
Submitted:
13 June 2025
Posted:
17 June 2025
You are already at the latest version
Abstract
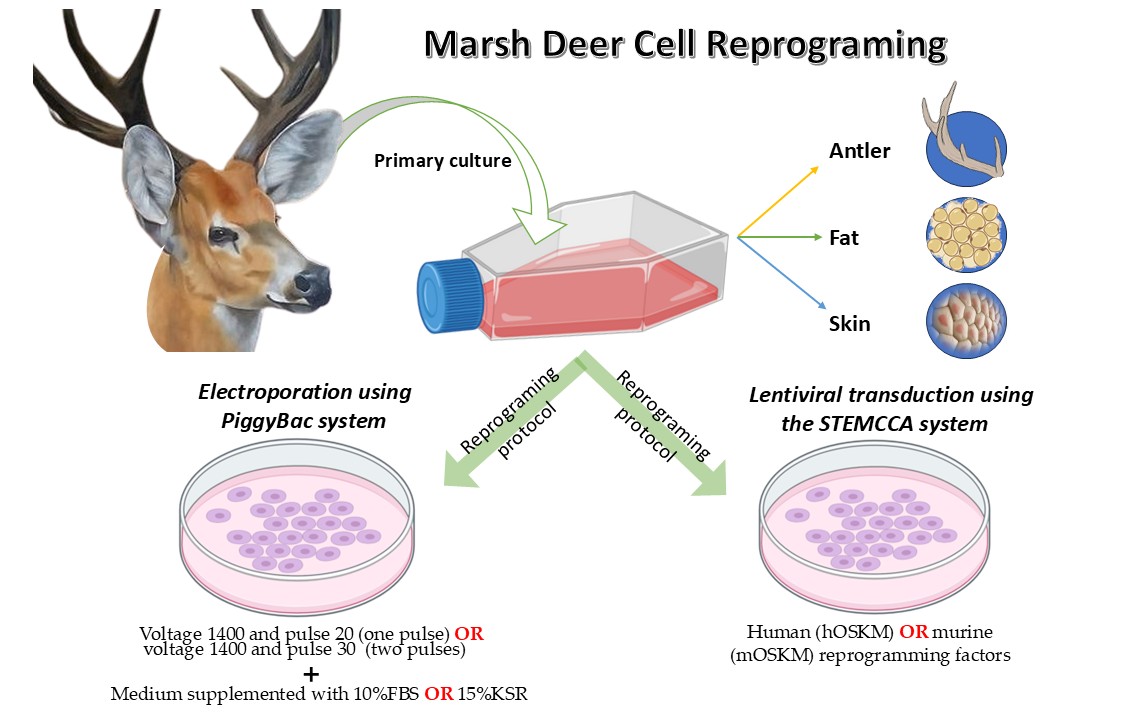
The capability to generate induced pluripotent stem cells (iPSCs) from adult somatic cells, enabling them to differentiate into any cell type has been demonstrated in several studies. In humans and mice, iPSCs have been shown to differentiate into primordial germ cells (PGCs), spermatozoa, and oocytes. However, research on iPSCs in deer is novel. Despite the necessity for establishing germplasm banks from endangered cervid species, the collection and cryopreservation of gametes and embryos have proven complex for this group. Therefore, the focus of this study was to establish protocols for deriving stable iPSC lines from Blastocerus dichotomus (Marsh deer) using primary cells derived from antler, adipose tissue, or skin, with the ultimate goal of producing viable gametes in the future. To achieve this, two main reprogramming approaches were tested: 1) transfection using PiggyBac transposons (plasmid PB-TET-MKOS) delivered via electroporation, and 2) lentiviral transduction using the STEMCCA system with either human (hOSKM) or murine (mOSKM) reprogramming factors. Both systems utilized murine embryonic fibroblasts (MEFs) as feeder cells. The PiggyBac system was further supplemented with a culture medium containing small molecules to aid reprogramming, including a GSK inhibitor, MEK inhibitor, ALK/TGF inhibitor, and thiazovivin. Initial colony formation was observed; however, these colonies failed to expand post-selection. Regardless of the reprogramming method employed, a low percentage of GFP-positive cells was detected, indicating an additional barrier to reprogramming cell lineages in this species. Despite these challenges, important insights were gained that will inform and guide future studies toward the successful generation of iPSCs in deer.
Keywords:
cervids
; electroporation
; genetic diversity
; lentiviral transduction
; PiggyBac transposons
; wild animals
1. Introduction
Most Neotropical deer species are experiencing population declines, rendering their conservation status markedly more critical than that of other mammalian taxa globally. While approximately 25% of all mammal species are currently classified as threatened and 15% as Data Deficient (DD) [1], this situation is more acute for Neotropical deer: 53% are categorized under a threat level, and 17.6% are classified as DD due to insufficient comprehensive studies to ascertain their true conservation status [2]. Declining wild populations typically face reduced genetic diversity and increased rates of consanguineous mating (inbreeding), thereby elevating their extinction risk [3,4].
The marsh deer (Blastocerus dichotomus), one of South America's largest terrestrial mammals, has suffered a reduction of approximately 65% in its natural habitat over the past four decades [5]. Moreover, the species is potentially extinct in countries such as Uruguay and Peru [6,7,8]. Beyond habitat loss and fragmentation, other anthropogenic pressures, including close proximity to domestic livestock (which facilitates pathogen transmission) and hunting [9], have exacerbated the decline of marsh deer populations. Consequently, B. dichotomus is classified as Vulnerable (VU) on the International Union for Conservation of Nature Red List [2].
In this context, the establishment of genome resource banks (GRBs) as repositories of genetic material such as semen, oocytes, embryos, and somatic cells (e.g., fibroblasts), alongside the application of assisted reproductive technologies (ARTs), presents promising strategies for conserving endangered species. These approaches can enhance reproductive management, preserve genetic variability, and facilitate more equitable genetic contributions among individuals [10,11]. Nevertheless, the acquisition of gametes from wild animals and the effective implementation of ARTs in these populations pose significant challenges. Furthermore, creating GRBs that comprehensively capture the extant genetic diversity of such populations remains a complex and demanding task.
The relative ease with which tissues and cells can be acquired and cryopreserved renders them a highly promising source of biological material. A particularly significant development in this context is the utilization of induced pluripotent stem cells (iPSCs). These cells are generated by reprogramming somatic cells to achieve a state of pluripotency analogous to that of embryonic stem cells (ESCs). Takahashi and Yamanaka [12] pioneered this methodology, successfully producing iPSCs through the induced expression of four key exogenous transcription factors: Oct4, Sox2, KLF4, and c-Myc. These factors act by modifying the endogenous transcriptional network of the cells, thereby facilitating both the establishment and subsequent maintenance of the pluripotent state.
Advancements in stem cell technology have introduced alternative methodologies wherein induced pluripotent stem cells (iPSCs) can be differentiated into functional gametes (oocytes, sperm, or spermatids) and, following fertilization, yield healthy animals [13,14]. The utilization of iPSCs for generating viable gametes offers significant advantages, including: Capacity for a single tissue fragment to serve as a virtually inexhaustible source of material, even post-mortem; Eliminating the need for repeated invasive procedures or chemical/physical restraints typically required for gamete and embryo retrieval, as somatic cells can be readily collected and cultured; Facilitates the collection of genetic material from free-ranging animals without necessitating the development of complex hormonal protocols or specialized gamete and embryo extraction methods; Supports the development and application of reproductive biotechnologies for embryo production; Enables the use of cells from juvenile animals that have not yet reached sexual maturity and are thus unable to produce gametes, or from senescent individuals whose gamete quality may be compromised; Mitigates the challenges associated with cryopreserving oocytes and embryos, which exhibit greater susceptibility to cryoinjuries compared to the more resilient iPSCs [15].
The efficiency of cellular reprogramming exhibits considerable variation across different cell types, being demonstrably lower in terminally differentiated cells compared to less-differentiated or undifferentiated adult progenitor cells [16]. Consequently, the selection of an appropriate cell source is a critical determinant in the development of iPSC lines, necessitating careful consideration of factors such as reprogramming amenability, tissue accessibility, and the facility of in vitro culture and maintenance.
In cervids, antler-derived stem cells present a unique and compelling opportunity [17]. Antlers, representing a rare instance of complete organ regeneration in mammals, serve as an invaluable model for investigating mechanisms of scar-free wound healing, resistance to tumorigenesis despite rapid cell proliferation, and potential therapeutic strategies for promoting limb regeneration [18]. These osseous structures, which undergo periodic regeneration from proliferating stem cell populations, typically originate from the frontal bone (pedicles) of adult male deer; however, in reindeer (Rangifer tarandus), antlers are present in both sexes [19]. Previous investigations in Cervus elaphus and Dama dama have demonstrated that the pedicle periosteum harbors cells expressing a panel of established stem cell markers (e.g., CD9, OCT-4, Nanog, STRO-1) [20,21]. Nevertheless, to date, the derivation of iPSC lines from this specific tissue source has not been reported.
Adipose tissue-derived stem cells (ADSCs) represent another promising avenue, as several studies have identified these cells as a highly suitable somatic cell source for the generation of induced pluripotent stem cells (iPSCs) [22]. ADSCs exhibit robust proliferative capacity and have demonstrated superior efficiency in iPSC line generation when compared to other cell lineages, such as mouse embryonic fibroblasts or adult neural stem cells. Furthermore, dermal tissue (skin) constitutes a readily accessible source of biological material [23,24]. Significantly, both skin and adipose tissues can be collected from individuals irrespective of sex or developmental stage, often through minimally invasive procedures
The successful derivation of iPSCs has been achieved in an extensive range of species. This includes laboratory animals such as mice [12] and rats [25]; domestic species like dogs [26], pigs [27], rabbits [28], sheep [29], horses [30], cattle [31], buffalo [32], quail [33], chickens [34], and goats [35]. Significantly, this technology has also been applied to various wild species, including rhesus monkeys (Macaca mulatta) [36], white rhinos (Ceratotherium simum) [37], mandrills (Mandrillus leucophaeus) [37], snow leopards (Panthera uncia) [38], Bengal tigers (Panthera tigris) [39], servals (Leptailurus serval) [39], jaguars (Panthera onca) [39], bats (Myotis lucifugus) [40], among other mammals, birds, and even amphibian, reptiles and fish [41].
Despite these widespread advancements, there are currently no published reports on iPSC derivation in any cervid species. Given that approximately one-third of global deer populations are classified as at risk of extinction [2], the development of iPSC lines with the capacity to differentiate into viable gametes could represent a pivotal conservation strategy. Therefore, the objective of the present study is to reprogram somatic cells derived from antlers, adipose, and dermal tissues of B. dichotomus, to contribute to the conservation of this specific species and to establish a foundational model for iPSC generation applicable to other deer species.
2. Materials and Methods
This work was evaluated and approved by the Ethics and Animal Welfare Committee (CEBEA) of the Faculty of Agricultural and Veterinary Sciences (FCAV) of São Paulo State University (UNESP), Jaboticabal campus, under protocol Nº. 025986/13. The study complies with the ethical principles in animal experimentation adopted by the Brazilian College of Animal Experimentation (COBEA).
Animal Biopsies
Biopsies were collected from animals at the Marsh Deer Conservation Center (Centro de Conservação do Cervo-do-Pantanal – CCCP) in Promissão, SP, Brazil, and the Deer Research and Conservation Centre (Núcleo de Pesquisa e Conservação de Cervídeos – NUPECCE) at the Department of Animal Science, FCAV – UNESP, Jaboticabal, SP, Brazil. The animals were anesthetized using a combination of 7.0 mg kg⁻¹ ketamine hydrochloride (Vetaset® – Fort Dodge – Brazil) and 1 mg kg⁻¹ xylazine (Rompum® – Bayer – Brazil) administered via intramuscular injection using anesthetic darts.
Antler biopsies were obtained from three males, with samples collected 45 to 60 days after antler growth began, approximately 0.5 cm below the tip. For skin biopsies, tissues were sampled from two females and one male; the samples were dissected from the proximal region of the pelvic limb, which had been previously shaved and disinfected with 70% alcohol, using sterile scissors and tweezers. Additionally, adipose tissue biopsies were taken from three females from the lateral area of the ear, just below the cartilage. This alternative site was selected due to the difficulty of safely obtaining fat from more commonly used regions, such as the caudal or abdominal areas, which are commonly used in other species [42,43,44].
Isolation of Somatic Cells
After collection, tissues were stored in a cryovial containing McCoy's solution (M4892, Sigma) with high concentrations of antibiotic (500mg L-1 of gentamicin and 20 mg L-1 of amphotericin B) and kept refrigerated on ice inside a Styrofoam box. Upon arrival at the laboratory, tissues were washed four times in PBS and mechanically separated into pieces of about 2 mm2 using a scalpel blade. After mechanical breakdown, cells were enzymatically digested with collagenase I (200 U ml-1, Sigma) inside an incubator at 37 °C for a period of 3 h for antler and skin, and 30 min for fat. After enzymatic digestion, cells were centrifuged for 5 min at 200 G and the pellet was suspended in 10 ml of complete culture medium composed by Dulbecco's modified Eagle's minimal essential medium (DMEM) with high glucose levels (4.5 g L-1), glutamine (0.6 g L-1), 1% essential amino acids, fetal bovine serum (FBS) in a proportion of 10% v/v, 50 mg L-1 of gentamicin sulfate and 2 mg l-1 amphotericin B. The sample was centrifuged again for 5 min at 200g and the pellet was resuspended in 5 ml of complete culture medium containing 10% of FBS. The medium exchange was performed 48 hours after the initial plating to remove non-adherent cells. For the maintenance of the culture, the medium was exchanged twice a week using complete culture medium supplemented with 10% fetal bovine serum (FBS).
Cells were cultured until they reached about 80% confluence and, on reaching this threshold, the cells were detached using trypsin 0.25%, centrifuged, and reseeded into new flasks. Cells of the second or third passage were cryopreserved in a medium composed of 80% FBS, 10% dimethyl sulfoxide (DMSO), 10% DMEM, 100 units/ml of penicillin and 100 ug/ ml streptomycin, using the slow cooling method (4 hours at 4 °C, 30 minutes in nitrogen steam and subsequently immersed in liquid nitrogen) and stored in cryogenic cylinders. At the time of performing the experiments, cells were thawed in a water bath at 37 °C and cultured. Cells of the fourth and fifth passage were used in further experiments.
Pluripotency Induction
Electroporation Using the PiggyBac System
The electroporation protocol used for the derivation of iPSC was similar to that previously used for the derivation of equine iPSC by Nagy and collaborators [30]. Therefore, to generate iPSC in Marsh Deer, we use the system based on transfections with transposon PiggyBac. The plasmids PB-TET- OSKM (excisable polycistronic vector with murine gene sequence), PBCAG-rtTA, pCyL43 PBase and PB-GFP were performed as previously described by Nagy and collaborators [30].
The cells were transfected using the electroporator device Neon® (Invitrogen), according to the manufacturer's instructions. For electroporation, we used 5x104 cells with a combination of DNA with equal proportions of the four plasmids (0,25 µg of each) for each well in 6-well plates. These plates were pre-gelatinized (0.1% gelatin) and plated with murine embryonic fibroblasts (MEF) inactivated by mitomycin C for 3 hours to serve as "feeders" of the transfected cells.
After transfection, cells were placed in medium composed by DMEM (Invitrogen, #11960-044), 2mM glutamax (Invitrogen), 0.1mM β-mercaptoethanol, 0.1mM non-essential amino acids, 1mM sodium pyruvate, 50U/ml penicillin/streptomycin, 1000U/ml leukemia inhibitory fator, 10ng/ml bFGF, 5µg/ml insulin, 1.5µg/ml doxycycline, 3µM GSK inhibitor (Stemgent #CHIR99021), 0,5µM MEK inhibitor (Stemgent #PD0325901), 2.5µM ALK/TGF inhibitor (Stemgent #A83-01), 1mM thiazovivin and contained either 10% FBS or 15% knockout serum replacement (KSR), depending on the tested group. We also tested two electroporation programs (see below), using protocol 1 that consists of pulse voltage equal 1400, pulse width equal 20, number of pulses equal 1, and protocol 2 that consists of pulse voltage equal 1400, pulse width equal 30 and number of pulses equal 2.
The plasmid PB-GFP, which encodes the green fluorescent protein (GFP), served as a reporter gene, enabling the quantification of cells that successfully integrated the plasmid. Protocol 1, employed in this study, was previously validated for iPSC derivation in equine species [30]. In contrast, Protocol 2 was specifically optimized for the cell type under investigation in this experiment through systematic evaluation of 24 pre-existing electroporation programs on the Neon® Transfection System. These optimization tests were conducted exclusively with the PB-GFP plasmid to accurately assess the efficiency of plasmid integration into the cells.
Cells were allocated to distinct experimental groups based on the electroporation protocol and culture medium composition: (G1) Protocol 1 with medium supplemented with Fetal Bovine Serum (FBS); (G2) Protocol 1 with medium supplemented with KnockOut™ Serum Replacement (KSR); (G3) Protocol 2 with medium supplemented with FBS; and (G4) Protocol 2 with medium supplemented with KSR. Following transfection, the cell culture plates were maintained for a minimum of 35 days to monitor for the emergence of colonies.
3. Results and Discussion
A total of twelve attempts to induce pluripotency via electroporation were conducted in six-well plates, each containing five wells with transfected cells and one negative control. These attempts were distributed as follows: five using Protocol 1 with FBS-containing medium (three for antler-derived cells, one for adipose-derived cells, and one for skin-derived cells); three using Protocol 2 with FBS-containing medium (two for antler-derived cells and one for adipose-derived cells); one using Protocol 1 with KSR-containing medium (antler-derived cells); and three using Protocol 2 with KSR-containing medium (two for antler-derived cells and one for adipose-derived cells).
Initial attempts to induce pluripotency utilized Protocol 1, as previously described for equine cells [30], which yielded approximately 40% GFP-positive cells. However, due to the absence of colonies that could be expanded into stable iPSC lines and a lower transfection efficiency (GFP-positive cells) compared to that observed in horses (approximately 90%), protocol optimization was undertaken. The implementation of Protocol 2 resulted in an approximate 20% increase in GFP-positive cells, reaching around 60%.
Following these attempts with Protocol 2, which still did not yield iPSC lines, FBS was replaced with KnockOut Serum Replacement (KSR). Previous studies have demonstrated that KSR can significantly enhance reprogramming efficiency and may even facilitate the conversion of partially reprogrammed iPSCs into a fully reprogrammed state [46,47]. Conversely, FBS used during the reprogramming phase can exert deleterious effects, such as altering the activities of H3K9 methyltransferases and demethylases, due to the presence of Bone Morphogenetic Protein (BMP) family growth factors in its composition [48].
KSR contains components such as thiamine and vitamin C, which are known to favor reprogramming. Vitamin C, in particular, has been extensively studied in the context of iPSC reprogramming. It possesses antioxidant properties [49], prevents the silencing of the Dlk1-Dio3 locus (which can be compromised by FBS, thereby impairing the generation of fully reprogrammed iPSCs from adult cells) [50], is linked to active DNA demethylation [51], and influences other epigenetic mechanisms [52]. Its application has led to improved iPSC quality in murine models, as assessed by the tetraploid complementation assay, an improvement attributed to a reduction in DNA methylation aberrations [50].
Despite these efforts and protocol modifications, stable iPSC lines could not be generated. Approximately 20 days post-transfection, emergent colonies were observed (Figure 1), some exhibiting irregular morphology (Figure 1A), with most being GFP-positive. These colonies were manually selected, dissected into four pieces using a scalpel blade, and transferred to new plates with feeder cells. However, no subsequent cell growth was observed after selection and passage. It is plausible that these cells underwent only partial reprogramming or became refractory to complete reprogramming due to various factors, including an inability to undergo mesenchymal-to-epithelial transition (MET), aberrant activation of differentiation-associated genes, or a failure to sustain pluripotency gene expression [53].
Multiple studies confirm that the somatic cell source significantly influences the efficiency and fidelity of reprogramming into induced pluripotent stem cells (iPSCs) [54,55,56,57]. Despite this, we were unable to establish iPSC lines from any of the Marsh deer tissue sources investigated. The protocol employed in this study mirrored that used by Nagy and collaborators [30] for the derivation of equine iPSCs. Notably, Nagy and collaborators [30] successfully reprogrammed cells only when using fetal fibroblasts; application of the same protocol to adult fibroblasts did not yield iPSC lines (Smith, personal communication). However, obtaining fetal fibroblasts from Marsh deer presents significant logistical challenges and is considered an impractical strategy for establishing germplasm banks.
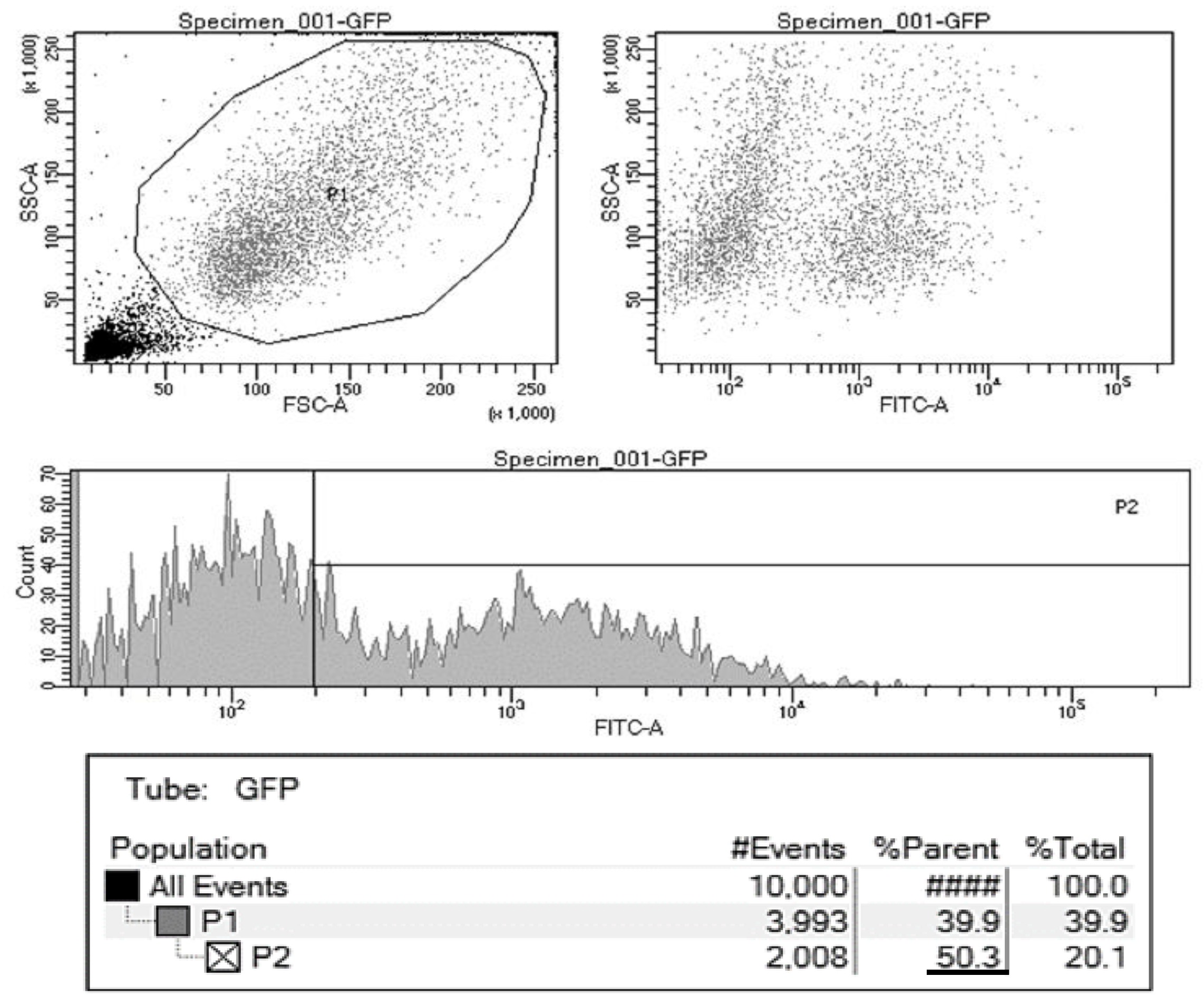
For attempts utilizing the STEMCCA lentiviral vector system, flow cytometry analysis of GFP reporter gene expression revealed results consistent with those previously observed using PiggyBac electroporation, with approximately 50% of cells expressing the GFP protein (Figure 2). In this lentiviral approach, each sample type (two skin, one antler, and one adipose-derived) was tested in a six-well plate format. Within each plate, three wells were dedicated to transduction with human OSKM factors, and three wells were used for murine OSKM factors. While previous studies have indicated that bovine cells reprogram more successfully with murine factors [45] and equine cells show a preference for human factors [58], no colony formation was observed in our Marsh deer cell experiments, irrespective of whether human or murine OSKM vectors were employed.
The challenges encountered in generating deer iPSCs were expected, given the well-documented difficulties in establishing pluripotent cell lines from domestic ungulates. Success in obtaining robust embryonic stem cell (ESC) lines from these species has been limited [59,60]. Similarly, for iPSCs, numerous studies report significant obstacles. These include failures in silencing exogenous transgene expression post-reprogramming, difficulties in maintaining cell cultures for extended periods, and the incomplete characterization of derived lines, often failing to meet all pluripotency criteria, particularly in vivo validation. Replicating culture conditions successfully employed for murine or human cells has not yielded comparable results in domestic ungulates, and identifying the optimal set of reprogramming factors for these species remains a major challenge [61].
A critical consideration in iPSC generation is the choice of exogenous reprogramming factors. Notably, iPSCs generated using the canonical Yamanaka factors (Oct4, Sox2, c-Myc, Klf4, commonly referred to as OSKM) exhibit higher rates of aberrant DNA methylation compared to cells derived via somatic cell nuclear transfer [62]. This observation suggests that OSKM factors alone may be insufficient to induce complete and faithful reprogramming. Consequently, an efficient and reliable strategy for iPSC generation might involve the incorporation of additional or alternative expression factors. Supporting this, Wang and collaborators [63] demonstrated accelerated reprogramming of both murine and human fibroblasts by co-expressing OSKM factors with two nuclear hormone receptors, Rarc and Lrh1. This approach significantly shortened the reprogramming timeline from the typical 3-4 weeks to 4-5 days. Further evidence comes from Nie and collaborators [64], who observed no colony formation when attempting to reprogram mature murine adipocytes using only OSKM factors delivered via the PiggyBac system. However, upon employing the six-factor combination (OSKM plus Lrh1 and Rarc) as described by Wang and collaborators [63], colonies began to emerge from day 16 onwards, with 86% of these successfully expanding into stable cell lines. The supplementary factors Lrh1 and Rarc are known to upregulate the expression of key pluripotency genes Oct4 and Nanog, thereby altering reprogramming kinetics and improving the efficiency of achieving a fully reprogrammed state.
In several species, OSKM alone have proven insufficient to induce pluripotency, necessitating the inclusion of additional factors. For instance, in cattle [31], wild felids such as the snow leopard [38], Bengal tiger, serval, and jaguar [39], as well as in the drill, Northern White Rhinoceros [37], and Ryukyu spiny rat [65], the addition of NANOG to the retroviral induction cocktail was required. Consistent with observations in our experiment, some of these studies reported initial colony formation using OSKM factors; however, these colonies could not be sustained through subsequent passages. Following the inclusion of NANOG, large, circular colonies were successfully generated, expanded over multiple passages, and established as iPSC lines, demonstrating pluripotency both in vitro and, in some in vivo assays.
Similarly, Talluri and collaborators [66], employing the PiggyBac transposon system for bovine fetal fibroblasts, failed to generate iPSC lines using only OSKM factors, requiring the co-expression of both NANOG and LIN28 for successful reprogramming. These same factors (OSKM, NANOG, and LIN28) were also essential for reprogramming Northern White Rhinoceros cells via electroporation-mediated transduction [67]. West and collaborators [68] utilized this expanded cocktail to produce porcine iPSCs, and notably, these lines demonstrated the capacity to contribute to chimeric offspring. In avian species, such as the Okinawa rail and Blakiston’s fish owl, reprogramming with OSKM factors delivered via PiggyBac transposase transduction was achieved using a modified OCT3/4 (M3O), plus the addition of NANOG, LIN28, and KLF2 [69].
However, the necessity to add or modify exogenous reprogramming factors is not attributable to a single determinant, as this requirement varies depending on the cell type, reprogramming methodology, culture medium composition, and other variables. For example, Kawaguchi and collaborators [70] successfully generated stable bovine iPSC lines from adipose-derived cells using the PiggyBac system with only OSKM factors. These lines were capable of contributing to the inner cell mass of host blastocysts and forming chimeric fetuses without the need for exogenous NANOG. However, when embryonic fibroblasts were transfected under identical conditions, the emergent colonies failed to establish stable cell lines capable of sustained propagation.
Another noteworthy observation is the apparently greater reprogramming efficiency achieved when employing two separate vectors, one encoding Oct4, Klf4, and Sox2 (OKS), and another encoding c-Myc, compared to a single polycistronic vector expressing all four factors (OSKM) [71]. In the aforementioned study on cattle by Kawaguchi and collaborators [70], it was reported that transfection with a single OSKM vector failed to yield colonies from adipose-derived cells. In contrast, co-transfection with separate OKS and c-Myc vectors resulted in the generation of iPSC lines with an efficiency of approximately 0.01%. Consequently, many specific aspects contribute to the successful generation of stable and reliable iPSC lines. This multiplicity of factors influencing cellular reprogramming, coupled with the gaps in fundamental biological knowledge regarding wild animals, highlights the considerable challenge associated with generating induced pluripotent stem cell (iPSC) lines for these species. Consequently, extensive testing and research are imperative before the ultimate goal can be achieved, particularly when considering the maximization of germplasm utilization from endangered species. Future studies may deepen our understanding of the mechanisms and optimize protocols for producing stable and reliable iPSC lines in cervid species.
4. Conclusions
To our knowledge, this study represents the first report in the literature on the utilization of antler-derived cells for cellular reprogramming to a pluripotent state. Although stable iPSC colonies were not obtained, significant advancements were made, and important obstacles were demonstrated. The initial stages, including the achievement of marsh deer cells, and the exploration of diverse methodological approaches (encompassing variations in culture medium with FBS or KSR, the use of transposon-based or lentiviral vectors, and different genetic material delivery methods such as electroporation or lentiviral transduction), form an essential foundation for refining future studies and achieving more advanced outcomes. Furthermore, an additional barrier to reprogramming in this species was clearly demonstrated, characterized by resistance to vector entry into the cells, as evidenced by GFP reporter gene expression levels. Consequently, future investigations can build upon the findings of this work to propose optimizations, such as the employment of different exogenous transcription factors and alternative culture medium supplementation.
Author Contributions
Conceptualization, L.D.R.; L.C.S, J.M.B.D. and F.F.B.; Methodology, L.D.R.; L.C.S and F.F.B. formal analysis, L.D.R. and J.T.; investigation, L.D.R. e F.F.B.; resources, J.M.B.D, L.C.S. and F.F.B..; writing—original draft preparation L.D.R.; writing—review and editing, L.D.R, F.F.B., E.D.P.S., J.T., J.M.B.D and L.C.S; visualization, L.D.R, F.F.B., E.D.P.S., J.T., J.M.B.D and L.C.S; supervision, J.M.B.D and L.C.S.. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP) process n◦ 2013/13972-0 and 2014/02439-2.
Institutional Review Board Statement
The animal study protocol was approved by the Ethics and Animal Welfare Committee (CEBEA) of the Faculty of Agricultural and Veterinary Sciences (FCAV) of São Paulo State University (UNESP), Jaboticabal campus, under protocol Nº. 025986/13 approved 04/12/2013.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Acknowledgments
We would like to thank the collaboration of the other members of the Deer Research and Conservation Center, the laboratory technician João Boer for helping with the equipment and preparing solutions, and the veterinarians Eveline Zanetti dos Santos and Juliana Linardi from the Pantanal Deer Conservation Center (CCCP) and the entire team of caretakers, who kept the animals healthy and in high welfare.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Schipper, J.; Chanson, J.S.; Chiozza, F.; Cox, N.A.; Hoffmann, M.; Katariya, V.; Lamoreux, J.; Rodrigues, A.S.L.; Stuart, S.N.; Temple, H.J.; et al. The status of the world’s land and marine mammals: Diversity, threat, and knowledge. Science 2008, 322, 225–230. [Google Scholar] [CrossRef] [PubMed]
- The IUCN Red List of Threatened Species. Available online: https://www.iucnredlist.org/ (accessed on 31 March 2025).
- Frankham, R.; Ballou, J.D.; Briscoe, D.A. A Primer of Conservation Genetics; Cambridge University Press: Cambridge, UK, 2004; p. 220. [Google Scholar]
- Keller, L.F.; Biebach, I.; Ewing, S.R.; Hoeck, P.E.A. The genetics of reintroductions: Inbreeding and genetic drift. In Reintroduction Biology: Integrating Science and Management; Ewen, J.G., Armstrong, D.P., Parker, K.A., Seddon, P.J., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2012; pp. 360–394. [Google Scholar]
- Weber, M.; Gonzalez, S. Latin American deer diversity and conservation: A review of status and distribution. Écoscience 2003, 10, 443–454. [Google Scholar] [CrossRef]
- Piovezan, U.; et al. Marsh deer Blastocerus dichotomus (Illiger, 1815). In Neotropical Cervidology: Biology and Medicine in Latin American Deer; Duarte, J.M.B., González, S., Eds.; Funep/IUCN: Jaboticabal, Brazil, 2010; pp. 66–76. [Google Scholar]
- González, S.; Lessa, E.P. Historia de la mastozoología en Uruguay. In Historia de la Mastozoología en Latinoamérica, las Guayanas y el Caribe; Ortega, J., Martínez, J.L., Tirira, D.G., Eds.; Editorial Murciélago Blanco y Asociación Ecuatoriana de Mastozoología: Quito, Ecuador; México D.F., Mexico, 2014; pp. 381–404. [Google Scholar]
- González, S.; Aristimuño, M.P.; Moreno, F. New record in Uruguay of the marsh deer (Blastocerus dichotomus Illiger, 1815) redefines its southern geographic distribution area. Front. Ecol. Evol. 2024, 12, 1419234. [Google Scholar] [CrossRef]
- González, S.; Duarte, J.M.B. Speciation, evolutionary history and conservation trends of Neotropical deer. Mastozool. Neotrop. 2020, 27, 37–47. [Google Scholar] [CrossRef]
- Holt, W.V.; Pickard, A.R. Role of reproductive technologies and genetic resource banks in animal conservation. Rev. Reprod. 1999, 4, 143–150. [Google Scholar] [CrossRef]
- Wildt, D.E. Genetic resource banks for conserving wildlife species: Justification, examples and becoming organized on a global basis. Anim. Reprod. Sci. 1992, 28, 247–257. [Google Scholar]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, K.; Ohta, H.; Kurimoto, K.; Aramaki, S.; Saitou, M. Reconstitution of the mouse germ cell specification pathway in culture by pluripotent stem cells. Cell 2011, 146, 519–532. [Google Scholar] [CrossRef]
- Hayashi, K.; Ogushi, S.; Kurimoto, K.; Shimamoto, S.; Ohta, H.; Saitou, M. Offspring from oocytes derived from in vitro primordial germ cell-like cells in mice. Science 2012, 338, 971–975. [Google Scholar] [CrossRef]
- Rola, L.D.; Buzanskas, M.E.; Melo, L.M.; Chaves, M.S.; Freitas, V.J.F.; Duarte, J.M.B. Assisted reproductive technology in Neotropical deer: A model approach to preserving genetic diversity. Animals 2021, 11, 1961. [Google Scholar] [CrossRef]
- Lee, G.; Papapetrou, E.P.; Kim, H.; Chambers, S.M.; Tomishima, M.J.; Fasano, C.A.; Ganat, Y.M.; Menon, J.; Shimizu, F.; Viale, A. Modeling pathogenesis and treatment of familial dysautonomia using patient-specific iPSCs. Nature 2009, 461, 402–406. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Berg, D.; Ba, H.; Sun, H.; Wang, Z.; Li, C. Deer antler stem cells are a novel type of cells that sustain full regeneration of a mammalian organ—Deer antler. Cell Death Dis. 2019, 10, 443. [Google Scholar] [CrossRef] [PubMed]
- Colitti, M.; Allen, P.; Price, J.S. Programmed cell death in the regenerating deer antler. J. Anat. 2005, 207, 339–351. [Google Scholar] [CrossRef]
- Kierdorf, U.; Kierdorf, H. Deer antlers—A model of mammalian appendage regeneration: An extensive review. Gerontology 2011, 57, 53–65. [Google Scholar] [CrossRef]
- Li, C.; Yang, F.; Sheppard, A. Adult stem cells and mammalian epimorphic regeneration—Insights from studying annual renewal of deer antlers. Curr. Stem Cell Res. Ther. 2009, 4, 237–251. [Google Scholar]
- Rolf, H.J.; Kierdorf, U.; Kierdorf, H.; Schulz, J.; Seymour, N.; Schliephake, H.; Napp, J.; Niebert, S.; Wolfel, H.; Wiese, K.G. Localization and characterization of STRO-1 cells in the deer pedicle and regenerating antler. PLoS ONE 2008, 3, e2064. [Google Scholar] [CrossRef]
- Tat, P.A.; Sumer, H.; Jones, K.L.; Upton, K.; Verma, P.J. The efficient generation of induced pluripotent stem (iPS) cells from adult mouse adipose tissue-derived and neural stem cells. Cell Transplant. 2010, 19, 525–536. [Google Scholar] [CrossRef] [PubMed]
- Coleman, S.R. Long-term survival of fat transplants: Controlled demonstrations. Aesthetic Plast. Surg. 1995, 19, 421–425. [Google Scholar] [CrossRef] [PubMed]
- Yarak, S.; Okamoto, O.K. Human adipose-derived stem cells: Current challenges and clinical perspectives. An. Bras. Dermatol. 2010, 85, 647–656. [Google Scholar] [CrossRef]
- Liao, J.; Cui, C.; Chen, S.; Ren, J.; Chen, J.; Gao, Y.; Li, H.; Jia, N.; Cheng, L.; Xiao, L. Generation of induced pluripotent stem cell lines from adult rat cells. Cell Stem Cell 2009, 4, 11–15. [Google Scholar] [CrossRef]
- Shimada, H.; Nakada, A.; Hashimoto, Y.; Shigeno, K.; Shionoya, Y.; Nakamura, T. Generation of canine induced pluripotent stem cells by retroviral transduction and chemical inhibitors. Mol. Reprod. Dev. 2009, 77, 2. [Google Scholar] [CrossRef] [PubMed]
- Ezashi, T.; Telugu, B.P.; Alexenko, A.P.; Sachdev, S.; Sinha, S.; Roberts, R.M. Derivation of induced pluripotent stem cells from pig somatic cells. Proc. Natl. Acad. Sci. USA 2009, 106, 10993–10998. [Google Scholar] [CrossRef]
- Táncos, Z.; Nemes, C.; Varga, E.; Bock, I.; Rungarunlert, S.; Tharasanit, T.; et al. Establishment of a rabbit induced pluripotent stem cell (RbiPSC) line using lentiviral delivery of human pluripotency factors. Stem Cell Res. 2017, 21, 16–18. [Google Scholar] [CrossRef] [PubMed]
- Bao, L.; He, L.; Chen, J.; Wu, Z.; Liao, J.; Rao, L.; et al. Reprogramming of ovine adult fibroblasts to pluripotency via drug-inducible expression of defined factors. Cell Res. 2011, 21, 600–608. [Google Scholar] [CrossRef] [PubMed]
- Nagy, K.; Sung, H.K.; Zhang, P.; Laflamme, S.; Vincent, P.; et al. Induced pluripotent stem cell lines derived from equine fibroblasts. Stem Cell Rev. 2011, 7, 693–702. [Google Scholar] [CrossRef]
- Ezashi, T.; Yuan, Y.; Roberts, R.M. Pluripotent stem cells from domesticated mammals. Annu. Rev. Anim. Biosci. 2016, 4, 223–253. [Google Scholar] [CrossRef]
- Macedo, G.G.; Costa, D.S.; Silva, S.E.M.; Alves, A.L.G.; Oliveira, M.F.; Martins, D.D.S.; et al. Induced pluripotent stem cells from large animals. Anim. Reprod. 2020, 17, e20200032. [Google Scholar]
- Manohar, M.; Lagutina, I.; Fulka, H.; Sung, L.-Y.; Eid, L.; Lazzari, G.; et al. Reprogramming of somatic cells in the sheep: Recent advances and future perspectives. Theriogenology 2016, 86, 109–117. [Google Scholar]
- Durnaoglu, S.; Genc, S.; Genc, K. Patient-specific pluripotent stem cells in neurological diseases. Stem Cells Int. 2011, 2011, 212487. [Google Scholar] [CrossRef]
- Carlin, R.; Davis, D.; Weiss, M.; Schultz, B.; Troyer, D. Expression of early transcription factors Oct-4, Sox-2 and Nanog by porcine umbilical cord (PUC) matrix cells. Reprod. Biol. Endocrinol. 2006, 4, 8. [Google Scholar] [CrossRef]
- Munoz, M.; Rodriguez, A.; De Frutos, C.; Caamaño, J.N.; Díaz, E.; Ikeda, S.; et al. Conventional and delayed embryo transfer in cattle—In vitro and in vivo embryo development and quality. Theriogenology 2015, 83, 467–476. [Google Scholar]
- Keefer, C.L. Artificial cloning of domestic animals. Proc. Natl. Acad. Sci. 2015, 112, 8874–8878. [Google Scholar] [CrossRef] [PubMed]
- de Souza, A.F.; Oba, E.; Milazzotto, M.P. Contribution of mitochondria to the study of oocyte and embryo quality. Anim. Reprod. 2022, 19, e20220006. [Google Scholar]
- Hirao, Y. In vitro growth of oocytes from domestic species. Anim. Sci. J. 2011, 82, 110–118. [Google Scholar]
- Costa, L.F.; Paulini, F.; Silva, R.C.; Andrade, E.R.; Lucci, C.M. Cryopreservation of ovarian tissue from white-tailed deer (Odocoileus virginianus): Effect of different concentrations of EG and DMSO. Theriogenology 2018, 120, 107–113. [Google Scholar]
- Comizzoli, P.; Songsasen, N.; Wildt, D.E. Protecting and extending fertility for females of wild and endangered mammals. Cancer Treat. Res. 2010, 156, 87–100. [Google Scholar]
- Comizzoli, P. Biobanking efforts and fertility preservation in wildlife species: Can they be improved? Asian J. Androl. 2015, 17, 640–645. [Google Scholar] [CrossRef]
- Barber, M.R.; Fayrer-Hosken, R.A. Possible mechanisms of mammalian immunocontraception. J. Reprod. Immunol. 2000, 46, 103–124. [Google Scholar] [CrossRef]
- Comizzoli, P.; Songsasen, N. Biobanking in wildlife: The end or a means to the end? Anim. Front. 2021, 11, 57–63. [Google Scholar]
- Tharasanit, T.; Comizzoli, P. Reproductive biotechnologies for endangered mammalian species. Vet. J. 2021, 269, 105604. [Google Scholar]
- Cox, J.F.; Alfaro, V. Interspecies somatic cell nuclear transfer: Advancements and future perspectives. Cell Reprogram. 2007, 9, 229–240. [Google Scholar]
- Comizzoli, P. Germplasm banks for conservation of wildlife species: Relevance of reproductive biotechnologies for preserving diversity. Theriogenology 2017, 109, 48–54. [Google Scholar]
- Songsasen, N.; Comizzoli, P.; Travis, A.J.; Wildt, D.E. Advances in reproductive technology and genome resource banking for wildlife species. Theriogenology 2012, 78, 165–183. [Google Scholar]
- Luvoni, G.C.; Chigioni, S.; Allievi, E.; Macis, D. Embryo technologies in dogs and cats. Theriogenology 2005, 64, 1665–1672. [Google Scholar]
- Galli, C.; Duchi, R.; Colleoni, S.; Lagutina, I.; Lazzari, G. Ovum pick-up, in vitro fertilization and embryo culture in cattle. Theriogenology 2007, 68, S59–S70. [Google Scholar]
- Comizzoli, P.; Holt, W.V. Breakthroughs and new thinking in gamete biology. Reprod. Domest. Anim. 2019, 54, 35–39. [Google Scholar]
- Campbell, M.L.; Venkatesh, D.; Ranganathan, V.; Comizzoli, P. Lessons from biobanking gametes and gonadal tissues for assisted reproduction in wildlife species. Cells 2022, 11, 2124. [Google Scholar]
- Luvoni, G.C. Conservation of endangered mammalian species through assisted reproduction: The role of in vitro embryo production. Anim. Reprod. 2020, 17, e20190062. [Google Scholar]
- Sato, M.; Hamatani, T. Concise review: Toward the generation of female gametes from induced pluripotent stem cells. Stem Cells 2020, 38, 6–13. [Google Scholar]
- Comizzoli, P. The crucial role of biobanking and fertility preservation as tools for biodiversity conservation and how to better organize and integrate them globally. Biopreserv. Biobank. 2021, 19, 260–268. [Google Scholar]
- Faunes, F.; Larrain, J. Conservation of pluripotency in embryonic and induced pluripotent stem cells. Stem Cells Int. 2012, 2012, 1–11. [Google Scholar]
- Schmidt, K.; Glage, S.; Neumann, A.; Schwinzer, R.; Dorsch, M.; Tychsen, B.; et al. An iPSC line from the endangered drill monkey (Mandrillus leucophaeus) for comparative stem cell studies. Stem Cell Res. 2020, 47, 101911. [Google Scholar]
- Verma, R.; Holland, M.K.; Temple-Smith, P.; Verma, P.J. Inducing pluripotency in somatic cells from the snow leopard (Panthera uncia), an endangered felid. Theriogenology 2012, 77, 220–228. [Google Scholar] [CrossRef] [PubMed]
- Verma, R.; Liu, J.; Holland, M.K.; Temple-Smith, P.; Williamson, M.; Verma, P.J. Derivation of induced pluripotent stem cells from the tiger (Panthera tigris), an endangered felid. Stem Cells Dev. 2013, 22, 169–176. [Google Scholar]
- Gouveia, C.; Huyser, C.; Egli, D.; Pepper, M.S. Lessons learned from somatic cell nuclear transfer. Int. J. Dev. Biol. 2020, 64, 453–465. [Google Scholar] [CrossRef]
- Figueiredo, M.A.P.; Silva, A.M.; Comizzoli, P.; Silva, A.R. Current advances in cryopreservation of wild carnivore semen for ex situ conservation. Theriogenology 2019, 130, 222–228. [Google Scholar]
- Sharma, R.; Bansal, A.K. Oxidative stress and antioxidant defence system in boar spermatozoa. J. Anim. Physiol. Anim. Nutr. 2020, 104, 759–768. [Google Scholar]
- Comizzoli, P. Biobanking efforts and fertility preservation in wildlife species: Can they be improved? Asian J. Androl. 2015, 17, 640–645. [Google Scholar] [CrossRef]
- Paris, M.C.J.; O’Brien, J.K.; Canfield, P.J.; Allen, W.R. Reproductive technologies and genetic management of wild equids: A review. Reprod. Fertil. Dev. 2005, 17, 661–671. [Google Scholar]
- Novakovic, S.; Vujovic, M.; Djordjevic, B.; Stanimirovic, Z. Challenges and perspectives in cryopreservation of reproductive cells and tissues of endangered animal species. Cryobiology 2017, 77, 1–5. [Google Scholar]
- Zhu, H.; Li, C.; Wang, D.; Lu, J.; Gao, Q.; Zhang, T. Advances in cryopreservation of mammalian oocytes and embryos: A review. Theriogenology 2018, 113, 90–96. [Google Scholar]
- Comizzoli, P. Biobanking in wildlife: Facing the frontiers of freeze-drying technologies for conservation purposes. Adv. Exp. Med. Biol. 2019, 1200, 389–403. [Google Scholar]
- Santiago-Moreno, J.; Esteso, M.C.; Castaño, C.; Toledano-Díaz, A.; López-Sebastián, A.; González-Bulnes, A. Cryopreservation of semen from endangered species. Anim. Reprod. Sci. 2012, 133, 209–215. [Google Scholar]
- Rall, W.F.; Fahy, G.M. Ice-free cryopreservation of mouse embryos at −196 °C by vitrification. Nature 1985, 313, 573–575. [Google Scholar] [CrossRef]
- Comizzoli, P.; Holt, W.V. Breakthroughs and new thinking in gamete biology. Reprod. Domest. Anim. 2019, 54, 35–39. [Google Scholar]
- Loux, S.C.; Grieger, D.M.; Newton, G.R.; Canisso, I.F. Development and application of sperm cryopreservation technologies in wildlife species. Anim. Reprod. Sci. 2020, 220, 106389. [Google Scholar]
Figure 1.
Photomicrography illustrating the colonies obtained using the protocol with PiggyBac transposons. (A) Irregular colony (100x magnification); (B) Regular colony (40x magnification).
Figure 1.
Photomicrography illustrating the colonies obtained using the protocol with PiggyBac transposons. (A) Irregular colony (100x magnification); (B) Regular colony (40x magnification).
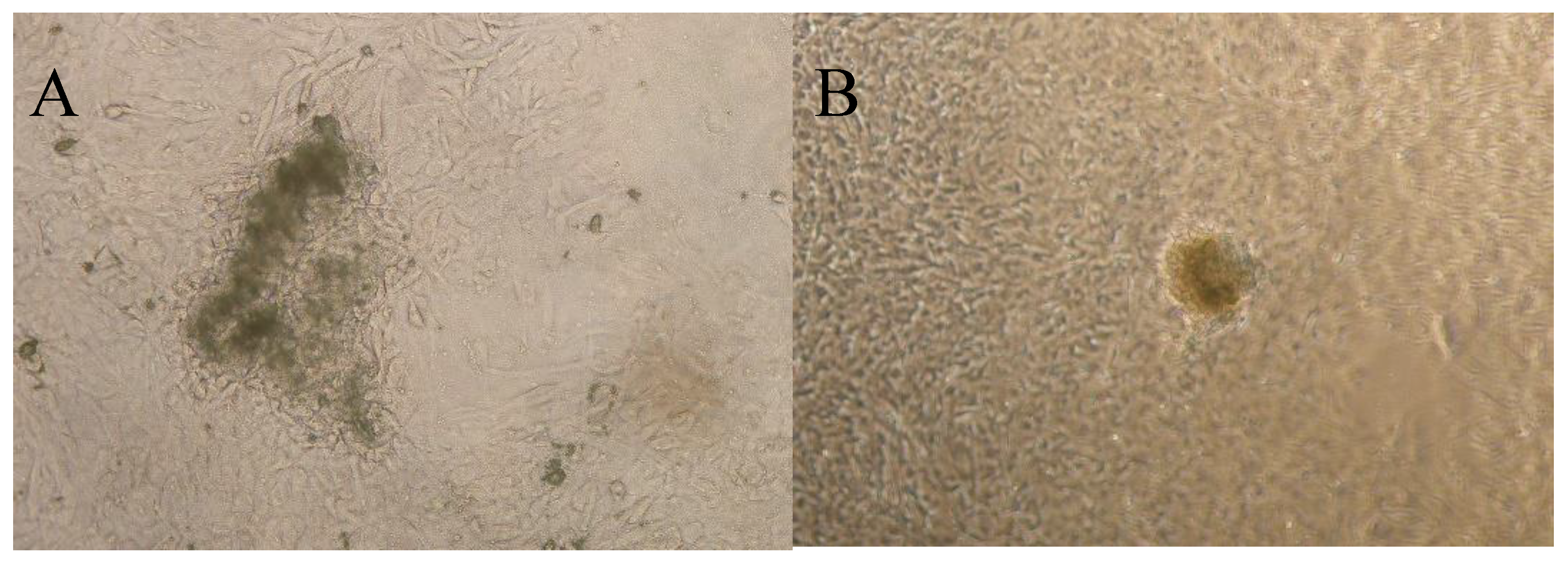
Figure 2.
Flow cytometry analysis demonstrated that 50.3% of cells exhibited GFP reporter gene expression.
Figure 2.
Flow cytometry analysis demonstrated that 50.3% of cells exhibited GFP reporter gene expression.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



