Submitted:
20 June 2025
Posted:
23 June 2025
You are already at the latest version
Abstract
Climbing vines induce increasing threats to forest growth under favourable environmental changes recently. In mangrove forests, the native vine Derris trifoliata became invasive and is now one of the main threats. Yet current management relies on manual removal with low efficiency. Exploring alternative cost-effective method is required. To assess the potential of a proposed biological control method, this study performed a pot-plant experiment using Cuscuta japonica to infect D. trifoliata and three common mangrove species in Beihai, China. Results showed that D. trifoliata had higher infection rate and high host mortality than mangrove. It also had significantly decreased moisture, nitrogen, phosphorous, potassium and increased soluble sugar and protein, whereas only moisture and one or two minerals of Excoecaria agallocha and Aegiceras corniculatum were influenced. Only Kandelia obovata had neither effective haustoria nor nutrients impact from the infection. This study indicated that C. japonica can do more damages to D. trifoliata than to mangrove species and has the potential to be used as biological control agent to the threatened mangrove forests of A. corniculatum and K. obovata with monitor and control. Further tests in the field are required to bring this method into practice.
Keywords:
mangrove
; holoparasite
; biological control
; nutrients
; Derris trifoliata
; Cuscuta japonica
; Excoecaria agallocha
; Kandelia obovata
; Aegiceras corniculatum
1. Introduction
Due to environmental changes such as annual rainfall and seasonality [1,2], lower salinity [3] and the increase of forest edges and canopy gaps [3,4,5,6], the abundance of liana and climbing vines in tropical and temperate forests are found increasing [2,4,5,7,8]. Such increase results in reduced tree growth, generation, and higher tree mortality [4,7,8,9]. In addition, native vines may become invasive and have a larger effect on forest edges than exotic vines [4,9].
Derris trifoliata (Leguminosae, Papilionoideae), is a common leguminous species widely adapted to fresh water ponds, riverine and saline estuarine wetlands of Asia and parts of Africa [3,10,11,12]. This perennial climbing vine is native to tropical and sub-tropical mangrove forests but has raised concerns for its rapid increase of abundance and tree canopy coverage [3,9,13,14]. It can breed via sexual reproduction and also through intense clonal growth especially at forest edges and in canopy gaps [9,12,15]. In recent years, D. trifoliata has been found causing mangrove density decline and tree death by covering over canopies and competing sunlight, moisture and nutrients [3,9,12,13]. D. trifoliata was identified as highly invasive plant in Bangladesh, densely covering 83% trees of the mangrove threatened by invasive plants [9]. In Indonesia, D. trifoliata gradually increased while mangroves declined from 1990 to 2020 in Segara Anakan Lagoon [3]. In China, the early notice of D. trifoliata threatening mangroves was in 2005 [16]. The process of D. trifoliata from occurring to covering mangrove trees to death took four to six years [17]. Although mangrove forest could expand into the adjacent mud flat, the speed of mangrove forest growth was still lower than that of D. trifoliata invasion during 2013-2022 [17]. This native species is now one of the main threats to Asian mangrove forests, especially to the common tree species in estuarine habitats [3,9,14].
Thus D. trifoliata management is required to control and prevent further spreading in order to protect the ecologically important mangrove forests [9,16,17]. A few methods were suggested include herbicide, manual cutting and biological control [7,16,18]. These methods had been widely discussed and tested in China with some experiences accumulated. Herbicide was usually not considered because of the potential water pollution to tidal zones [19,20]. Manual removal was the most commonly used method in practice by cutting down stems or pulling out root manually [7,18,19,20]. It was recommended to hang the roots up in trees for drying out and preventing sprout tillers, instead of leaving roots in soil or water [20]. However, manual removal costs lots of money, labour and time with low efficiency [7,18]. Another proposed method is wild population exploitation to extract chemicals and produce biological pesticide or medicine [16,19,20]. Yet the practice of this method is not found. Only crude ideas about biological control were given, such as natural enemy [16], allelopathy and alternative plant species [18]. Further study in searching for biological control agents is required [20].
Native parasitic plants can suppress invasive species and it is encouraged to explore the potential of being biological control agents in cost-effective invasion management [21]. In central and eastern European grasslands, root hemiparasitic Rhinanthus alectorolophus suppressed native invasive species Calamagrostis epigejos by disrupting its clonal spread, and it was officially used in C. epigejos-infected grasslands ecological restoration in the Czech Republic [21]. There are more experiments found native parasitic plants have greater impacts on invasive hosts than on other native plants, for instance, hemiparasitic vine Cassytha pubescens on exotic invader Ulex europaeus in Australia, root hemiparasite Pedicularis palustris on native invasive Phragmites australis in Europe, and holoparasitic Cuscuta species on several invasive weeds and vine in China [21,22,23].
Studies of the generalist parasitic Cuscuta species shed light on the searching of biological control agents for D. trifoliata management because of their habit of investing on most rewarding hosts, which are usually fast-growing [24,25]. Because genus Cuscuta is a group of ephemeral holoparasitic vines with no leaves and roots, they live completely on host resources via haustoria on stems [26]. In addition, there is a large number of species in this group hence it is probable to find some in the same area of D. trifoliata [26]. In China, there were several Cuscuta species can suppress invasive hosts while benefit native communities. Cuscuta australis can widely inhibits the growth of Ipomoea cairica, Mikania micrantha, Wedelia trilobata [27], Xanthium spinosum [22], young Bidens pilosa [28] and Humulus scandens mono-cultures or dominated plant community [29]. Cuscuta chinensis caused more damage to exotic invasive hosts than to native non-invasive hosts [30]. Cuscuta campestris restrained the growth of invasive M. micrantha and Solanum rostratum [31,32]. Cuscuta reflexa can also suppress M. micrantha [33]. C. japonica produced allelo-chemicals against M. micrantha, acted as strong carbon sink for Ambrosia trifida, severely reduced the growth of Solidago canadensis and could biologically control these invasive weeds [34,35,36]. Therefore, it worths to explore the effects of Cuscuta species on the invasive D. trifoliata.
This case study in Beihai China is motivated by the potential of developing a biological control method using Cuscuta species for D. trifoliata management. Beihai is located in the Beibu Gulf in the southwest of China. Common mangrove species in the estuarine habitats include the native Aegiceras corniculatum, Kandelia obovata, Avicennia marina, Excoecaria agallocha, Bruguiera gymnorhiza, and the exotic Sonneratia apetala and Laguncularia racemosa. In Beihai, D. trifoliata is often found on A. corniculatum and K. obovata, E. agallocha and S. apetala in the field [17,37]. In a pilot study, C. australis and C. japonica, identified based on floral morphology, were collected from field bush on shore next to mangrove forests and fixed to D. trifoliata, but only C. japonica managed to develop haustoria and survive.
In this study, a pot-plant experiment was performed using C. japonica to infect D. trifoliata and three common mangrove species threatened by D. trifoliata in Beihai, China. The aim of this study is to assess the effectiveness and risk of this proposed biological control method in answering two questions:
- (1)
- Does C. japonica successfully infect D. trifoliata and have significant negative effects?
- (2)
- Will C. japonica also infect the mangrove covered by D. trifoliata and cause significant negative effects?
2. Materials and Methods
2.1. Plant Material Preparation
There were four host species in the C. japonica infection experiment, including D. trifoliata, E. agallocha, K. obovata and A. corniculatum. C. japonica was sourced from Beihai Coastal National Wetland Park. According to field observation in 2023 and 2024, the growing season of D. trifoliata was from April to September, and that of E. agallocha was from June to September. The flowering season for D. trifoliata was from April to August, for E. agallocha was in May, June and August, for C. japonica was in October and November. Relatively intensive defoliation of D. trifoliata and E. agallocha was in May and July. D. trifoliata had root nodules for nitrogen fixation [12,15].
In order to minimise the impacts of intra-specific variations in parasite responses [12,23], seedlings of each host species were selected from the same source near Beihai with similar age, height and ground diameter. D. trifoliata seeds were collected in November 2023 from single wild population then cultivated into seedlings each planted in a 7.5 L pot with commercial vegetable soil and fermented goat faecal soil (v/v = 1:1, Dewoduo Fertilizer, Hebei, China). In April 2024, K. obovata (30 cm in height) and A. corniculatum (25 cm in height) were purchased from a local commercial nursery (Bagui Company, Guangxi), while the seedlings of E. agallocha (35 cm in height) were collected from a single location in the Beihai Coastal National Wetland Park. Each mangrove species was transplanted into a plastic box (80 cm × 40 cm × 34 cm) which was filled with source field soil and placed in a larger box (120 cm × 50 cm × 38 cm) with freshwater at a 20 cm depth.
All seedlings were planted on an open roof top for at least four months with sufficient natural sunlight, time irrigation of freshwater, and weekly supply of liquid fertiliser (Dewoduo Fertilizer, Hebei, China; N:P:K = 2:1:1, 1:300 diluted, 150 ml each individual).
2.2. Experimental Design
Individuals of each host species were randomly divided into infection group and control group with the same group size of 30 individuals for D. trifoliata, and 10 individuals for E. agallocha, K. obovata and A. corniculatum. The infection experiment started from August 2024 during the growing season of both C. japonica and the hosts species.
Pre-infection sampling for chemical analysis was performed on the day before infection (Day 0). On the day of infection (Day 1), C. japonica stems with buds (25 cm long) were collected from the field then using thin metal wires to fix one stem to each D. trifoliata and two stems to each mangrove host in the infection groups. Planting conditions remained the same, including the management of irrigation and fertiliser.
Infection groups were observed every 2-3 days for the survival (alive =1, dead = 0) of each C. japonica stem to calculate the parasite survival rate for each host species. Post-infection sampling for chemical analysis and histology was carried out when host leaves turned yellow or when there was sufficient infected host biomass, which was on Day 75 for D. trifoliata, Day 96 for E. agallocha, and Day 112 for K. obovata and A. corniculatum. Because all host species were inactive in winter and started sprouting in spring, C. japonica were allowed to continue growing on hosts after sampling to observe the survival (alive =1, dead = 0) of host species until the end of May 2025.
2.3. Sampling and Analysis
For histology, two to three specimens having haustoria of C. japonica firmly attached to the stem and petiole of four host species were harvested respectively, trimmed into 5-7 mm long pieces and immediately preserved in 50% FAA fixative solution (biosharp® Beijing Labgic Technology Co. Ltd, China) at room temperature for one week then stored at 4 ℃. After haustoria specimens of all host species were collected and fixed, they were sent to a testing company (Wuhan ProNets Testing Technology Co. LTD, Hubei, China) for paraffin sectioning, Safranin-O/fast green staining, and light microscope image capturing.
Pre-infection sampling collected six samples from D. trifoliata and three samples from E. agallocha, K. obovata and A. corniculatum respectively. Each sample of host species contained at least 50 g of leaves and stems. Six samples (about 100 g) of C. japonica were collected from the field source.
For the post-infection chemical analysis sampling, leaves and stems with C. japonica were harvested from all plants of each infection group. C. japonica and the host were carefully separated as soon as possible. Host specimen was mixed within group then allocated 45-50 g to each paper sample bag. It was the same for C. japonica to fill 90-100 g per bag. There were six samples for D. trifoliata and three samples for E. agallocha, K. obovata and A. corniculatum respectively. Since D. trifoliata forms nodules in fixing nitrogen [12], the six samples of C. japonica from D. trifoliata were analysed separately from the six samples from the three mangrove species. For each control group, same number of samples of at least 50 g each were collected.
To determine moisture content, the wet weight of each sample was measured to the nearest 0.01 g (Yingheng electronic balance YH-A50002, Guangdong, China) then inactivated enzymes at 105
℃
for 30 min, and oven-dried at 70
℃
(Taisite drying oven WGL-230B, Tianjin, China) to the consistent dry weight. Moisture content was represented as (weight loss/wet weight) x 100%.
The dry samples were stored in zip-lock bag and sent to a testing company (Wuhan ProNets Testing Technology Co. LTD, Hubei, China) for grinding and analysing total nitrogen content (Kjeldahl method), total phosphorous content (vanadium molybdate blue colorimetric method), total potassium content (flame photometry), soluble sugar content (anthrone method) and soluble protein content (Bradford assay).
2.4. Statistical Analysis
All statistical analysis were performed in SPSSAU online version 25.0 (The SPSSAU project,
https://www.spssau.com
) and α = 0.05. Two-way analysis of variances (ANOVAs) was used to analyse the effects and interactions of infection and host species on host chemical parameters. Independent samples t-tests or non-parametric tests were used to compare the differences of chemicals between infection group and control group of each host species. Welch’s ANOVA was used to compare host and parasite chemical contents between pre-infection and post-infection sampling.
3. Results
3.1. Survival Rates of Cuscuta japonica and Hosts
The observation of the survival of C. japonica stems lasted from Day 1 to Day 38 until all survived stems had successfully established haustoria and started growing in length. There were originally 30 stems on D. trifoliata and 20 stems on E. agallocha, K. obovata, A. corniculatum, and the survival rates on Day 38 were 53%, 30%, 20% and 0% respectively (Figure 1). Parasite survival rate of A. corniculatum had already reached 0% on Day 21. Stems on D. trifoliata had the highest survival rate throughout the records. Because C. japonica stems did not survive or grow well on A. corniculatum and K. obovata to provide sufficient post-infection samples, more stems were fixed after Day 38 to force infection establishment.
After post-infection sampling, the infection group of D. trifoliata were completely defoliated on Day 101 then C. japonica died afterwards, but three D. trifoliata individuals were sprouting in 2025. Leaves of
E. agallocha
continued yellowing after Day 96 and all individuals had new leaves back in 2025, while C. japonica increased shortly then mostly dried out in winter and remained several stems in May 2025. All individuals of
K. obovata
were green throughout the whole time and C. japonica only had several stems on since infection establishment. The infection on
A. corniculatum remained a low level but it rapidly increased from April 2025 causing leaf senescence at the end of May. In summary, by the end of May 2025, the survival rate of D. trifoliata was 10% while the other three host species had 100% survived from the infection experiment in 2024.
3.2. Haustoria on Different Host Species
Images of sectioned haustoria from each host species are presented (Figure 2). They included the haustoria on both stems and petioles of host species, except for the petiole of A. corniculatum which was too short to bear a haustorium. Images showed that C. japonica managed to reach the host vascular bundles of D. trifoliata (Figure 2a, b), E. agallocha (Figure 2c, d) and A. corniculatum (Figure 2g). In K. obovata, parasite was able to penetrate host tissues but showed no signs of invading the host vascular structures (Figure 2e, f). It is noticed that the structure of vascular bundle in the petiole of D. trifoliata was similar to stem structure with a single large bundle instead of several small vascular bundles like the petioles of E. agallocha and K. obovata (Figure 2a, c, e).
3.3. Infection Responses of Host Species
In the results of two-way ANOVA of post-infection chemicals, there were species × infection interactions for total nitrogen content, total phosphorous content, total potassium content and soluble sugar content, which indicated that these chemicals in different host species were affected by infection in different ways (Table 1). Moisture content was independently affected by host species and infection without interaction while soluble protein content only had significant variations among host species (Table 1).
Among the
four
host species, all chemicals of D. trifoliata were significantly influenced by infection in t-tests, with decreases in moisture content by 4%, total nitrogen content by 14%, total phosphorous content by 27% and total potassium content by 49%, but increases in soluble sugar content by 49% and soluble protein content by 20% (
Figure 3
;
Table S2
). In contrast, all chemicals of
K. obovata
show no significant difference between infection group and control group in non-parametric tests (
Figure 3
;
Table S3
). And
E. agallocha
had moisture content, total nitrogen and phosphorous content decreased by 2%, 13% and 32% respectively (
Figure 3
;
Table S3
). The moisture content and total potassium content of
A. corniculatum
decreased by 2% and 12% (
Figure 3
;
Table S3
).
3.4. Other Special Features in Chemical Analysis
As a leguminous plant, D. trifoliata (n=12) had the highest total nitrogen content than both C. japonica (n=6) and other host species (n=6) in either pre-infection (Welch F4, 12.904=120.601, P<0.0001) or post-infection sampling (Welch F5, 15.306=98.323, P<0.0001) (Figure 4; Table S4). Besides, the total nitrogen content of C. japonica on D. trifoliata (g/100g dry matter) increased significantly (2.23 ± 0.32) while those infecting other host species had significantly lower total nitrogen content (1.07 ± 0.13) compared to its pre-infection level (1.54 ± 0.33; n=6, F2, 15=26.734, P<0.0001) (Figure 4). All chemicals of C. japonica, except total nitrogen content, were higher than those of host species in both pre-infection and post-infection sampling (Figure 3; Table S4).
4. Discussion
In answering the first question of this study, results indicated that C. japonica can successfully infect D. trifoliata and have significant negative effects. For the other research question, results showed that C. japonica can infect different mangrove species to different degrees and cause significant negative effects to a few nutrients of some species.
4.1. Infection and Effects of Cuscuta to Different Host Species
53% of C. japonica stems survived and established haustoria on D. trifoliata, and resulted in a 90% death in 101 days. In contrast, there were less individuals infected and no death in mangrove species. It indicated that C. japonica restrained and caused more damage to native invasive species than to other native species. It is similar to the characteristic of C. australis, C. campestris and C. chinensis that they suppressed exotic invasive plants but benefited native species in previous studies [27,30,31]. Histology of haustoria anatomy of different species explained their different infection responses. The hyphae of C. japonica penetrated into tissues but cannot reach the vascular bundles of K. obovata. That can explain why this species had no significant nutrients variations between control and infection group, because there was no transfer of nutrients in dysfunctional haustoria [38]. All nutrients of the infected D. trifoliata measured in this study were significant different, whereas only moisture and one or two minerals of E. agallocha and A. corniculatum were influenced (Figure 3). Compared to mangrove species, D. trifoliata had thinner cortex in stem and larger vascular bundle in petiole, hence it was easier to be penetrated and infected [24,26].
The moisture content of D. trifoliata measured in this study (around 68.94-70.56%, Table S4) was similar to the result of a previous study in Thailand (67.10% ± 0.02%) [10]. Infection caused significant reduced moisture content of host species except K. obovata, and the decrease in D. trifoliata was more severe than mangrove species. Because C. japonica, as holoparasite, mainly obtains water via xylem from the host [39]. Parasitic plants also keep higher transpiration rate and lower water potential than hosts to form a bidirectional flow bringing in water and nutrients [40]. For the mineral nutrients (total nitrogen, total phosphorous and potassium), C. australis caused a notable decrease in exotic invasive hosts [27], which is similar to the results of the C. japonica on D. trifoliata in this study. Total nitrogen contents of D. trifoliata and E. agallocha decreased by 14% and 13% respectively. It was found that Cuscuta species not only obtained nitrogen mainly from the host phloem, but also developed xylic hyphae (Figure 2d) connecting to host xylem for nitrogen uptake from root [34,39,41]. In this study, although D. trifoliata had the highest total nitrogen content, C. japonica also gained more nitrogen from it than from mangroves. Infection can have additional adverse effect to nodulated plants because the inadequate supply of resource to the root restricted nitrogen fixation [42]. Restricted nitrogen fixation may produce stress to the maintenance of D. trifoliata population [12]. In each host species, the nitrogen of post-infection sampling was higher than that of pre-infection sampling in this study (Figure 4), because host seedlings were allowed to grow with regular nitrogen supply from frequent fertilising.
Although parasite hyphae reached the vascular bundle of host species except K. obovata according to histology, only D. trifoliata had soluble sugar and soluble protein significantly influenced. And interestingly, they were increasing instead of decreasing. It is contrary to the impact of the same parasite to S. canadensis, with a decrease of 44.47% in soluble sugar and 43.45% in soluble protein [36]. However, in another study in India, protein increased dramatically in all five host plants infected by C. reflexa, which was considered as the stimulation of plant defence mechanism [43]. Examples of both increase and decrease soluble protein in different host-parasite researches were given in this study [43]. Therefore, the response of host plant to parasite infection is diverse according to different host species with different abilities and mechanisms against parasite defence. In comparing to the 20% rise of soluble protein, there was a remarkable 49% increase in soluble sugar of the infected D. trifoliata. It may be explained by Cuscuta stimulated sugar accumulation as a result of the gene expression disorders in the galactose metabolism pathway [22]. Also, carbohydrates accumulation may be due to decreased total nitrogen content because less carbohydrates are utilised in nitrogen metabolise [44]. Sugar accumulation plays a signalling function which can induce leaf senescence [45]. It may work together with the Cuscuta driven excessive autophagy [22] to induce the premature plant senescence of D. trifoliata in this study.
In summary, the higher infection rate and mortality, and higher loss of moisture and nutrients in D. trifoliata supports the view that C. japonica can do more damages to this invasive vine than to mangrove species and has the potential to be used as biological control agent for D. trifoliata management.
4.2. Risk Assessment and Control of Cuscuta japonica in Mangroves
Although all mangrove seedlings survived in the experiment, it is aware that C. japonica did manage to infect the three mangrove species to some extent. Considering the influences of host age upon Cuscuta’s performance [28], this study provided young and relatively vulnerable mangrove hosts for C. japonica than in the field and showed the consequences of a worse scenario of the proposed biological control method. Implications from the results are 1) E. agallocha is susceptible to infection and not suitable for the method; 2) long-term infection may affect A. corniculatum hence C. japonica should be eliminated after D. trifoliata wilts; 3) the method may have a low risk to K. obovata and can be used with monitoring.
This experiment used mangrove seedlings and they were eventually infected by C. japonica. However, such infection is less likely to happen in the field because mangrove seedlings experience daily tidal floods in Beihai which is fatal to C. japonica. In field observation, C. japonica did not live below the tidal line in mangrove trees and was decimated by days of heavy rain. The effect of C. japonica on adult mangrove trees may be different to the experiment results from seedlings as tree trunk and branches are lignified with higher resistance to infection [24]. In our infection trial test, C. japonica stems did not survive on adult A. corniculatum and K. obovata in the field. Therefore, C. japonica is less likely to do harm to mangrove seedlings in tidal field, but may infect adult trees which requires further study in the field.
Although parasitic vines are common in shore vegetations adjacent to mangroves in Beihai, to our best knowledge, Cuscuta species has not been discussed in previous mangrove studies hence they are probably alien to mangrove. It is important to assess the risk before introducing biological control agent to the field and controlling method should be prepared beforehand. In previous studies, Cuscuta species can be effectively controlled by spraying glyphosate [46] and 6% Tusite AS [47]. In mangrove forest, there is a potential to spray sea water or saturated salt water instead to reduce noxious chemicals.
4.3. The Potential of Proposed Method and Further Research
Considering the effectiveness and safety of the proposed method, it has the potential to be used on the threatened mangrove forests of A. corniculatum and K. obovata before carefully tested. In Guangxi, K. obovala and A. corniculatum were found severely degraded due to a 13.8% annual increase of D. trifoliata from 2013 to 2022 [17]. This method, if applicable, can not only be used in Beihai, but also in mangroves dominated by these two tree species of other areas. In China, A. corniculatum was the most frequently reported mangrove species threatened by D. trifoliata due to their same preference to hypo-saline estuarine habitats [16,19,20,48]. In an Indonesian lagoon, mangrove tree communities with A. corniculatum, Avicennia alba and Sonneratia caseolaris experienced areas decline and patches fragmentation from D. trifoliata [3].
Apart from the choice and potential of biological control agent, the possible scenario of applying this method is also considered based on the dispersal pattern of D. trifoliata. Using satellite images, two types of areas were identified as in different phrases of D. trifoliata spreading on mangroves [17]. Phrase one was at the beginning of spread, from D. trifoliata being absent to dotted present along mangrove edges and gaps, then spreading out in circles and causing a relatively lower tree loss [17]. Phrase two was that D. trifoliata patches on mangrove forests rapidly expanded over mangrove canopy which induced high tree loss each year [17]. It was suggested that these two types should be managed in different ways [17]. Manual clear is more suitable to tackle the phrase one scenario because mangrove edges or gaps may be accessible to human by foot or by boat. For the condition of phrase two, mangrove canopy is not accessible to human and it needs remotely applied methods. In this situation, biological control using C. japonica can be a choice, for it is possible to place parasite stems, monitor, spray and control Cuscuta remotely using small unmanned aircraft.
This study used only fresh water in the planting system and may not reflect the parasite effects under the field condition with salinity around 8-12 ‰ [49]. The host biomass and nutrition availability to C. japonica may change along salinity and host species, and alter the host-parasite interactions [40]. In addition, other important traits of plant growth and some chemical signals in responding parasitic infection can be included in future study, such as total biomass, root biomass and nodules, proline, abscisic acid and mannitol [23,36,40]. Moreover, host size and host age can influence parasite effect and should be considered [23,28]. Therefore, future experiments on infecting adult hosts in saline environment are important in unfolding further understanding in how C. japonica can be used in biological control of D. trifoliata.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: Two-way ANOVA results for the effects of Cuscuta japonica infection and host species on moisture content (%), total nitrogen content (g/100g), total phosphorous content (g/100g), total potassium content (g/kg), soluble sugar content (mg/g) and soluble protein content (mg/g). Significant results are in bold; Table S2 Results of independent samples t-tests for post-infection sampled chemicals of Derris trifoliata (n=6): moisture content (%), total nitrogen content (g/100g), total phosphorous content (g/100g), total potassium content (g/kg), soluble sugar content (mg/g) and soluble protein content (mg/g). Significant results are in bold; Table S3 Results of non-parametric tests for post-infection sampled chemicals of a) Excoecaria agallocha (n=3), b) Kandelia obovata (n=3) and c) Aegiceras corniculatum (n=3): moisture content (%), total nitrogen content (g/100g), total phosphorous content (g/100g), total potassium content (g/kg), soluble sugar content (mg/g) and soluble protein content (mg/g). Significant results are in bold; Table S4 Results of Welch’s ANOVA for pre-infection and post-infection sampled (pooled samples of control and infection groups) chemicals of Derris triofiliata (n=12), Excoecaria agallocha (n=6), Kandelia obovata (n=6), Aegiceras corniculatum (n=6), Cuscuta japonica infecting Derris triofiliata (n=6) and other host species (n=6): moisture content (%), total nitrogen content (g/100g), total phosphorous content (g/100g), total potassium content (g/kg), soluble sugar content (mg/g) and soluble protein content (mg/g). Different superscript letters signify significant differences (vertically).
Author Contributions
Conceptualization and methodology, all authors; investigation, H. W. and Y. X.; formal analysis, validation, visualization and writing - original draft preparation, H. W.; writing - review and editing, all authors; project administration, H. W.; funding acquisition, H. W. and W. L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Special Funding for Science and Technology Bases and Talents of Guangxi Province [grant number AD22080040], Research Start-up Fund of Guangxi Mangrove Coastal Wetland Conservation and Sustainable Use for Qualified Scientists [grant number 2022GMRC-04], Guangxi Forestry Scientific Research [grant number 2025KX No.06], the Innovation and Development fund of Guangxi Academy of Sciences [grant number 2024YGFZ504-101], The Basic Scientific Foundation of Guangxi Mangrove Research Center [grant number GMRC-202403]. The APC was funded by Special Funding for Science and Technology Bases and Talents of Guangxi Province [grant number AD22080040].
Data Availability Statement
The datasets presented in this article are not readily available because the data are part of an ongoing study. Requests to access the datasets should be directed to the correspondence.
Acknowledgments
Many thanks to the Beihai Coastal National Wetland Park of Guangxi for permitting and assisting in collecting Excoecaria agallocha seedlings for the experiment.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- DeWalt, S.J.; Schnitzer, S.A.; Chave, J.; Bongers, F.; Burnham, R.J.; Cai, Z.; Chuyong, G.; Clark, D.B.; Ewango, C.E.N.; Gerwing, J.J.; et al. Annual rainfall and seasonality predict pan-tropical patterns of liana density and basal area. Biotropica 2010, 42, 309–317. [Google Scholar] [CrossRef]
- Parolari, A.J.; Paul, K.; Griffing, A.; Condit, R.; Perez, R.; Aguilar, S.; Schnitzer, S.A. Liana abundance and diversity increase with rainfall seasonality along a precipitation gradient in Panama. Ecography 2020, 43, 25–33. [Google Scholar] [CrossRef]
- Prayudha, B.; Siregar, V.; Ulumuddin, Y.I.; Prasetyo, L.B.; Agus, S.B.; Suyadi; Suyarso; Salatalohi, A. ; Anggraini, K. Mangrove forest encroachment by Nypa frutican, Derris trifoliata, and Acanthus spp. in Segara Anakan Lagoon. IOP Conference Series: Earth and Environmental Science 2023, 1251, 012017. [Google Scholar] [CrossRef]
- Matthews, E.R.; Schmit, J.P.; Campbell, J.P. Climbing vines and forest edges affect tree growth and mortality in temperate forests of the U.S. Mid-Atlantic States. For. Ecol. Manag 2016, 374, 166–173. [Google Scholar] [CrossRef]
- Schnitzer, S.A.; DeFilippis, D.M.; Visser, M.; Estrada-Villegas, S.; Rivera-Camaña, R.; Bernal, B.; Peréz, S.; Valdéz, A.; Valdéz, S.; Aguilar, A.; et al. Local canopy disturbance as an explanation for long-term increases in liana abundance. Ecol. Lett. 2021, 24, 2635–2647. [Google Scholar] [CrossRef]
- Campbell, M.J.; Edwards, W.; Magrach, A.; Alamgir, M.; Porolak, G.; Mohandass, D.; Laurance, W.F. Edge disturbance drives liana abundance increase and alteration of liana-host tree interactions in tropical forest fragments. Ecol. Evol. 2018, 8, 4237–4251. [Google Scholar] [CrossRef]
- Paul, G.S.; Yavitt, J.B. Tropical vine growth and the effects on forest succession: a review of the ecology and management of tropical climbing plants. Bot. Rev. 2011, 77, 11–30. [Google Scholar] [CrossRef]
- Becknell, J.; Vargas G, G.; Wright, L.; Woods, N.-F.; Medvigy, D.; Powers, J. Increasing liana abundance and associated reductions in tree growth in secondary seasonally dry tropical forest. Front. For. Glob. Change 2022, 5, 838357. [Google Scholar] [CrossRef]
- Biswas, S.R.; Choudhury, J.K.; Nishat, A.; Rahman, M.M. Do invasive plants threaten the Sundarbans mangrove forest of Bangladesh? For. Ecol. Manag 2007, 245, 1–9. [Google Scholar] [CrossRef]
- Polyium, U.; Thaisong, P.N. Phytochemical and nutritional values of local plants in the Phraek Nam Daeng Community Samut Songkhram Province Thailand. AMM 2018, 879, 101–107. [Google Scholar] [CrossRef]
- Raju, A.J.S.; Kumar, R. Pollination ecology of Derris trifoliata (Fabaceae), a mangrove associate in Coringa Mangrove Forest, Andhra Pradesh, India. J. Threat. Taxa 2016, 8, 8788–8796. [Google Scholar] [CrossRef]
- Zhang, Y.; Xin, K.; Liao, B.; Ai, X.; Sheng, N. The genetic and environmental adaptation of the associated liana species Derris trifoliata Lour. (Leguminosae) in mangroves. Forests 2021, 12, 1375. [Google Scholar] [CrossRef]
- Zhu, Y.; Liu, K.; Liu, L.; Myint, S.W.; Wang, S.; Cao, J.; Wu, Z. Estimating and mapping mangrove biomass dynamic change using WorldView-2 images and digital surface models. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2020, 13, 2123–2134. [Google Scholar] [CrossRef]
- Sheng, N.; Xin, K.; Zhang, C.; Hua, G. Dispersal pattern of Derris trifoliata in mangroves. Journal of Hainan Normal University(Natural Science) 2021, 34. [Google Scholar] [CrossRef]
- Aluri, J.S.R.; Kumar, R.; Chappidi, P. Reproductive biology of mangrove plants Clerodendrum inerme, Derris trifoliata, Suaeda maritima, Suaeda monoica, Suaeda nudiflora. Transylv. Rev. Syst. Ecol. Res. 2016, 18, 31–68. [Google Scholar] [CrossRef]
- Huang, X.; Zhong, C.; Chen, S.; Liu, Y.; Liang, S. Mangrove forest threatened by Derris trifoliata. Wetland Science & Management 2015, 11, 26–29. [Google Scholar] [CrossRef]
- Lyu, T.; Zhong, S.; Jiang, W.; Ling, Z.; Chu, A. Effects of Deris trfoliata on mangrove degradation in the Lianzhou Bay of Guangxi based on China’s high-resolution remote sensing data. Wetland Science 2025, 23, 11–21. [Google Scholar] [CrossRef]
- Zhang, Y.; Liao, B.; Yang, L.; Jiang, Z.; Xin, K. Research summary on native associated liana species Derris trifoliata in mangrove forests. Wetland Science 2022, 20, 421–426. [Google Scholar] [CrossRef]
- Huang, D.; Sun, X.; Guo, X.; Gou, Z. Risk analysis of mangrove associate plant Derris trifoliate in Hainan. Tropical Forestry 2019, 47, 62–65. [Google Scholar] [CrossRef]
- Li, L.; Lin, J. The distribution and preventing strategy of Derris trifoliate in Hainan Dongzhaigang National Nature Reserve. Tropical Forestry 2019, 47, 36–38. [Google Scholar] [CrossRef]
- Těšitel, J.; Cirocco, R.M.; Facelli, J.M.; Watling, J.R. Native parasitic plants: Biological control for plant invasions? Appl. Veg. Sci. 2020, 23, 464–469. [Google Scholar] [CrossRef]
- Wang, Y.; Bu, H.; Gu, X.; Liu, W.; Wang, X. Integrated omic analysis provides insights into how Cuscuta australis inhibits the growth and reproduction of Xanthium spinosum. BMC Plant Biol. 2025, 25, 657. [Google Scholar] [CrossRef] [PubMed]
- Cirocco, R.M.; Facelli, J.M.; Watling, J.R. The impact of a native hemiparasite on a major invasive shrub is affected by host size at time of infection. J. Exp. Bot. 2020, 71, 3725–3734. [Google Scholar] [CrossRef] [PubMed]
- Zan, Q.; Wang, B.; Wang, Y.; Zhang, J.; Liao, W.; Li, M. The harm caused by Mikania micrantha and its control by Cuscuta campestris. Chinese Journal of Plant Ecology 2003, 27, 822–828. [Google Scholar] [CrossRef]
- Koch, A.M.; Binder, C.; Sanders, I.R. Does the generalist parasitic plant Cuscuta campestris selectively forage in heterogeneous plant communities? New Phytol. 2004, 162, 147–155. [Google Scholar] [CrossRef]
- Furuhashi, T.; Katsuhisa, F.; and Weckwerth, W. The parasitic mechanism of the holostemparasitic plant Cuscuta. J. Plant Interact 2011, 6, 207–219. [Google Scholar] [CrossRef]
- Yu, H.; Liu, J.; He, W.-M.; Miao, S.-L.; Dong, M. Cuscuta australis restrains three exotic invasive plants and benefits native species. Biol. Invasions 2011, 13, 747–756. [Google Scholar] [CrossRef]
- Li, J.; Yang, B.; Yan, Q.; Zhang, J.; Yan, M.; Li, M. Effects of a native parasitic plant on an exotic invader decrease with increasing host age. AoBP 2015, 7, plv031. [Google Scholar] [CrossRef]
- Wu, A.P.; Zhong, W.; Yuan, J.R.; Qi, L.Y.; Chen, F.L.; Liang, Y.S.; He, F.F.; Wang, Y.H. The factors affecting a native obligate parasite, Cuscuta australis, in selecting an exotic weed, Humulus scandens, as its host. Sci. Rep. 2019, 9, 511. [Google Scholar] [CrossRef]
- Li, J.; Jin, Z.; Song, W. Do native parasitic plants cause more damage to exotic invasive hosts than native non-invasive hosts? An implication for biocontrol. PloS One 2012, 7, e34577. [Google Scholar] [CrossRef]
- Yu, H.; Yu, F.H.; Miao, S.L.; Dong, M. Holoparasitic Cuscuta campestris suppresses invasive Mikania micrantha and contributes to native community recovery. Biol. Conserv. 2008, 141, 2653–2661. [Google Scholar] [CrossRef]
- Wang, W.B.; Gao, F.F.; Shao, M.N.; Liu, M.C.; Zhai, H.F.; Qu, B.; Feng, Y.L. First record of field dodder (Cuscuta campestris) parasitizing invasive buffalobur (Solanum rostratum). J. Plant Pathol. 2020, 102, 703–707. [Google Scholar] [CrossRef]
- Wang, H.; Ze, S.; Ji, M.; Zhao, N.; Hu, L.; Xie, S. Impacts of different fertilizer processing on Cuscuta Reflexa infecting Mikania micrantha. Jiangsu Agriculture Science 2020, 48, 98–101. [Google Scholar] [CrossRef]
- Wang, W.; Gao, F.; Feng, W.; Wu, Q.; Feng, Y. The native stem holoparasitic Cuscuta japonica suppresses the invasive plant Ambrosia trifida and related mechanisms in different light conditions in northeast China. Front. Plant Sci. 2022, 13. [Google Scholar] [CrossRef]
- Li, Q.; Xiao, H. Study on the allelochemicals of inhibiting Mikania micrantha growth from Cuscuta japonica. Ecology and Environmental Sciences 2012, 21, 434–439. [Google Scholar] [CrossRef]
- Jiang, H.; Fang, F.; Guo, S. Influences of parasitism by Cuscuta japonica plants on eco-physiological characteristics of Solidago canadensis. Acta Ecologica Sinica 2008, 28, 399–406. [Google Scholar] [CrossRef]
- Liu, W.; Li, L.; Xue, Y.; Liang, Q.; Tao, Y.; Wu, H.; Jiang, W. Phenomenon and mechanisms of Sonneratia apetala introduction and spread promoting excessive growth of Derris trifoliata. Forests 2024, 15, 525. [Google Scholar] [CrossRef]
- Facelli, E.; Wynn, N.; Tsang, H.T.; Watling, J.R.; Facelli, J.M. Defence responses of native and invasive plants to the native generalist vine parasite Cassytha pubescens – anatomical and functional studies. Aust. J. Bot. 2020, 68, 300–309. [Google Scholar] [CrossRef]
- Jeschke, W.D.; Räth, N.; Bäumel, P.; Czygan, F.-C.; Proksch, P. Modelling the flow and partitioning of carbon and nitrogen in the holoparasite Cuscuta reflexa Roxb. and its host Lupinus albus L.: I. Methods for estimating net flows. J. Exp. Bot. 1994, 45, 791–800. [Google Scholar] [CrossRef]
- Zagorchev, L.; Stöggl, W.; Teofanova, D.; Li, J.; Kranner, I. Plant parasites under pressure: effects of abiotic stress on the interactions between parasitic plants and their hosts. Int. J. Mol. Sci. 2021, 22, 7418. [Google Scholar] [CrossRef]
- Jhu, M.Y.; Sinha, N.R. Cuscuta species: Model organisms for haustorium development in stem holoparasitic plants. Front. Plant Sci. 2022, 13, 1086384. [Google Scholar] [CrossRef] [PubMed]
- Jeschke, W.D.; Bäumel, P.; Räth, N.; Czygan, F.-C.; Proksch, P. Modelling of the flows and partitioning of carbon and nitrogen in the holoparasite Cuscuta reflexa Roxb. and its host Lupinus albus L.: II. Flows between host and parasite and within the parasitized host. J. Exp. Bot. 1994, 45, 801–812. [Google Scholar] [CrossRef]
- Nikam, S.S.; Pawar, S.B.; Kanade, M.B. Study of Cuscuta reflexa Roxb. with reference to host diverstiy, anatomy and biochemistry. Central European Journal of Experimental Biology 2014, 3, 6–12. [Google Scholar]
- Taiz, L.; Zeiger, E. Plant Physiology, 5th Edition ed.; Sinauer Associates Inc.: Sunderland, 2010. [Google Scholar]
- Wingler, A.; Roitsch, T. Metabolic regulation of leaf senescence: interactions of sugar signalling with biotic and abiotic stress responses. Plant Biol. 2008, 10, 50–62. [Google Scholar] [CrossRef]
- Hock, S.M.; Wiecko, G.; Knezevic, S.Z. Glyphosate dose affected control of field dodder (Cuscuta campestris) in the tropics. Weed Technol. 2008, 22, 151–155. [Google Scholar] [CrossRef]
- Du, X.; Huang, M.; Ma, Y.; Guo, C.; Liang, H.; Tian, H.; Jiang, X.; Nong, G. Occurrence and Growth Dynamics of Semen cuscutae in Gardens of Guangxi and Evaluation of Herbicides for Its Control. Journal of Southern Agriculture 2012, 42, 748–751. [Google Scholar]
- Li, H.; Ouyang, M.; Zeng, T.; Wu, C.; Liu, J.; Liu, Y. The research on the mangrove resources in Dajiaoshan Seaside Park, Nansha, Guangzhou. Journal of Guangdong University of Education 2019, 39, 43–47. [Google Scholar] [CrossRef]
- Sheng, N. Spatial distribution characteristics of Derris trifoliata in mangroves and its influencing factors. Mater Degree, Hainan Normal University, Haikou, China, 21. 20 April.
Figure 1.
Survival rates of Cuscuta japonica stems of Derris trifoliata, Excoecaria agallocha, Kandelia obovata and Aegiceras corniculatum from Day 1 to Day 38 of infection.
Figure 1.
Survival rates of Cuscuta japonica stems of Derris trifoliata, Excoecaria agallocha, Kandelia obovata and Aegiceras corniculatum from Day 1 to Day 38 of infection.
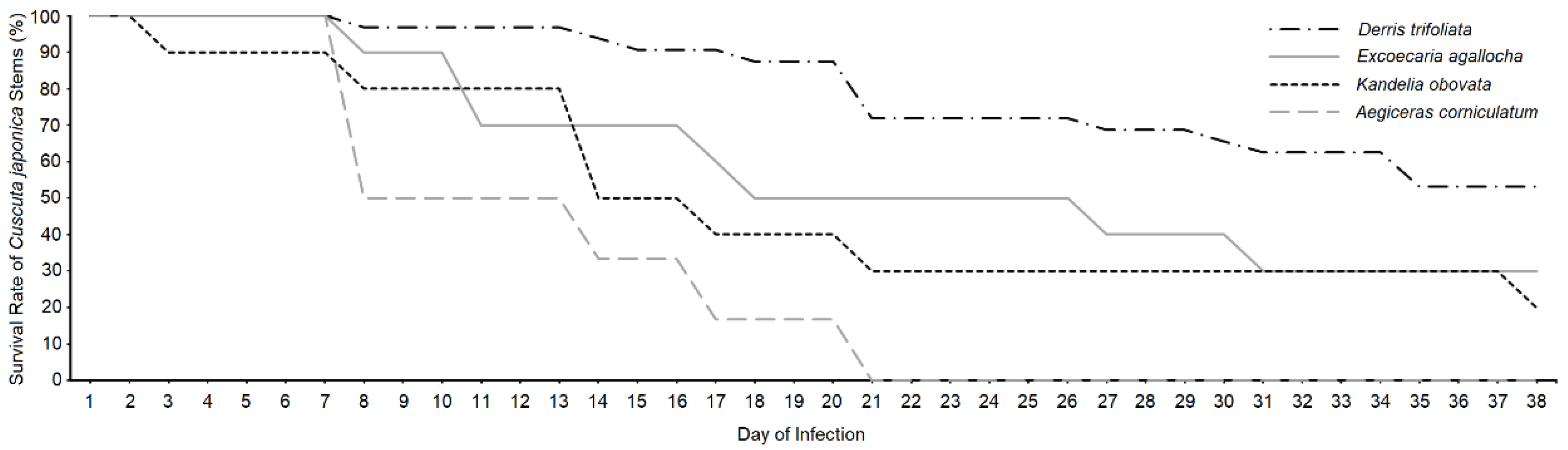
Figure 2.
Safranin-O/fast green staining light microscopy of Cuscuta japonica haustoria on the a) petiole and b) stem of Derris trifoliata, c) petiole and d) stem of Excoecaria agallocha, e) petiole and f) stem of Kandelia obovata, and the g) stem of Aegiceras corniculatum. HP, host pith; HVB, host vascular bundle; PS, parasite stem; PH, parasite haustorium.
Figure 2.
Safranin-O/fast green staining light microscopy of Cuscuta japonica haustoria on the a) petiole and b) stem of Derris trifoliata, c) petiole and d) stem of Excoecaria agallocha, e) petiole and f) stem of Kandelia obovata, and the g) stem of Aegiceras corniculatum. HP, host pith; HVB, host vascular bundle; PS, parasite stem; PH, parasite haustorium.
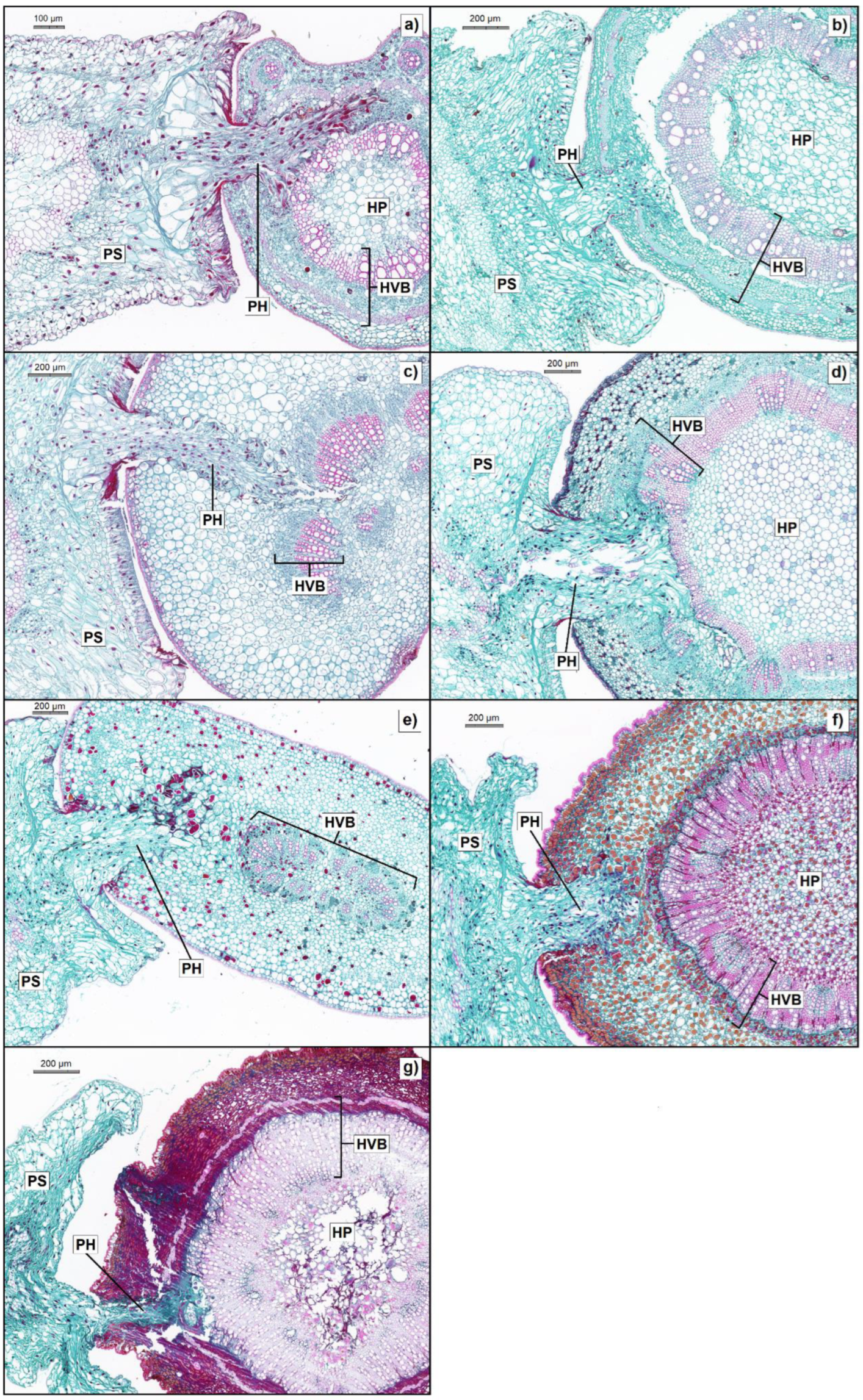
Figure 3.
Results of post-infection sampled (a) moisture content, (b) total nitrogen content, (c) total phosphorous content, (d) total potassium content, (e) soluble sugar content and (f) soluble protein content of Cuscuta japonica (grey boxes, n = 6) and its infected (grey boxes) or uninfected (black boxes) host including Derris trifoliata (n = 6), Excoecaria agallocha (n = 3), Kandelia obovata (n = 3) and Aegiceras corniculatum (n = 3). Asterisks signify significant differences between infection status of a host species.
Figure 3.
Results of post-infection sampled (a) moisture content, (b) total nitrogen content, (c) total phosphorous content, (d) total potassium content, (e) soluble sugar content and (f) soluble protein content of Cuscuta japonica (grey boxes, n = 6) and its infected (grey boxes) or uninfected (black boxes) host including Derris trifoliata (n = 6), Excoecaria agallocha (n = 3), Kandelia obovata (n = 3) and Aegiceras corniculatum (n = 3). Asterisks signify significant differences between infection status of a host species.
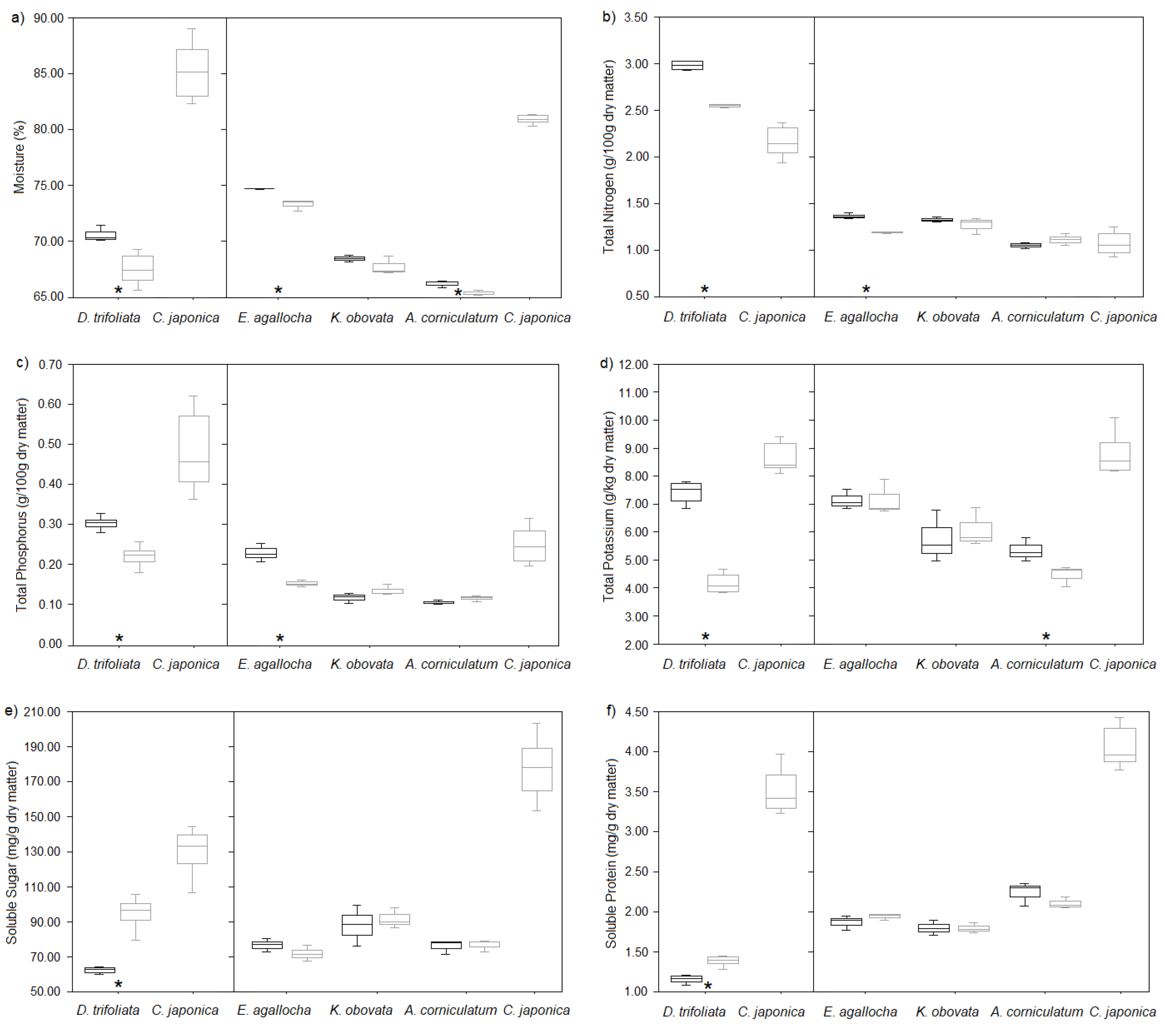
Figure 4.
Total nitrogen content of Cuscuta japonica (CJ, grey dotted bars, n=6) and host species Derris trifoliata (DT, n = 12), Excoecaria agallocha (n = 6), Kandelia obovata (KO, n = 6) and Aegiceras corniculatum (AC, n = 6) in pre-infection (open bars) and post-infection (grey bars) sampling. CJ-DT = C. japonica infecting D. trifoliata; CJ-others = C. japonica infecting other host species. Different letters signify significant differences in each sampling separately.
Figure 4.
Total nitrogen content of Cuscuta japonica (CJ, grey dotted bars, n=6) and host species Derris trifoliata (DT, n = 12), Excoecaria agallocha (n = 6), Kandelia obovata (KO, n = 6) and Aegiceras corniculatum (AC, n = 6) in pre-infection (open bars) and post-infection (grey bars) sampling. CJ-DT = C. japonica infecting D. trifoliata; CJ-others = C. japonica infecting other host species. Different letters signify significant differences in each sampling separately.
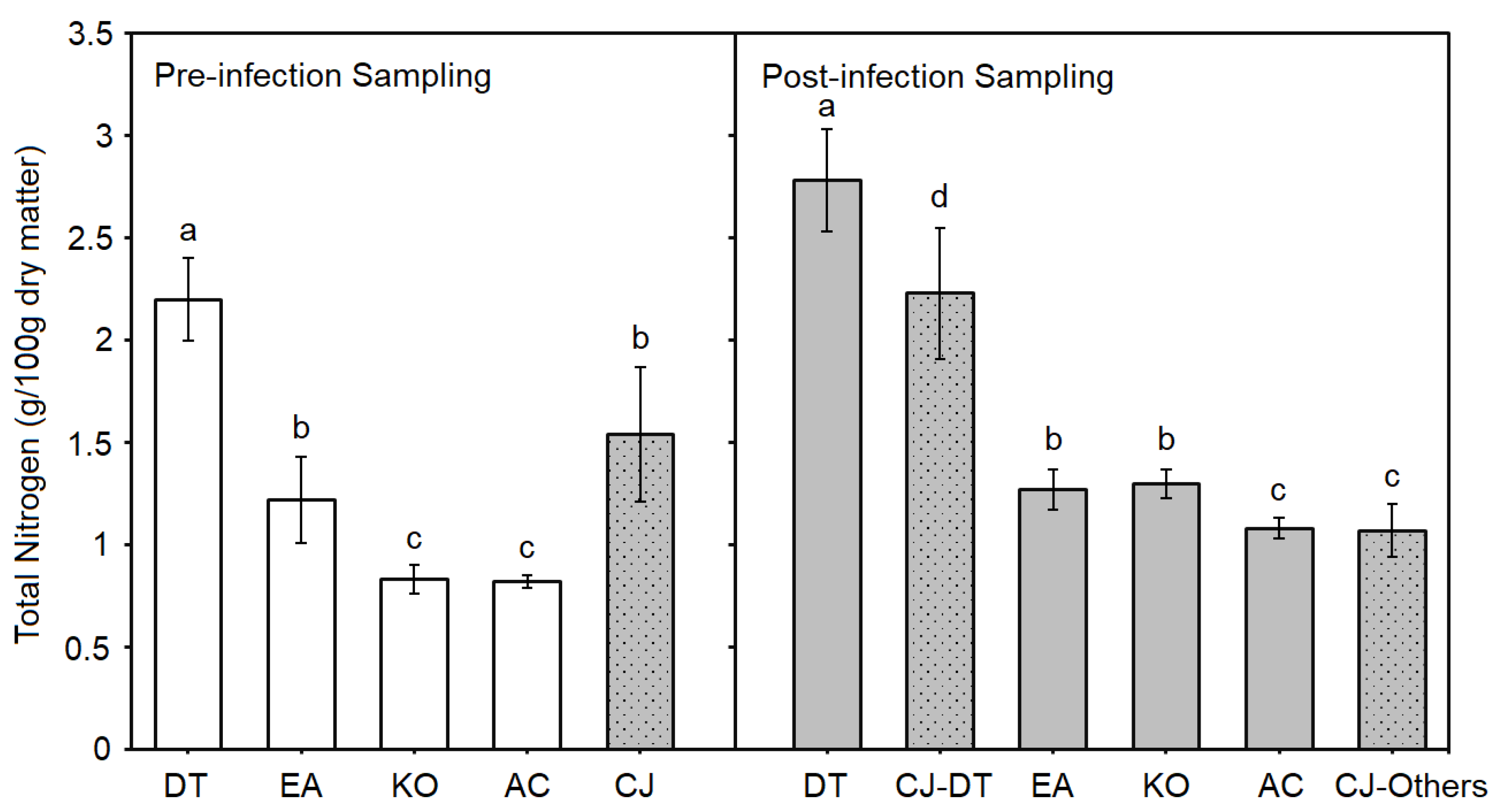

Table 1.
Two-way ANOVA results (P-values) for the effects of infection by Cuscuta japonica (IC) and host species (HS) on moisture content (%), total nitrogen content (g/100g), total phosphorous content (g/100g), total potassium content (g/kg), soluble sugar content (mg/g) and soluble protein content (mg/g). Significant effects are in bold; F and sum of square values are presented in Table S1.
Table 1.
Two-way ANOVA results (P-values) for the effects of infection by Cuscuta japonica (IC) and host species (HS) on moisture content (%), total nitrogen content (g/100g), total phosphorous content (g/100g), total potassium content (g/kg), soluble sugar content (mg/g) and soluble protein content (mg/g). Significant effects are in bold; F and sum of square values are presented in Table S1.
| Moisture | Nitrogen | Phosphorous | Potassium | Soluble Sugar | Soluble Protein | |
|---|---|---|---|---|---|---|
| HS | 0.000 | 0.000 | 0.000 | 0.001 | 0.003 | 0.000 |
| IC | 0.000 | 0.000 | 0.000 | 0.001 | 0.006 | 0.191 |
| HS × IC | 0.078 | 0.000 | 0.000 | 0.000 | 0.000 | 0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



