
Submitted:
23 June 2025
Posted:
24 June 2025
You are already at the latest version
Abstract
This study focuses on the identification of three lactic acid bacteria isolates obtained from traditional Algerian fermented wheat, as well as the evaluation of their biological activities mainly probiotic, antimicrobial, anti-inflammatory, and antioxidant properties. The identification of these isolates was carried out through phenotypic and genotypic characterizations. It was found that isolate LB3 was Lactiplantibacillus plantarum, however isolates LB1 and LB2 were identified as Weissella confusa. Moreover, the strains LB1, LB2 and LB3 are capable of maintaining their growth at pH 3 and in bile salts after 4 h with survival rates between 41% and 90%. Furthermore, their co-aggregation capacity with Staphylococcus aureus indicated a percentage higher than 50%. The three strains demonstrated strong inhibitory effects against pathogenic bacteria, showing inhibition of 5 % to 40%. They exhibited significant anti-inflammatory activity ranging from 20% to 39%. All strains revealed powerful ability to scavenge DPPH·, a greater ferric reducing activity, and high levels of glutathione. Additionally, they are capable of exerting a protective effect against plasma lipid peroxidation with inhibition rates ranging from 20% to 39%. These findings suggest that these strains possess promising probiotic potential for future therapeutic agent to be used in the development of novel functional fermented foods.
Keywords:
lactic acid bacteria
; probiotic
; traditional fermented wheat
; antioxidant activity
; antibacterial activity
; anti-inflammatory activity
1. Introduction
The gut microbiota is crucial in maintaining the organism's homeostasis. The imbalance of the latter induces an alteration in the functioning of the immune system, which promotes the development of certain pathologies such as allergies, chronic inflammatory diseases, and intestinal infections [1,2]. The causes of this dysbiosis are complex and probably attributable to several factors such as the environment, genetics, and diet. Indeed, its modulation through diet can be a key factor in ensuring and restoring its balance and consequently maintaining the host's health and well-being. For this reason, researchers are very interested in biological products such as beneficial bacteria, primarily probiotics. These are living microorganisms that, when administered in adequate quantities, would have a beneficial effect on the host's health [3].
These are often praised for their beneficial effects on health, as well as their ability to influence diseases such as cancer and obesity, which are associated with imbalances in the gut flora [4]. The effectiveness of these is mainly linked to the selection of the species and even the specific bacterial strain. According to Yadav and Shukla [5], lactic acid bacteria (LAB) stand out among the selected live bacterial strains for their crucial role in maintaining intestinal balance [6]. Due to their numerous health benefits, lactic acid bacteria are considered promising probiotic candidates. However, it is necessary to emphasize that not all bacteria can be considered probiotics, and it is essential to examine their probiotic properties as well as their safety. These microbes should have considerable resistance to persist in the stomach’s highly acidic conditions [7]. Furthermore, their ability to adhere in the gastrointestinal tract (intestinal mucosa and epithelial cells) prevents their elimination by intestinal motility. This adhesion allows them to reproduce, colonize, and affect the immune system, while competitively eliminating pathogens [8].
This ability demonstrates their aptitude for food bioconservation and can be used as starter culture in the fermentation process under controlled conditions. During fermentation, LAB synthesize a number of compounds, such as exopolysaccharides, aromatic compounds, organic acids, etc., which extend the shelf life of the food and improve its sanitary, sensory, and nutritional properties, as well as increase the antioxidant capacity of the fermented food. This increase is mainly due to the depolymerization of phenolic compounds [9]. Therefore, the valorization and investigation of probiotic potential of LAB isolated from various local fermented foods and products have awakened scientific interest in recent years. Fermented wheat, which is one of the food substrates stimulating their development, appears as a promising resource. It provides a natural support for the separation and analysis of LAB.
In Algeria, Hamoum, a traditionally fermented wheat, is naturally produced in an underground silo known as Matmora [10]. In addition to its sensory and gustatory attributes, fermentation enhances the nutritional properties of wheat and stimulates the multiplication of beneficial microorganisms. According to Nithya et al. [11], fermented wheat is recognized for its preventive virtues against intestinal pathophysiological dysfunctions. Lactic bacteria derived from fermented wheat could represent a promising solution commonly used in the medical and food sectors. In this context, the present study focuses on the identification of three selected LAB isolates obtained from traditional Algerian fermented wheat, as well as the in vitro evaluation of their probiotic, antibacterial, anti-inflammatory, and antioxidant properties.
2. Materials and Methods
2.1. Lactic Acid Bacteria Isolates
Three bacterial isolates were obtained from traditional fermented wheat, harvested in Rahwia,Tiaret region (Algeria). The pure culture isolates, previously stored at -20 °C, were transferred into deMan, Rogosa, Sharpe (MRS) broth and incubated at 37 °C for 24 h, either under aerobic conditions (LB1, LB2) or under anaerobic conditions (LB3).
2.2. Pathogenic Strains
The strains used for the evaluation of antibacterial activity are Escherichia coli (E.coli) ATCC 10536, Bacillus subtilis (B.subtilis) ATCC 6633, and Bacillus cereus (B.cereus) ATCC 10876, and Staphylococcus aureus (S.aureus) ATCC6528 provided by SAIDAL and the Mostapha Bacha Hospital in Algiers, as well as an isolate of Pseudomonas aeruginosa (P. aeruginosa) from a canine infection at the Veterinary Institute of Tiaret.
2.3. Phenotypic Identification of the Isolates
Morphological properties of the three strains were examined on MRS agar following 24 hours of incubation [12]. The catalase test and the fermentation type were evaluated according to the method described by Delarass [13]. Proliferation at various temperatures was recorded in MRS broth following incubation at 10°C, 30°C, and 45°C for 72 hours [14]. The resistance of bacterial isolates to NaCl was assessed in MRS broth with 6.5% and 9.5% NaCl at 30 °C for 48 hours [15].
2.4. Molecular Identification of Bacteria Using 16S rRNA Gene Sequencing
The genomic DNA of LAB isolates from traditional Algerian fermented wheat was extracted according to the optimized protocol at the Laboratory of Biotechnology and Bio-Geo- Resource Valorization. Higher institute of Biotechnology Sidi Thabet, Tunis, using the phenol-chloroform technique. The amplification and sequencing of the 16S rRNA gene were carried out utilizing universal primers (27f 5'AGAGTTTGATCMTGGCTCAG 3' and 1492R 5' GGTTACCTTGTTACGACTT3'). One microliter of extracted DNA was diluted to 1/10 and added to the reaction mixture comprising 2.5µL of PCR reaction (10×), 0.2 µL of deoxyribonucleoside triphosphates, 0.3 µL primer, and 0.2 µL of DNA polymerase, then adjusted to 25 µL with distilled water. PCR reactions were done on a thermocycler (T100 BIO RAD, USA) utilizing this program: hybridization at 95 °C for 3 min, this was succeeded by denaturation at 95°C for 30 s (35 cycles), the protocol was then continued with hybridization step at 57.5°C for 1 minute, 1 minute of elongation at 72°C, and conducted with 10 minutes of elongation cycle.
Sequencing was performed using the automated Sanger technique. The sequence obtained from the gene coding for 16S rRNA was compared with the homologous sequences of reference microbial species listed in Genbank, using BLAST NCBI (http://blast.ncbi.nlm.nih.gov). An evolutionary tree was also developed using MEGA 11 software to identify the most similar bacterial species using the neighbor-joining method [16]. The nucleotide sequences have been added in the GenBank with accession numbers PV652951, PV653095, and PV653185 for the isolated bacteria LB1, LB2, and LB3, respectively.
2.5. Probiotic Properties Assessment
2.5.1. Tolerance to Acidity
The tolerance of the strains to acidity was performed according to Anandharaj et al. [17] MRS medium was calibrate to pH 3 and pH 6.5, inoculated with overnight cultures, and incubated at 37°C. The growth was established over 3 h. The survival rate (%) was determined using the pour plate count method on MRS agar after incubation of 0 and 3 h.
2.5.2. Bile Salts Tolerance
The resistance ability of strains to bile salt was carried out by the method of Anandharaj et al [17]. The MRS medium supplemented with 0.3% (w/v) bile salts were inoculated with 18h culture of bacterial isolates (108 CFU/ml) and the incubation was carried out at 37°C. The viability rate (%) was assessed utilizing the colony counting technique on MRS agar after incubation of 0 and 4 h.
2.5.3. Antibiotic Sensitivity
Antibiotic sensitivity of strains was determined according to Tarique et al. [18]. The bacterial suspensions of each isolate regulated to 106 CFU/ml were introduced and kept at 37°C for incubation after the placement of antibiotic discs. The inhibition diameters were evaluated follwing this classification, resistant less than 1.5 cm, intermediate between 1.6 and 2.0 cm, sensitive more than 2.0 cm) [19].
2.5.4. Auto-Aggregation
The surface cell auto-aggregation was carried out by the modified protocol of Abdulla et al. [20]. The overnight cultures of bacterial isolates were subjected to centrifugation for 15 minutes at 4500 g (4°C), then rinsed with phosphate-buffered saline (PBS) (pH 7.2) and regulated to 108 CFU/ml (A0). A volume of 4 ml of suspended cells were mixed, maintained at 37°C for 3 h incubation, and the absorbance of the supernatant was determined at 625 nm (A1). The percentage of bacterial cells auto-aggregation was measured with the following equation (1):
A% =1- (A1/A0) ×100,
A1 is absorbance at 3h and A0 is absorbance at 0 h.
2.5.5. Co-Aggregation
The co-aggregation rates between the isolated strains and two pathogenic bacteria (S. aureus and E.coli) were performed by the protocol of Abushelaibi et al. [21]. The same volume of lactic and pathogenic strains suspensions were mixed, vortexed, and then subjedted to 3 hours of incubation at 37°C. The absorbance of the selected LAB strains (Ax), the pathogenic strains (Ay), and the mixture [A(x+y)] was recorded at 620 nm. The rate of co-aggregation was measured with the formula (2):
%= {(AX0 + AY0)/2 - A(x+y) / (AX0 + AY0)/2} x 100,
Where AX0 : Absorbance in the presence of LAB strain at 0h; AY0; absorbance in the presence of pathogenic bacteria at 0 h; A(x+y): Absorbance of the mixture after 3h of incubation).
2.5.6. Hemolytic Activity
The bacterial cultures were streaked on the surface of horse blood agar (5% v/v) and incubated for 48 h at 37°C. The degradation of the blood was noted by the formation of distinct halos (β hemolysis) or greenish areas (α hemolysis) surrounding the colonies [22].
2.5.7. Proteolytic Activity
The proteolytic property of the strains was relaized by the disc technique on MRS agar containing 10% skim milk. Discs impregnated with 15 μL of each bacterial culture were placed on the agar, then incubated at 30 °C and 37 °C for 24 h. The diameters of the hydrolysis zones were measured to classify proteolytic activity: very high for a halo greater than 10 mm, moderate for a halo of 3 to 10 mm, and low for a halo less than 3 mm [23].
2.6. Evaluation of the Biological Activity of LAB Isolates
2.6.1. Determination of Minimum Inhibition Percentages by Using Microdilution Method
Supernatants of bacterial isolates, diluted to different concentrations, were then deposited in the wells of a microplate, with 100µl of each generated concentration. Each well was inoculated with 106 CFU/ml of a suspension of pathogenic bacteria. The wells in the first row of the same microplate received Mueller Hinton medium inoculated with the same concentration of bacteria and without supernatant. The prepared microplates underwent to 21 hours of incubation at 37°C. After incubation, 20 μl of 2,3,5 triphenyltetrazolium was added to observe the bacterial proliferation.The minimum percentage of inhibition (I%) was determined from the first well without bacterial growth [24].
2.6.2. Assessment of Anti-Inflammatory Property
The anti-inflammatory property of the LAB isolates was carried out according to Kar et al. [25]. Overnight cultures of each bacterial isolate were prepared. After centrifugation at 6000 rpm for 10 min at 4°C, the bacterial cell pellets were subjected to three successive washes with 500μl of sterile PBS (20mM, pH=7.4). The latter were reconstituted in 500 μl of the same buffer. The total number of cells was adjusted to 109 CFU/ml (OD≈1.2) [26]. The capacity to inhibit protein denaturation was carried out by preparing three solutions: Test solution 1: composed of 450μl of BSA (at 5% v/v) and 50μl of bacterial suspension; Solution 2: composed of 450μl of BSA and 50μl of bidistilled water as a control; Solution 3: includes 450μl of BSA and 50μl of sodium diclofenac (100 mg/mL). After adjusting each solution to pH 6.3, they were incubated for 20 minutes at 37°C and then subjected to 75°C for 3 minutes. The absorbance was determined after cooling process, and the formula (3) was used to evaluate the rate of protein denaturation inhibition.
I% = 100 × [1- (A2 /A1)],
Where A1 is absorbance test solution and A2 is absorbance of positive control
2.6.3. Evaluation of Ferric Reducing Antioxidant Power (FRAP)
This test was decribed by Benzie and Strain [27], uses solution prepared from acetate buffer at pH 3.6, 10 mM 2,4,6 -Tris(2-pyridyl)-s-triazine, and 20 mM FeCl3, all was subjected to 37°C of incubation. After that, a volume of 100 μL of each solution was adjusted to 900 µL of the FRAP solution and kept at room temperature for 30 min of incubation.The concentration of ferric-reducing power for each bacterial suspension was calculated using a calibration curve performed with FeSO4.
2.6.4. Evaluation of 2,2-diphényl-1-picrylhydrazyl (DPPH•) Scavenging
This method is evaluated according to the protocol described by Sanchez-Moreno et al. [28]. A volume of 750 μl of each bacterial suspension was added to 750 μL of the methanolic DPPH• solution at 4 mg/mL. The reaction mixture was incubated at room temperature and in the dark for 50 min, then the absorbance was measured at 517 nm. The ability of the bacterial isolates to scavenge DPPH• was expressed as a percentage of inhibition, calculated using the following equation (4):
where A1 : absorbance of the control; A2 : absorbance of the sample.
% inhibition of the DPPH• radical = [(A1-A2) / A1] × 100,
2.6.5. Lipid Peroxidation Evaluation
The evaluation of lipid peroxidation was performed by the protocol of Bekkouche et al. [29]. 160 μL of bacterial suspensions, 40 μL of copper sulfate solution (CuSO4 at 0.33 mg/mL), and 160 μL of human plasma were combined and underwent to 50 °C of incubation for 12 hours. Two controls were used: negative control (160 μL of plasma + 160 μL of distilled water) and a positive control (160 μL of plasma + 160 μL of distilled water + 40 μL of CuSO4). After 12 h of incubation, 200 mL of the reaction solution was added to 800 μL of a thiobarbituric acid mixture (0.375% (w/v)), trichloroacetic acid (20%), 2,6-di-tert-butyl-4-methylphenol (0.01%), and HCl (1N) (20%) and subjected to 15 minutes of incubation at 100°C. After cooling, 2 mL of butanol-1 was used to extract the complex that had formed. After centrifugation at 4000 rpm during 10 minutes, the absorbance was read at 532 nm. Malondialdehyde (MDA) concentration was determined utilizing 1,3,3-tetraethoxypropane curve , and MDA inhibition rate was evaluated using the equation (5):
MDA (%) = [(A0-A1) /A0]x 100,
Where A0 : MDA concentration of the positive control; A1 : MDA concentration of the sample.
2.6.6. Determination of Reduced Glutathione (GSH) Levels
Preparation of intracellular extracts of LAB isolates
According to Su et al. [26] modified, overnight cultures of each bacterial isolate were centrifuged at 6000 rpm for 10 minutes at 4 °C, the cells were rinsed three times with sterile PBS, then adjusted to 10⁸ CFU/ml. Cell suspensions were subjected to ultrasonic disruption. Sonication was conducted for five intervals of one minute each in an ice bath. Cell debris was eliminated using centrifugation at 10,000 rpm for 20 minutes at 4°C, and the resultant intracellular extracts were utilized to quantify total proteins and GSH.
GSH Assay
500 μL of intracellular extract from bacterial isolates were mixed to 750 μL of PBS (0.05N, pH 8) and 250 μL of Ellman's reagent. The mixture was kept 15 minutes at room temperature. The optical density reading was performed at 412 nm. The concentration of GSH was determined using a calibration curve of reduced GSH.
2.7. Statistical Analysis
The statistical analysis was performed using one-way analysis of variance (ANOVA) to compare the means followed by the Tukey test (p<0.05) using 8 version of the Statistica software, published by Statsoft in Tulsa, Oklahoma. All data are expressed as means ± standard error of the mean (SE).
3. Results
3.1. Identification of Bacterial Isolates
3.1.1. Morphological, Biochemical, and Physiological Characteristics
The LAB isolates exhibited morphological diversity. The colonies of isolates LB1 and LB2 were small, round, smooth, and white, whereas LB3 was medium-sized, smooth, and white. At the microscopic examination, the three strains were Gram-positive: LB1 and LB2 were cocci (isolated or in pairs), while LB3 was rod shape (isolated or in clusters). The results of the biochemical characteristics indicated that these bacteria were catalase negative. The strains LB1 and LB2 hydrolyzed arginine (ADH+), unlike the strain LB3 which, did not hydrolyze arginine. Regarding fermentation, LB1 and LB2 were heterofermentative, while LB3 was homofermentative. Physiological identification revealed that the three strains showed growth at the tested temperature, they were all capable of growing in NaCl concentrations 6.5% (Table 1).
3.1.2. Molecular Identification
The sequence similarity results revealed a 99% homology with the sequences of Weissella confusa (isolates LB1 and LB2), while isolate LB3 belongs to Lactiplantibacillus plantarum (isolate LB3), the phylogenetic tree is shown in Figure 1.
3.2. Tolerance to Acidic pH and Bile Salts
The results revealed that the isolates LB1 and LB3 exhibited better tolerance, with high survival rates estimated at 54% and 52.93±0.003%, respectively, at pH 3 for 3 h, compared to strain LB2 and normal conditions (pH 6.5). Furthermore, the results regarding bile salt resistance (0.3%) demonstrated that all strains obtained from fermented wheat revealed a high ability to persist 4h in the presence of 0.3% bile salts, indicating survival rates of 89%, 76%, and 98%, respectively (Table 2).
2.3. Antibiotic Sensitivity
The findings indicated that the LAB isolates resisted metronidazole, ceftazidime, and streptomycin, with an absence of inhibition zones. On the other hand, they did not show resistance to the antibiotic chloramphenicol, with inhibition diameters greater than 15mm (Table 3).
2.4. Co-Aggregation and Auto-Aggregation
The findings revealed that all bacterial strains exhibited a strong auto-aggregation capacity, with rates varying between 64.5±0.76 % and 73.83±0.33%. However, the co-aggregation results mentioned in Table 4 indicated that strain LB3 exhibited a high co-aggregation potential with E. coli, evaluated at 27.85±0.41%. While strains LB1 and LB2 had lower co-aggregation ability, at 6.78±0.16 % and 6.19±0.6% respectively. As for co-aggregation with S. aureus, all selected strains LB1, LB2, and LB3 demonstrated a considerable capacity ranging from 50.2±0.55 to 60 ± 1.2%.
2.5. Hemolytic Activity
The results of the hemolytic activity indicated that no hemolytic activity was detected. No zone of hemolysis was observed around the colonies, indicating that all the isolates are characterized by a γ-hemolysis type.
2.6. Proteolytic Activity
The results indicated that all the strains showed strong proteolytic activity on MRS agar supplemented with 10% skim milk at a temperature of 30 and 37°C with values ranging between 19.3±0.3 mm and 23.5 ± 1.42 mm (Table 5).
2.7. Determination of Minimum Inhibition Percentages (MIP)
The results of the MIP evaluation by microdilution showed that each strain had a different spectrum of antibacterial activity. The supernatant of the LB3 strain revealed the strongest inhibitory effect against B. cereus (MIP of 5%), followed by B.subtilis, E.coli, and P. aeruginosa (MPI of 10%). In contrast, the supernatants of isolates LB1 and LB2 recorded MIP between 30 and 40% (Figure 2).
2.8. Anti-Inflammatory Activity
The results revealed that strain LB3 was the most active with an inhibition rate of 40%. While the LB1 and LB2 isolates showed inhibitions of 25% and 33%, respectively. However, this capacity remains significantly lower than that observed with the Diclofenac standard (Figure 3).
2.9. FRAP
The results indicated that the LB3 isolate exhibited antioxidant activity, reflected by a higher and significant ferric-reducing power with a concentration of 311.99 ± 1.55 μmol/L. This value is higher than that of the other bacterial isolates, which range between 146.97 ± 1.25 and 285.18 ± 1.58 μmol/L (Table 6).
2.10. DPPH•
The results of the DPPH• radical inhibition percentages showed that all strains had high and varied inhibition rates, ranging from 80.58 ± 0.15% to 86.75 ± 0.65%, indicating a strong antioxidant capacity. However, strain LB3 had an inhibition rate (80.58 ± 0.15%) slightly lower than those of isolates LB1 and LB2, which exhibited nearly identical and the highest inhibitions (Table 6).
2.11. GSH Concentrations
The concentrations of GSH in the three intracellular extracts were very high and similar, with rates of 1.02 ± 0.03, 1.08 ± 0.05, and 1.12 ± 0.04 μmol/mg of protein, respectively (Table 6).
2.12. Lipid Peroxidation Inhibition
From these results, it appears that the three bacterial strains (LB1, LB2, and LB3) possessed a protective antioxidant capacity against the plasma lipid peroxidation ,with values ranging between 27.83 ±0.0025% and 39 ± 0.002%. We observed that strain LB1 was the most active strain with a percentage of around 39.18 ±0.002%, almost identical to that of strain LB3 (Table 6).
4. Discussion
This study aims to identify phenotypic and genotypic characteristics of certain lactic acid bacteria isolated from traditional Algerian fermented wheat and to investigate their probiotic and biological properties. The obtained results regarding the phenotypic characteristics of the bacterial isolates coded LB1, LB2 confirm the phenotypic identification characteristics of Weissella, and for the isolate coded LB3, affirm its belonging to the genus Lactobacillus. These are identical to the main identification characteristics presented by Kandler and Weiss [30]. Moreover, the results of 16S rRNA gene sequencing confirmed that these bacteria belong to the two genera, namely Weissella and Lactobacillus. The genera we identified in our samples corroborate with those of Tahlaïti et al. [31], as well as Chadli et al. [32], who studied the molecular identification of LAB derived from Algerian Hamoum. These authors found that most of the isolates belong to, Leuconostoc Pediococcus, Lactobacillus, and Enterococcus.
The resistance of bacterial isolates to acidic pH can be attributed to their ability to produce acids and other metabolites that stabilize their environment. This tolerance is reinforced by internal pH regulation mechanisms, such as proton exchange, allowing lactobacilli to survive in an acidic environment [33]. These findings are consistent with previous studies, which have demonstrated that Lb. plantarum, Weissella confusa, and Weissella cibaria have high survival rate in acidic environment [31,34]. We also noticed the tolerance of these bacteria to bile salts (0.3%). These results corroborate with those obtained by Angelescu et al. [35] and Chadli et al. [32], who observed strong survival rates of LAB in the presence of bile salts.
The high auto-aggregation of the three strains can be attributed to several factors, including the production of polysaccharides, electrostatic interactions between surface charges, as well as Van der Waals forces and hydrophobic bonds [36]. Anandharaj et al. [17] reported that Lactobacillus and Weissella sp. exhibit auto-aggregation rates between 18 and 79%. Similarly, the significant co-aggregation capacity between probiotic bacteria and Gram-positive pathogens is explained by their morphological similarity, particularly the presence of peptidoglycan and their hydrophobic nature [37]. This co-aggregation allows probiotics to release antimicrobial agents near pathogens, creating a hostile environment that inhibits their growth [33]. Furthermore, the results of the proteolytic activity corroborate with those obtained by N’tcha et al. [38], who revealed that Lb. fermentum, Lb. casei, Leuconostoc mesenteroides, and Streptococcus thermophilus exhibit proteolytic activity, with proteolysis diameters ranging between 24 ± 5.30 mm and 27.5 ± 10.61 mm.
On the other hand, the findings of the antibacterial property of the LAB isolates are in agreement with those of Tahlaiti et al. [31], who revealed that the lactic strains Lb. plantarum (M6, R27), Lb. brevis (BL8), and Pediococcus acidilactici (M54) from Algerian fermented wheat (Hamoum) exhibit high efficacy effect on E. coli, P. aeruginosa and S.aureus. This result could be attributed to the synthesis of antibacterial metabolites particularly bacteriocins ,organic acids and hydrogen peroxide [39] as well as exopolysaccharides (EPS) [40]. Also, it is observed that protein denaturation, related to inflammation, is influenced by lactic acid bacteria, whose protective capacity varies according to the strains and the bioactive molecules produced. Studies, such as those by Khan et al. [40] and Jain and Mahta [41], showed that strains such as Lb. agilis, Lb. casei, and Enterococcus faecium inhibit the denaturation of bovine serum albumin, with significant anti-inflammatory effects.
At the same time, the results of the antioxidant activity reveal that the LAB isolates exhibit significant antioxidant activity. This is reflected by a strong ability to scavenge the DPPH° radical, a power to reduce iron, an inhibition of lipid peroxidation, and a capacity to produce GSH. Indeed, these isolates exhibit a DPPH° scavenging capacity comparable to that reported by Riane et al. [42] for lactic strains isolated from fermented milk (72% to 83%). Our results are also similar to those of Duz et al. [43], these authors observed a maximum activity of 90.34 ± 0.40% for the Lb. plantarum IH14L strain. Previous studies have notably shown that the antioxidant activity of certain Lactobacillus species (Lb. rhamnosus, Lb. helveticus, Lb. sakei, and Lb. plantarum) is linked to the production of EPS on the cell surface [44]. EPS could trap free radicals by releasing active hydrogen or by combining with them to form stable compounds, and they also possess iron-reducing capacity [45]. At the same time, the results of plasma lipid peroxidation inhibition are similar to those of Lin and Chang [46], who exhibited that Bifidobacterium longum and Lb. acidophilus inhibit the plasma lipid peroxidation with rates ranging from 11 to 29%. Another study conducted by Zhang et al. [47] revealed that lactic acid bacteria exert an antioxidant activity against lipid oxidation by inducing the expression of antioxidant genes using adaptive mechanisms, particularly by chelating transition metals, which promote the formation of free radicals and lipid peroxidation. EPS are also involved in inhibiting the formation of MDA. According to the research done by Li et al. [46], the EPS of Lb. plantarum LP6 have the ability to enhance antioxidant enzymatic activities, maintain cellular integrity, and inhibit lipid oxidation of PC12 cells exposed to H2O2. Also, the inhibitory effect on MDA formation by lactic acid bacteria related to particular constituents to each bacterium, such as GSH, antioxidant enzymes, vitamins, amino acids, etc., and the different associated redox reactions [48].
5. Conclusions
This research suggests that the lactic acid strains identified as Weissella confusa and Lactiplantibacillus plantarum, obtained from traditional Algerian fermented wheat, exhibit considerable probiotic potential due to their technological and functional properties. These strains demonstrate a high tolerance to 0.3% of bile salts and in acidic environment, as well as a remarkable ability to aggregate and co-aggregate, and possess antibacterial action against pathogenic microorganisms. The latter possess antioxidant and anti-inflammatory activities, which could be exploited for therapeutic or nutraceutical purposes. Whether to serve as a starter culture or for the creation of novel fermented foods with beneficial properties.
Author Contributions
Conceptualization, R.B. and R.B.; methodology, R.B.; software, D.B.; validation, R.B., H.C. and C.A.; formal analysis, C.F.; investigation, D.B.,F.B. and C.F; resources, R.B. and C.A.; data curation, R.B.; writing—original draft preparation, R.B., R.B., D.B., and F.B; writing—review and editing, R.B. and H.C.; visualization, R.B.; supervision, C.A.; project administration, R.B. and R.B.; funding acquisition, R.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Algerian Ministry of Higher Education and Scientific Research.
Data Availability Statement
Not applicable.
Acknowledgments
The authors gratefully acknowledge Professor Cherif Ameur for his invaluable support and for generously allowing them to conduct a part of experiments in his laboratory.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| LAB | Lactic Acid Bacteria |
| CFU | Colony Forming Unit |
| ADH | Arginine Dihydrolase |
| FRAP | Ferric Reducing Antioxidant Power |
| DPPH | 2,2-diphényl-1-picrylhydrazyl |
| GSH | Reduced glutathione |
| MDA | Malondialdehyde |
References
- Delzenne, N. M.; Cani, P. D.; Bibiloni, R.; Knauf, C.; Waget, A.; Neyrinck, A. M.; Burcelin, R. Changes in gut microbiota control metabolic endotoxemia-induced inflammation in high-fat diet–induced obesity and diabetes in mice. Diabetes 2008, 57, 1470–1481. [Google Scholar] [CrossRef]
- Bandopadhyay, P.; Ganguly, D. Gut dysbiosis and metabolic diseases. Prog.Mol.Bio. Trans. Sci. 2022, 191, 153–174. [Google Scholar] [CrossRef]
- FAO /WHO. Probiotics in food. Health and nutritional properties and guidelines for the evaluation. Rome: Food and agriulture organization of the united nation and world health organization FAO food and nutrition paper no.85.
- Sivamaruthi, B. S.; Kesika, P.; Chaiyasut, C. The role of probiotics in colorectal cancer management. J. Evid-Based Complementary Altern. Med. 2020, 12, 2936. [Google Scholar] [CrossRef]
- Yadav, R.; Shukla, P. Probiotics for human health: current progress and applications. Rec. Adv. Appl. Microbiol. 2017, 133–147. [Google Scholar] [CrossRef]
- Zhao, D.; Du, R.; Chen, L.; Pei, F. Regulating gut microbiota by lactic acid bacteria: Effects based on probiotic characteristics and their metabolites. Lausanne: Front. Med. SA 2024, 1–276. [Google Scholar] [CrossRef]
- Brodkorb, A.; Egger, L.; Alminger, M.; Alvito, P.; Assunção, R.; Ballance, S. T.; Bohn, C.; Bourlieu-Lacanal, R.; Boutrou, F.; Carri`ere, A.; Clemente, M.; Corredig, D.; Dupont, C.; Recio, I. INFOGEST static in vitro simulation of gastrointestinal food digestion. Nat. protoc. 2019, 14, 991–1014. [Google Scholar] [CrossRef] [PubMed]
- Ashaolu, T. J. Immune boosting functional foods and their mechanisms: A critical evaluation of probiotics and prebiotics. Biomed. Pharmacother. 2020, 130, 110625. [Google Scholar] [CrossRef]
- Lortal, S.; El Mecherfi, K. E.; Mariotti, F.; Eutamène, H.; Rul, F.; Champomier-Vergès, M.C.; Savary-Auzeloux, I. Aliments fermentés & bénéfices santé : un défi pour la recherche. Cah. Nut. Diét. 2020, 55, 136–148. [Google Scholar]
- Ben Mehel, B. B.; Bousbahi, S.; Gérard, P.; Bousbahi, S. Impact nutritionnel d’un blé fermenté type Hamoum sur la translocation bactérienne intestinale chez le rat malnutri en phase de réalimentation. Nut. Clini. Métabol. 2019, 33, 102. [Google Scholar] [CrossRef]
- Nithya, A.; Misra, S.; Panigrahi, C.; Dalbhagat, C. G.; Mishra, H. N. Probiotic potential of fermented foods and their role in non-communicable diseases management: An understanding through recent clinical evidences. Food Chem. Advanc. 2023, 3, 100381. [Google Scholar] [CrossRef]
- Labioui, H.; Elmoualdi, L.; El Yachioui, M.; Ouhssine, M. Sélection de souches de bactéries lactiques antibactériennes. Bull. Soc.Pharm. Bordeaux, 2005, 144, 237–250. [Google Scholar]
- Delarras, C. Pratique en microbiologie de laboratoire: recherche de bactéries et de levures-moisissures; Lavoisier-Tec & Doc, Paris, 2014; pp.476.
- Badis, A.; Laouabdia-Sellami, N.; Guetarni, D.; Kihal, M.; Ouzrout, R. Caractérisation phénotypique des bactéries lactiques isolées à partir du lait cru de chève de deux populations caprines locales « arabia et kabyle ». Sci. Technol. 2005, 23, 30–37. [Google Scholar]
- Idoui, T.; Boudjerda, J.; Leghouchi, E.; Karam, N. E. Lactic acid bacteria from" Sheep's Dhan", a traditional butter: Isolation, identification and major technological traits. Grasas y aceites 2009, 60, 177–183. [Google Scholar] [CrossRef]
- Lemoine, F.; Correia, D.; Lefort, V.; Doppelt-Azeroual, O.; Mareuil, F.; Cohen-Boulakia, S. .NPhylogeny.fr: New generation phylogenetic services for non-specialists. Nucleic Acids Research 2019, 47, W260–w265. [Google Scholar] [CrossRef]
- Anandharaj, M.; Sivasankari, B.; Santhanakaruppu, R.; Manimaran, M.; Rani, R. P.; Sivakumar, S. Determining the probiotic potential of cholesterol-reducing Lactobacillus and Weissella strains isolated from gherkins (fermented cucumber) and south Indian fermented koozh. Rese. Microbiol. 2015, 166, 428–439. [Google Scholar] [CrossRef]
- Tarique, M.; Abdalla, A.; Masad, R.; Al-Sbiei, A.; Kizhakkayil, J.; Osaili, T.; Olaimat, A.; Liu, S.Q.; Fernandez-Cabezudo, M.; al-Ramadi, B.; Ayyash, M. Potential probiotics and postbiotic characteristics including immunomodulatory effects of lactic acid bacteria isolated from traditional yogurt-like products. Lebensm. Wiss. Technol. 2022, 159. [Google Scholar] [CrossRef]
- Vlková, E.; Rada, V.; Popelářová, P.; Trojanová, I.; Killer, J. Antimicrobial susceptibility of bifidobacteria isolated from gastrointestinal tract of calves. Livest. Sci. 2006, 105, 253–259. [Google Scholar] [CrossRef]
- Abdulla, A.A.; Abed, T.A.; Saeed, A.M. Adhesion, autoaggregation and hydrophobicity of six Lactobacillus strains. Br. Microbiol. Res. J. 2014, 4, 381–391. [Google Scholar] [CrossRef]
- Abushelaibi, A.; Al-Mahadin, S.; El-Tarabily, K.; Shah, N.P.; Ayyash, M. Characterization of potential probiotic lactic acid bacteria isolated from camel milk. LWT – Food. Sci. Technol. 2017, 79, 316–325. [Google Scholar] [CrossRef]
- Argyri, A.A.; Zoumpopoulou, G.; Karatzas, K.-A.G.; Tsakalidou, E.; Nychas, G-J.E.; Panagou, E.Z.; Tassou, C.C. Selection of po-tential probiotic lactic acid bacteria from fermented olives by in vitro tests. Food. Microbiol. 2013, 33, 282–291. [Google Scholar] [CrossRef]
- Alapont, C.; Martínez-Culebras, P. V.; López-Mendoza, M. C. Determination of lipolytic and proteolytic activities of mycoflora isolated from dry-cured teruel ham. J. Food. Sci. Technol. 2015, 52, 5250–5256. [Google Scholar] [CrossRef] [PubMed]
- Chen, C. C.; Lai, C. C.; Huang, H. L.; Huang, W. Y.; Toh, H. S.; Weng, T. C.; Tang, H. J. Antimicrobial activity of Lactobacillus species against carbapenem-resistant Enterobacteriaceae. Front. Microbiol. 2019, 10, 789. [Google Scholar] [CrossRef]
- Kar, B.; Nepal, A.; Kumar, R.B.; Dolai, N.; Bhattacharya, S.; Upal, K. Antioxidant and anti-inflammatory properties hymenodictyonexcelsum bark. Orient. Pharm. Exp. Med. 2013, 13, 103–114. [Google Scholar] [CrossRef]
- Su, J.; Wang, T.; Li, Y.; Li, J.; Zhang, Y.; Wang, Y.; Li, H. Antioxidant properties of wine lactic acid bacteria: Oenococcusoeni. Appl. Microbiol. Biotechnol. 2015, 99, 5189–5202. [Google Scholar] [CrossRef] [PubMed]
- Benzie, I. F. F.; Strain, J. J. The Ferric Reducing Ability of Plasma (FRAP) as a Measure of “Antioxidant Power”: The FRAP Assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef]
- Sánchez-Moreno, C.; Larrauri, J. A.; Saura-Calixto, F. A procedure to measure the antiradical efficiency of polyphenols. J. Sci. Food Agric. 1998, 76, 270–276. [Google Scholar] [CrossRef]
- Bekkouch, O.; Harnafi, M.; Touiss, I.; Khatib, S.; Harnafi, H.; Alem, C.; Amrani, S. In Vitro Antioxidant and In Vivo Lipid-Lowering Properties of Zingiber officinale Crude Aqueous Extract and Methanolic Fraction: A Follow-Up Study. J. Evid. Based Complementary Altern. Med. 2019, 9734390. [Google Scholar] [CrossRef]
- Kandler, O., Weiss, N. (1986). Regular, non-sporing gram-positive rods. In: Bergey’s manual of systematic bacteriology, Vol. 1, edited by Krieg, N. et al. Baltimore, MD: Williams and Wilkins Co, 1208 -1234.
- Tahlaïti, H.; Dalache, F.; Homrani, A.; Nemmiche, S. Caractérisation et criblage du potentiel probiotique de bactéries lactiques isolées à partir de blé " Hamoum " traditionnellement fermenté. J. sud-asiatique Biol. Exp. 2017, 7, 181–190. [Google Scholar]
- Chadli, A.; Benbouziane, B.; Bouderoua, K.; Bentahar, M. C.; Benabdelmoumene, D. Assessment of potential probiotic properties and biotechnological activities of lactobacillus strains isolated from traditional algerian fermented wheat ELHAMOUM. Asi J. D. Food. Res. 2024, 1–7. [Google Scholar] [CrossRef]
- de Souza, B. M. S.; Borgonovi, T. F.; Casarotti, S. N.; Todorov, S. D.; Penna, A. L. B. Lactobacillus casei and Lactobacillus fermentum strains isolated from mozzarellacheese: probiotic potential, safety, acidifying kinetic parameters and viability under gastrointestinal tract conditions. Probiotics Antimicrob. Proteins 2019, 11, 382–396. [Google Scholar] [CrossRef]
- Lakra, A. K.; Domdi, L.; Hanjon, G.; Tilwani, Y. M.; Arul, V. Some probiotic potential of Weissella confusa MD1 and Weissella cibaria MD2 isolated from fermented batter. L.w.t. 2020, 125, 109261. [Google Scholar] [CrossRef]
- Angelescu, I. R.; Zamfir, M.; Stancu, M. M.; Grosu-Tudor, S. S. Identification and probiotic properties of lactobacilli isolated from two different fermented beverages. Ann. Microbiol. 2019, 69, 1557–1565. [Google Scholar] [CrossRef]
- Doghri, I.; Portier, E.; Desriac, F.; Zhao, J. M.; Bazire, A.; Dufour, A.; Lanneluc, I. Anti-biofilm activity of a low weight proteinaceous molecule from the marine bacterium Pseudoalteromonas sp. IIIA004 against marine bacteria and human pathogen biofilms. Microorganisms 2020, 8, 1295. [Google Scholar] [CrossRef] [PubMed]
- Arif, M.; Akteruzzaman, M.; Islam, S. S.; Das, B. C.; Siddique, M. P.; Kabir, S. L. Dietary supplementation of Bacillus-based probiotics on the growth performance, gut morphology, intestinal microbiota and immune response in low biosecurity broiler chickens. Vet. Anim. Sci. 2021, 14, 100216. [Google Scholar] [CrossRef] [PubMed]
- N’tcha, C.; Haziz, S.; Agbobatinkpo, P.; Vieira-Dalodé, G.; Boya, B.; Codjia, J. C.; Baba-Moussa, L. Probiotic properties of lactic acid bacteria isolated from a beninese traditional beer’s ferment. Int. J. Appl. Biol. Pharm. Technol. 2016, 7, 314–330. [Google Scholar]
- Arena, M. P.; Capozzi, V.; Russo, P.; Drider, D.; Spano, G.; Fiocco, D. Immunobiosis and probiosis: antimicrobial activity of lactic acid bacteria with a focus on their antiviral and antifungal properties. Appli. microbiol. biotechnol. 2018, 102, 9949–9958. [Google Scholar] [CrossRef] [PubMed]
- Khan, A. N.; Yasmin, H.; Ghazanfar, S.; Hassan, M. N.; Keyani, R.; Khan, I.; Ahmad, A. Antagonistic, Anti-oxidant, Anti-inflammatory and Anti-diabetic Probiotic Potential of Lactobacillus agilis Isolated from the Rhizosphere of the Medicinal Plants. Saudi J. Biol. Sci. 2021, 28, 6069–6076. [Google Scholar] [CrossRef]
- Jain, N.; Mehta, A. Evaluation of Probiotic Properties and Antimicrobial Activity of Enterococcus faecium BM10 KY788342 and Lactobacillus casei GM10 KY794586. Food Pharma. International 2017, 1, 84–92. [Google Scholar] [CrossRef]
- Riane, K.; Sifour, M.; Ouled-Haddar, H.; Idoui, T.; Bounar, S.; Boussebt, S. Probiotic properties and antioxidant efficiency of Lactobacillus plantarum 15 isolated from milk. J. Microbiol. Biotechnol. Food Sci. 2021, 516–520. [Google Scholar] [CrossRef]
- DÜz, M.; DoĞan, Y. N.; DoĞan, İ. Antioxidant activitiy of Lactobacillus plantarum, Lactobacillus sake and Lactobacillus curvatus strains isolated from fermented Turkish Sucuk. An. Acad. Bras. Ciênc. 2020, 92, e20200105. [Google Scholar] [CrossRef]
- Wang, K.; Niu, M.; Song, D.; Song, X.; Zhao, J.; Wu, Y.; Niu, G. Preparation, partial characterization and biological activity of exopolysaccharides produced from Lactobacillus fermentum S1. J. Biosci. Bioeng. 2020, 129, 206–214. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Ji., J.; Chen, X.; Jiang, M.; Rui, X.; Dong, M. Structural elucidation and antioxidant activities of exopolysaccharides from Lactobacillus helveticus MB2-1. Carbohydr. Polym. 2014, 102, 351–359. [Google Scholar] [CrossRef] [PubMed]
- Lin, M. Y.; Chang, F. J. Antioxidative effect of intestinal bacteria Bifidobacterium longum ATCC 15708 and Lactobacillus acidophilus ATCC 4356. Dig. Dis. Sci. 2000, 45, 1617–1622. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Hu., P.; Lou., L.; Zhan., J.; Fan., M.; Li., D.; Liao, Q. Antioxidant activities of lactic acid bacteria for quality improvement of fermented sausage. J. Food Sci. 2017, 82, 2960–2967. [Google Scholar] [CrossRef]
- Lee, J. Y.; Kang, C. H. Probiotics alleviate oxidative stress in H2O2-exposed hepatocytes and t-BHP-induced C57BL/6 mice. Microorganisms 2022, 10, 234. [Google Scholar] [CrossRef]
Figure 1.
Phylogenetic tree illustrating the relative positions of lactic acid bacteria obtained from traditional algerian fermented wheat. Methanobrevibacter intestini served as an outgroup.
Figure 1.
Phylogenetic tree illustrating the relative positions of lactic acid bacteria obtained from traditional algerian fermented wheat. Methanobrevibacter intestini served as an outgroup.

Figure 2.
Minimum percentage of inhibition (%) of bacterial isolates against pathogenic bacteria.The results are expressed as means ± SE (n=3); letters indicate significantly different between strains (p < 0.05).
Figure 2.
Minimum percentage of inhibition (%) of bacterial isolates against pathogenic bacteria.The results are expressed as means ± SE (n=3); letters indicate significantly different between strains (p < 0.05).
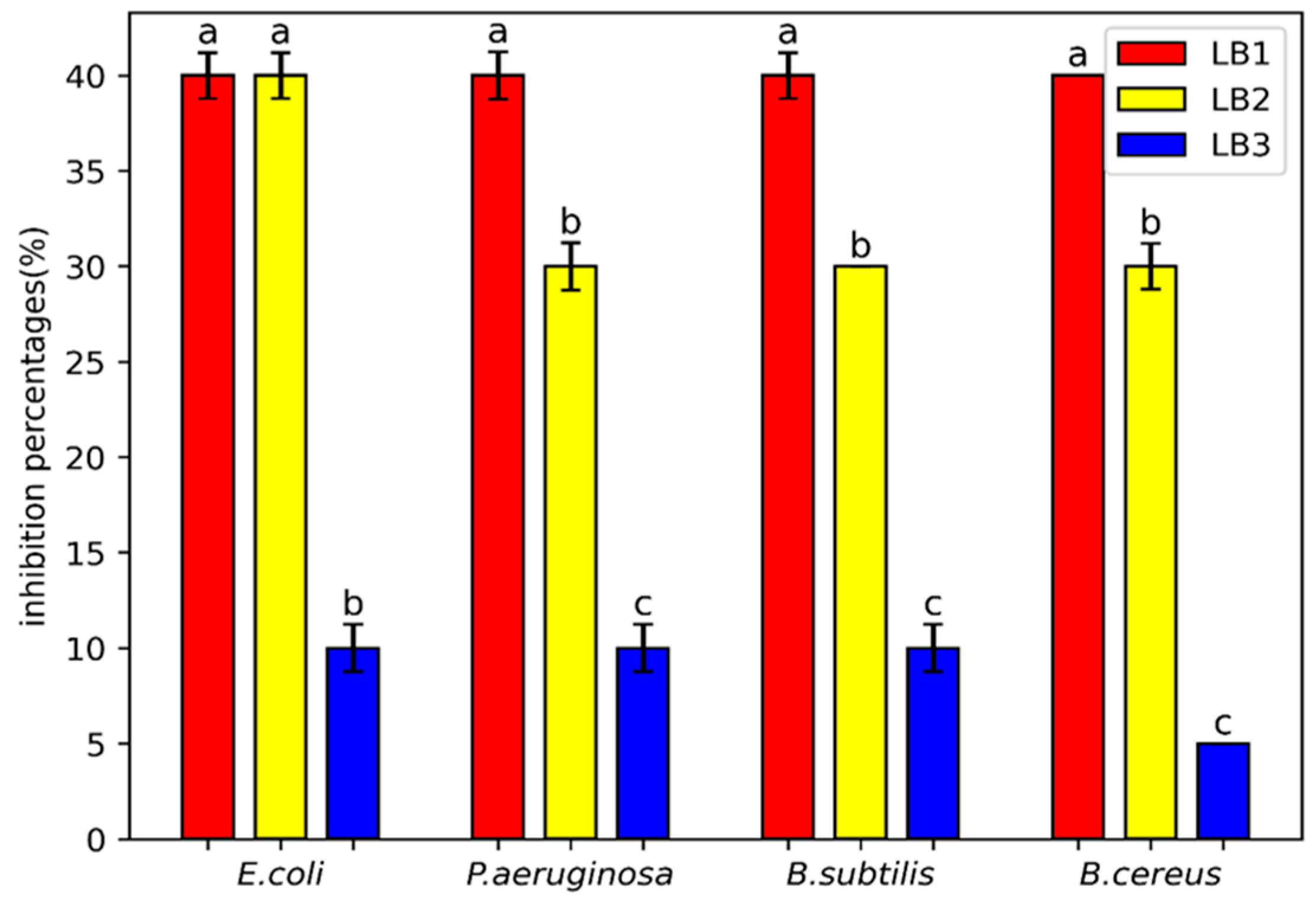
Figure 3.
Anti-inflammatory activity of the bacterial isolates. The results are expressed as means ± SE (n=3); letters indicate significantly different between strains (p < 0.05).
Figure 3.
Anti-inflammatory activity of the bacterial isolates. The results are expressed as means ± SE (n=3); letters indicate significantly different between strains (p < 0.05).
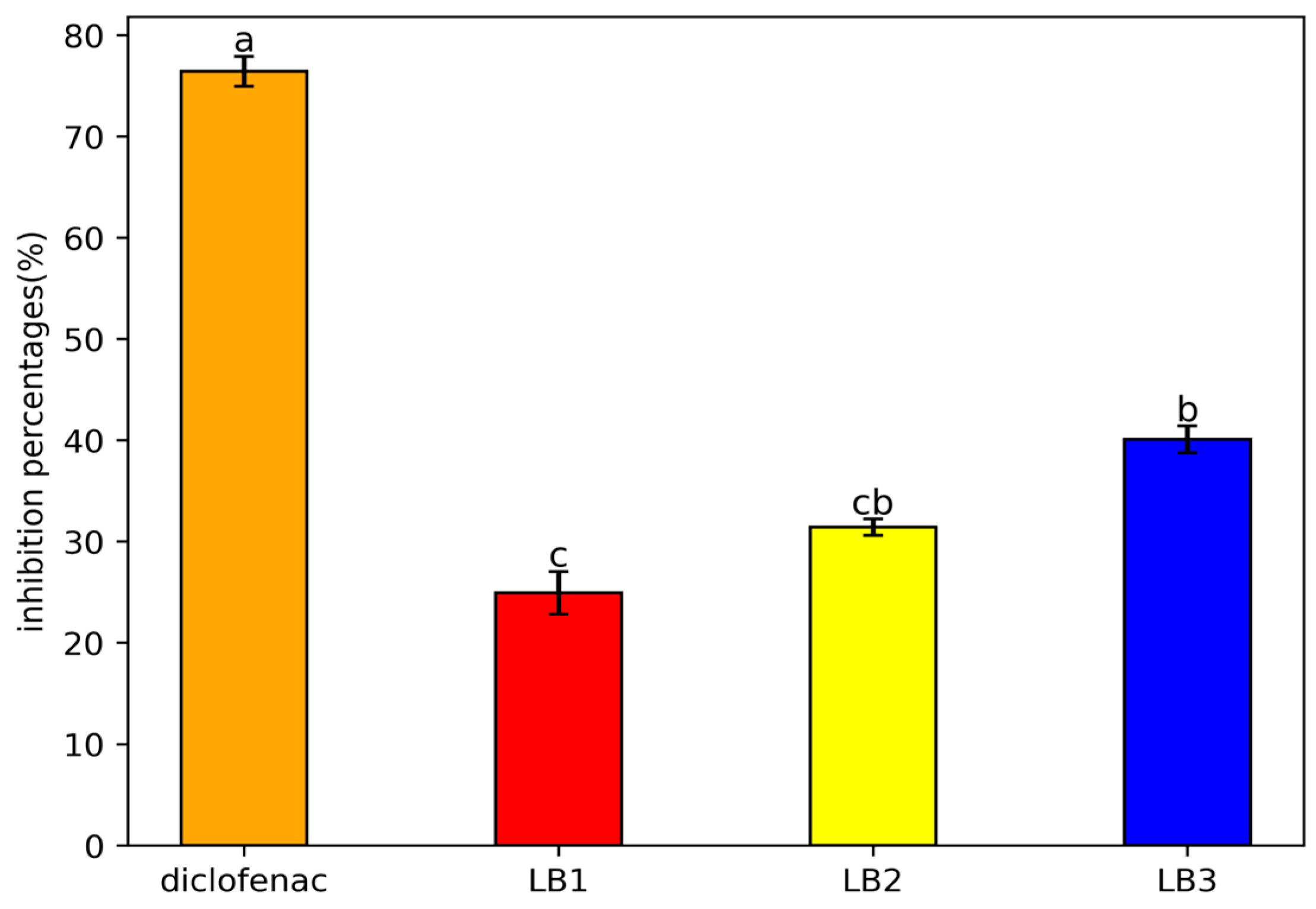

Table 1.
Morphological, biochemical, and physiological characteristics of bacterial isolates from traditional fermented wheat.
Table 1.
Morphological, biochemical, and physiological characteristics of bacterial isolates from traditional fermented wheat.
| Tests | LB1 | LB2 | LB3 |
| Gram | +* | + | + |
| Cell shape | cocci | cocci | rod |
| Catalase | - | - | - |
| Fermentation Type | Heterofermentative | Heterofermentative | Homofermentative |
| Arginine dihydrolase | + | + | - |
| Growth at temperature | |||
| 10°C | + | + | + |
| 30°C | + | + | + |
| 45°C | + | + | + |
| Growth in NaCl (%) | |||
| 6.5 | + | + | + |
| 9.5 | - | - | - |
*(+) positive results. (-) negative results; .
Table 2.
Survival rate of all strains expressed as percentage of viability under acidic pH and bile salts conditions at 37°C.
Table 2.
Survival rate of all strains expressed as percentage of viability under acidic pH and bile salts conditions at 37°C.
| Bacterial strains | pH 3 (3h) | Bile salts 0.3 % (4 h) |
| LB1 | 54.13± 0.001*a | 88.84±1.03b |
| LB2 | 40.23±0.03b | 75.74±0.71c |
| LB3 | 52.93±0.003a | 97.54±0.65a |
The results are expressed as means ± SE (n=3); *letters indicate significantly different between strains (p < 0.05).
Table 3.
Antibiotic susceptibility of selected strains.
| Antibiotics | LB1 | LB2 | LB3 |
| Chloramphenicol (30µg) | I | S | S |
| Colistin (10µg) | I | I | I |
| Metronidazole (6µg) | R* | R | R |
| Streptomycin(10µg) | R | R | R |
| Penicillin(10µg) | S | I | R |
| Ceftazidime (10µg) | R | R | R |
| Gentamicin (10µg) | S | S | I |
*R: Resistant; I: Intermediate; S: Sensitive.
Table 4.
Auto-aggregation and Co-aggregation rates of selected strains.
| Strains | Auto-aggregation % | Co-aggregation % | |
| E.coli | S. aureus | ||
| LB1 | 65.5±0.2b* | 6.78±0.16b | 50.62± 0.36b |
| LB2 | 64.5±0.76b | 6.19±0.6b | 50.2±0.55b |
| LB3 | 73.83±0.33a | 27.85±0.41a | 60 ± 1.3a |
The results are expressed as means ± SE (n=3); *letters indicate significantly different between different strains (p < 0.05).
Table 5.
Diameters (mm) of proteolysis obtained by bacterial strains.
| Strains | T30°C | T37°C |
| LB1 | 23.5 ± 1.42 a* | 0.44 b |
| LB2 | 19.75 ± 0.85 a | 20 ± 0.54b |
| LB3 | 19.5 ± 0.22 a | 19.3 ± 0.2b |
The results are expressed as means ± SE (n=3); *letters indicate significantly different between strains (p < 0.05).
Table 6.
Antioxidative activities of bacterial strains.
| LB1 | LB2 | LB3 | |
| FRAP (µmol/L) | 171.36±17.77b* | 146.97 ±1.25b | 311.99± 1.55a |
| DPPH (%) | 84.45 ± 0.42a | 86.75 ± 0.65a | 80.58 ± 0.15b |
| GSH (µmol/mg) | 1.02±0.03a | 1.08±0.05a | 1.12± 0.04b |
| Lipid peroxidation (%) | 39.18 ±0.002a | 25.63 ±0.003b | 36 ± 0.001a |
The results are expressed as means ± SE (n=3);* letters indicate significantly different between strains (p < 0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



