
Submitted:
30 June 2025
Posted:
30 June 2025
You are already at the latest version
Abstract
Agronomic effectiveness of biofertilizers is influenced by strain origin, genetic identity, crop genotype, soil type and environmental conditions. For best results both the plant and rhizobia strain must be adapted to the common harsh soil conditions. While plant varieties have changed over the years, complementary research on new strains effectiveness under varying soil fertility conditions has lagged in southern Africa. Seven field experiments were established in the main soybean producing areas of Zimbabwe in the north, central and north-east regions to evaluate agronomic benefits of new rhizobia strains against the current exotic commercial strain (MAR 1491). One site was irrigated (site 3) and the other six sites were rainfed (site 1, 2, 4, 5, 6, and 7). While trends in inoculation response varied from site to site due to site conditions, inoculation with the strains NAZ15, NAZ 25, and NAK128 consistently yielded high grain yields which were similar to the current commercial strain MAR 1491 and to application of mineral fertilizer (51.75 and 100 kg N ha-1). Grain yield levels were generally below 2 t ha-1 for site 2, 3, and 5, above 2t ha-1 for sites 1, 4, and 6, while for the irrigated site 3 they ranged upwards of 3 t ha-1. When irrigated all strains except NAK9 performed similarly in terms of grain yields and aboveground N uptake. We recommend further testing on the inclusion of the indigenous strains NAZ15, NAZ 25 and NAK128 in multi-strain commercial inoculant production targeting application in regions and soils where they excel beyond the current exotic strain MAR 1491.
Keywords:
Agricultural sustainability
; field trials
; low input farming systems
; microbial inoculants
; symbiotic nitrogen fixation
; tropical soils
1. Introduction
Production of soybean in smallholder farming areas of Zimbabwe was popularized in the nineties due to the activity and support of the Soybean Promotion Taskforce [1]. From 2020 to 2023, the average national hectarage dedicated to soybean production was 41,234 ha, producing on average 63,588 t of grain, with average national yields of 1.6 t ha-1 [2]. This production quantity represents only 29% the annual national requirement for food, animal feed and other industries (220,000 t; [3]). The average yield per hectare includes large scale commercial production but soybean yield in smallholder farming areas is usually much less than the 1.6 t ha-1 national average due to multiple factors which include rainfall, inherent soil fertility, use of fertilizers and inoculation. An average yield of 1.6 t ha-1 is however low compared to potential yields of above 5 t ha-1 under irrigation in tropical environments [4]. The soybean crop is a strategic choice for national, regional and global food and nutrition security, because of high protein (35-45%) and oil (15-25%) content. Soybean seeds also contain 23-33% carbohydrates, 4% fiber, and provide essential vitamins and minerals like potassium (K), magnesium (Mg), calcium (Ca), zinc (Zn), iron (Fe), and copper (Cu), along with antioxidants [5,6]. However, with rising population and the risks associated with climate change and variability, it is important to investigate strategies that improve soybean productivity as well as benefits realized from symbiotic N2 fixation.
The use of N2 fixing rhizobium inoculants is recommended to economically and sustainably supply the N needs of a soybean crop as opposed to using inorganic nitrogenous fertilizers. Benefits of inoculants include maintenance of high plant N supply due to symbiotic N2 fixation and high grain yields. Nitrogen fixation is affected by several factors, including the presence, abundance, and efficiency of rhizobia; the amount of nitrogen in the soil; the plant's genetic makeup and age; its interaction with rhizobia; and shifts in the soil’s physical and chemical properties [5,7]. Grain yield benefits could be constrained by harsh environmental conditions that affect growth of the host legume, the microsymbiont (rhizobia) and the bacteria/plant symbiotic relationship [8]. Before rhizobium can form nodules with soybean, it must survive adverse soil conditions such as moisture stress, soil acidity and high temperatures, known to reduce rhizobia survival in tropical soils [9,10]. Results from numerous field studies around the world show that increasing inoculant dosage could improve survival and therefore have greater yield impacts [11,12]. However, in terms of economics and agronomy, if available, providing a better suited rhizobium strain is preferred because it is more economical as opposed to providing a less suited strain in greater quantity to improve effectiveness.
In addition to the physical-environment limitations to rhizobium survival, poor soil fertility limits N2 fixation by limiting nutrient availability for both rhizobia in soil and a growing legume [13,14]. Earlier studies in Zimbabwe reported low support for rhizobia populations in soils due to poor water holding capacity and low fertility [15]. A more recent study by Chiduwa [16] reported similar lack of persistence in soil of the introduced MAR1491 commercial rhizobia strain. To be agronomically effective, rhizobia that is introduced through commercial inoculants must be adapted to the common harsh soil conditions which will improve survivability within the period from inoculation to successful nodule formation (nodulation). Rhizobia strain selection is found on environmental preference; the principle that certain strains are better suited for survival, nodulation and symbiotic N2 fixation in particular environments than others. A particular rhizobia’s ability to form nodules relates to plant yield response through nodulation intensity and expression of the Nod genes within the plant [17].
Despite the intensive soybean breeding for high yields and disease resistance that has taken place since the eighties in southern Africa, there is limited published research on isolation and selection of effective indigenous strains that could be more adapted to nodulation efficiency, effectiveness and N2 fixation under local conditions. To fill this gap, recently in South Africa, rhizobia strains have been isolated and evaluated for ability to nodulate with the various soybean cultivars [18]. However, the main limitation of the research reported was the lack of yield data from field trials.
In Germany, three indigenous Bradyrhizobium isolates, GMF14, GMM36, and GEM96) were tested in comparison to an exotic Bradyrhizobium strain (USDA110), in combination with different soybean cultivars, under greenhouse and field conditions. Regardless of soybean cultivar, inoculation with GEM96 resulted in the highest shoot N content of 21.89 g kg–1, which was significantly similar to the USDA110 strain (21.91 g kg–1). Additionally, the GEM96 strain promoted greater nodule formation, nodule mass, grain yield (1.38 t ha⁻¹), and protein content (412 g kg⁻¹), matching the performance of the USDA110 strain. These findings underscore the effectiveness of locally adapted Bradyrhizobium strains in enhancing soybean productivity [19]. A report on the higher competency of 4 native rhizobia isolates on soybean growth and nodulation, compared with commercial strains, under glasshouse conditions in Ethiopia, is also available [5].The presence of effective indigenous rhizobia populations, which can relate with soybean was confirmed in Zimbabwe [20]. However, there is lack of data on soybean yield response to inoculation with newly isolated rhizobia under different fertility conditions of Zimbabwe. This is despite the addition of newly isolated rhizobia strains to the Soil Productivity Research Laboratory (SPRL) Microbial Culture Bank through two parallel isolation studies under the N2 Africa project (https://n2Africa.org). Currently soybean rhizobia inoculants in Zimbabwe are produced from a Bradyrhizobium diazoefficiens USDA110 strain (MAR 1491) originally isolated over sixty-five years ago in Florida, USA.
The objectives of this study were to evaluate the effect of inoculation with recently isolated rhizobia strains on soybean grain yield and to assess how different soil fertility conditions influence the inoculation response.
2. Materials and Methods
2.1. Experimental Sites and Rhizobia Strain Details
Field experiments were established in north, central and north-east regions of Zimbabwe (Figure 1), which collectively account for approximately 94% of the total annual soybean production of Zimbabwe. Sites 1 – 3 were located at the Soil Productivity Research Laboratory (SPRL), Kushinga Agricultural College (KPC), and the Horticultural Research Institute (HRI), all situated in Marondera district and classified as agro-ecological region 2a. This region is characterized by a mean annual rainfall of between 750 – 1000 mm with mean maximum annual temperatures that range 23 – 27oC. This region constitutes the most agriculturally productive region in the country, second only to agroecological region I. Sites 4, 5, 6, and 7 were in Wedza, Makoni, Mhondoro and Chinhoyi districts respectively, and represent communal farming areas. Sites 6 and 7 are classified as agro-ecological region IIb (750 – 1000mm annual rainfall, 25 – 28o C maximum temperatures), while sites 5 and 4 are classified as region III (650 – 800 mm annual rainfall, 25 – 28o C maximum temperatures) and IV (450 – 650 mm annual rainfall, 27 – 29o C maximum temperatures), respectively. All experimental sites were rainfed to mimic smallholder farmer conditions, except for site 3 (HRI) were drip irrigation was installed to supplement rain during the trial duration. Site 3 represented more commercial and resource endowed farmers.
The treatments consisted of 6 rhizobia strains, 5 of which had been isolated in an earlier phase of the N2 Africa project, a positive control with inorganic N fertilizer (at 51.75 or 100 kg N ha-1 for rainfed and irrigated conditions respectively) and a negative control without any N fertilizer or inoculant applied. Rhizobia strains NAZ 15, 21, and 25 were isolated in 2011 from Goromonzi, Wedza and Murehwa soils in Zimbabwe (and later deposited in the SPRL Microbial Culture Bank and renamed MAR 1667, 1673, and 1677 respectively). Rhizobia strains NAK 9 and 128 were isolated in Kenya within the same project. Rhizobium strain MAR1491 is the current commercial strain for soybean rhizobia inoculants in Zimbabwe, originally isolated in United States of America (USDA 110) and served as a reference strain.
Each strain was cultured in a standardized mother culture solution and mixed with a uniform carrier material, following the SPRL inoculation batch preparation protocol. This facilitated random quality control and quality assurance protocols for purity and rhizobial population analysis. Results from serial dilutions and plating on Yeast extract mannitol (YEM) agar plates revealed that each inoculant sachet had at least 109 viable rhizobial cells g-1 of carrier media and no other identified species.
2.2. Trial Establishment and Setup
At all the farmer sites (sites 4-7) land was tilled using ox-drawn ploughs while tractors were used for the on-station sites (1-3). A basal dressing of single super phosphate (SSP) fertilizer was applied to all plots at a rate of 200 kg ha-1 (19% P2O5) to supply 16.6 kg P ha-1. Trials were arranged in a randomized complete block design (RCBD) with eight treatments and 4 replicates with blocking against the slope at all sites. Each treatment plot measured 3 x 4 m2. Inoculants with specific strains were applied as a slurry to soybean seeds using clean and sanitized implements treated with 70% ethanol. At each site, inoculated seeds were immediately planted, one treatment at a time to avoid rhizobia cross contamination. Soybean was planted with an inter-row spacing of 0.3 m x 0.1 m within row. Two seeds per station were planted and later thinned to one plant per station. The -N treatment control plots did not receive any N fertilizer from inoculant or a mineral form. At the rainfed sites (1, 2, 4-7) the +N treatment received ammonium nitrate as a top-dressing fertilizer at a rate of 150 kg ha-1 (51.75 kg N ha-1), which was split applied into two equal applications at 7 and 30 days after planting. At the irrigated site 3, the rate of N was doubled to 100 kg N ha-1 and similarly split applied as at other sites. All the plots at all the sites were routinely weeded as needed using hand hoes throughout the experiment period. Implements were sanitized between treatments. Application rates of N and P employed in this study were typical of what most smallholder farmers would apply as general recommendation rates for Zimbabwe based on soybean removal rates of 60kg N ha-1 and 15.3 kg P ha-1 from soil [21].
2.3. Soil Sampling and Soil Chemical Analysis
At all sites, soil samples were collected for baseline soil fertility analyses. Composite samples were derived from ten subsamples (0 - 15 cm depth) before the establishment of the experiments. The composite samples were analyzed for pH, organic carbon (OC), mineral nitrogen (N), available phosphorus (P) and exchangeable potassium, calcium and magnesium (K, Ca, and Mg) [22]. Mineral N was measured as ammonium and nitrate nitrogen after extraction in 2M KCl (10 g soil :100 ml) for 1 hour. Available P was determined colorimetrically [23] after extraction using the resin bag extraction method [24]. Exchangeable cations were determined after extraction in 1M NH4OAc for 30 mins followed by filtering, and Mg content was read on an atomic absorption spectrophotometer, while K and Ca were read on a flame photometer [24].
2.4. Plant Material Sampling and Analysis
Due to logistical constraints, nodulation and biomass assessments were performed only for the sites 1-3 at the flowering stage, N uptake and estimation of N2 fixation was done for only the irrigated site 3, and grain yield and grain N content was assessed for all sites. For sites 1 and 2, an average of three plants were randomly sampled per plot. For site 3, ten plants per plot were randomly sampled. Whole plants were dug up carefully to preserve the roots and the above ground biomass was separated from the roots at the base of the stem. The roots were washed over of a 1 mm sieve, blot dried using clean paper towels, then root nodules were removed and counted. The fresh mass of the nodules was thereafter recorded. The above-ground plant biomass was dried to a constant mass in an oven at 70°C and recorded.
At all sites, grain yields were determined after harvesting pods at physiological maturity from a net plot of 1m2 avoiding the plot borders by at least a meter. The pods were dried (to a constant mass), shelled and the grain was weighed. Plant and grain samples were ground and analyzed for total N using the micro-Kjeldahl technique [22]. A 0.2 g plant sample was digested with concentrated H2SO4 (98%) in a Gerhadt Kjeldatherm® digester, then distilled in 50% NaOH with N collection in a boric acid mixture then titrated with 0.07 M H2SO4. Nitrogen uptake in grain was calculated by multiplying the %N content with the total grain mass.
For the irrigated site 3, nitrogen uptake was calculated by multiplying the average %N content in all aboveground plant parts with the total aboveground mass (including grain). Nitrogen derived from fixation (Dff) was estimated by subtracting the average N uptake of the uninoculated control from the N uptake in the inoculated treatments. The assumptions of this estimation were that the uninoculated control plants had poor nodulation and ineffective nodules. The N uptake from uninoculated plants was assumed to be mainly from available nitrogen in the soil and represented on average N uptake from soil by inoculated plants assuming that all plants had similar root morphology and soil N uptake [25].
2.5. Statistical Analysis
All statistical analysis was performed in IBM SPSS software (Version 29.0.2.0 (20)). Data on biomass, nodule counts and mass, total grain N uptake, grain yields were assessed for normality using the Shapiro-Wilk test and homogeneity of variance was assessed using the Levene statistic. For normally distributed data, a one-way ANOVA was run and where there were significant differences, Tukey HSD was used to separate significantly different means at 95% confidence interval. Generally, nodule counts, and nodule mass data were not normally distributed, and the independent samples Kruskal-Wallis test was employed to assess significance with pairwise separation of means thereafter. For normally distributed data with unequal variances (grain yield: site 4, grain P uptake: site 3), the Brown-Forsythe robust test of equality of means was used with mean separation using the Games-Howell statistic.
3. Results
3.1. Study Site Soil Characteristics
Soil fertility characteristics varied substantially across the experimental sites (Table 1). Soils at the study sites ranged from slightly acidic to acidic, apart from site 7 that was neutral on the CaCl2 scale (Table 1). Sites 1 and 4 had the most infertile soils that had very low organic carbon content at 0.35 and 0.23% soil OC, respectively. Mineral N ranged from 5 to 44 ppm with Wedza recording the lowest. Available P was also very low at the same two sites, however, sites 5 and 6 were acutely deficient in available P. Only site 2 had high available P at 37 ppm. Exchangeable K levels were low at site 1, site 4, and site 5.
3.2. Nodulation Counts and Nodule Mass Soybean During Flowering at the Researcher Managed Sites
The +AN and -N treatments had negligible nodule counts and fresh nodule mass at the rainfed sites (1 and 2), while at the irrigated site (3), they had fresh nodule mass that was comparable to that of the NAZ 21 and NAK 9 treatments. The strains NAZ 15, NAK 128 and MAR 1491 had significantly higher nodule counts and nodule mass than the uninoculated +AN and -N treatments, at all the three sites (Table 2). The NAZ 25 and NAK 9 treatments had comparable nodule counts to the +AN and -N treatments across the same three sites. Although the strain NAZ 25 had lower nodule counts than the NAZ 15, NAK 128 and MAR 1491 treatments, at the three sites, it had a considerable nodule mass which was comparable to that of the former treatments at all sites except at site 1 and for NAZ15.
3.3. Biomass Production of Soybean During Flowering at the Researcher Managed Sites
Biomass production during flowering was significantly influenced by treatment (P < 0.05) at the rain-fed sites 1 and 2 (Figure 2a,b) but not at the irrigated site 3 (P = 0.715) (Figure 2c). The rhizobia strains NAZ 15, NAK9, NAK128 produced soybean biomass yields that were similar to that of the commercial strain MAR1491 at site 1 (Figure 2a). At the same site, NAK128 and MAR1491 produced significantly higher biomass than NAZ21, NAZ25, +AN, and -N treatments. At site 2, while NAK128 produced the highest biomass, it was similar to the NAZ15, NAZ 21, NAZ 25, and NAK 9 treatments (Figure 2b). At both sites: 1 and 2, the +N treatment was not significantly different to the -N treatment and generally both treatments produced very low biomass compared to the rest of the inoculated treatments.
3.4. Soybean Grain Yields at All Sites
Grain yields across all the treatments were < 2 t ha-1 at site 1, site 4 and site 6, while only site 3 and site 5 had the +N and all rhizobium inoculated treatments yielding > 2 t ha-1 of grain (Table 3). Inoculation or fertilization did not significantly affect (P = 0.57) soybean grain yields at site 6, while there were significant differences (P < 0.05) at the other sites. Inoculation with the strains NAZ15, NAZ 25, and NAK128 consistently resulted in significantly (P< 0.05) higher grain yields than the -N treatment at sites 1 - 5 except for NAK128 at site 3. At the same time, the treatments were comparable to the commercial strain MAR 1491 and the +AN treatments. Plants inoculated with NAK 9 did not show significant grain yield advantage compared to the -N treatment at sites 1, 2, 3, and 7. At site 7 and out of all the treatments, only the NAZ 15 and NAK 128 produced significantly less (P = 0.001) grain yields than the commercial strain MAR1491 (Table 3). Grain yields of soybean that received AN fertilizer were not significantly different from the -N control at sites 1, 4, 6, and 7.
Generally, a near similar trend to that of grain yields was observed for grain N uptake (Table 3). Inoculation or fertilization did not significantly (P = 0.068) affect soybean grain N uptake at site 6. At sites 1 - 3, and 5, The +AN treatment plots had significantly higher grain N uptake than the -N control. At the same sites, the +AN treatment plots were not significantly different in grain N uptake to the NAZ 15, NAZ 21, NAZ 25, NAK 128, and MAR 1491 treatment plots. At site 4, the NAZ 15, NAZ 21, and NAZ 25 treatment plots yielded significantly (P < 0.001) higher (at least 67% more) grain N uptake than the +AN treatment. Correlation analysis of pooled data from rainfed sites revealed that there was significant (P < 0.01) positive correlation between grain yields and soil organic matter, initial available N, and exchangeable Ca and Mg (Figure 3). The same trend was true for grain N uptake and the soil properties.
3.5. Nitrogen Uptake and Estimated Nitrogen Derived from Fixation at Site 3
While biomass production was not significantly different across treatments during the flowering stage at site 3 (Figure 2c), there were significant differences between treatments at the physiological maturity stage. The range of aboveground biomass ranged from 4.1 to 5.5 t ha-1 for the fertilized / inoculated treatments. The NAZ21, NAZ25, NAK128, MAR 1491, and +N treatments produced significantly more (roughly double) aboveground biomass than the -N treatment (Figure 4a). Except for NAK128, all inoculated treatments and the +N treatment had comparable shoot N uptake (Figure 4b). The NAZ21 and +AN treatments had the highest N uptake which was significantly different to that of the NAK128 and -N treatments. Estimated nitrogen derived from symbiotic N2 fixation followed a similar trend to N uptake, the NAZ21 treatment had the highest amount of N2 derived from fixation which was comparable to the rest of the inoculated treatments except for the NAK 128 treatment.
4. Discussion
The near zero nodule counts observed in the un-inoculated treatments at site 1 and 2 suggest that indigenous soybean-nodulating Bradyrhizobia populations were very low in the soils. This is because the soils were acidic, had low OC content and undergo periods of low soil moisture, all which limits microbial survival [10]. However, at site 3, the +N and -N treatments had nodulation counts and nodule mass that was comparable to that of NAZ21, NAZ25. Partly a reason for this could be that the site is regularly irrigated which reduces soil moisture stress on soil microbes. In other part, the increased productivity of irrigated land compared to non-irrigated could lead to more microbial diversity and populations due to increased C input in the form of plant roots and residues. Land management significantly affects rhizobia survival and diversity and activities that lead to increased soil OC such as manure and biochar application are reported to positively affect rhizobia abundance [15,26].
In soils such as site 1 and 2, inoculation response can be very high at low soil N conditions, since competition with native strains will be minimal. Significantly higher nodule counts in inoculated treatments suggest that all the strains tested were very effective at forming nodules with soybean. Though the strain NAZ 25 did not necessarily produce high nodule counts at site 1, it had considerable nodule mass at the three sites, indicating its potential as an efficient N2 fixing strain. Due to the high energy requirements of N2 fixation, the number of nodules is largely regulated by the host plant, through a pathway known as autoregulation of nodulation (AON) [27]. Environmental conditions such as soil nutrient deficiency that negatively affect photosynthesis may therefore lead to greater nodulation restrictions by the plant. This happens by limiting the plant’s carbon resources allocated to the roots [28]. This AON is likely the reason that the same Bradyrhizobium strain differently nodulated the same host in different environments. At the same time, while nodule count is a good indicator of potential N2 fixation, other studies have shown that taproot based instead of total nodule counts is a much better indicator of potential N2 fixation [29].
The low biomass observed in the -N controls show that lack of an N source whether through rhizobia symbiosis or mineral fertilizer will result in limited vegetative growth of soybean. Most soils in the smallholder areas are inherently deficient in nitrogen or depleted through extractive production methods [10]. At the national general recommendation rates for nitrogen employed in this study, increases in soybean grain after application of 57.5 kg N ha-1 (rainfed sites) or 100 kg N ha-1 (irrigated site) were comparable to most of the inoculated treatments. This confirms that inoculants can indeed replace mineral N fertilizer in these cropping systems. Despite the high need for N under such N limiting systems, rhizobia being supported by host plants are likely to adjust to meet high plant N requirements.
Soybean grain yields at the rain fed sites were within the range of reported Zimbabwe national average of 1.6 t ha-1 [30]. A soybean yield gap decomposition highlighted multiple factors driving suboptimal yields in similar smallholder farm sites in Malawi, Mozambique and Zambia, including crop management, and confirming genotype x environment and microbe (G x E x M) interaction effects [31]. The overall low grain yields are majorly attributable to poor soil fertility limiting crop productivity and to some extent, the erratic and low rainfall that characterizes some of these areas. For example, site 4 had the poorest soil fertility, with very low OC, mineral N, K and available P, which led to the lowest grain yields across all the treatments (< 1.1 t ha-1). In addition, reduced rainwater contributed to the low yields. At this site soybean exhibited yield non-responsiveness to the addition of ammonium nitrate fertilizer.
The low soil OC content at sites (0.23 – 1.35%) reported in this study are in line with those reported by Manzeke-Kangara et al. [32] of 0.4 - 1.5% in Southern Africa and those reported by Nyawasha et al. [33] of < 0.4% OC in North East Zimbabwean smallholder farming areas. Low soil OC levels have been shown to result in less efficient N2 fixation in inoculated Trifolium repens grown in South Africa [34]. Some studies have highlighted the synergic response on N, P, and OC showing that balanced fertilization is critical in increasing N-use efficiency in such low fertility soils [35,36]. Except for NAK9 inoculated plots, grain yields from inoculated or +N plots at the irrigated site 3 were within range of expected commercial yield level of 3.5 t ha-1.
The trend in soybean grain yields with inoculation strain type was different at each site suggesting that strain performance was influenced by site specific factors, including rainfall and soil fertility. It is logistically impractical to tailor the strain used in commercial inoculant production for each smallholder setting, and selection of a strain that is consistent across the ecological zones in Zimbabwe is therefore essential. While inoculation with NAZ15, NAZ 25, and NAK128 showed consistently good soybean grain yields on average, their performance was similar to the current commercial strain (MAR 1491). This implies that there is limited justification to update the commercial inoculant strain solely on the basis of N2 fixation. Strain selection should consider additional traits such as ecological fitness which were not assessed in this work. Additionally, introducing a multi-strain inoculant with at least two strains (e.g. NAZ25 and MAR1491) may provide better agronomic efficiency of seed inoculation in different areas of Zimbabwe. The MAR 1491 (USDA 110) strain is used as a commercial strain in several African countries for example in Kenya [37] and Zimbabwe. The lack of differences between grain yields of inoculated soybean and the +N control implies that there could be no need to apply inorganic fertilizer when soybean has been inoculated with an efficient N2 fixing rhizobium strain. At the same time, inoculation may yield better yield outcomes than inorganic N addition under severely soil fertility limiting conditions, as observed for site 4 where inoculated treatments produced double the grain yield and grain N uptake (Table 3). The use of general N and P rates was a limitation in this study, however the data generated is more realistic to current farmer rates. While further studies should employ higher rates that are in line with soil nutrient deficiencies with proper correction of soil pH, a key limitation is water availability. Higher inorganic fertilizer rates increase chances of crop failure by osmotic stress when water is limited.
Soil pH and available P were surprisingly not correlated with grain yield and grain N uptake for the rain fed sites indicating that when P was supplied at 16.6 kg P ha-1 in the tested soils, P supply was not a major bottleneck to crop productivity. However, this does not imply that soil P was not a limiting factor because total plant P uptake ranges 15 – 20 kg P ha-1 for a soybean grain yield of 2.5 t ha-1 [38] and most of the sites were deficient in P (Table 1). A low rate of P supply (16.6 kg P ha-1) was used because it represents the ‘general’ recommendation for soybean and is realistic to what most smallholder farmers would apply without a soil test. This general recommendation is based on a soybean removal rates of 60kg N ha-1, 15.3 kg P ha-1, and 66 kg K ha-1 from soil [21]. Soil OC, initial available N, and exchangeable Ca and Mg were all positively correlated with grain yields. Of all the tested soil attributes, soil OC had the highest moderately strong relationship with grain yield (Spearman coefficient: 0.645). Similarly in an Africa wide study by Kebonye et al. [39] plant biomass and soil OC were found to be highly correlated, explained by soil OC creating favorable conditions for nutrient uptake and releasing nutrients through mineralization. Our results highlight the importance of soil OC management and improvement in symbiotic N2 fixing systems which could result in increased inoculation response.
The high grain yields (2.9 – 3.3 t ha-1) and similarity of grain yields in inoculated treatments at the irrigated site 3 (except for the NAK 9 treatment) shows that these strains have high inoculation response when water is not limiting. The reduction/elimination of soil moisture stress during the critical root nodulation stage enhances nodulation success by improving inoculant strain survival. The relatively high nodule numbers and fresh nodule mass per plant of inoculated plants corroborates this. All inoculated treatments at site 3 had an N uptake in aboveground biomass that was significantly higher than that of the -N treatment indicating that there was indeed increased N supply due to inoculation. Estimates of Ndff suggest that the NAZ21 and NAZ25 strains may have superior to MAR1491 N2 fixing ability under irrigated conditions.
5. Conclusions
We concluded that some strains native to Zimbabwe (NAZ15, NAZ 25) can enhance nodulation capacity, biomass N uptake and grain yield of soybean, comparable to the exotic strain, therefore recommend inclusion of NAZ15, NAZ 25 and NAK128 in commercial multi-strain inoculants. Introducing a multi-strain inoculant may provide better agronomic efficiency of seed inoculation in different areas of Zimbabwe and further testing is recommended. Results from this study also revealed that while type of strain is important, inoculation response is influenced by the soil fertility profiles in different parts of Zimbabwe. There is need to couple soil organic and nutrient management with inoculants for improved N2 fixation. Further studies of inoculation coupled with biochar addition to soils with very low levels of soil organic matter are recommended.
Author Contributions
Conceptualization, A.T., C.S.M., and M.S.C.; methodology, A.T., G.K., C.M., C.S.M., T.P.T. and M.S.C.; validation, A.T., G.K., C.M., C.S.M., and M.S.C.; formal analysis, A.T.; investigation, A.T., G.K., C.M., C.S.M.; resources, A.T. and M.S.C.; data curation, A.T. and G.K.; writing—original draft preparation, A.T.; writing—review and editing, A.T., G.K., C.M., C.S.M., T.P.T. and M.S.C.; visualization, A.T.; supervision, A.T.; project administration, A.T., G.K. and M.S.C.; All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded in part by the Bill & Melinda Gates Foundation through Grant OPP1020032 to Wageningen University (N2Africa: Putting Nitrogen Fixation to Work for Smallholder Farmers in Africa (www.N2Africa.org)) and by the Government of Zimbabwe through the Chemistry and Soil Research Institute.
Data Availability Statement
Data reported in this manuscript is available upon request from the authors.
Acknowledgments
We acknowledge the support given by staff at Soil Productivity Research Laboratory in establishing field trials and collecting data. We thank management at Horticulture Research Institute and at Kushinga P. Agricultural College for allowing us to use their facilities and land for trials. We also thank the farmers who allowed us to establish our trials on their land.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Abbreviations
The following abbreviations are used in this manuscript:
| SPRL | Soil Productivity Research Laboratory |
| CSRI | Chemistry and Soil Research Institute |
| HRI | Horticultural Research Institute |
| KPC | Kushinga Phikelela Agricultural College |
| SNF | Symbiotic Nitrogen Fixation |
| OC | Organic Carbon |
References
- Giller, K.E. The successful intensification of smallholder farming in Zimbabwe. LEISA Magazine 2008, 24.2, 30–31. [Google Scholar]
- FAOSTAT. Crops and livestock products. Food and Agriculture Organization of the United Nations: 2025.
- Basera, J. Hapson. Soyabean: A strategic crop. In Field Crops, Seed Co Zimbabwe: Harare, Zimbabwe, 2025.
- Zambon, L.M.; Umburanas, R.C.; Schwerz, F.; Sousa, J.B.; Barbosa, E.S.T.; Inoue, L.P.; Dourado-Neto, D.; Reichardt, K. Nitrogen balance and gap of a high yield tropical soybean crop under irrigation. Front Plant Sci 2023, 14, 1233772. [Google Scholar] [CrossRef] [PubMed]
- Desta, M.; Akuma, A.; Minay, M.; Yusuf, Z.; Baye, K. Effects of Indigenous and Commercial Rhizobia on Growth and Nodulation of Soybean (Glycine max L) under Greenhouse Condition. The Open Biotechnology Journal 2023, 17. [Google Scholar] [CrossRef]
- Sharma, S.; Kaur, M.; Goyal, R.; Gill, B.S. Physical characteristics and nutritional composition of some new soybean (Glycine max (L.) Merrill) genotypes. J Food Sci Technol 2014, 51, 551–557. [Google Scholar] [CrossRef]
- Nzeyimana, F.; Onwonga, R.N.; Ayuke, F.O.; Chemining'wa, G.N.; Nabahungu, N.L.; Bigirimana, J.; Noella Josiane, U.K. Determination of abundance and symbiotic effectiveness of native rhizobia nodulating soybean and other legumes in Rwanda. Plant Environ Interact 2024, 5, e10138. [Google Scholar] [CrossRef]
- Romanova, I.; Syromiatnykov, Y.; Yakovlieva, A.; Voinash, S.; Orekhovskaya, A.; Vanzha, V.; Akhtyamova, L.; Vashchilin, V.; Arabov, F.; Mehdiyeva, A.; et al. Efficiency of symbiosis between Bradyrhizobium bacteria and soybean plants under various biopreparations. BIO Web of Conferences 2025, 181. [Google Scholar] [CrossRef]
- Hungriaa, M.; Vargas, M.A.T. Environmental factors affecting N2 fixation in grain legumes in the tropics, with an emphasis on Brazil. Field Crops Research 2000, 65, 151–164. [Google Scholar] [CrossRef]
- Tumbure, A.; Dube, S.; Tauro, T.P. Insights of Microbial Inoculants in Complementing Organic Soil Fertility Management in African Smallholder Farming Systems. In Towards Sustainable Food Production in Africa, 2023. 59–83. [CrossRef]
- Jarecki, W.; Borza, I.M.; Rosan, C.A.; Vicas, S.I.; Domuța, C.G. Soybean Response to Seed Inoculation with Bradyrhizobium japonicum and/or Nitrogen Fertilization. Agriculture 2024, 14. [Google Scholar] [CrossRef]
- Kanonge-Mafaune, G.; Chiduwa, M.S.; Chikwari, E.; Pisa, C. Evaluating the effect of increased rates of rhizobial inoculation on grain legume productivity. Symbiosis 2018, 75, 217–227. [Google Scholar] [CrossRef]
- Chekanai, V.; Chikowo, R.; Vanlauwe, B. Response of common bean (Phaseolus vulgaris L.) to nitrogen, phosphorus and rhizobia inoculation across variable soils in Zimbabwe. Agric Ecosyst Environ 2018, 266, 167–173. [Google Scholar] [CrossRef]
- Chibeba, A.M.; Kyei-Boahen, S.; Guimaraes, M.F.; Nogueira, M.A.; Hungria, M. Feasibility of transference of inoculation-related technologies: A case study of evaluation of soybean rhizobial strains under the agro-climatic conditions of Brazil and Mozambique. Agric Ecosyst Environ 2018, 261, 230–240. [Google Scholar] [CrossRef] [PubMed]
- Zengeni, R.; Mpepereki, S.; Giller, K.E. Manure and soil properties affect survival and persistence of soyabean nodulating rhizobia in smallholder soils of Zimbabwe. Applied Soil Ecology 2006, 32, 232–242. [Google Scholar] [CrossRef]
- Chiduwa, M. Improving the legume-rhizobium symbiosis in Zimbabwean agriculture: A study of rhizobia diversity & symbiotic potential focussed on soybean root nodule bacteria. In School of Agricultural Sciences; Murdoch University: Murdoch, Australia, 2021. [Google Scholar]
- Szczerba, A.; Plazek, A.; Kopec, P.; Wojcik-Jagla, M.; Dubert, F. Effect of different Bradyrhizobium japonicum inoculants on physiological and agronomic traits of soybean (Glycine max (L.) Merr.) associated with different expression of nodulation genes. BMC Plant Biol 2024, 24, 1201. [Google Scholar] [CrossRef]
- Ndhlovu, K.; Bopape, F.L.; Diale, M.O.; Mpai, T.; Morey, L.; Mtsweni, N.P.; Gerrano, A.S.; Vuuren, A.v.; Babalola, O.O.; Hassen, A.I. Characterization of Nodulation-Compatible Strains of Native Soil Rhizobia from the Rhizosphere of Soya Bean (Glycine max L.) Fields in South Africa. Nitrogen 2024, 5, 1107–1123. [Google Scholar] [CrossRef]
- Omari, R.A.; Yuan, K.; Anh, K.T.; Reckling, M.; Halwani, M.; Egamberdieva, D.; Ohkama-Ohtsu, N.; Bellingrath-Kimura, S.D. Enhanced Soybean Productivity by Inoculation With Indigenous Bradyrhizobium Strains in Agroecological Conditions of Northeast Germany. Front Plant Sci 2021, 12, 707080. [Google Scholar] [CrossRef]
- Musiyiwa, K.; Mpepereki, S.; Giller, K.E. Physiological diversity of rhizobia nodulating promiscuous soyabean in Zimbabwean soils. Symbiosis 2005, 40, 97–107. [Google Scholar]
- FAO. Fertilizer use by crop in Zimbabwe; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2006. [Google Scholar]
- Anderson, J.; Ingram, J. Tropical Soil Biology and Fertility: A Handbook of Methods. Soil Science 1994, 157, 265. [Google Scholar] [CrossRef]
- Murphy, J.; Riley, J.P. A modified single solution method for the determination of phosphate in natural waters. Analytica Chimica Acta 1962, 27, 31–36. [Google Scholar] [CrossRef]
- Okalebo, R.J.; Gathua, K.W.; Woomer, P.L. Laboratory methods of soil and plant analysis: A working manual, 2nd ed.; TSBF-CIAT and SACRED Africa, Kenya: Nairobi, Kenya, 2002. [Google Scholar]
- Tumbure, A.; Wuta, M.; Mapanda, F. Preliminary evaluation of the effectiveness ofRhizobium leguminosarumbv.viceaestrains in nodulating hairy vetch (Vicia villosa) in the sandy soils of Zimbabwe. South African Journal of Plant and Soil 2013, 30, 233–239. [Google Scholar] [CrossRef]
- Wang, W.; Shi, F.; Du, J.; Li, L.; Bai, T.; Xing, X. Soil factors that contribute to the abundance and structure of the diazotrophic community and soybean growth, yield, and quality under biochar amendment. Chemical and Biological Technologies in Agriculture 2023, 10. [Google Scholar] [CrossRef]
- Shen, D.; Bisseling, T. Soybean breeders can count on nodules. Trends Plant Sci 2025, 30, 10–12. [Google Scholar] [CrossRef] [PubMed]
- Abd-Alla, M.H.; Al-Amri, S.M.; El-Enany, A.-W.E. Enhancing Rhizobium–Legume Symbiosis and Reducing Nitrogen Fertilizer Use Are Potential Options for Mitigating Climate Change. Agriculture 2023, 13, 2092. [Google Scholar] [CrossRef]
- Zubrod, M. Counting the nodules that count, relationships between seed nitrogen and root nodules. Natural Sciences Education 2022, 51. [Google Scholar] [CrossRef]
- Chikwanha, H. High production costs render soyabean uncompetitive. In The Herald; Zimpapers: Harare, Zimbabwe; p. 2024.
- Omondi, J.O.; Chiduwa, M.S.; Kyei-Boahen, S.; Masikati, P.; Nyagumbo, I. Yield gap decomposition: quantifying factors limiting soybean yield in Southern Africa. npj Sustainable Agriculture 2024, 2. [Google Scholar] [CrossRef]
- Manzeke-Kangara, M.G.; Ligowe, I.S.; Tibu, A.; Gondwe, T.N.; Greathead, H.M.R.; Galdos, M.V. Soil organic carbon and related properties under conservation agriculture and contrasting conventional fields in Northern Malawi. Frontiers in Soil Science 2025, 4. [Google Scholar] [CrossRef]
- Nyawasha, R.W.; Falconnier, G.N.; Todoroff, P.; Wadoux, A.M.J.C.; Chikowo, R.; Coquereau, A.; Leroux, L.; Jahel, C.; Corbeels, M.; Cardinael, R. Drivers of soil organic carbon stocks at village scale in a sub-humid region of Zimbabwe. Catena 2025, 252. [Google Scholar] [CrossRef]
- Swanepoel, P.A.; Botha, P.R.; Truter, W.F.; Surridge-Talbot, A.K. The effect of soil carbon on symbiotic nitrogen fixation and symbioticRhizobiumpopulations in soil withTrifolium repensas host plant. African Journal of Range & Forage Science 2011, 28, 121–127. [Google Scholar] [CrossRef]
- Nezomba, H.; Mtambanengwe, F.; Tittonell, P.; Mapfumo, P. Practical assessment of soil degradation on smallholder farmers' fields in Zimbabwe: Integrating local knowledge and scientific diagnostic indicators. Catena 2017, 156, 216–227. [Google Scholar] [CrossRef]
- Chipomho, J.; Rugare, J.T.; Mabasa, S.; Zingore, S.; Mashingaidze, A.B.; Chikowo, R. Short-term impacts of soil nutrient management on maize (Zea mays L.) productivity and weed dynamics along a toposequence in Eastern Zimbabwe. Heliyon 2020, 6, e05223. [Google Scholar] [CrossRef]
- Mutuma, S.P.; Okello, J.J.; Karanja, N.K.; Woomer, P.L. Smallholder farmers’ use and profitability of legume inoculants in Western Kenya. African Crop Science Journal 2014, 22, 205–213. [Google Scholar]
- GRDC. Soybean nutrition and fertiliser. In GRDC Grownotes, Grains Research and Development Corporation: Australia, 2016.
- Kebonye, N.M.; John, K.; Delgado-Baquerizo, M.; Zhou, Y.; Agyeman, P.C.; Seletlo, Z.; Heung, B.; Scholten, T. Major overlap in plant and soil organic carbon hotspots across Africa. Sci Total Environ 2024, 951, 175476. [Google Scholar] [CrossRef]
Figure 1.
Experimental site locations.
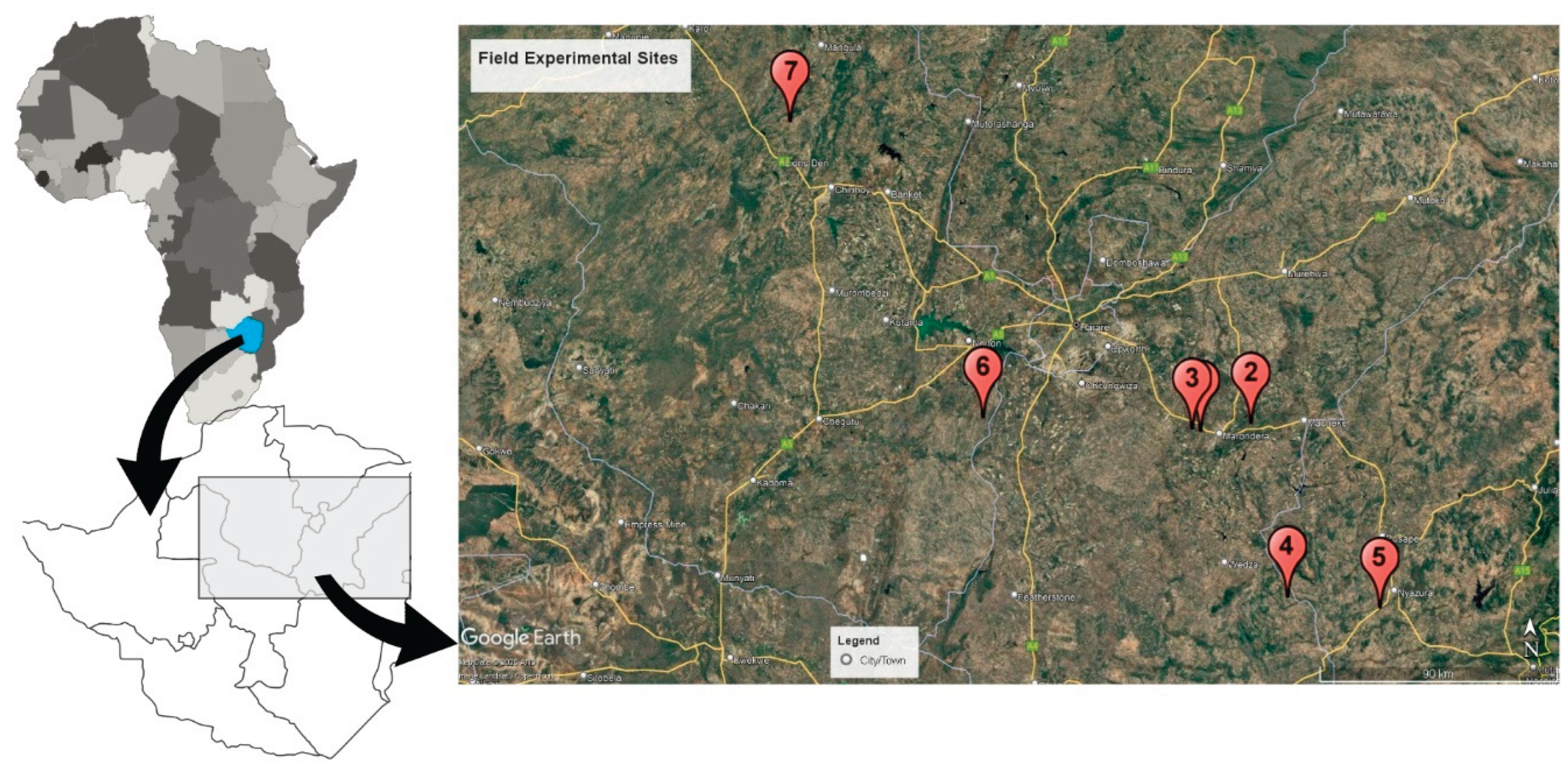
Figure 2.
Biomass production of soybean at the flowering stage at researcher managed sites. Error bars are 2x standard error of means (n = 4). Treatments with the same letter are not significantly different from each other at 0.05 probability level.
Figure 2.
Biomass production of soybean at the flowering stage at researcher managed sites. Error bars are 2x standard error of means (n = 4). Treatments with the same letter are not significantly different from each other at 0.05 probability level.
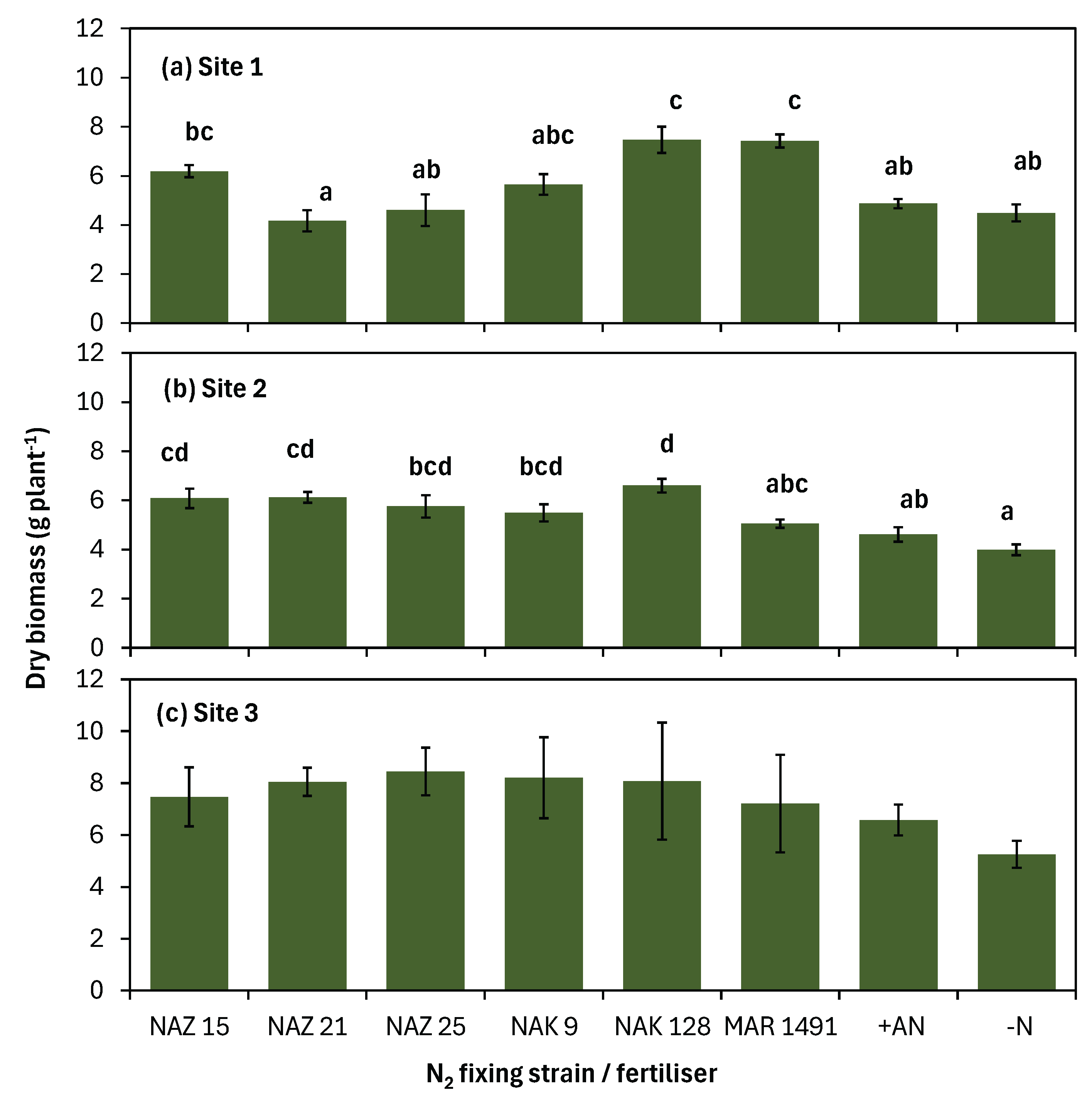
Figure 3.
Correlation matrix of grain yield, grain N uptake and soil properties constructed from pooled data from rainfed sites (n =48).
Figure 3.
Correlation matrix of grain yield, grain N uptake and soil properties constructed from pooled data from rainfed sites (n =48).
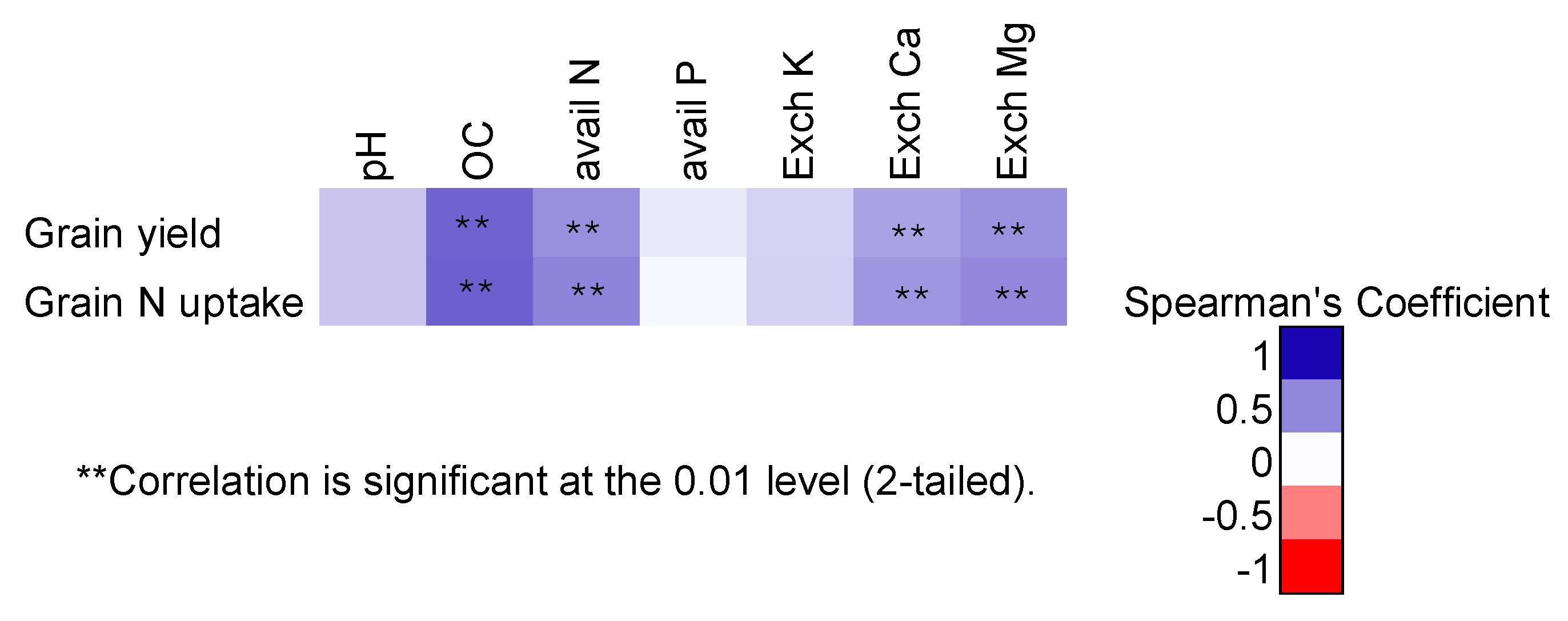
Figure 4.
Aboveground biomass production, plant N uptake and estimated N derived from fixation (Ndff) in aboveground biomass of soybean at site 3. Error bars are 2 x standard error of means (SEM), n = 4.
Figure 4.
Aboveground biomass production, plant N uptake and estimated N derived from fixation (Ndff) in aboveground biomass of soybean at site 3. Error bars are 2 x standard error of means (SEM), n = 4.
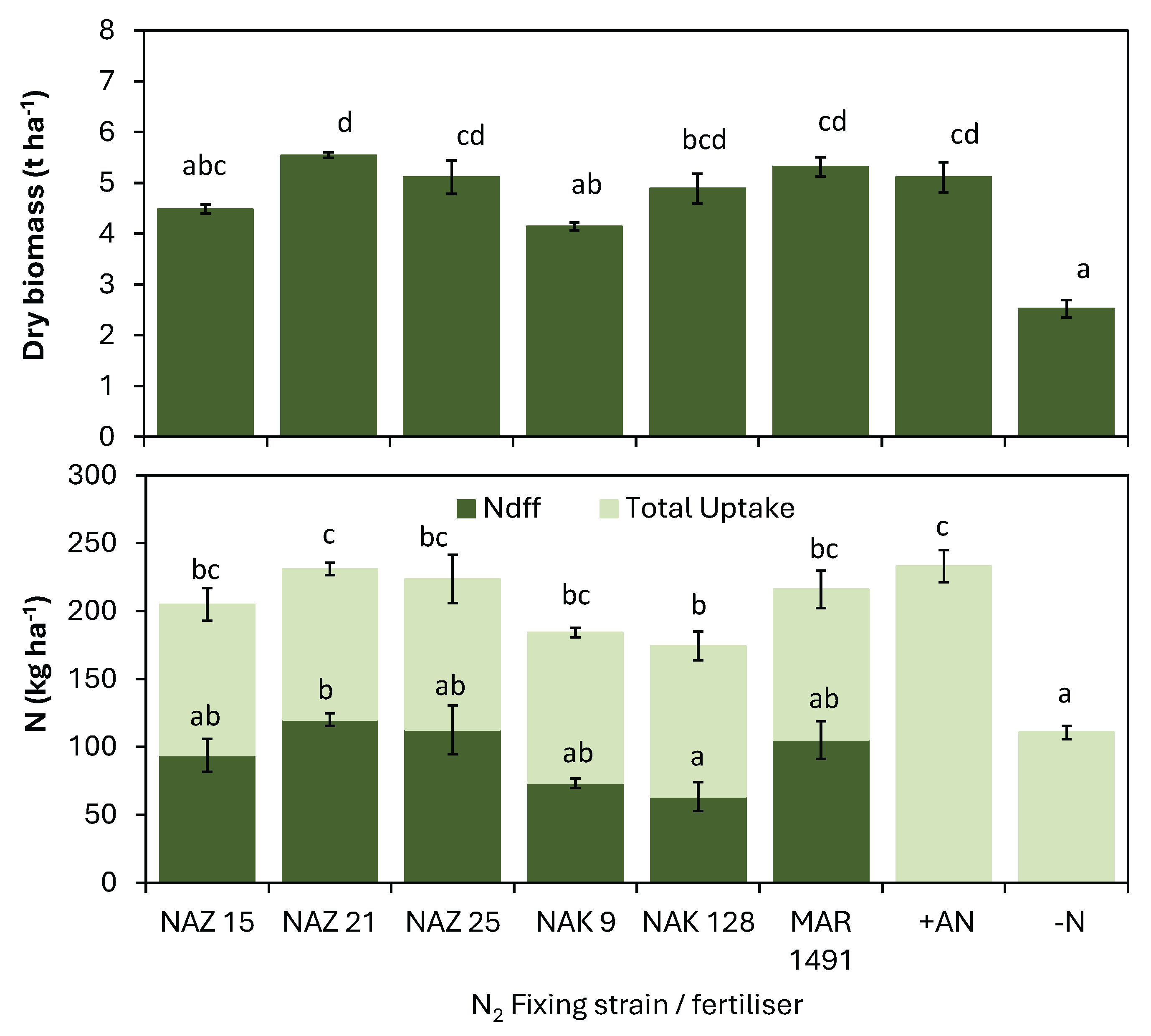

Table 1.
Selected soil chemical characteristics at the seven experimental sites prior to trial establishment.
Table 1.
Selected soil chemical characteristics at the seven experimental sites prior to trial establishment.
| Site | Details /Names | pH 1 | Soil OC % | Mineral N (ppm) | Avail P (ppm) | Exchangeable bases (me %) | ||
|---|---|---|---|---|---|---|---|---|
| K | Ca | Mg | ||||||
| 1 | SPRL | 4.5 | 0.35 | 22 | 5 | 0.09 | 0.64 | 0.38 |
| 2 | Kushinga P.C. | 5.3 | 1.08 | 11 | 37 | 0.42 | 2.4 | 1.51 |
| 3 | Horticulture Research Institute | 4.9 | 1.28 | 23 | 19.6 | 1.02 | 3.63 | 0.20 |
| 4 | Wedza | 5.0 | 0.23 | 5 | 4 | 0.05 | 0.73 | 0.35 |
| 5 | Rusape/Makoni | 5.1 | 1.35 | 44 | 2 | 0.25 | 9.41 | 4.1 |
| 6 | Mhondoro | 4.7 | 1.06 | 29 | 2 | 0.56 | 5.35 | 2.83 |
| 7 | Chinhoyi | 5.6 | 1.20 | 28 | 9 | 0.74 | 6.43 | 4.3 |
1CaCl2 scale, Avail – available, me% - milliequivalents per 100g.
Table 2.
Nodule counts and nodule fresh mass of soybean inoculated with various strains of rhizobia, at the researcher managed sites in Marondera, Zimbabwe.
Table 2.
Nodule counts and nodule fresh mass of soybean inoculated with various strains of rhizobia, at the researcher managed sites in Marondera, Zimbabwe.
| Site 1 | Site 2 | Site 3 | ||||
|---|---|---|---|---|---|---|
| N fixing strain / fertilizer | Nodules plant-1 | Fresh nodule mass plant-1 (g) | Nodules plant-1 | Fresh nodule mass plant-1 (g) | Nodules plant-1 | Fresh nodule mass plant-1 (g) |
| NAZ 15 | 36d ±1 | 0.98c ±0.07 | 25d ±1 | 0.38bc ±0.09 | 25b ±2 | 0.79cd ±0.11 |
| NAZ 21 | 24bcd ±2 | 0.57ab ±0.06 | 13cd ±1 | 0.38bc ±0.05 | 10a ±2 | 0.42abc ±0.05 |
| NAZ 25 | 7ab ±1 | 0.59ab ±0.03 | 11abc ±1 | 0.40bc ±0.04 | 17ab ±4 | 0.82cd ±0.16 |
| NAK 9 | 17abc ±2 | 0.60ab ±0.05 | 11abc ±1 | 0.22b ±0.02 | 10a ±2 | 0.43abc ±0.04 |
| NAK 128 | 27cd ±4 | 0.84bc ±0.07 | 12bcd ±1 | 0.43c ±0.04 | 22ab ±2 | 1.00d ±0.10 |
| MAR 1491 | 24bcd ±1 | 0.88bc ±0.04 | 16cd ±1 | 0.23bc ±0.04 | 21ab ±2 | 0.73bcd ±0.10 |
| +AN | 1a ±0 | 0.00a ±0 | 0.4ab ±0 | 0.04a ±0.02 | 12a ±4 | 0.37a ±0.05 |
| -N | 2a ±1 | 0.15a ±0.06 | 0.1a ±0 | 0.01a ±0.01 | 10a ±2 | 0.40ab ±0.08 |
| Significance | ** | ** | ** | *** | ** | *** |
Superscript letters represent statistical significance at α= 0.05, n=4. Numbers after ± are standard error of means (SEM).
Table 3.
Soybean grain yields and grain N uptake after inoculating with varying rhizobia strains.
| N fixing strain / fertilizer | Site 1 | Site 2 | Site 5 | Site 4 | Site 6 | Site 7 | Site 3 |
|---|---|---|---|---|---|---|---|
| Grain yield (t ha-1) | |||||||
| NAZ 15 | 1.84b ±0.14 | 1.47b ±0.08 | 2.15ab ±0.16 | 0.81b ±0.09 | 1.06 ±0.04 | 1.12a ±0.12 | 3.31b ±0.21 |
| NAZ 21 | 1.31ab ±0.17 | 2.10c ±0.07 | 2.56b ±0.22 | 0.86b ±0.13 | 1.13 ±0.14 | 1.63ab ±0.11 | 3.28b ±0.10 |
| NAZ 25 | 1.70b ±0.12 | 1.63b ±0.06 | 2.41b ±0.30 | 1.08b ±0.16 | 1.28 ±0.21 | 1.73ab ±0.24 | 3.27b ±0.25 |
| NAK 9 | 1.08a ±0.12 | 1.45ab ±0.09 | 2.88b ±0.17 | 0.70b ±0.08 | 1.30 ±0.23 | 1.60ab ±0.19 | 2.43a ±0.13 |
| NAK 128 | 1.69b ±0.16 | 1.59b ±0.12 | 2.34b ±0.18 | 0.77b ±0.06 | 0.75 ±0.08 | 0.83a ±0.16 | 2.90ab ±0.07 |
| MAR 1491 | 1.63ab ±0.14 | 1.70bc ±0.13 | 1.99ab ±0.28 | 0.63ab ±0.10 | 1.59 ±0.30 | 2.35b ±0.36 | 3.26b ±0.17 |
| +AN | 1.83b ±0.08 | 1.75bc ±0.08 | 2.29b ±0.25 | 0.30ab ±0.03 | 1.07 ±0.16 | 1.61ab ±0.15 | 3.47b ±0.18 |
| -N | 1.39ab ±0.06 | 1.02a ±0.08 | 1.23a ±0.19 | 0.16a ±0.06 | 0.77 ±0.15 | 1.49ab ±0.20 | 1.61a ±0.07 |
| Significance | *** | *** | *** | *** | ns | * | * |
| Grain N Uptake (kg N / ha-1) | |||||||
| NAZ 15 | 107.7b ±7.6 | 86.4b ±4.2 | 145.2ab ±13.6 | 52.4c ±5.4 | 62.1 ±7.6 | 70.9ab ±11.6 | 106.20b ±7.95 |
| NAZ 21 | 77.1ab ±9.8 | 103.0b ±8.6 | 156.2b ±14.3 | 58.7c ±10.3 | 60.9 ±6.3 | 93.0ab ±5.9 | 106.87b ±6.60 |
| NAZ 25 | 99.1ab ±7.2 | 95.5b ±7.2 | 148.3ab ±16.4 | 64.4c ±7.3 | 73.6 ±12.8 | 103.8ab ±14.5 | 104.58b ±9.72 |
| NAK 9 | 61.6a ±6.4 | 68.2ab ±9.0 | 182.4b ±12.6 | 42.7bc ±5.1 | 71.6 ±17.3 | 100.3ab ±13.8 | 77.33b ±3.72 |
| NAK 128 | 99.4ab ±9.3 | 92.9b ±12.9 | 141.4ab ±15.4 | 41.8bc ±4.8 | 37.6 ±4.8 | 52.9a ±11.5 | 66.35ab ±10.33 |
| MAR 1491 | 95.0ab ±8.0 | 94.4b ±6.3 | 121.0ab ±21.9 | 41.3bc ±7.8 | 96.1 ±21.2 | 126.5b ±20.9 | 97.25b ±8.30 |
| +AN | 113.8b ±2.7 | 86.8b ±6.5 | 150.1b ±15.4 | 17.3ab ±1.9 | 51.2 ±5.7 | 100.2ab ±13.8 | 106.91b ±7.58 |
| -N | 67.8a ±12.9 | 43.4a ±3.4 | 76.9a ±11.5 | 10.0a ±3.6 | 42.7 ±10.7 | 91.4ab ±15.4 | 50.80a ±1.45 |
| Significance | * | *** | *** | *** | ns | * | *** |
Numbers after ± are standard error of means (SEM), n = 4, ns – not significant/.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



