
Submitted:
26 June 2025
Posted:
27 June 2025
You are already at the latest version
Abstract
After decades of use of neonicotinoid insecticides, it is proven that sublethal doses have behavioural and physiological effects on both Apis mellifera honey bee individuals and colonies. This work aimed a) to ring test the methodology of digital measurement of hypopharyngeal glands (HPGs) of honey bees and to validate the method against the subjectivity of the assessors and b) to assess possible differences between laboratory and field studies when examining the impact of sub-lethal levels of the neonicotinoid imidacloprid on worker honey bee HPGs size. We did this by employing young honey bees reared in colonies that were fed for one month with sugar solution and pollen paste, both contaminated with imidacloprid at concentrations of 2 ng/kg and 3 ng/kg, respectively. Newly emerged bees were marked and then maintained as adults in either their own colonies or in laboratory cages. Digital measurements of HPG acini diameter were performed by 10 independent assessors using the same software at days 10 and 15 post-emergence. We found that HPGs glands have a significantly greater diameter under field conditions than in laboratory conditions, and that fifteen-day-old bees had smaller acini diameter compared to 10-day-old workers when reared in the laboratory but not in actual colonies. Imidacloprid had a significant negative effect on acini diameter of 10-day old workers, both under laboratory and field conditions, but not on 15-day old workers in both laboratory and field conditions. The reliability of measurements undertaken by all assessors was greater than 95% and there was no significant difference among the assessors. In conclusion, we found that digital measurement of HPG acini was a highly reliable method for detecting changes in the development of the glands and followed the same pattern under laboratory and field conditions, always independently by human error. Furthermore, the digital measurement of the HPG diameter at day 10 of the bees' life is a very reliable tool to be used for lower tier risk assessment and extrapolated for higher tier assessment.
Keywords:
apis mellifera
; ring testing
; lab
; field
; neonicotinoid
; sublethal
; hypopharyngeal gland
; assessor
1. Introduction
The honey bee (Apis mellifera L.) is regarded as one of the most important social insects in the world, not only for its pollination services [1,2,3] but also for its economic value of beekeeping products. However, dramatic and worrying losses of honey bee colonies have been observed worldwide over the last decade [4,5,6]. It is believed that multiple causes are behind colony losses [7,8,9,10]. For example, in recent years pesticides, in particular neonicotinoids, have been blamed for elevated losses [11,12,13,14,15,16,17]. Honeybees can be exposed to neonicotinoids in several ways: consumption of contaminated nectar and pollen [18], contact with pesticides exuded from plants, and exposure to contaminated air during sowing [19,20,21,22]. Through a nation-wide biomonitoring study with bees, imidacloprid was found to widely contaminate bee-collected pollen samples (12% of positive samples, N = 554) with high concentrations (average = 2 μg/kg, max = 19 μg/kg), leading to detrimental risk for honeybees (Hazard Quotient = 5054, higher than the 1000 safety threshold) [18]. Residues found in pollen was usually higher than those found in nectar (e.g., imidacloprid residues in treated sunflowers were 1.9 μg/kg in nectars and 3.3 μg/Kg in pollen [23] and from maize dressed seeds ranged between 1 to11 μg/kg [24]. Residues of imidacloprid and its metabolite 6-chloronic have been found in pollen pellets in concentrations that ranged from 1.1 to 5.7 μg/kg and from 0.6 to 9.3 μg/kg, respectively [25]. In some cases, exudes from plants germinated from neonicotinoid- coated seeds can contain insecticide residues as high as 200 mg/l for imidacloprid [21,26].
Concentrations in nectar and pollen of below 10 μg/kg usually induces intoxication which does not result in lethal consequences, but rather impairs behaviour and physiology of individual honey bees [27], and explecity learning and orientation [28,29,30,31], dance communication [28], flight [29], foraging activity [30,31], homing behaviour [32], neurophysiology [33,34] and reproduction [35,36]. Negative effects have even been seen at the colony level [37]. More specifically, the negative effects on tissue development, such as worker hypopharyngeal glands (HPGs), which produce secretions used to nourish developing bees [38] are regarded as an indicator of colony failure [10,33] or bad nutrition for the colony and the queen [14,39].
HPG development begins very early in the life of the adult worker honey bees, reaching their full development at nursing age of 8–12 days old [40,41]. Afterwards, they decrease in size and their function changes, producing digestive enzymes like invertase and glucose oxidase [42]. Thus, HPGs are important not only to colony growth but also for food processing. Size of HPGs typically refers to the size of the gland lobes - the acini. According to [41] acini diameter reflects the amount of proteins produced in the glands. A decrease in intake pollen by workers may result in development of smaller or even atrophic HPGs [43,44], and therefore less protein production and reared brood [45]. Several researchers have shown that chemical stressors, have a decreasing effect on acini diameter of the HPGs [10,33,46] by using an optical microscopic methodology to assess acini size. Based on this, the methodology of measuring the diameter of the acini has been used by many researchers in the past, examining the functional status of the HPGs and assessing the effects of pesticides and food supplies on physiological effects [33,47,48].
Wilde et al., [14] showed that the protein profiles of tissue extracts from worker A. mellifera were significantly reduced after the administration of sublethal doses of imidacloprid. Because several of these proteins belong to the same group of major proteins as royal jelly, these reduced protein fractions likely signify reduced quality of royal jelly. Furthermore, De Smet et al., [34] also demonstrated a context-dependent effects of imidacloprid exposure on honey bees, where increased levels of the detoxification genes encoding, CYP9Q3 and CYT P450, were correlated with decreased HPGs acini diameters.
The importance of the status of HPGs for risk assessment of pesticides was recognized in the Bee Guidance Document [49], where the implementation of a simple and cost-effective first tier was suggested for adult workers. The use of HPG size or function was suggested as an approximation to brood care studies which could not be performed in a lower tier assessment. The objective of this work was to evaluate the microscopic/ digital measurement of acini diameter recommended by the Bee Guidance Document by performing a ring test to be applied collectively in the future for lower tier risk assessment purposes. We assessed possible differences between laboratory and field studies when examining the impact of the stressor imidacloprid on worker honey bees’ HPGs development; we also evaluated operator (assessor) variability when employing digital measuring.
2. Materials and Methods
2.1. Experimental Set-Up and Pesticide Exposure
All honeybees used in this study were Apis mellifera macedonica originally collected from their natural distribution area and have previously been confirmed by MtDNA analysis. Two honey bee colonies were used from the apiary placed in Nea Moudania, Greece during the early summer and each was assigned to the control or insecticide treatment group. The insecticide colony was administered imidacloprid weekly, and for 4 weeks. Pure analytical imidacloprid was added to the sugar solution and pollen paste, (after it was diluted well in water) in a concentration of 2 ng/kg and 3 ng/kg, respectively (as average residue concentrations); 500 ml of sugar solution and 150 g of pollen paste was given once a week. Small amounts of food were provided to the colonies therefore they had the chance to also forage elsewhere (as not all bees of a colony are foraging on the same source). After 30 days newly emerged honey bees from the treatment colony as well as from the uncontaminated one (control) were subjected to a further feeding under laboratory conditions. One brood frame per colony was transferred to a laboratory incubator (set to 34.5˚C and 70% RH) to collect emerging worker bees. A number of 100 bees were marked and returned back to their perspective colony. A number of bees were also collected and kept in cages for the laboratory assay; bees from the treatment colony were also given imidacloprid contaminated food during the laboratory rearing period. Control bees, collected from control colonies, were kept under the same conditions as the treated ones but fed uncontaminated food.
The laboratory assay followed the description of Hatjina et al., [33] where wooden, mesh sided cages (10×10×10 cm) with removable metal floor sheets that facilitated cleaning, were used accommodating about 60 bees. The bees were fed sugar solution (33 %w/v) via gravity feeders and pollen pastry on pre-weighted plastic plates, both ad libitum, with new food added every second day and with the same concentration of imidacloprid as fed the colonies. Pollen paste was given to the bees only till day 10, as it has been shown before that after day 10 of their life they consume very small amounts of pollen [33]. Day 10 was also chosen to sample the bees and see the difference on the development of HPGs. Three cages were used for imidacloprid treated bees and three cages for control/ untreated bees. The cages were kept in an incubator in darkness at 34–35 °C and relative humidity of about 70 %.
2.2. HPGs Dissection and Measurement
At day 10 and 15 post marking and caging, a sample of 5 alive honey bees from each cage from the laboratory were collected and a sample of 5 marked alive honey bees from both colonies were captured; all sample bees were kept in the freezer at -20o C till analysis. Eight (8) groups of samples bees were resulted from the experimental set up each consisted of 5 bees: 10 days control lab, 10 days imidacloprid lab, 15 days control lab, 15 days imidacloprid lab, 10 days control field, 10 days imidacloprid field, 15 days control field and 15 days imidacloprid field.
The sampled bees were decapitated under a dissecting microscope in insect saline solution consisting of: 7.5g NaCl, 2.38 g Na2HPO4, 2.72g KH2PO4, dissolved in 1L of distilled water [50]. The removed parts of the HPGs were placed in a mixture of the electrophoresis stain made by using 0.1 g of Coomassie Brilliant Blue R250, 45 ml methanol, 10 ml acetic acid + distilled water to a total of 100 ml) [51] for 4 seconds and then on a microscopic slide (without a cover slip). The glands were subsequently photographed with a Sony CCD-Iris Hi resolution colour video camera under a Leica MZ6 binocular microscope. The Image Pro-Plus software was used to measure the diameters of the acini (lobes) later under the magnification of 2X. The short diameter (width) of the acinus was chosen to be measured, as this was apparent most of the time (Figure 1).
Ten assessors (testers/researchers) from ten different laboratories received the same microphotographs plus a calibrated image. The open-source ImageJ software (http://imagej.en.softonic.com/) chosen to be measured. At least 5 acini were randomly chosen from each photograph, and a minimum of 100 acini had to be measured per sample.
2.3. Statistical Analysis
Initially the inter-rater reliability (= the degree of agreement among independent observers who assess the same phenomenon) of the assessor’s measurements was assessed as reliability percentage by applying the following formula: 1- standard deviation/ mean of diameter of acini per group *100.
A full factorial General Linear Model Analysis was also performed on all data, using all variables as follows: the ‘treatment’ (Control versus imidacloprid), the ‘age’ (10-days old versus 15-days old) and the ‘condition’ (laboratory versus field) as fixed effect factors and the ‘assessor’ as a random effect factor. A Tukey-B post hoc test was applied to mean values per group for the factor ‘treatment’ to detect the significant subsets. Statistical analysis was performed using the R software version of 4.1.1 (R Core Team, 2021).
3. Results
3.1. Reliability of Assessors
The reliability of the assessors was very high, always higher than 95% in all groups, both in laboratory and field conditions regardless of age and treatment (Table 1). Furthermore, given that StDev values were always very small compared to the Mean values pre group we regarded the data as uniform among the assessors and we treated the data of all assessors in each group as one pooled data set.
Plotting all control data from both lab and field conditions and from both age groups against all data from the treated groups we showed that overall, the diameter of the HPG acini was always higher in control than in imidacloprid treated bees independently of the assessor (Figure 1).
3.2. Effect of Treatment in Different Age Groups and Test Conditions
GLM analysis results revealed that all factors had a significant overall effect: Imidacloprid decreased significantly the diameter of the acini (‘treatment’: P< 0.001; Table S1, Figure 1; Figure 2); 15-days old bees as an overall had reduced diameter of acini (‘age’: P< 0.005, Table S1; Figure 3) and bees in the laboratory had smaller HPGs than the bees in the field (‘condition’: P<0.001, Table S1; Figure 3). GLM analysis has confirmed the effect of the treatment and the rest of the variables was independent from the assessor (‘treatment’ X ‘assessor’: P<0.001, Table S1; Figure 1) similar results of what we show on the reliability of the assessors’ performance (Table 1, Figure 2). The overall differences among the ‘assessors’ were found to be significant, but they did not alter the effect of any of the other variables (Table S1).
The effect of imidacloprid treatment was different among the age groups, both in the laboratory and in the field, with the 10-days age bees having significantly smaller acini after treatment with imidacloprid in the lab and in the field, but the same was not true for the 15-days old bees (both in the lab and in the bees) (‘treatment’ X ‘age’: P<0.001, Table S1, Figure 3).
Although the overall effect of age showed that the 15-days old bees had smaller HPG acini, this was not the same if comparing bees reared in the laboratory and in the field (‘age’ X ‘condition’: P<0.001, Table S1, Figure 3). In particular, 15-days control bees in the laboratory had significantly smaller acini than the 10-days control bees but 15-days old bees reared in the field had the same diameter of the acini with the 10-days old bees reared in the field (Table S1, Figure 3). Furthermore, the reduction of the diameter of the acini on the 10-days old bees in the laboratory is similar to the reduction induced also by the treatment of imidacloprid to the same age group (Figure 3).
4. Discussion
Size of HPGs reflects the amount of proteins produced in the glands [41], and thus the amount and quality of the RJ produced for feeding the brood. Any stressor affecting the size of the glands is believed to affect the amount of the brood food available [38]. Furthermore, as already mentioned above, the use of neonicotinoid pesticides affects the size of the glands [33], induce histopathological changes [46], and reduces the quality of the brood food by reducing the proteolytic activity and the protein content especially of RJ major proteins [14]. Therefore, digital measurements of the diameter of the HPG acini could be used in risk assessment methodology to assess the effect of any neurotoxic pesticide even microbial bioinsecticides [52]. One of the main objectives of this study was to evaluate the reliability of digital measurements of the diameter of the HPG acini, performed by different assessors. We observed that all assessors concluded the same results (Table 1; Figure 2). The reliability percentages for all groups of age and test conditions were > 95%, a percentage that is highly accurate and an excellent proof of the consistency of the method, with little influence by assessor bias. Furthermore, although some differences were observed among the assessors, as expected given the human variable, these differences did not affect the overall result, in all groups and ages of bees.
The natural effect of age on the development of the acini has already been known for decades, as the structure of HPGs start to develop from the pupae around a week before the emergence of the young worker; they continue to change structure in their adult life, reaching their peak at the nurse stage 6-10 days post emergence [42]. When assessing this natural effect of age on non-treated workers, particularly 10- and 15-days post-emergence, we discovered that the expected peak reduction of the acini size was only present in workers kept under laboratory conditions and not on those from the field (Figure 3). Under laboratory conditions the 10-day old bees had significantly larger HPGs and overall size, as also shown in previous experiments [38]. Then, the acini naturally decrease as bees stop nursing and start foraging. The bees sampled from the field had no significant difference in diameter between the two groups of age. As expected, environmental conditions are of significant importance, and the size of the acini could vary [53]. We hypothesised that the respected colonies were still investing in brood-rearing instead of investing in foraging for nectar, as it was not the main honey flow period yet, therefore their HPGs were still well developed [53]. We also showed that HPG diameter was significantly larger on workers from the field for both 10-days and 15-days old compared to the those reared in the laboratory, which is in disagreement with the size of the acini diameter presented in De Smet et al., [34]. This may also support the hypothesis that the different environmental conditions and needs of the colonies influence the development of the acini, and therefore the age when workers undergo transition from one to another task (e.g., from nursing to foraging [40]. There is also evidence that co-exposure to stressors as well as environmental and endogenous factors modulate the honeybee response to pesticides and that honeybee workers are not all equal regarding the risk posed by exposure to pesticides [54].
The reason for overall bigger HPG acini in bees from the field and absence of decrease after the 10th day can possibly be explained by multiple reasons, such as the presence of brood in the colony and the large number of nurse bees during the summer season. Shown by Smodiš Škerl & Gregorc [55] the size of the acini can be affected positively and the HPGs secretion period can be extended in actively nursing workers. Therefore, the absence of brood in the laboratory, yet the presence of a large number of brood cells in the colonies when the experiment was conducted (July-August) can possibly explain the absence of the decreasing effect that was observed in the field. Moreover, the experimental workers from the field had access to a richer and more diverse source of pollen diet than bees maintained in the laboratory that were given a specific pollen paste until 10 days post-emergence. A multi- flower pollen is important for the development of the individual bees as it can stimulate and affect the total number of proteins, carbohydrates and lipids and furthermore have an impact on brood and colony development and immunity [56]. Studies show that specific genus of plants (Rubus, Castanea sp., Asparagus sp.) have a positive effect on the size of the acini of HPGs [44,52]. Furthermore, the phytochemicals that bees collect from the nectar and pollen offers them benefits of improved longevity and pathogen tolerance, and some compounds (p-coumaric acid or kaempferol) have been shown that can increase the size of the acini [57]. Exposure to fresh pollen in the field gives them the advantage of a rich pollen and nectar diet that stimulates the development of HPGs, while the stimulating effect of stored pollen used in the laboratory is reduced [56]. Overall, the absence of brood and the restricted source of proteins in the laboratory can further influence the size of the HPG acini.
The effect of imidacloprid was found significant only on the 10-days old bees, while not on the 15-days old bees both under laboratory and field conditions. The decreasing effect on the 10-days old bees is in agreement with previous findings [33,34]. Furthermore, De Smet et al., [34] showed that the decreasing effect of imidacloprid on 10-days old bees was observed in both laboratory and field conditions; results of this study confirm the findings. However, even if the imidacloprid did not show a decreasing effect on the 15-days old bees in this study, this result was again similar to the bees reared in the laboratory with the bees reared in the field. Furthermore, the reduction of HPGs on treated bees could not be associated with the food consumption. As stated above, the fact that the bees in the laboratory were given contaminated pollen only till day 10 of their life, and bees in the field were given small quantities of contaminated food, might imply that after the exposure to the stressors ceased (e,g, imidacloprid) the bees try to detoxify themselves. Some evidence of detoxification was found during the study by De Smet et al., [34]; however, this hypothesis needs to be further investigated and verified, as there is no knowledge on what the time frame is honeybees need to detoxify from different chemical stressors.
Our work demonstrated that results from the two different tests, laboratory and field, were in agreement. Both showed a negative effect of imidacloprid on 10-day old bees in the laboratory and in the field, but not on 15-day old bees. These results, together with the ones by De Smet et al., [34] support that Tier 1 laboratory studies can be used as a proxy for Tier 2 studies, at least in respect of the HPGs development, at day 10 of the bees’ life. The sublethal effects caused by the synergy of insecticides, fungicides and parasites occurring naturally in the field [10] could also be compensated by richer nutrition, or amplified by poor nutrition. Considering also that the development of the glands in the honey bee worker, as a social insect, is very complex [40], the assessment of the acini at day 10 of their life is a reasonable recommendation, if the HPG measurement is going to be part of the risk assessment methodology. It is also of great importance to note that all other stressors acting synergistically in the honey bee health, immunity, productivity (e.g., pathogens and nutrition) might have higher impact later on the bees’ life (Straub et al., 2019), as well as on the reproductive individuals [36] given that the exposure in the field is continuous and not only for the first days of the bees’ adult life. However, the digital measurements of HPGs of the honey bees at day 10 of their life can be proposed as a very robust method to be considered in risk assessment approaches for revealing sub-lethal effects of stressors.
5. Conclusions
Hypopharyngeal glands of nursing worker honey bees, which are responsible for producing royal jelly, are considered as a robust indicator of developmental/ physiological failure due to pesticide exposure [10,33,34,46. Neurotoxic pesticides such as imidacloprid reduce the size of the lobes of the HPGs, which are called acini.
Digital measurements of the diameter of the acini were shown to be a reliable methodological tool for risk assessment, as all results were independent from the assessor as the human factor.
Similar trends in acini reduction were found both in the laboratory and field tests, when the same concentration of the toxic substance was administered, but exposure in the field was probably lower. Thus, measurement of the HPG size in the laboratory can be used as a proxy for field studies.
As acini size 15 days post-emergence are subject to environmental conditions (including pollen availability, brood presence, foraging activity, diverse food sources) and only to age, the recommendation for using the HPG acini diameter as a methodological tool is restricted to 10 day old individuals.
Environmental Risk Assessment (ERA) schemes could benefit from the results of the presented work, providing insight into scenarios more likely encountered by worker honey bees in the real world.
Supplementary Materials
The following supporting information can be downloaded at: Preprints.org, Table S1 Table S1. GLM analysis results on the data originated from the digital measurements on the honey bee HPG acini diameter, using the ‘treatment’ (control versus imidacloprid), ‘age’ (10-days old versus 15-days old) and ‘condition’ (laboratory versus field) as fixed effect factors; “assessor” was also used as a random effect factor.
Author Contributions
conceptualization: F.H. and N.S.D.; data curation: A.D.M., F.H. and M.T.; formal analysis: A.D.M., F.H. and M.T.; funding acquisition: F.H. and N.S.D.; investigation: F.H., M.T., L.B., A.B., S.B., M.C., J.H.L., S.K., D.L., A.M., M.P., M.T.R., F.S., N.S.D., S.T. and G.R.W.; methodology: F.H. and A.B.; writing – original draft: F.H., M.T., N.S.D. and G.R.W. All authors have read and agreed to the published version of the manuscript.”
Funding
This work was supported by: the EC-Greek Government project (EC Dir. 797/07 “Effect of neonicotinoids and air pollution on honey bees”); the Alabama Agricultural Experiment Station, USDA NIFA Multi-state Project NC1173; COLOSS Association support to APITOX Task Force. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Data Availability Statement
The data are available through the ZENODO public repository. https://zenodo.org/badge/DOI/10.5281/zenodo.15644265.svg (accessed on 11 June 2025).
Acknowledgments
We would like to thank all technicians and researchers in our institutions who helped to maintain the honey bee colonies. The authors have reviewed and edited the output and take full responsibility for the content of this publication.”
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| HPG | Hypopharyngeal Glands |
| GLM | General Linear Model |
References
- Aguilar, R.; Ashworth, L.; Galetto, L.; Aizen, M. A. Plant Reproductive Susceptibility to Habitat Fragmentation: Review and Synthesis through a Meta-analysis. Ecol. Lett. 2006, 9, 968–980. [Google Scholar] [CrossRef] [PubMed]
- Bommarco, R.; Marini, L.; Vaissière, B. E. Insect Pollination Enhances Seed Yield, Quality, and Market Value in Oilseed Rape. Oecologia 2012, 169, 1025–1032. [Google Scholar] [CrossRef] [PubMed]
- Potts, S. G.; Biesmeijer, J. C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W. E. Global Pollinator Declines: Trends, Impacts and Drivers. Trends Ecol. Evol. 2010, 25, 345–353. [Google Scholar] [CrossRef]
- Neumann, P.; Carreck, N. L. Honey Bee Colony Losses. J. Apicult. Res. 2010, 49, 1–6. [Google Scholar] [CrossRef]
- Odoux, J.-F.; Aupinel, P.; Gateff, S.; Requier, F.; Henry, M.; Bretagnolle, V. ECOBEE: A Tool for Long-Term Honey Bee Colony Monitoring at the Landscape Scale in West European Intensive Agroecosystems. J. Apicult. Res. 2014, 53, 57–66. [Google Scholar] [CrossRef]
- Jacques, A.; Laurent, M.; EPILOBEE Consortium; Ribière-Chabert, M. ; Saussac, M.; Bougeard, S.; Budge, G. E.; Hendrikx, P.; Chauzat, M.-P. A Pan-European Epidemiological Study Reveals Honey Bee Colony Survival Depends on Beekeeper Education and Disease Control. PLoS ONE 2017, 12, e0172591. [Google Scholar] [CrossRef] [PubMed]
- Goulson, D.; Nicholls, E.; Botías, C.; Rotheray, E. L. Bee Declines Driven by Combined Stress from Parasites, Pesticides, and Lack of Flowers. Science 2015, 347, 1255957. [Google Scholar] [CrossRef]
- Chmiel, J. A.; Daisley, B. A.; Pitek, A. P.; Thompson, G. J.; Reid, G. Understanding the Effects of Sublethal Pesticide Exposure on Honey Bees: A Role for Probiotics as Mediators of Environmental Stress. Front. Ecol. Evol. 2020, 8, 22. [Google Scholar] [CrossRef]
- del Águila Conde, M.; Febbraio, F. Risk Assessment of Honey Bee Stressors Based on in Silico Analysis of Molecular Interactions. EFS2 2022, 20, (EU–FORA. [Google Scholar] [CrossRef]
- Bruckner, S.; Straub, L.; Neumann, P.; Williams, G. R. Negative but Antagonistic Effects of Neonicotinoid Insecticides and Ectoparasitic Mites Varroa Destructor on Apis Mellifera Honey Bee Food Glands. Chemosphere 2023, 313, 137535. [Google Scholar] [CrossRef]
- Blacquière, T.; Smagghe, G.; Van Gestel, C. A. M.; Mommaerts, V. Neonicotinoids in Bees: A Review on Concentrations, Side-Effects and Risk Assessment. Ecotoxicology 2012, 21, 973–992. [Google Scholar] [CrossRef] [PubMed]
- Simon-Delso, N.; Amaral-Rogers, V.; Belzunces, L. P.; Bonmatin, J. M.; Chagnon, M.; Downs, C.; Furlan, L.; Gibbons, D. W.; Giorio, C.; Girolami, V.; Goulson, D.; Kreutzweiser, D. P.; Krupke, C. H.; Liess, M.; Long, E.; McField, M.; Mineau, P.; Mitchell, E. A. D.; Morrissey, C. A.; Noome, D. A.; Pisa, L.; Settele, J.; Stark, J. D.; Tapparo, A.; Van Dyck, H.; Van Praagh, J.; Van Der Sluijs, J. P.; Whitehorn, P. R.; Wiemers, M. Systemic Insecticides (Neonicotinoids and Fipronil): Trends, Uses, Mode of Action and Metabolites. Environ. Sci. Pollut. Res. 2015, 22, 5–34. [Google Scholar] [CrossRef] [PubMed]
- Bonmatin, J.-M.; Giorio, C.; Girolami, V.; Goulson, D.; Kreutzweiser, D. P.; Krupke, C.; Liess, M.; Long, E.; Marzaro, M.; Mitchell, E. A. D.; Noome, D. A.; Simon-Delso, N.; Tapparo, A. Environmental Fate and Exposure; Neonicotinoids and Fipronil. Environ. Sci. Pollut. Res. 2015, 22, 35–67. [Google Scholar] [CrossRef] [PubMed]
- Wilde, J.; Frączek, R. J.; Siuda, M.; Bąk, B.; Hatjina, F.; Miszczak, A. The Influence of Sublethal Doses of Imidacloprid on Protein Content and Proteolytic Activity in Honey Bees ( Apis Mellifera L.). J. Apicult. Res. 2016, 55, 212–220. [Google Scholar] [CrossRef]
- Zaluski, R.; Justulin, L. A.; Orsi, R. D. O. Field-Relevant Doses of the Systemic Insecticide Fipronil and Fungicide Pyraclostrobin Impair Mandibular and Hypopharyngeal Glands in Nurse Honeybees (Apis Mellifera). Sci. Rep. 2017, 7, 15217. [Google Scholar] [CrossRef]
- Al Naggar, Y.; Baer, B. Consequences of a Short Time Exposure to a Sublethal Dose of Flupyradifurone (Sivanto) Pesticide Early in Life on Survival and Immunity in the Honeybee (Apis Mellifera). Sci. Rep. 2019, 9, 19753. [Google Scholar] [CrossRef]
- Paleolog, J.; Wilde, J.; Gancarz, M.; Wiącek, D.; Nawrocka, A.; Strachecka, A. Imidacloprid Pesticide Causes Unexpectedly Severe Bioelement Deficiencies and Imbalance in Honey Bees Even at Sublethal Doses. Animals 2023, 13, 615. [Google Scholar] [CrossRef]
- Tosi, S.; Costa, C.; Vesco, U.; Quaglia, G.; Guido, G. A 3-Year Survey of Italian Honey Bee-Collected Pollen Reveals Widespread Contamination by Agricultural Pesticides. Sci. Total Envir. 2018, 615, 208–218. [Google Scholar] [CrossRef]
- Girolami, V.; Marzaro, M.; Vivan, L.; Mazzon, L.; Greatti, M.; Giorio, C.; Marton, D.; Tapparo, A. Fatal Powdering of Bees in Flight with Particulates of Neonicotinoids Seed Coating and Humidity Implication. J. Appl. Entom. 2012, 136, (1–2). [Google Scholar] [CrossRef]
- Greatti, M.; Sabatini, A. G.; Barbattini, R.; Rossi, S.; Stravisi, A. Risk of Environmental Contamination by the Active Ingredient Imidacloprid Used for Corn Seed Dressing. Preliminary Results. Bull. Insectology 2003, 56, 69–72. [Google Scholar]
- Marzaro, M.; Vivan, L.; Targa, A.; Mazzon, L.; Mori, N.; Greatti, M.; Toffolo, E. P.; Bernardo, A. D.; Giorio, C.; Marton, D.; Girolami, V. Lethal Aerial Powdering of Honey Bees with Neonicotinoids from Fragments of Maize Seed Coat. Bull. Insectology 2001, 64, 119–126. [Google Scholar]
- Tapparo, A.; Marton, D.; Giorio, C.; Zanella, A.; Soldà, L.; Marzaro, M.; Vivan, L.; Girolami, V. Assessment of the Environmental Exposure of Honeybees to Particulate Matter Containing Neonicotinoid Insecticides Coming from Corn Coated Seeds. Environ. Sci. Technol. 2012, 46, 2592–2599. [Google Scholar] [CrossRef]
- Schmuck, R.; Schöning, R.; Stork, A.; Schramel, O. Risk Posed to Honeybees ( Apis Mellifera L, Hymenoptera) by an Imidacloprid Seed Dressing of Sunflowers. Pest Management Science 2001, 57, 225–238. [Google Scholar] [CrossRef] [PubMed]
- Bonmatin, J. M.; Marchand, P. A.; Charvet, R.; Moineau, I.; Bengsch, E. R.; Colin, M. E. Quantification of Imidacloprid Uptake in Maize Crops. J. Agric. Food Chem. 2005, 53, 5336–5341. [Google Scholar] [CrossRef]
- Chauzat, M.-P.; Martel, A.-C.; Cougoule, N.; Porta, P.; Lachaize, J.; Zeggane, S.; Aubert, M.; Carpentier, P.; Faucon, J.-P. An Assessment of Honeybee Colony Matrices, Apis Mellifera (Hymenoptera: Apidae) to Monitor Pesticide Presence in Continental France. Envir. Toxicol. Chem. 2011, 30, 103–111. [Google Scholar] [CrossRef]
- Girolami, V.; Mazzon, L.; Squartini, A.; Mori, N.; Marzaro, M.; Di Bernardo, A.; Greatti, M.; Giorio, C.; Tapparo, A. Translocation of Neonicotinoid Insecticides From Coated Seeds to Seedling Guttation Drops: A Novel Way of Intoxication for Bees. J. Econ. Entomol. 2009, 102, 1808–1815. [Google Scholar] [CrossRef]
- Tosi, S.; Sfeir, C.; Carnesecchi, E.; vanEngelsdorp, D.; Chauzat, M.-P. Lethal, Sublethal, and Combined Effects of Pesticides on Bees: A Meta-Analysis and New Risk Assessment Tools. Sci. Total Environ. 2022, 844, 156857. [Google Scholar] [CrossRef] [PubMed]
- Kirchner, W. H. 17. Mad-Bee-Disease? Subletale Effekte von Imidacloprid (Gaucho®) Auf Das Verhalten von Honigbienen. Apidologie, 0089. [Google Scholar]
- Tosi, S.; Burgio, G.; Nieh, J. C. A Common Neonicotinoid Pesticide, Thiamethoxam, Impairs Honey Bee Flight Ability. Sci. Rep. 2017, 7, 1201. [Google Scholar] [CrossRef]
- Colin, M. E.; Bonmatin, J. M.; Moineau, I.; Gaimon, C.; Brun, S.; Vermandere, J. P. A Method to Quantify and Analyze the Foraging Activity of Honey Bees: Relevance to the Sublethal Effects Induced by Systemic Insecticides. Arch. Environ. Contam. Toxicol. 2004, 47, 387–395. [Google Scholar] [CrossRef]
- Ramirez-Romero, R.; Chaufaux, J.; Pham-Delègue, M.-H. Effects of Cry1Ab Protoxin, Deltamethrin and Imidacloprid on the Foraging Activity and the Learning Performancesof the Honeybee Apis Mellifera, a Comparative Approach. Apidologie 2005, 36, 601–611. [Google Scholar] [CrossRef]
- Henry, M.; Béguin, M.; Requier, F.; Rollin, O.; Odoux, J.-F.; Aupinel, P.; Aptel, J.; Tchamitchian, S.; Decourtye, A. A Common Pesticide Decreases Foraging Success and Survival in Honey Bees. Science 2012, 336, 348–350. [Google Scholar] [CrossRef] [PubMed]
- Hatjina, F.; Papaefthimiou, C.; Charistos, L.; Dogaroglu, T.; Bouga, M.; Emmanouil, C.; Arnold, G. Sublethal Doses of Imidacloprid Decreased Size of Hypopharyngeal Glands and Respiratory Rhythm of Honeybees in Vivo. Apidologie 2013, 44, 467–480. [Google Scholar] [CrossRef]
- De Smet, L.; Hatjina, F.; Ioannidis, P.; Hamamtzoglou, A.; Schoonvaere, K.; Francis, F.; Meeus, I.; Smagghe, G.; De Graaf, D. C. Stress Indicator Gene Expression Profiles, Colony Dynamics and Tissue Development of Honey Bees Exposed to Sub-Lethal Doses of Imidacloprid in Laboratory and Field Experiments. PLoS ONE 2017, 12, e0171529. [Google Scholar] [CrossRef]
- Williams, G. R.; Troxler, A.; Retschnig, G.; Roth, K.; Yañez, O.; Shutler, D.; Neumann, P.; Gauthier, L. Neonicotinoid Pesticides Severely Affect Honey Bee Queens. Sci. Rep. 2015, 5, 14621. [Google Scholar] [CrossRef] [PubMed]
- Straub, L.; Villamar-Bouza, L.; Bruckner, S.; Chantawannakul, P.; Gauthier, L.; Khongphinitbunjong, K.; Retschnig, G.; Troxler, A.; Vidondo, B.; Neumann, P.; Williams, G. R. Neonicotinoid Insecticides Can Serve as Inadvertent Insect Contraceptives. Proc. R. Soc. B. 2016, 283, 20160506. [Google Scholar] [CrossRef]
- Sandrock, C.; Tanadini, M.; Tanadini, L. G.; Fauser-Misslin, A.; Potts, S. G.; Neumann, P. Impact of Chronic Neonicotinoid Exposure on Honeybee Colony Performance and Queen Supersedure. PLoS ONE 2014, 9, e103592. [Google Scholar] [CrossRef]
- Crailsheim, K.; Schneider, L. H. W.; Hrassnigg, N.; Bühlmann, G.; Brosch, U.; Gmeinbauer, R.; Schöffmann, B. Pollen Consumption and Utilization in Worker Honeybees (Apis Mellifera Carnica): Dependence on Individual Age and Function. J. Insect Physiol. 1992, 38, 409–419. [Google Scholar] [CrossRef]
- Milone, J. P.; Chakrabarti, P.; Sagili, R. R.; Tarpy, D. R. Colony-Level Pesticide Exposure Affects Honey Bee (Apis Mellifera L.) Royal Jelly Production and Nutritional Composition. Chemosphere 2021, 263, 128183. [Google Scholar] [CrossRef]
- Crailsheim, K.; Stolberg, E. Influence of Diet, Age and Colony Condition upon Intestinal Proteolytic Activity and Size of the Hypopharyngeal Glands in the Honeybee (Apis Mellifera L.). J. Insect Physiol. 1989, 35, 595–602. [Google Scholar] [CrossRef]
- Knecht, D,; Kaatz, H. H. Patterns of larval food production by hypopharyngeal glands in adult worker honey bees. Apidologie 1990, 21, 457–458. [Google Scholar] [CrossRef]
- Deseyn, J.; Billen, J. Age-Dependent Morphology and Ultrastructure of the Hypopharyngeal Gland of Apis Mellifera Workers (Hymenoptera, Apidae). Apidologie 2005, 36, 49–57. [Google Scholar] [CrossRef]
- DeGrandi-Hoffman, G.; Chen, Y.; Huang, E.; Huang, M. H. The Effect of Diet on Protein Concentration, Hypopharyngeal Gland Development and Virus Load in Worker Honey Bees (Apis Mellifera L.). J. Insect Physiol. 2010, 56, 1184–1191. [Google Scholar] [CrossRef]
- Di Pasquale, G.; Salignon, M.; Le Conte, Y.; Belzunces, L. P.; Decourtye, A.; Kretzschmar, A.; Suchail, S.; Brunet, J.-L.; Alaux, C. Influence of Pollen Nutrition on Honey Bee Health: Do Pollen Quality and Diversity Matter? PLoS ONE 2013, 8, e72016. [Google Scholar] [CrossRef] [PubMed]
- Herbert, E. W. Jr.; Shimanuki, H.; Caron, D. Optimum protein levels required by honey bees (Hymenoptera, Apidae) to initiate and maintain brood rearing. Apidologie 1977, 8, 141–146. [Google Scholar] [CrossRef]
- Ibrahim, E.-D. S.; Abd Alla, A. E.; El-Masarawy, M. S.; Salem, R. A.; Hassan, N. N.; Moustafa, M. A. M. Sulfoxaflor Influences the Biochemical and Histological Changes on Honeybees (Apis Mellifera L.). Ecotoxicology 2023, 32, 674–681. [Google Scholar] [CrossRef]
- Renzi, M. T.; Rodríguez-Gasol, N.; Medrzycki, P.; Porrini, C.; Martini, A.; Burgio, G.; Maini, S.; Sgolastra, F. Combined Effect of Pollen Quality and Thiamethoxam on Hypopharyngeal Gland Development and Protein Content in Apis Mellifera. Apidologie 2016, 47, 779–788. [Google Scholar] [CrossRef]
- Ahmad, S.; Khan, K. A.; Ghramh, H. A.; Li, J. Phosphoproteomics Analysis of Hypopharyngeal Glands of the Newly Emerged Honey Bees (Apis Mellifera Ligustica). J. King Saud Univ. - Science 2022, 34, 102206. [Google Scholar] [CrossRef]
- European Food Safety Authority. Guidance on the Risk Assessment of Plant Protection Products on Bees (Apis Mellifera, Bombus Spp. and Solitary Bees). EFS2 2013, 11. [Google Scholar] [CrossRef]
- Berger, B.; Carmargo Abdalla, F. Programmed germ cell differentiation during ovary stages of oogenesis in caged virgin and fecundated queens of Apis mellifera Linné, 1758. Braz. J Morphol. Sci. 2005, 22, 1–4. [Google Scholar]
- Rhodes, J.W.; Somerrville, D. C. Introduction and early performance of queen bees. 2003. [Google Scholar]
- Omar, E. M.; Abdu_Allah, G.; Tawfik, A. Impacts of Sub-Lethal Concentrations of Two Macrocyclic Lactone Insecticides on Nurse Bees (Apis Mellifera L.) Hypopharyngeal Glands Development. PREPRINT (Version 1) , 2022. 17 October. [CrossRef]
- Hrassnigg, N.; Crailsheim, K. Adaptation of Hypopharyngeal Gland Development to the Brood Status of Honeybee (Apis Mellifera L.) Colonies. J. Insect Physiol. 1998, 44, 929–939. [Google Scholar] [CrossRef]
- Poquet, Y.; Vidau, C.; Alaux, C. Modulation of Pesticide Response in Honeybees. Apidologie 2016, 47, 412–426. [Google Scholar] [CrossRef]
- Škerl, M. I. S.; Gregorc, A. Characteristics of hypopharyngeal glands in honeybees (Apis mellifera carnica) from a nurse colony. Slov. Vet. Res. 2015, 52, 67–74. [Google Scholar]
- Bryś, M. S.; Skowronek, P.; Strachecka, A. Pollen Diet—Properties and Impact on a Bee Colony. Insects 2021, 12, 798. [Google Scholar] [CrossRef]
- Niño, E. L.; Yokota, S.; Stacy, W. H. O.; Arathi, H. S. Dietary Phytochemicals Alter Hypopharyngeal Gland Size in Honey Bee (Apis Mellifera L.) Workers. Heliyon 2022, 8, e10452. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Microphotographs of HPGs acini used for the digital measurements; a. control; b. treated with imidacloprid.
Figure 1.
Microphotographs of HPGs acini used for the digital measurements; a. control; b. treated with imidacloprid.
Figure 2.
Boxplots for the honey bee HPG diameter of acini for all data (lab & field), grouped by assessor and treatment (Control, Imidacloprid).
Figure 2.
Boxplots for the honey bee HPG diameter of acini for all data (lab & field), grouped by assessor and treatment (Control, Imidacloprid).
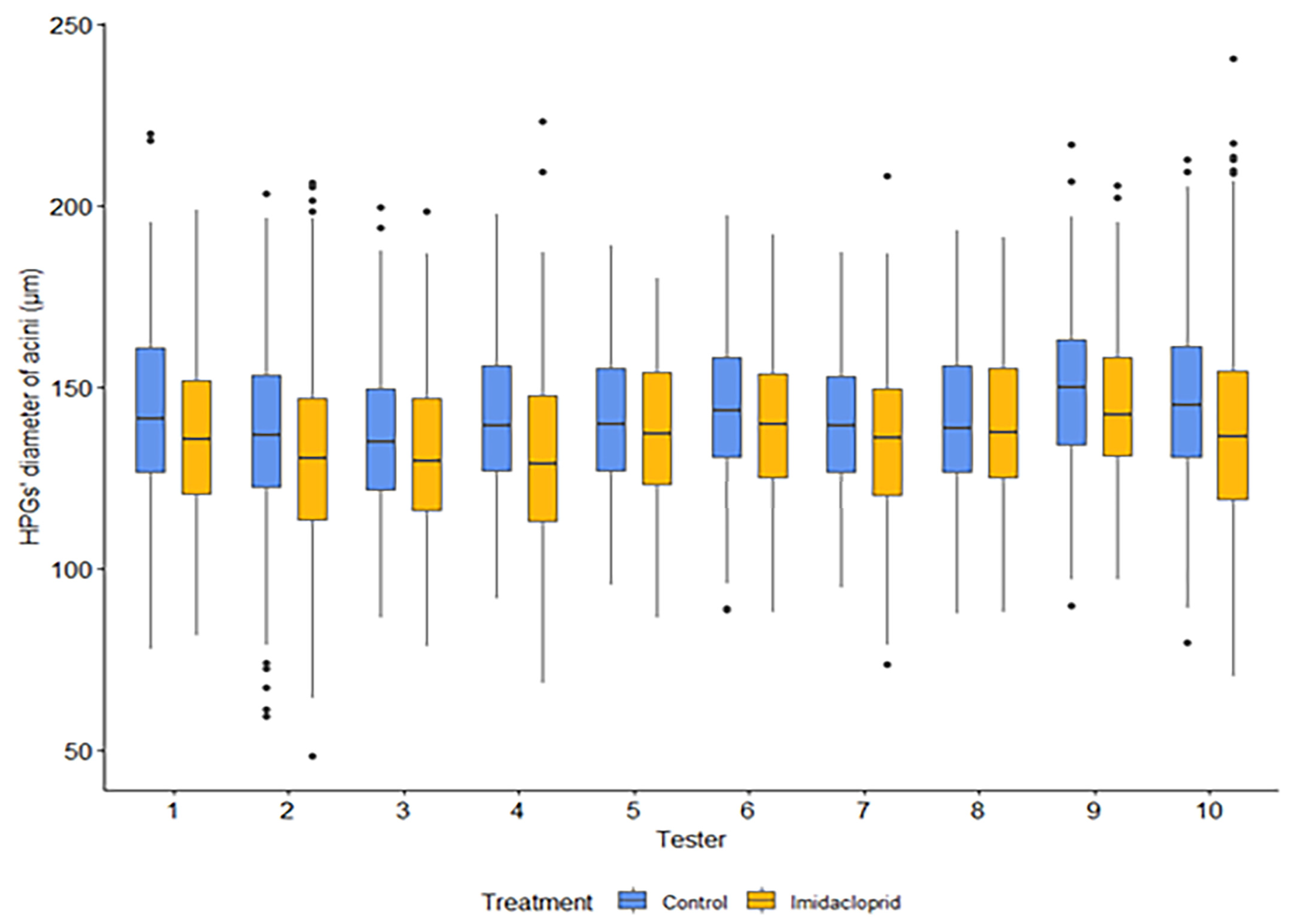
Figure 3.
Bars represented the diameter mean values (and SE) of the HPG acini of all treated groups, of both 10 and 25 days old bees in laboratory and field conditions (C= control, Im =imidacloprid). Groups with different letters are significantly different for P<0.001.
Figure 3.
Bars represented the diameter mean values (and SE) of the HPG acini of all treated groups, of both 10 and 25 days old bees in laboratory and field conditions (C= control, Im =imidacloprid). Groups with different letters are significantly different for P<0.001.
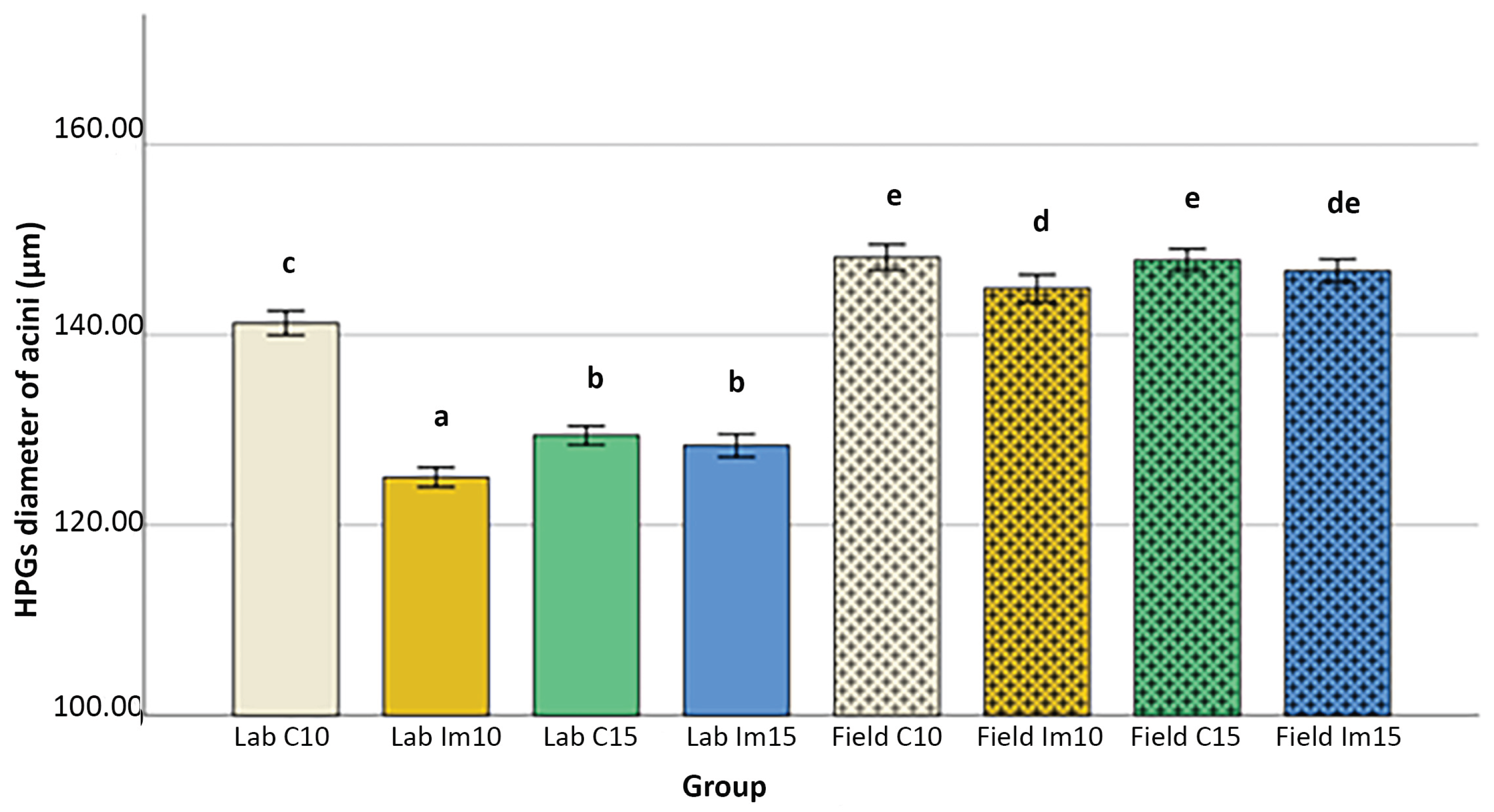

Table 1.
Mean values of the diameter of the honey bee HPG acini (μm) for each assessor and mean value, standard deviation and reliability percentage for each bee sampling group (C=Control, I= Imidacloprid).
Table 1.
Mean values of the diameter of the honey bee HPG acini (μm) for each assessor and mean value, standard deviation and reliability percentage for each bee sampling group (C=Control, I= Imidacloprid).
| Laboratory | Field | |||||||
|---|---|---|---|---|---|---|---|---|
| Assessor | C-10 | I-10 | C-15 | I-15 | C-10 | I-10 | C-15 | I-15 |
| T1 | 138.94 | 123.66 | 130.33 | 124.15 | 148.46 | 147.34 | 151.02 | 146.82 |
| T2 | 138.46 | 121.53 | 125.80 | 126.31 | 142.42 | 135.38 | 144.15 | 142.76 |
| T3 | 135.27 | 120.07 | 125.99 | 125.41 | 142.28 | 139.38 | 142.28 | 140.31 |
| T4 | 143.38 | 119.14 | 123.79 | 122.32 | 145.73 | 141.26 | 148.65 | 143.42 |
| T5 | 140.67 | 124.02 | 128.94 | 128.28 | 146.96 | 145.12 | 145.59 | 150.76 |
| T6 | 143.23 | 129.66 | 129.97 | 131.57 | 151.20 | 149.06 | 153.36 | 149.80 |
| T7 | 138.88 | 124.34 | 130.17 | 129.95 | 145.52 | 143.97 | 147.40 | 146.89 |
| T8 | 140.32 | 126.11 | 132.14 | 133.00 | 148.94 | 146.51 | 141.88 | 150.17 |
| T9 | 146.96 | 134.87 | 137.37 | 132.87 | 158.16 | 156.11 | 157.06 | 158.21 |
| T10 | 145.76 | 127.36 | 131.49 | 129.29 | 154.72 | 149.47 | 150.76 | 144.83 |
| Mean Val. | 141.19 | 125.08 | 129.60 | 128.32 | 148.44 | 145.36 | 148.22 | 147.40 |
| St.Dev. | 3.61 | 4.71 | 3.85 | 3.69 | 5.10 | 5.83 | 4.93 | 5.12 |
| Reliability | 97.44% | 96.23% | 97.03% | 97.12% | 96.56% | 95.99% | 96.67% | 96.52% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



