
Submitted:
16 June 2025
Posted:
16 June 2025
You are already at the latest version
Abstract
Pericon (Tagetes lucida) is an important medicinal and traditional plant used widely in ceremonies and considered as biocultural heritage. In Yautepec (Pericon Hill, in Nahuatl), Morelos, people report the extinction of this emblematic species. The extinction causes are until now not known. Nevertheless, the decline of populations may be attributable to a combination of factors, including alterations in land use, the utilisation of herbicides, habitat degradation, and overexploitation of resources. This decline could be exacerbated by the species ' inability to adapt to environmental changes. Genetic diversity is crucial to maintain the population’s adaptability in the face of diseases, environmental changes and other stressors and has an impact on population persistence. We tested the hypothesis that reduced genetic diversity in pericon populations caused the extinction of this species in Yautepec hills. The genetic diversity of six populations of Tagetes lucida were evaluated using PCR-based inter-simple sequence repeat (ISSR) markers. Ten ISSR primers produced a total of 1677 fragments with 100% polymorphism. The mean values of PIC, EMR and Rp were 0.34, 2.51 and 2.18, respectively. The results indicate that evaluated populations of Tagetes lucida have a high genetic diversity (79%), showing that the genetic factors are not the drivers of the species local extinction, and suggesting, that nongenetic factors such as overexploitation, overkill by herbicides, habitat destruction and fragmentation, the introduction of invasive species, rapid climate change, or pollution, reduce the growth rates of species and cause their populations to decline. These results show that T. lucida is a sensitive species that may require urgent conservation interventions.
Keywords:
Tagetes lucida
; polymorphism
; genetic erosion
; climate change
1. Introduction
Plants have been used by humans since immemorial time. On the one hand, natural elements have been used to satisfy humans’ necessities such as food, clothing, building materials and traditional medicine. On the other hand, plants have been included in cosmogonies of original civilisations being incorporated in the culture and beliefs, conferring them spiritual value [1,2,3,4]. In this sense, in Mexico several plant species that were used by Mesoamerican groups before the Spanish conquest, are still used due to their socio-cultural importance, for example the milpa and agave now considered part of Mexico biocultural heritage [5,6,7,8,9].
Pericon or yauhtli (Tagetes lucida) is considered a biocultural heritage in Central Mexico, specifically in Yautepec (pericon hill in Nahuatl), Morelos. This is because it has been used since ancient Mesoamerican civilisations in religious ceremonies, traditional medicine, food and clothing. Tagetes lucida is a plant that belongs to the Asteraceae family. It is native to Mexico, South and Central America [10,11,12]. It is a wild plant that is characterised by its yellow colouration and intense aniseed odour [13]. From an ecological perspective, the function of this plant within its ecosystem is of key importance. This is due to the fact that it plays a key role in attracting pollinators and deterring herbivores. Pericon is a perennial species that thrives in a distinct ecological niche: the ecotone between the tropical dry forest and the oak-pine forest, but also it grows in corn fields, in open fields and on roads.
Currently, the loss of species occur at a rate that exceeds the rate of origin of new species, and this is a consequence mainly due to human activities [14], such as changes in land use, climate change, overexploitation, use of herbicides and deforestation [15,16,17]. The rate of these changes has obligated to study the health of species, this includes the distribution and species richness, genetic erosion and other studies that allow to create the basis for management and conservation programs. In this sense, genetic erosion is referred to loss of genetic diversity within a species, often magnified or accelerated by human activities [18,19].
The deciduous forest is the main predominant ecosystem in Yautepec, Morelos and is one of the most coveted ecosystems for agricultural activity and urban development due to environmental conditions such as climate, soil type and vegetation [20,21]. Since colonial times, sugar cane production has played an important role in the regional economy, however, this activity has affected land use, and it also has had an important impact on the growth and distribution of the population since the 16th century. During the Porfiriato territorial expansion, emigration and immigration triggered the Zapatista movement in Morelos [22]. In 1940 green revolution was implemented in Mexican agriculture with technological packs for increased production, this modernization caused a dependency on seeds, pesticides, herbicides and fertilizers [23,24]. These are examples of several important historical processes that have influenced changes in Yautepec territory.
To know the perception of Yautepec inhabitants about the pericon populations growth, a series of interviews were conducted with social actors (chronicler, historian, pericon sellers, communal landowners, Mexica dancers, local authorities and a civil organization). The questions include in this interview were: Do you think that the pericon is still growing as it did years ago, or has it decreased? Why? More than 50% of the social actors consider pericon populations to have decreased in the last 50 years. They mention “The field has decreased, everything has changed, the climate is very changing, there is a water shortage”, “Due to urban sprawl and the use of herbicides in the field”, “the field where pericon grew in, now it is occupied by a lot of condominiums or another type of crops”.
In this sense, Yautepec inhabitants perceive a decrease in pericon populations, and also an increase in the temperature caused by climate changes that could encourage the migration of pericon populations to colder zones. Furthermore, pericon populations have been exposed to herbicides applications and land burning, and commercial and traditional exploitation could influence the erosion of this species.
It is hypothesized that species with such specific habitat requirements, such as Pericon, are particularly vulnerable to habitat loss and deterioration [25,26,27]. Changes in land use, overexploitation, the use of herbicides and deforestation have been identified as potential risks to this species [15,16,17], with the resultant genetic erosion potentially leading to a decline in genetic diversity, which in turn affects the adaptability of the species, as has been observed in the case of Pinus remota (Pinaceae) [28] and wild amaranth species [29]. This study sets out to investigate whether the loss of genetic diversity was the driving force behind the disappearance of Tagetes lucida in the hills of Yautepec.
Molecular markers have become a simple and accurate tool for the estimation of genetic diversity and relationship among genotypes of any organism [30,31]. Maturase K (matK) and chloroplast ribulose biphosphate carboxylase (rbcL) have been utilised to establish relationships between genera, given that these genes possess conserved regions which facilitate primer design and enable the differentiation of genera by virtue of their variable regions [30,31,32,33]. The use of nuclear internal transcribed spacer (ITS) and chloroplast DNA markers has been demonstrated to facilitate the differentiation between species and populations [33,34,35,36]. The molecular markers employed for the purpose of determining relationships between populations within a species include random amplified polymorphic DNA (RAPD), amplified fragment length polymorphic (AFLP), restriction fragment length polymorphic (RFLP) and simple sequence repeat (SSR), single nucleotide polymorphic (SNP), sequence-characterised amplified regions (SCARs) and inter simple sequence repeat (ISSR) [37,38,39,40]. One of the most frequently used markers in the genus Tagetes and in wild species is the Inter-Simple Sequence Repeat (ISSR) [41,42,43], a multilocus marker system that has demonstrated a higher rate of reproducibility due to the utilisation of longer primers (16-25 mers). This variety of markers has been demonstrated to be expeditious, economical, highly variable, and amenable to reproducibility without the necessity for prior sequence information regarding the amplified locus [44,45].
The present study sought to ascertain whether the genetic diversity of pericon was the causative agent of its extinction in the hills of Yautepec. In order to achieve this objective, an evaluation of the genetic diversity of six populations of Tagetes lucida was conducted using PCR-based Inter-Simple Sequence Repeat (ISSR) markers.
2. Results
2.1. Polymorphism and Efficiency of ISSR Markers
Of the 30 primers tested only 10, PrPha1, ISSR-02, ISSR-02b, ISSR1, ISSR4*, ISSR6, ISSR 10, ISSR-18, ISSR-31 and UBC889, showed highly polymorphism (Table 1). These ISSR primers were used to characterize and evaluate the genetic diversity in 180 genotypes of Tagetes lucida. ISSR primers produced 1677 fragments, all of them were polymorphic. The number of bands (TNB) per primer ranged from 131 (ISSR6) to 179 (ISSR10 and ISSR02b) with an average of 168 bands per primer (Table 2).
The Polymorphism Information Content (PIC) was calculated for each ISSR marker and ranged from 0.294 (ISSR-18) to 0.371 (UBC-889) with an average of 0.338 (Table 2). The effective multiple ratio (EMR) ranged from 1.044 (ISSR-6) to 4.888 (UBC-889) with an average of 2.511. The resolving power ranged from 1.333 (ISSR-6) to 3.5 (ISSR4*) with an average of 2.177.
2.2. Genetic Diversity Analysis
Molecular variance analysis (AMOVA) was performed to evaluate the variation between and within the Tagetes lucida populations (Table 3). According to the combination of the marker data, the results revealed 79% of the variation within regions, while the variation between regions was 21% with a significant PhiPT value (PhiPT=0.20, ƿ=0.001).
The observed number of alleles (Na) ranged from 1.31 (Ocuituco) to 1.54 (Oaxaca) with an average of 1.42. The effective number of alleles (Ne) ranged from 1.33 (Tepoztlan 2) to 1.39 (Oaxaca) with an average of 1.37. Shanon’s information index (I) ranged from 0.31 (Tepoztlan 2) to 0.38 (Tepoztlan 1) with a mean of 0.33. Expected heterozygosity (He) ranged from 0.20 (Tepoztlan 2) to 0.24 (Oaxaca). The percentage of polymorphic loci (PPL) of Tagetes lucida ranged from 64.23% (Ocuituco) to 75.71% (Oaxaca) with an average of 70.24% (Table 4).
2.3. Genetic Distance Cluster Analysis
The genetic distance between 6 regions, including 180 accessions of Tagetes lucida, ranged from a minimum of 0.02 to a maximum of 0.08. The minimum genetic distance was obtained for the paired regions of Tepoztlan1 and Yautepec. On the other hand, the maximum genetic distance was obtained for the paired regions of Oaxaca and Ocuituco (Table 5).
2.4. Cluster Analysis
The dendrogram divided the 6 populations into four main clusters (Figure 1). The first cluster included Yautepec and Tepoztlan1. The second cluster included Puente de Ixtla and Tepoztlan2. The third cluster consisted of Oaxaca and the fourth cluster of Ocuituco.
3. Discussion
Molecular markers reveal sites of variation in the DNA [46], In this work ISSR were used because these markers facilitate the determination of the genetic diversity among populations of the same species. The data showed that ISSR markers were an efficient tool for estimating genetic diversity in six populations of Tagetes lucida. Evaluated pericon populations showed a high percentage of polymorphism with an average of 100%, thus Tagetes lucida has important genetic diversity despite its traditional use, climate change, land use change, herbicide use and urbanization [17]. This result is higher compared to Namita et al. [43], Panwar et al. [44] and Majumder et al. [47] who reported 60.48%, 92.73% and 60.6% using ISSR markers for different species of Tagetes, in fact the result was higher than the reported by Shahzadi et al. [48] with 95.21% using RAPD markers in Tagetes. The mean values of PIC and Rp (0.34 and 2.18, respectively) confirm that ISSR markers are efficient for analyzing the genetic diversity of Tagetes lucida. These values are comparable to the results obtained by Panwar et al.[44] for Tagetes erecta (PIC=0.34) and Abd-dada et al. [45] for an endemic plant of Morocco (Euphorbia resinifera O. Berg) (Rp=2.8), based on ISSR markers.
According to the analysis of molecular variance (AMOVA), the variation within the population was higher (79%) than between them (21%). A high level of genetic diversity within the population should help the plants to cope with local environmental changes[45], as well as facilitate conservation and management programmes. The highest values of indices related to genetic variation were obtained for Tepoztlan1 (Ne= 1.43, I= 0.38 and He= 0.25) and Oaxaca (Na= 1.54 and PPL= 75.71%). This result indicates, on the one hand, that Tepoztlan1 and Oaxaca could be an important source of diversity for breeding conservation projects. The high genetic diversity observed in the Oaxaca population could have been due to the fact that the region is an open zone where several populations of Tagetes lucida converge. According to the distances and the dendogram, pericon is not vulnerable to habitat fragmentation, but it could be to overexploitation, because the distances between Yautepec, Tepoztlan1, Puente de Ixtla and Tepoztlan2 are smaller, despite habitat fragmentation, these populations are located in places where inhabitants collect pericon for different uses year by year. The opposite occurs in the Oaxaca and Ocuituco regions, where the dendogram showed differentiation from the other populations, which could be due to both populations being located on the road.
It is reported that fragmentation and small and dispersed populations cause genetic isolation, loss of genetic diversity and low genetic flow among them, and it could lead to a serious erosion of the genetic pool [28], in this investigation it was expected that diversity genetic would be low in the Tagetes lucida population as a consequence of loss and habitat fragmentation. ISSR markers amplified genomic neutral regions, so with this markers it is obtained a neutral genetic diversity, these markers have the disadvantage of amplifying non coding regions, thus to gain a better understanding of the adaptative genetic load and its influence on the adaptation of species to environmental and habitat changes it is necessary to use molecular markers that facilitate the determination of adaptative genetic diversity that will allow to study the capacity of the plants to respond to environmental changes [49,50]. Tagetes lucida populations may have high genetic diversity among them as it is reported in this work using ISSR as molecular markers, however this does not mean that the species could survived to accelerated socio-environmental changes. The changes in Yautepec have occurred in an accelerate way and included pesticides use, in this regard inhabitants of Morelos mention the affectation of herbaceous plants due to use of glyphosate, they observed a decrease in wild plants such as pericon related with the use of this kind of products, so these activities are very abrupt and plants may have not opportunity to adapt and thus got extinct.
On the other hand, it is necessary to explore the use of other molecular markers, such as microsatellites. Since they are codominant, they will provide more precise data on gene flow and its directionality. Likewise, the use of chloroplast markers will provide ancestral signals about past population dynamics that could be linked to climatic changes during the Pleistocene and that may have contributed to the current distribution of the species.
Conservation Recommendations. The results obtained in this study may be useful for determining specific actions for the conservation of the species. Particularly in the state of Morelos, where traditional use of Tagetes lucida is intense, it is necessary to propose conservation strategies with the community, the stakeholders are an important referent in decision-making. Some propose could be to produce plants with seeds from wild populations because the annual extraction of wild plants from their populations could be silently impacting the populations. If it is possible to produce plants from wild seeds, it is possible to reduce the extraction of wild plants and promote their conservation.
4. Materials and Methods
4.1. Obtention of Plant Material
Tagetes lucida was collected from five different geographical points of the state of Morelos (Yautepec, Ocuituco, Tepoztlán (east), Tepoztlán (west), Puente de Ixtla) and one from Oaxaca, ranging from 1222 to 2135 m above sea level (Figure 2, Table 6). The sampling mode used in this study is simple random and it was based on NOM-126-ECOL-2000 during the months of September to November of 2020-2021. The collection consisted of five pericon leaves of plants in flowering state per individual (one plant), with 30 individuals per population.
4.3. ISSR Analysis
A random set of 30 primers (Table 7) was tested for analysis of genetic diversity among six Tagetes lucida populations. The selective criteria was to select ISSR that showed most polymorphism. DNA amplification was carried out in 25 µl per reaction. PCR amplification reactions contained 80 ng genomic DNA, 10 µl 1X PCR buffer, 2.5 Mm MgCl2, 0.5 Mm dNTP’s (dATP, dCTP, dGTP and dTTP), 0.6 pmol/µl primer, 3 U/µl TaqDNA Polymerase (BIOLINE, London, UK) and 29.75 µl pure water. Amplification was performed in a thermocycler (Multi GeneTM) with the following programme: denaturation at 94°C for 5 min; followed by 45 cycles at 94°C for 45 s, a hybridization at 52 to 57°C (depending on primer) for 1 min; an extension step at 72°C for 2 min and a final extension at 72°C for 10 min. DNA amplifications were stored at 4°C until visualization by polyacrylamide gel electrophoresis with ethidium bromide solution for 30 min. The approximate molecular weight of the DNA amplifications was determined using a 100 pb ladder (ROCHE DNA Molecular weight Marker XIV).
4.4. Statical Analysis
4.4.1. Genetic Diversity and Frequency Analysis
The band profile was scored as present (1) or absent (0) for each entry and a binary qualitative data matrix was constructed. Weak bands with negligible intensity were excluded from scoring. Genetic parameters such as observed number of alleles (Na), effective number of alleles (Ne), Shannon’s information index (I), expected heterozygosity (He) and percentage of polymorphic loci (PPL) were calculated using GenAlex version 6.5. The analysis of molecular variance for the 180 accessions was calculated using the PopGen software platform.
4.4.2. Marker Efficiency Analysis
Polymorphism information for each molecular marker was measured by calculating several parameters including: polymorphism information content (PIC), effective multiple ratio (EMR), marker index (MI) and resolving power (Rp) using iMEC program (https://irscope.shinyapps.io/iMEC/). The percentage polymorphic band (PPB) was calculated according to the methodology of [32].
4.4.3. Marker Efficiency Analysis
Calculations for multivariate analysis were performed using PopGen. Clustering was performed to determine the genetic distance between individuals and to verify the consistency of population genetic variation. The dissimilarity index for cluster analysis was performed using the Unweighted Paired Group Method using Arithmetic Averages (UPGMA) cluster analysis and a dendrogram was constructed.
5. Conclusions
The present study evaluated the genetic diversity of Tagetes lucida, a wild species that could be a vulnerable plant. The results of this study confirmed the efficiency of using ISSR markers to analyze genetic diversity between and within populations of Tagetes lucida. The level of polymorphism within populations of Tagetes lucida was significantly high, showing that the decline in populations was not due to the plants’ inability to adapt to changing conditions, but that they may have been victims of a drastic process of overexploitation and pesticides use. The determination of adaptive genetic diversity may provide information on the capacity of these populations to adapt to the changes occurring in their habitat and the pressure on them imposed by collection for ceremonial and medicinal uses. The results of this work should be useful for designing management and conservation programmes for Tagetes lucida through decision making with stakeholders. The plant needs to be propagated and reintroduced into the hills where it naturally grew. Existing populations need to be managed in a sustainable way, through the sustainable collection of flowers for ceremonial and medicinal use.
Author Contributions
Conceptualization, K.B.T., M.M.C.R, L.A.A.M. and J.M.M.; methodology, J.M.M, K.B.T. and M.M.C.R.; validation, M.M.C.R.; formal analysis, J.M.M. and M.M.C.R.; investigation, K.B.T, M.M.C.R. and J.M.M.; resources, K.B.T., M.M.C.R.; data curation, J.M.M. and M.M.C.R.; writing—original draft preparation, K.B.T, M.M.C.R and J.M.M.; writing—review and editing, K.B.T., M.M.C.R. and J.M.M.; visualization, K.B.T., M.M.C.R. and J.M.M.; supervision, K.B.T., M.M.C.R.; project administration, K.B.T., M.M.C.R.; funding acquisition, K.B.T., M.M.C.R. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Instituto Politécnico Nacional through Secretaría de Investigación y Posgrado (SIP 20240980, SIP 20232022, SIP 20220841, SIP 20200336) and CONAHCyT mobility scholarship.
Acknowledgments
The authors thank the molecular biology laboratory of the Centro Interdisciplinario de Investigación para el Desarrollo Integral Regional Unidad Durango for the facilities, and Margarita Ortega and Abimael for their technical assistance. Thanks to CONAHCyT for the scholarship (781734) awarded to JMM.
Data Availability Statement
The original contributions presented in this study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author(s).
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| ISSR | Inter Simple Sequence Repeat |
| PIC | Polymorphic Index Content |
| EMR | Effective Multiple Ratio |
| Rp | Resolving power |
| AMOVA | Analysis of Molecular Variance |
| PCR | Polymerase Chain Reaction |
| MI | Marker Index |
| TNB | Total Number of Bands |
| NPB | Number of Polymorphism Bands |
| PPB | Percent of Polymorphic Band |
| df | degree of freedom |
| SS | Sum of Squares |
| MS | Mean of Squares |
| Est. Var. | Estimated Variance components |
| Var | Total Variance |
| Na | Observed number of alleles |
| Ne | Effective number of alleles |
| I | Shannon’s information index |
| He | Expected heterozygosity |
| PPL | Percentage of Polymorphic Loci |
| UPGMA | Unweighted Paired Group method using Arithmetic Averages |
| DNA | Deoxyribonucleic Acid |
| RAPD | Random Amplified Polymorphic DNA |
| SILEX | Silica Matrix Extraction |
| CTAB | Cetyltrimethylammonium Bromide |
| EDTA | Ethylenediaminetetraacetic acid |
| PEG | Polyethylene glycol |
References
- Olko, J.; Radding, C. Living with Nature, Cherishing Language: Indigenous Knowledges in the Americas through History; 2023; ISBN 9783031387395.
- Ouré, R.B.; Kpéra, G.N.; Djagoun, C.A.M.S.; Kassa, B.; Natta, A.; Djego, J.G.; Eniang, E.A.; Mensah, G.A.; Sinsin, B. Does Crocodile Conservation Matter in Sacred Natural Sites of Benin (Western Africa)? West African J. Appl. Ecol. 2023, 31, 69–85. [Google Scholar]
- Villa, R.A. Toad in the Road: Biocultural History and Conservation Challenges of the Sonoran Desert Toad. J. Psychedelic Stud. 2023, 7, 68–79. [Google Scholar] [CrossRef]
- Nunes, C.B.; Vieira, K.C.; Pereyra, P.E.R.; Hallwass, G.; Cunha, C. V.; Silvano, R.A.M. ‘From the Sky to the Ground’: Fishers’ Knowledge, Landscape Analysis and Hydrological Data Indicate Long-Term Environmental Changes in Amazonian Clear Water Rivers. Sci. Total Environ. 2023, 904, 166763. [Google Scholar] [CrossRef]
- Rangel-Landa, S.; Casas, A.; Rivera-Lozoya, E.; Torres-García, I.; Vallejo-Ramos, M. Ixcatec Ethnoecology: Plant Management and Biocultural Heritage in Oaxaca, Mexico; Journal of Ethnobiology and Ethnomedicine, 2016; Vol. 12; ISBN 1300201601013.
- Moreno-Calles, A.I.; Valencia-Vázquez, F. Nopales, Tsituni, Colorines y Capulines. Legado Biocultural Que Comemos y Cuidamos En La Ciudad de Morelia. Rev. Etnobiología 2024, 22, 36–48. [Google Scholar]
- Vázquez-Delfin, P.; Casas, A.; Vallejo, M. Adaptation and Biocultural Conservation of Traditional Agroforestry Systems in the Tehuacán Valley: Access to Resources and Livelihoods Strategies. Heliyon 2022, 8. [Google Scholar] [CrossRef] [PubMed]
- Jiménez, E.S. El Maguey Mexiquense : Patrimonio Biocultural y Sustentabilidad En El Aprovechamiento de Una Planta Ancestral y Bien Común The Mexiquense Maguey : Biocultural Heritage and Sustainability in the Use of an Ancestral Plant and Common Good. 2024, 1982–2011.
- Pensado-Leglise, M. del R.; Luna-Vargas, S.; Bustamante-Ramírez, H.A. Conservation of Biocultural Diversity in the Huasteca Potosina Region, Mexico. Diversity 2022, 14. [Google Scholar] [CrossRef]
- Kumar, A.; Pratap, B.; Gautam, D.K.; Yadav, V. Variability , Heritability and Genetic Advance Studies in French Marigold Variability , Heritability and Genetic Advance Studies in French Marigold ( Tagetes Patula L .). 2019.
- Batish, D.R.; Arora, K.; Singh, H.P.; Kohli, R.K. Potential Utilization of Dried Powder of Tagetes Minuta as a Natural Herbicide for Managing Rice Weeds. Crop Prot. 2007, 26, 566–571. [Google Scholar] [CrossRef]
- Soule, J.A. Infrageneric Systematics of Tagetes. In Proceedings of the Proceedings of the International Compositae Conference, Compositae: Systematics, Kew, UK; 1994; pp. 435–443.
- Domínguez-García, M.V.; Flores-Merino, M.V.; Montenegro-Morales, L.P.; Bedolla-Barajas, M. La Magia Ceremonial y Medicinal Del Yauhtli o Pericón (Tagetes Lucida Cav.). In Principios activos de plantas usadas en la medicina tradicional mexicana; Domínguez-García, M.V., Ed.; Universidad Autónoma del Estado de México: Toluca, México, 2023; pp. 123–142. [Google Scholar]
- Frankham, R.; Ballou, J.D.; Briscoe, D.A. A Primer of Conservation Genetics; Cambridge University Press: Cambridge, 2004; ISBN 9780521538275. [Google Scholar]
- Mendoza-Ponce, A.; Corona-Núñez, R.; Kraxner, F.; Leduc, S.; Patrizio, P. Identifying Effects of Land Use Cover Changes and Climate Change on Terrestrial Ecosystems and Carbon Stocks in Mexico. Glob. Environ. Chang. 2018, 53, 12–23. [Google Scholar] [CrossRef]
- Sáenz-Romero, C.; Kremer, A.; Nagy, L.; Újvári-Jármay, É.; Ducousso, A.; Kóczán-Horváth, A.; Hansen, J.K.; Mátyás, C. Common Garden Comparisons Confirm Inherited Differences in Sensitivity to Climate Change between Forest Tree Species. PeerJ 2019, 7, e6213. [Google Scholar] [CrossRef]
- Kurpis, J.; Serrato-Cruz, M.A.; Feria Arroyo, T.P. Modeling the Effects of Climate Change on the Distribution of Tagetes Lucida Cav. (Asteraceae). Glob. Ecol. Conserv. 2019, 20, e00747. [Google Scholar] [CrossRef]
- Rogers, D.L. Genetic Erosion. Nativ. plants 2004, 113–122. [Google Scholar] [CrossRef]
- Ahuja, M.R.; Mohan Jain, S. Genetic Diversity and Erosion in Plants: Indicators and Prevention; 2015; Vol. 1; ISBN 9783319256351.
- García Flores, A.; Valle Marquina, R.; Monroy Martínez, R. El Patrimonio Biocultural de La Selva Baja Caducifolia, Sierra de Huautla, Morelos: The Biocultural Heritage of the Deciduous Lowland Forest, Sierra de Huautla, Morelos. Inventio 2021, 17, 1–13. [Google Scholar] [CrossRef]
- Cheryl2 E., M. El Desarrollo de Las Haciendas En El Valle de Yautepec, 1610-1760. In Historia de Morelos. Tierra, gente, tiempos del Sur.; Crespo, H., Ed.; México, 2018; pp. 237–263.
- Sánchez-Salazar, M.T. La Actividad Cañero-Azucarera Como Factor Fundamental En La Organización Espacial Del Estado de Morelos. Investig. geográficas 1987, 103–123. [Google Scholar] [CrossRef]
- Martínez-Castro, César; Ríos-Castillo, Maricela; Castillo-Leal, M. La Revolución Verde y Sus Consecuencias Socioeconómicas En La Agricultura Mexicana. Ra Ximhai 2019, 15, 101–116. [CrossRef]
- Romero Polanco, E. Un Siglo de Agricultura En México; UNAM, Ed.; México, D.F., 2002; ISBN 970-701-291-9.
- García-Gonzáles, R.; Carrasco, B.; Peñailillo, P.; Letelier, L.; Herrera, R.; Lavandero, B.; Moya, M.; Caligari, P.D.S. Genetic Variability and Structure of Gomortega Keule (Molina) Baillon (Gomortegaceae) Relict Populations: Geographical and Genetic Fragmentation and Its Implications for Conservation. Botany 2008, 86, 1299–1310. [Google Scholar] [CrossRef]
- Fuentes, Glenda; Cisternas, Anita; Valencia, Graciela; Mihic, Maritza; Ruiz, Eduardo; Hasbún, Rodrigo; Valenzuela, Sofía; and M. Baeza, C. Diversidad Taxonómica y Genética Del Sitio Prioritario Península de Hualpén, Región Del Bío-Bío, Chile. Implicancias Para La Conservación. Gayana Bot. 2017, 74, 94–110.
- Bekessy, S.A.; Allnutt, T.R.; Premoli, A.C.; Lara, A.; Ennos, R.A.; Burgman, M.A.; Cortes, M.; Newton, A.C. Genetic Variation in the Vulnerable and Endemic Monkey Puzzle Tree, Detected Using RAPDs. Heredity (Edinb). 2002, 88, 243–249. [Google Scholar] [CrossRef]
- Naydenov, K.D.; Mladenov, I.; Alexandrov, A.; Naydenov, M.K.; Gyuleva, V.; Goudiaby, V.; Nikolić, B.; Kamary, S. Patterns of Genetic Diversity Resulting from Bottlenecks in European Black Pine, with Implications on Local Genetic Conservation and Management Practices in Bulgaria. Eur. J. For. Res. 2015, 134, 669–681. [Google Scholar] [CrossRef]
- Ruiz Hernández, V.C.; Legaría Solano, J.P.; Sahagún Castellanos, J.; De la O Olan, M.; Ruiz Hernández, V.C.; Legaría Solano, J.P.; Sahagún Castellanos, J.; de la O Olan, M. Variabilidad Genética En Algunas Especies Cultivadas y Silvestres de Amaranto. Rev. Mex. Ciencias Agrícolas 2018, 9, 405–416. [Google Scholar] [CrossRef]
- Thormann, C.E.; Ferreira, M.E.; Camargo, L.E.A.; Tivang, J.G.; Osborn, T.C. Comparison of RFLP and RAPD Markers to Estimating Genetic Relationships within and among Cruciferous Species. Theor. Appl. Genet. 1994, 88, 973–980. [Google Scholar] [CrossRef] [PubMed]
- do Val, A.D.; Ferreira, J.L.; Vieira Neto, J.; Pasqual, M.; de Oliveira, A.F.; Borém, A.; Cançado, G.M. Genetic Diversity of Brazilian and Introduced Olive Germplasms Based on Microsatellite Markers. Genet. Mol. Res. 2012, 11, 556–571. [Google Scholar] [CrossRef] [PubMed]
- Yong, W.T.L.; Mustafa, A.A.; Derise, M.R.; Rodrigues, K.F. DNA Barcoding Using Chloroplast MatK and RbcL Regions for the Identification of Bamboo Species in Sabah. Adv. Bamboo Sci. 2024, 7, 100073. [Google Scholar] [CrossRef]
- Li, F.W.; Kuo, L.Y.; Rothfels, C.J.; Ebihara, A.; Chiou, W.L.; Windham, M.D.; Pryer, K.M. Rbcl and Matk Earn Two Thumbs up as the Core DNA Barcode for Ferns. PLoS One 2011, 6, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Group, C.P.W.; Hollingsworth, P.M.; Forrest, L.L.; Spouge, J.L.; Hajibabaei, M.; Ratnasingham, S.; van der Bank, M.; Chase, M.W.; Cowan, R.S.; Erickson, D.L.; et al. A DNA Barcode for Land Plants. Proc. Natl. Acad. Sci. U. S. A. 2009, 106, 12794–12797. [Google Scholar] [CrossRef]
- Shneyer, V.S.; Rodionov, A. V. Plant DNA Barcodes. Biol. Bull. Rev. 2019, 9, 295–300. [Google Scholar] [CrossRef]
- Ismail, M.; Ahmad, A.; Nadeem, M.; Javed, M.A.; Khan, S.H.; Khawaish, I.; Sthanadar, A.A.; Qari, S.H.; Alghanem, S.M.; Khan, K.A.; et al. Development of DNA Barcodes for Selected Acacia Species by Using RbcL and MatK DNA Markers. Saudi J. Biol. Sci. 2020, 27, 3735–3742. [Google Scholar] [CrossRef]
- Antil, S.; Abraham, J.S.; Sripoorna, S.; Maurya, S.; Dagar, J.; Makhija, S.; Bhagat, P.; Gupta, R.; Sood, U.; Lal, R.; et al. DNA Barcoding, an Effective Tool for Species Identification: A Review. Mol. Biol. Rep. 2023, 50, 761–775. [Google Scholar] [CrossRef]
- Sun, P.; Clamp, J.C.; Xu, D.; Huang, B.; Shin, M.K.; Turner, F. An ITS-Based Phylogenetic Framework for the Genus Vorticella: Finding the Molecular and Morphological Gaps in a Taxonomically Difficult Group. Proc. R. Soc. B Biol. Sci. 2013, 280. [Google Scholar] [CrossRef]
- M, Idrees, I.M. Molecular Markers in Plants for Analysis of Genetic Diversity: A Review. Eur. Acad. Res. 2014, 2, 1513–1540.
- Mukkamula, N.; Beema, N.; Dasari, S.; Kiran Kumar, B. Assessment of Genetic Diversity in Maerua Oblongifolia (Forssk.) A. Rich. Accessions Based on RAPD Markers. Ecol. Genet. Genomics 2023, 27, 100171. [Google Scholar] [CrossRef]
- Velasco-Ramírez, A.P.; Torres-Morán, M.I.; Molina-Moret, S.; de Jesús Sánchez-González, J.; Santacruz-Ruvalcaba, F. Efficiency of Rapd, Issr, Aflp and Istr Markers for the Detection of Polymorphisms and Genetic Relationships in Camote de Cerro (Dioscorea Spp.). Electron. J. Biotechnol. 2014, 17, 65–71. [Google Scholar] [CrossRef]
- Choi, T.Y.; Lee, S.R. A Review of Intraspecific Genetic Diversity on Wild Plants in Korea Estimated from Varying Nuclear DNA Markers. J. Asia-Pacific Biodivers. 2021, 14, 622–627. [Google Scholar] [CrossRef]
- Namita; Panwar, S.; Sonah, H.; Singh, K.P.; Sharma, T.R. Genetic Diversity Analysis of Marigold (Tagetes Sp) Genotypes Using RAPD and ISSR Markers. Indian J. Agric. Sci. 2013, 83, 484–490.
- Panwar, S.; Singh, K.P.; Namita; Janakiram, T.; Sonah, H.; Sharma, T.R. DNA Fingerprinting in African Marigold (Tagetes Erecta L.) Genotypes Using ISSR and URP Markers. Indian J. Hortic. 2018, 75, 105–110. [CrossRef]
- Abd-dada, H.; Bouda, S.; Khachtib, Y.; Bella, Y.A.; Haddioui, A. Use of ISSR Markers to Assess the Genetic Diversity of an Endemic Plant of Morocco (Euphorbia Resinifera O. Berg). J. Genet. Eng. Biotechnol. 2023, 21. [Google Scholar] [CrossRef] [PubMed]
- Jones, N.; Ougham, H.; Thomas, H.; PaÅ-akinskien, I. Markers and Mapping Revisited: Finding Your Gene. New Phytol. 2009, 183, 935–966. [Google Scholar] [CrossRef]
- Majumder, J.; Singh, S.; Kumari, M.; Verma, M. Variability and Correlation Studies on Induced Mutants of Marigold (Tagetes Erecta L.) for Different Traits and Assessing Them Using Molecular Markers. Plant Tissue Cult. Biotechnol. 2018, 28, 223–236. [Google Scholar] [CrossRef]
- Shahzadi, I.; Ahmad, R.; Waheed, U.; Shah, M.M. Genetic Diversity Analysis of Tagetes Species Using Pcr Based Molecular Markers. Pakistan J. Bot. 2016, 48, 1593–1599. [Google Scholar]
- Chung, M.Y.; Merilä, J.; Li, J.; Mao, K.; López-Pujol, J.; Tsumura, Y.; Chung, M.G. Neutral and Adaptive Genetic Diversity in Plants: An Overview. Front. Ecol. Evol. 2023, 11. [Google Scholar] [CrossRef]
- Teixeira, J.C.; Huber, C.D. The Inflated Significance of Neutral Genetic Diversity in Conservation Genetics. Proc. Natl. Acad. Sci. U. S. A. 2021, 118, 1–10. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Genetic relationships amoung all populations of Tagetes lucida.
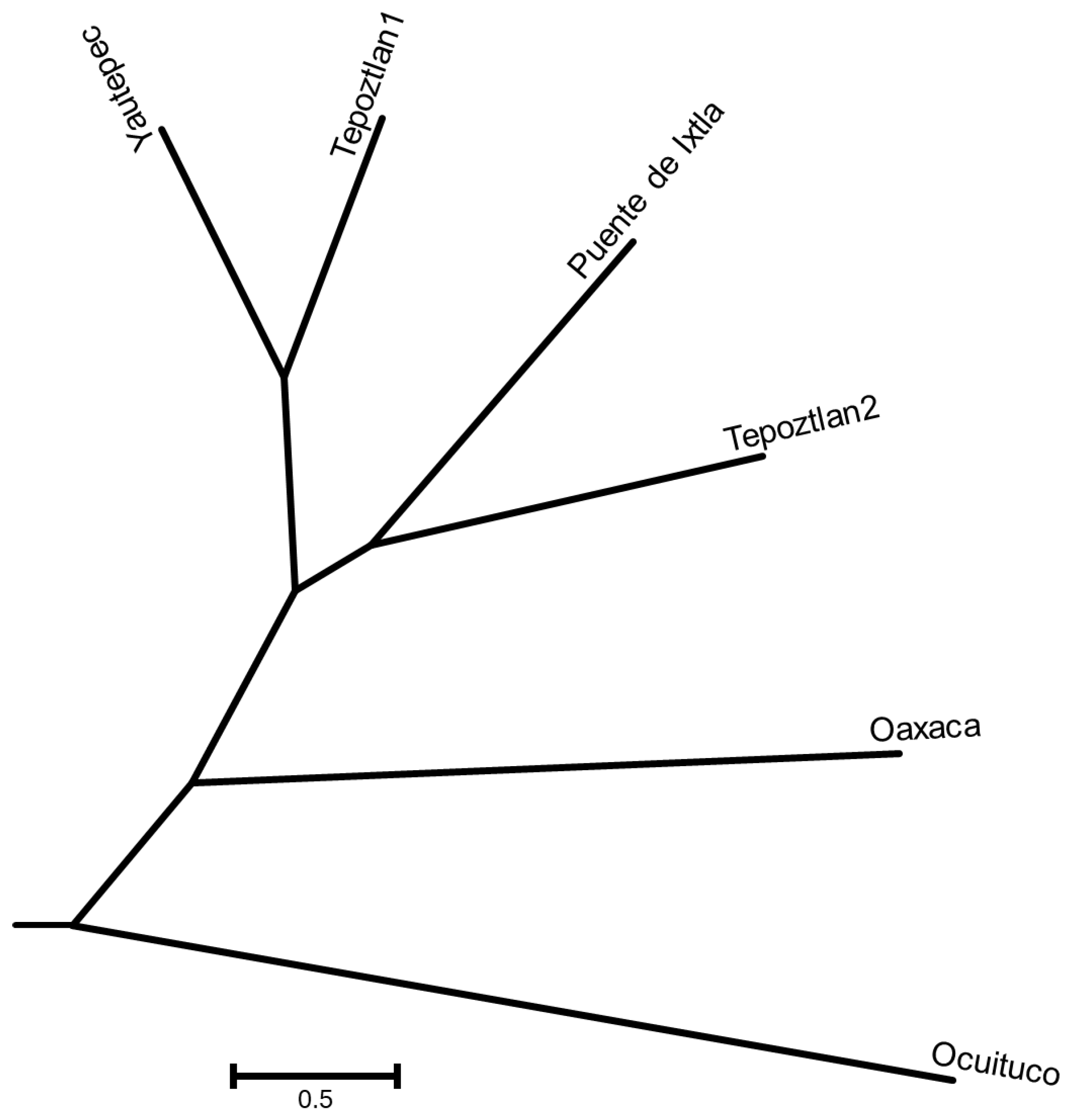
Figure 2.
Geographic distribution of the Tagetes lucida populations.
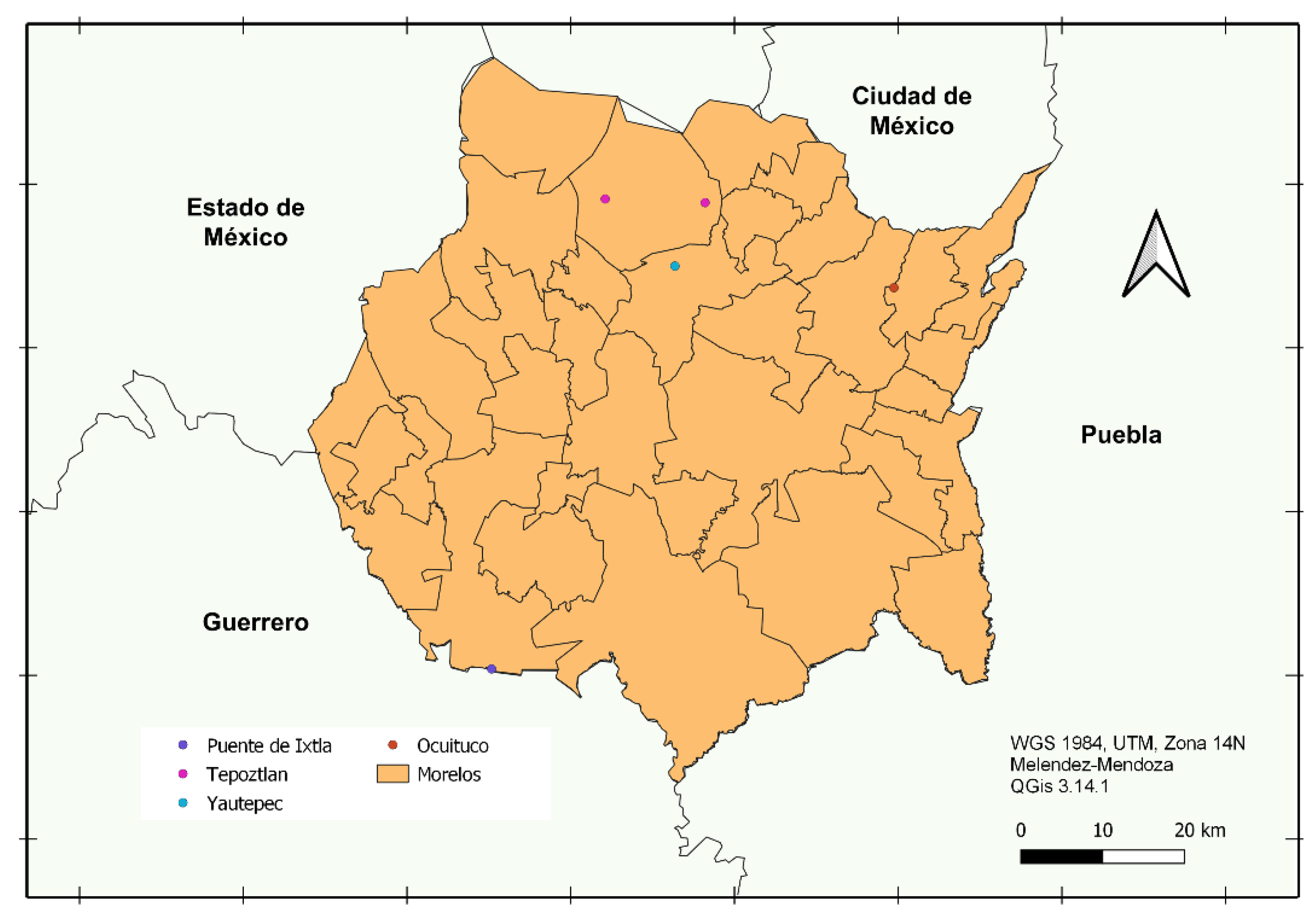

Table 1.
Selected ISSR primers for analysis in Tagetes lucida.
| ISSR primers | Sequence (5’-3’) | Ta (°C) | |
|---|---|---|---|
| PrPha1 | GAGCAACAACAACAACAA | 52 | |
| ISSR-02 | CACACACACACACACAGC | 57 | |
| ISSR-02b | CACACACACACACACAGG | 57 | |
| ISSR1 | BDBACAACAACAACAACA | 52 | |
| ISSR4* | CTCCTCCTCCTCCTCCTC | 57 | |
| ISSR6 | ACAACAACAACAACABDB | 52 | |
| ISSR10 | CACACACACACACACACAG | 57 | |
| ISSR-18 | GAGAGAGAGAGAGAGAC | 52 | |
| ISSR-31 | AGAGAGAGAGAGAGAGGC | 57 | |
| UBC889 | CTTACACACACACACA | 52 | |
A: adenine; T: timine; C: citocine and G: guanine; Ta: annealing temperature.
Table 2.
Efficacy of primer polymorphism in Tagetes lucida.
| Primer | Sequence (5’-3’) | TNB | NPB | PPB(%) | PIC | EMR | Rp |
|---|---|---|---|---|---|---|---|
| ISSR6 | ACAACAACAACAACABDB | 131 | 131 | 100 | 0.30 | 1.04 | 1.33 |
| ISSR10 | CACACACACACACACACAG | 179 | 179 | 100 | 0.35 | 3.27 | 3.27 |
| ISSR02 | CACACACACACACACAGC | 172 | 172 | 100 | 0.36 | 2.49 | 2.04 |
| ISSR02b | CACACACACACACACAGG | 179 | 179 | 100 | 0.37 | 3.22 | 2.04 |
| ISSR18 | GAGAGAGAGAGAGAGAC | 150 | 150 | 100 | 0.29 | 1.42 | 1.81 |
| ISSR1 | BDBACAACAACAACAACA | 165 | 165 | 100 | 0.31 | 1.69 | 1.68 |
| ISSR31 | AGAGAGAGAGAGAGAGGC | 175 | 175 | 100 | 0.34 | 2.01 | 1.81 |
| ISSR4* | CTCCTCCTCCTCCTCCTC | 175 | 175 | 100 | 0.32 | 2.33 | 3.50 |
| UBC889 | CTTACACACACACACA | 176 | 176 | 100 | 0.37 | 4.89 | 2.67 |
| PrPha1 | GAGCAACAACAACAACAA | 175 | 175 | 100 | 0.36 | 2.75 | 1.61 |
| Mean | 168 | 168 | 100 | 0.34 | 2.51 | 2.18 |
TNB: total number of bands; NPB: number of polymorphism bands; PPB: percent of polymorphic band; PIC: polymorphism information content; EMR: effective multiple ratio; MI: marker index; Rp: resolving power.
Table 3.
Analysis of molecular variance (AMOVA) based on ISSR data in Tagetes lucida population.
| Source | df | SS | MS | Est. Var. | Var |
|---|---|---|---|---|---|
| Among Pops | 5 | 342.500 | 68.500 | 2.026 | 21% |
| Within Pops | 174 | 1341.133 | 7.708 | 7.708 | 79% |
| Total | 179 | 1683.633 | 9.734 | 100% |
df: degree of freedom; SS: sum of squares; MS: mean of squares; Est. Var.: estimated variance components; Var: total variance; PhiPT=0.20, ƿ= 0.001.
Table 4.
Genetic variation among different regions of Tagetes lucida populations.
| Na | Ne | I | He | PPL | |
|---|---|---|---|---|---|
| Yautepec | 1.40 | 1.37 | 0.33 | 0.22 | 70.00% |
| Ocuituco | 1.31 | 1.38 | 0.33 | 0.22 | 64.29% |
| Tepoztlan1 | 1.47 | 1.43 | 0.38 | 0.25 | 72.86% |
| Puente de Ixtla | 1.34 | 1.36 | 0.31 | 0.21 | 65.71% |
| Oaxaca | 1.54 | 1.39 | 0.36 | 0.24 | 75.71% |
| Tepoztlan2 | 1.47 | 1.33 | 0.31 | 0.20 | 72.86% |
| Mean | 1.42 | 1.37 | 0.33 | 0.22 | 70.24% |
Na: observed number of alleles; Ne: effective number of alleles; I: Shannon’s information index; He: expected heterozygosity; PPL: percentage of polymorphic loci.
Table 5.
Pairwise population matrix of Nei genetic distance among 6 regions including 180 accessions of Tagetes lucida.
Table 5.
Pairwise population matrix of Nei genetic distance among 6 regions including 180 accessions of Tagetes lucida.
| Yautepec | Ocuituco | Tepoztlan1 | Puente de Ixtla | Oaxaca | Tepoztlan2 | |
|---|---|---|---|---|---|---|
| Yautepec | 0.000 | |||||
| Ocuituco | 0.040 | 0.000 | ||||
| Tepoztlan1 | 0.022 | 0.034 | 0.000 | |||
| Puente de Ixtla | 0.024 | 0.063 | 0.036 | 0.000 | ||
| Oaxaca | 0.049 | 0.088 | 0.054 | 0.054 | 0.000 | |
| Tepoztlan2 | 0.041 | 0.070 | 0.039 | 0.029 | 0.036 | 0.000 |
Table 6.
Tagetes lucida populations used for molecular analysis.
| Site | Code | Accesion | No. of sample | Altitude (m) | Latitude, Longitude | Voucher Nr. |
|---|---|---|---|---|---|---|
| Tepoztlan1 | TlTepoz1 | Wild | 30 | 1633 | 18.976, -99.150 | 30944 |
| Tepoztlan2 | TlTepoz2 | Wild | 30 | 1601 | 18.970, -99.034 | 30945 |
| Yautepec | TlYaute | Wild | 30 | 1222 | 18.902, -99.069 | 30950 |
| Ocuituco | TlOcui | Wild | 30 | 1753 | 18.878, -98.815 | 30949 |
| Puente de Ixtla | TlPte | Wild | 30 | 1233 | 18.470, -99.272 | 30951 |
| Oaxaca | TlOax | Wild | 30 | 2135 | 17.263, -96.539 | 30954 |
Table 7.
ISSR primers screening for analysis of genetic diversity among six Tagetes lucida populations.
Table 7.
ISSR primers screening for analysis of genetic diversity among six Tagetes lucida populations.
| ISSR primers | Sequence (5’-3’) | Ta (°C) |
|---|---|---|
| PrPha1 | GAGCAACAACAACAACAA | 52 |
| PrPha2 | AGAGAGAGAGAGAGAGTG | 52 |
| PHA 2 | CTCGTGTGTGTGTGTGTGT | 52 |
| PHA 3 | AGAGAGAGAGAGAGAGCG | 52 |
| PHA 4 | AGAGAGAGAGAGAGAG | 52 |
| PHA 5 | CCACCACCACCACCA | 52 |
| PHA 6 | GAAGAAGAAGAAGAA | 54 |
| ISSR1 | BDBACAACAACAACAACA | 52 |
| ISSR2 | CACCACCACCACCACCACG | 57 |
| ISSR 3 | ACCACCACCACCACCACCG | 57 |
| ISSR3* | CTCTCTCTCTCTCTCTAG | 52 |
| ISSR4 | GACAGACAGACAGACAAG | 52 |
| ISSR4* | CTCCTCCTCCTCCTCCTC | 57 |
| ISSR5 | GAGGAGGAGGAGGAGGAGC | 57 |
| ISSR6 | ACAACAACAACAACABDB | 52 |
| ISSR7 | WBGACAGACAGACAGACA | 52 |
| ISSR8 | GAGAGAGAGAGAGAGAGAC | 57 |
| ISSR9 | AGCAGCAGCAGCAGCGA | 57 |
| ISSR10 | CACACACACACACACACAG | 57 |
| ISSR-02 | CACACACACACACACAGC | 57 |
| ISSR-02b | CACACACACACACACAGG | 57 |
| ISSR-03 | GAGAGAGAGAGAGAGACT | 57 |
| ISSR-06 | AGAGAGAGAGAGAGAGCT | 57 |
| ISSR-12 | AGAGAGAGAGAGAGAGC | 57 |
| ISSR-18 | GAGAGAGAGAGAGAGAC | 57 |
| ISSR-31 | AGAGAGAGAGAGAGAGGC | 57 |
| UBC889 | CTTACACACACACACA | 52 |
| UBC880 | GGAGAGGAGAGGAGA | 57 |
| ISSR-15 | GTGGTGGTGGC | 52 |
| ISSR-08 | CCCGTGTGTGTGTGTGT | 52 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



