
Submitted:
25 June 2025
Posted:
26 June 2025
You are already at the latest version
Abstract
To accelerate maturation of intestinal function and promote growth and development, the effect of maternal clofibrate on intestinal fatty acid metabolism was investigated in postnatal piglets. 27 pregnant sows were fed either 0, 0.25, or 0.5% clofibrate during late gestation and early lactation. [1-14C]-Oleic acid metabolism was measured in vitro in intestinal mucosa of piglets with/without L-carnitine and/or malonate. Clofibrate increased oleic acid metabolism on d1, and the increase was higher from 0.5 than 0.25% of maternal clofibrate (p < 0.005). Flux to CO2 increased with age while flux to acid soluble products (ASP) remained constant after d1. Flux to esterified products (ESP) increased on d7, but the increase was dampened by clofibrate (p < 0.0001). Carnitine increased flux to CO2 and malonate decreased it (p < 0.0001), but neither affected ASP or ESP. Intestinal NEFA and TG levels decreased linearly, and CPT activity increased quadratically with age. Clofibrate increased ACOX abundance but decreased the ratio of CPT1A and CPT1B on d1. Postnatal age increased FABP2 but decreased PPARα and RXRα. In conclusion, maternal supplementation of clofibrate promotes intestinal energy generation from fat oxidation in postnatal piglets, but the promotion is influenced by age, in which ACOX, FABP2 and CPT1 might play modulatory roles.
Keywords:
clofibrate
; fatty acid
; intestinal mucosa
; maternal
; suckling piglets
1. Introduction
The permeability of placenta to fatty acids (FAs) is highly restricted, consequentially; the primary energy substrate for developing fetus is glucose. Ketone bodies may be used as fuel and lipogenic substrates by the fetus when the mother is under fasting conditions using FA for ketogenesis [1]. However, milk lipids become the principal substrate for oxidative metabolism after birth. More than 55% of the dietary energy for neonates is milk fat [2] depending on species, suggesting that highly efficient oxidation of FA from milk fat is very important for the newborns’ development and growth. Unlike human infants, newborn piglets and rats [3] have very limited fat stores. Energy is derived primarily from absorbed FA from the milk lipids, implying that intestine plays a critical role for neonates to acquire energy efficiently after birth.
Lipids and other nutrients from milk are taken primarily from the small intestinal lumen into the lymph system/bloodstream via the intestinal epithelial cells. This process undergoes significant development throughout life with key changes happening during fetal development and neonatal growth. The changes during development affect the efficiency of lipid uptake depending on species and energy needs at different stages. Fetal and neonatal pigs, like immature infants, are enriched in fetal-type cells in the developing intestine [4]. The fetal-type cells need to mature into adult enterocytes with fully functional absorptive cells at birth and during the suckling period. This transformation is crucial for their survival, development and growth after birth, as they have limited energy storage and developmental delays. To have adult absorptive and secretory functions, therefore, the energy requirement of intestine is very high [5] including the needs for synthetic or digestive enzymes and the fast renewal of enterocytes.
The role of peroxisome proliferator-activated receptor alpha (PPARα) in FA oxidation in the liver has been well established, as the liver is the primary location of FA metabolic and ketogenic pathways to generate energy for use by other tissues. Compared to liver, the intestine utilizes lipids for energy itself [6]. This characteristic has driven us to evaluate the regulatory function of PPARα in the intestine. So far, most studies examining the effects of PPARα activation on intestinal lipid metabolism were performed in rodents [7,8,9,10]. Specifically, the expression of genes associated with PPARα activation by clofibrate, its pharmaceutical agonist, was determined in the developing small intestine in rat [11]. The results revealed that the PPARα target genes such as acyl-CoA oxidase 1 (ACOX 1) and FA-binding proteins (FABPs) might be coordinately regulated during postnatal development. Moreover, increased FA oxidation was observed also in enterocytes treated with the PPARα agonist bezafibrate [8]. As non-peroxisome-proliferating species [12], however, the kinetic responses of intestinal FA oxidative metabolism of pigs to PPARα activation have not been explored during neonatal-suckling period. Optimizing the metabolism of FA in the small intestine is beneficial for the survival and development of newborn pigs, because high neonatal piglet mortality has remained a major economic and animal welfare problem in the swine industry worldwide for decades.
The high mortality observed in neonatal swine is associated with the low energy intake and utilization after birth [13]. Increasing milk lipid utilization can improve growth and development, subsequently reducing the high mortality. Results from our previous studies showed that supplementation of clofibrate to neonatal piglets promoted hepatic and extrahepatic PPARα-dependent gene expression and increased FA oxidation [14,15]. Specifically, we found that maternal clofibrate could be transferred to the fetus across the placenta membrane and increase FA oxidation at birth. However, the stimulated FA oxidation is diminished with time [16], suggesting that sustained PPARα activation is required to maintain the induced high FA oxidation rate after birth. In addition, lactation transfer of clofibrate was observed in suckling rats with increased hepatic and renal cytochrome P450 4A mRNA levels [17,18]. With this knowledge, we hypothesized that supplementation of clofibrate to sows during late gestation and early lactation may stimulate the utilization of milk FA in the rapidly developing intestine of their offspring. To test our hypothesis, the capacity of energy generation from fatty acids was evaluated over time in intestine of suckling piglets from sows treated with or without clofibrate during late gestation and early lactation.
2. Results
2.1. Fatty Acid Oxidation
2.1.1. Effects of Maternal Supplementation of Clofibrate on Fatty Acid Metabolism in the Intestinal Mucosa of Piglets During the Neonatal-Suckling Period
- Significant interactions between maternal supplementation of clofibrate and postnatal age were observed for the 14C accumulation in CO2 (p < 0.005), ASP (p < 0.05), and ESP (p < 0.0001) as well as the total oxidation (p < 0.05) and total metabolism (p < 0.0001) from oleic acid (Table 1).
- The 14CO2 accumulation was stimulated linearly with the dose of maternal clofibrate on d1 (p < 0.0005), but the stimulation was diminished after d7 depending on the dose. The accumulation rate measured on d1 was 1.1 and 1.6-fold greater in piglets from sows with 0.25 and 0.5% clofibrate than controls. The difference was not detectable on d7, while the accumulation was 60% higher on d19 from 0.25% and 44% and 72% higher on d 14 and 19 from 0.5% clofibrate treated sows than from the controls (p < 0.05).
- The 14C accumulation in ASP on d1 was 1.3 and 1.7-fold higher in pigs from sows with 0.25 and 0.5% clofibrate than from the controls (p < 0.05), but the maternal supplementation had no effects on the 14C accumulation in ASP in pigs after d1. The 14C accumulation in ASP was higher from d1 than all other ages (p < 0.0001), and no differences were observed between all other ages.
- The 14C accumulation in ESP in piglets from control sows increased from d1 to 7 but decreased greatly after d7 (p < 0.005). The accumulation was on average 61% and 81% lower from d 14 and 19 than d1 and d7. No difference was detected between d 14 and 19. Maternal clofibrate had no impact on the accumulation in ESP on d 1, 14 and 19, but decreased the accumulation in ESP on d 7 (p < 0.0001). The decrease was greater from maternal clofibrate level 0.5% than 0.25%.Third bullet.
- The 14C accumulation in total oxidation (CO2 + ASP) was 37% and 79% higher in piglets on d1 from sows with 0.25 % and 0.5 % clofibrate than from the controls (p < 0.05), but supplementation had no effects on the 14C accumulation after d1. The 14C accumulation in CO2 + ASP was higher from d1 than that measured from other ages (p < 0.0001), but no differences were observed between all other ages.
- The 14C accumulation in the total metabolism (CO2+ASP+ESP) in piglets from control sows increased from d1 to 7 (p < 0.005) but had no difference on d 14 and 19. The accumulation was 3.1 and 4.6-fold higher on average from d1 and 7 than d14 and 19 (p < 0.0001). Maternal supplementation of 0.5% clofibrate increased the total metabolism on day 1 (p < 0.005) but decreased the accumulation in ESP on d 7. The decrease was greater from maternal clofibrate level 0.5% than 0.25%.
2.1.1. Effects of Maternal Supplementation of Clofibrate on Fatty Acid Metabolism in the In-Testinal Mucosa of Piglets During the Neonatal-Suckling Period
- The % CO2 in total FA oxidation increased, and % of ASP in Total FA oxidation decreased linearly with maternal clofibrate dose (p < 0.05) and postnatal age (p < 0.0001). No interactions (p > 0.1) were detected between maternal clofibrate and postnatal age (Table 2).
- The % of CO2, ASP and ESP in total FA metabolites had significant (p <0.005) interactions (Table 2) between the maternal clofibrate and age. The % of CO2 in total metabolites increased with age from d1 to d14 (p < 0.0001), but the increase was greater in pigs from sows fed 0.25% clofibrate on d7. The % CO2 in pigs on d14 and 19 from sows fed 0.5% clofibrate were higher than that from control sows (p < 0.05). The % of CO2 in pigs from sows fed 0.25% clofibrate was lower at d 19 compared to d14 (p< 0.005), but the % measured in pigs from sows fed 0 or 0.5% clofibrate showed no difference. The % of ASP in total metabolites measured in piglets from control sows was decreased from d1 to d7 and then increased from d7 to d19 (p < 0.0001). Maternal clofibrate increased the % of ASP, but the increase varied with age and clofibrate dose. The % was higher in piglets on d1 and 14 from sows fed 0.25% clofibrate than control sows, and on d7 and d14 from sows fed 0.5% clofibrate than the controls (p < 0.05). The % of ESP in total metabolites in pigs from control sows increased from d1 to d7 but decreased after d7 (p < 0.005). As opposed to the % of ASP, maternal supplementation of clofibrate decreased the % of ESP and the decrease varied with the age and dose of maternal clofibrate supplementation. The decrease in % of ESP from 0.5% of clofibrate was similar as 0.25% of clofibrate on d1 and d7 but was greater than 0.25% of clofibrate on d14 and 19 (p < 0.05).
- There was no interaction between maternal clofibrate and age on the ratio of CO2 and ASP (C/A). Maternal clofibrate also had no impact on the ratio, but the ratio increased with age. On average the ratio increased by 2.75-fold after d7 compared to d1 (p < 0.0001). The ratio of oxidized and metabolized products (O/M) in piglets from the control sows decreased by 34% from d1 to d7 but increased by 106% from d7 to d14 and by 37% from d14 to d19 (p < 0.001). Maternal clofibrate had no impact on the ratio at d1 and d7 but increased greatly after d7. The ratio in piglets from clofibrate-fed sows was on average 13% and 39% higher than control sows (p < 0.005).
2.1.3. Effects of Carnitine and Malonate on Fatty Acid Metabolism in the Intestinal Mucosa of Piglets During the Neonatal-Suckling Period
- No interaction (Figure 1A) was detected between maternal clofibrate (p = 0.9) and the tissue treatment with carnitine or malonate for CO2 production. However, the supplementation of carnitine increased 14C accumulation in CO2 by 18% compared to control (p < 0.05). Supplementation of malonate decreased 14C accumulation in CO2 by 61% compared to control (p < 0.0001). No improvement was observed after adding carnitine to the treatment with malonate. Supplementation of carnitine or/and malonate in the mucosa incubation had no impact on the accumulation in ASP (p > 0.1). No interaction was detected between maternal clofibrate (p > 0.1) and the tissue treatment with carnitine or malonate. The tissues from all pigs treated with carnitine and malonate had no impact on the ESP production (p > 0.1).
- Total oxidized products (CO2 + ASP) and the total metabolites (CO2+ASP+ESP) were not affected by supplementation of carnitine or/and malonate (p > 0.1). No interaction was detected between maternal clofibrate (age) and the tissue treatment with carnitine or malonate (Figure 1B). Addition of carnitine increased the ratio of O/M regardless of malonate treatment (p < 0.0001), but no interaction was detected between maternal clofibrate (age) and the tissue treatment with carnitine or malonate.
2.1.4. Effects of CARNITINE and malonate on Distribution (%) of CO2, ASP and ESP in Total Fatty Acid Oxidation and Metabolism in the Intestinal Mucosa of Piglets During the Neonatal-Suckling Period
- There was no interaction between maternal clofibrate treatments and the treatments with carnitine and/or malonate (p > 0.05). However, significant interactions were detected between adding carnitine or/and malonate and postnatal age for % of CO2 and ASP (p < 0.0 1) in total oxidation and the C/A (p < 0.0001) as well as the % of CO2 (p < 0.0001) and ASP (p < 0.01) in the total metabolism. The interaction for ESP % in the total metabolism and the O/M also tended to be significant (p = 0.052).
- Addition of carnitine in the incubation medium had no influence on the % of CO2 and ASP in the total oxidized products (p > 0.1), but addition of malonate reduced % of CO2 (Figure 2A) and increased % of ASP (Figure 2B) in all ages (p < 0.05). The decrease was greater from the addition of carnitine+malonate than malonate when compared to the control. The ratio of CO2/ASP in control group increased quadratically with age, but the increase was reduced by the addition of carnitine. Addition of malonate inhibited the ratio increase with age and kept the ratio with no difference from the d1.
- A similar pattern of % of CO2 (Figure 3A) as in oxidation was observed from control group. However, the % of ASP (Figure 3B) significantly decreased after d1, and the decrease was greater from d7 than d14 and 19. Addition of carnitine or/and malonate increased % of ESP (Figure 3C) and the increase was greater from carnitine+malonate than carnitine or malonate only. No difference was observed between d14 and 19. The % of ESP increased on d7 and decreased after d7. The increase was reduced by the addition of carnitine and increased by the addition of malonate. No impacts were detected after d7. The ratio of oxidation and metabolism followed the same pattern as observed in % ASP.
2.2. NEFA and TG Concentrations
Maternal clofibrate supplementation had no impact on NEFA and TG concentration in the intestinal mucosa (p > 0.1). The average NEFA and TG level in the tissue were 52.94 ± 5.47 (µmol/g) and 2.39 ± 0.30 (μg/mg protein) respectively. However, both NEFA (Figure 4A) and TG (Figure 4B) varied greatly with postnatal age, in which a significant linear decrease with postnatal age was observed in the TG concentration and a quadratic response to the postnatal age was observed in NEFA concentration (p < 0.005). No interaction (p > 0.05) was detected between maternal clofibrate and postnatal age.
2.3. CPT Enzyme Activity
Maternal supplementation of clofibrate had no significant impact on the activities of CPT1, CPT2 and Total (CPT1 + CPT2), except for the trends test using orthogonal polynomial coefficients that showed the CPT1 activity tended increase linearly with maternal clofibrate dose (p=0.067). Postnatal age had a greater impact on CPT2 activity and the total activity of CPT (p < 0.001), and the impact followed a quadratic pattern (p < 0.01). The highest activity was observed in piglets at 14 days of age (Figure 4C).
2.4. Gene Expression (qPCR)
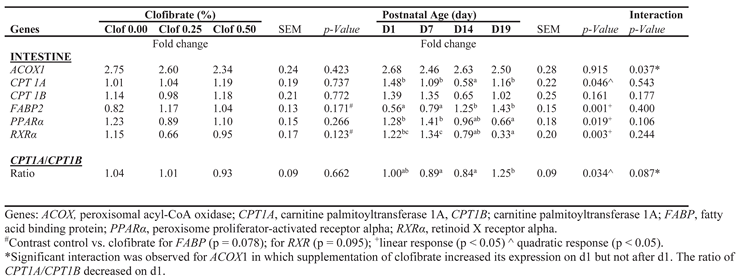
Maternal supplementation of clofibrate had no impact on the expression of genes examined in intestine mucosa of pigs (Table 3). However, postnatal age had an impact on the expression of CPT1A (p < 0.05), FABP (p < 0.001), PPARα (p < 0.05) and Retinoid X receptor alpha (RXRα; p < 0.005). The expression FABP was on average 90% higher in pigs at d14 and 19 than d1 and 7, while the expression of CPT1A was 53% lower on d14 than on the average of d1, d7 and d19. PPARα and RXRα were on average 50% and 74% lower in pigs at d19 than d1 and d7. In addition, the interaction between maternal clofibrate and postnatal age was significant for ACOX expression (p < 0.05) and tended to be significant for the ratio of CPT1A and CPT1B (p = 0.087), in which maternal supplementation of clofibrate increased ACOX expression but decreased the ratio of CPT1A and CPT1B on d1.
3. Discussion
3.1. The Effect of Maternal Clofibrate on Intestinal Fatty Acid Metabolism in Suckling Piglets
Development and growth of the intestine is rapid after birth, and increasing FA oxidation could be important for the intestine to meet the energy requirement and promote the fetal-type enterocyte maturation because milk fat is the primary energy source at birth. In this study, we evaluated the effects of PPARα activation on intestinal FA metabolism in piglets by feeding the sows clofibrate (a PPARα agonist) during late gestation and early lactation. The effects of clofibrate varied with dose and piglet postnatal age. In general, the intestinal oxidation of oleate to CO2 increased linearly with the postnatal age. Maternal supplementation of clofibrate promoted the increase throughout the entire postnatal period. Consistent with the increase in CO2 production, the TG and NEFA levels decreased with postnatal age, demonstrating the role and importance of TG and NEFA in intestinal energy supplies. Furthermore, stimulation of CO2 production was impacted by clofibrate doses and by postnatal age despite clofibrate being undetectable in the milk. Similar results were not observed in liver metabolism with the same treatments after d1 [14]. Given the detection limit of measuring clofibrate, whether the variation was associated with a potential very trace amount of clofibrate in the milk was not known. In addition, the variations in CO2 production were not in conjunction with an increase in CPT1 activity and gene expression of PPARα and RXRα in the present study although a higher CPT2 activity was observed on d14. CPT1A and CPT1B are present on the mitochondrial membrane in most tissues, while pig has an atypical molecular structure of CPT1, and the sensitivity of CPT1A and CPT1B to malonyl-CoA inhibition are different from other animals [19], in which CPT1A has a high sensitivity to malonyl-CoA inhibition and CPT1B has a low sensitivity to malonyl-CoA inhibition [20]. Therefore, we examined the CPT1A and CPT1B gene expression and were unable to detect any changes in intestine of piglets from sows with clofibrate supplementation in early lactation. However, we noticed that the ratio of CPT1A/CPT1B decreased with postnatal age. The interaction between maternal clofibrate and piglet age for the ratio tended to be significant (p = 0.087) and the ratio was lower on d1 from clofibrate than control. The changes in CPT1A and CPT1B ratio might influence the sensitivity of CPT1 to malonyl-CoA inhibition. The relative decrease in highly malonyl-CoA sensitive CPT1A might be associated with the CO2 increase. In addition, interaction between maternal clofibrate and postnatal age for ACOX1 expression was detected with a higher expression on d1. Also, FABP2 expression increased with age, indeed, suggesting that ACOX1 and FABP play a role in intestinal FA oxidation as observed in rats [11]. Furthermore, it is possible that genes associated with TCA that were not examined in this study contributed to the increase in CO2 production. CO2 production is indicative of increased ATP production through the TCA, and it has been reported that intermediates of the TCA were altered with clofibrate treatment [21].
Compared to CO2, ASP production had completely different responses. The dose-dependent stimulation of oleate oxidation to ASP by maternal clofibrate was detected only on day1. Combing CO2 and ASP together, the effect of maternal clofibrate on FA oxidation followed the same pattern as observed in ASP production. This result was consistent with that observed in liver of piglets receiving clofibrate from maternal supplementation, demonstrating clofibrate transferred from sows to fetal piglets via the placenta. After d1, ASP production decreased dramatically, and the reduced ASP production was not affected by maternal clofibrate supplementation or by further advancement in postnatal age. The lack of response to maternal clofibrate could be due to a low mammary transfer efficiency or dose of clofibrate supplementation, because the dose used in previous study with lactating rats was at least 7-fold [18] higher than the dose we fed to the sows. In addition, ketogenic capability was observed in the small intestine of rats [22] and can be induced by ketogenic diet [23], but the expression of ketogenic rate-limiting enzyme mitochondrial 3-hydroxy-3-methylglutaryl-CoA synthase (HMGCS) was not detectable in the small intestine of neonatal pigs [24]. Butyrate was converted into ketone bodies observed in colonocytes with increasing concentrations of [1-14C] butyrate [25], but gene expression almost was not detected in duodenum, jejunum and ileum until 8 weeks of age. Therefore, the reduced ASP production appeared to be associated with the inherent defect in ketogenic ability in intestine of neonatal piglets. In addition, the PPARα, and RXRα significantly decreased after d7. The reduced ASP production appeared to be associated with the lack of ketogenic ability and gene expression, while the high ASP production on d1 could be related to the high-level TG and NEFA as well as accumulation of the absorbed soluble carnitine esters after birth.
A previous study with hepatocytes isolated from suckling newborn and 15 d-old pigs showed that neonatal piglets have a huge rate of hepatic fatty acid esterification [26]. The rate of fatty acid esterification was associated with the rapid increase in fat- stores after birth [27]. Indeed, the hepatic esterification increased greatly with the postnatal age [14]. A similar increase in ESP production on d2 was also observed in a previous study with pig intestinal cells [28]. It is very interesting, however, that the intestinal esterification rate increased after birth and decreased greatly after 7 days, and maternal clofibrate had a great impact on the increase of the FA esterification on d7. The high ESP measured in pigs from control sows was reduced greatly compared to pigs from sows with clofibrate. The reduction was much greater from high maternal clofibrate than low maternal clofibrate, suggesting that maternal clofibrate could affect the ESP after birth and the degree of effect depended on the dosage of maternal clofibrate. However, no clofibrate effect was detected when the ESP decreased quickly with age after d7. These changes were consistent with the decrease in PPARα, RXRα expression and increase in FABP expression with age. The results from this study highlighted the high esterification capacity for FA in the newborn pig intestine, which may preserve this major colostrum FA for delivery to other tissues. Nutritional control of intestinal FA esterification, such as dietary fat intake, has been described in the literature [29]. Milk fat usually is high after farrowing and in the first several days after farrowing depending on species. Sow milk fat peaked around d3 and then decreased as lactation progressed [30]. We were interested in the potential influence of the decreased ESP on FA oxidation. The decrease in esterification on d7 and after d7 clearly illustrate that it has limited impact on overall oxidation. Combining the oxidative products and ESP, the total metabolites were increased at d1 primarily due to an increase in oxidation induced by clofibrate, while the total metabolism was decreased at d7 primarily due to a decrease in esterification.
Although the stimulatory effect of clofibrate on fatty acid oxidation had a significant interaction with the postnatal age, the interaction did not affect oxidative product distributions between CO2 and ASP. In general, the CO2 increased with clofibrate dose and postnatal age while the ASP decreased with clofibrate and postnatal age. Both linear and quadratic responses were detected. However, the distributions of CO2, ASP and ESP in total metabolism were impacted significantly by maternal clofibrate and postnatal age. In general, the % of oxidative products on average increased and the % of ESP decreased in piglets from sows receiving clofibrate during late gestation and early lactation. The effect of maternal clofibrate on the distribution of metabolic products appeared to be different from that in liver [14], implying the tissue specificity in which the absorption of dietary FA and converting them into TG for transport to other tissues is the primary pathway but not for producing ketone bodies and FA synthesis. The limited FA synthetic activity and rapid reduction in FA esterification are also the key factors in altering the metabolic allocations.
3.2. The effect of Providing Carnitine and Inhibiting TCA Activity on Intestinal Fatty Acid Metabolism in Suckling Pigs
Providing carnitine increased CO2 production and the oxidative proportion of the total metabolism, suggesting that the stimulatory role of carnitine might be limited to the entry of FA into mitochondria. This was confirmed by inhibiting TCA cycle activity via the addition of malonate, an inhibitor of succinate dehydrogenase. As the product of β-oxidation, acetyl-CoA can be metabolized completely via TCA cycle or be converted to acetate, acetyl-carnitine, HMG-CoA for ketogenesis or malonyl-CoA for FA synthesis, which comprises most of the ASP. We expected that ASP would increase when carnitine was added into the system and both ASP and ESP would increase when TCA cycle was inhibited. However, no influences on ASP and ESP were detected by the addition of carnitine or/and malonate. Available evidence in the literature showed that lipogenesis in pigs is extremely low in tissues other than adipose [31]. Consistent with this observation, the activities of enzymes related to lipogenesis are also very low in intestine [32]. In addition to lipogenesis, the gene of the key enzyme HMGCS for ketogenic pathway is undetectable in the small intestine of neonatal pigs [24]. The lack of lipogenic and ketogenic activities appeared to be associated with the lower response of ASP to carnitine and malonate. Although the status of carnitine acetyltransferase and acetyl-hydrolase in pig intestine was not evaluated in this study, and are not available in literature, our results illustrated and supported that the energy generation pathway is the main flux within intestinal FA metabolism. Energy generated from FA oxidation is the primary fate in intestinal mucosal fatty acid metabolism such as the TG/FA absorption and FABP synthesis.
4. Materials and Methods
4.1. Animals and Treatments
The animal study was described previously [14]. Briefly, a total of 27 gestating sows (in 3 blocks and 9 sows each block) were divided into three groups and fed a standard commercial corn/soybean-meal diet supplemented with three levels of clofibrate: 0% (Control), 0.25% (Clof 0.25), and 0.5% (Clof 0.5) of feed. The treated sows received either clofibrate dissolved in 15 mL of ethanol or 15 mL of ethanol vehicle. The sows were fed twice daily, but the ethanol with or without clofibrate was given to all the sows in the morning via mixing with and feeding in small amount of feed. The clofibrate supplementation started from one-week pre farrowing and ended with one-week post farrowing. The Institutional Animal Care and Use Committee (IACUC) at North Carolina State University, IACUC ID 16-142, approved animal care and all experimental procedures.
Piglets with average body weight from each litter at each time point were selected on days 1, 7, 14, and 19. The selected piglets were euthanized via American Veterinary Medical Association approved exsanguination while under anesthesia. Intestinal mucosa was collected from the proximal and distal ends of the small intestine from the piglets at each time point in 30-centimeter segments. The segment was rinsed with 0.9% NaCl solution and then opened lengthwise. With the segment on a smooth flat surface, a glass microscope slide was used to scrape the mucosa free. The first 30 centimeters of the proximal end was used for fatty acid oxidation measurement and the second 30 centimeters were frozen in liquid nitrogen for later analysis.
4.2. Fatty Acid Metabolism Measurements
The collected mucosa was homogenized in a handheld glass homogenizer (7 mL, Dounce Tissue Grinder; Vineland, NJ 08360, USA) in a buffer containing 220 mM mannitol, 70 mM sucrose, 2 mM N-2-hydroxyethylpiperazine-N-2-ethanesulfonic acid, 0.1mM EDTA with a ratio of 1:4 (w:v). FA oxidation was measured in a reaction buffer incubated with the fresh mucosa homogenate using [1-14C]-oleic acid as a substrate (1 mM; 0.25μCi/μmole). The measurements were performed in the presence or absence of L-carnitine (1 mM) with or without malonate (5 mM), an inhibitor of tricarboxylic acid cycle (TCA), in a reaction buffer reported by [33]. The CO2, acid soluble products (ASP) and esterification products (ESP) generated from the measurements and the homogenate protein were determined following the procedures described previously [14].
4.3. Non-Esterified Fatty Acids (NEFA) and Triglycerides (TG) Assays
TG and NEFA were determined in the frozen mucosa using the commercial kits from MyBioSource Inc (MBS9719080 and MBS2556986; San Diego, CA, USA) by following the kit instructions.
4.4. Enzymatic Assay
Specific activity of carnitine palmitoyl transferase (CPT) was determined in homogenate from the frozen mucosa samples with and without addition of malonyl-CoA (10 mM). The determination was performed following the procedure as described previously [34].
4.5. RNA Isolation and RT-qPCR
Total RNA was isolated from the mucosa using Tri reagent with 50 mg of the frozen samples following the manufacturer’s procedure. Quantification and quality control were performed using a Nanodrop spectrophotometer (ND-1000ThermoFisher, Wilmington, DE), and 0.8% agarose gel electrophoresis post DNase treatment. cDNA synthesis was performed using Super Script III Reverse Transcriptase as described previously [15]. Primers for RT-qPCR (Supplemental Table 1) were created using BLAST Primer Designer (https://www.ncbi.nlm.nih.gov/tools/primer-blast/). The RT-PCR reaction and the ct values were normalized to a housekeeping gene (RPL9) and a plate normalizer was included on each plate to account for run differences.
4.6. Chemicals
All chemicals for this study were sourced from Sigma-Aldrich, Inc (St. Louis, MO 63103), except for clofibrate that came from Cayman Chemical (Ann Arbor, MI 48108), Superscript from Thermo Fisher Scientific (Waltham, MA02451), and TurboDNase from Ambion. The 14C radiolabeled oleic acid and acetyl-CoA were purchased from American Radiolabeled Chemicals (St Louis, MO 63146) and SYBR Green from BioRad (Hercules, CA94547).
4.7. Statistical Analysis
Data from enzymatic and RT-qPCR assays were analyzed according to a 3 (Control, Clof 0.25 and Clof 0.5) x 4 (d1, 7, 14 and 19) factorial randomized complete block design (blocked by sow (litter)). Data from in vitro fatty acid oxidation was analyzed according to split-plot design. The main plot was the 3 maternal dietary treatments on animals at 4 ages, and the sub-plot was the 4 treatments with a 2 (± carnitine) x 2 (± malonate) factorial design on the tissues. Analyses were performed using the General Linear Models (GLM) procedure of SAS (SAS software 9.4; Cary, NC USA). Testing for trends (linear and quadratic) was performed also with the contrast statement using orthogonal polynomial coefficients. The least square means (Lsmeans) were calculated and the interactions between clofibrate x postnatal age, and between (clofibrate x postnatal age) x treatments were tested. The data from main effects were reported only if the interaction was not significant. Data are presented as Lsmeans ± standard error means unless specified otherwise. Difference was reported as significant when p < 0.05 and as trends when 0.05 < p < 0.1.
5. Conclusions
Maternal supplementation of clofibrate at late gestation and early lactation improves energy generation via increasing β-oxidation activity, decreasing FA esterification and promoting TG and NEFA utilization. However, the improvement varies with the increase in postnatal age, in which the alterations of gene expression of ACOX and FABP as well as ratio of CPT1A/CPT1B may play regulatory roles. Addition of carnitine increases FA oxidation, but the increase is limited by the activity of TCA cycle. In addition, the dose of clofibrate and the efficiency in delivery of clofibrate via milk apparently need to be investigated in the future.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1: Primers used in this study with expected amplicon size post RT-qPCR. Gene Bank numbers demonstrate the genes of the pig genome referenced.
Author Contributions
Conceptualization, B.P., L.X. and J.O.; methodology, B.P., J.H., F.W, P.M. and L.Y.; formal analysis, B.P., J.H., F.W., P.M. and L.Y.; investigation, B.P., J.Z.; data curation, B.P., J.H., J.Z and F.W.; writing—original draft preparation, B.P; writing—review and editing, B.P, L.X., H.L. and J.O.; supervision, L.X. and H.L.; project administration, L.X.; funding acquisition, L.X. and J.O. All authors have read and agreed to the published version of the manuscript.
Funding
This work is supported by Animal Nutrition, Growth and Lactation [grant no. 2015-67015-23245 /project accession no. 1005855] and [2023-67015-39663/project accession no 1030033] from the USDA National Institute of Food and Agriculture, and by the North Carolina Agricultural Research Hatch projects 1016618 and 02780. Funds received from the grants cover the costs of publishing this article in open access.
Institutional Review Board Statement
The study was conducted in accordance with the protocol submitted to the Institutional Animal Care and Use Committee (IACUC) at North Carolina State University, IACUC ID 16-142, approved animal care and all experimental procedures.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data is contained within the article and will be available in PubAg.
Acknowledgments
The authors wish to express thanks to Libby Pratt, Zoe Olmstead, and Megan Wallace for help with animal feeding, sample collection, and FA analysis.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Abbreviations
The following abbreviations are used in this manuscript:
| ACOX1 | Acyl-CoA oxidase 1 |
| ASP | Acid soluble products |
| ESP | Esterification products |
| FA | Fatty acid |
| FABPs | FA-binding proteins |
| HMGCS | 3-hydroxy-3-methylglutaryl-CoA synthase |
| NEFA | Non-Esterified Fatty Acids |
| RXRα | Retinoid X receptor alpha |
| TCA | Tricarboxylic acid cycle |
| TG | Triglycerides |
References
- Herrera E, Amusquivar E. Lipid metabolism in the fetus and the newborn. Diabetes Metab Res Rev. 2000, 16(3):202-10. [CrossRef]
- Girard J, Ferré P, Pégorier JP, Duée PH. Adaptations of glucose and fatty acid metabolism during perinatal period and suckling-weaning transition. Physiol Rev. 1992, 72(2):507-62. [CrossRef]
- Hahn P, Koldovskv O: Utilization of Nutrients During Postnatal Development. Oxford,Pergamon Press, 1966, p 17.
- Weström B, Arévalo Sureda E, Pierzynowska K, Pierzynowski SG, Pérez-Cano FJ. The Immature Gut Barrier and Its Importance in Establishing Immunity in Newborn Mammals. Front Immunol. 2020, 9;11:1153. [CrossRef]
- Kimura RE. Neonatal intestinal metabolism. Clin Perinatol. 1996, 23(2):245-63. [CrossRef]
- Girard J, Duée PH, Ferré P, Pégorier JP, Escriva F, Decaux J F. Fatty acid oxidation and ketogenesis during development. Reprod. Nutr. Develop. 1985, 25(1 B), 303-319. [CrossRef]
- Small,G. M., T. J. Hocking, A. P. Strudee, K. Burdett, and M. J. Connock. 1981. Enhancement by dietary clofibrate of peroxisomal palmityl-CoA oxidase in kidney and small intestine of albino mice and liver of genetically lean and obese mice. Life Sci. 1981, 28:1875–82. [CrossRef]
- Kimura R, Takahashi N, Murota K, Yamada Y, Niiya S, Kanzaki N, Murakami Y, Moriyama T, Goto T, Kawada T. Activation of peroxisome proliferator-activated receptor-alpha (PPARalpha) suppresses postprandial lipidemia through fatty acid oxidation in enterocytes. Biochem Biophys Res Commun. 2011, 24;410(1):1-6. [CrossRef]
- Kimura R, Takahashi N, Lin S, Goto T, Murota K, Nakata R, Inoue H, Kawada T. DHA attenuates postprandial hyperlipidemia via activating PPARalpha in intestinal epithelial cells. J Lipid Res. 2013, 54(12):3258-68. [CrossRef]
- Karimian Azari E, Leitner C, Jaggi T, Langhans W, Mansouri A. Possible role of intestinal fatty acid oxidation in the eating-inhibitory effect of the PPAR-alpha agonist Wy-14643 in high-fat diet fed rats. PLoS One. 2013, 17;8(9):e74869. [CrossRef]
- Mochizuki K, Suruga K, Yagi E, Takase S, Goda T. The expression of PPAR-associated genes is modulated through postnatal development of PPAR subtypes in the small intestine. Biochim Biophys Acta. 2001, 1531(1-2):68-76. [CrossRef]
- Ringseis R, Eder K. 2009. Influence of pharmacological PPARalpha activators on carnitine homeostasis in proliferating and non-proliferating species. Pharmacol. Res. 2009, 60:179–84. [CrossRef]
- Shim K, Jacobi S, Odle J, Lin X. Pharmacologic activation of peroxisome proliferator-activating receptor-α accelerates hepatic fatty acid oxidation in neonatal pigs. Oncotarget. 2018, 8;9(35):23900-23914. [CrossRef]
- Zhao J, Pike B, Wang F, Yang L, Meisner P, Huang Y, Odle J, Lin X. Effects of maternal feeding of clofibrate on hepatic fatty acid metabolism in suckling piglet. J Anim Sci Biotechnol. 2024, 5;15(1):163. [CrossRef]
- Pike B, Zhao J, Hicks JA, Wang F, Hagen R, Liu HC, Odle J, Lin X. Intestinal Carnitine Status and Fatty Acid Oxidation in Response to Clofibrate and Medium-Chain Triglyceride Supplementation in Newborn Pigs. Int J Mol Sci. 2023, 23;24(7):6066. [CrossRef]
- Lin X, Jacobi S, Odle J. Transplacental induction of fatty acid oxidation in term fetal pigs by the peroxisome proliferator-activated receptor alpha agonist clofibrate. J Anim Sci Biotechnol. 2015, 26;6(1):11. [CrossRef]
- Simpson AE, Brammar WJ, Pratten MK, Cockcroft N, Elcombe CR. Placental transfer of the hypolipidemic drug, clofibrate, induces CYP4A expression in 18.5-day fetal rats. Drug Metab Dispos. 1996, 24(5):547-54.
- Simpson AE, Brammar WJ, Pratten MK, Elcombe CR. Translactational induction of CYP4A expression in 10.5-day neonatal rats by the hypolipidemic drug clofibrate. Biochem Pharmacol. 1995, 22;50(12):2021-32. [CrossRef]
- Nicot C, Hegardt FG, Woldegiorgis G, Haro D, Marrero PF. Pig liver carnitine palmitoyltransferase I, with low Km for carnitine and high sensitivity to malonyl-CoA inhibition, is a natural chimera of rat liver and muscle enzymes. Biochemistry. 2001, 20;40(7):2260-6.
- Relat J, Nicot C, Gacias M, Woldegiorgis G, Marrero PF, Haro D. Pig muscle carnitine palmitoyltransferase I (CPTI beta), with low Km for carnitine and low sensitivity to malonyl-CoA inhibition, has kinetic characteristics similar to those of the rat liver (CPTI alpha) enzyme. Biochemistry. 2004 Oct 5;43(39):12686-91.
- Voltti H, Hassinen IE. Effect of clofibrate on the hepatic concentrations of citric acid cycle intermediates and malonyl-CoA in the rat. Life Sci. 1981, 5;28(1):47-51. [CrossRef]
- Békési A, Williamson DH. An explanation for ketogenesis by the intestine of the suckling rat: the presence of an active hydroxymethylglutaryl-coenzyme A pathway. Biol Neonate. 1990, 58(3):160-5. [CrossRef]
- Wallenius V, Elias E, Elebring E, Haisma B, Casselbrant A, Larraufie P, Spak E, Reimann F, le Roux CW, Docherty NG, Gribble FM, Fändriks L. Suppression of enteroendocrine cell glucagon-like peptide (GLP)-1 release by fat-induced small intestinal ketogenesis: a mechanism targeted by Roux-en-Y gastric bypass surgery but not by preoperative very-low-calorie diet. Gut. 2020, 69(8):1423-1431. [CrossRef]
- Adams SH, Alho CS, Asins G, Hegardt FG, Marrero PF. Gene expression of mitochondrial 3-hydroxy-3-methylglutaryl-CoA synthase in a poorly ketogenic mammal: effect of starvation during the neonatal period of the piglet. Biochem J. 1997, 15;324 ( Pt 1)(Pt 1):65-73. [CrossRef]
- Darcy-Vrillon B, Cherbuy C, Morel MT, Durand M, Duée PH. Short chain fatty acid and glucose metabolism in isolated pig colonocytes: modulation by NH4+. Mol Cell Biochem. 1996, 23;156(2):145-51. [CrossRef]
- Pégorier JP, Duée PH, Girard J, Peret J. Metabolic fate of non-esterified fatty acids in isolated hepatocytes from newborn and young pigs. Evidence for a limited capacity for oxidation and increased capacity for esterification. Biochem J. 1983, 15;212(1):93-7. [CrossRef]
- Manners MJ, Mccrea MR. Changes in the chemical composition of sow-reared piglets during the 1st month of life. Br J Nutr. 1963, 17:495-513. [CrossRef]
- Cherbuy C, Guesnet P, Morel MT, Kohl C, Thomas M, Duée PH, Prip-Buus C. Oleate metabolism in pig enterocytes is characterized by an increased oxidation rate in the presence of a high esterification rate within two days after birth. J Nutr. 2012, 142(2):221-6. [CrossRef]
- Trotter PJ, Storch J. Nutritional control of fatty acid esterification in differentiating Caco-2 intestinal cells is mediated by cellular diacylglycerol concentrations. J Nutr. 1993, 123(4):728-36. [CrossRef]
- Suarez-Trujillo A, Luecke SM, Logan L, Bradshaw C, Stewart KR, Minor RC, Ramires Ferreira C, Casey TM. Changes in sow milk lipidome across lactation occur in fatty acyl residues of triacylglycerol and phosphatidylglycerol lipids, but not in plasma membrane phospholipids. Animal. 2021, 15(8):100280. [CrossRef]
- Huang WY, Kummerow FA. Cholesterol and fatty acid synthesis in swine. Lipids. 1976, 11(1):34-41. [CrossRef]
- Qureshi AA, Burger WC, Elson CE, Benevenga NJ. Effects of cereals and culture filtrate of Trichoderma viride on lipid metabolism of swine. Lipids. 1982, 17(12):924-34. [CrossRef]
- Lin X, Adams SH, Odle J. Acetate represents a major product of heptanoate and octanoate beta-oxidation in hepatocytes isolated from neonatal piglets. Biochem J. 1996, 15;318 (Pt 1) (Pt 1):235-40. [CrossRef]
- 34. Xi L, Matsey G, Odle J.The effect of 5-aminoimidazole-4-carboxamide ribonucleoside (AICAR) on fatty acid oxidation in hepatocytes isolated from neonatal piglets. J Anim Sci Biotechnol. 2012, 17;3(1):30. [CrossRef]
Figure 1.
Effects of carnitine and malonate on intestinal oleic acid oxidation in suckling piglets. Oleic acid oxidation was measured with or without carnitine (1 mM) and/or malonate (5 mM) in intestinal mucosa of pigs from sows with or without clofibrate (Con, control; Car, carnitine; Mal, malonate; CarMal, carnitine+malonate). Columns represent oxidative products (µmol/g protein . h)and are least square means ± standard error means (CO2; ASP, acid soluble product; ESP, esterified products; TTO, total oxidation product; TTM, total metabolic products; TTO/TTM, the ratio of TTO and TTM). abcColumns with differing letters denote significant differences (p < 0.05).
Figure 1.
Effects of carnitine and malonate on intestinal oleic acid oxidation in suckling piglets. Oleic acid oxidation was measured with or without carnitine (1 mM) and/or malonate (5 mM) in intestinal mucosa of pigs from sows with or without clofibrate (Con, control; Car, carnitine; Mal, malonate; CarMal, carnitine+malonate). Columns represent oxidative products (µmol/g protein . h)and are least square means ± standard error means (CO2; ASP, acid soluble product; ESP, esterified products; TTO, total oxidation product; TTM, total metabolic products; TTO/TTM, the ratio of TTO and TTM). abcColumns with differing letters denote significant differences (p < 0.05).
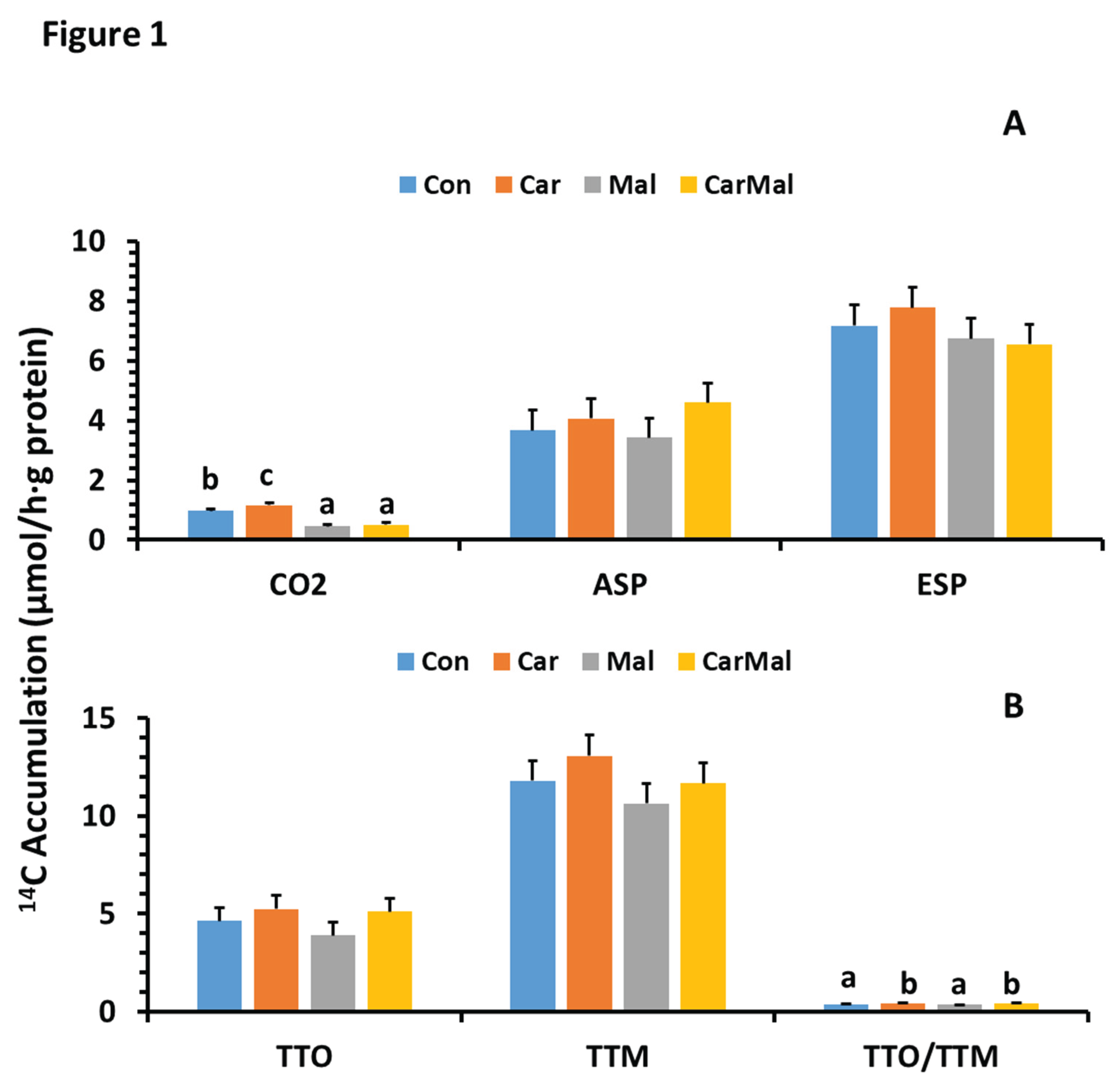
Figure 2.
Effects of carnitine and malonate on the distribution of intestinal oleic acid oxidation in piglets during the suckling period. Oleic acid oxidation was measured with or without carnitine (1 mM) or/and malonate (5 mM) in intestinal mucosa of pigs from sows with and without clofibrate (Con, control; Car, carnitine; Mal, malonate; CarMal, carnitine+malonate). Columns represent % of total oxidative products and are least square means ± standard error means for CO2 (A) and ASP, acid soluble product (B). Columns lacking a common letter are different (p < 0.05).
Figure 2.
Effects of carnitine and malonate on the distribution of intestinal oleic acid oxidation in piglets during the suckling period. Oleic acid oxidation was measured with or without carnitine (1 mM) or/and malonate (5 mM) in intestinal mucosa of pigs from sows with and without clofibrate (Con, control; Car, carnitine; Mal, malonate; CarMal, carnitine+malonate). Columns represent % of total oxidative products and are least square means ± standard error means for CO2 (A) and ASP, acid soluble product (B). Columns lacking a common letter are different (p < 0.05).
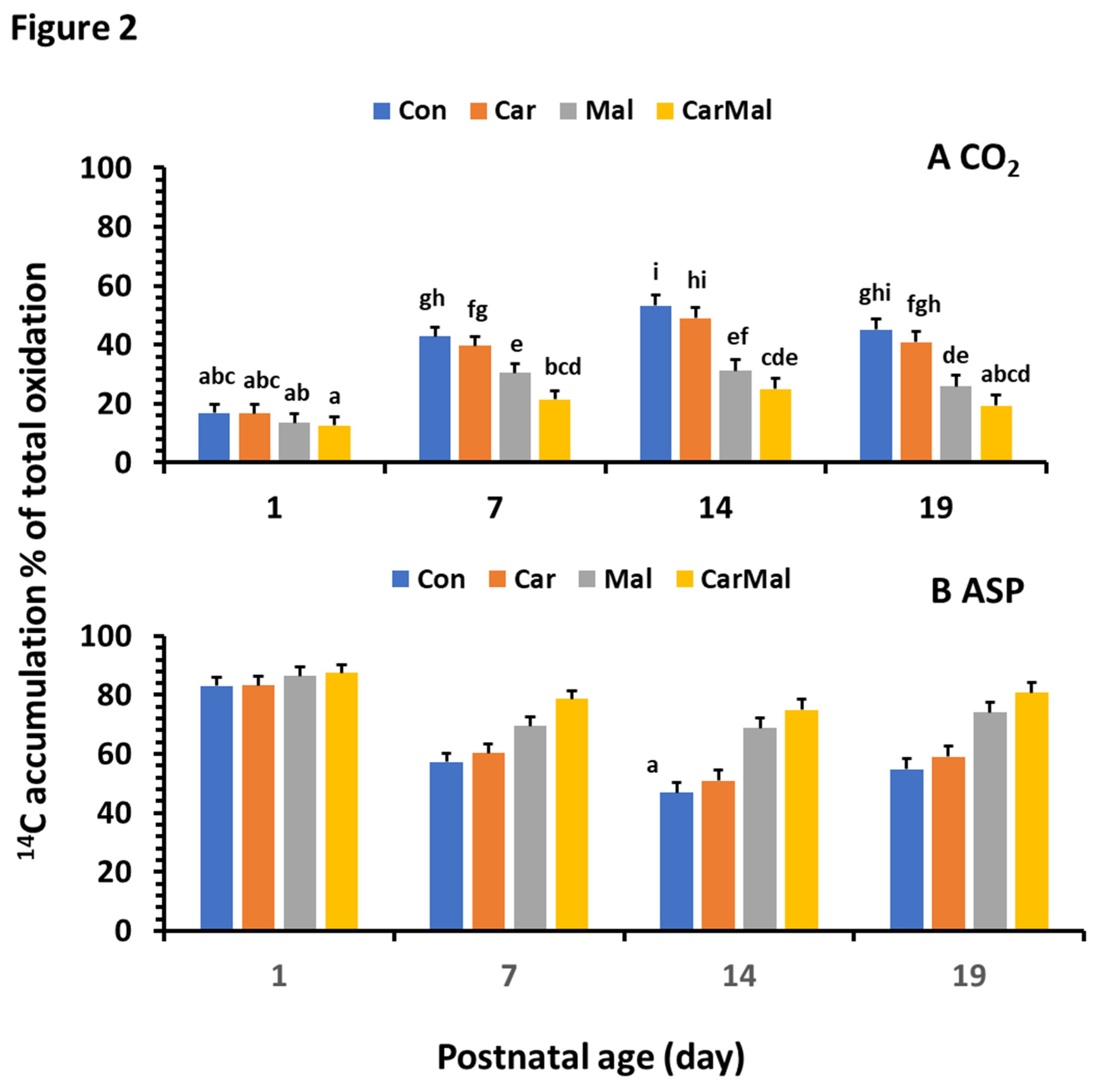
Figure 3.
Effects of carnitine and malonate on the distribution of intestinal oleic acid metabolism in piglets during the suckling period. Oleic acid oxidation was measured with or without carnitine (1 mM) or/and malonate (5 mM) in intestinal mucosa of pigs from sows with and without clofibrate (Con, control; Car, carnitine; Mal, malonate; CarMal, carnitine+malonate). Columns represent % of total metabolic products and are least square means ± standard error means for CO2 (A); ASP, acid soluble product (B); and ESP, esterified products (C). Columns lacking a common letter are different (p < 0.05).
Figure 3.
Effects of carnitine and malonate on the distribution of intestinal oleic acid metabolism in piglets during the suckling period. Oleic acid oxidation was measured with or without carnitine (1 mM) or/and malonate (5 mM) in intestinal mucosa of pigs from sows with and without clofibrate (Con, control; Car, carnitine; Mal, malonate; CarMal, carnitine+malonate). Columns represent % of total metabolic products and are least square means ± standard error means for CO2 (A); ASP, acid soluble product (B); and ESP, esterified products (C). Columns lacking a common letter are different (p < 0.05).
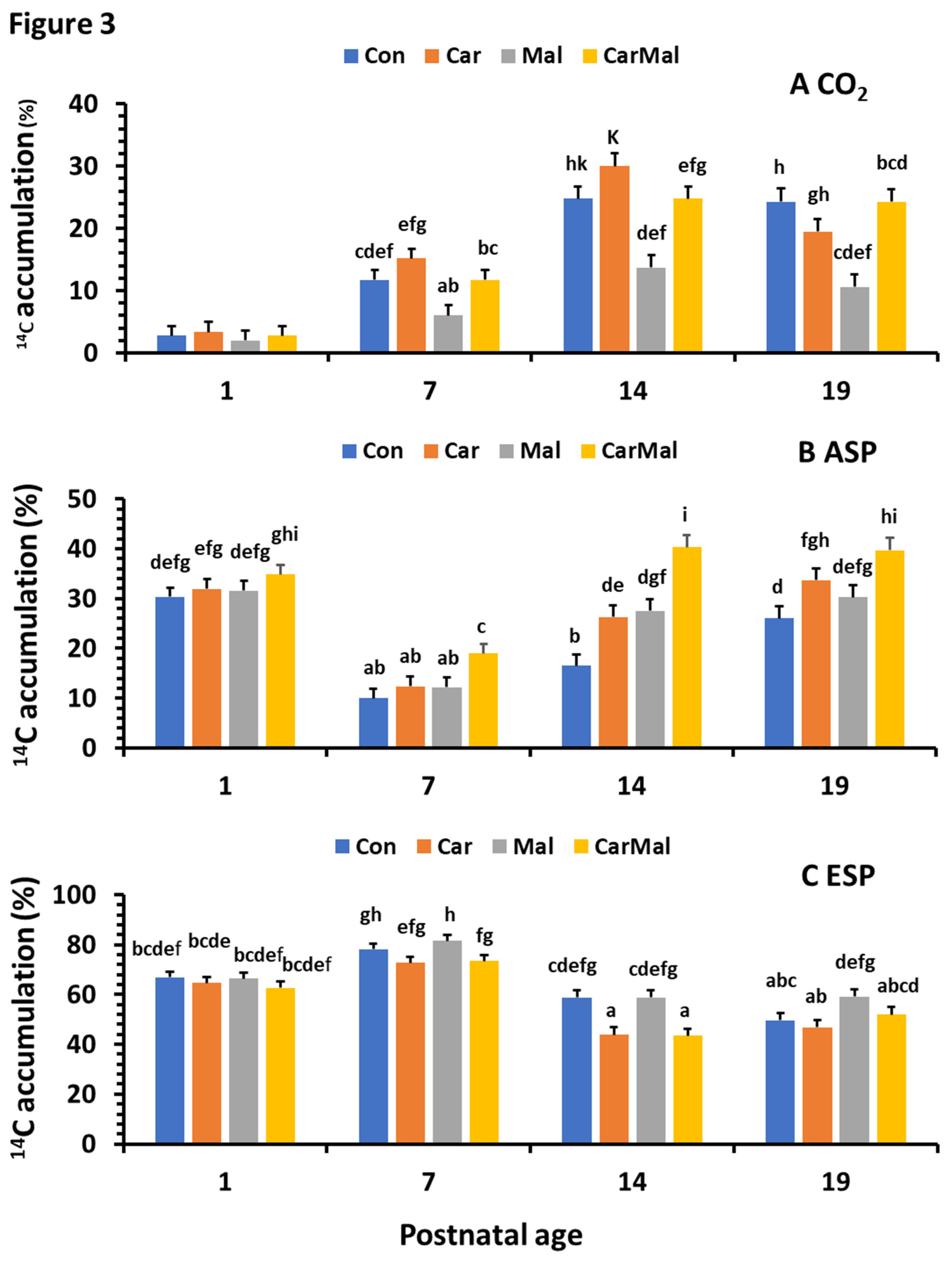
Figure 4.
Effect of postnatal age on intestinal NEFA, TG and CPT activity in suckling piglets. NEFA (A) and TG (B) were determined using the commercial kits from MyBioSource Inc (MBS9719080 and MBS2556986; San Diego, CA, USA). The type and color of the markers represented data from maternal treatments (Clof 0, control; Clof 0.25 clofibrate 0.25; Clof 0.5 clofibrate 0.5) and the curve was from regression analysis with data from all treatments (R2 = 0.2 and p < 0.0001) for NEFA and (R2=0.17 and p < 0.001) for TG). CPT activity (C; R2=0.1 and p < 0.05) was measured using radio-enzymatic assay [34].
Figure 4.
Effect of postnatal age on intestinal NEFA, TG and CPT activity in suckling piglets. NEFA (A) and TG (B) were determined using the commercial kits from MyBioSource Inc (MBS9719080 and MBS2556986; San Diego, CA, USA). The type and color of the markers represented data from maternal treatments (Clof 0, control; Clof 0.25 clofibrate 0.25; Clof 0.5 clofibrate 0.5) and the curve was from regression analysis with data from all treatments (R2 = 0.2 and p < 0.0001) for NEFA and (R2=0.17 and p < 0.001) for TG). CPT activity (C; R2=0.1 and p < 0.05) was measured using radio-enzymatic assay [34].

Table 1.
Effects of maternal clofibrate and postnatal age on oleic acid metabolism in small-intestinal mucosa of suckling pigs.
Table 1.
Effects of maternal clofibrate and postnatal age on oleic acid metabolism in small-intestinal mucosa of suckling pigs.
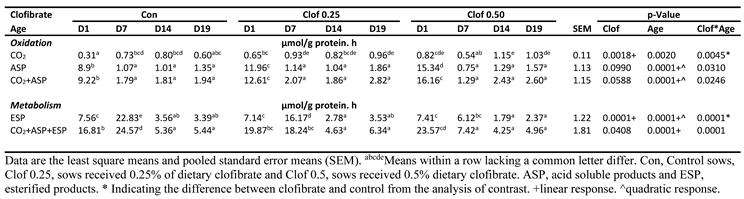 |
Table 2.
Effects of maternal clofibrate and postnatal age on product distribution from oleic acid metabolism in small-intestinal mucosa from suckling pigs.
Table 2.
Effects of maternal clofibrate and postnatal age on product distribution from oleic acid metabolism in small-intestinal mucosa from suckling pigs.
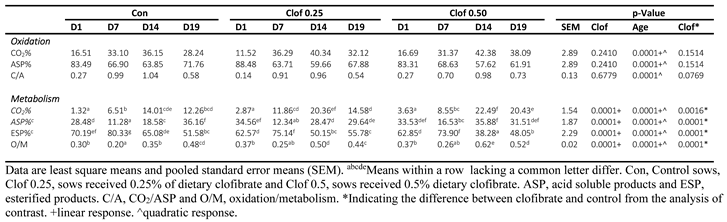 |
Table 3.
Effect of maternal clofibrate and postnatal age on intestinal mucosa gene expressions of piglets during neonatal suckling period.
Table 3.
Effect of maternal clofibrate and postnatal age on intestinal mucosa gene expressions of piglets during neonatal suckling period.
 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



