
Submitted:
27 June 2025
Posted:
27 June 2025
You are already at the latest version
Abstract
Early diarrhea and weaning stress are the main factors that seriously affect the high-efficiency healthy breeding. The beneficial microorganism has the characteristics of improving feed digestibility, improving host growth performance, enhancing immunity, and reducing diarrhea. As a carrier of probiotics, fermented milk can promote the establishment of a beneficial microbial community in the gut of piglets. The purpose of this study was to investigate the effects of supplemental lactic acid bacteria fermented milk (LFM) on growth performance, immune inflammation, and fecal microbiota structure in growing and weaned piglets. Sixteen litters of the same batch of (Duroc×Landrace×Yorkshire) piglets with 7-day-old (initial BW =2.85 ± 0.09 kg) were randomly divided into the control group (CON) and the fermented milk group (FM) with 8 litters in each group. Piglets in the FM group were supplemented twice a day with 150 ml, 300 ml, and 450 ml of FM per litter at 7-13 days, 14-20 days and 21-28 days of age, respectively, while the CON group was added with an equal amount of drinking water. The results showed that adding FM significantly increased the average daily feed intake of piglets at 7-28 days of age, significantly increased the average daily gain of piglets after weaning, and significantly reduced the diarrhea rate. In addition, consumption of FM significantly reduced serum proinflammatory cytokine levels and increased serum levels of anti-inflammatory cytokines and immunoglobulin G. Additionally, FM consumption significantly decreased the relative abundance of Erysipelotrichaceae UCG-003 and Intestinimonas, which were positively related to the diarrhea rate of piglets, and increased the relative abundance of Ligilactobacillus and Holdemania in fecal microbiota of suckling piglets. In addition, the relative abundance of Campylobacter, Clostridium, p-251-o5_norank, and other harmful bacteria in weaned piglets was significantly reduced by FM. These findings support the benefits of FM for regulating gut microbiota balance and improving weaning health in piglets.
Keywords:
fermented milk
; weaned piglets
; growth performance
; fecal microbiota
; immune inflammation
1. Introduction
In the lactation period, the body organs of piglets have not been fully developed, and the ability to adapt to the external environment is poor. At this stage, piglets are difficult to raise, and their health status directly affects the economic benefits of pig breeding [1]. At the weaning stage, early weaning usually leads to delayed development of the gastrointestinal tract and dysbiosis of the intestinal microbiota, resulting in decreased digestibility, diarrhea, intestinal inflammation, and stunted growth, generally known as weaning stress syndrome [2]. At the same time, the transition from sow milk to solid feed at weaning may induce intestinal microbiota dysbiosis in piglets, resulting in post-weaning diarrhea [3], which significantly increased the mortality of piglets after weaning [4].
Acidifiers, as a safe additive can be used to replace antibiotics, can improve the intestinal health of weaned piglets, reduce intestinal pH, and facilitate the colonization of lactic acid bacteria in the intestines of piglets [5]. Adding mannan oligosaccharides to the diet of piglets can improve the growth performance and nutrient digestibility of piglets [6]. At the same time, many studies have shown that oligosaccharides have positive effects on promoting the health of piglets [7]. In addition, infant formula is often supplemented with oligosaccharides and functional oligosaccharides, such as galactooligosaccharides, fructooligosaccharides, and galactose, which are beneficial to intestinal development [8]. A functional fermented yoghurt was developed with three strains of lactic acid bacteria from pigs as fermentation strains and adding acidifiers and functional oligosaccharides, The results showed that supplementation with FM could improve the growth performance of weaned piglets and increase the abundance of beneficial bacteria in the intestine. Fermented yoghurt is a good culture of probiotics because of its specific chemical and physical properties, including low pH, high buffer capacity, and watery texture [9]. Feeding fermented yoghurt to piglets can improve their growth performance and reduce diarrhea rates. Feeding the fermented milk to piglets after 2 weeks of weaning improved intestinal health and immune tolerance [10]. The research on using FM as a carrier of probiotics has become a hot topic recently. Although many functions of fermented yoghurt have been recognized, the application of fermented yoghurt in the pig industry still lacks in-depth research [11].
On the basis of the previous research of the research group, this experiment fed the basic diet and FM at the same time to investigate the effects of FM on the growth performance, immune function and fecal microbiota of suckling and weaning piglets. This study provides a theoretical basis for the application of FM in animal husbandry.
2. Materials and Methods
2.1. Preparation of Lactic Acid Bacteria Fermented Milk
Fermented milk was prepared with 15% milk powder from cows, 8% sucrose, and 2% functional oligosaccharides (1.5% fructooligosaccharides and 0.5% galactose) as substrates (Lin et al., 2022). Three strains of lactic acid bacteria from pigs (Lactobacillus johnsonii LAB19, Enterococcus hirae LAB20, and Enterococcus faecium LAB8 in a ratio of 3:2:3) were used as a composite strain, adding 0.25%-0.5% complex acidifier to adjust the pH of yoghurt to about 4.0, aseptic package, and stored in a 4°C refrigerated reserve. The main nutritional components of fermented milk were as follows: dry matter 23.45%, milk protein 3.24%, milk fat 3.12%, lactose 5.60%, and viable bacteria 4.50 × 109 CFU/mL.
2.2. Animals and Experimental Design
The experiment was conducted in Zhenjiang, Jiangsu Province, China. Sixteen litters of the same batch of (Duroc×Landrace×Large white) piglets with 7-day-old (initial BW =2.85 ± 0.09 kg) were randomly divided into control group (CON) and fermented milk group (FM) with 8 litters in each group. The 21-day feeding trial was conducted in two phases. During the pre-weaning period (days 7–21), all piglets had ad libitum access to sow milk. In addition, the control (CON) group received creep feed, whereas the fermented milk (FM) group received creep feed supplemented with fermented milk twice daily per litter at 07:00 and 17:00. Specifically, piglets in the FM group were supplemented with 150 mL of fermented milk per feeding per litter from days 7 to 13, and 300 mL per feeding per litter from days 14 to 20. Following weaning (days 21–28), CON piglets continued on creep feed alone, while FM piglets received creep feed plus 450 mL of fermented milk per feeding per litter twice daily. Throughout the experimental period, the automatic drinking system was inspected daily at fixed time points (07:00, 12:00, and 17:00) to ensure ad libitum access to feed and water for all piglets. The nutritional levels of CP, digestible energy, minerals and vitamins, and calcium were kept consistent in the diets of each group, and the nutritional levels were based on the National Research Council (NRC) in 2012 and Nutrient Requirements of Swine in China (2004).
2.3. Sampling
On days 21, 24, and 28, at 07:00-08:00, fresh fecal samples of approximately 5 g were collected from 1 piglet per litter randomly, dispensed into 50 mL centrifuge tubes, and immediately frozen with liquid nitrogen for further analysis. On days 21 and 28 of the experiment, 5 mL of blood from the anterior jugular vein of one piglet per litter was randomly collected, and after centrifugation at 3500 r/min for 10 min, the supernatant was dispensed into 1.5 mL centrifuge tubes and stored at -20°C. From 18 to 21 days and 25 to 28 days of age, 100 g of fecal samples were collected daily in a new self-sealing bag, and 10 mL of 10% hydrochloric acid was added dropwise to fix the nitrogen for 3 consecutive days. The fecal samples were put into a refrigerator for freezing and preservation at -20°C, and the fecal samples were used for the determination of the nutrient apparent digestibility.
2.4. Growth Performance
Individual body weight (BW) was recorded at the age of 7, 14, 21 and 28 days after 12 h of fasting. The feed intake of piglets in each pen was recorded as the amount of feed provided during the experiment minus the amount of surplus and waste. The average daily gain (ADG) = (test final weight –test initial weight)/number of test days; The average daily feed intake (ADFI) = (feed intake during test – feed remaining during test)/number of test days; The ratio of feed to gain (F: G) = average daily feed intake/average daily gain.
2.5. Diarrhoea-Related Indicators
During the experimental period, at 08:30 and 17:30 each day, each piglet was individually inspected for faecal contamination and erythema in the perianal region, and faecal consistency was assessed; faecal scores were then assigned based on stool consistency. The diarrhea scores were assigned as follows: normal: cylindrically shaped (0 points); soft feces: thin, soft, and shapely (1 point); mild diarrhoea: sticky, unshapely, porridge-like thin stool (2 points); severe diarrhoea: liquid, unshapely, watery thin stool (3 points) [12]. Among these, two points or more were recorded as diarrhea, and the treatment was recorded in detail. Diarrhoea rate = number of pigs with diarrhoea/(number of test pigs * total days) * %; diarrhoea index = sum of fecal score/(number of test pigs * total days).
2.6. Apparent Digestibility of Nutrients
The fecal samples were removed from the refrigerator, thawed to room temperature, dried in a thermostatic oven at 65°C to an air-dried state, and crushed through a 40-mesh sieve. Apparent digestibility of nutrients was determined by using the endogenous indicator acid insoluble ash (AIA). Crude fat (ether extract, EE) content was determined by Soxhlet extraction, and crude protein (CP) content was determined by Kjeldahl nitrogen determination in both ration and fecal samples.
2.7. Serum Biochemical Indicators
Serum samples stored in the refrigerator at -20°C were removed and thawed at -4°C. Serum immuno-inflammatory indicators were measured by the ELISA kit according to the manufacturer’s instructions (Enzyme-Linked Biotechnology, Shanghai, China). Immunoglobulin A (IgA), Immunoglobulin G (IgG), interleukin-1β (IL-1β), interleukin-6 (IL-6), interleukin-8 (IL-8), interleukin-10 (IL-10), and tumor necrosis factor-α (TNF-α) levels in serum and ileum were determined using ELISA kits according to the manufacturer’s instructions.
2.8. 16S rRNA Gene Sequencing of Fecal Microbiota
The fecal samples were thawed at 4°C, and 1.5 g of feces was taken into a 2 mL sterilized centrifuge tube and centrifuged at 9000 rpm at 4°C for 5 min. The microbial nucleic acids were extracted according to the methods of Dai [13]. After passing the quality control of the Nano-Drop 2100 Nucleic Acid Protein Detector (Thermo Fisher Scientific, USA), the sample was stored at -20°C. The V4-V5 regions of the 16S rRNA gene were amplified by PCR using bacterial primers 515F (5‘-GTGCCAGCMGCCGCGG-3’) and 907R (5‘-CCGTCAATTCMTTTRAGTTT-3’). The DNA from the amplified products was subsequently purified using the AxyPrep DNA Recovery Kit (AxyPrep Biosciences, USA), and the paired-end libraries were constructed according to Illumina’s Genomic DNA Library Preparation Procedure and analyzed by sequencing on the Illumina Mi-seq platform following the manufacturer’s standard procedure. Species taxonomic annotations were obtained from Silva database comparisons at a 70% confidence level. MOTHUR (version:1.30.1) was used to calculate the sequencing depth and α-diversity index of the microbial communities and to perform principal co-ordinate analysis (PCoA).
2.9. Data Analysis
Experimental data were analyzed using SPSS 20.0 software (SPSS Inc., Chicago, IL, USA), with all values expressed as mean ± standard error of mean (Mean ± SEM). Individual body weight and mean daily weight gain were analyzed using a mixed model, with the nest factor as the test factor. Mean feed intake and feed-to-weight ratio were analyzed using a mixed model with FM treatment as the main factor. Other variables were analyzed using a t-test with litter as the experimental unit (n = 8), where only one piglet per litter was sampled to represent the litter mean. All images were generated using GraphPad Prism 9.5 (GraphPad Software, Inc., La Jolla, CA, USA), and significant differences were declared when P < 0.05, and differences of 0.05 ≤ P < 0.10 were considered trends.
3. Results
3.1. Effect of Supplemental Fermented Milk on Growth Performance and Diarrhea Indexes of Suckling and Weaning Piglets
As shown in Table 1, FM significantly increased ADFI of piglets at 7-14 days of age (p < 0.05). In addition, FM significantly increased ADG of piglets at 22-28 days of age (p < 0.05). As shown in Table 2, the diarrhea rate (p = 0.092) and diarrhea index (p = 0.079) of piglets fed with FM at 7-14 days of age tended to decrease. There were no significant changes (p > 0.05) in diarrhea rate and diarrhea index between the two groups at 15-21 days of age (p > 0.05). FM supplementation significantly reduced the diarrhea rate and diarrhea index of piglets at 22-28 days of age (p < 0.05).
3.2. Effect of Supplemental Fermented Milk on Nutrient Digestibility of Lactating-Weaning Piglets
As shown in Table 3, the apparent digestibility of crude protein (p = 0.078) and crude fat (p = 0.065) in piglets fed with FM at 18-21 days of age showed a decreasing trend as compared to the CON group. The apparent digestibility of crude fat (p = 0.062) in 25-28 days old piglets fed with FM tended to decrease. There were no significant differences in the apparent digestibility of dry matter, crude ash, or crude fiber between the two groups (p > 0.05).
3.3. Effect of Supplemental Fermented Milk on Serum Inflammatory Indexes in Suckling and Weaning Piglets
As shown in Table 4, the level of serum IL-1β in piglets fed with FM at 21 days of age was significantly higher than that in the control group (p < 0.05). At the 28th day of age, compared with the CON group, the level of IL-10 were significantly increased with FM treatment (p < 0.05), while the level of IL-6 were significantly decreased (p < 0.05).
3.4. Effect of Supplemental Fermented Milk on Serum Immunoglobulin Content in Suckling and Weaning Piglets
As showed in Figure 1, there were no significant changes in IgA and IgG levels between the two groups at 21 days of age (p > 0.05). FM Supplementation significantly increased the level of IgG at 28 days of age (p < 0.05), but the level of IgA had no significant change (p > 0.05).
3.5. Effect of Supplemental Fermented Milk on the Structure of Fecal Microbiota in Suckling and Weaning Piglets
Fecal microbial α-diversity indices are shown in Figure 2A, Figure 2B, and Figure 2C. At the 21, 24, and 28 days of age, the ACE index of piglets in the supplemented FM was significantly lower (p < 0.05) than the control group. At 24 and 28 days of age, the Shannon index was significantly lower (p < 0.05) in piglets in the FM supplementation group compared to the CON group. To analyze the differences between groups, we performed between-group PCoA principal coordinate analysis of genus-level microorganisms (Figure 2D, Figure 2E, and Figure 2F). The representative contribution of the control group to the total microorganisms detected at 21, 24, and 28 days of age was 13, 18, and 21%, respectively, and the supplemented fermented milk group to the total microorganisms detected was 15%, 12%, and 12%, respectively. To identify species that differed between groups, we analyzed species differences in the abundance of microbiota in fecal samples at the phylum level, family level, and genus level in piglets at the 21, 24, and 28 days of age (Figure 2G, Figure 2I, and Figure 2H). At the phylum level, phylum Firmicutes was the most predominant of the fecal microbiota in the two groups, followed by Bacteroidota and Spirochaetota; at the family level, Oscillospiraceae, Lachnospiraceae, Ruminococcaceae, and Muribaculaceae dominated the fecal microbiota of both groups; at the genus level, two groups of fecal microbiota were dominated by Eubacterium coprostanoligenes group_norank and Muribaculaceae_norank.
In order to identify the different strains between the two groups, Linear discriminant analysis Effect Size (LEfSe) was performed to analyze the microbial groups with significant differences between the two groups (Linear discriminant analysis, LDA > 3). As shown in Figure 3B, at the 21 day of age, the relative abundances of genera Lactobacillus royalei and Haldemannia were significantly higher in the FM group compared with the CON group (p < 0.05), while the genera Erysipelotrichaceae UCG-003, Burkholderiacea_Caballeronia_Paraburkholderia, Lachnospiraceae_XPB1014_group and Intestinimonas in the CON group were significantly higher compared with the FM group (p < 0.05). Nineteen bacterial taxa including genera p-251-o5_norank, UCG-005, CAG-873, Treponema, and phylum Spirochaetota were significantly higher in the control group at day 24 of age (Figure 3D). As showed in Figure 3E, at 28 day of age, the abundance of genus Megasphaerobacter was significantly higher (p < 0.05) in the FM group compared with the CON group; twenty-one bacterial taxa including phyla Desulfobacterota, Campylobacterota and Fusobacteriota, families Bacteroidales RF16 group and Anaerovoracaceaegenera, Campylobacter, Shuttleworthia and p-251-o5_norank were significantly higher in the CON group than the FM group (p < 0.05).
3.6. Correlation Analysis of Fecal Microbiota and Differential Phenotypic Indicators in Suckling and Weaning Piglets
In order to identify the key species causing changes in the production performance of weaned piglets, spearman correlation coefficient was used to evaluate the correlation between intestinal microbiota and differential phenotypic indicators. As can be seen in Figure 4A, at the 21 day of age, the diarrheal rate of piglet was positively (p < 0.05) correlated with the abundances of genera Erysipelotrichaceae UCG-003 and Intestinimonas; serum IgG level was negatively correlated (p < 0.05) with genus Holdermannia. In additation, serum IL-1β level and genus Lachnospiraceae XPB1014 group showed negative (p < 0.05) correlation. At the 28th day of age of piglets, the genus p-251-o5_norank was positively (p < 0.05) correlated with piglet diarrheal rate (Figure 4B), serum IL-6 and TNF-α levels; the phylum Fusobacteriota, the genera Fusobacterium and p-251-o5_norank were negatively correlated with serum IL-10 (p < 0.05), the phylum Campylobacterota, the genera Campylobacter and Dimlma were negatively correlated with serum IgA level (p < 0.05).
4. Discussion
The shortening of the lactation period and the insufficient intake of colostrum due to the weaning of early piglets prevented them from obtaining sufficient immunoglobulins and nutrients, coupled with the fact that their gastrointestinal digestive system is still in a state of development, which can easily lead to indigestion and diarrhea, and affect subsequent growth and development, production performance, and even death [14]. In this study, FM was supplemented before and after the weaning of piglets to evaluate its effects on growth performance, diarrhea, immunoinflammatory parameters, and fecal microbiota composition. FM is a healthy and green functional food, rich in beneficial lactobacilli, which has the function of regulating the dynamic balance of the gastrointestinal microbiota and maintaining the healthy state of the intestinal tract [15]. Lactobacilli involved in fermentation can break down proteins, lactose, and produce immunologically active substances, such as immunomodulatory factors, antioxidant substances, and antimicrobial peptides, which can promote the development of the body’s immune organs [16]. Our study demonstrated that supplemental feeding with fermented milk moderately enhanced the growth performance of piglets during both the pre- and post-weaning phases, while also significantly reducing the incidence of diarrhea in weaned piglets. These findings align with previous research [17]. The observed benefits may be attributed to the improved palatability and nutritional value of fermented milk, which likely promotes better appetite in piglets. Additionally, the abundant and stable lactic acid bacteria in the fermented milk are well-adapted to the piglets’ gastrointestinal environment and may support the development of the immune system by promoting the production of immunomodulatory factors and antioxidant peptides [18]. It can be well adapted to the complex internal environment in the piglets’ intestines, promoting the efficiency of intestinal digestion and absorption of nutrients. We also found that FM supplementation significantly increased serum immunoglobulin and anti-inflammatory cytokine levels in piglets after weaning, which is consistent with Griet’s findings [19]. This may be related to the acidifying agents and probiotics in fermented milk, which can promote the production of anti-inflammatory cytokines by stimulating macrophages and enhancing phagocytosis of pathogenic microorganisms, as well as positively affect intestinal tight junction proteins by decreasing the levels of pro-inflammatory cytokines, and consequently, intestinal tight junction proteins [20]. In addition, the addition of FM positively affected the intestinal microbiota and regulated the intestinal microecological balance, which in turn improved the intestinal internal environment of weaned piglets [21].
The FM (fermented-milk) group showed non-significant but consistent decreases in CP and CF digestibility at 18–21 days, this may result from the high lactic-acid content and bioactive peptides in the FM supplement, which lower intestinal pH and increase digesta viscosity—factors known to impair pancreatic enzyme activity and slow enzyme–substrate diffusion. Additionally, enhanced proliferation of probiotic bacteria can incorporate dietary protein and fat into microbial biomass that is then excreted, appearing as reduced ‘apparent’ digestibility [22,23]. Future studies should directly measure luminal pH, viscosity, and enzyme activities to confirm these mechanisms.
In this study, oral administration of FM significantly reduced the concentration of IL-6 and significantly increased the concentration of IL-10 in piglets after weaning, which may be an important mechanism of action of FM in alleviating weaning-induced intestinal barrier damage. Previous studies have shown that weaning of piglets causes activation of inflammatory pathways such as NF-κB, and T cell subsets such as CD4+ and CD8+ are released in response to infections and stress, leading to an increase in the expression of intestinal inflammatory factor genes [3,24]. It is well known that overexpression of pro-inflammatory cytokines (TNF-α, IL-1β, IL-6, and IL-8) induces intestinal mucosal damage, alters intestinal epithelial permeability, and triggers an inflammatory response, thereby exacerbating diarrhea [25]. On the other hand, IL-10 can attenuate inflammatory responses by decreasing pro-inflammatory cytokine expression [26]. The results of this study also showed that FM supplementation attenuated diarrhea in piglets after weaning. In summary, FM supplementation can alleviate intestinal mucosal damage and reduce piglet diarrhea by inhibiting the overproduction of pro-inflammatory cytokines and promoting the secretion of anti-inflammatory factors.
Serum immunoglobulin content is an important basis for the stability of piglets through the weaning period. The increase in immunoglobulin concentration can promote and regulate the immune function of antibodies, which is closely related to the gastrointestinal mucosal immune barrier to protect the intestinal surface from the damage of pathogenic bacteria, and thus reduce the adverse effects of weaning stress on piglets [27,28]. It was found that supplementing the diet of weaned piglets with probiotic-based feed additives can significantly increase the serum IgG level of weaned piglets and improve their immune response ability [29]. The addition of lactic acid acidifier of can improve the immune resistance ability of the piglets, so as to improve the intestinal immunity of piglets [30]. The present study showed that the additional intake of FM significantly increased the content of IgG in serum at the age of 28 days. Supplemental FM appears to enhance the immune function of weaned piglets, a benefit that may be attributable to the bioactive small molecules present in FM (e.g., organic acids, metabolites, and soluble peptides), which have been shown to elevate immunoglobulin and cytokine levels [31]. In conclusion, our results demonstrated that supplementation with FM improves the immune function of the organism and thus reduces the adverse effects of weaning stress in piglets.
The dynamic balance of the microecology of the intestinal tract is a prerequisite for the normal functioning of the body, but with the weaning of piglets, dietary and environmental changes, the gastrointestinal barrier function and immune function of piglets will be disturbed to a certain extent. The composition and quantity of microorganisms in piglet feces can reflect the balance of intestinal microbiota [32,33]. It has been reported that Firmicutes and Bacteroidetes are the two most dominant phylum regardless of the age or different intestinal segments of pigs [34]. After oral administration of lactic acid bacteria to weaned piglets, the dominant phylum in the gastrointestinal tract were Firmicutes, Bacteroides, Proteobacteria, and Clostridium [35]. This is consistent with the results of the present experiment, indicating that supplemental feeding of fermented milk had no impact on the predominant bacterial species in the feces of weaned piglets. The addition of the acidifier provide an acidic environment, that increase the abundance of Lactobacillus in the intestine [36]. By feeding probiotic complexes, the intestinal metabolism of piglets could be promoted to produce lactic acid, thus reducing the pH in the intestine and inhibiting the growth and reproduction of harmful microbiota, and that a reasonable microbial community structure could prevent the development of intestinal diseases, improve performance, and reduce diarrhea-associated indices [37]. The combined supplementation of Lactobacillus fermentum and Chlorella lactis in the diets of weaned piglets resulted in a decreasing trend in the α-diversity of the intestinal microbiota of piglets, which is similar to the effect of antibiotics, and this may be due to the inhibitory effect of probiotics on the pathogens and the regulation of the intestinal microbiota [17]. The results of this experiment showed that the supplemental feeding of FM could reduce the richness and diversity of fecal microbial communities in weaned piglets, without affecting the diversity estimation of fecal microbial communities, which may be due to the fact that FM contains a large number of lactobacilli as the main beneficial bacteria and thus occupies ecological positions in the intestine, reflecting that the addition of lactobacilli to the FM can improve the structure of the microbial communities in the intestine of the piglets, reduce the number of harmful bacteria, and maintain the intestinal structure of the intestinal microbial community of piglets.
In this experiment, supplemental feeding of FM could significantly increase the relative abundance of Lactobacillus and Hordelmannia in the fecal microbiota of suckling piglets and decrease the relative abundance of Erysipelotrichaceae UCG-003 and Intestinimonas. Lactobacillus has been reported to have a strong effect on the overall microbial composition and fecal microbiota maturation of piglets and to promote performance and resistance to diarrhea [38]. Erysipelotrichaceae UCG-003 have a high abundance in the mammalian intestinal tract and have been reported to be associated with host metabolic disorders and inflammatory diseases, which can be induced by a reduction in the number of adherent bacteria that It can cause bacterial overgrowth by reducing the number of adherent bacteria, which induces inflammatory T-cells, leading to intestinal inflammation [39]. Intestinimonas is a conditionally pathogenic bacterium and acts as a pro-inflammatory microorganism, causing an increase in inflammatory mediators released by immune cells. The damage leads to intestinal mucosal shielding and immune dysfunction in pigs, leading to diarrhea [40]. Intestinal inflammation has been similarly linked to the production of the Erysipelotrichaceae UCG-003, which is positively correlated [41]. This is consistent with the results of the present experiment, in which Intestinimonas and Erysipelotrichaceae UCG-003 were significantly and positively correlated with diarrhea in piglets prior to weaning, and supplementation with FM was able to reduce the concentration of Intestinimonas and Erysipelotrichaceae UCG-003 in the fecal microbiota of piglets. The relative abundance of p-251-o5_norank, Campylobacter and Fusobacterium in the fecal microbiota of piglets was reduced by supplemental FM after piglets were weaned. Campylobacter is reported to be one of the most common bacteria causing diarrhea worldwide, inducing microbiota dysbiosis and metabolic dysfunction, causing intestinal dysfunction [42]; Fusobacterium causes intestinal inflammation and mucosal damage, which in turn increases intestinal permeability, making it easier for harmful substances to pass through the intestinal wall, triggering symptoms such as diarrhea [43]. In this experiment, p-251-o5_norank was the genus with the highest relative abundance in the level of fecal microbiota genus of weaned piglets in the CON group, and its relative abundance was significantly and positively correlated with the rate of diarrhea and inflammation in weaned piglets. In conclusion, supplemental feeding of fermented milk could promote the increase of the relative abundance of beneficial intestinal microbiota, inhibit the growth and reproduction of harmful microbiota, such as Campylobacter and Clostridium difficile, regulate the intestinal microecological balance, and then improve the intestinal environment of weaned piglets. A major limitation of the present study is the absence of functional assays linking FM-induced shifts in microbial taxa to intestinal barrier integrity and immune cell activity. To address this, future work should employ targeted metabolomic analysis of key microbial metabolites (e.g., short-chain fatty acids, tryptophan catabolites) and quantify epithelial tight-junction gene expression (e.g., CLDN1, OCLN) or permeability markers (e.g., serum zonulin). Moreover, absolute qPCR using strain-specific primers for each of the three probiotics would verify successful colonization and allow correlation of probiotic load with changes in both resident community structure and host endpoints. Such studies will be essential to establish causal mechanistic links between probiotic supplementation, microbial function, and host physiology.
5. Conclusions
In summary, the results showed that the addition of FM had positive effects on the growth performance, immunity, and intestinal microbiota of piglets, and these findings provide new ideas for the use of probiotic-fermented milk to ameliorate weaning stress in piglets under actual commercial conditions.
Author Contributions
Conceptualization, Y.S. and Y.L.; methodology, H.Z., Y.R. and D.T.; software, H.Z. and Y.S.; formal analysis, H.Z. and Y.R.; investigation, H.Z., Y.R. and D.T.; data curation, H.Z. and Y.R.; writing—original draft preparation, H.Z. and Y.R.; writing—review and editing, Y.S. and Y.L.; visualization, Y.R.; supervision, Y.S. and Y.L.; project administration, Y.S.; funding acquisition, Y.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Key R&D Program of China (2022YFD1300402).
Institutional Review Board Statement
This work was approved by the Animal Care and Use Committee of Nanjing Agricultural University (SYXK2019-0066) and implemented based on the standard of Experimental Animal Care and Use Guidelines of China (EACUGC2018-01).
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in the study are included in the article, further inquiries can be directed to the author.
Conflicts of Interest
D.T. is from Shangdang Lvzhou Ecological Pig Farm. The farm had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results. The other authors declare no conflicts of interest.
References
- Thompson, L. H., Hanford, K. J. & Jensen, A. H. Estrus and fertility in lactating sows and piglet performance as influenced by limited nursing. J Anim Sci 53, 1419-1423 (1981). https://doi.org/10.2527/jas1982.5361419x. [CrossRef]
- Meng, Q. et al. Weaning Alters Intestinal Gene Expression Involved in Nutrient Metabolism by Shaping Gut Microbiota in Pigs. Front Microbiol 11, 694 (2020). https://doi.org/10.3389/fmicb.2020.00694. [CrossRef]
- Gresse, R. et al. Gut Microbiota Dysbiosis in Postweaning Piglets: Understanding the Keys to Health. Trends Microbiol 25, 851-873 (2017). https://doi.org/10.1016/j.tim.2017.05.004. [CrossRef]
- Xiong, X. et al. Nutritional Intervention for the Intestinal Development and Health of Weaned Pigs. Front Vet Sci 6, 46 (2019). https://doi.org/10.3389/fvets.2019.00046. [CrossRef]
- Long, S. et al. Mixed organic acids as antibiotic substitutes improve performance, serum immunity, intestinal morphology and microbiota for weaned piglets. Animal Feed Science and Technology 235, 23-32 (2018).
- Houdijk, J. G. M., Bosch, M. W., Verstegen, M. W. A. & Berenpas, H. J. Effects of dietary oligosaccharides on the growth performance and faecal characteristics of young growing pigs. Animal Feed Science and Technology 71, 35-48 (1998).
- Tian, S., Wang, J., Yu, H., Wang, J. & Zhu, W. Changes in Ileal Microbial Composition and Microbial Metabolism by an Early-Life Galacto-Oligosaccharides Intervention in a Neonatal Porcine Model. Nutrients 11 (2019). https://doi.org/10.3390/nu11081753. [CrossRef]
- Wang, J., Tian, S., Yu, H., Wang, J. & Zhu, W. Response of Colonic Mucosa-Associated Microbiota Composition, Mucosal Immune Homeostasis, and Barrier Function to Early Life Galactooligosaccharides Intervention in Suckling Piglets. J Agric Food Chem 67, 578-588 (2019). https://doi.org/10.1021/acs.jafc.8b05679. [CrossRef]
- Karimi, R., Mortazavian, A. M. & Cruz, A. G. Viability of probiotic microorganisms in cheese during production and storage: a review. Dairy Sci Technol 91, 283-308 (2011).
- Lin, A., Yan, X., Wang, H., Su, Y. & Zhu, W. Effects of lactic acid bacteria-fermented formula milk supplementation on ileal microbiota, transcriptomic profile, and mucosal immunity in weaned piglets. J Anim Sci Biotechnol 13, 113 (2022). https://doi.org/10.1186/s40104-022-00762-8. [CrossRef]
- Rateliffe, B., Cole, C. B., Fuller, R. & Newport, M. J. The effect of yoghurt and milk fermented with a porcine intestinal strain of Lactobacillus reuteri on the performance and gastrointestinal flora of pigs weaned at two days of age. Food Microbiology 3, 203-211 (1986).
- Marquardt, R. R. et al. Passive protective effect of egg-yolk antibodies against enterotoxigenic Escherichia coli K88+ infection in neonatal and early-weaned piglets. FEMS Immunol Med Microbiol 23, 283-288 (1999). https://doi.org/10.1111/j.1574-695X.1999.tb01249.x. [CrossRef]
- Dai, Z. L., Zhang, J., Wu, G. & Zhu, W. Y. Utilization of amino acids by bacteria from the pig small intestine. Amino Acids 39, 1201-1215 (2010). https://doi.org/10.1007/s00726-010-0556-9. [CrossRef]
- Chen, J. et al. Dietary chlorogenic acid improves growth performance of weaned pigs through maintaining antioxidant capacity and intestinal digestion and absorption function. J Anim Sci 96, 1108-1118 (2018). https://doi.org/10.1093/jas/skx078. [CrossRef]
- Chen, S. et al. Differential Effects of Lactobacillus casei Strain Shirota on Patients With Constipation Regarding Stool Consistency in China. J Neurogastroenterol Motil 25, 148-158 (2019). https://doi.org/10.5056/jnm17085. [CrossRef]
- Peters, A. et al. Metabolites of lactic acid bacteria present in fermented foods are highly potent agonists of human hydroxycarboxylic acid receptor 3. PLoS Genet 15, e1008145 (2019). https://doi.org/10.1371/journal.pgen.1008145. [CrossRef]
- Wang, S. et al. Combined supplementation of Lactobacillus fermentum and Pediococcus acidilactici promoted growth performance, alleviated inflammation, and modulated intestinal microbiota in weaned pigs. BMC Vet Res 15, 239 (2019). https://doi.org/10.1186/s12917-019-1991-9. [CrossRef]
- De Filippis, F., Pasolli, E. & Ercolini, D. The food-gut axis: lactic acid bacteria and their link to food, the gut microbiome and human health. FEMS Microbiol Rev 44, 454-489 (2020). https://doi.org/10.1093/femsre/fuaa015. [CrossRef]
- Griet, M. et al. Soluble factors from Lactobacillus reuteri CRL1098 have anti-inflammatory effects in acute lung injury induced by lipopolysaccharide in mice. PLoS One 9, e110027 (2014). https://doi.org/10.1371/journal.pone.0110027. [CrossRef]
- Woo, I. K., Hyun, J. H., Jang, H. J., Lee, N. K. & Paik, H. D. Probiotic Pediococcus acidilactici Strains Exert Anti-inflammatory Effects by Regulating Intracellular Signaling Pathways in LPS-Induced RAW 264.7 Cells. Probiotics Antimicrob Proteins (2024). https://doi.org/10.1007/s12602-024-10263-x. [CrossRef]
- Lin, A. et al. Effects of lactic acid bacteria-fermented formula milk supplementation on colonic microbiota and mucosal transcriptome profile of weaned piglets. Animal 17, 100959 (2023). https://doi.org/10.1016/j.animal.2023.100959. [CrossRef]
- Liu, H. et al. Protective approaches and mechanisms of microencapsulation to the survival of probiotic bacteria during processing, storage and gastrointestinal digestion: A review. Crit Rev Food Sci Nutr 59, 2863-2878 (2019). https://doi.org/10.1080/10408398.2017.1377684. [CrossRef]
- Guarner, F. & Malagelada, J. R. Gut flora in health and disease. Lancet 361, 512-519 (2003). https://doi.org/10.1016/s0140-6736(03)12489-0. [CrossRef]
- Lallès, J. P., Bosi, P., Smidt, H. & Stokes, C. R. Nutritional management of gut health in pigs around weaning. Proc Nutr Soc 66, 260-268 (2007). https://doi.org/10.1017/s0029665107005484. [CrossRef]
- Conway, S., Hart, A., Clark, A. & Harvey, I. Does eating yogurt prevent antibiotic-associated diarrhoea? A placebo-controlled randomised controlled trial in general practice. Br. J. Gen. Pract. 57, 953-959 (2007). https://doi.org/10.3399/096016407782604811. [CrossRef]
- Ouyang, W., Rutz, S., Crellin, N. K., Valdez, P. A. & Hymowitz, S. G. Regulation and functions of the IL-10 family of cytokines in inflammation and disease. Annu Rev Immunol 29, 71-109 (2011). https://doi.org/10.1146/annurev-immunol-031210-101312. [CrossRef]
- Pourcyrous, M., Nolan, V. G., Goodwin, A., Davis, S. L. & Buddington, R. K. Fecal short-chain fatty acids of very-low-birth-weight preterm infants fed expressed breast milk or formula. J Pediatr Gastroenterol Nutr 59, 725-731 (2014). https://doi.org/10.1097/mpg.0000000000000515. [CrossRef]
- Sangild, P. T., Vonderohe, C., Melendez Hebib, V. & Burrin, D. G. Potential Benefits of Bovine Colostrum in Pediatric Nutrition and Health. Nutrients 13 (2021). https://doi.org/10.3390/nu13082551. [CrossRef]
- Yu, X. et al. Protective effects of Bacillus licheniformis on growth performance, gut barrier functions, immunity and serum metabolome in lipopolysaccharide-challenged weaned piglets. Front Immunol 14, 1140564 (2023). https://doi.org/10.3389/fimmu.2023.1140564. [CrossRef]
- Mahony, J., McDonnell, B., Casey, E. & van Sinderen, D. Phage-Host Interactions of Cheese-Making Lactic Acid Bacteria. Annu Rev Food Sci Technol 7, 267-285 (2016). https://doi.org/10.1146/annurev-food-041715-033322. [CrossRef]
- Jintao, W. Effects of Fermented Feed on Growth Performance,Plasma Biochemical Parameters and Digestibility of Dietary Components in Weaned Piglets. Journal of the Chinese Cereals and Oils Association (2009).
- Jinlong, Z. et al. Effects of fermented product of mulberry leaf on growth performance, serum biochemical indexes and intestinal morphology of broiler chickens. Feed Industry (2017).
- Yu, X. & Yingting, L. Research Progress of Oligosaccharides on Intestinal Flora in Animals. Feed Review (2011).
- Ramadan, Z. et al. Fecal microbiota of cats with naturally occurring chronic diarrhea assessed using 16S rRNA gene 454-pyrosequencing before and after dietary treatment. J. Vet. Intern. Med. 28, 59-65 (2014). https://doi.org/10.1111/jvim.12261. [CrossRef]
- Zhang, D. et al. Changes in the diversity and composition of gut microbiota of weaned piglets after oral administration of Lactobacillus or an antibiotic. Appl Microbiol Biotechnol 100, 10081-10093 (2016). https://doi.org/10.1007/s00253-016-7845-5. [CrossRef]
- Hai-Feng, W., Hai-Xia, C., Wen-Ming, Z. & Jian-Xin, L. Effects of Compound Acidifiers on Growth Performance and Intestinal Microbial Communities of Weaned Piglets. Chin J Anim Sci (2011).
- Mishra, D. K., Verma, A. K., Agarwal, N., Mondal, S. K. & Singh, P. Effect of Dietary Supplementation of Probiotics on Growth Performance, Nutrients Digestibility and Faecal Microbiology in Weaned Piglets. Animal Nutrition & Feed Technology 14, 283 (2014).
- Wang, G. et al. Lactobacillus reuteri improves the development and maturation of fecal microbiota in piglets through mother-to-infant microbe and metabolite vertical transmission. Microbiome 10, 211 (2022). https://doi.org/10.1186/s40168-022-01336-6. [CrossRef]
- Wu, J. et al. Isolation and genomic characterization of five novel strains of Erysipelotrichaceae from commercial pigs. Bmc Microbiol 21, 125 (2021). https://doi.org/10.1186/s12866-021-02193-3. [CrossRef]
- Hasan, S. et al. Late gestation diet supplementation of resin acid-enriched composition increases sow colostrum immunoglobulin G content, piglet colostrum intake and improve sow gut microbiota. Animal 13, 1599-1606 (2019). https://doi.org/10.1017/s1751731118003518. [CrossRef]
- Yan, Y. et al. A Combination of Baicalin and Berberine Hydrochloride Ameliorates Dextran Sulfate Sodium-Induced Colitis by Modulating Colon Gut Microbiota. J Med Food 25, 853-862 (2022). https://doi.org/10.1089/jmf.2021.K.0173. [CrossRef]
- Allos, B. M. Campylobacter jejuni Infections: update on emerging issues and trends. Clin Infect Dis 32, 1201-1206 (2001). https://doi.org/10.1086/319760. [CrossRef]
- Sun, C., Song, R., Zhou, J., Jia, Y. & Lu, J. Fermented Bamboo Fiber Improves Productive Performance by Regulating Gut Microbiota and Inhibiting Chronic Inflammation of Sows and Piglets during Late Gestation and Lactation. Microbiol Spectr 11, e0408422 (2023). https://doi.org/10.1128/spectrum.04084-22. [CrossRef]
Figure 1.
Effects of supplementing fermented milk on immunoglobulin of suckling and weaning piglets. (A) Serum IgA content, (B) Serum IgG content, with * indicating p < 0.05.
Figure 1.
Effects of supplementing fermented milk on immunoglobulin of suckling and weaning piglets. (A) Serum IgA content, (B) Serum IgG content, with * indicating p < 0.05.
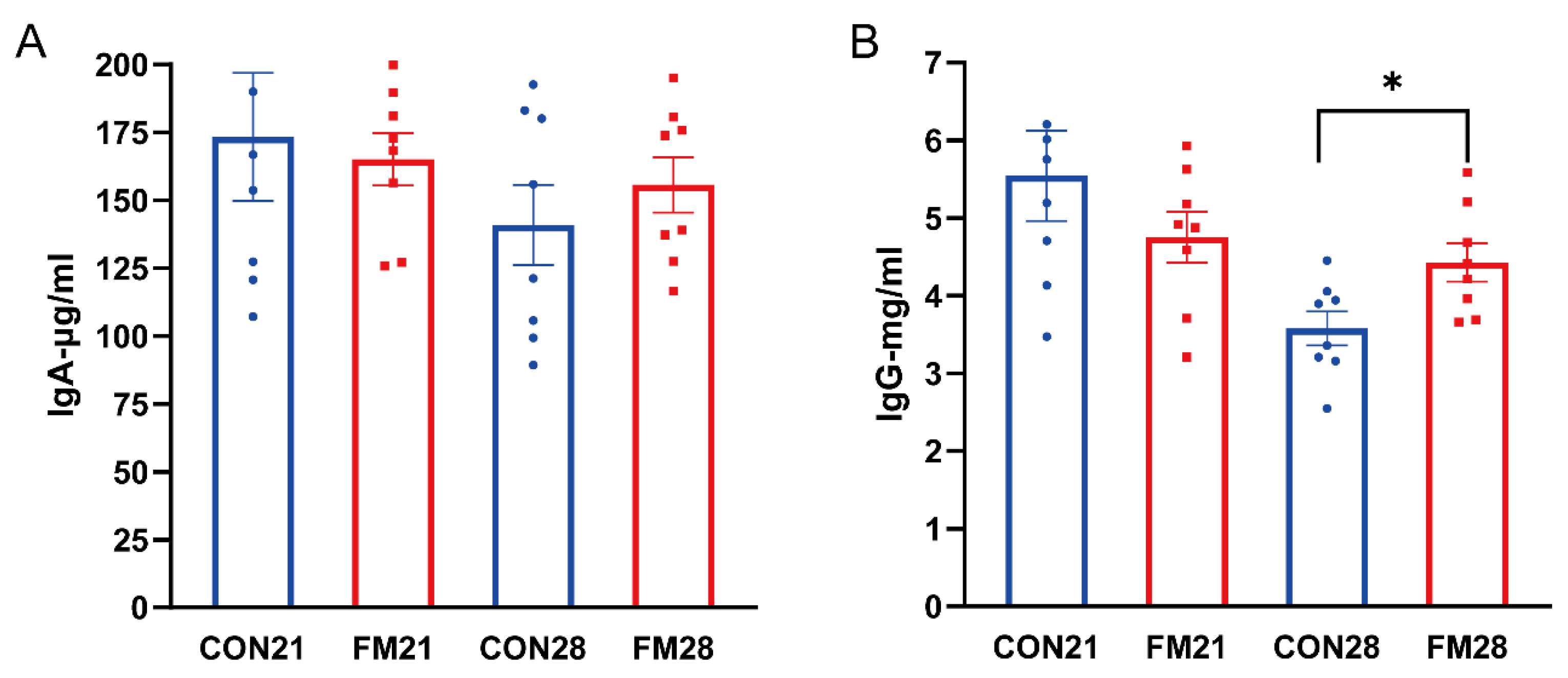
Figure 2.
Microbial composition in feces of piglets. (A-C) α-diversity indices of fecal microbiota of piglets on day 21, day 24 and day 28, respctively; (D-F) Principal component analysis of fecal microbiota of piglets on day 21, day 24 and day 28, respctively; (GHI) Relative abundance of fecal microbiota at the phylum, family and genus levels, with * indicating P < 0.05.
Figure 2.
Microbial composition in feces of piglets. (A-C) α-diversity indices of fecal microbiota of piglets on day 21, day 24 and day 28, respctively; (D-F) Principal component analysis of fecal microbiota of piglets on day 21, day 24 and day 28, respctively; (GHI) Relative abundance of fecal microbiota at the phylum, family and genus levels, with * indicating P < 0.05.
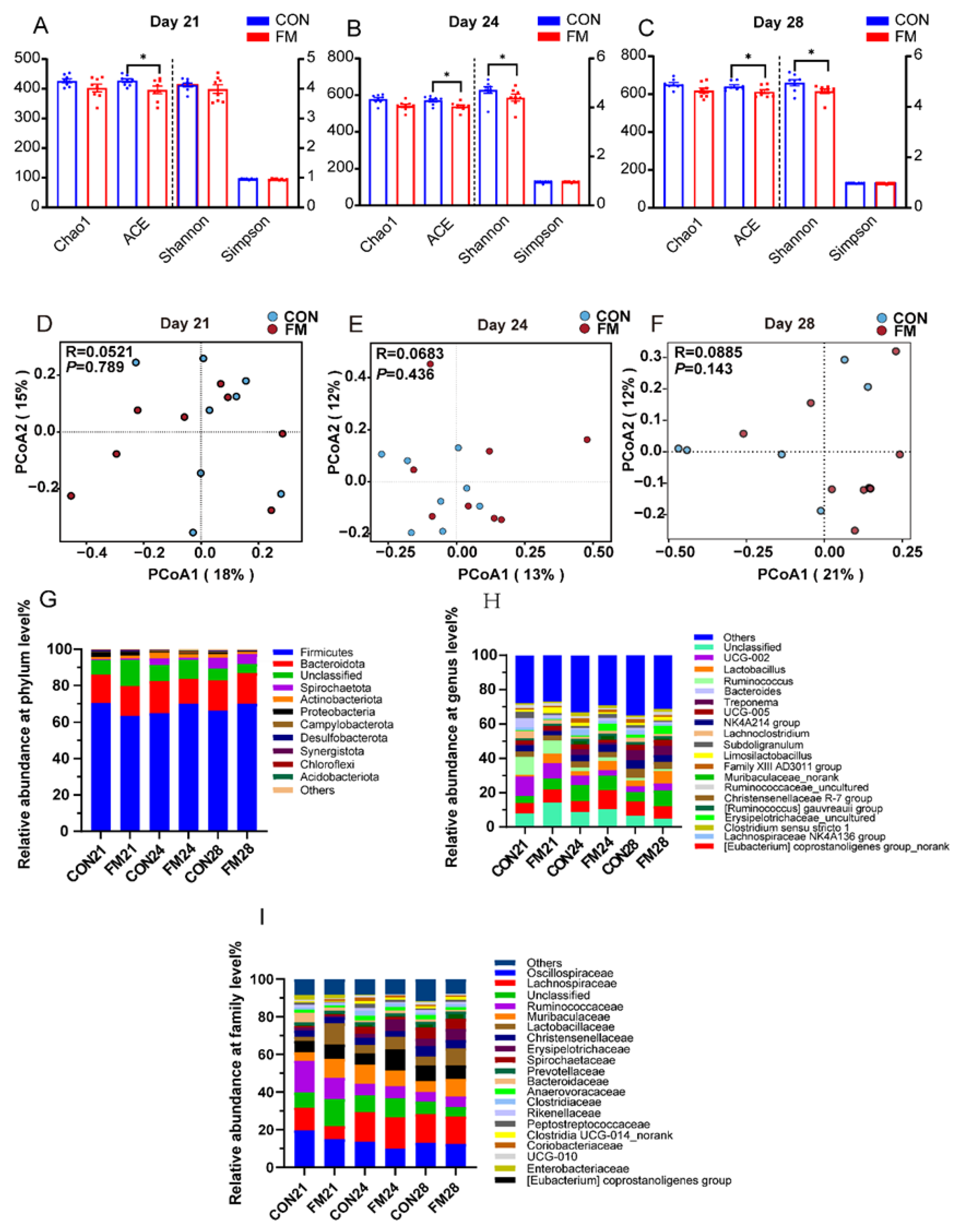
Figure 3.
LEfSe analysis of fecal microbiota. (A, C, E) Histogram of the distribution of LDA values of fecal microbiota of piglets on day 21, day 24 and day 28, respctively; (B, D, F) Evolutionary branching of fecal microbiota of piglets on day 21, day 24 and day 28, respctively; (G, H, I) Relative abundance of fecal microbiota with significant differences between the control (CON) and fermented milk (FM) groups on day 21 day 24 and day 28, respctively (LDA Score>2, p < 0.05).
Figure 3.
LEfSe analysis of fecal microbiota. (A, C, E) Histogram of the distribution of LDA values of fecal microbiota of piglets on day 21, day 24 and day 28, respctively; (B, D, F) Evolutionary branching of fecal microbiota of piglets on day 21, day 24 and day 28, respctively; (G, H, I) Relative abundance of fecal microbiota with significant differences between the control (CON) and fermented milk (FM) groups on day 21 day 24 and day 28, respctively (LDA Score>2, p < 0.05).
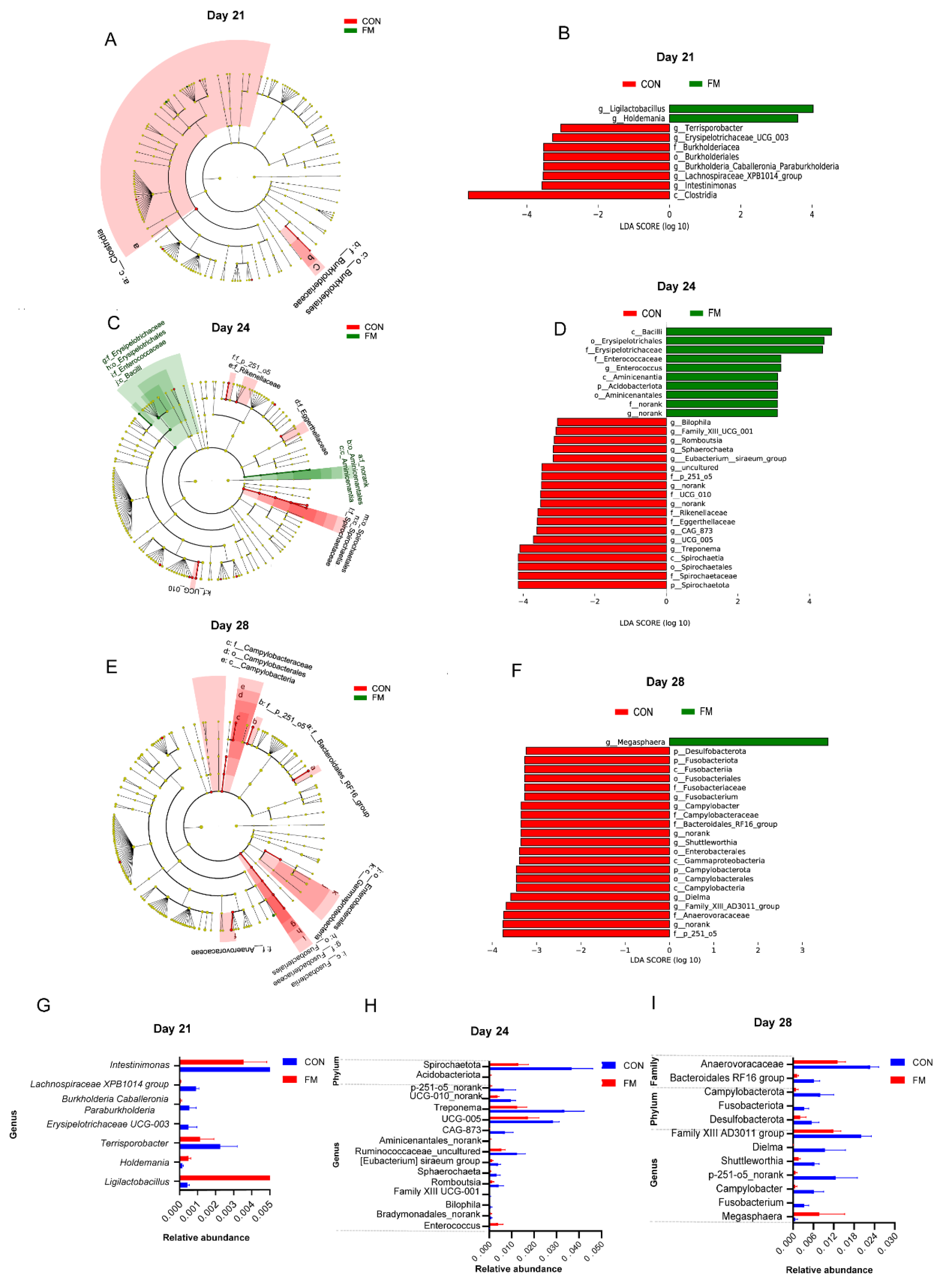
Figure 4.
Correlation analysis of fecal microbiota with differential phenotypic indicators in piglets on day 21 (A) and day 28 (B). * indicates significant correlation (p < 0.05); ** indicates highly significant correlation (p<0.01).
Figure 4.
Correlation analysis of fecal microbiota with differential phenotypic indicators in piglets on day 21 (A) and day 28 (B). * indicates significant correlation (p < 0.05); ** indicates highly significant correlation (p<0.01).
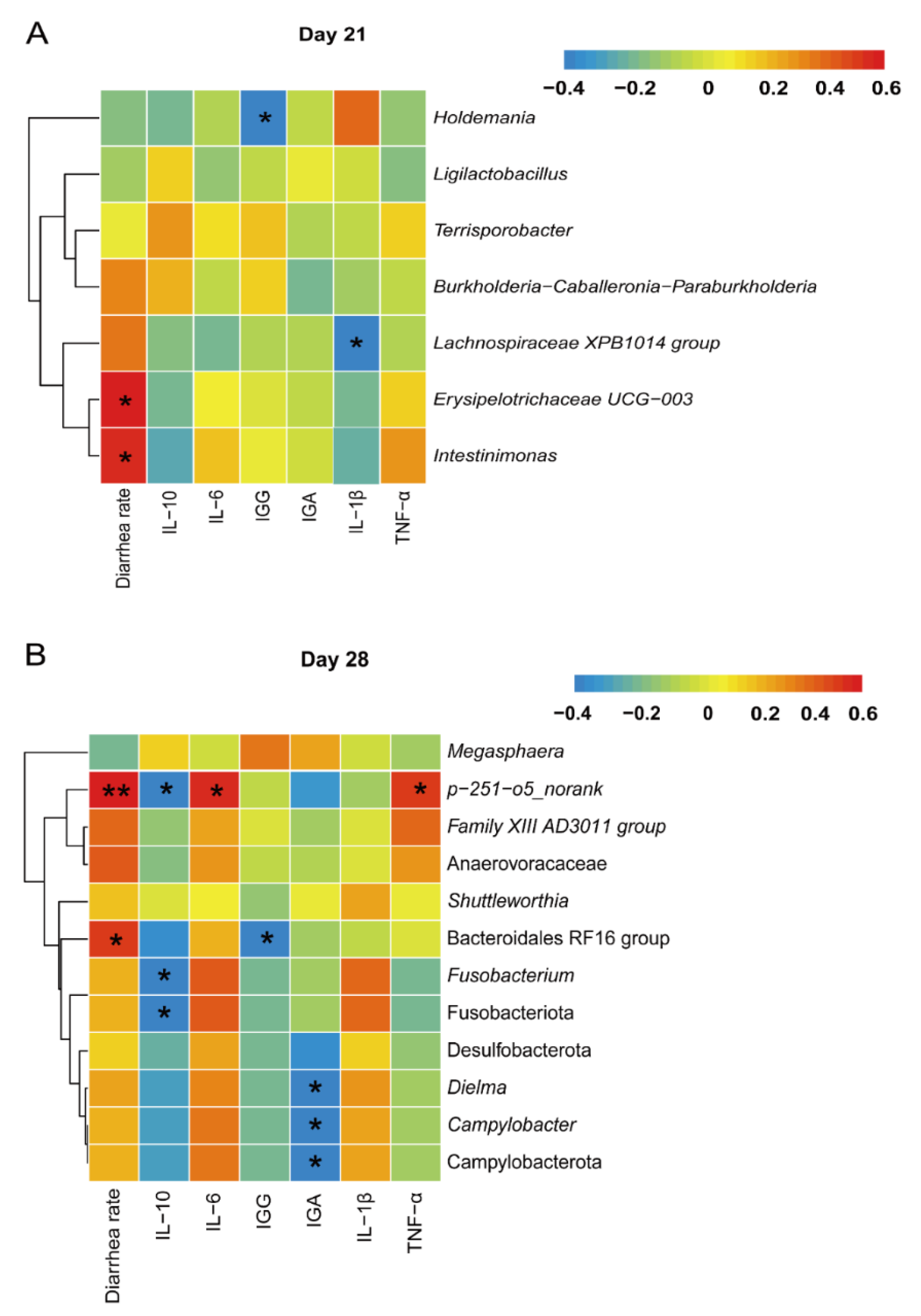

Table 1.
Growth performance of suckling and weaning piglets in the CON and FM groups.
| Items | CON | FM | P-value | |
|---|---|---|---|---|
| BW (kg) | Day 7 | 2.71 ± 0.14 | 2.99 ± 0.09 | 0.114 |
| Day 14 | 4.40 ± 0.17 | 4.58 ± 0.10 | 0.563 | |
| Day 21 | 6.25 ± 0.18 | 6.44 ± 0.16 | 0.450 | |
| Day 28 | 6.16 ± 0.23 | 6.65 ± 0.134 | 0.099 | |
| ADG (g) | Days 7-14 | 247.98 ± 11.94 | 226.61 ± 11.39 | 0.222 |
| Days 15-21 | 257.03 ± 10.68 | 264.97 ± 14.47 | 0.658 | |
| Days 22-28 | -10.95 ± 12.41 | 30.82 ± 11.27 | 0.027 | |
| ADFI (g) | Days 7-14 | 1.08 ± 0.07 | 2.43 ± 0.19 | < 0.001 |
| Days 15-21 | 3.18 ± 0.36 | 5.13 ± 0.37 | 0.002 | |
| Days 22-28 | 7.85 ± 0.79 | 10.52 ± 2.42 | 0.038 | |
CON, control group; FM, lactic acid bacteria-fermented milk group; n = 8; BW, body weight; ADG, average daily gain; ADFI, average daily feed intake.
Table 2.
Indicators of diarrhea in suckling and weaning piglets in the CON and FM groups.
| Items | CON | FM | P-value | |
|---|---|---|---|---|
| Diarrhea rate (%) | Days 7-14 | 1.09 ± 0.45 | 1.8 ± 1.8 | 0.092 |
| Days 15-21 | 3.63 ± 1.18 | 2.06 ± 1.01 | 0.331 | |
| Days 22-28 | 10.99 ± 2.32 | 4.76 ± 0.07 | 0.032 | |
| Diarrhea index | Days 7-14 | 0.036 ± 0.016 | 0.004 ± 0.004 | 0.079 |
| Days 15-21 | 0.109 ± 0.035 | 0.062 ± 0.030 | 0.329 | |
| Days 22-28 | 0.330 ± 0.069 | 0.141 ± 0.73 | 0.031 | |
CON, control group; FM, lactic acid bacteria-fermented milk group; n = 8.
Table 3.
Apparent digestibility (%) of nutrients in suckling and weaning piglets in the CON and FM groups.
Table 3.
Apparent digestibility (%) of nutrients in suckling and weaning piglets in the CON and FM groups.
| Items | CON | FM | P-value | |
|---|---|---|---|---|
| Days 18-21 | DM | 75.85 ± 1.00 | 73.91 ± 1.66 | 0.256 |
| CP | 71.07 ± 0.89 | 69.22 ± 1.58 | 0.078 | |
| EE | 66.01 ± 2.97 | 64.73 ± 1.45 | 0.065 | |
| CF | 33.08 ± 2.08 | 31.68 ± 3.15 | 0.366 | |
| Ash | 47.83 ± 2.56 | 42.78 ± 2.14 | 0.420 | |
| Days 25-28 | DM | 74.18 ± 1.26 | 74.05 ± 1.17 | 0.136 |
| CP | 67.18 ± 2.48 | 66.54 ± 3.17 | 0.589 | |
| EE | 66.56 ± 1.03 | 63.46 ± 2.78 | 0.062 | |
| CF | 31.95 ± 0.94 | 31.76 ± 1.26 | 0.469 | |
| Ash | 44.99 ± 2.41 | 41.45 ± 3.00 | 0.549 | |
CON, control group; FM, lactic acid bacteria-fermented formula milk supplementation group; n = 8; DM, dry matter; CP, crude protein; EE, ether extract; CF, crude fibre; Ash, crude ash.
Table 4.
Indicators of serum inflammation in suckling and weaning piglets in the CON and FM groups.
| Items | CON | FM | P-value | |
|---|---|---|---|---|
| Day 21 | IL-1β (pg/mL) | 8.14 ± 0.60 | 11.95 ± 1.19 | 0.024 |
| IL-6 (pg/mL) | 77.28 ± 30.27 | 62.49 ± 7.49 | 0.643 | |
| IL-10 (pg/mL) | 16.88 ± 1.72 | 16.98 ± 1.13 | 0.963 | |
| TNF-α (pg/mL) | 11.08 ± 1.69 | 9.09 ± 0.57 | 0.284 | |
| Day 28 | IL-1β (pg/mL) | 10.64 ± 0.61 | 9.66 ± 0.83 | 0.355 |
| IL-6 (pg/mL) | 77.09 ± 8.59 | 53.06 ± 5.82 | 0.036 | |
| IL-10 (pg/mL) | 6.11 ± 2.93 | 14.48 ± 1.18 | 0.019 | |
| TNF-α (pg/mL) | 9.87 ± 1.51 | 10.09 ± 0.65 | 0.894 | |
CON, control group; FM, lactic acid bacteria-fermented formula milk supplementation group; n = 8; IL-1β, Interleukin-1β; IL-6, Interleukin-6; IL-10, Interleukin-10; TNF-α, tumor necrosis factor.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



