
Submitted:
19 June 2025
Posted:
20 June 2025
You are already at the latest version
Abstract
Calcium (Ca²⁺) is an essential nutrient for plants and enhances their resistance to heavy metal stress. This study employed hydroponic experiments to investigate how varying Ca²⁺ levels (0, 0.5, 2, and 5 mM) regulate maize seedling growth, physiological-biochemical traits, and cellular antioxidant defense systems under cadmium (Cd) stress, thereby alleviating Cd toxicity. The results showed that Ca²⁺ supplementation mitigated the inhibitory effects of Cd stress on root length, dry weight, and photosynthetic pigment content. Additionally, Ca²⁺ application increased the activities of superoxide dismutase (SOD) and catalase (CAT), as well as the contents of glutathione (GSH) and ascorbic acid (ASA) in leaves, with maximum increases of 14.9%, 65.39%, 146%, and 135%, respectively, compared to the control (CK). Under Ca²⁺ treatment, peroxidase (POD) activity significantly decreased, with a maximum reduction of 34.38% compared to CK, while malondialdehyde (MDA) content markedly increased, reaching 19.84% higher than CK. Principal component analysis (PCA) revealed that the 5 mM Ca²⁺ treatment achieved the highest composite score, indicating optimal antioxidant capacity in maize seedlings and reduced membrane lipid peroxidation caused by excessive Cd accumulation. This study enhances the understanding of calcium’s role in plant stress responses and provides insights for safe maize production in Cd-contaminated environments.
Keywords:
cadmium stress
; calcium
; maize seedlings
; antioxidant system
1. Introduction
The farmland soil environment is a critical factor influencing crop yield and quality. With the rapid industrialization in China, heavy metal pollution has emerged as a prominent environmental concern. Cadmium (Cd), characterized by high mobility and a high exceedance rate in soil monitoring sites, readily migrates from farmland to crops, subsequently entering the food chain and posing significant risks to human health. It is therefore recognized as one of the most hazardous heavy metals [1]. Cd absorbed through roots or foliage accumulates in plant tissues, interfering with diverse physiological and biochemical processes. This leads to metabolic disorders, growth inhibition, and severe oxidative stress [2]. The detrimental effects of Cd on plants include suppression of seed germination, structural damage to roots and cytoskeletal microtubules, inhibition of photosynthesis, and disruption of redox systems [3]. Studies have shown that Cd stress reduces wheat growth, yield, and photosynthetic efficiency while increasing malondialdehyde (MDA) and hydrogen peroxide (H₂O₂) content [4]. Furthermore, under high Cd concentrations, excessive reactive oxygen species (ROS), including H₂O₂, OH⁻, O₂·⁻, and O₂, are generated, inducing membrane lipid peroxidation and ultimately impairing crop growth [5]. Plants alleviate Cd toxicity by enhancing the activities of antioxidant enzymes, including superoxide dismutase (SOD), peroxidase (POD), catalase (CAT), ascorbate peroxidase (APX), and glutathione reductase (GR), as well as non-enzymatic antioxidants such as glutathione (GSH), ascorbic acid (ASA), mannitol, and flavonoids. These mechanisms prevent excessive reactive oxygen species (ROS) accumulation and maintain cellular redox balance [6]. A study on Aster subulatus revealed that under Cd stress, although biomass was significantly reduced, the pronounced increase in POD and CAT activities allowed the plant to sustain normal growth and demonstrate effective detoxification capabilities [7]. Calcium (Ca²⁺) enhances POD and SOD activities, promoting the elimination of intracellular peroxides via antioxidant groups. This process reduces malondialdehyde (MDA) content by suppressing lipid peroxidation, enhances plasma membrane stability, and alleviates Cd-induced cytotoxicity in rice cells [8] .
Calcium is an essential nutrient for plant growth, playing a critical role in regulating physiological and biochemical processes and maintaining cellular functions. As a second messenger, Ca²⁺ enhances plant resilience to abiotic stress through stress signaling pathways, thereby exerting protective and growth-promoting effects [9,10]. Studies indicate that exogenous calcium application mitigates cadmium (Cd) toxicity by alleviating Cd-induced physiological impairments, such as root growth inhibition, reactive oxygen species (ROS) accumulation, oxidative damage, and root tip cell death [11]. Research has shown that under varying Cd stress levels, exogenous calcium supplementation promotes plant height and biomass accumulation. Concurrently, it increases malondialdehyde (MDA) content while reducing antioxidant enzyme activities and Cd content in various plant tissues [12]. Furthermore, the accumulation and distribution of Cd within plants are influenced by the concentration and chemical forms of exogenous calcium. Therefore, it is imperative to investigate the specific mechanisms through which exogenous calcium regulates growth and Cd detoxification in maize seedlings.
Maize, a staple crop extensively cultivated in China, serves as both a primary forage resource and a vital industrial feedstock. Recognized for its high tolerance to diverse environmental stressors, it has been widely used as a key crop species in heavy metal contamination studies [2,13]. Recent studies demonstrate that calcium (Ca²⁺) supplementation alleviates cadmium (Cd)-induced photosynthetic decline in maize and effectively reduces oxidative damage in seedlings under both Cd stress and low-temperature stress [14].
The escalating cadmium (Cd) contamination in agricultural soils has caused substantial crop yield losses. Based on the dual roles of exogenous calcium (Ca²⁺) in regulating plant growth and alleviating cadmium-induced toxicity, and considering its critical multifunctional roles in plant growth and metabolic processes, we hypothesize that Ca²⁺ supplementation can effectively mitigate Cd-induced adverse effects on maize seedling growth. This study investigates the effects of varying calcium levels on maize seedling growth, physiological and biochemical parameters, and antioxidant enzyme activities under Cd stress, aiming to determine the extent to which exogenous Ca²⁺ alleviates Cd toxicity. Given the current lack of mechanistic understanding of Cd toxicity mitigation, this work will provide critical insights for future research on potential molecular mechanisms underlying heavy metal stress alleviation in diverse plant species.
2. Materials and Methods
2.1. Seedling Treatment
The test maize variety is the self-cross variety “Hongdan 3”, developed through multi-generation selection at the Honghe Prefecture Agricultural Research Institute in Yunnan,China. Seeds with plump kernels and uniform size were selected. After washing the seeds with deionized water, they were soaked in tap water, germinated in the dark, and sprouted in vermiculite. The maize seedlings that grew uniformly were transplanted to a solar greenhouse and cultivated with a 1/2 Hogland nutrient solution (pH 5.5~6.5).
2.2. Selection of Cd Levels
Different concentrations of Cd chloride were used (0, 50, 100, 150, 200, 250, 300, 350μM). The preliminary results showed that 250 μM Cd inhibited the growth and germination of 50% of maize seedlings.
2.3. Experimental Design
After growing in 1/2 Hoagland nutrient solution for 5 days, the plants were transferred to containers filled with full-strength Hoagland solution and subjected to the following treatments: Control (CK): 0 μM Cd; k-1: 0.5 mM Ca + 250 μM Cd; k-2: 2 mM Ca + 250 μM Cd; k-3: 5 mM Ca + 250 μM Cd. Both Cd and Ca were added simultaneously to the solution. A total of 40 L nutrient solution was placed in lidless rectangular containers, with three plants cultivated per 200 mL of solution, resulting in 60 plants in total. The experiment included three replicates and lasted for 30 days.
2.4. Plant Sample Collection and Measurements
At the seedling stage (4–6 leaf stage; 30 days post-emergence), plant materials were collected to assess growth parameters, physiological and biochemical traits, and antioxidant defense system enzymes. Seedlings were rinsed with distilled water, and growth attributes were recorded. Maize leaf samples were frozen at –80°C for subsequent analysis of physiological/biochemical traits and antioxidant activity. Sampled seedlings were oven-dried at 70°C until constant weight to determine root dry weight (RDW) and shoot dry weight (SDW).
2.5. Growth Parameter Measurements
Stem length (SL) was measured from the base to the tip of the top leaf for each plant. Roots were carefully removed from the nutrient solution to record root length (RL).
2.6. Determination of Photosynthetic Pigments
Chlorophyll content was determined according to Arnon [15], and carotenoid content was measured using the method of Davis [16]. Leaf samples (0.1 g) were homogenized in 5 mL of 80% (v/v) acetone, filtered, and absorbance was recorded at 645, 663, and 480 nm using a UV spectrophotometer (UV2700, Shimadzu, Japan). The formulas for photosynthetic pigment calculations were as follows:
Chl. a (mg/g fresh weight) = [12.7(OD₆₆₃) – 2.69(OD₆₄₅)] × V/(1000 × W)
Chl. b (mg/g fresh weight) = [22.9(OD₆₄₅) – 4.68(OD₆₆₃)] × V/(1000 × W)
Carotenoids (mg/g fresh weight) = [(OD₄₈₀ + 0.114(OD₆₆₃) – 0.638(OD₆₄₅)] / 2500
where V represents the volume of acetone (mL), and W denotes the fresh weight (g) of the leaf sample.
2.7. MDA Determination
Malondialdehyde (MDA) content was determined via the thiobarbituric acid method [17]. Briefly, 0.1 g of tissue was homogenized in 10% trichloroacetic acid (TCA) and centrifuged at 12,000 × *g* for 10 min. Subsequently, 2 mL of supernatant was mixed with 2 mL of 10% TCA in a fresh centrifuge tube, boiled for 15 min, and cooled on ice. After centrifugation at 3,000 × *g* for 10 min, the absorbance of the supernatant was measured at 532 nm and 600 nm.
2.8. Determination of Antioxidant Enzyme Activities
Enzyme extraction: Leaf samples (0.1 g) were ground in a pre-cooled mortar with ice-cold 50 mM phosphate buffer (pH 7.0, containing 1% PVP, 100 μM EDTA, and 5 mM DTT). The homogenate was centrifuged at 12,000 × *g* for 20 min at 4°C, and the supernatant was collected as the crude enzyme extract.
Superoxide dismutase (SOD) activity was assayed by the nitroblue tetrazolium (NBT) photoreduction method [18], with one unit defined as the amount of enzyme required to inhibit 50% of NBT photoreduction.
Peroxidase (POD) activity was measured via the guaiacol colorimetric method [19], where one unit corresponds to a ΔA₄₇₀ of 0.01 per minute. Catalase (CAT) activity was determined by monitoring the decrease in absorbance at 240 nm, with one unit defined as a reduction of 0.1 absorbance units per minute.
2.9. Determination of Oxidative Substances
Glutathione (GSH) content was measured by the DTNB method[19]. Samples (0.5 g) were divided into two portions: one for dry weight determination and the other for GSH extraction. The tissue was homogenized in 5 mL of 5% trichloroacetic acid (TCA), centrifuged at 15,000 × *g* for 10 min, and the supernatant was adjusted to 5 mL. A reaction mixture containing 0.25 mL supernatant, 2.6 mL of 150 mM NaH2PO4 (pH 7.7), and 0.18 mL DTNB reagent was incubated at 30°C for 5 min, and absorbance was measured at 412 nm.
Ascorbic acid content was determined following Nino and Shah [20]. Fresh maize tissue (0.1 g) was ground with thiobarbituric acid (TBA) and centrifuged at 10,000 × *g* for 10 min. An aliquot (500 μL) of the supernatant was mixed with 2 mL of diluted sulfuric acid, incubated at 37°C for 30 min, and centrifuged at 12,000 × *g*. Absorbance was measured at 520 nm using a spectrophotometer (Hitachi U-2910, Tokyo, Japan).
2.10. Determination of Cd and Ca Content in Plants
Dried shoots and roots of maize were ground to a fine powder. Samples (0.2 g) were digested with 2 mL H2O2 and 6 mL HNO3 in a microwave digestion system (Labtech, Ethos One) under the following conditions: preheating for 10 min, heating at 190°C for 15 min, and cooling for 20 min. The digestate was diluted to 50 mL with deionized water. Cd and Ca2+ concentrations were quantified by inductively coupled plasma optical emission spectrometry (ICP-OES, PerkinElmer). The Cd translocation factor was calculated as the ratio of Cd accumulation in shoots to that in roots [21].
2.11. Data Processing
Data were statistically analyzed and visualized using Excel 2017. One-way ANOVA with Duncan’s test (*p* < 0.05) was performed in IBM SPSS Statistics 26. Correlation analysis, principal component analysis (PCA), and plotting were conducted using R language.
3. Results and Analysis
3.1. Effects of Different Calcium Levels on Growth Characteristics of Maize Seedlings Under Cadmium Stress
Under Cd stress, plant height, root length, above-ground dry weight (ADW), and root dry weight (RDW) of maize seedlings increased with rising Ca²⁺ concentrations, reaching the highest values in the K-3 treatment (1.99%, 10.60%, 15.72%, and 3.64% higher than CK, respectively). Root length and ADW in K-3 differed significantly (*p* < 0.05) from CK, K-1, and K-2, while its plant height and RDW showed no significant difference compared to CK. Under K-1 treatment, plant height, root length, and ADW were significantly lower than CK, though RDW reduction was not statistically significant. These results indicate that low Ca²⁺ concentrations exacerbate Cd stress, whereas high Ca²⁺ levels mitigate Cd toxicity.
3.2. Effects of Different Calcium Levels on Photosynthetic Pigment Content in Maize Seedlings Under Cd Stress
Under Cd stress, chlorophyll *a*, chlorophyll *b*, and carotenoid contents in Ca-treated seedlings exhibited an initial decrease followed by an increase with rising Ca²⁺ levels (Table 2). Chlorophyll *a* content in all Ca treatments was significantly lower (*p* < 0.05) than CK, with K-2 showing the lowest value (15.94% reduction). Chlorophyll *b* content in all treatments except K-2 was significantly higher (*p* < 0.05) than CK, peaking in K-1 (42.05% increase). Carotenoid content in K-1 and K-2 was significantly lower than CK, with K-2 showing the lowest value (22.27% reduction). Overall, 5 mM Ca²⁺ treatment (K-3) enhanced chlorophyll *a* and carotenoid accumulation.

Table 1.
Effects of different Ca2+ levels on the growth and photosynthetic pigments in maize under Cd stress.
Table 1.
Effects of different Ca2+ levels on the growth and photosynthetic pigments in maize under Cd stress.
| Treatment | Shoot Length (cm) |
Root Length (cm) |
Shoot Dry Weight (g) |
Root Dry Weight (g) |
|---|---|---|---|---|
| CK | 45.27±0.23a | 31.98±0.23c | 15.46±0.26b | 3.57±0.13a |
| K-1 | 43.55±0.15b | 30.75±0.36b | 14.23±0.32c | 3.39±0.19a |
| K-2 | 45.19±0.81a | 33.72±0.09b | 16.03±0.38b | 3.47±0.09a |
| K-3 | 46.17±0.20a | 35.37±0.13a | 17.89±0.31a | 3.70±0.11a |
Means provided with error bars; in columns different letter indicates significance (P<0.05).
3.3. Effects of Different Calcium Levels on MDA Content in Maize Under Cd Stress
Significant differences (*p* < 0.05) were observed in GSH, AsA, and MDA contents among treatments (Figure 1). GSH content in all Ca treatments was significantly higher than CK, peaking in K-2 (146% increase, Figure 1A). AsA content increased significantly (*p* < 0.05) in all Ca²⁺ treatments compared to CK, with K-3 showing the highest value (135% increase, Figure 1B). MDA content in Ca treatments was significantly elevated (*p* < 0.05) relative to CK, reaching a maximum in K-2 (23.44% increase), though no significant difference existed between K-2 and K-3 (Figure 1C).
3.4. Antioxidant Enzyme Activities
POD activity in Ca-treated seedlings under Cd stress was significantly lower (*p* < 0.05) than CK, showing an initial increase followed by a decline with rising Ca²⁺ levels. K-2 exhibited the highest POD activity (11.13% reduction vs. CK), while K-3 showed the lowest (34.38% reduction). No significant difference was observed between K-1 and K-2 (Figure 1D).
SOD activity decreased initially and then increased with Ca²⁺ levels, peaking in K-1 (23.68% increase vs. CK), while K-2 was significantly lower than CK (Figure 2E).
CAT activity in all Ca treatments was significantly higher (*p* < 0.05) than CK, peaking in K-2 (66.80% increase), with no significant difference between K-2 and K-3 (Figure 1F).
3.5. Cd and Ca Accumulation in Plant Tissues Under Varying Calcium Levels
Cd content in shoots and roots increased significantly (*p* < 0.05) with Ca²⁺ supply, peaking in K-3 (3.43 μg g⁻¹ and 20.71 μg g⁻¹, respectively). The Cd translocation factor followed K-2 > K-3 > K-1, with K-1 being 29.14% lower than K-2, indicating that low Ca²⁺ inhibits Cd translocation from roots to shoots. The 5 mM Ca²⁺ treatment significantly increased (*p* < 0.05) Ca content in shoots and roots by 48.05% and 52.42%, respectively, compared to CK (Figure 2).
3.6. Correlation Matrix
Under cadmium stress, peroxidase (POD) exhibited a highly significant positive correlation (*p*< 0.001) with chlorophyll *b*, ascorbic acid (AsA), and root Cd content. Chlorophyll *a* showed a significant positive correlation (*p* < 0.001) with root Ca, shoot Cd, catalase (CAT), malondialdehyde (MDA), and glutathione (GSH). Superoxide dismutase (SOD) was significantly positively correlated (*p* < 0.001) with chlorophyll *b*. AsA displayed a highly significant positive correlation (*p* < 0.001) with root Cd content. CAT showed a significant negative correlation (*p* < 0.001) with MDA and GSH. Root Ca content was negatively correlated (*p* < 0.001) with shoot Ca and GSH, and negatively correlated (*p* < 0.01) with shoot Cd, CAT, and MDA. Shoot Ca exhibited a significant negative correlation (*p* < 0.01) with MDA and GSH. Root Cd was negatively correlated (*p* < 0.001) with shoot Cd, CAT, and MDA. Shoot Cd content showed a highly significant negative correlation (*p* < 0.001) with CAT, MDA, and GSH(Figure 3).
3.7. Principal Component Analysis (PCA)
PCA results revealed substantial differences in the effects of calcium treatments on maize seedling growth and physiological traits. Under different calcium treatments, the first principal component (PC1) accounted for 76.87% of the variance, while the second principal component (PC2) explained 21.69%, cumulatively contributing 98.56% of the total variance(Figure 4A). Physiological and biochemical traits of maize seedlings varied markedly across treatments. For PC1, GSH exhibited the highest contribution rate, followed by CAT. For PC2, root Cd contributed most significantly, followed by AsA(Figure 4B). Based on comprehensive evaluation, the 5 mM Ca²⁺ treatment ranked highest, indicating its pronounced impact on photosynthetic pigment content and antioxidant metabolism in maize under Cd stress.
4. Discussion
Cd is a harmful element that adversely affects plant growth and development. It readily accumulates in plant tissues, causing physiological and metabolic disorders, growth inhibition, and severe oxidative stress [2]. As an essential mineral element for plants, Ca²⁺ also functions as a versatile signaling ion, playing critical roles at multiple sites within various signaling cascade networks. Consequently, Ca²⁺ enhances plant stress resistance [21,22]. Exogenous application of can improve plant adaptability to abiotic stresses and mitigate damage caused by adverse conditions [23].
Under heavy metal stress, the redox balance in plants is disrupted, leading to excessive production of reactive oxygen species (ROS), which triggers membrane lipid peroxidation and damages the cellular membrane system [24]. To counteract oxidative damage, plants employ both enzymatic (SOD, POD, PPO, APX, GR) and non-enzymatic (AsA-GSH cycle) mechanisms to scavenge excess ROS. Under normal conditions, enzymes such as SOD, POD, and CAT exhibit strong ROS-scavenging capacity, enabling timely removal of reactive oxygen species. Under stress conditions, the production of free radicals in plant cells increases significantly. These free radicals initiate membrane lipid peroxidation, leading to membrane damage and reduced activity of ROS-scavenging enzymes. Glutathione (GSH) and ascorbic acid (AsA) are essential small-molecule antioxidants in plants. Together, they constitute the non-enzymatic antioxidant system and participate in cellular redox processes [25]. AsA is catalyzed by APX to convert H₂O₂ into harmless H₂O₂, while GSH is regenerated via GR to eliminate ROS. Additionally, GSH promotes the synthesis of non-protein thiols (NPTs) and phytochelatins (PCs), thereby inactivating free metal ions [26]. Exogenous Ca²⁺ application enhances SOD and CAT activities in tomatoes increases AsA and GSH contents in Stachys lanata, and alleviates Cd stress, which aligns with the findings of this study. POD catalyzes the decomposition of peroxides (including H₂O₂) and participates in lignin biosynthesis [27]. Since Cd transport is influenced by xylem stability [28]. POD activity may indirectly affect Cd accumulation. This study revealed a highly significant positive correlation between POD activity and root Cd content, as well as a significant positive correlation with shoot Cd content. Medium-to-high Ca²⁺ treatments enhanced SOD and CAT activities, increased antioxidant contents (e.g., GSH and AsA), and improved membrane stability. This may be attributed to the Ca²⁺-calmodulin (CaM) complex regulating enzyme activities (including SOD, POD, and CAT) or Ca²⁺ reducing Cd’s ability to displace metal ions at enzyme active sites, thereby mitigating Cd-induced inhibition of antioxidant systems. Future studies should analyze spectral characteristics of maize antioxidant enzymes and related gene expression under exogenous Ca²⁺ treatment to elucidate specific mechanisms of Ca²⁺-mediated enzyme regulation.
Malondialdehyde (MDA) is a product of lipid peroxidation in organism metabolism and biological membrane systems. Its concentration reflects the degree of cell membrane lipid peroxidation [29]. Under Cd stress, an increased MDA trend was observed in maize seedlings, indicating aggravated oxidative damage. This aligns with the findings of LiNa [30], who reported that high calcium concentrations elevated MDA levels in Panax notoginseng under 0.6 mg/kg Cd stress. This phenomenon may result from calcium chloride (CaCl₂) exacerbating Cd toxicity in plants.
In this experiment, 5 mM Ca²⁺ treatment significantly increased the CH1b content in maize seedlings, while the Car content showed no significant change (Table 2), and CH1a exhibited a downward trend. These results are consistent with the findings of Alrashidi et al. [31], where foliar application of calcium fertilizer on tomatoes significantly enhanced CH1b content. This phenomenon may be attributed to exogenous calcium regulating the activity of CH1b synthase or acting as a signaling molecule to promote CH1b synthesis [32]. In this experiment, CH1a showed a significant positive correlation with CAT and GSH, while CH1b was positively correlated with ASA, indicating that the photosynthetic system and antioxidant system can mutually influence each other. This is because photosynthetic pigments not only participate in photosynthesis but also eliminate reactive oxygen species (ROS) in plant cells, thereby alleviating oxidative stress [33]. Furthermore, based on the results of this study, supplemental application of calcium fertilizer in agricultural practices-particularly in Cd-contaminated farmlands—can improve the photosynthetic performance of maize seedlings and enhance their productivity under heavy metal stress conditions.
Relevant studies have shown that the addition of exogenous calcium can reduce cadmium (Cd) content and its translocation capacity in plants such as rice [33] and pak choi [34]. However, other studies have reported that exogenous calcium increased Cd content in leaves and stems, as well as translocation coefficients, in rice during both the tillering and maturity stages [35], which may be attributed to differences in plant cultivars and environmental conditions. In this experiment, medium-to-high calcium treatments promoted Cd uptake and translocation (Table 3). This phenomenon could be explained by Cd stress accelerating the synthesis of glutathione (GSH) and phytochelatins (PCs) in plants. These compounds bind to Cd to form highly soluble complexes, facilitating their transport via the xylem and rapid accumulation in shoots [36]. Concurrently, calcium treatment enhanced Cd translocation from roots to shoots, as evidenced by a highly significant increase in the translocation coefficient and the expression levels of Cd transport-related genes (OsNRAMP5 and OsHMA2) (p<0.01) [37]. This study has not yet elucidated the specific regulatory mechanisms by which exogenous calcium influences plant Cd uptake. Further research should focus on the subcellular distribution of Cd and the responses of related transporter protein genes.
5. Conclusions
Cadmium stress induced an increasing trend in malondialdehyde (MDA) content in maize seedlings, indicating aggravated oxidative damage that inhibited seedling growth. The addition of exogenous Ca²⁺ alleviated the inhibitory effects of Cd stress on root length, dry weight, and photosynthetic pigment content, while enhancing the activities of superoxide dismutase (SOD) and catalase (CAT) in leaves. Under Cd stress, medium-to-high Ca²⁺ concentrations elevated glutathione (GSH) and ascorbic acid (AsA) levels in seedlings, thereby reducing Cd-induced lipid peroxidation in cell membranes. Principal component analysis (PCA) revealed that the 5 mM Ca²⁺ treatment achieved the highest comprehensive score, optimizing all measured maize parameters. This study not only advances the understanding of calcium’s role in plant stress response mechanisms but also provides novel strategies for safe maize production in Cd-contaminated soils, demonstrating significant theoretical innovation and practical value.
References
- Li, Y.N.; Tan, M.T.; Wu, H.F.; Zhang, A.Y.; Xu, J.S.; Meng, Z.J. Transfer of Cd along the food chain: The susceptibility of Hyphantria cunea larvae to Beauveria bassiana under Cd stress. Journal of HazardousMaterials. 2023, 453, 131420. [Google Scholar] [CrossRef] [PubMed]
- Muhammad, A.; Naz, S.; Kumar, R.; Sardar, H.; Nawaz, M.A.; Kumar, A. Unraveling the mechanisms of cadmium toxicity in horticultural plants: implications for plant health. South African Journal Of Botany. 2023, 12, 433–442. [Google Scholar] [CrossRef]
- Rasafi, T.E.; Oukarroum, A.; Haddioui, A.; Song, H.; Rinklebe, J.; Nanthi, B. Cadmium stress in plants: a critical review of the effects, mechanisms, and tolerance strategies. Critical Reviews in Environmental Science and Technology. 2022, 52, 675–726. [Google Scholar] [CrossRef]
- Yang, J.K.; Zhu, L.N.; Yang, Q.Y. Effects of Silicon-Calcium-Magnesium Fertilizer and Modified Humic Acid on Soil Cadmium Chemical Fractions and Accumulation in Wheat. Journal of Ecology and Rural Environment. 2021, 37, 808–816. [Google Scholar] [CrossRef]
- Maleki, M.; Ghorbanpour, M.; Kariman, K. Physiological and antioxidative responses of medicinal plants exposed to heavy metals stress. Plant Gene. 2017, 11, 247–54. [Google Scholar] [CrossRef]
- Huang, Y.X.; Wu, X.; Cao, Y.; Yan, X.Y.; Li, L. Research Progress on Effect and Mechanism of Exogenous Calcium Induced Plant Response to Cadmium Stress. Journal of Agricultural Science and Technology. 2024. [CrossRef]
- Chen, W.; Jiang, W.Y.; Yang, Y.X.; Liao, J.; Liang, X.L.; Wang, H.J. Physiological response and cadmium accumulation and translocation characteristics of Aster subulatus Michx. to cadmium stress. Southwest China Journal of Agricultural Sciences. 2022, 35, 2860–2866. [Google Scholar] [CrossRef]
- Ma, Q.; Chen, B.; Zhao, Y.Y.; Zeng, Q.R.; Deng, X. Effect of Combined Application of Calcium Magnesium Phosphate-potassium Carbonate-lime on Cadmium Accumulation in Rice (Oryza sativa L.). Chinese Journal of Soil Science. 2024, 55, 819–829. [Google Scholar] [CrossRef]
- El-Beltagi, H.S.; Mohamed, H.I. Alleviation of cadmium toxicity in Pisum sativum L. seedlings by calcium chloride. Notulae Botanicae Horti Agrobotanici Cluj-Napoca 2013, 41, 157–168. [Google Scholar] [CrossRef]
- Malik, Z., Afzal, S., Danish, M., Abbasi, G.H., Bukhari, S.A.H., Khan, M.I., 2020. Role of nitric oxide and calcium signaling in abiotic stress tolerance in plants. Protective Chemical Agents in the Amelioration of Plant Abiotic Stress: Biochemical and Molecular Perspectives 2020, 563–581. [CrossRef]
- Yan, X. 2019. Toxicity of cadmium on roots of different genotypes of rice and regulation of calcium [D]. Chengdu: Sichuan Agricultural University.
- Wang, F.; Li, Y.S.; Wang, H.N.; Peng, Y.L.; Fang, Y.F.; Wang, W.; Ma, Y.Z. Effect of calcium on growth and physiological characteristics of maize seedling under lead stress. Shuitu Baochi Xuebao (Journal of Soil and Water Conservation) 2016, 30, 202–207. [Google Scholar]
- Shi, X.; Meng, H.L.; Wang, T.T.; Tang, Y.H. Effects of Bacterial fertilizer on the Growth and Absorption of Cadmium and Lead in Maize. Bangladesh J. Bot. 2024, 53, 297–304. [Google Scholar] [CrossRef]
- Saleem, M.; Parveen, A.; Khan, S.; Hussain, I.; Wang, X.K.; Alshaya, H. Silicon Fertigation Regimes Attenuates Cadmium Toxicity and Phytoremediation Potential in Two Maize (Zea mays L.) Cultivars by Minimizing Its Uptake and Oxidative Stress. Sustainability 2022, 14, 2–18. [Google Scholar] [CrossRef]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1. [Google Scholar] [CrossRef] [PubMed]
- Davies, B. Chemistry and biochemistry of plant pigments. London: Academic Press. 1976, 38-165.
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts: Kinetics and stoichiometry of fatty acid peroxidation. Arch Biochem Biophys. 1968, 125, 189–98. [Google Scholar] [CrossRef] [PubMed]
- Li, H.S.; Sun, Q.; Zhao, S.J. Experimental Principles and Techniques of Plant Physiology and Biochemistry [M]. Beijing: Higher Education Press. 2000, 167-169.
- Chen, J.X.; Wang, X.F. . Plant Physiology Experiment Guidance [M]. Guangzhou: South China University of Technology Press. 2002. [Google Scholar]
- Nino, H.; Shah, W. Vitamins In: Fundamentals of Clinical Chemistry. Tietz, NW. WB Saunders, Philadelphia.
- Lamia, S.; Muhammad, H.; Yoshiyuki, M. Role of calcium signaling in cadmium stress mitigation by indol-3-acetic acid and gibberellin in chickpea seedlings. Environmental Science & Pollution Research. [CrossRef]
- Shumayla, T.S.; Sharma, Y. Expression of TaNCL2—a ameliorates cadmium toxicity by increasing calcium and enzymatic antioxidants activities in Arabidopsis. Chemosphere 2023, 329, 138636. [Google Scholar] [CrossRef]
- Sun, L.; Liu, M.H.; Yi, L.T. Effects of exogenous calcium on growth and physiological characteristics of poplars under cadmium stress. Asian Journal of Ecotoxicology, (in Chinese). 2024, 19, 390–403. [Google Scholar] [CrossRef]
- Wang, Y.X.; Zheng, W.Y.; Hou, L.; Liu, C.T.; bHe, C.Z.; Chen, Z.X. Study on Physiological Response and Tolerance of Populus yunnanensis Seedlings to Cadmium and Zinc Stress. Journal of southwest foredtry university, 2024, 44, 31–39. [Google Scholar] [CrossRef]
- Mishra, B.; Sangwan, R.S.; Mishra, S.; Jadaun, J.S.; Sabir, F.; Sangwan, N.S. Effect of cadmium stress on inductive enzymatic and nonenzymatic responses of ROS and sugar metabolism in multiple shoot cultures of Ashwagandha (Withania somnifera Dunal). Protoplasma 2014, 1031–1045. [Google Scholar] [CrossRef]
- Qin, S.; Liu, H.; Nie, Z.; Gao, W.; Li, C.; Lin, Y.; Zhao, P. AsA-GSH Cycle and Antioxidant Enzymes Play Important Roles in Cd Tolerance of Wheat. Bulletin of Environmental Contamination and Toxicology, 2018, 101, 684–690. [Google Scholar] [CrossRef]
- Tang, X.Y.; Li, H.Y.; Chen, F.; Xu, X.X.; Qi, X.; Zhang, S. Effect of Foliar Spraying Calcium Oxide Nanoparticles on Cadmium Accumulation and Physiological Response in Coriander and Spinach under Cadmium Stress. Journal of Agro-Environment Science. 2024. [Google Scholar]
- Long, C.Y. Release effects of exogenous SA,GSH,Ca and CA on Stachys lanata under Cd stress, Sichuan Agricultural University, 2019. 6.
- Mazhar, M.W.; Ishtiaq, M.; Maqbool, M.; Ajaib, M.; Hussain, I.; Hussain, T.; Parveen, A. Synergistic application of calcium oxide nanoparticles and farmyard manure induces cadmium tolerance in mung bean (Vigna radiata L.) by influencing physiological and biochemical parameters. PLoS One 2023, 18, e0282531. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Li, T.; Mei, X.Y.; Ruan, M.J.; Wu, Y.Q.; Zu, Y.Q. Effects of Two Exogenous Calcium on Physiological and Biochemical Characteristics and Cadmiun Accumulation of Panax notoginseng of Cadmium Stress. Guizhou Agricultural Sciences China. 2019, 47, 131–137. [Google Scholar]
- Wu, J.Y.; Pan, Q.W.; Muhammad, F.; Zhang, L.l.; Gong, H.; Liu, L.J.; Yang, G.; Wang, W.T.; Pu, Y.Y.; Fang, Y.; Ma, L.; Sun, W.C. Effects of exogenous calcium and calcium inhibitor on physiological characteristics of winter turnip rape (Brassica rapa) under low temperature stress. Junyan et al. BMC Plant Biology. 2024, 24, 937. [Google Scholar] [CrossRef]
- Mourato, M.; Reis, R.; Louro, L.M. Characterization of plant antioxidative system in response to abiotic stresses: a focus on heavy metal toxicity[M]. Advances in Selected Plant Physiology Aspects. 2012, 23–44. [Google Scholar]
- Liu, J.L.; Feng, X.Y.; Qiu, G.Y. Inhibition Roles of Calcium in Cadmium Uptake and Translocation in Rice: A Review. International journal of molecular sciences 2023, 24. [Google Scholar] [CrossRef]
- Treesubsuntorn, C.; Thiravetyan, P. Calcium acetate-induced reduction of cadmium accumulation in Oryza sativa: expression of auto-inhibited calcium-ATPase and cadmium transporters. Plant Biology. 2019, 21, 862–872. [Google Scholar] [CrossRef]
- Mishra, N.; Jiang, C.K.; Chen, L. Achieving abiotic stress tolerance in plants through antioxidative defense mechanisms. Frontiers in plant science. 2023, 14, 1110622. [Google Scholar] [CrossRef]
- Cheng, Y.W.; Wang, N.; Liu, R.X. Cinnamaldehyde Facilitates Cadmium Tolerance by Modulating Ca2+ in Brassica rapa. Water, Air, & Soil Pollution 2021, 232. [Google Scholar]
- Ye, W.L.; Wu, F.; Zhang, G.Y.; Fang, Q.; Lu, H.J.; Hu, H.X. Calcium decreases cadmium concentration in root but facilitates cadmium translocation from root to shoot in rice. J. Plant Growth Regul. 2020, 39, 422–429. [Google Scholar] [CrossRef]
Figure 1.
Effect of calcium(Ca;CK-0mM Ca+0μM Cd,k-1: 0.5mM Ca+250μM Cd; k-2: 2mM Ca+ 250 μM Cd; k-3: 5mM Ca+250 μM Cd) and cadmium(Cd) treatments on the A) glutathione (GSH), B) ascorbic acid(ASA), C) Malondialdehyde(MDA), D) peroxidase (POD), E) superoxide dismutase(SOD),F) catalase(CAT) of maize seedlings. Date are the mean±standard error(n=3).Differernt letters indicate significant differences among treatments(ANOVA,Duncan`s test, (P<0.05).
Figure 1.
Effect of calcium(Ca;CK-0mM Ca+0μM Cd,k-1: 0.5mM Ca+250μM Cd; k-2: 2mM Ca+ 250 μM Cd; k-3: 5mM Ca+250 μM Cd) and cadmium(Cd) treatments on the A) glutathione (GSH), B) ascorbic acid(ASA), C) Malondialdehyde(MDA), D) peroxidase (POD), E) superoxide dismutase(SOD),F) catalase(CAT) of maize seedlings. Date are the mean±standard error(n=3).Differernt letters indicate significant differences among treatments(ANOVA,Duncan`s test, (P<0.05).
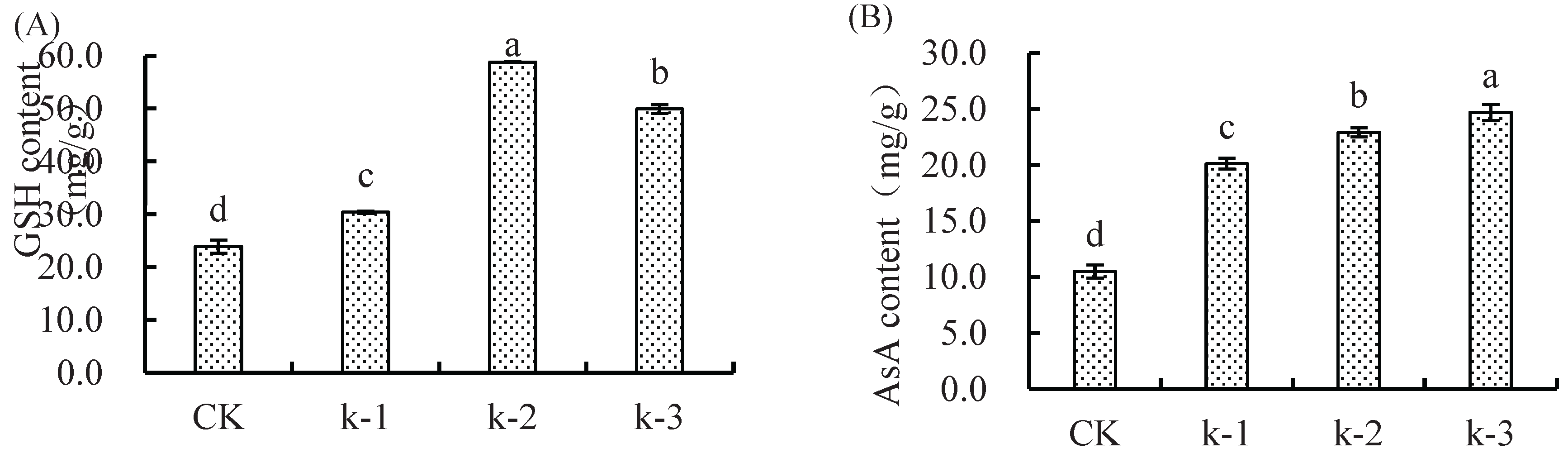
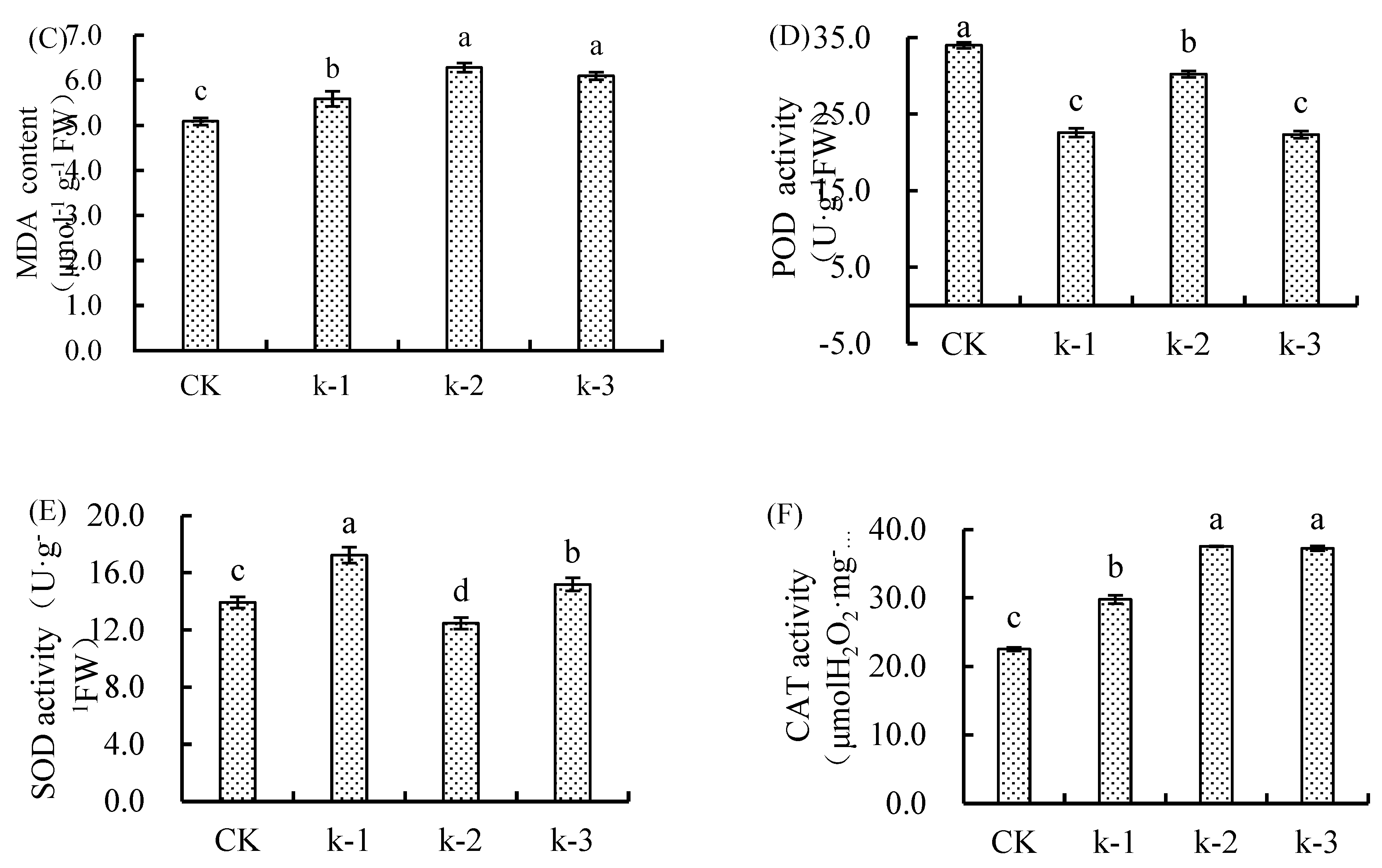
Figure 2.
Ca2+ content of above-ground parts and roots of maize seedlings under Cd stress at different Ca2+ levels.
Figure 2.
Ca2+ content of above-ground parts and roots of maize seedlings under Cd stress at different Ca2+ levels.
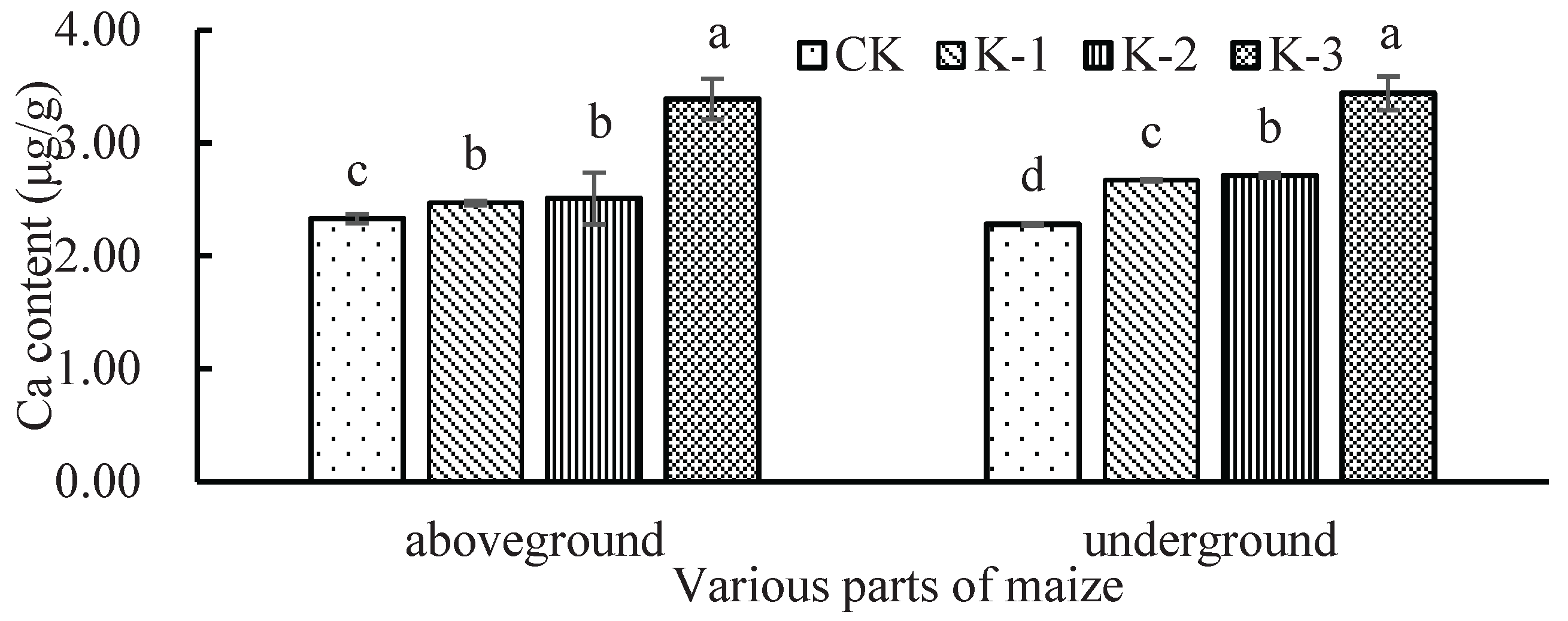
Figure 3.
Correlation between morphological and physiological biochemical traits of maize seedlings under Cd stress.
Figure 3.
Correlation between morphological and physiological biochemical traits of maize seedlings under Cd stress.
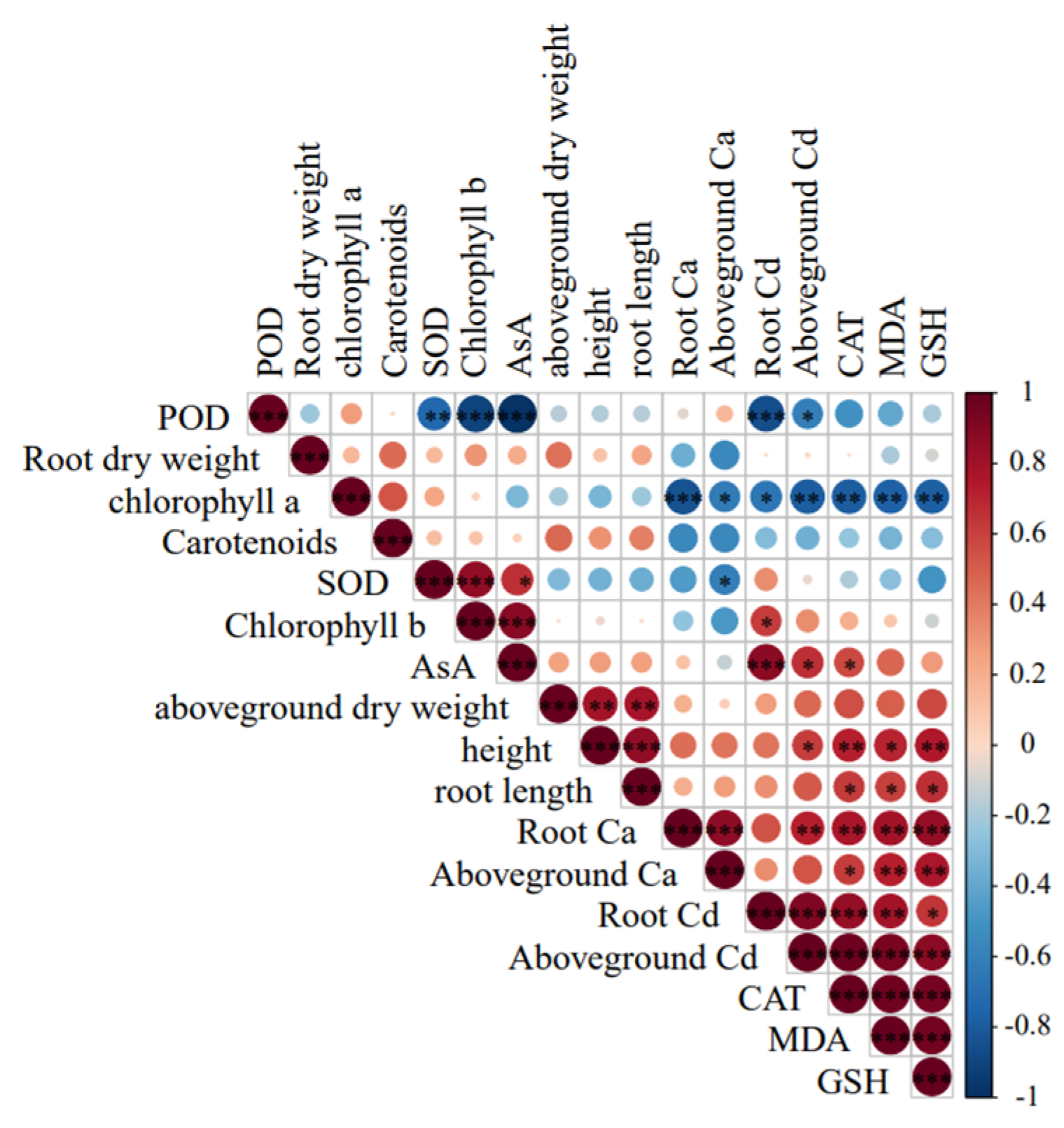
Figure 4.
PCA of growth and physiological and biochemical indexes of maize at different Ca2+ levels under Cd stress.
Figure 4.
PCA of growth and physiological and biochemical indexes of maize at different Ca2+ levels under Cd stress.
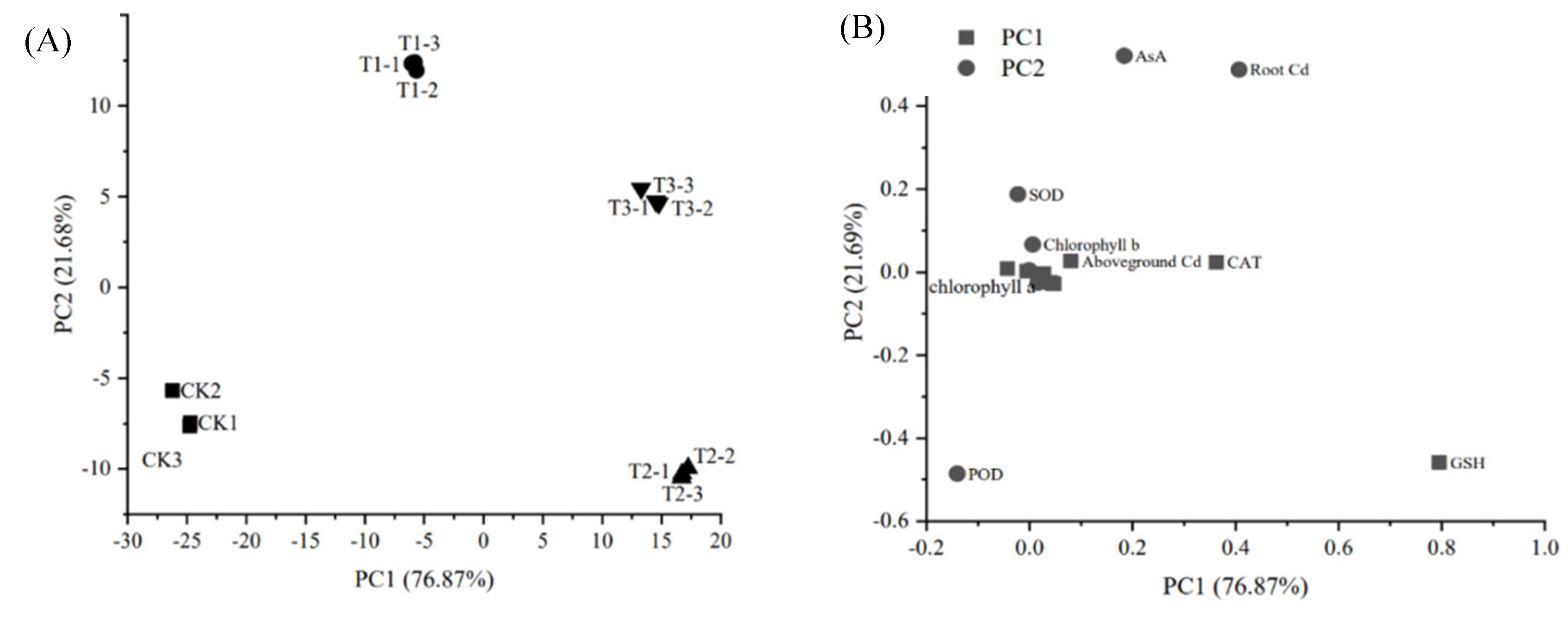
Table 2.
Effects of different Ca2+ levels on photosynthetic pigments in maize seedlings under Cd stress.
Table 2.
Effects of different Ca2+ levels on photosynthetic pigments in maize seedlings under Cd stress.
| Treatment | Chl a (mg g-1 FW) | Chl b (mg g-1 FW) | Caro. (mg g-1 FW) |
|---|---|---|---|
| CK | 14.37±0.10a | 3.02±0.12b | 2.20±0.10a |
| K-1 | 13.17±0.30b | 4.13±0.06a | 1.83±0.11b |
| K-2 | 12.08±0.37c | 2.96±0.18b | 1.71±0.11b |
| K-3 | 13.11±0.68d | 4.29±0.13a | 2.25±0.12a |
Means provided with error bars; in columns different letter indicates significance (P<0.05) treatments. Abbreviations: Chlorophyll a (Chl a); Chlorophyll b (Chl b); Carotenoids (Caro).
Table 3.
Effects of Cd concentration and Cd transport rate in different parts of maize at different Ca2+ levels under Cd stress.
Table 3.
Effects of Cd concentration and Cd transport rate in different parts of maize at different Ca2+ levels under Cd stress.
| Treatment CK |
Cd Concentration(μg g-1) | Transfer Coefficient(IF) | |
|---|---|---|---|
| Above-Ground Parts | Roots | Cd Above-Ground Parts/Cd Roots | |
| K-1 | 2.18±0.04c | 18.13±0.31b | 0.1201±0.0025b |
| K-2 | 3.37±0.11b | 19.88±0.21c | 0.1695±0.0005a |
| K-3 | 3.43±0.10a | 20.71±0.14a | 0.1656±0.0010a |
Means provided with error bars; in columns different letter indicates significance (P<0.05) treatments.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



