all detected peaks were calculated based on the retention time of linear alkanes (C7-C30) analyzed on the same day. Library matching for putative compound identification was performed against commercially available and in-house databases. A minimum mass spectral similarity of 800 and a retention index within ± 30 units was required to assign a putative identity. The compounds trans-catechin, gallic acid, protocatechoic acid and pyrogallol were putatively identified by using a Target Analyte Search in ChromaTOF® with a S/N of 750, a minimum area of 100, and a minimum height of 25. All analytes were putatively identified according to level 2 of the metabolomics standards initiative [20]. Analytes of interest were normalized to the total peak area of all detected peaks in those samples [21].
2.3. Electrochemical Analysis
2.3.1. Cyclic Voltammogram
Electrochemical experiments were conducted as previously published [22,23]. The solutions of different extracts were previously prepared by dissolving 100 mg of the target extract in 10 mL of 10 mM CuSO4 solution. Cyclic voltammograms were recorded at room temperature in a CH I660 equipment (BioLogic Science Instruments, France) working with a conventional three-electrode cell using a platinum wire auxiliary electrode, an Ag/AgCl reference electrode, and a glassy carbon electrode (GCE) as working electrode. The study of the cyclic voltammogram signals corresponding to the cathodic region attributed to the reduction of Cu2+ to Cu0 is observed at −0.65 V versus Ag/AgCl. As a result of the reduction, a deposit of Cu0 takes place in the GCE (stripping). In the subsequent anodic scan, the deposit of solid Cu0 is oxidized to Cu 2+, producing the stripping signal corresponding to the oxidation peak at +0.20 V. The study of the modification of both signals indicates the chelation of copper exerted by the different extracts.
2.3.2. Electrochemical Antioxidant Activity Determination
The total antioxidant capacity was measured using the BRS device following the manufacturer’s instructions (BQC Redox Technologies, Oviedo, Asturias, Spain). This portable instrument enables the assessment of the total antioxidant capacity (TAC). Disposable strips interacting with the sample generate an electrochemical signal proportional to the redox activity [24]. Measurements were expressed as BRS values in microcoulombs (µC), and experiments were performed in triplicate.
2.4. Antioxidant Analysis
2.4.1. ABTS Assay
As a initial approach the assay was performed at a concentration of 1mg/ml for each aqueous extract using the ABTS Assay Kit (Bioquochem, Asturias, Spain) according to the manufacturer’s instructions using the spectrophotometer Genesys 20 (Thermo Scientific). Trolox was used as a standard, and data were shown as Trolox Equivalents Antioxidant Capacity (TEAC) (µM Trolox). The ABTS+ radical scavenging capacity was calculated as follows:
Where is the blank-corrected absorbance of standard 1 (0 µM Trolox) and is the blank-corrected absorbance measured for each sample.
For a more in-depth study, the 2,2'-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid) (ABTS) assay was performed on each A. occidentale aqueous extract as previously published [25]. First, different concentrations of each aqueous extract were tested to assess the IC50 value. The ABTS+ radical was produced by reacting 2,2’-azino-bis(3-ethylbenzthiazoline-6-sulphonic acid (ABTS) 7mM with K2S2O8 2.45mM (both reagents dissolved in water) at a volume ratio of 1:1. After mixing, the reaction was incubated in the dark at room temperature for 16 hours. The radical was diluted in ethanol to reach a final absorbance of 0.750 ± 0.025 at 734 nm. For this experiment, different concentrations of the aqueous extracts were tested: 0.0001, 0.001, 0.01, 0.1, 1, 10, 100, 1000 and 10000 µg/mL in a 96-well plate. 150 µL of each solution was added to the plate, followed by 50 µL of the ABTS⁺ radical. The plate was then incubated for 30 minutes at room temperature. After incubation, the absorbance was measured at a wavelength of 734 nm using a Varioskan LUX plate reader (Thermo Fisher Scientific, Vantaa, Finland)
2.4.2. FRAP Assay
A initial assay was performed at a concentration of 1 mg/mL for each aqueous extract, using the FRAP Assay Kit (Bioquochem, Asturias, Spain), according to the manufacturer's instructions using the spectrophotometer Genesys 20 (Thermo Scientific). A more detailed study was performed, in which the ferric reducing antioxidant power (FRAP) assay of four different extracts of aerial parts of A. occidentale was evaluated, as previously described in the literature [25]. The FRAP reagent was prepared by mixing 10 mM 2,4,6-tripyridyl-S-triazine in 40 mM HCl, 20 mM FeCl₃·6H₂O and 300 mM acetate buffer, all dissolved in Milli-Q water, in a proportion of 1:10:10. Briefly, 180 µL of the FRAP reagent was added to each well of a 96-well plate, followed by the addition of 20 µL of each aqueous extract solution. Concentrations of 0.0001, 0.001, 0.01, 0.1, 1, 10, 100, 1000 and 10000 µg/ml of the aqueous extract were tested. After a 30-minute incubation period in the dark, the absorbance at 593 nm was measured using a Varioskan LUX plate reader (Thermo Fisher Scientific, Vantaa, Finland).
2.4.3. DPPH Assay
The assay was initially performed at a concentration of 1 mg/mL for each aqueous extract using the DPPH Assay Kit (Bioquochem, Asturias, Spain) according to the manufacturer’s instructions using the spectrophotometer Genesys 20 (Thermo Scientific). . Trolox was used as a standard, and data are shown as Trolox Equivalents Antioxidant Capacity (TEAC) (µM Trolox). The DPPH radical scavenging capacity was calculated as follows:
Where is the blank-corrected absorbance of standard 1 (0 µM Trolox) and is the blank-corrected absorbance measured for each sample.
A more detailed study was conducted in which 2, 2-diphenyl-1-picrylhydrazyl (DPPH) assay was performed for each A. occidentale aqueous extract as published [25]. The DPPH radical was generated by preparing a DPPH 0.2 mM solution in absolute ethanol. Different concentrations of the aqueous extract were tested: (0.0001, 0.001, 0.01, 0.1, 1, 10, 100, 1000, 10000) µg/mL in a 96-well plate. 150 µL of each aqueous extract solution was added to the plate, followed by the addition of 50µL of the DPPH radical. The plate was then incubated for 30 minutes at room temperature. Absorbance was measured at a wavelength of 517nm using the Varioskan LUX plate reader (Thermo Fisher Scientific, Vantaa, Finland).
2.5. Cell Culture
Human lens epithelial cells (HLEC, line SRA 01/04) were kindly donated by Dr. Venkat Reddy, (Kellogg Eye Center, University of Michigan). Mouse embryonic fibroblasts (MEFs) from mice WT were generously donated by Dr. Masaaki Komatsu (Juntendo University, Japan). Human retinal pigment epithelial cells (ARPE19) and SH-SY5Y human neuroblastoma cells were purchased from the American Type Culture Collection (ATCC). These three cell lines were cultured in Dulbecco's Modified Eagle Medium (DMEM) (GIBCO) containing 10% fetal bovine serum (FBS), 1% penicillin-streptomycin mix (10.000 U/ml), 1% MEM non-essential amino acids (GIBCO) and 1% sodium pyruvate 100mM (GIBCO) in an incubator at 37°C in a humid atmosphere containing 5% CO2.
2.6. Analysis of Cytotoxicity and Cell Viability Against Oxidative Stress
2.6.1. Cytotoxicity Activity
A density of 5 × 10³ cells per well was used to seed the SH-SY5Y cells in a 96-well plate, which was then grown in the incubator for 24 hours. The cells were exposed to different concentrations (ranging from 0.01 to 1,000 µg/mL) of an aqueous leaf and bark extract, which had been resuspended in 50 mM PBS for 24 hours. Following this pretreatment with the plant extracts, the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT) assay was conducted to assess cell proliferation and viability. 5 μL of 20 mg/mL of MTT was added to the wells, and the plate was incubated at 37 °C for 2 h; after this, 200 μL of dimethylsulfoxide was added to solubilize the formazan crystals, and the plate was left in the dark at room temperature for 30 minutes. Absorbance was measured at 570 nm using a Varioskan LUX plate reader (Thermo Fisher Scientific, Vantaa, Finland). Dose-response curves were plotted using linear regression analysis and LD50 (inhibition of cell viability) concentrations were calculated.
2.6.2. Neuroprotection Assay Against H2O2-Induced Cytotoxicity
As in the previous experiment, 5 × 10³ SH-SY5Y cells/well were seeded in 96-well plates and grown in the incubator for 24 hours. Cells were pretreated with cell media or 5, 10, 20, 50, 100, and 200 µg/mL of the PBS 1X-resuspended aqueous bark and leaves extract. After 24 hours, except for control wells, cells were exposed for four hours to H202 at a concentration of 500 µM. The MTT viability assay was performed according to the conditions described above. Cell viability values were expressed as a percentage in comparison to the viability of the untreated cells.
2.6.3. Determination of the Levels of Intracellular ROS
Dichlorofluorescein diacetate (DCFH2-DA) was used to determine intracellular ROS levels as previously reported [25]. Intracellular esterases cleave DCFH2-DA to form the fluorescent compound DCF. The fluorescent intensity is proportional to the intracellular ROS level. 5x10⁵ SH-SY5Y cells were seeded in 96-well culture black, clear-bottom plates and, after 24 hours, were treated with the test extract for additional 24 hours. Then, the cells were loaded with 10 µM DCFH₂DA in FBS-free culture medium for 50 minutes in the dark at 37 °C in a CO₂ incubator. Finally, the cells were exposed to 1 mM H₂O₂ for four hours. After the incubation period, the cells were twice washed with Hank's buffer. Fluorescence was measured using a Varioskan LUX microplate reader (Thermo Fisher Scientific, Vantaa, Finland) with excitation at 485 nm and emission at 535 nm wavelengths. Results are expressed as a percentage of the fluorescent increase compared to the control. All experiments were performed at least four times in sextuplicate for each extract concentration.
2.6.4. Determination of the Mitochondrial Membrane Potential
96-well plates were seeded with 5 × 10³ SH-SY5Y cells per well and incubated for 24 hours. The cells were pretreated with different concentrations (5, 10, 25, 50, 100 and 200 µg/mL) of the aqueous leaf and bark extract for 24 hours. Following the treatment with plant extracts, cells were treated with 500 µM H2O2 for 4 h, after which they were washed twice and incubated for 20 minutes with 5 µM JC-1 in culture media in the dark. The plates were then washed three times with Hank's buffer and left in 200 µL of fresh buffer. The absorbance of each well was measured at excitation wavelengths of 535 nm and emission wavelengths of 595 nm, as well as at emission wavelengths of 485 nm and 535 nm, to evaluate the presence of JC-1 aggregates and assess red and green color, respectively.
Flow cytometry studies were also conducted to evaluate the mitochondrial membrane potential. 6-well plates were seeded with 5 × 10⁵ SH-SY5Y cells per well. After 24 hours, the cells were treated with 100 or 200 µg/mL of aqueous extract from the leaves or bark and incubated for a further 24 hours. Then, cells were treated with 400 µM H₂O₂ for 4 h, after which they were washed twice with Hank's buffer and incubated for 20 minutes in the dark with 5 µM JC-1 in culture media. After staining, the cells were washed three times with Hank's buffer, resuspended in 500 µL of culture medium and analyzed using flow cytometry with an Accuri C6 flow cytometer (BD Biosciences, Ann Arbor, Michigan, USA).
2.7. Quantitative Real-Time PCR Assays
100 mm TC-treated culture dishes were seeded with either HLECs, MEF or ARPE19 cells. Cells were left growing until they reached a confluence of 80%. Then, the cells were washed with PBS 1X, and fresh media containing a concentration of 1mg/ml of the extract of leaves, bark, fruit, or cashew nut of A. occidentale was added. Cells were incubated at 37°C for 18h with each plant extract or mock solution. Then, the cells were washed once with cold PBS 1X, scraped in the same buffer and the cell pellets obtained by low-speed centrifugation were stored at -80°C until processing. Total RNA was isolated according to the manufacturer’s instructions. 1 μg of total RNA of each sample was transcribed using the NZY Reverse Transcriptase kit (NZYTech, Lisboa, Portugal). Quantitative real-time PCR (qRT-PCR) was employed to assess mRNA expression levels of the following genes: HMOX1, NQO1, GCLC and GCLM. The primers used for each gene are shown in Table 1. qPCR experiments were performed employing QuantStudio™ 5 Real-Time PCR System. The relative expression of the gene GAPDH was used as housekeeping to normalize target expression vs sample condition.
2.8. Statistical Analysis
All numerical results are reported as the mean ±SEM from a minimum of three independent experiments. In all instances “n” refers to individual experiments, indicated in the corresponding figure legends. GraphPad InStat software (GraphPad) was used for analysis of statistical significance. Two-tailed Student's t test was used to evaluate single comparisons between different experimental groups. Differences were considered statistically significant for a value of p < 0.05 and denoted by an asterisk in the graph. Comparisons were made between control and treated groups or the entire intragroup using one-way ANOVA or two-way ANOVA and Dunnett’s multiple comparisons test using GraphPad Prism 10.0 (GraphPad-Software, La Jolla, CA, USA).
3. Results
3.1. Metabolomics Revealed Differential Polyphenolic Content in the Extracts of the Aerial Parts of A. occidentale
The FT-IR analysis showed meaningful differences between the spectra of the tested materials. The relative intensities differed, and each spectrum provided essential information about specific functional groups ( Suppl. Figure S1). The spectra of the four extracts were integrated to generate a composite FT-IR spectrum, which represents the plant's phytochemical profile, capturing both shared and unique functional groups (Figure 1A). The leaf extract presented a modest signal in the region of ~1600 cm⁻¹, suggesting the presence of aromatic C=C or any carbonyl groups (e.g. phenolic acids). In the case of the bark spectrum, a prominent peak around the region of ~1600 cm⁻¹ was detected, which may indicate the presence of carbonyl groups (C=O) or aromatic C=C. Additionally, the spectral complexity in the region 1000-1300 cm⁻¹ might be potentially attributable to lignin, a characteristic component of plant bark. The spectrum corresponding to the fruit sample showed a flat signal profile up to 1300 cm⁻¹ area and the signal increased around 500-800 cm⁻¹. This event can be associated with the presence of sugars, polysaccharides, or organic acids, which are classic components of fruits. The lack of signal in the 1600 cm¹ region of the spectrum suggests the absence of conjugated carbon-carbon or phenolic compounds. The spectrum corresponding to the nut extract showed a low profile in the 1600 cm⁻¹ region. Still, it contained structures in the 1000–1300 cm⁻¹ region, indicating that the sample could be rich in fats, proteins or simple phenols.
Next, we analyzed the presence of polyphenols in the extracts of the aerial parts of A. occidentale (Figure 1B). The leaf extract was highly enriched in polyphenols with 19.08 ± 0.67 µg of gallic acid equivalents (GAE)/mL, followed by the bark extract, with a content of polyphenols of 7.41 ± 0.55 µg GAE/mL The values for fruit and cashew nut were low. The content of polyphenols in the fruit and cashew nut extract were 0.35 ± 0.09 µg GAE/mL and 0.30 ± 0.16 µg GAE/ml, respectively. GCxGC-TOFMS revealed the presence of specific polyphenols in the different extracts (Figure 1C-F; Suppl. Figure S1, Suppl Material). Phenol, benzoic acid, and gallic acid were identified in all four samples ( Suppl. Figure S1). In the leaves aqueous extract trans-catechin, protocatechoic acid, and pyrogallol were all putatively identified. Interestingly, trans-catechin was only found in the leaf extract (Figure 1C; Suppl. Figure S1).
3.2. An Array of Antioxidant Capacity Assays Revealed Differential Antioxidant Potential of the Aerial Parts of A. occidentale
To gain information about the electroactivity of the extracts, the cyclic voltammetry technique based on copper chelation was carried out [26]. Cyclic voltammetry revealed differential electroactive properties of the different samples (Figure 2A-E). A cathodic shift to a higher reduction potential of the signal of Cu+2 reduction compared with the copper control profile was observed in all the samples. This perturbation of the cathodic signal is attributed to the chelation of Cu+2 by the metabolites present in the extracts. The modification was lower in the case of nut extract (Figure 2D). When the oxidation of the stripping Cu0 was analyzed, extract of leaves, bark and fruit showed a splitting of the signal confirming the chelation detected in the cathodic signal (Figure 2A-C). In the case of nuts extract, this anodic signal was very similar to the copper control (Figure 2D), suggesting no evidence of coordination of this extract with copper. Electrochemical quantification of the total antioxidant capacity revealed significant differences (Fig2E). The value for the leaves extract was 1039 ± 54.50 µC, and the bark extract was approximately half (627.2 ± 45.85 µC). The values for the fruit and the cashew nut were significantly lower, with 114.2 ± 7.82 µC and 64.02 ± 2.97 µC, respectively, although the fruit extract was significantly different to the nut sample.
Next, we carried out a set of broadly-used assays to measure scavenging and antioxidant capacity: 2,2′-azino-bis-3-ethylbenzthiazoline-6-sulphonic acid (ABTS) (Fig3 A,B) and Ferric Reducing Antioxidant Power (FRAP) assay (Figure 3C,D) are two methods based on electron transfer capacity while 2,2-diphenyl-1-picryl-hydrazyl-hydrate (DPPH) (Fig3 E,F) is based on hydrogen atom transfer capacity. Inhibition curves were plotted for dose-dependent experiments, and IC50 values were determined along with Trolox Equivalent Antioxidant Capacity (TEAC) value, a well-defined measurement of antioxidant strength. Quantitative parameters corresponding to these antioxidant assays are shown in Table 2.
Leaf extract had the higher ABTS•+ radical scavenger activity (644.40 ± 27.60 (TEAC (µM Trolox)). The bark extract showed slightly lower activity (482.70 ± 13.68 TEAC), meanwhile the scavenging activity of the fruit and nut samples were only 69.97 ± 2.12 and 32.34 ± 7.71 TEAC, respectively (Figure 3A). Both leaves and bark extracts of A. occidentale exhibited high antioxidant activity with IC50 values of 12.3 ± 2.1 and 13.8 ± 4.1 µg/m, respectively (Figure 3B). On the contrary, low inhibitory activities against the radical ABTS•+ were found in fruit and nut extracts. In the case of the fruit extract the IC50 value was 447.2 ± 15.0 µg/mL, while the value for the nut sample was undefined and > 1000 µg/mL (Figure 3B, Table 2).
In alignment with the previous results, similar antioxidant behavior was found when the FRAP assay was carried out. The FRAP assay using 1 mg/ml concentration revealed differential antioxidant properties for leaves and bark extracts (Figure 3C). FRAP value of leaves extract was 470.30 ± 40.44 µM Fe2+ and bark extract showed an antioxidant value of 331.10 ± 19.41 µM Fe2+. These FRAP values were comparatively higher than those observed for the fruit and nut extracts, which were 67.10 ± 5.69 and 0.62 ± 0.62 µM Fe2+, respectively. Leaf and bark extracts were the most antioxidants samples, with values of 0.13 and 0.12 of TEAC, respectively. The antioxidant capacity of fruit and nut extract was close to 0 in this assay, with values of 0.03 and 0.01 of TEAC. (Figure 3D, Table 2).
The DPPH assay showed a similar pattern on radical scavenging capacity (Figure 3E,F). TEAC values for DPPH using 1mg/ml concentration were 513.30 ± 5.64 µM Trolox for leaves extract, 364.7 ± 2.52 µM Trolox for bark extract, 16.95 ± 7.03 µM Trolox for fruit extract and close to 0 µM Trolox for nut extract (Figure 3E). In this case, the IC50 values of the leaves (58.67 µg/mL) and bark (79.52 µg/mL) extracts were significantly higher when compared to fruit and nut samples, the latter with IC values higher than 1000 µg/mL (Figure 3F, Table 2).
In sum, a set of different assays that quantify the H-donating activity and electron transfer ability revealed that leaf and bark aqueous extracts of A. occidentale had significantly higher antioxidant potential than fruit and nut aqueous extracts.
3.3. Leaf and Bark Extracts of A. occidentale Are Protective Against Oxidative Stress in SH-SY5Y Cells
To in-depth evaluate the potential bioactivity of the most potent extracts of A. occidentale, cytotoxicity assays were conducted in SH-SY5Y cells, a human neuroblastoma cell line commonly used in the context of oxidative stress and neurodegeneration [25,26,27] (Figure 4). The dose-response curve revealed the LD50 values, which represents the concentration required to reduce cell viability to 50% compared to the control, were 275 µg/mL and 363 µg/mL for leaf and bark extracts, respectively. Given the low toxicity of the extract, further experiments were carried out to evaluate the potential shielding capacity of the extracts in these human-origin neuronal cells. SH-SY5Y were treated with different concentrations of leaves or bark extracts (ranging from 5 to 200 µg/mL) for 24h prior to H2O2 500 µM treatment for another 4 h. As expected, H2O2 treatment significantly decreased cell viability of SH-SY5Y (p< 0.05). Lower concentrations of the leaf extract could not counteract effectively the H2O2-derived cytotoxicity while higher concentrations restored significantly cell viability (Figure 5). A dose-dependent experiment revealed similar behavior for the bark extract (Figure 5). Exposure to 500 µM H₂O₂ significantly reduced SH-SY5Y cell viability. When cells were pre-treated with low concentrations of the aqueous bark extract no significant protection against oxidative damage was observed. However, treatment with concentrations 10µg/mL and above led to a statistically significant recovery of cell viability.
As expected, under H₂O₂-derived oxidative stress, the level of ROS increased dramatically. Both leaf and bark extracts effectively reduce intracellular ROS levels in a dose-dependent manner (Figure 6). High levels of ROS leads to a reduction in mitochondrial membrane potential causing mitochondrial dysfunction and cell damage [2,25]. Thus, we evaluated the impact of leaf and bark extracts of Anacardium occidentale at the subcellular level using the fluorescent probe 5,5′,6,6′-tetrachloro-1,1′,3,3′-tetraethylbenzimidazolylcarbocyanine iodide (JC-1). JC-1 is commonly used to assess the polarization state of the mitochondrial membrane. In healthy mitochondria, JC-1 forms aggregates inside the organelle and displays red fluorescence. However, in depolarized mitochondria, the JC-1 dye fluoresces green and remains in the cytoplasm of the cells. Thus, the ratio of the red/green fluorescence allows us to infer mitochondrial function[28].
H₂O₂-treated SH-SY5Y cells reduced to 60% the ratio of fluorescence when the dye was measured in 96-well plates (Fig7 A). Interestingly, the pretreatment with any concentration of the leaves' extract resulted in a significant recovery in the mitochondrial membrane potential. Also, pretreatment with bark extract at any tested concentration significantly alleviates the H₂O₂-mitochondrial derived damage. To confirm that the A. occidentale extract might provide mitochondrial protection, flow cytometry experiments using JC-1 were conducted. The FL1 channel captured the green fluorescence that represents damaged mitochondria, while the FL2 channel measured the JC-1 emission in red (healthy mitochondria). H₂O₂-treated cells showed an increase in the ratio BL-1A/BL-2A. Interestingly, the pretreatment with the plant extracts reduced the ratio (Figure 7C). In the experiment using leaf extract, both concentrations were successful in decreasing the BL-1A/BL-2A value. In contrast, only 200 µg/mL of bark extract was protective in terms of mitochondrial function in the flow cytometry analysis.
Altogether, these results indicate that leaf and bark extracts present low cytotoxicity and SH-SY5Y cells exposed to these extracts are more resistant to H₂O₂-derived mitochondrial damage, counteracting oxidative stress.
3.4. Leaf and Bark Extracts (But Not Fruit and Nut Extracts) Trigger the Expression of NRF2-Target Cytoprotective Genes Against Oxidative Stress
While the direct ROS scavenger capacity of phytocompounds present in the extracts of the aerial parts of A. occidentale could be beneficial to combat cellular oxidative stress, the activation of cellular antioxidant response through the expression of cytoprotective genes could be a secondary protective mechanism.
Given that natural compounds present in plant extracts have been shown to trigger the NRF2 signaling route, enhancing the function of antioxidant enzymes[29], we evaluated if extract of different aerial part of the A. occidentale might induce the expression of well-described NRF2 target genes (Figure 8A-H and Suppl. Figure S4). Both leaf and bark extracts impacted the expression of HMOX1, NQO1, GCLC and GCLM, in different cell lines including MEFs, HLECs, and ARPE-19. On the contrary, neither fruit nor nut extracts did alter significantly the expression of these cytoprotective genes.
Leaf and bark extracts induced upregulation of HMOX1 (Figure 8 A,E). This transcriptional increment was statistically significant in MEFs cells (~3-fold in leaves and 8-fold change compared to untreated cells) while high variability was found in HLECs exposed to these extracts (~9-fold in leaves and 12-fold change compared to untreated cells). Fruit and nut extracts did not induce a significant increase in gene expression. Analogously, leaf extract promoted the upregulation of NQO1 (~5-fold compared to the untreated group), and the bark extract resulted in the highest expression (~12-fold compared to untreated cells) in HLECs (Figure 8A). In MEFs, extracts from leaves and bark significantly induced the expression of the NQO1 gene. (~3-fold change, p<0.01 and ~4-fold change, p<0.05, respectively) (Figure 8D).
In the case of genes related to glutathione metabolism, HLEC cells exposed to leaf and bark extracts displayed 4-fold and 7-fold increases in GCLC, respectively (Figure 8C). An approximately 8-fold increase of GCLC expression was detected in MEFs cells treated with leaf and bark extracts, although the variability was higher in this cell type (Figure 8G). GCLC increased slightly in cells treated with fruit and cashew extracts, although this response was not statistically significant in any cell type. Although the upregulation of GLCM took place when both cell types were treated with leaf and bark extracts, only the aqueous leaf extract significantly stimulated GLCM in a statistically significant manner in HLECs (~4-fold change, p<0.05). Neither fruit nor nut samples impacted the expression of this gene. Similar trends were also observed for all the analyzed genes in ARPE-19 cells ( Suppl. Figure S4).
Altogether, these results suggest that phytocompounds present in leaf and bark samples were able to transcriptionally modulate core genes involved in antioxidant defense.
4. Discussion
Although A. occidentale is widely used in herbal remedies by communities in tropical regions, its folkloric use is mainly based on perceived effectiveness against specific ailments, and there is a limited number of studies that provide research-based evidence to authenticate its pharmacological potential. Contemporary research provides fragmented knowledge on the different aerial parts of the plant, with numerous methods of extraction documented in the literature. Some studies shed light on the nutraceutical applications of the cashew nut, a food product that is consumed worldwide, with potential biological applications including modulation of inflammatory and metabolic processes microbiota activity [30,31,32]. The fruit has also been the subject of study, and anti-inflammatory and wound-healing properties were reported in mice [33]. Bark has been suggested as a source of bioactive compounds for dermatology applications [34], and the greater heterogeneity in the biological formulations reported in the literature associated with A. occidentale leaves suggests a higher bioactive and/or therapeutic potential in this aerial part. Antidiabetic, antimalarial, and antimicrobial properties were also reported for leaf extracts [35,36,37]. However, despite the promising therapeutic potential of A. occidentale, a comprehensive and systematic characterization of aerial part extracts has not been reported. Our study presents a thorough analysis of the potential of this medicinal plant, which is used in traditional medicine worldwide, employing a multidisciplinary approach. The arrays of techniques used in our study range from metabolomics, electrochemical analysis to assays that evaluate biological potential to combat oxidative stress. Our comparative analysis revealed that both leaf and bark aqueous extracts are not toxic and confer shielding capacity against oxidative stress (Figure 9). On the contrary, neither fruit nor nut aqueous extracts had a remarkable capacity to scavenge free radicals or trigger the expression of cytoprotective genes.
A battery of assays was used to provide phytochemical profiles and insights into the chemical compounds underlying the antioxidant properties of the different extracts of A. occidentale. Different metabolomic profiles were found, and the TPC assay revealed that the aqueous extract of A. occidentale leaves and bark exhibited the highest polyphenol content. These results were further corroborated by metabolomic experiments, which detected the presence of trans-catechin, gallic, benzoic acid or protocatechuic acid or pyrogallol. In alignment with our findings, other studies have identified similar polyphenols in mostly ethanolic extracts of nuts and leaves of A. occidentale or related species of the Anacardiaceae family [38,39,40]. In our study, cyclic voltammetry and spectrophotometric methods revealed a higher antioxidant capacity in both leaf and bark samples, thereby highlighting the potential therapeutic relevance of these aerial parts. Leaves are a well-recognized source of polyphenols and the presence of bioactive compounds in leaves makes the consumption of leafy vegetables recommended due to their high nutritional benefits [41,42]. Bark extract displayed strong electrolytic activity, which may suggest a significant content of polyphenols, tannins, or flavonoids. As identified in our study, the presence of therapeutically active polyphenols in the bark of numerous plant species has been previously reported [43,44,45,46]. Thus, it is expected to have a relative presence of polyphenols in leaf or bark extract and our findings are consistent with other results found in the literature for different plant species. For example, leaf extracts of Moringa oleifera exhibit greater antioxidant power than stem extracts, with stem extracts in turn showing more antioxidant capacity than branch extracts and Barleria prionitis L. leaves extracts have higher phenolic content and radical scavenger activity than flower and stem extracts [47,48].
On the contrary, metabolomic studies conducted with aqueous extract of A. occidentale fruit revealed a less promising metabolomic profile in terms of potential antioxidant capacity and minimal presence of total polyphenols in the sample. The FTIR assay suggests the presence of polysaccharides or organic acids and, similarly, the spectrum obtained from the cyclic voltammetry assay indicates a high oxidation capacity of the sample, possibly due to the presence of a large quantity of oxidizable sugars. These compounds are consistent with those typically found in fruits, as commonly accumulate high levels of sugars and organic acids such as citric acid, tartaric acid, and ascorbic acid. Also, these results may explain the presence of pectin, a polysaccharide with industrial and biotechnological interest, which has previously been identified in the fruit of A. occidentale [49]. The findings related to the cashew aqueous extract exhibited a pattern similar to that observed with the fruit. The FTIR and TPC analysis indicated the absence of polyphenolic compounds and a certain chemical diversity. Surprisingly, scientific literature has not extensively documented the identification of pharmacologically relevant compounds in the cashew itself. Interestingly, anacardic acid—a molecule with promising therapeutic potential—is typically found in the cashew nutshell under normal conditions, rather than in the cashew kernel. Anacardic acid is thought to exert neuroprotective effects, modulate autophagic processes and possess antibacterial properties [50,51,52,53]. Note that the limited activity in the aqueous extract of fruit or nut could be attributed to the extraction method, as the presence of polyphenols in plant fruits is well documented in scientific literature [54]. It is possible that the polyphenols or other bioactive molecules present in the fruit and nuts are not fully water-soluble, or that the water temperature or pH may have hindered their extraction. Furthermore, the solvent used in maceration, or any extraction method, can be key for the obtention of molecules of interest. In this context, it may be of interest to explore alternative extraction methods, such as infusion, decoction, ultrasound-assisted extraction, or Soxhlet extraction in future experiments. Further experiments should be performed in the future using different organic solvents (or a mixture of these solvents) such as ethanol, hexane, ethyl acetate or methanol.
In recent years, significant efforts have been devoted to identifying phytocompounds—particularly secondary metabolites—that may be useful in the context of nervous system-related or neurodegenerative diseases, where oxidative stress plays a critical role. In such scenarios, plant-derived compounds have gained special relevance due to their demonstrated strong antioxidant properties. In our study, further characterization revealed that the aqueous extracts from leaf and bark demonstrated a high degree of safety, exhibiting cytotoxicity only at very high concentrations, and were protective against oxidative stress and preserved proper mitochondrial function in vitro. In our study, leaf and bark extracts were neuroprotective in SH-SY5Y cells—a cell line commonly used as a model for the study of neurodegenerative diseases and disorders associated with the nervous system. Of note, there is increasing evidence that phytocompounds derived from the Anacardiaceae family might be neuroprotective. Methanolic bark extracts of Anacardium microcarpum, a closely related species to A. occidentale, provided neuroprotection in 6-hydroxydopamine-induced damage in chicken brain slices, and hydroalcoholic and methanolic extracts of the same plant demonstrated antioxidant and protective properties in vivo, showing protective effects against paraquat-induced stress in the Drosophila melanogaster model [55,56]. Hexanolic extract of the plant Spondias mombin (L) (Anacardiaceae) has been shown to ameliorate oxidative status and provide neuroprotection at the behavioral level in zebrafish [57]. Vernicidin B, a flavonoid isolated from the ethanolic fraction of the plant Toxicodendron vernicifluum (Anacardiaceae), demonstrated neuroprotective properties by decreasing the production of ROS and reducing apoptosis induced by exposure to H₂O₂[58]. Finally, the neuroprotective properties of leaf extracts of A. occidentale in vitro in HT22 and Neuro-2a cells were reported by improving neuroinflammation markers and various parameters related to neural function in rats [59,60].
Traditionally, the antioxidant capacity of a given plant extract has been directly linked, at least in part, to the polyphenol content of the sample. These bioactive molecules possess radical scavenging capacity and inherent antioxidant properties. However, growing literature suggests that polyphenols can modulate different cellular processes such as autophagy, fat and glucose metabolism, inflammation, and the response to oxidative stress [61]. The transcription factor NRF2 is a master regulator of genes related to the response to stress induced by free radicals. The development of therapeutic strategies targeting this transcription factor could be instrumental in addressing damage associated with oxidative stress and as recently described, in neurodegenerative diseases [62]. The idea that plant extracts may activate the NRF2 route is a concept that is beginning to be explored in the field of phytotherapy. In our study, both leaf and bark extract enhanced NRF2-targets (NQO1, HMOX1, GCLC and GCLM) in different cell types, suggesting that the impact of bioactive phytocompounds in the modulation of these cytoprotective genes is universal. In this regard, it is noteworthy that other plant extracts were reported to activate NRF2, including saponin extract of Panax notoginseng, olive leaf extract Olea europaea or leaves' ethanolic extract of Rosmarinus officinalis L. [63,64,65]. Nevertheless, although our findings suggest the importance of NRF2 as a mediator of the response derived of leaf and bark extract of A. occidentale, we cannot discard a more complex scenario involving, for example, other transcription factors.
Taken together, our systematic characterization points out that the extracts of A. occidentale arise as a potential therapeutic remedy against oxidative stress, within the context of neuroprotection. One of the upcoming challenges will be the identification, isolation, and validation of phytocompounds with high antioxidant power or the formulation of the extracts for therapeutic purposes. The development of these therapeutic strategies derived from A. occidentale might be a formulation to consider in pathological contexts with chronic oxidative stress, such as aging or aging-related diseases.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1. Metabolites identified in the A.occidentale leaf extract by GC×GC-TOFMS; Table S2. Metabolites identified in the A.occidentale bark extract by GC×GC-TOFMS; Table S3. Metabolites identified in the A.occidentale fruit extract by GC×GC-TOFMS; Table S4. Metabolites identified in the A.occidentale nut extract by GC×GC-TOFMS.
Author Contributions
Conceptualization (APM, LG-M, GC-T and EB); methodology (JLL, RTG, GC-T and EB); formal analysis (JLL, AL, RTG, GC-T and EB); investigation (APM, LG-M, JLL, RTG, AD-B, AL, IG-R, SS, VM-M, MS,JJH); resources (JLL, JJH, GC-T and EB); writing—original draft (APM); visualization (JLL, SS, AD-B, GC-T and EB); writing—review and editing (LG-M, JLL, SS, AD-B, AL,GC-T and EB); supervision (GC-T and EB); project administration (GC-T and EB); funding acquisition (LG-M, GC-T and EB). All authors read and approved the final manuscript.
Funding
This work was supported by RYC2018-024434-I, MINECO PID2023-151873OB-I00 (to EB) and IDOC23-06 (to LGM).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Hajam, Y.A.; Rani, R.; Ganie, S.Y.; Sheikh, T.A.; Javaid, D.; Qadri, S.S.; Pramodh, S.; Alsulimani, A.; Alkhanani, M.F.; Harakeh, S.; et al. Oxidative Stress in Human Pathology and Aging: Molecular Mechanisms and Perspectives. Cells 2022, 11, 552. [Google Scholar] [CrossRef] [PubMed]
- Yan, M.H.; Wang, X.; Zhu, X. Mitochondrial defects and oxidative stress in Alzheimer disease and Parkinson disease. Free radical biology & medicine 2013, 62, 90–101. [Google Scholar] [CrossRef]
- Liguori, I.; Russo, G.; Curcio, F.; Bulli, G.; Aran, L.; Della-Morte, D.; Gargiulo, G.; Testa, G.; Cacciatore, F.; Bonaduce, D.; et al. Oxidative stress, aging, and diseases. Clinical interventions in aging 2018, 13, 757–772. [Google Scholar] [CrossRef]
- Yeung, A.W.K.; Tzvetkov, N.T.; Georgieva, M.G.; Ognyanov, I.V.; Kordos, K.; Jóźwik, A.; Kühl, T.; Perry, G.; Petralia, M.C.; Mazzon, E.; et al. Reactive Oxygen Species and Their Impact in Neurodegenerative Diseases: Literature Landscape Analysis. Antioxidants & redox signaling 2021, 34, 402–420. [Google Scholar] [CrossRef]
- Böhm, E.W.; Buonfiglio, F.; Voigt, A.M.; Bachmann, P.; Safi, T.; Pfeiffer, N.; Gericke, A. Oxidative stress in the eye and its role in the pathophysiology of ocular diseases. Redox Biology 2023, 68, 102967. [Google Scholar] [CrossRef]
- Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Squadrito, F.; Altavilla, D.; Bitto, A. Oxidative Stress: Harms and Benefits for Human Health. Oxidative Medicine and Cellular Longevity 2017, 2017, 8416763. [Google Scholar] [CrossRef]
- Vomund, S.; Schäfer, A.; Parnham, M.J.; Brüne, B.; von Knethen, A. Nrf2, the Master Regulator of Anti-Oxidative Responses. International journal of molecular sciences 2017, 18. [Google Scholar] [CrossRef]
- Birben, E.; Sahiner, U.M.; Sackesen, C.; Erzurum, S.; Kalayci, O. Oxidative stress and antioxidant defense. The World Allergy Organization journal 2012, 5, 9–19. [Google Scholar] [CrossRef]
- Ngo, V.; Duennwald, M.L. Nrf2 and Oxidative Stress: A General Overview of Mechanisms and Implications in Human Disease. Antioxidants (Basel, Switzerland) 2022, 11. [Google Scholar] [CrossRef]
- Schmidlin, C.J.; Dodson, M.B.; Madhavan, L.; Zhang, D.D. Redox regulation by NRF2 in aging and disease. Free Radical Biology and Medicine 2019, 134, 702–707. [Google Scholar] [CrossRef]
- Masyita, A.; Mustika Sari, R.; Dwi Astuti, A.; Yasir, B.; Rahma Rumata, N.; Emran, T.B.; Nainu, F.; Simal-Gandara, J. Terpenes and terpenoids as main bioactive compounds of essential oils, their roles in human health and potential application as natural food preservatives. Food chemistry: X 2022, 13, 100217. [Google Scholar] [CrossRef] [PubMed]
- Heinrich, M.; Mah, J.; Amirkia, V. Alkaloids Used as Medicines: Structural Phytochemistry Meets Biodiversity—An Update and Forward Look. Molecules 2021, 26, 1836. [Google Scholar] [CrossRef] [PubMed]
- Rana, A.; Samtiya, M.; Dhewa, T.; Mishra, V.; Aluko, R.E. Health benefits of polyphenols: A concise review. Journal of food biochemistry 2022, 46, e14264. [Google Scholar] [CrossRef] [PubMed]
- Garnatje, T.; Peñuelas, J.; Vallès, J. Ethnobotany, Phylogeny, and ‘Omics’ for Human Health and Food Security. Trends in Plant Science 2017, 22, 187–191. [Google Scholar] [CrossRef]
- Guerra, R.N.M.; Oliveira, A.S.; Farias, J.R.; Franco, D.C.G.; Santos, P.G.; Barbosa, N.T.; Muniz, S.B.; Abreu, A.G.; Nascimento, F.R.F. Anacardiaceae Family: Effect of Isolated Compounds and Other Identified Phytochemicals Against Clinically Relevant Candida Species—A Short Review. Antibiotics 2025, 14, 308. [Google Scholar] [CrossRef]
- Méril-Mamert, V.; Ponce-Mora, A.; Sylvestre, M.; Lawrence, G.; Bejarano, E.; Cebrián-Torrejón, G. Antidiabetic Potential of Plants from the Caribbean Basin. Plants 2022, 11, 1360. [Google Scholar] [CrossRef]
- Ukwenya, V.O.; Alese, M.O.; Ogunlade, B.; Folorunso, I.M.; Omotuyi, O.I. Anacardium occidentale leaves extract and riboceine mitigate hyperglycemia through anti-oxidative effects and modulation of some selected genes associated with diabetes. Journal of diabetes and metabolic disorders 2023, 22, 455–468. [Google Scholar] [CrossRef]
- Thesnor, V.; Molinié, R.; Giebelhaus, R.T.; de la Mata Espinosa, A.P.; Harynuk, J.J.; Bénimélis, D.; Vanhoye, B.; Dunyach-Rémy, C.; Sylvestre, M.; Cheremond, Y.; et al. Antibacterial Activity and Untargeted Metabolomics Profiling of Acalypha arvensis Poepp. Molecules 2023, 28, 7882. [Google Scholar] [CrossRef]
- Giebelhaus, R.T.; Nguyen, G.; Schmidt, S.A.; Wang, S.; Mesfin, E.Y.; Nam, S.L.; de la Mata, A.P.; Harynuk, J.J. GC×GC-TOFMS Analysis of Fecal Metabolome Stabilized Using an At-Home Stool Collection Device. Applied Biosciences 2024, 3, 348–359. [Google Scholar] [CrossRef]
- Sumner, L.W.; Amberg, A.; Barrett, D.; Beale, M.H.; Beger, R.; Daykin, C.A.; Fan, T.W.; Fiehn, O.; Goodacre, R.; Griffin, J.L.; et al. Proposed minimum reporting standards for chemical analysis Chemical Analysis Working Group (CAWG) Metabolomics Standards Initiative (MSI). Metabolomics : Official journal of the Metabolomic Society 2007, 3, 211–221. [Google Scholar] [CrossRef]
- Nam, S.L.; Giebelhaus, R.T.; Tarazona Carrillo, K.S.; de la Mata, A.P.; Harynuk, J.J. Evaluation of normalization strategies for GC-based metabolomics. Metabolomics : Official journal of the Metabolomic Society 2024, 20, 22. [Google Scholar] [CrossRef] [PubMed]
- Brinvillier, D.; Barrast, M.; Couderc-Murillo, P.; Bono-Yagüe, J.; Rousteau, A.; Gómez Escribano, A.P.; Palmeira-Mello, M.V.; Doménech-Carbó, A.; Passe-Coutrin, N.; Sylvestre, M.; et al. Spectroscopic, Electrochemical, and Biological Assays of Copper-Binding Molecules for Screening of Different Drugs and Plant Extracts against Neurodegenerative Disorders. ACS Omega 2022, 7, 16260–16269. [Google Scholar] [CrossRef] [PubMed]
- Matignon, L.; Lo, M.M.; Monpierre, M.; Correia, M.V.; Valencia, D.P.; Palmeira-Mello, M.V.; Sylvestre, M.-N.; Pruneau, L.; Sylvestre, M.; Domenech, A.; et al. Phytochemical and Biological Study of Trophic Interaction between Pseudosphinx Tetrio L. Larvae and Allamanda Cathartica L. Plants 2023, 12, 520. Larvae and Allamanda Cathartica L. Plants 2023, 12, 520. [Google Scholar] [PubMed]
- Corpas, F.J.; Rodríguez-Ruiz, M.; Campos, M.J.; Taboada, J.; Palma, J.M. Electrochemical Detection of Total Antioxidant Capacity (TAC) in Plant Tissues from Different Origins. Methods in molecular biology (Clifton, N.J.) 2024, 2798, 1–9. [Google Scholar] [CrossRef]
- Nieto, C.I.; Cornago, M.P.; Cabildo, M.P.; Sanz, D.; Claramunt, R.M.; Torralba, M.C.; Torres, M.R.; Martínez Casanova, D.; Sánchez-Alegre, Y.R.; Escudero, E.; et al. Evaluation of the Antioxidant and Neuroprotectant Activities of New Asymmetrical 1,3-Diketones. Molecules 2018, 23, 1837. [Google Scholar] [CrossRef]
- Angelé-Martínez, C.; T., N.K.V.; S., A.F.; N., A.J.; and Brumaghim, J.L. Reactive oxygen species generation by copper(II) oxide nanoparticles determined by DNA damage assays and EPR spectroscopy. Nanotoxicology 2017, 11, 278–288. [Google Scholar] [CrossRef]
- Pang, Q.Q.; Kim, J.H.; Kim, H.Y.; Kim, J.-H.; Cho, E.J. Protective Effects and Mechanisms of Pectolinarin against H2O2-Induced Oxidative Stress in SH-SY5Y Neuronal Cells. Molecules 2023, 28, 5826. [Google Scholar] [CrossRef]
- Salvioli, S.; Ardizzoni, A.; Franceschi, C.; Cossarizza, A. JC-1, but not DiOC6(3) or rhodamine 123, is a reliable fluorescent probe to assess delta psi changes in intact cells: implications for studies on mitochondrial functionality during apoptosis. FEBS letters 1997, 411, 77–82. [Google Scholar] [CrossRef]
- Wang, T.; Liu, M.; Li, X.; Zhang, S.; Gu, H.; Wei, X.; Wang, X.; Xu, Z.; Shen, T. Naturally-derived modulators of the Nrf2 pathway and their roles in the intervention of diseases. Free Radical Biology and Medicine 2024, 225, 560–580. [Google Scholar] [CrossRef]
- Menezes, F.; da Cruz Almeida É, T.; da Silva Vieira, A.R.; de Souza Aquino, J.; Dos Santos Lima, M.; Magnani, M.; de Souza, E.L. Impact of Cashew (Anacardium occidentale L.) by-Product on Composition and Metabolic Activity of Human Colonic Microbiota In Vitro Indicates Prebiotic Properties. Current microbiology 2021, 78, 2264–2274. [Google Scholar] [CrossRef]
- Siracusa, R.; Fusco, R.; Peritore, A.F.; Cordaro, M.; D’Amico, R.; Genovese, T.; Gugliandolo, E.; Crupi, R.; Smeriglio, A.; Mandalari, G.; et al. The Antioxidant and Anti-Inflammatory Properties of Anacardium occidentale L. Cashew Nuts in a Mouse Model of Colitis. Nutrients 2020, 12, 834. [Google Scholar] [CrossRef] [PubMed]
- Dias, C.C.Q.; Madruga, M.S.; Pintado, M.M.E.; Almeida, G.H.O.; Alves, A.P.V.; Dantas, F.A.; Bezerra, J.K.G.; de Melo, M.; Viera, V.B.; Soares, J.K.B. Cashew nuts (Anacardium occidentale L.) decrease visceral fat, yet augment glucose in dyslipidemic rats. PloS one 2019, 14, e0225736. [Google Scholar] [CrossRef] [PubMed]
- da Silveira Vasconcelos, M.; Gomes-Rochette, N.F.; de Oliveira, M.L.; Nunes-Pinheiro, D.C.; Tomé, A.R.; Maia de Sousa, F.Y.; Pinheiro, F.G.; Moura, C.F.; Miranda, M.R.; Mota, E.F.; et al. Anti-inflammatory and wound healing potential of cashew apple juice (Anacardium occidentale L.) in mice. Experimental biology and medicine (Maywood, N.J.) 2015, 240, 1648–1655. [Google Scholar] [CrossRef]
- Borges, J. Cashew tree (Anacardium occidentale): Possible applications in dermatology. Clinics in dermatology 2021, 39, 493–495. [Google Scholar] [CrossRef]
- Jaiswal, Y.S.; Tatke, P.A.; Gabhe, S.Y.; Vaidya, A.B. Antidiabetic activity of extracts of Anacardium occidentale Linn. leaves on n-streptozotocin diabetic rats. Journal of traditional and complementary medicine 2017, 7, 421–427. [Google Scholar] [CrossRef]
- Kaushik, M.; Hoti, S.L.; Saxena, J.K.; Hingamire, T.; Shanmugam, D.; Joshi, R.K.; Metgud, S.C.; Ungar, B.; Singh, I.; Hegde, H.V. Antimalarial Activity of Anacardium occidentale Leaf Extracts Against Plasmodium falciparum Transketolase (PfTK). Acta parasitologica 2023, 68, 832–841. [Google Scholar] [CrossRef]
- Sunderam, V.; Thiyagarajan, D.; Lawrence, A.V.; Mohammed, S.S.S.; Selvaraj, A. In-vitro antimicrobial and anticancer properties of green synthesized gold nanoparticles using Anacardium occidentale leaves extract. Saudi journal of biological sciences 2019, 26, 455–459. [Google Scholar] [CrossRef]
- Sruthi, P.; Roopavathi, C.; Madhava Naidu, M. Profiling of phenolics in cashew nut (Anacardium occidentale L.) testa and evaluation of their antioxidant and antimicrobial properties. Food Bioscience 2023, 51, 102246. [Google Scholar] [CrossRef]
- Désiré, G.N.S.; Simplice, F.H.; Guillaume, C.W.; Kamal, F.Z.; Parfait, B.; Hermann, T.D.S.; Hervé, N.A.H.; Eglantine, K.W.; Linda, D.K.J.; Roland, R.N.; et al. Cashew (Anacardium occidentale) Extract: Possible Effects on Hypothalamic–Pituitary–Adrenal (HPA) Axis in Modulating Chronic Stress. Brain sciences 2023, 13, 1561. [Google Scholar] [CrossRef]
- Lima Júnior, J.P.d.; Franco, R.R.; Saraiva, A.L.; Moraes, I.B.; Espindola, F.S. Anacardium humile St. Hil as a novel source of antioxidant, antiglycation and α-amylase inhibitors molecules with potential for management of oxidative stress and diabetes. Journal of Ethnopharmacology 2021, 268, 113667. [Google Scholar] [CrossRef]
- Amat-ur-Rasool, H.; Symes, F.; Tooth, D.; Schaffert, L.-N.; Elmorsy, E.; Ahmed, M.; Hasnain, S.; Carter, W.G. Potential Nutraceutical Properties of Leaves from Several Commonly Cultivated Plants. Biomolecules 2020, 10, 1556. [Google Scholar] [CrossRef] [PubMed]
- Masiala, A.; Vingadassalon, A.; Aurore, G. Polyphenols in edible plant leaves: an overview of their occurrence and health properties. Food & function 2024, 15, 6847–6882. [Google Scholar] [CrossRef]
- Yazaki, Y. Utilization of flavonoid compounds from bark and wood: a review. Natural product communications 2015, 10, 513–520. [Google Scholar] [CrossRef] [PubMed]
- Faggian, M.; Bernabè, G.; Ferrari, S.; Francescato, S.; Baratto, G.; Castagliuolo, I.; Dall’Acqua, S.; Peron, G. Polyphenol-Rich Larix decidua Bark Extract with Antimicrobial Activity against Respiratory-Tract Pathogens: A Novel Bioactive Ingredient with Potential Pharmaceutical and Nutraceutical Applications. Antibiotics 2021, 10, 789. [Google Scholar] [CrossRef]
- Wafa, D.; Nicolas, B. Pine Bark Phenolic Extracts, Current Uses, and Potential Food Applications: A Review. Current Pharmaceutical Design 2020, 26, 1866–1879. [Google Scholar] [CrossRef]
- Her, Y.; Lee, T.K.; Sim, H.; Lee, J.C.; Kim, D.W.; Choi, S.Y.; Hong, J.K.; Lee, J.W.; Kim, J.D.; Won, M.H.; et al. Pinus thunbergii bark extract rich in flavonoids promotes hair growth in dorsal skin by regulating inflammatory cytokines and increasing growth factors in mice. Molecular medicine reports 2022, 25. [Google Scholar] [CrossRef]
- Shih, M.-C.; Chang, C.-M.; Kang, S.-M.; Tsai, M.-L. Effect of Different Parts (Leaf, Stem and Stalk) and Seasons (Summer and Winter) on the Chemical Compositions and Antioxidant Activity of Moringa oleifera. International journal of molecular sciences 2011, 12, 6077–6088. [Google Scholar] [CrossRef]
- Sunil, K.J.; Mukesh, K.D.; Sanjeeb, D.; Arti Raj, V.; Ch, V.R. A comparative study on total phenolic content, reducing power and free radical scavenging activity of aerial parts of Barleria prionitis. International Journal of Phytomedicine 2010, 2, 155–159. [Google Scholar]
- Tamiello-Rosa, C.S.; Cantu-Jungles, T.M.; Iacomini, M.; Cordeiro, L.M.C. Pectins from cashew apple fruit (Anacardium occidentale): Extraction and chemical characterization. Carbohydrate research 2019, 483, 107752. [Google Scholar] [CrossRef]
- Liu, Y.; Zhao, Z.; Guo, J.; Ma, Y.; Li, J.; Ji, H.; Chen, Z.; Zheng, J. Anacardic acid improves neurological deficits in traumatic brain injury by anti-ferroptosis and anti-inflammation. Experimental neurology 2023, 370, 114568. [Google Scholar] [CrossRef]
- da Silva, W.M.B.; Pinheiro, S.O.; Alves, D.R.; de Menezes, J.; Magalhães, F.E.A.; Silva, F.C.O.; Marinho, M.M.; Marinho, E.S.; de Morais, S.M. Anacardic Acid Complexes as Possible Agents Against Alzheimer's Disease Through Their Antioxidant, In vitro, and In silico Anticholinesterase and Ansiolic Actions. Neurotoxicity research 2021, 39, 467–476. [Google Scholar] [CrossRef] [PubMed]
- Seong, Y.A.; Shin, P.G.; Yoon, J.S.; Yadunandam, A.K.; Kim, G.D. Induction of the endoplasmic reticulum stress and autophagy in human lung carcinoma A549 cells by anacardic acid. Cell biochemistry and biophysics 2014, 68, 369–377. [Google Scholar] [CrossRef] [PubMed]
- Hollands, A.; Corriden, R.; Gysler, G.; Dahesh, S.; Olson, J.; Raza Ali, S.; Kunkel, M.T.; Lin, A.E.; Forli, S.; Newton, A.C.; et al. Natural Product Anacardic Acid from Cashew Nut Shells Stimulates Neutrophil Extracellular Trap Production and Bactericidal Activity. The Journal of biological chemistry 2016, 291, 13964–13973. [Google Scholar] [CrossRef] [PubMed]
- Williamson, G. The role of polyphenols in modern nutrition. Nutrition Bulletin 2017, 42, 226–235. [Google Scholar] [CrossRef]
- Martins, I.K.; de Carvalho, N.R.; Macedo, G.E.; Rodrigues, N.R.; Ziech, C.C.; Vinadé, L.; Filho, V.M.B.; Menezes, I.A.; Franco, J.; Posser, T. Anacardium microcarpum Promotes Neuroprotection Dependently of AKT and ERK Phosphorylation but Does Not Prevent Mitochondrial Damage by 6-OHDA. Oxidative Medicine and Cellular Longevity 2018, 2018, 2131895. [Google Scholar] [CrossRef]
- Müller, K.R.; Martins, I.K.; Rodrigues, N.R.; da Cruz, L.C.; Barbosa Filho, V.M.; Macedo, G.E.; da Silva, G.F.; Kamdem, J.P.; de Menezes, I.R.A.; Franco, J.L.; et al. Anacardium microcarpum extract and fractions protect against paraquat-induced toxicity in Drosophila melanogaster. EXCLI journal 2017, 16, 302–312. [Google Scholar] [CrossRef]
- Santo, G.D.; de Veras, B.O.; Rico, E.; Magro, J.D.; Agostini, J.F.; Vieira, L.D.; Calisto, J.F.F.; Mocelin, R.; de Sá Fonseca, V.; Wanderley, A.G. Hexane extract from SpoSndias mombin L. (Anacardiaceae) prevents behavioral and oxidative status changes on model of Parkinson's disease in zebrafish. Comparative biochemistry and physiology. Toxicology & pharmacology : CBP 2021, 241, 108953. [Google Scholar] [CrossRef]
- Zhong, T.; Li, M.; Wu, H.; Wang, D.; Liu, J.; Xu, Y.; Fan, Y. Novel Flavan-3,4-diol vernicidin B from Toxicodendron Vernicifluum (Anacardiaceae) as potent antioxidant via IL-6/Nrf2 cross-talks pathways. Phytomedicine : international journal of phytotherapy and phytopharmacology 2022, 100, 154041. [Google Scholar] [CrossRef]
- Oyagbemi, A.A.; Kolawole, A.A.; Elizabeth, A.O.; Oluwaseun, A.K.; Racheal, F.O.; Olanrewaju, E.O.; Olabisi, A.T.; Seun, O.B.; Olubunmi, F.O.; Omolola, O.I.; et al. Leaf extract of Anacardium occidentale ameliorates biomarkers of neuroinflammation, memory loss, and neurobehavioral deficit in N(ω)-nitro-L-arginine methyl ester (L-NAME) treated rats. Biomarkers : biochemical indicators of exposure, response, and susceptibility to chemicals 2023, 28, 263–272. [Google Scholar] [CrossRef]
- Duangjan, C.; Rangsinth, P.; Zhang, S.; Wink, M.; Tencomnao, T. Anacardium Occidentale L. Leaf Extracts Protect Against Glutamate/H(2)O(2)-Induced Oxidative Toxicity and Induce Neurite Outgrowth: The Involvement of SIRT1/Nrf2 Signaling Pathway and Teneurin 4 Transmembrane Protein. Frontiers in pharmacology 2021, 12, 627738. [Google Scholar] [CrossRef]
- Ponce-Mora, A.; Salazar, N.A.; Domenech-Bendaña, A.; Locascio, A.; Bejarano, E.; Gimeno-Mallench, L. Interplay Between Polyphenols and Autophagy: Insights From an Aging Perspective. Frontiers in bioscience (Landmark edition) 2025, 30, 25728. [Google Scholar] [CrossRef] [PubMed]
- Mayer, C.; Riera-Ponsati, L.; Kauppinen, S.; Klitgaard, H.; Erler, J.T.; Hansen, S.N. Targeting the NRF2 pathway for disease modification in neurodegenerative diseases: mechanisms and therapeutic implications. Frontiers in pharmacology 2024, 15, 1437939. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zheng, G.; Yang, S.; Liu, S.; Wu, Y.; Miao, Y.; Liang, Z.; Hua, Y.; Zhang, J.; Shi, J.; et al. The plant extract PNS mitigates atherosclerosis via promoting Nrf2-mediated inhibition of ferroptosis through reducing USP2-mediated Keap1 deubiquitination. British journal of pharmacology 2024, 181, 4822–4844. [Google Scholar] [CrossRef] [PubMed]
- HAS, A.L.; Alotaibi, M.F.; Bin-Jumah, M.; Elgebaly, H.; Mahmoud, A.M. Olea europaea leaf extract up-regulates Nrf2/ARE/HO-1 signaling and attenuates cyclophosphamide-induced oxidative stress, inflammation and apoptosis in rat kidney. Biomedicine & pharmacotherapy = Biomedecine & pharmacotherapie 2019, 111, 676–685. [Google Scholar] [CrossRef]
- Wang, H.; Cheng, J.; Yang, S.; Cui, S.W.; Wang, M.; Hao, W. Rosemary extract reverses oxidative stress through activation of Nrf2 signaling pathway in hamsters fed on high fat diet and HepG2 cells. Journal of Functional Foods 2020, 74, 104136. [Google Scholar] [CrossRef]
Figure 1.
Metabolomics reveals differential polyphenol composition in aqueous extracts of the aerial part of Anacardium occidentale. Composite of FT-IR spectra profiles (A), total phenolic content for leaf (black), bark (blue), fruit (pink) and nut (green) aqueous extract of Anacardium occidentale (B), GC×GC-TOFMS profiles for (C) leaf, (D) bark, (E) fruit and (F) nut extracts. Specific polyphenols are indicated in each profile. n = 3; *, p < 0.05; **, p < 0.01; ***, p < 0.001, t test.
Figure 1.
Metabolomics reveals differential polyphenol composition in aqueous extracts of the aerial part of Anacardium occidentale. Composite of FT-IR spectra profiles (A), total phenolic content for leaf (black), bark (blue), fruit (pink) and nut (green) aqueous extract of Anacardium occidentale (B), GC×GC-TOFMS profiles for (C) leaf, (D) bark, (E) fruit and (F) nut extracts. Specific polyphenols are indicated in each profile. n = 3; *, p < 0.05; **, p < 0.01; ***, p < 0.001, t test.
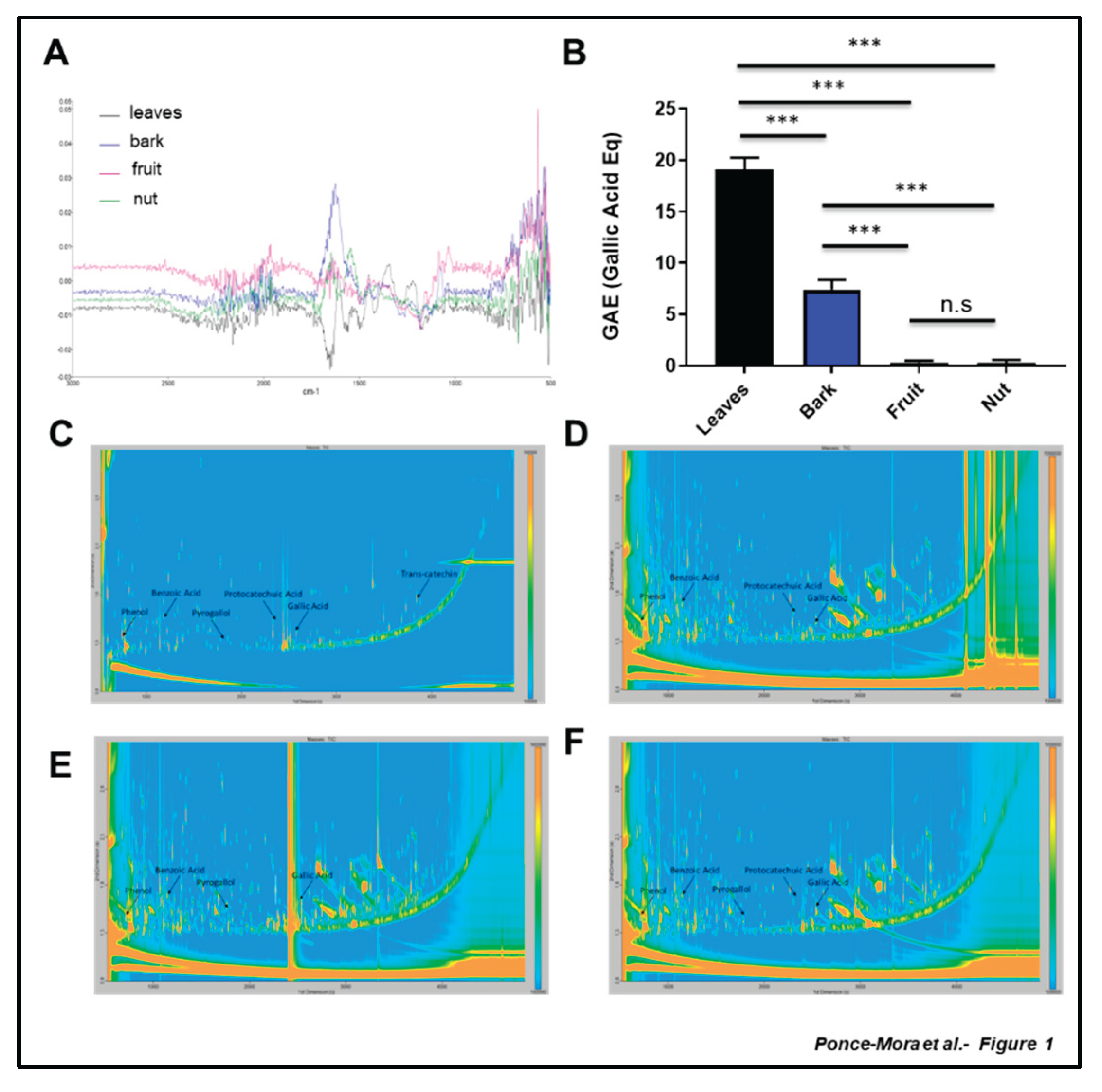
Figure 2.
Electrochemical analysis reveals differential antioxidant capacity of the aerial part of Anacardium occidentale. Cyclic voltammograms of films originating from leaf (A), bark (B), fruit (C) and nut (D) aqueous extract of Anacardium occidentale. Overlapping of the different profiles is shown (E). Total Antioxidant activity was using the BRS device (F). Measurements are expressed in microcoulombs (µC). n = 3; *, p < 0.05; **, p < 0.01; ***, p < 0.001, t test.
Figure 2.
Electrochemical analysis reveals differential antioxidant capacity of the aerial part of Anacardium occidentale. Cyclic voltammograms of films originating from leaf (A), bark (B), fruit (C) and nut (D) aqueous extract of Anacardium occidentale. Overlapping of the different profiles is shown (E). Total Antioxidant activity was using the BRS device (F). Measurements are expressed in microcoulombs (µC). n = 3; *, p < 0.05; **, p < 0.01; ***, p < 0.001, t test.
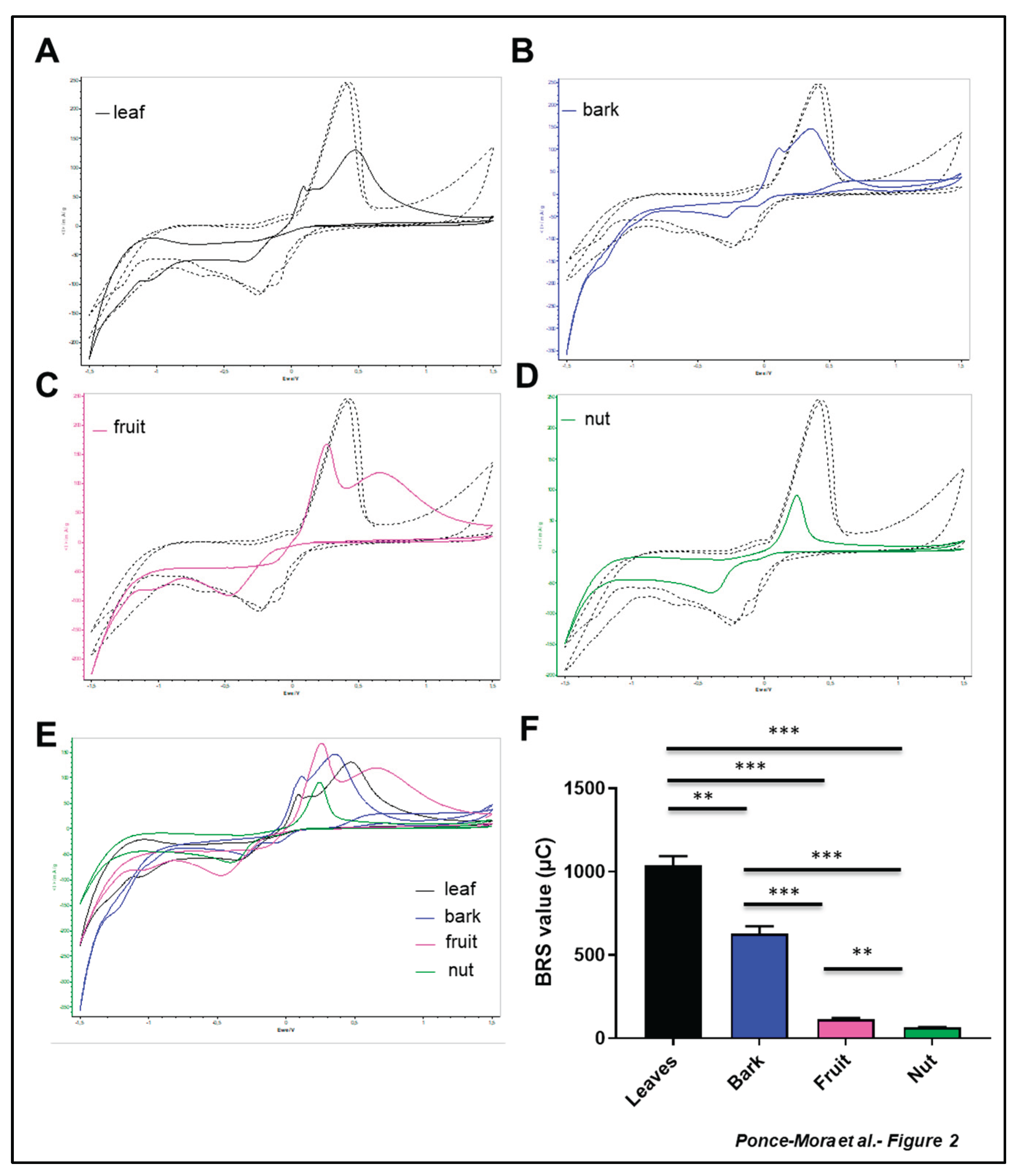
Figure 3.
Antioxidant capacity assay of aerial part of A. occidentale. (A-B) ABTS (2,2'-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid)), (C-D) Ferric Reducing Antioxidant Potential (FRAP) and (E-F) 2,2-diphenyl-1-picrylhydrazyl (DPPH) assays were performed in leaf (black), bark (blue), fruit (pink) and nut (green) aqueous extracts of A.occidentale. TEAC: Trolox Equivalents Antioxidant Capacity n = 3; *, p < 0.05; **, p < 0.01; ***, p < 0.001, t test.
Figure 3.
Antioxidant capacity assay of aerial part of A. occidentale. (A-B) ABTS (2,2'-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid)), (C-D) Ferric Reducing Antioxidant Potential (FRAP) and (E-F) 2,2-diphenyl-1-picrylhydrazyl (DPPH) assays were performed in leaf (black), bark (blue), fruit (pink) and nut (green) aqueous extracts of A.occidentale. TEAC: Trolox Equivalents Antioxidant Capacity n = 3; *, p < 0.05; **, p < 0.01; ***, p < 0.001, t test.
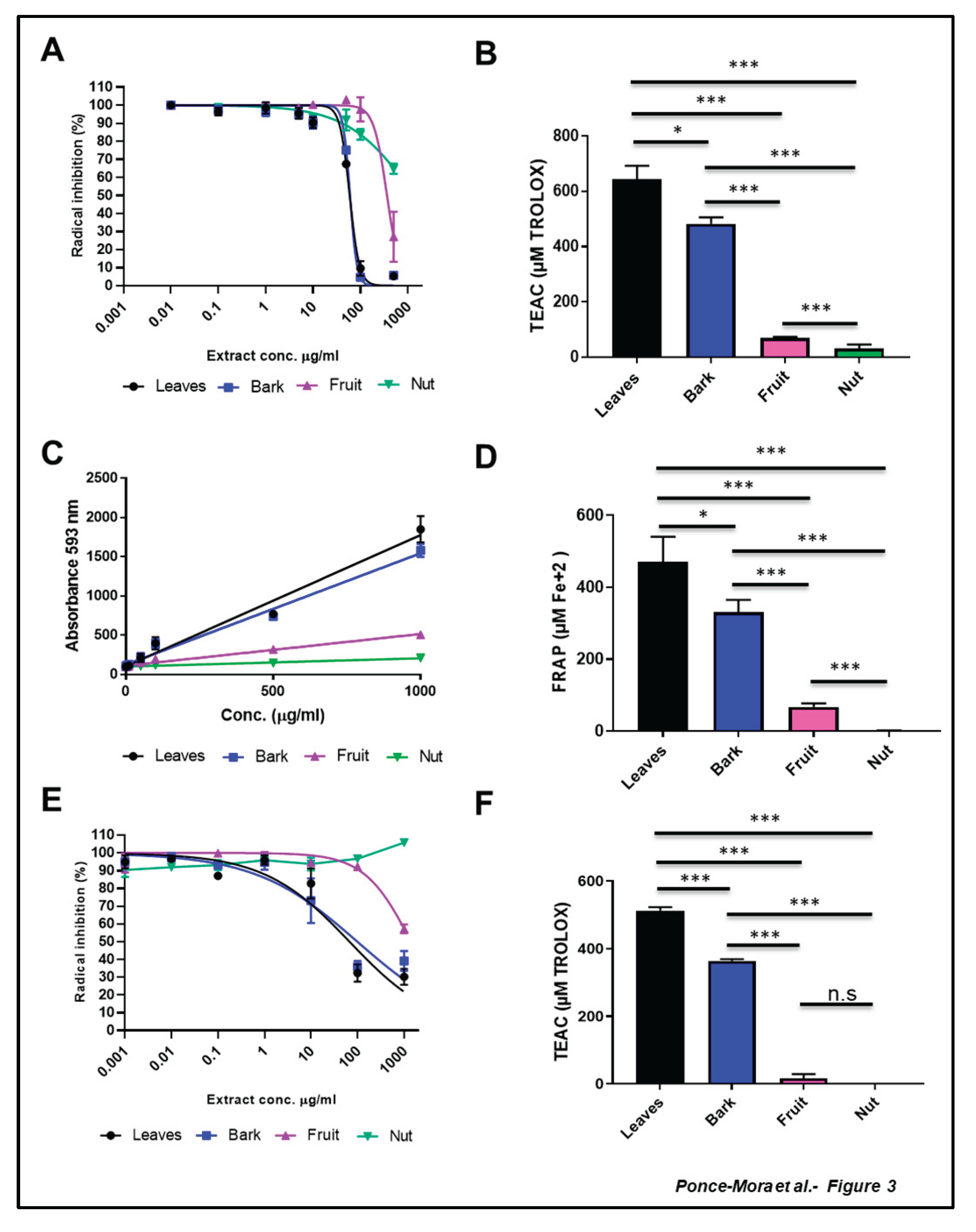
Figure 4.
Cytotoxicity of leaves and bark extracts in SH-SY5Y cells. Cell viability was analyzed using MTT assay. Extracts were administered at different concentrations for 24 h prior to H2O2 500 µM treatment for other 4 h. Data are presented as Mean ± SD of three independent experiments carried out by sextuplicate.
Figure 4.
Cytotoxicity of leaves and bark extracts in SH-SY5Y cells. Cell viability was analyzed using MTT assay. Extracts were administered at different concentrations for 24 h prior to H2O2 500 µM treatment for other 4 h. Data are presented as Mean ± SD of three independent experiments carried out by sextuplicate.
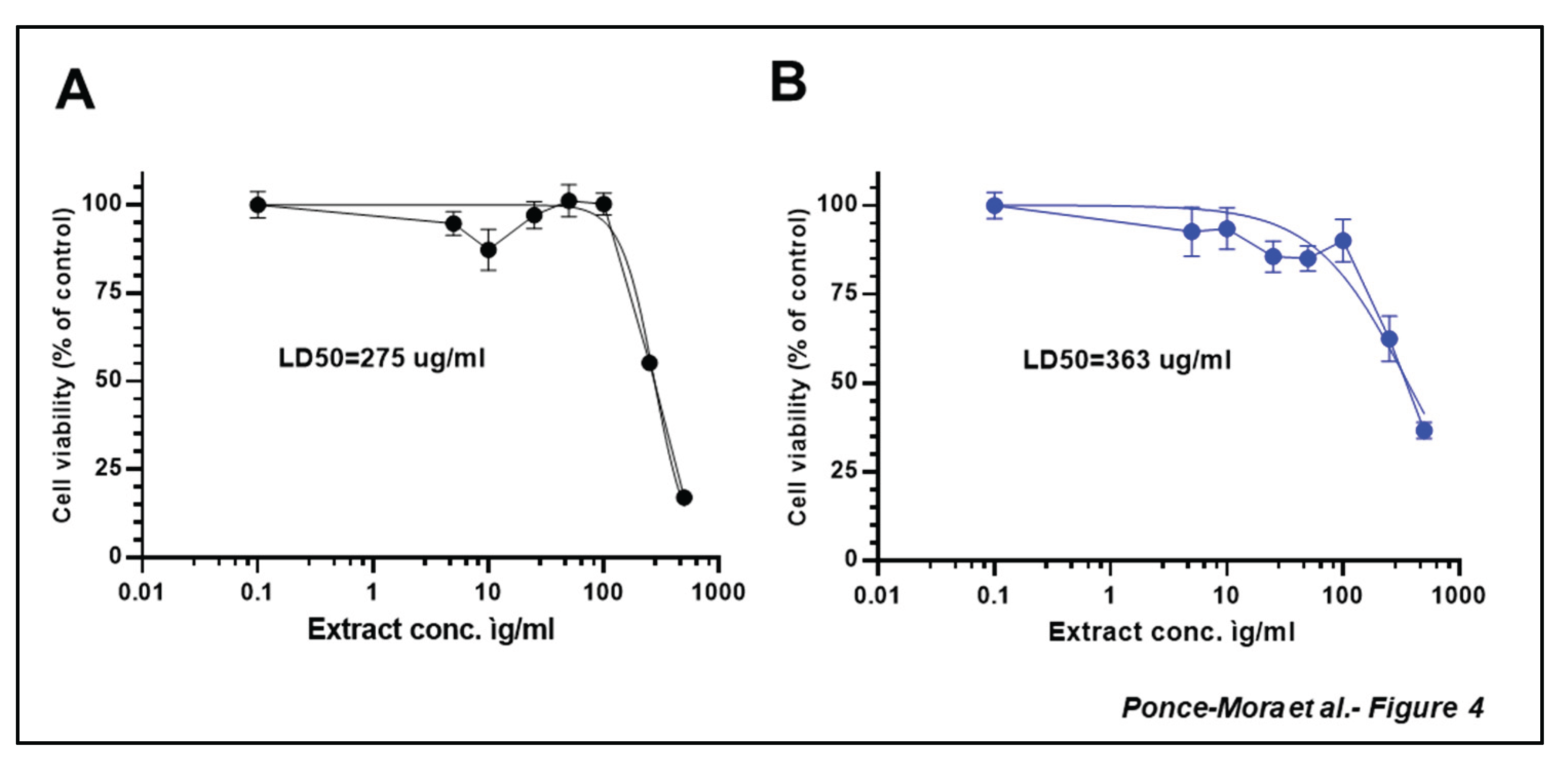
Figure 5.
Neuroprotection effect of leaves and bark extracts against H2O2-induced cytotoxicity in SH-SY5Y cells. Cell viability was analyzed using MTT assay. Extracts were administered at different concentrations for 24 h prior to H2O2 500 µM treatment for other 4 h. Data are presented as Mean ± SD of three independent experiments carried out by sextuplicate. # p < 0.001 vs. vehicle-treated control. *** p < 0.001 vs. only H2O2 treated cells.
Figure 5.
Neuroprotection effect of leaves and bark extracts against H2O2-induced cytotoxicity in SH-SY5Y cells. Cell viability was analyzed using MTT assay. Extracts were administered at different concentrations for 24 h prior to H2O2 500 µM treatment for other 4 h. Data are presented as Mean ± SD of three independent experiments carried out by sextuplicate. # p < 0.001 vs. vehicle-treated control. *** p < 0.001 vs. only H2O2 treated cells.
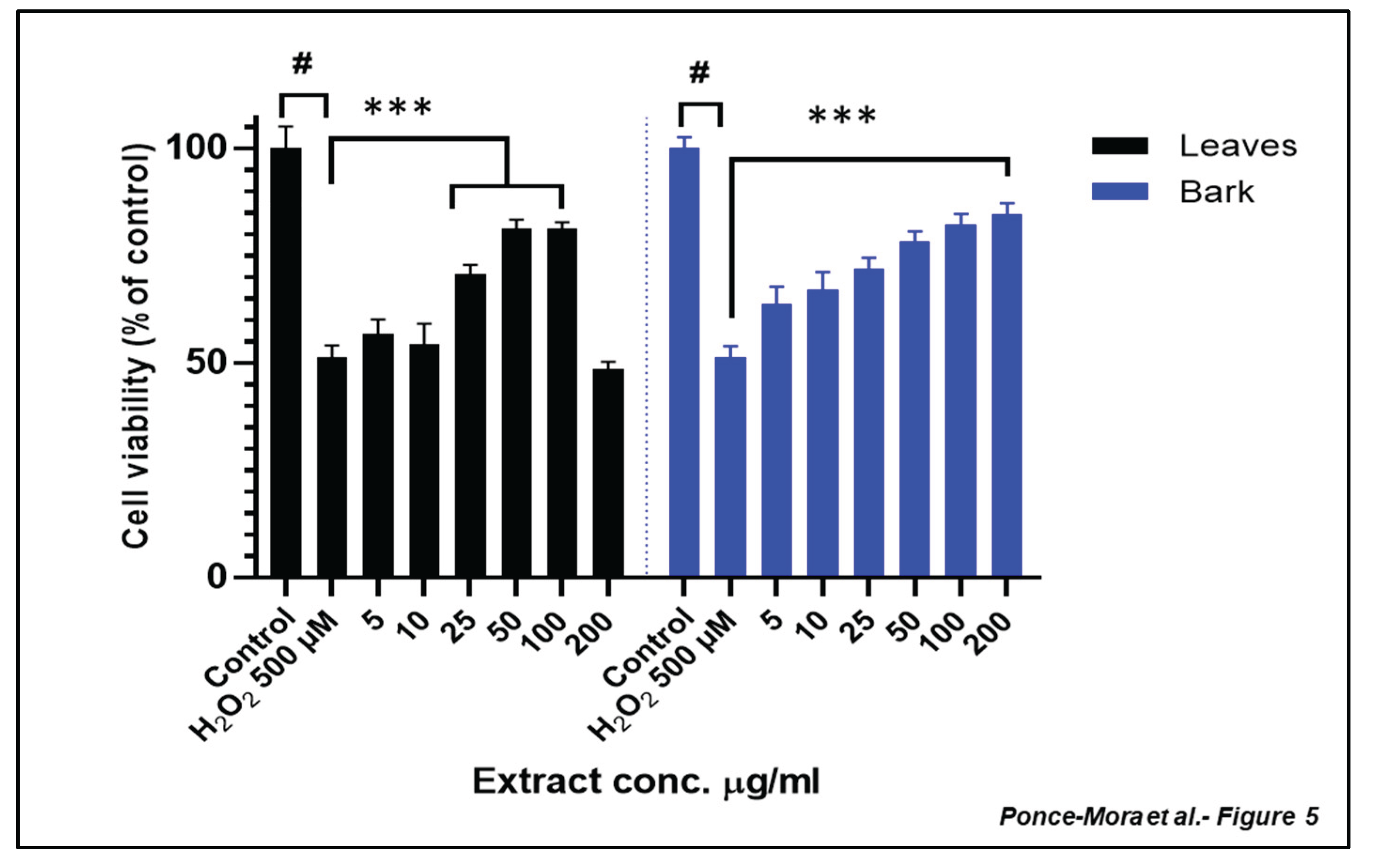
Figure 6.
Reduction of intracellular ROS production treated with leaf and bark extract against H2O2 induced cytotoxicity in SH-SY5Y cells. (A) Leaf and (B) bark extracts inhibit H2O2 intracellular ROS production. Cells were pre-treated with the extracts before exposure to 1mM H2O2 for 4 h, and intracellular ROS levels were determined. Each bar represents the relative intracellular ROS level expressed as percentage of control and significance of difference indicated with * (p < 0.05), **(p < 0.01), *** (p < 0.001), **** (p < 0.0001) when cells treated with extracts are compared to control and H2O2 vs. control cells.
Figure 6.
Reduction of intracellular ROS production treated with leaf and bark extract against H2O2 induced cytotoxicity in SH-SY5Y cells. (A) Leaf and (B) bark extracts inhibit H2O2 intracellular ROS production. Cells were pre-treated with the extracts before exposure to 1mM H2O2 for 4 h, and intracellular ROS levels were determined. Each bar represents the relative intracellular ROS level expressed as percentage of control and significance of difference indicated with * (p < 0.05), **(p < 0.01), *** (p < 0.001), **** (p < 0.0001) when cells treated with extracts are compared to control and H2O2 vs. control cells.
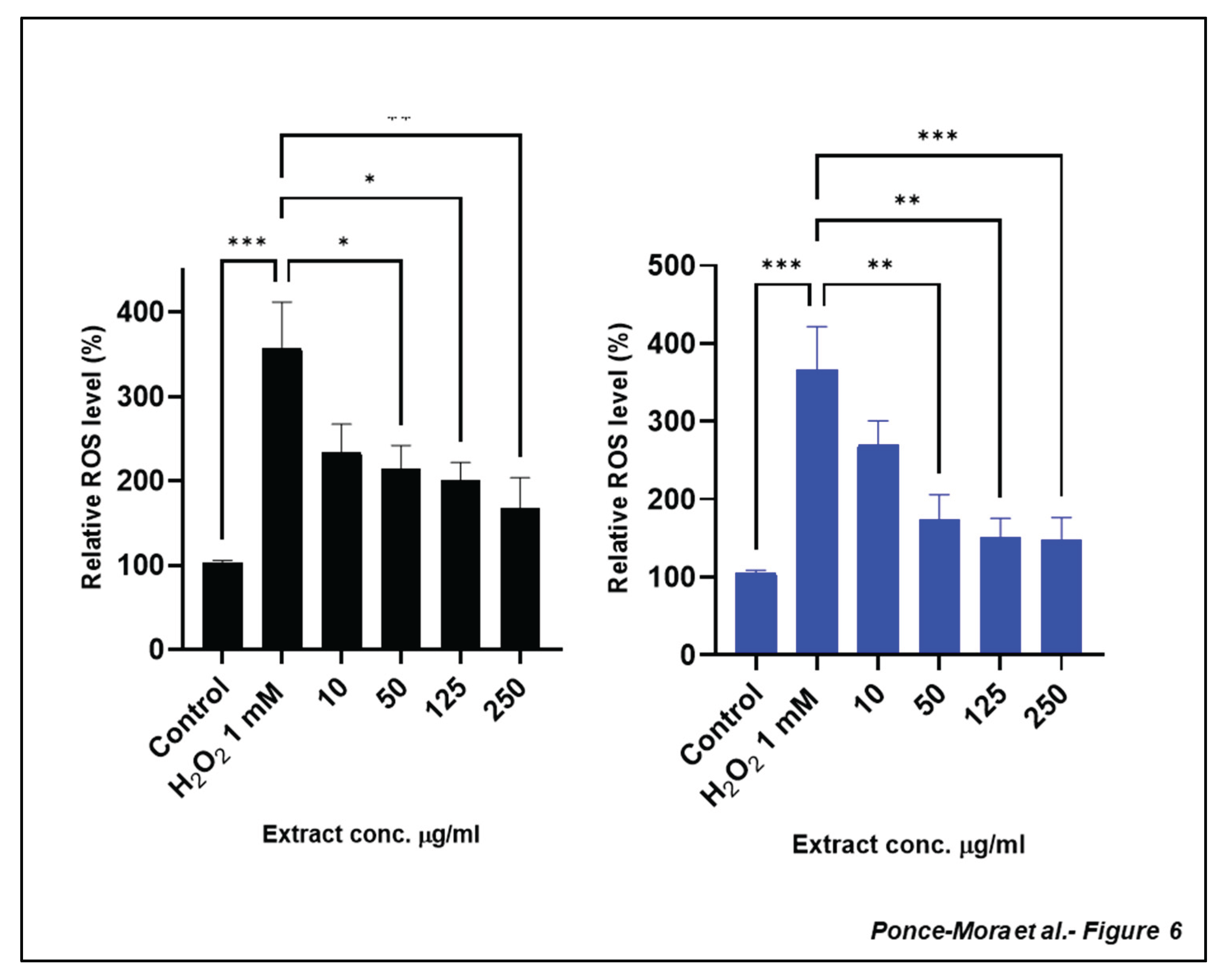
Figure 7.
MMP loss prevention by leaves and bark extracts in SH-SH5Y under exposure to H2O2. Cells were treated with extracts for 24 h prior to exposure to 250µM H2O2. MMP is expressed as the JC-1 fluorescence ratio in terms of red fluorescence to green fluorescence (Aggregates:Monomers). Data are presented as the Mean ± SEM of four independent experiments carried out in sextuplicate. # p < 0.001 vs. vehicle-treated control. **** p < 0.0001 vs. cells treated only with H2O2.
Figure 7.
MMP loss prevention by leaves and bark extracts in SH-SH5Y under exposure to H2O2. Cells were treated with extracts for 24 h prior to exposure to 250µM H2O2. MMP is expressed as the JC-1 fluorescence ratio in terms of red fluorescence to green fluorescence (Aggregates:Monomers). Data are presented as the Mean ± SEM of four independent experiments carried out in sextuplicate. # p < 0.001 vs. vehicle-treated control. **** p < 0.0001 vs. cells treated only with H2O2.
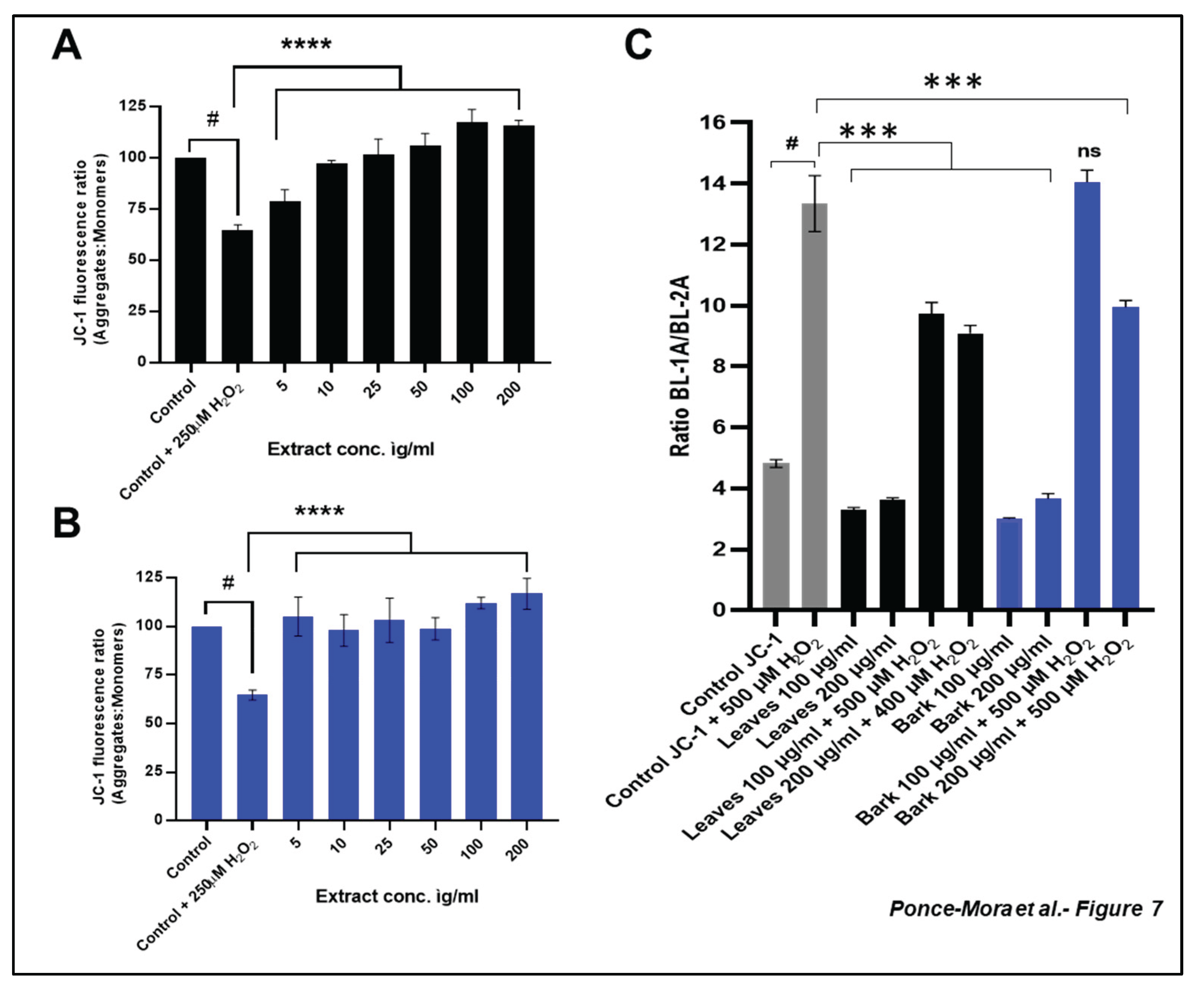
Figure 8.
Leaves and bark aqueous extracts of Anacardium occidentale trigger the expression of NRF2-targets genes. Changes in NRF2-targets genes were analyzed in human lens epithelial cells (HLECs) and mouse embryonic fibroblast (MEFs) treated for 24 hours with 1mg/ml of leaf (black), bark (blue), fruit (pink) and nut (green) aqueous extract. qPCR analysis was carried out against the indicated gene in (A-D) HLECs and (E-H) MEFs s. n >4; *, p < 0.05; *, p < 0.01, t test.
Figure 8.
Leaves and bark aqueous extracts of Anacardium occidentale trigger the expression of NRF2-targets genes. Changes in NRF2-targets genes were analyzed in human lens epithelial cells (HLECs) and mouse embryonic fibroblast (MEFs) treated for 24 hours with 1mg/ml of leaf (black), bark (blue), fruit (pink) and nut (green) aqueous extract. qPCR analysis was carried out against the indicated gene in (A-D) HLECs and (E-H) MEFs s. n >4; *, p < 0.05; *, p < 0.01, t test.
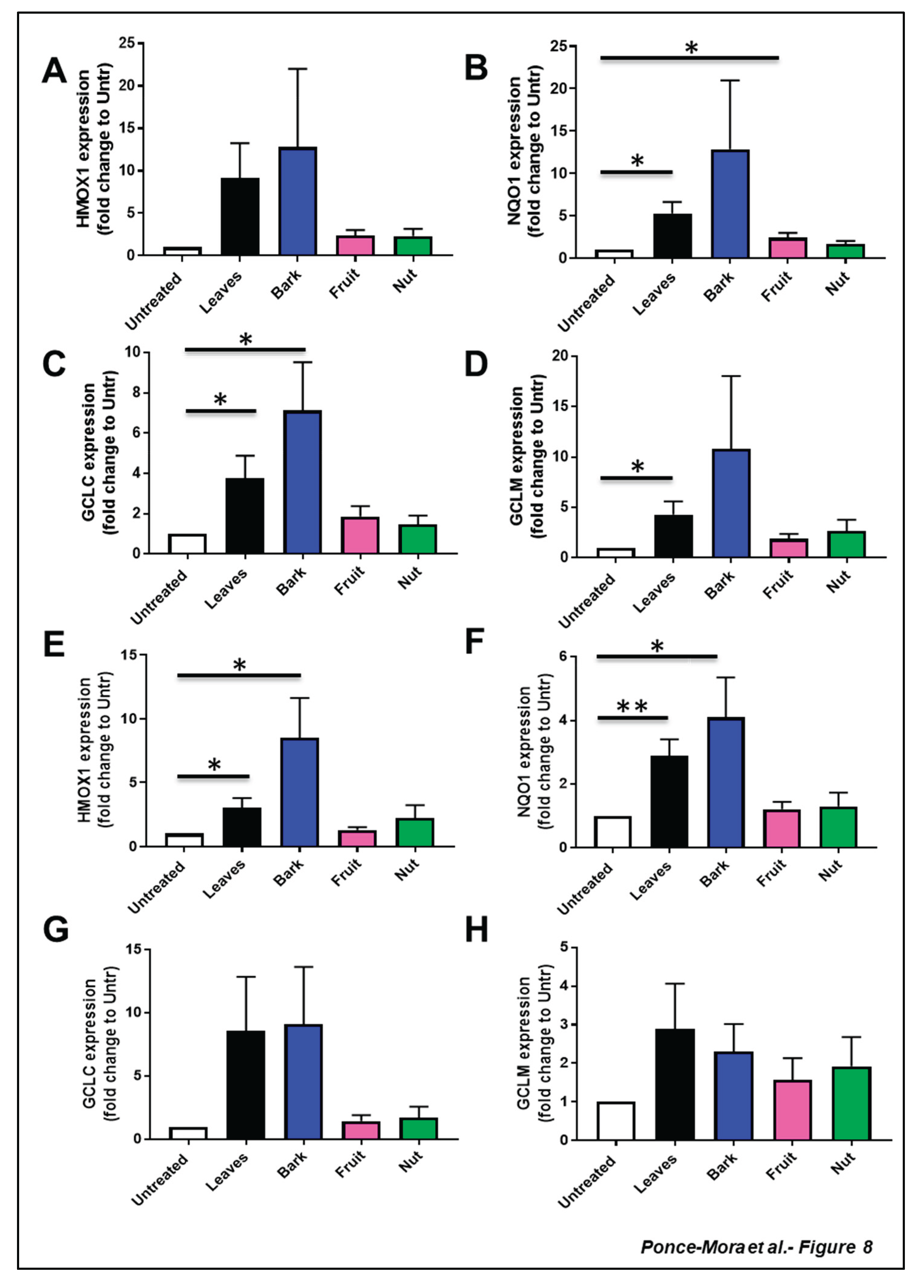
Figure 9.
Working model. Leaves and bark (but not fruit and nut) aqueous extracts of Anacardium occidentale have high ROS scavenger capacity and trigger the expression of cytoprotective genes related to NRF2 pathway.
Figure 9.
Working model. Leaves and bark (but not fruit and nut) aqueous extracts of Anacardium occidentale have high ROS scavenger capacity and trigger the expression of cytoprotective genes related to NRF2 pathway.
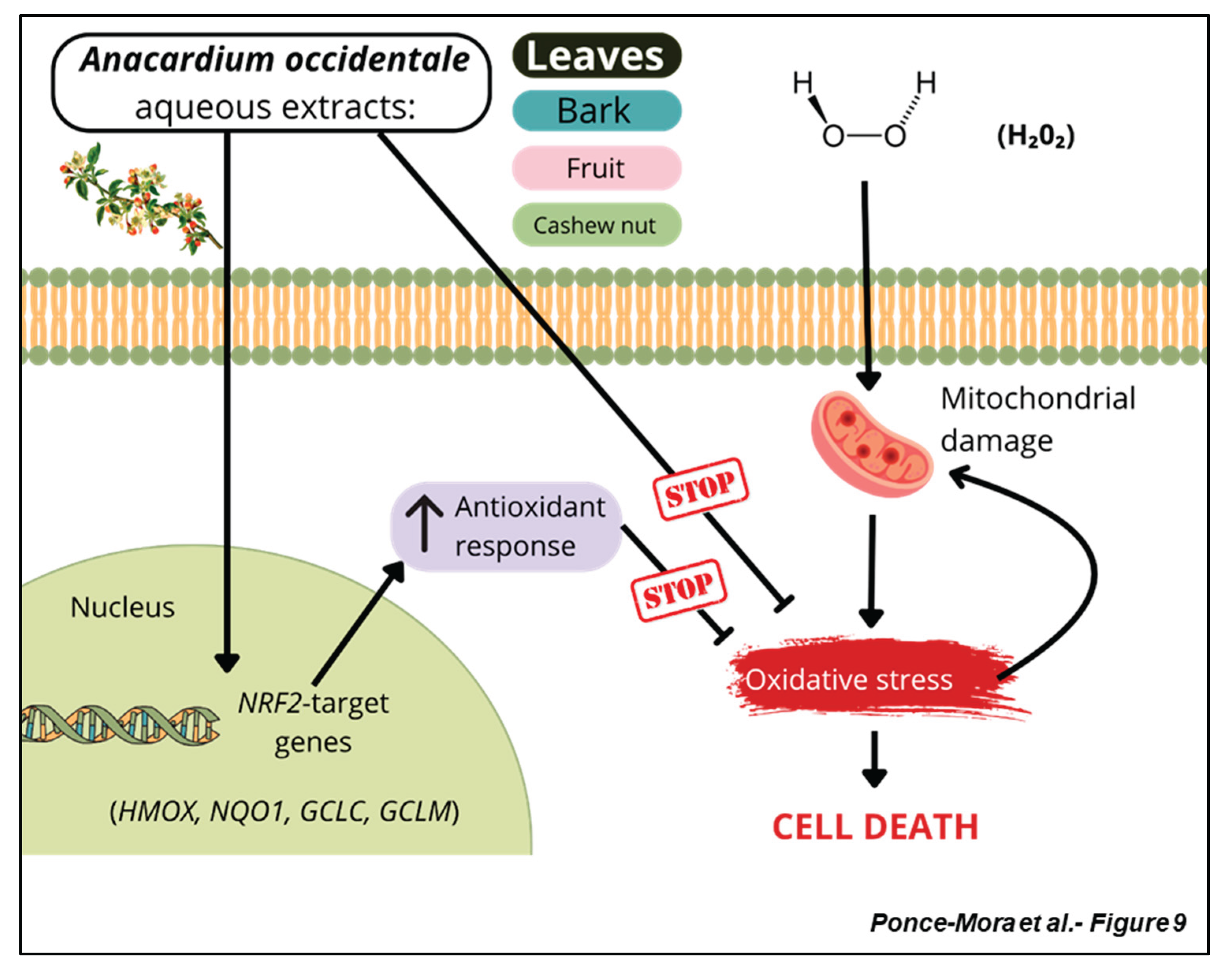

Table 1.
Primers used for quantitative real-time PCR assays.
| Gene | Sequence (5’-3’) | Length (bp) | Tm (oC) | GC content (%) | Species |
|---|---|---|---|---|---|
| Nrf2 FW | CAGAAGGAACAGGAGAAGGC | 20 | 55,3 | 55 | Mus musculus |
| Nrf2 REV | CTTGTTTGGGAATGTGGGC | 19 | 54,6 | 52,6 | Mus musculus |
| Hmox FW | TGCTCGAATGAACACTCTGG | 20 | 54,9 | 50 | Mus musculus |
| Hmox1 REV | TGGTCTTTGTGTTCCTCTGTC | 21 | 54,7 | 47,6 | Mus musculus |
| Gclm FW | ATGACCCGAAAGAACTGCTC | 20 | 55 | 50 | Mus musculus |
| Gclm REV | ATGATTCCCCTGCTCTTCAC | 20 | 54,6 | 50 | Mus musculus |
| Gclc FW | ACCATCACTTCATTCCCCAG | 20 | 54,6 | 50 | Mus musculus |
| Gclc REV | TTCTTGTTAGAGTACCGAAGCG | 22 | 54,5 | 45,5 | Mus musculus |
| Nqo1 FW | GGATTTGCCTACACATATGCTG | 22 | 53,9 | 45,5 | Mus musculus |
| Nqo1 REV | TGAATCGGCCAGAGAATGAC | 20 | 54,8 | 50 | Mus musculus |
| Gapdh FW | CCTGCTTCACCACCTTCTTGA | 21 | 57,2 | 52,4 | Mus musculus |
| Gapdh REV | TGTGTCCGTCGTGGATCTGA | 20 | 58,1 | 55 | Mus musculus |
| NRF2 FW | ATGACAATGAGGTTTCTTCGG | 21 | 52,9 | 42,9 | Homo sapiens |
| NRF2 REV | CAATGAAGACTGGGCTCTC | 19 | 52,9 | 52,6 | Homo sapiens |
| HMOX1 FW | AACTCCCTGGAGATGACTC | 19 | 53,3 | 52,6 | Homo sapiens |
| HMOX1 REV | CTCAAAGAGCTGGATGTTGAG | 21 | 53,4 | 47,6 | Homo sapiens |
| GCLM FW | GTTGACATGGCCTGTTCAG | 19 | 53,9 | 52,6 | Homo sapiens |
| GCLM REV | AACTCCATCTTCAATAGGAGGT | 22 | 53,1 | 40,9 | Homo sapiens |
| GCLC FW | AAGTGGATGTGGACACCAG | 19 | 54,7 | 52,6 | Homo sapiens |
| GCLC REV | CTGTCATTAGTTCTCCAGATGC | 22 | 53,1 | 45,5 | Homo sapiens |
| NQO1 FW | ACATCACAGGTAAACTGAAGG | 21 | 52,3 | 42,9 | Homo sapiens |
| NQO1 REV | TCAGATGGCCTTCTTTATAAGC | 22 | 52,5 | 40,9 | Homo sapiens |
| GAPDH FW | TCCTTCCTGGGCATGGAG | 18 | 56,9 | 61,1 | Homo sapiens |
| GAPDH REV | AGGAGGAGCAATGATCTTGATCTT | 24 | 55,8 | 41,7 | Homo sapiens |
Table 2.
Overview of the antioxidant parameters obtained by the ABTS, FRAP and DPPH assays.
| Sample | IC50 (µg/ml) (ABTS) | TEAC50 (ABTS) | Trolox equivalents (FRAP) | IC50 (DPPH) | |
|---|---|---|---|---|---|
| Trolox | 5.8 ± 1.3 | 1 | 1 | 5.62 | |
| Leaves | 12.3 ± 2.1 | 0.5 | 0.13 | 58.67 | |
| Bark | 13.8 ± 4.1 | 0.4 | 0.12 | 79.52 | |
| Fruit | 447.2 ± 15.0 | 0.01 | 0.03 | > 1000 | |
| Nut | >1000 | n.d | 0.01 | > 1000 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



