
Submitted:
13 June 2025
Posted:
16 June 2025
You are already at the latest version
Abstract
Aromatic compounds are vital in both natural and synthetic chemistry, traditionally sourced from non-renewable petrochemicals. However, plant biomass, particularly lignin, offers a renewable alternative source for aromatic compounds. Lignin, a complex polymer in plant cell walls, is the largest renewable source of aromatic compounds, though its degradation remains challenging. Lignin can be chemically degraded through oxidation, acid hydrolysis or solvolysis. Microorganisms, including fungi, offer a sustainable alternative in breaking down lignin into valuable compounds. Fungi possess unique enzymes capable of converting aromatic compounds derived from lignin into biofuels, bioplastics, and chemical building blocks. However, their aromatic metabolic pathways are less studied compared to bacterial systems. Recent advances in genomics, proteomics, and metabolic engineering are helping to reveal these pathways, offering new opportunities to optimize fungi for lignin bioconversion. By manipulating these pathways, researchers aim to enhance lignin valorization, contributing to sustainable biobased industries. This review highlights recent progress in understanding fungal aromatic metabolism, focusing on how Aspergillus niger converts plantderived aromatic compounds into useful products and the versatility of aromatic metabolism within the Aspergillus genus. Addressing the current knowledge gaps in fungal pathways could unlock their potential in sustainable technologies, promoting eco-friendly production of biofuels and chemicals from renewable resources.
Keywords:
aromatic metabolism
; aspergilli
; benzoic acids
; cinnamic acids
; phenolic compounds
1. Introduction
Aromatic compounds play crucial roles in plants and provide structural support (e.g., lignin), defense against herbivores and pathogens (e.g., phenolics), and contribute to pigmentation, fragrance, and flavor, aiding in pollination and protection against environmental stress. Additionally, they are also involved in metabolic processes and allelopathy, helping plants compete and thrive in their ecosystems [1]. Aromatic compounds are also prevalent in everyday life and are found in a wide range of man-made products such as beverages, cosmetics, foods, fuels, lubricants, medicines, paints, and plastics [2]. Most commercial aromatic compounds utilized today are synthesized from base chemicals like benzene, toluene and xylene which are typically extracted from nonrenewable fossil resources. Therefore, developing methods to utilize the wealth of natural aromatic compounds, producing those from renewable sources is essential for sustainable practices and reducing dependence on fossil fuels.
One of the most promising renewable sources for aromatic compounds is the complex plant polymer lignin. Between 15-30% of the dry weight of plant biomass consists of lignin depending on the species. Lignin is mostly built from the aromatic compounds coniferyl alcohol (guaiacyl (G)-units), p-coumaryl alcohol (p-hydroxyphenyl (H)-units) and sinapyl alcohol (syringyl (S)-units), derived from hydroxycinnamic acids, and the ratios of building blocks used in lignin varies per plant species. Additionally, different building blocks such as caffeoyl alcohol and vanillyl alcohol have been observed in lignin [3]. Hydroxycinnamic acids such as ferulic acid (3-methoxy-4-hydroxycinnamic acid) and p-coumaric acid (p-hydroxycinnamic acid) can be linked to complex plant cell wall structures like hemi-cellulose and pectin and have ester linkages with arabinose and galactose [11,12,13]. Currently, much research is performed in the utilization of lignin through chemical, enzymatic and biotransformation approaches [4,5,6,7].
Fungi are able to degrade and metabolize a wide range of monomeric aromatic compounds efficiently but the aromatic metabolic pathways in fungi are poorly studied. Moreover, most existing studies focus on different fungal species, making it challenging to determine which specific pathways are present within a single species. Hence, a complete fungal aromatic metabolic pathway overview is non-existing. In order the fill up the gaps, a comparison between the aromatic metabolic pathways of bacteria, yeast and fungi was made and showed that many pathways are shared between them [8]. Before 2020, the best described aromatic metabolic pathways were from Aspergillus japonicus and were obtained by using UV spectra, TLC, GLC and dioxygenase activity [9]. However, this study was before the genome era and no genes were identified. Despite that many fungal genomes are now sequenced and published, the number of identified fungal genes encoding enzymes converting aromatic compounds is remarkably low [8,10]. It was also revealed that unique and unexplored aromatic metabolic pathways exist in fungi.
Recently, multiple genes and enzymes involved in the metabolism of monomeric aromatic compounds, mainly in Aspergilli species, have been identified. With the identification of these genes, better predictions and correlations can be made of which metabolic pathways are present in fungi. In this review, the recent advances and insight in the monomeric aromatic metabolic pathways in fungi from the past years, with a main focus on Aspergillus niger, are highlighted and discussed.
2. Metabolism of Hydroxycinnamic Acids
The monomeric aromatic compounds ferulic acid (3-methoxy-4-hydroxycinnamic acid) and p-coumaric acid (p-hydroxycinnamic acid) are hydroxycinnamic acids that are part of complex sugars and are linked to arabinose and galactose residues via ester linkages [11,12,13]. These linkages are part of the plant cell wall and contribute to the cell wall rigidity and aid in the plants resistance against environmental stress and pathogens. Multiple fungi are able to hydrolyze ester bonds between hydroxycinnamic acids and polysaccharides using feruloyl esterases (Fae’s). These enzymes play a crucial role in the degradation on complex plant biomass and it has been shown that Fae’s can be used to releases ferulic acid from agricultural waste streams such as wheat bran, maize bran and sugar beet pulp [14,15,16]. Fae’s have been identified and studied in many fungi but mainly in the Aspergilli; Aspergillus niger, Aspergillus oryzae and Aspergillus nidulans [17]. At this moment, three Fae’s have been identified and characterized in A. niger [18,19,20]. Homologs of feruloyl esterase’s have been observed and studied in multiple fungi including Aspergillus, Fusarium, Penicillium, Pleurotus, Trichoderma, and Talaromyces species [17].
Multiple pathways that can convert ferulic acid have been described. A. oryzae can release ferulic acid from the cell wall of rice endosperm during brewing processes. After the release, ferulic acid is decarboxylated to 4-vinylguaiacol (Figure 1) [21]. 4-vinylguaiacol can give an undesired flavor in distilled products such as beer, awamori and sake [22,23,24,25,26]. In Aspergillus luchuensis, decarboxylation of hydroxycinnamic acids to its vinyl form is catalyzed by phenolic acid decarboxylase (Pad). This conversion was also observed in the A. niger diploid strain DAR2 (derived from A. niger C28B25). This strain is able to decarboxylate ferulic acid to 4-vinylguaiacol and further to vanillic acid, possibly with vanillin as intermediate (Figure 1) [27]. A close homolog of AlPad is present in A. niger (NRRL3_08440) and could be also involved in the decarboxylation of ferulic acid. This pathway has also been observed other fungi such as Fusarium solani (Mart.) Sacc., Colletotrichum gloeosporioides, Isaria farinosa and many yeast species [25,26,28,29] indicating that this pathway is relatively common. Interestingly, the A. niger C28B25, the parental strain of DAR2, is degrading ferulic acid to vanillic acid without the non-oxidative decarboxylation step to vinyl-guaiacol indicating that other pathways are present in A. niger [27].
Decarboxylation of hydroxycinnamic acid to its vinyl form has been observed in many fungi. In Saccharomyces cerevisiae, hydroxycinnamic acids and cinnamic acid are decarboxylated by ferulic acid decarboxylase 1 (Fdc1) and flavin prenyltransferase 1 (Pad1) respectively [30,31]. Homologs of Fdc1 and Pad1 are found in multiple Aspergillus species and have be shown to do this catalysis in vitro [32,33]. However, deletion of these genes in A. niger resulted in abolished growth on cinnamic acid but no phenotypes were observed on the hydroxycinnamic acids; ferulic acid, p-coumaric acid and caffeic acid [32]. This gave new indications that these enzymes are involved in vivo in cinnamic acid decarboxylation but not hydroxycinnamic acid decarboxylation [32]. In addition this was also supported by transcriptome data, showing that fdcA or padA are not induced when grown on caffeic acid, ferulic acid or p-coumaric acid. Therefore, FdcA of A. niger was renamed to cinnamic acid decarboxylase A (CdcA) [32].
Recently, it has been shown that A. niger degrades the hydroxycinnamic acids; caffeic acid, ferulic acid, p-coumaric acid and m-coumaric acid through the peroxisomal CoA-dependent beta-oxidative metabolic pathway resulting in the formation of protocatechuic acid (3,4-dihydroxybenzoic acid), vanillic acid (4-hydroxy-3-methoxybenzoic acid), p-hydroxybenzoic acid and m- hydroxybenzoic acid, respectively (Figure 1) [34]. Also, dihydrocaffeic acid (3-(3,4-dihydroxyphenyl)propanoic acid) and phloretic acid (3-(4-hydroxyphenyl)propanoic acid) are degraded through this pathway [34]. The CoA-dependent beta-oxidative metabolic pathway is well-known for the degradation of fatty acids but was not previously known to be involved in the degradation of hydroxycinnamic acids. Interestingly, also in plants it has been shown that hydroxycinnamic acid can be degraded through this pathway [35]. Multiple genes involved in the degradation of hydroxycinnamic acids in A. niger were identified using whole genome transcriptome data [34].
In A. niger, the first step of the CoA-dependent beta-oxidative metabolic pathway is catalyzed by the hydroxycinnamate-CoA synthase (HscA), and deletion of it results in strongly reduced growth on hydroxycinnamic acids but appears not to be required for fatty acid degradation [34]. The second and third step of this pathway is catalyzed by fatty acid oxidase A (FoxA) and is was identified in multiple fungal species to be involved in the fatty acid degradation [34,36,37,38,39,40,41]. The fourth step is catalyzed by 3-ketoacyl-CoA thiolase (KatA) (Figure 1). Deletion of these related genes resulted in clear phenotypes on several hydroxycinnamic acids, however growth was not abolished on ferulic acid or p-coumaric acid indicating that another pathway, functional redundancy or alternative genes are present in A. niger [34]. The final step of the CoA-dependent oxidative pathway is proposed to be catalyzed by a thioesterase. Four thioesterases (TheA, TheB, TheC and TheD) have been identified and individual deletion of these genes did not results in reduced growth on ferulic acid, p-coumaric acid and caffeic acid. Deletion of theB results in growth reduction on m-coumaric acid but the growth recovers over time and it was suggested that there is functional redundancy in the final step. The redundancy of the thioesterases could be further studied by making a quadruple deletion of the genes and analyze the growth on hydroxycinnamic acids. Interestingly, deletion of the CoA-dependent beta-oxidative genes did not result in reduce growth on sinapic acid (3,5-dimethoxy-4-hydroxycinnamic acid) or cinnamic acid indicating that they are degraded through other pathway(s) in A. niger.
In A. niger, o-coumaric acid (2-hydroxycinnamic acid) can be converted to 4-hydroxycoumarin but is appears not to be further degraded [42]. In A. fumigatus, melilotic acid (3-(2-hydroxyphenyl)propanoic acid) is converted through esterification to o-coumaric acid and further to 4-hydroxycoumarin and dicoumarol [43,44]. Interestingly, a CoA-dependent beta-oxidative pathway was suggested for the conversion of o-coumaric acid to 4-hydroxycoumarin with o-coumaryl-SCoA, o-hydroxyphenyl-β-hydroxypropionyl-SCoA, o-hydroxyphenyl-β-ketopropionic acid-SCoA as intermediates [44]. As mentioned before, in A. niger dihydrocaffeic acid (3-(3,4-dihydroxyphenyl)propanoic acid) and phloretic acid (3-(4-hydroxyphenyl)propanoic acid) are converted through the CoA-dependent beta-oxidative pathway and it is possible that both compounds are esterified to caffeic acid and p-coumaric acid, respectively. However, the involved genes of this pathway still need to be identified.
An alternative metabolic pathway for the metabolism of hydroxycinnamic acid is a CoA-independent non-oxidative pathway which has been observed in A. flavus [45]. p-coumaric acid is converted to β-hydroxy-(p-hydroxyphenyl)propionic acid and p-hydroxybenzoylacetic acid followed by a conversion to p-hydroxybenzoic acid. In A. luchuensis, it is proposed that ferulic acid in converted to vanillin through a CoA-dependent non-oxidative pathway having feruloyl-CoA and 4-hydroxy-3-methoxyphenyl-β-hydroxypropionyl-CoA (HMPKP-SCoA) as intermediate [24]. In A. brasiliensis ATCC 16404, the 2’ position of the aliphatic chain of ferulic acid is methoxylated to (E)-3-(4-hydroxy-3-methoxyphenyl)-2-methoxyacrylic acid [46].
3. Metabolism of Cinnamic Acid
The decarboxylation of cinnamic acid has been observed in multiple Aspergilli and Trichoderma species including A. niger, A. japonicus, A. flavus, A. oryzae, A. wentii, T. reesei, T. viride, T. koningii but also Penicillium species and the yeast S. cerevisiae [9,30,32,47,48,49]. As mentioned before, A. niger decarboxylates cinnamic acid to styrene by CdcA and PadA (Figure 2). Both genes are also involved in the degradation of sorbic acid and are regulated by the sorbic acid regulator A (SdrA) [30,32,50]. Interestingly, all three genes are clustered on the genome of many Aspergilli and a strong correlation was found in Aspergilli species between the ability to grow on cinnamic acid and sorbic acid and this gene cluster [32]. For example, A. nidulans is unable to grow on cinnamic acid due to a mutation resulting in a truncated padA.
In A. japonicus and A. niger, cinnamic acid can also be converted to benzoic acid [9,32,51]. Deletion of benzoate-4-monooxygenase (bphA), p-hydroxy-m-hydroxylase (phhA), protocatechuate 3,4-dioxygenase (prcA) or the combination protocatechuate hydroxylase (phyA)/prcA or hydroxyquinol 1,2-dioxygenase (hqdA)/prcA in A. niger results in reduced growth on cinnamic acid indicating that it is converted towards benzoic acid [51,52]. In addition, A. niger ΔprcAΔphyA transformant grown on cinnamic acid and cinnamyl alcohol resulted in accumulation of protocatechuic acid [51]. Currently, it remains unknown whether styrene is an intermediate of this conversion pathway or that an alternative cinnamic acid metabolic pathway is present in A. niger. However, in other fungi cinnamic acid pathways towards benzoic acid and p-hydroxybenzoic acid have been observed. In Phomopsis liquidambari, it is suggested that cinnamic acid is decarboxylated to styrene followed by the conversion to benzaldehyde and benzoic acid by a putative laccase [53]. In the yeast Yarrowia lipolytica OKYL029, cinnamic acid is hydroxylated to p-coumaric acid and further converted to p-hydroxybenzoic acid [54]. Deletion of a cytochrome P450 sharing homology with trans-cinnamate 4-monooxygenases (YALI1_B28430g, TCM1) resulted in a blockage of p-coumaric acid formation however cinnamic acid was still converted through an unknown pathway. The conversion of cinnamic acid to p-coumaric acid was also suggested in A. japonicus [9] and observed in A. niger [42]. However, in the latter also melilotic acid (3-(2-hydroxyphenyl)propanoic acid), o-coumaric acid and p-hydroxybenzoic acid were also detected (Figure 2) [42].
In several fungi other alternative cinnamic acid metabolic pathways have been observed. In Neurospora crassa and Mucor sp. JX23, cinnamic acid is converted towards acetophenone [55,56]. In A. japonicus and Schizophyllum commune, cinnamic acid can be reduced to cinnamaldehyde and cinnamyl alcohol [9,57]. The genes encoding these enzymes still remains to be identified.
4. Metabolism of Benzoic Acid and Related Aromatic Compounds
In fungi, multiple aromatic compounds are metabolized and funneled towards protocatechuic acid [8,51]. In A. niger, all p-hydroxyphenyl and 3,4-dihydroxyphenyl units, which are derived from H-unit monolignol, are converted towards protocatechuic acid, indicating that this pathway plays an important role in the degradation of hydroxylated aromatic compounds in fungi.
One of the aromatic compound that is converted to protocatechuic acid is benzoic acid (benzene carboxylic acid), a commonly used food preservative with anti-microbial properties [58,59]. In multiple Aspergillus species, benzoic acid can be hydroxylated by benzoate-4-monooxygenase (BphA) to p-hydroxybenzoic acid [9,52,60,61,62,63,64]. This pathway is continued by conversion of p-hydroxybenzoic acid to protocatechuic acid catalyzed by PhhA, with further conversion to 3-carboxy-cis,cis-muconic acid by PrcA [52,63]. Interestingly, deletion of prcA in A. niger did not abolish growth on protocatechuic acid, indicating that an alternative pathway is present. Protocatechuic acid can also be converted to hydroxyquinol (1,2,4-trihydroxybenzene) by protocatechuate hydroxylase (PhyA) and is further converted by hydroxyquinol 1,2-dioxygenase (HqdA) [51,52]. Deletion of phyA did not result in growth reduction on protocatechuic acid, revealing that this metabolic pathway may have a minor role in protocatechuic acid metabolism, as also observed in A. nidulans [65]. In A. niger, benzoic acid can also be hydroxylated to m-hydroxybenzoic acid and further to protocatechuic acid [66]. BphA is not able to meta-hydroxylate benzoic acid to m-hydroxybenzoic acid [67] while deletion of bphA results in severely reduced growth on benzoic acid [52]. Therefore, conversion of benzoic acid to m-hydroxybenzoic acid is likely to be a minor pathway in A. niger.
With this knowledge, an A. niger cell factory was constructed that can accumulate protocatechuic acid from benzoic acid, benzaldehyde, benzyl alcohol, p-anisic acid, p-anisaldehyde, p-anisyl alcohol, p-hydroxybenzoic acid, p-hydroxybenzaldehyde, p-hydroxybenzyl alcohol, m-hydroxybenzoic acid, protocatechuic aldehyde, p-coumaric acid, caffeic acid, cinnamic acid and cinnamyl alcohol [51]. However, accumulation of protocatechuic acid was not observed on ferulic acid, vanillin, veratric acid, p-cresol, or anethole indicating that these are converted through alternative pathways. This study contributed greatly to mapping out which aromatic compounds are ultimately converted into protocatechuic acid (Figure 3).
Several aromatic compounds were not tested for protocatechuic acid accumulation with the A. niger cell factory [51], but were observed in other studies to be converted to protocatechuic acid. In A. niger, the chlorinated derivatives 2-chlorobenzoate and 3-chlorobenzoate are both observed to be converted to p-hydroxybenzoic acid and further to protocatechuic acid [68]. Mandelic acid is also converted to protocatechuic acid with benzoyl formate, benzaldehyde, benzoic acid and p-hydroxybenzoic acid as intermediate [69]. The polyethylene terephthalate (PET) derived compounds; dimethyl-terephtahalate, monomethyl-terephthalate and terephthalate are all suggested to be converted into protocatechuic acid [70]. In A. flavus, conversion of p-coumaryl alcohol to p-coumaric aldehyde followed by conversion to p-coumaric acid followed by the conversion to p-hydroxybenzoic acid and protocatechuic acid was observed [45]. Interestingly, A. japonicus can convert veratric acid into protocatechuic acid [9] but is in A. niger converted to vanillic acid [71]. Aspergillus fumigatus is able to degrade p-cresol which is suggested to subsequently be converted p-hydroxybenzyl alcohol followed by p-hydroxybenzaldehyde, p-hydroxybenzoic acid and finally to protocatechuic acid [72]. In another suggested pathway, p-cresol is converted to 4-methylcatechol, (3,4-dihydroxybenzyl alcohol), protocatechualdehyde and protocatechuic acid which is then converted by prcA to 3-oxoadipate. No evidence was found in A. niger that p-cresol is converted through these pathways (Lubbers and de Vries, 2021). In A. japonicus, p-anisic acid can also be reduced to the corresponding aldehyde and alcohol form [9] but was not observed in A. niger [51]. In Aspergillus sojae, protocatechuic acid is decarboxylated to catechol [64].
5. Metabolism of Guaiacyl Units and Related Aromatic Compounds
Guaiacyl units, a key building block of lignin in gymnosperms (softwoods), feature a methoxy group at the meta-position and a hydroxyl group at the para position on their aromatic ring (Figure 4). Coniferyl alcohol, ferulic acid, guaiacol, vanillic acid and vanillin are all guaiacyl units. As mentioned previously, in A. niger ferulic acid is converted to vanillic acid through the CoA-dependent beta-oxidative metabolic pathway. Recently, the gene and enzyme, vanillate hydroxylase A (VhyA), involved in the conversion of vanillic acid to methoxyhydroquinone was identified [71]. This pathway was also observed in A. flavus, Paecilomyces variotii and Sporotrichum pulverulentum [73,75,76]. Methoxyhydroquinone is further processed through ring cleavage by methoxyhydroquinone 1,2-dioxygenase (MhdA) [71]. Deletion of vhyA in A. niger showed that, when grown on coniferyl alcohol, ferulic acid, vanillin, vanillyl alcohol, veratryl alcohol, veratric aldehyde or veratric acid, the fungus converts these substrates into vanillic acid, which subsequently accumulates due to the deletion of vhyA [71].
In A. niger, vanillin is converted to vanillic acid by vanillin dehydrogenase (VdhA) (Figure 4). Deletion of vdhA abolished growth on vanillin and vanillin did not accumulate in the ΔvdhA mutant when grown on ferulic acid or coniferyl alcohol, indicating that ferulic acid is not degraded to vanillin in A. niger. It has been shown in A. niger and A. japonicus that vanillin can also be reduced to vanillyl alcohol [9,71]. In A. luchuensis, vanillin is converted to vanillic acid but is also converted vanillin-glucoside [24]. Aspergillus carbonarius and several Trichoderma species have been observed to produce vanillin from vanillic acid while none of the other tested Aspergillus and Penicillium species were not able to do this conversion and indicates that they are lacking a vanillate reductase [77]. Demethylation of vanillin to p-hydroxybenzaldehyde was observed in A. japonicus [9].
Other pathways have been observed in fungi in which vanillic acid is decarboxylated to guaiacol, protocatechuic acid, vanillin or/and vanillyl alcohol [77]. Interestingly, A. flavus, A. niger and A. nidulans are only able to convert vanillic acid to methoxyhydroquinone while multiple Aspergillus and Penicillium species are able to convert vanillic acid to methoxyhydroquinone but also to protocatechuic acid or guaiacol.
In A. japonicus, it was suggested that veratric acid is demethylated to vanillic acid followed by an additional demethylation to protocatechuic acid (Figure 4) [9]. Conversion of vanillic acid to protocatechuic acid has also been reported in F. solani [28]. In A. niger, veratric acid, veratric aldehyde and veratryl alcohol are converted to vanillic acid but the conversion towards protocatechuic acid was not observed [51,71]. The gene encoding veratric acid demethylase remains to be identified. As observed with other aromatic acids, reduction to its aldehyde and alcohol form has been observed, both A. flavus and A. japonicus are able to reduce veratric acid towards veratric aldehyde and veratryl alcohol [9,73].
6. Conversion of Syringyl Units
While syringyl units, in combination with guaiacyl units, are the predominant building blocks of lignin in hardwoods, the degradation of syringyl units by fungi remains less understood. Syringyl units such as sinapyl alcohol, sinapic acid, syringic acid and syringic aldehyde, feature two methoxy groups at both meta positions and a hydroxyl group at the para position on their aromatic ring (Figure 5). It is observed that A. niger can degrade sinapic acid (3,5-dimethoxy-4-hydroxycinnamic acid) and syringic acid (3,5-dimethoxy-4-hydroxybenzoic acid) (Lubbers et al., unpublished data). However, it remains unknown how these are degraded. Sinapic acid is not degraded through the CoA-dependent beta oxidative pathway since deletion of hcsA, foxA, or katA did not result in any growth phenotype [34]. No other new studies have become available concerning the degradation of syringyl units by Aspergilli and therefore the metabolic pathways remains unknown.
7. Gallic Acid Metabolic Pathways
Gallic acid (3,4,5-trihydroxybenzoic acid) is a commonly observed metabolite in the degradation of syringyl units [8] and is part of the polymeric aromatic compound tannic acid. It has been shown that many Aspergillus species can release gallic acid from tannic acid using tannases [78,79,80]. Gallic acid has anti-oxidative and antimicrobial properties and is applied in cosmetic, food and beverage, agricultural and pharmaceutical industry [81].
Currently, not many gallic acid metabolic pathways have been observed in fungi [8]. In A. oryzae, gallic acid is converted to pyrogallol (1,2,3-trihydroxybenzene), progallin A (ethyl 3,4,5-trihydroxybenzoate) and methyl gallate (Figure 5) [82]. It was suggested that these compounds were further processed by a ring opening reaction. In A. niger and A. nidulans, gallic acid is converted into 1,2,3,5-tetrahydroxybenzene, catalyzed by PhyA [65,83]. Deletion of 17 putative dioxygenase genes, including prcA, hqdA, and crcA, did not impair growth on gallic acid, suggesting that gallic acid is metabolized through an alternative pathway [83]. In A. nidulans, it has been proposed that 1,2,3,5-tetrahydroxybenzene is further converted to 5-hydroxydienelactone by a DUF3500 containing protein (AN10530) functioning as a putative dioxygenase [65]. Homologs of this gene are present in A. niger and remain to be studied.
8. Salicylic Acid Metabolic Pathways
Salicylic acid (2-hydroxybenzoic acid) is a well-known signaling molecule in plants and is involved the defense against pathogens [84]. Currently, not many different fungal salicylic acid metabolic pathways have been elucidated. The most observed conversion is the hydroxylation of the carboxylic group of salicylic acid resulting in the formation of catechol (1,2-dihydroxybenzene). This conversion is catalyzed by the salicylic acid hydroxylase (ShyA) [85]. This conversion was observed in several aspergilli including A. niger, A. japonicus, A. nidulans and A. terreus but also in other fungi such as Epichloë festucae and Sclerotinia sclerotiorum (Figure 6) [9,63,85,86,87,88]. Alternatively, it has been proposed in A. nidulans and Fusarium graminearum that salicylic acid can also be hydroxylated to 2,3-dihydroxybenzoic acid (o-pyrocatechuic acid) [63,89] followed by a decarboxylation step to catechol catalyzed by 2,3-dihydroxybenzoic acid decarboxylase (DhbA) [85,90,91]. In A. niger, no evidence was found for the hydroxylation of salicylic acid to 2,3-dihydroxybenzoic acid since the deletion of shyA resulted in abolished growth on salicylic acid and deletion of dhbA did not result in reduced growth on salicylic acid [85]. Deletion of catechol 1,2-dioxygenase (CrcA) revealed that salicylic acid, 2,3-dihydroxybenzoic acid and catechol are converted towards cis,cis-muconic acid. Cis-cis-muconic acid is an interesting compound since it a valuable chemical building block that can be used for polymer and drug production [92]. In A. fumigatus, phenol can be ortho-hydroxylated to catechol and cis,cis-muconic acid but also para-hydroxylated to hydroquinone (1,4-dihydoxybenzene) and hydroxyquinol [72].
Recently, an additional salicylic metabolic pathway was observed in A. terreus in which salicylic acid is hydroxylated to gentisic acid (2,5-dihydroxybenzoic acid) (Figure 6) [86]. Gentisic acid is further converted to maleylpyruvate by a putative gentisate 1,2-dioxygenase (ATEG_06714). In N. crassa, salicylic acid can also be reduced to salicylic aldehyde and salicyl alcohol [93]. Those pathways had not been observed in Aspergilli.
9. Ring Cleavage Pathways
The cleavage of the aromatic ring is a critical step in the detoxification and utilization of aromatic compounds as a carbon source. In fungi, most aromatic compounds are mainly converted towards catechol, hydroxyquinol, gallic acid, gentisic acid, protocatechuic acid or pyrogallol. These aromatic compounds are targeted by dioxygenases that catalyze the ring cleavage. The cleaved compounds are converted in multiple steps to acetyl-CoA, fumarate, oxaloacetate, pyruvate, or succinate and enter the TCA cycle [8,94]. In fungi, most aromatic compounds are cleaved by intradiol dioxygenases and uses non-heme Fe(III) to cleave the aromatic nucleus ortho to the hydroxyl substituents [95]. Extradiol dioxygenases using non-heme Fe(II) or other divalent metal ions to cleave the aromatic nucleus meta to the hydroxyl substituents, are less common in fungi [8].
Recently, multiple intradiol dioxygenases have been described in A. niger. Currently, four dioxygenases have been studied i.e. the catechol 1,2-dioxygenase (CrcA), hydroxyquinol 1,2-dioxygenase (HqdA), protocatechuic 3,4-dioxygenase (PrcA) and NRRL3_05330, a putative hydroxyquinol 1,2-dioxygenase [51,52,85,96]. Also, a methoxyhydroquinone 1,2-dioxygenase (MhdA) was identified which shares similarities with homogentisate 1,2-dioxygenases [71]. The role of NRRL3_05330 remains unknown since deletion did not result in any clear phenotype [85].
Protocatechuic acid is one of the main intermediates in the degradation of hydroxy phenolic compounds. It has been shown for several Aspergilli, that the aromatic ring of protocatechuic acid is cleaved into 3-carboxy-cis,cis-muconic acid and further converted through the β-ketoadipate pathway to acetyl-CoA and succinyl-CoA [63,94]. Several genes in this pathway were identified in A. nidulans and deletion of these genes resulted in reduced growth on protocatechuic acid [63]. More recently, the β-ketoadipate pathway has been further studied in A. niger resulting in the identification of four enzymes (3-carboxy-cis,cis-muconate cyclase (CmcA; NRRL3_02586), 3-carboxymuconolactone hydrolase/decarboxylase (ChdA; NRRL3_01409), β-ketoadipate:succinyl-CoA transferase (KstA; NRRL3_01886) and β-ketoadipyl-CoA thiolase (KctA; NRRL3_01526)) [94]. In addition, an essential protein (NRRL3_00837) was found involved in the β-ketoadipate pathway but its function remains unknown [94].
In A. nidulans and A. niger, catechol is cleaved by CrcA into cis,cis-muconic acid with subsequential conversion towards 3-oxoadipate. In A. nidulans, several genes involved in the degradation of catechol have been identified with whole-genome transcriptomics and proteomics [63]. Deletion of AN3895 (muconate isomerase), AN4061 (muconolactone isomerase), AN4531 (3-oxoadipate enol-lactone hydrolase) results in the formation of cis,cis-muconic acid, muconolactone and 3-oxoadipate enol-lactone, respectively, when grown on salicylic acid. Homologs of these genes were also identified in A. niger with whole-genome transcriptomics and were strongly induced by salicylic acid [85].
10. Regulation of Aromatic Pathways
Transcriptional regulators are important for the regulation of metabolic pathways in fungi. Many transcription factors involved in the regulation of sugar metabolic pathways have been described [97]. However, only a few transcriptional regulators involved in the regulation of metabolic pathways for aromatic compounds have been identified. Recently, a regulator/repressor (TanR/TanX) complex is involved in the regulation of gallic acid metabolic genes has been identified [83]. Deletion of tanR resulted in abolished growth on gallic acid and tannic acid indicating that this transcription factor regulates genes involved in the regulation of genes involved in the degradation of gallic acid and tannic acid. Whole-genome transcriptomic data of A. niger ΔtanX grown on fructose, revealed that phyA is 2300 times more expressed than the parental strain. As mentioned above, cinnamic/sorbic acid metabolic pathway genes cdcA and padA are regulated by SdrA. More recently, it was shown that the weak acid regulator A and B (WarA and WarB) also play a role in the degradation of cinnamic acid and sorbic acid, however the exact role of WarA and WarB needs to be further studied [98].
Fatty acid regulator A (FarA) has a role in the degradation of fatty acids [99,100] but also hydroxycinnamic acids in A. niger [101]. Deletion of farA results in reduced growth on p-coumaric acid and abolished growth on ferulic acid and caffeic acid and the fatty acids valeric acid and oleic acid. A new far-like protein (FarD) was identified and deletion of it results in abolished growth on caffeic acid and ferulic acid and reduce growth on p-coumaric acid and p-hydroxybenzoic acid. Deletion of farA in A. nidulans results in reduced expression of foxA [99], which is also important for the degradation of hydroxycinnamic acids [34]. A putative FarA binding site has been observed in the promoter of hscA [101]. The exact role of FarA in the degradation of aromatic compounds need to be studied further but it is highly likely that it regulates multiple genes involved in the CoA-dependent beta-oxidative metabolic pathway.
Recently, it was observed that the regulator FarB has a role in the degradation of benzoic acid, vanillic acid and the short fatty acid valeric acid, but not on p-coumaric acid, caffeic acid, protocatechuic acid or p-hydroxybenzoic acid [101]. It is possible that FarB is regulating bphA since no phenotypes are not observed on p-hydroxybenzoic acid and protocatechuic acid. Currently, it is remains unknown which genes are regulated by FarB.
11. Future Perspectives; Strategies for Studying and Identifying Aromatic Metabolic Pathways in Filamentous Fungi
The aromatic metabolic pathways of filamentous fungi belonging to the Aspergillus family have been better studied compared to other fungal species. However, still many aromatic pathways are have gaps or are unstudied. In addition, many of the enzymes involved in these pathways remain to be identified.
To study aromatic metabolic pathways in fungi, deletion of the ring cleaving dioxygenases is a straight forward approach, as these enzymes play critical roles in the metabolic pathway and deleting them causes severe phenotypes on aromatic compounds (Table 1) [8,65,71,83]. In A. niger, deletion of prcA gave clear phenotypes on many aromatic compounds related to the H-unit, while deletion of hqdA alone did not result in any phenotypes on the tested aromatic compounds [52]. However, the combined deletion of prcA and hqdA resulted in more severe phenotypes and even revealed the existence of an alternative pathway in A. niger. Deletion of mhdA, revealed that aromatic compound related to the G-unit are degraded to methoxyhydroquinone [71]. This pathway is important to created aromatic compounds such as vanillin and vanillic acid. Deletion of crcA, revealed that salicylic acid, 2,3-dihydroxybenzoic acid are degraded to catechol and that there is no alternative enzyme or pathway present in A. niger [85]. It appears that in A. niger, ortho-hydroxylated benzoic acids are converted to catechol, while meta- and para-hydroxylated benzoic acids are converted to protocatechuic acid, and hydroxyquinol. Meta-methoxylated compounds such as ferulic acid, vanillic acid and veratric acid are converted to methoxyhydroquinone.
For the identification of genes encoding aromatic compound converting enzymes, whole genome transcriptomics is a strong tool that can aid in the identification of genes after exposure to aromatic compounds. Currently, most genes involved in the aromatic metabolism of A. niger and A. nidulans were identified using this approach [34,52,63,65,71,83,86]. In A. niger, a short exposure of 2h to an aromatic compound can results in a strong induction of aromatic compound metabolic genes. For example, salicylic acid metabolic genes; shyA, dhbA and crcA, are strongly upregulated by salicylic acid and by not p-hydroxyphenyl or guaiacylic aromatic compounds such as benzoic acid or p-coumaric acid, ferulic acid or vanillic acid. Similar observations were made for vhyA and mhdA, that are strongly induced by guaiacylic aromatic compounds while the CoA-dependent β-oxidative genes hcsA, foxA, katA, theA, theB, theC and theD are strongly induced by hydroxycinnamic acids. This demonstrated that these genes are not only rapidly upregulated but also induced by specific aromatic compounds.
12. Conclusions
In this review, we highlight recent advances in understanding aromatic metabolism in Aspergillus niger. It is important to note that Aspergillus species, in general, possess a remarkable diversity of aromatic metabolic pathways, reflecting their adaptability to different environmental conditions and substrates. Given the already substantial versatility observed in Aspergillus species, it is anticipated that even greater metabolic diversity will be uncovered when studying basidiomycetes, which are known for their complex and highly specialized systems for degrading aromatic compounds such as lignin.
Further exploration of fungal aromatic metabolism will not only enhance biotechnological production but also uncover new opportunities for bio-based solutions in pharmaceuticals, food, and fragrance industries. Currently, research is primarily focused on the production of aromatic compounds such as vanillin, gallic acid, catechol, and protocatechuic acid using microorganisms, given their widespread applications across various industries [81,102,103,104]. Advances in metabolic engineering, synthetic biology, and systems biology will enable the optimization of fungal strains for higher yields and broader compound diversity. Additionally, a deeper understanding of regulatory networks and enzyme mechanisms involved in aromatic biosynthesis could lead to innovative bioprocesses, reducing reliance on petrochemical-based production and promoting sustainability. As research continues to uncover novel and expand fungal aromatic metabolic pathways, the potential for industrial applications will continue to expand, reinforcing fungi as valuable cell factories for aromatic compounds.
Data Availability Statement
Not applicable.
Acknowledgments
I thank Arthur F. J. Ram and Han de Winde (Department of Molecular Biotechnology, Institute for Biology, Leiden University) for their support, discussions and revisions on the manuscript.
Conflicts of Interest
The author declare no conflicts of interest.
References
- Li, Z.H.; Wang, Q.; Ruan, X.; Pan, C. De; Jiang, D.A. Phenolics and plant allelopathy. Molecules 2010, 15, 8933–8952. [Google Scholar] [CrossRef] [PubMed]
- Myers, R.L. The 100 Most Important Chemical Compounds. 100 Most Important Chem. Compd. 2007. [Google Scholar] [CrossRef]
- Chen, F.; Tobimatsu, Y.; Havkin-Frenkel, D.; Dixon, R.A.; Ralph, J. A polymer of caffeyl alcohol in plant seeds. Proc. Natl. Acad. Sci. U. S. A. 2012, 109, 1772–1777. [Google Scholar] [CrossRef]
- Zhao, L.; Zhang, J.; Zhao, D.; Jia, L.; Qin, B.; Cao, X.; Zang, L.; Lu, F.; Liu, F. Biological degradation of lignin: A critical review on progress and perspectives. Ind. Crops Prod. 2022, 188, 115715. [Google Scholar] [CrossRef]
- Atiwesh, G.; Parrish, C.C.; Banoub, J.; Le, T.A.T. Lignin degradation by microorganisms: A review. Biotechnol. Prog. 2022, 38. [Google Scholar] [CrossRef] [PubMed]
- Madadi, M.; Abbas, A. Lignin Degradation by Fungal Pretreatment: A Review. J. Plant Pathol. Microbiol. 2017, 08. [Google Scholar] [CrossRef]
- Pollegioni, L.; Tonin, F.; Rosini, E. Lignin-degrading enzymes. FEBS J. 2015, 282, 1190–1213. [Google Scholar] [CrossRef]
- Lubbers, R.J.M.; Dilokpimol, A.; Visser, J.; Mäkelä, M.R.; Hildén, K.S.; de Vries, R.P. A comparison between the homocyclic aromatic metabolic pathways from plant-derived compounds by bacteria and fungi. Biotechnol. Adv. 2019, 37, 0–1. [Google Scholar] [CrossRef] [PubMed]
- Milstein, O.; Vered, Y.; Shragina, L.; Gressel, J.; Flowers, H.M.; Hüttermann, A. Metabolism of lignin related aromatic compounds by Aspergillus japonicus. Arch. Microbiol. 1983, 135, 147–154. [Google Scholar] [CrossRef]
- Mäkelä, M.R.; Marinović, M.; Nousiainen, P.; Liwanag, A.J.M.; Benoit, I.; Sipilä, J.; Hatakka, A.; de Vries, R.P.; Hildén, K.S. Aromatic metabolism of filamentous fungi in relation to the presence of aromatic compounds in plant biomass. Adv. Appl. Microbiol. 2015, 91, 63–137. [Google Scholar] [CrossRef]
- Ralet, M.C.; Thibault, J.F.; Faulds, C.B.; Williamson, G. Isolation and purification of feruloylated oligosaccharides from cell walls of sugar-beet pulp. Carbohydr. Res. 1994, 263, 227–241. [Google Scholar] [CrossRef]
- Colquhoun, I.J.; Ralet, M.C.; Thibault, J.F.; Faulds, C.B.; Williamson, G. Structure identification of feruloylated oligosaccharides from sugar-beet pulp by NMR spectroscopy. Carbohydr. Res. 1994, 263, 243–256. [Google Scholar] [CrossRef] [PubMed]
- Ralet, M.C.; Faulds, C.B.; Williamson, G.; Thibault, J.F. Degradation of feruloylated oligosaccharides from sugar-beet pulp and wheat bran by ferulic acid esterases from Aspergillus niger. Carbohydr. Res. 1994, 263, 257–269. [Google Scholar] [CrossRef]
- Faulds, C.B.; Williamson, G. Release of ferulic acid from wheat bran by a ferulic acid esterase (FAE-III) from Aspergillus niger. Appl. Microbiol. Biotechnol. 1995, 43, 1082–1087. [Google Scholar] [CrossRef] [PubMed]
- Bonnin, E.; Saulnier, L.; Brunel, M.; Marot, C.; Lesage-Meessen, L.; Asther, M.; Thibault, J.F. Release of ferulic acid from agroindustrial by-products by the cell wall-degrading enzymes produced by Aspergillus niger I-1472. Enzyme Microb. Technol. 2002, 31, 1000–1005. [Google Scholar] [CrossRef]
- Williamson, G.; Faulds, C.B.; Kroon, P.A. Specificity of ferulic acid (feruloyl) esterases. Biochem. Soc. Trans. 1998, 26, 205–209. [Google Scholar] [CrossRef]
- Dilokpimol, A.; Mäkelä, M.R.; Aguilar-Pontes, M.V.; Benoit-Gelber, I.; Hildén, K.S.; de Vries, R.P. Diversity of fungal feruloyl esterases: updated phylogenetic classification, properties, and industrial applications. Biotechnol. Biofuels 2016, 9, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Dilokpimol, A.; Mäkelä, M.R.; Mansouri, S.; Belova, O.; Waterstraat, M.; Bunzel, M.; de Vries, R.P.; Hildén, K.S. Expanding the feruloyl esterase gene family of Aspergillus niger by characterization of a feruloyl esterase, FaeC. N. Biotechnol. 2017, 37, 200–209. [Google Scholar] [CrossRef]
- de Vries, R.P.; Visser, J. Regulation of the feruloyl esterase (faeA) gene from Aspergillus niger. Appl. Environ. Microbiol. 1999, 65, 5500–5503. [Google Scholar] [CrossRef]
- de Vries, R.P.; vanKuijk, Patricia, A.; C.M. Kester, H.; Visser, J. The Aspergillus niger faeB gene encodes a second feruloyl esterase involved in pectin and xylan degradation and is specifically induced in the presence of aromatic compounds. 2002, 386, 377–386.
- Todokoro, T.; Negoro, H.; Kotaka, A.; Hata, Y.; Ishida, H. Aspergillus oryzae FaeA is responsible for the release of ferulic acid, a precursor of off-odor 4-vinylguaiacol in sake brewing. J. Biosci. Bioeng. 2022, 133, 140–145. [Google Scholar] [CrossRef] [PubMed]
- Maeda, M.; Tokashiki, M.; Tokashiki, M.; Uechi, K.; Ito, S.; Taira, T. Characterization and induction of phenolic acid decarboxylase from Aspergillus luchuensis. J. Biosci. Bioeng. 2018, 126, 162–168. [Google Scholar] [CrossRef]
- Maeda, M.; Motosoko, M.; Tokashiki, T.; Tokashiki, J.; Mizutani, O.; Uechi, K.; Goto, M.; Taira, T. Phenolic acid decarboxylase of Aspergillus luchuensis plays a crucial role in 4-vinylguaiacol production during awamori brewing. J. Biosci. Bioeng. 2020, 130, 352–359. [Google Scholar] [CrossRef] [PubMed]
- Taira, J.; Toyoshima, R.; Ameku, N.; Iguchi, A.; Tamaki, Y. Vanillin production by biotransformation of phenolic compounds in fungus, Aspergillus luchuensis. AMB Express 2018, 8. [Google Scholar] [CrossRef] [PubMed]
- Edlin, D.A.N.; Narbad, A.; Dickinson, J.R.; Lloyd, D. The biotransformation of simple phenolic compounds by Brettanomyces anomalus. FEMS Microbiol. Lett. 1995, 125, 311–315. [Google Scholar] [CrossRef]
- Linke, D.; Riemer, S.J.L.; Schimanski, S.; Nieter, A.; Krings, U.; Berger, R.G. Cold generation of smoke flavour by the first phenolic acid decarboxylase from a filamentous ascomycete – Isaria farinosa. Fungal Biol. 2017, 121, 763–774. [Google Scholar] [CrossRef]
- Baqueiro-Peña, I.; Rodríguez-Serrano, G.; González-Zamora, E.; Augur, C.; Loera, O.; Saucedo-Castañeda, G. Biotransformation of ferulic acid to 4-vinylguaiacol by a wild and a diploid strain of Aspergillus niger. Bioresour. Technol. 2010, 101, 4721–4724. [Google Scholar] [CrossRef]
- Nazareth, S.; Mavinkurve, S. Degradation of ferulic acid via 4-vinylguaiacol by Fusarium solani (Mart.) Sacc. Can. J. Microbiol. 1986, 32, 494–497. [Google Scholar] [CrossRef]
- Zhang, P.H.; Yu, X.Y.; Weng, L.X.; Sun, L.L.; Mao, Z.C.; Zhang, Y.L. Degradation of ferulic acid by the endophytic fungus Colletotrichum gloeosporioides TMTM-13 associated with Ostrya rehderiana Chun. ACS Omega 2019, 4, 21000–21004. [Google Scholar] [CrossRef]
- Plumridge, A.; Melin, P.; Stratford, M.; Novodvorska, M.; Shunburne, L.; Dyer, P.S.; Roubos, J.A.; Menke, H.; Stark, J.; Stam, H.; et al. The decarboxylation of the weak-acid preservative, sorbic acid, is encoded by linked genes in Aspergillus spp. Fungal Genet. Biol. 2010, 47, 683–692. [Google Scholar] [CrossRef]
- Plumridge, A.; Stratford, M.; Lowe, K.C.; Archer, D.B. The weak-acid preservative sorbic acid is decarboxylated and detoxified by a phenylacrylic acid decarboxylase, PadA1, in the spoilage mold Aspergillus niger. Appl. Environ. Microbiol. 2008, 74, 550–552. [Google Scholar] [CrossRef]
- Lubbers, R.J.M.; Dilokpimol, A.; Navarro, J.; Peng, M.; Wang, M.; Lipzen, A.; Ng, V.; Grigoriev, I. V.; Visser, J.; Hildén, K.S.; et al. Cinnamic acid and sorbic acid conversion are mediated by the same transcriptional regulator in Aspergillus niger. Front. Bioeng. Biotechnol. 2019, 7, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Tian, G.; Liu, Y. Mechanistic insights into the catalytic reaction of ferulic acid decarboxylase from Aspergillus niger: A QM/MM study. Phys. Chem. Chem. Phys. 2017, 19, 7733–7742. [Google Scholar] [CrossRef] [PubMed]
- Lubbers, R.J.M.; Dilokpimol, A.; Visser, J.; de Vries, R.P. Aspergillus niger uses the peroxisomal CoA-dependent β-oxidative genes to degrade the hydroxycinnamic acids caffeic acid, ferulic acid, and p-coumaric acid. Appl. Microbiol. Biotechnol. 2021, 105, 4199–4211. [Google Scholar] [CrossRef]
- Widhalm, J.R.; Dudareva, N. A familiar ring to it: Biosynthesis of plant benzoic acids. Mol. Plant 2015, 8, 83–97. [Google Scholar] [CrossRef] [PubMed]
- Fosså, A.; Beyer, A.; Pfitzner, E.; Wenzel, B.; Kunau, W.H. Molecular cloning, sequencing and sequence analysis of the fox-2 gene of Neurospora crassa encoding the multifunctional β-oxidation protein. MGG Mol. Gen. Genet. 1995, 247, 95–104. [Google Scholar] [CrossRef]
- Requena, N.; Füller, P.; Franken, P. Molecular characterization of GmFOX2, an evolutionarily highly conserved gene from the mycorrhizal fungus Glomus mosseae, down-regulated during interaction with rhizobacteria. Mol. Plant-Microbe Interact. 1999, 12, 934–942. [Google Scholar] [CrossRef]
- Maggio-Hall, L.A.; Keller, N.P. Mitochondrial β-oxidation in Aspergillus nidulans. Mol. Microbiol. 2004, 54, 1173–1185. [Google Scholar] [CrossRef]
- Klose, J.; Kronstad, J.W. The multifunctional β-oxidation enzyme is required for full symptom development by the biotrophic maize pathogen Ustilago maydis. Eukaryot. Cell 2006, 5, 2047–2061. [Google Scholar] [CrossRef]
- Wang, Z.Y.; Soanes, D.M.; Kershaw, M.J.; Talbot, N.J. Functional analysis of lipid metabolism in Magnaporthe grisea reveals a requirement for peroxisomal fatty acid β-oxidation during appressorium-mediated plant infection. Mol. Plant-Microbe Interact. 2007, 20, 475–491. [Google Scholar] [CrossRef]
- Boisnard, S.; Espagne, E.; Zickler, D.; Bourdais, A.; Riquet, A.L.; Berteaux-Lecellier, V. Peroxisomal ABC transporters and β-oxidation during the life cycle of the filamentous fungus Podospora anserina. Fungal Genet. Biol. 2009, 46, 55–66. [Google Scholar] [CrossRef]
- Bocks, S. . Fungal Metabolism-I. the transformations of coumarin, o-coumaric acid and trans-cinnamic acid by Aspergillus niger. Phytochemistry 1967, 6, 127–130. [Google Scholar] [CrossRef]
- Bye, A.; King, H.K. The biosynthesis of 4-hydroxycoumarin and dicoumarol by Aspergillus fumigatus Fresenius. Biochem. J. 1970, 117, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Bye, A.; Ashton, W.M.; King, H.K. A proposed mechanism for the conversion of o-coumaric acid to 4-hydroxy-coumarin in Aspergillus fumigatus fresenius. Biochem. Biophys. Res. Commun. 1968, 32, 94–97. [Google Scholar] [CrossRef] [PubMed]
- Iyayi, C.B.; Dart, R.K. The degradation of p-coumaryl alcohol by Aspergillus flavus. J. Gen. Microbiol. 1982, 128, 1473–1482. [Google Scholar] [CrossRef]
- Catarine Santos Rodrigues, C.; Latércia Tranches Dias, A.; de Oliveira Silva, E. Unprecedented derivatization of ferulic acid through selective methoxylation by Aspergillus brasiliensis ATCC 16404. Biocatal. Biotransformation 2019, 37, 233–237. [Google Scholar] [CrossRef]
- Pinches, S.E.; Apps, P. Production in food of 1,3-pentadiene and styrene by Trichoderma species. Int. J. Food Microbiol. 2007, 116, 182–185. [Google Scholar] [CrossRef]
- Pagot, Y.; Belin, J.M.; Husson, F.; Spinnler, H.E. Metabolism of phenylalanine and biosynthesis of styrene in Penicillium camemberti. J. Dairy Res. 2007, 74, 180–185. [Google Scholar] [CrossRef]
- Lafeuille, J.L.; Buniak, M.L.; Vioujas, M.C.; Lefevre, S. Natural formation of styrene by cinnamon mold flora. J. Food Sci. 2009, 74. [Google Scholar] [CrossRef]
- Stratford, M.; Plumridge, A.; Pleasants, M.W.; Novodvorska, M.; Baker-Glenn, C.A.G.; Pattenden, G.; Archer, D.B. Mapping the structural requirements of inducers and substrates for decarboxylation of weak acid preservatives by the food spoilage mould Aspergillus niger. Int. J. Food Microbiol. 2012, 157, 375–383. [Google Scholar] [CrossRef]
- Lubbers, R.J.M.; de Vries, R.P. Production of protocatechuic acid from p-hydroxyphenyl (H) units and related aromatic compounds using an Aspergillus niger cell factory. MBio 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Lubbers, R.J.M.; Dilokpimol, A.; Peng, M.; Visser, J.; Mäkelä, M.R.; Hildén, K.S.; De Vries, R.P. Discovery of novel p-hydroxybenzoate-m-hydroxylase, protocatechuate 3,4 ring-cleavage dioxygenase, and hydroxyquinol 1,2 ring-cleavage dioxygenase from the filamentous fungus Aspergillus niger. ACS Sustain. Chem. Eng. 2019, 7, 19081–19089. [Google Scholar] [CrossRef]
- Xie, X.G.; Dai, C.C. Biodegradation of a model allelochemical cinnamic acid by a novel endophytic fungus Phomopsis liquidambari. Int. Biodeterior. Biodegrad. 2015, 104, 498–507. [Google Scholar] [CrossRef]
- Konzock, O.; Tous-Mohedano, M.; Cibin, I.; Chen, Y.; Norbeck, J. Cinnamic acid and p-coumaric acid are metabolized to 4-hydroxybenzoic acid by Yarrowia lipolytica. AMB Express 2023, 13. [Google Scholar] [CrossRef]
- Said, S.; Neves, F.M.; Griffiths, A.J.F. Cinnamic acid inhibits the growth of the fungus Neurospora crassa, but is eliminated as acetophenone. Int. Biodeterior. Biodegrad. 2004, 54, 1–6. [Google Scholar] [CrossRef]
- Ma, L.; Liu, X.; Liang, J.; Zhang, Z. Biotransformations of cinnamaldehyde, cinnamic acid and acetophenone with Mucor. World J. Microbiol. Biotechnol. 2011, 27, 2133–2137. [Google Scholar] [CrossRef]
- Nimura, Y.; Tsujiyama, S. ichi; Ueno, M. Bioconversion of cinnamic acid derivatives by Schizophyllum commune. J. Gen. Appl. Microbiol. 2010, 56, 381–387. [Google Scholar] [CrossRef] [PubMed]
- Krebs, H.A.; Wiggins, D.; Stubbs, M. Studies on the mechanism of the antifungal action of benzoate. Biochem. J. 1983, 214, 657–663. [Google Scholar] [CrossRef]
- del Olmo, A.; Calzada, J.; Nuñez, M. Benzoic acid and its derivatives as naturally occurring compounds in foods and as additives: Uses, exposure, and controversy. Crit. Rev. Food Sci. Nutr. 2017, 57, 3084–3103. [Google Scholar] [CrossRef]
- van Gorcom, R.F.M.; Boschloo, J.G.; Kuijvenhoven, A.; Lange, J.; van Vark, A.J.; Bos, C.J.; van Balken, J.A.M.; Pouwels, P.H.; van den Hondel, C.A.M.J.J. Isolation and molecular characterisation of the benzoate-para-hydroxylase gene (bphA) of Aspergillus niger: A member of a new gene family of the cytochrome P450 superfamily. MGG Mol. Gen. Genet. 1990, 223, 192–197. [Google Scholar] [CrossRef]
- Boschloo, J.G.; Moonen, E.; van Gorcom, R.F.M.; Hermes, H.F.M.; Bos, C.J. Genetic analysis of Aspergillus niger mutants defective in benzoate-4-hydroxylase function. Curr. Genet. 1991, 19, 261–264. [Google Scholar] [CrossRef]
- Boschloo, J.G.; Paffen, A.; Koot, T.; van den Tweel, W.J.J.; van Gorcom, R.F.M.; Cordewener, J.H.G.; Bos, C.J. Genetic analysis of benzoate metabolism in Aspergillus niger. Appl. Microbiol. Biotechnol. 1990, 34, 225–228. [Google Scholar] [CrossRef]
- Martins, T.M.; Hartmann, D.O.; Planchon, S.; Martins, I.; Renaut, J.; Silva Pereira, C. The old 3-oxoadipate pathway revisited: New insights in the catabolism of aromatics in the saprophytic fungus Aspergillus nidulans. Fungal Genet. Biol. 2015, 74, 32–44. [Google Scholar] [CrossRef]
- Yuasa, K.; Ishlzuka, K.; Kaburaki, S.; Sakasal, T. Metabolism of phenylalanine in Aspergillus sojae. Agric. Biol. Chem. 1975, 39, 2199–2206. [Google Scholar] [CrossRef]
- Martins, T.M.; Bento, A.; Martins, C.; Tomé, A.S.; Moreira, C.J.S.; Silva Pereira, C. Bringing up to date the toolkit for the catabolism of aromatic compounds in fungi: The unexpected 1,2,3,5-tetrahydroxybenzene central pathway. Microb. Biotechnol. 2024, 17, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Bocks, S.M. Fungal metabolism-III. The hydroxylation of anisole, phenoxyacetic acid, phenylacetic acid and benzoic acid by Aspergillus niger. Phytochemistry 1967, 6, 785–789. [Google Scholar] [CrossRef]
- Faber, B.W.; Van Gorcom, R.F.M.; Duine, J.A. Purification and characterization of benzoate-parahydroxylase, a cytochrome P450 (CYP53A1), from Aspergillus niger. Arch. Biochem. Biophys. 2001, 394, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Shailubhai, K.; Sahasrabudhe, S.R.; Vora, K.A.; Modi, V.V. Degradation of chlorinated derivatives of phenoxyacetic acid and benzoic acid by Aspergillus niger. FEMS Microbiol. Lett. 1983, 18, 279–282. [Google Scholar] [CrossRef]
- Jamaluddin, M.; Rao, P. V.; Vaidyanathan, C.S. Involvement of the protocatechuate pathway in the metabolism of mandelic acid by Aspergillus niger. J. Bacteriol. 1970, 101, 786–793. [Google Scholar] [CrossRef]
- Ganji, S.H.; Karigar, C.S.; Pujar, B.G. Metabolism of dimethylterephthalate by Aspergillus niger. Biodegradation 1995, 6, 61–66. [Google Scholar] [CrossRef]
- Lubbers, R.J.M.; Dilokpimol, A.; Nousiainen, P.A.; Cioc, R.C.; Visser, J.; Bruijnincx, P.C.A.; de Vries, R.P. Vanillic acid and methoxyhydroquinone production from guaiacyl units and related aromatic compounds using Aspergillus niger cell factories. Microb. Cell Fact. 2021, 20, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Jones, K.H.; Trudgill, P.W.; Hopper, D.J. Metabolism of p-cresol by the fungus Aspergillus fumigatus. Appl. Environ. Microbiol. 1993, 59, 1125–1130. [Google Scholar] [CrossRef] [PubMed]
- Palazzolo, M.A.; Mascotti, M.L.; Lewkowicz, E.S.; Kurina-Sanz, M. Self-sufficient redox biotransformation of lignin-related benzoic acids with Aspergillus flavus. J. Ind. Microbiol. Biotechnol. 2015, 42, 1581–1589. [Google Scholar] [CrossRef]
- Raman, T.S.; Shanmugasundaram, E.R. Metabolism of some aromatic acids by Aspergillus niger. J. Bacteriol. 1962, 84, 1339–1340. [Google Scholar] [CrossRef]
- Buswell, J.A.; Eriksson, K.E. Vanillate hydroxylase from Sporotrichum pulverulentum. Methods Enzymol. 1988, 161, 274–281. [Google Scholar] [CrossRef]
- Rahouti, M.; Seigle-Murandi, F.; Steiman, R.; Eriksson, K.E. Metabolism of ferulic acid by Paecilomyces variotii and Pestalotia palmarum. Appl. Environ. Microbiol. 1989, 55, 2391–2398. [Google Scholar] [CrossRef] [PubMed]
- Guiraud, P.; Steiman, R.; Seigle-Murandi, F.; Benoit-Guyod, J.L. Metabolism of vanillic acid by Micromycetes. World J. Microbiol. Biotechnol. 1992, 8, 270–275. [Google Scholar] [CrossRef]
- Seth, M.; Chand, S. Biosynthesis of tannase and hydrolysis of tannins to gallic acid by Aspergillus awamori - Optimisation of process parameters. Process Biochem. 2000, 36, 39–44. [Google Scholar] [CrossRef]
- Lokeswari, N.; Jaya Raju, K. Optimization of gallic acid production from Terminalia chebula by Aspergillus niger. E-Journal Chem. 2007, 4, 287–293. [Google Scholar] [CrossRef]
- Mukherjee, G.; Banerjee, R. Biosynthesis of tannase and gallic acid from tannin rich substrates by Rhizopus oryzae and Aspergillus foetidus. J. Basic Microbiol. 2004, 44, 42–48. [Google Scholar] [CrossRef]
- Badhani, B.; Sharma, N.; Kakkar, R. Gallic acid: A versatile antioxidant with promising therapeutic and industrial applications. R. Soc. Chem. Adv. 2015. [Google Scholar] [CrossRef]
- Guo, D.; Zhang, Z.; Liu, D.; Zheng, H.; Chen, H.; Chen, K. A comparative study on the degradation of gallic acid by Aspergillus oryzae and Phanerochaete chrysosporium. Water Sci. Technol. 2014, 70, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Arentshorst, M.; Falco, M. Di; Moisan, M.C.; Reid, I.D.; Spaapen, T.O.M.; van Dam, J.; Demirci, E.; Powlowski, J.; Punt, P.J.; Tsang, A.; et al. Identification of a conserved transcriptional activator-repressor module controlling the expression of genes involved in tannic acid degradation and gallic acid utilization in Aspergillus niger. Front. Fungal Biol. 2021, 2, 1–16. [Google Scholar] [CrossRef]
- Shah, J. The salicylic acid loop in plant defense. Curr. Opin. Plant Biol. 2003, 6, 365–371. [Google Scholar] [CrossRef]
- Lubbers, R.J.M.; Dilokpimol, A.; Visser, J.; Hildén, K.S.; Mäkelä, M.R.; de Vries, R.P. Discovery and functional analysis of a salicylic acid hydroxylase from Aspergillus niger. Appl. Environ. Microbiol. 2021, 87, 1–14. [Google Scholar] [CrossRef]
- Martins, T.M.; Martins, C.; Guedes, P.; Silva Pereira, C. Twists and turns in the salicylate catabolism of Aspergillus terreus, revealing new roles of the 3-hydroxyanthranilate pathway. mSystems 2021, 6. [Google Scholar] [CrossRef]
- Penn, C.D.; Daniel, S.L. Salicylate degradation by the fungal plant pathogen Sclerotinia sclerotiorum. Curr. Microbiol. 2013, 67, 218–225. [Google Scholar] [CrossRef]
- Ambrose, K. V.; Tian, Z.; Wang, Y.; Smith, J.; Zylstra, G.; Huang, B.; Belanger, F.C. Functional characterization of salicylate hydroxylase from the fungal endophyte Epichloë festucae. Sci. Rep. 2015, 5, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Rocheleau, H.; Al-harthi, R.; Ouellet, T. Degradation of salicylic acid by Fusarium graminearum. Fungal Biol. 2019, 123, 77–86. [Google Scholar] [CrossRef]
- Santha, R.; Savithri, H.S.; Rao, N.A.; Vaidyanathan, C.S. 2,3-Dihydroxybenzoic acid decarboxylase from Aspergillus niger: A novel decarboxylase. Eur. J. Biochem. 1995, 230, 104–110. [Google Scholar] [CrossRef]
- Subba Rao, P. V; Moore, K.; Towers, G.H.N. O-Pyrocatechuic acid carboxy-lysase from Aspergillus niger. Arch. Biochem. Biophys. 1967, 466–473. [Google Scholar]
- Choi, S.; Lee, H.N.; Park, E.; Lee, S.J.; Kim, E.S. Recent advances in microbial production of cis,cis-muconic acid. Biomolecules 2020, 10, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Bachman, D.M.; Dragoon, B.; John, S. Reduction of salicylate to saligenin by Neurospora. Arch. Biochem. Biophys. 1960, 91, 326. [Google Scholar] [CrossRef]
- Sgro, M.; Chow, N.; Olyaei, F.; Arentshorst, M.; Geoffrion, N.; Ram, A.F.J.; Powlowski, J.; Tsang, A. Functional analysis of the protocatechuate branch of the β-ketoadipate pathway in Aspergillus niger. J. Biol. Chem. 2023, 299, 105003. [Google Scholar] [CrossRef] [PubMed]
- Vaillancourt, F.; Bolin, J.; Eltis, L. The ins and outs of ring-cleaving dioxygenases. Crit. Rev. Biochem. Mol. Biol. 2006, 41, 241–267. [Google Scholar] [CrossRef]
- Semana, P.; Powlowski, J. Four aromatic intradiol ring cleavage dioxygenases from Aspergillus niger. Appl. Environ. Microbiol. 2019, 85. [Google Scholar] [CrossRef]
- Kowalczyk, J.E.; Benoit, I.; De Vries, R.P. Regulation of plant biomass utilization in Aspergillus; 1st ed.; Elsevier Inc., 2014; Vol. 88; ISBN 9780128002605.
- Seekles, S.J.; van Dam, J.; Arentshorst, M.; Ram, A.F.J. Natural variation and the role of Zn2 Cys6 Transcription factors SdrA, WarA and WarB in sorbic acid resistance of Aspergillus niger. Microorganisms 2022, 10. [Google Scholar] [CrossRef]
- Hynes, M.J.; Murray, S.L.; Duncan, A.; Khew, G.S.; Davis, M.A. Regulatory genes controlling fatty acid catabolism and peroxisomal functions in the filamentous fungus Aspergillus nidulans. Eukaryot. Cell 2006, 5, 794–805. [Google Scholar] [CrossRef]
- Hynes, M.J.; Murray, S.L.; Khew, G.S.; Davis, M.A. Genetic analysis of the role of peroxisomes in the utilization of acetate and fatty acids in Aspergillus nidulans. Genetics 2008, 178, 1355–1369. [Google Scholar] [CrossRef]
- Arentshorst, M.; Reijngoud, J.; van Tol, D.J.C.; Reid, I.D.; Arendsen, Y.; Pel, H.J.; van Peij, N.N.M.E.; Visser, J.; Punt, P.J.; Tsang, A.; et al. Utilization of ferulic acid in Aspergillus niger requires the transcription factor FarA and a newly identified Far-like protein (FarD) that lacks the canonical Zn(II)2Cys6 domain. Front. Fungal Biol. 2022, 3, 1–17. [Google Scholar] [CrossRef]
- Kakkar, S.; Bais, S. A review on protocatechuic acid and its pharmacological potential. ISRN Pharmacol. 2014, 2014, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Kaur, B.; Chakraborty, D. Biotechnological and molecular approaches for vanillin production: A review. Appl. Biochem. Biotechnol. 2013, 169, 1353–1372. [Google Scholar] [CrossRef] [PubMed]
- Kocaçalişkan, I.; Talan, I.; Terzi, I. Antimicrobial activity of catechol and pyrogallol as allelochemicals. Zeitschrift fur Naturforsch. - Sect. C J. Biosci. 2006, 61, 639–642. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Metabolic pathway of the hydroxycinnamic acids, ferulic acid, p-coumaric acid and caffeic acid in A. niger. Pathways observed in A. niger are marked with black arrows. Unidentified pathways in A. niger are marked with dashed arrows. Boxes near an arrow represents identified enzymes catalyzing the reactions. Pathways that were not observed in A. niger but in other Aspergilli are marked with blue (A. flavus), green (A. japonicus) yellow (A. parasiticus), orange (A. fumigatus), cyan (A. luchuensis) and purple (A. brasilliensis) arrows. HMPKP-SCoA, 4-hydroxy-3-methoxyphenyl-β-hydroxypropionyl-CoA; HMPKP-SCoA, 4-hydroxy-3-methoxyphenyl-β-ketopropionic acid-CoA; 4HPKP-SCoA, 4-hydroxyphenyl-β-hydroxypropionyl-CoA; 4HPKP-SCoA, 4-hydroxyphenyl-β-ketopropionic acid-CoA; HHPKP-SCoA, 3,4-dihydroxyphenyl-β-hydroxypropionyl-CoA; HHPKP-SCoA, 3,4-dihydroxyphenyl-β-ketopropionic acid-CoA.
Figure 1.
Metabolic pathway of the hydroxycinnamic acids, ferulic acid, p-coumaric acid and caffeic acid in A. niger. Pathways observed in A. niger are marked with black arrows. Unidentified pathways in A. niger are marked with dashed arrows. Boxes near an arrow represents identified enzymes catalyzing the reactions. Pathways that were not observed in A. niger but in other Aspergilli are marked with blue (A. flavus), green (A. japonicus) yellow (A. parasiticus), orange (A. fumigatus), cyan (A. luchuensis) and purple (A. brasilliensis) arrows. HMPKP-SCoA, 4-hydroxy-3-methoxyphenyl-β-hydroxypropionyl-CoA; HMPKP-SCoA, 4-hydroxy-3-methoxyphenyl-β-ketopropionic acid-CoA; 4HPKP-SCoA, 4-hydroxyphenyl-β-hydroxypropionyl-CoA; 4HPKP-SCoA, 4-hydroxyphenyl-β-ketopropionic acid-CoA; HHPKP-SCoA, 3,4-dihydroxyphenyl-β-hydroxypropionyl-CoA; HHPKP-SCoA, 3,4-dihydroxyphenyl-β-ketopropionic acid-CoA.
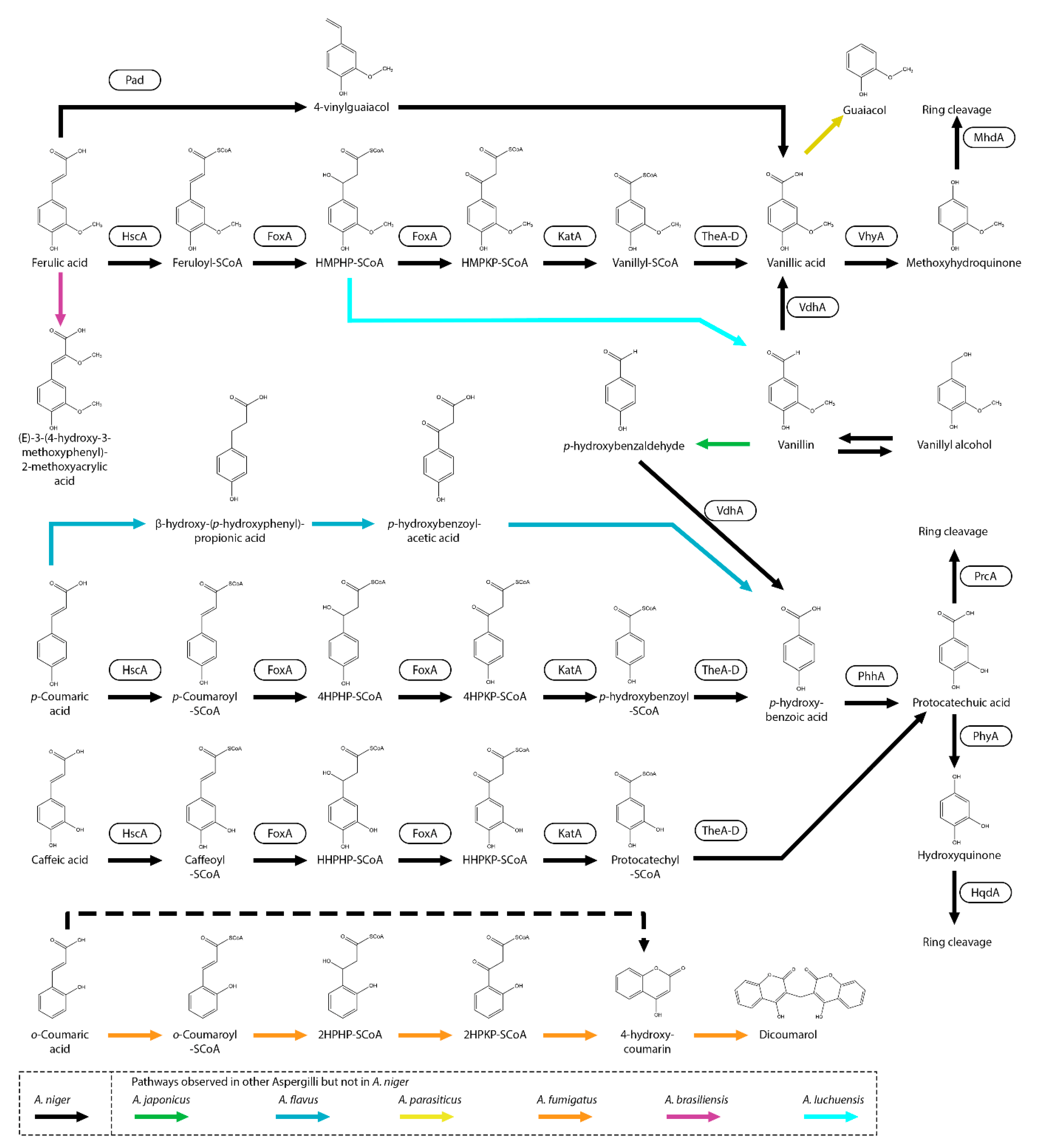
Figure 2.
Cinnamic acid metabolic pathway of A. niger. Boxes near an arrow represents the enzymes catalyzing the reactions. Suggested pathways are marked with dashed arrows. Pathways that were not observed in A. niger but in other Aspergilli are marked with green (A. japonicus) arrows.
Figure 2.
Cinnamic acid metabolic pathway of A. niger. Boxes near an arrow represents the enzymes catalyzing the reactions. Suggested pathways are marked with dashed arrows. Pathways that were not observed in A. niger but in other Aspergilli are marked with green (A. japonicus) arrows.
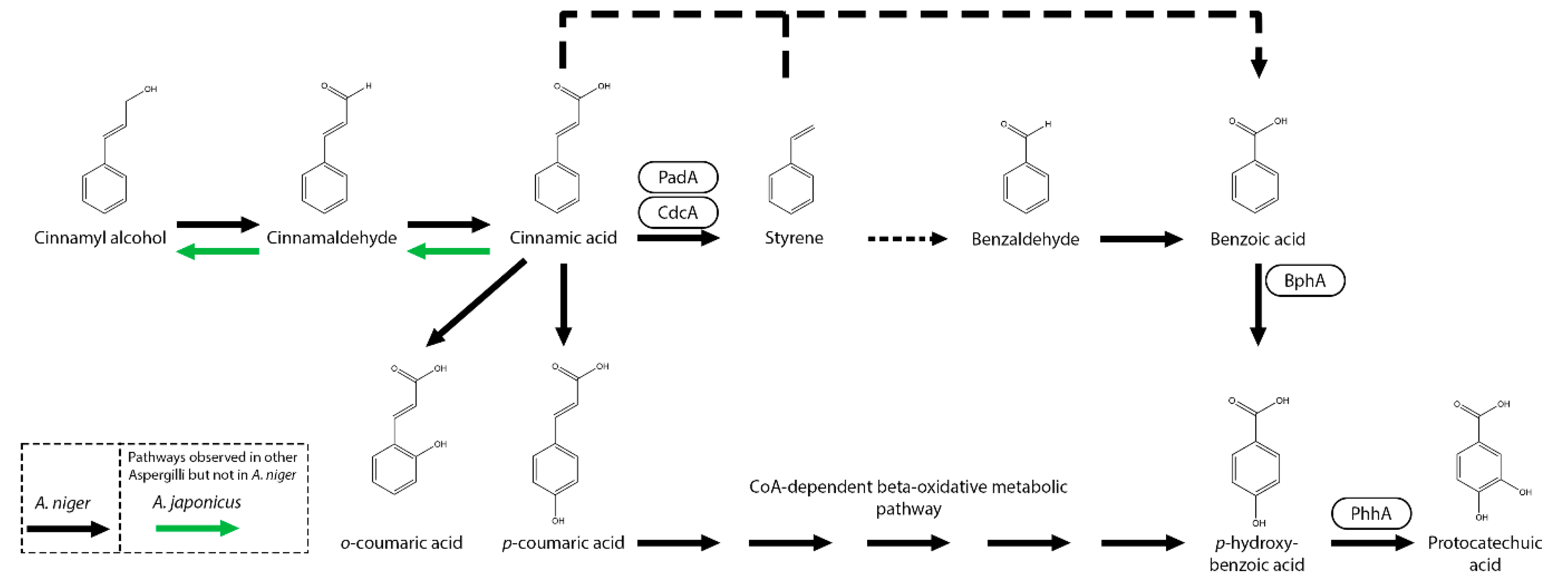
Figure 3.
Benzoic acid and related compounds metabolic pathway of A. niger. Black arrow represent observed A. niger metabolic pathways and boxes near an arrow represents the enzymes catalyzing the reactions. Pathways that were not observed in A. niger but in other Aspergilli are marked with green (A. japonicus), blue (A. flavus), orange (A. fumigatus) or red (A. sojae) arrows.
Figure 3.
Benzoic acid and related compounds metabolic pathway of A. niger. Black arrow represent observed A. niger metabolic pathways and boxes near an arrow represents the enzymes catalyzing the reactions. Pathways that were not observed in A. niger but in other Aspergilli are marked with green (A. japonicus), blue (A. flavus), orange (A. fumigatus) or red (A. sojae) arrows.
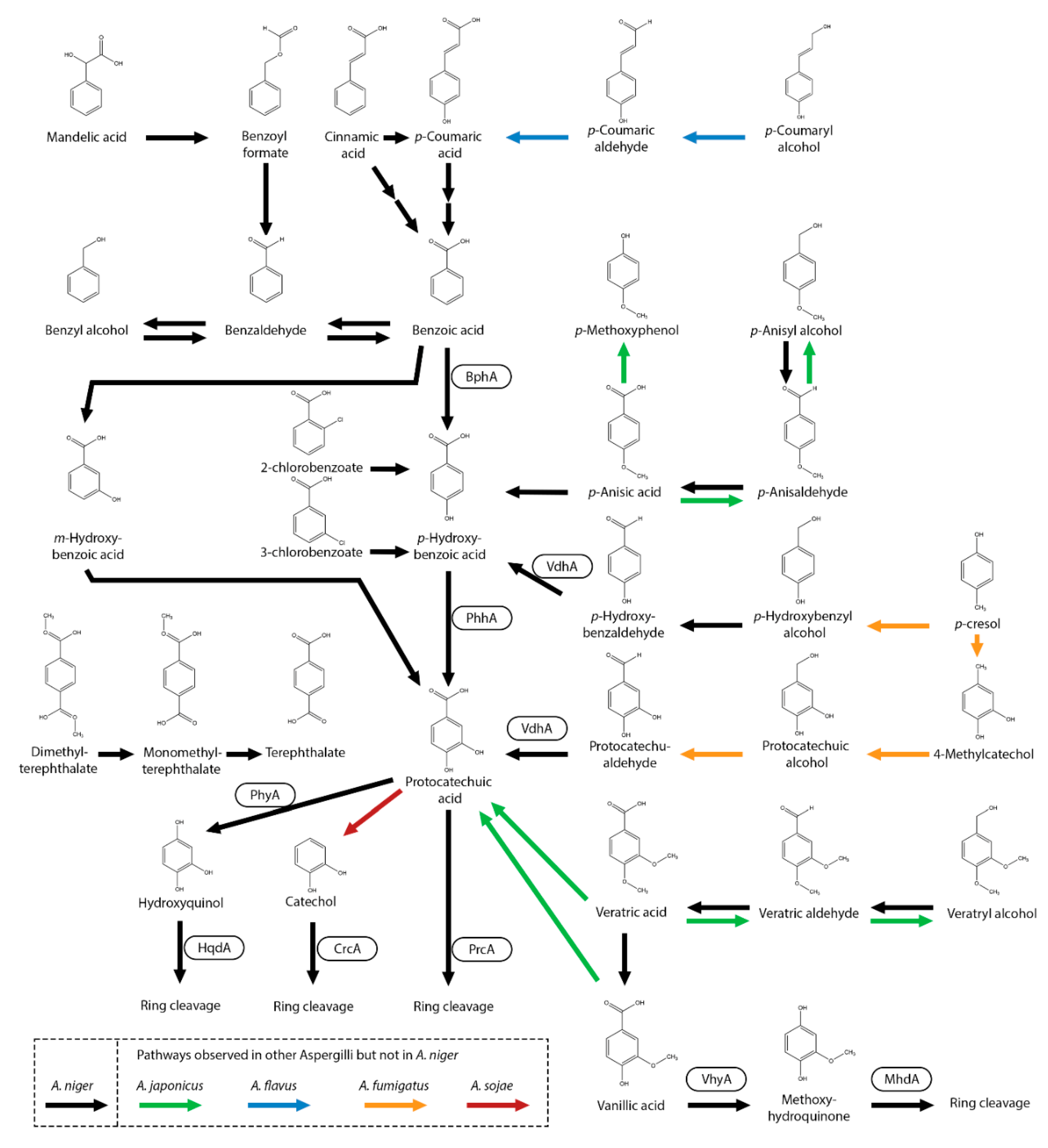
Figure 4.
Vanillic acid and related compounds metabolic pathway of A. niger. Black arrow represents pathways observed in A. niger. Boxes near an arrow represents the enzymes catalyzing the reactions. Pathways that were not observed in A. niger but in other Aspergilli are marked with green (A. japonicus), pink (A. carbonarius), Cyan (A. luchuensis) and yellow (A. parasiticus) arrows.
Figure 4.
Vanillic acid and related compounds metabolic pathway of A. niger. Black arrow represents pathways observed in A. niger. Boxes near an arrow represents the enzymes catalyzing the reactions. Pathways that were not observed in A. niger but in other Aspergilli are marked with green (A. japonicus), pink (A. carbonarius), Cyan (A. luchuensis) and yellow (A. parasiticus) arrows.
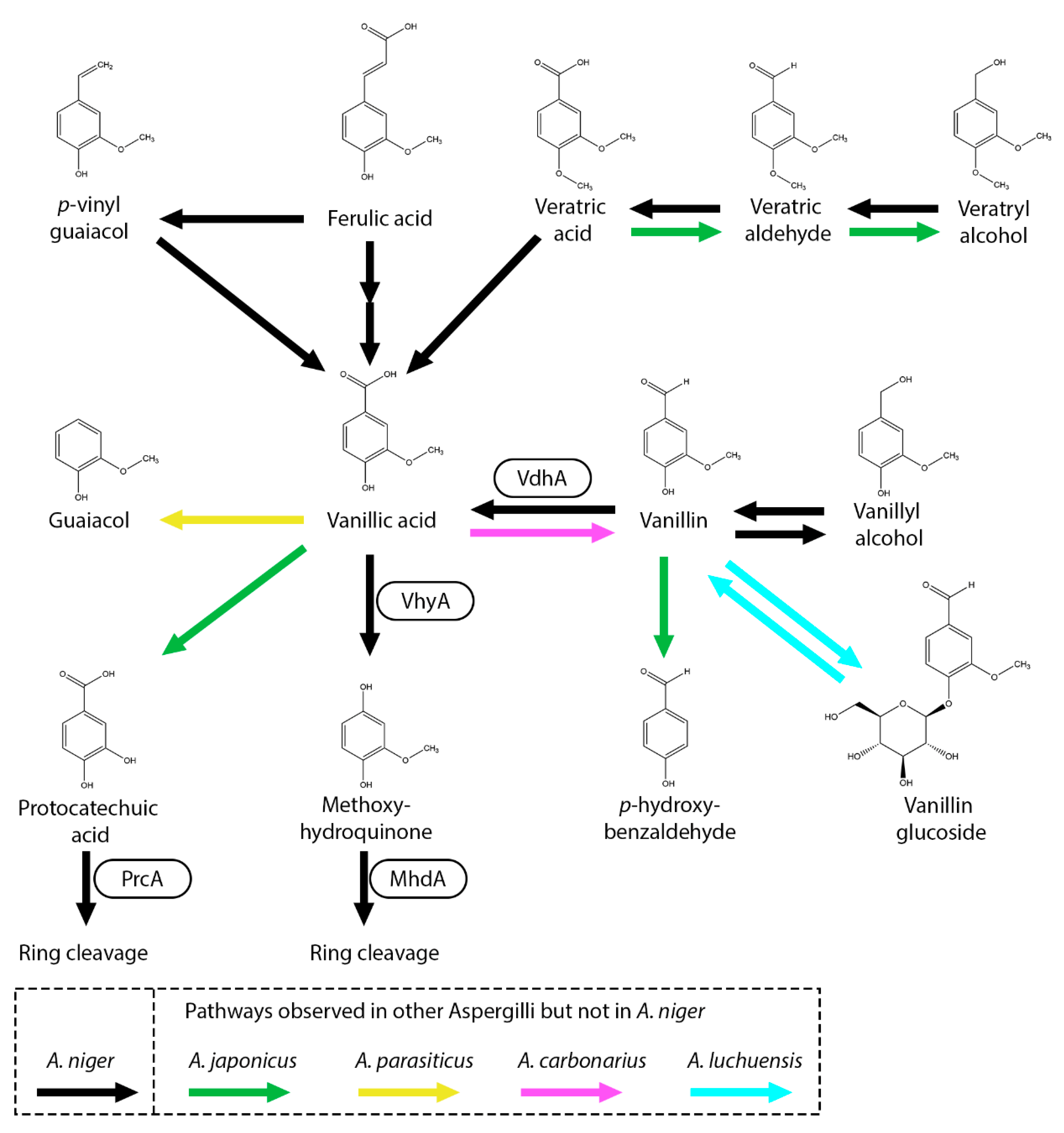
Figure 5.
Gallic acid and related compounds metabolic pathway of A. niger. Black arrow represents pathways observed in A. niger. Boxes near an arrow represents the enzymes catalyzing the reactions. Pathways that were not observed in A. niger but in other Aspergilli are marked with gray (A. oryzae) and light blue (A. nidulans) arrows.
Figure 5.
Gallic acid and related compounds metabolic pathway of A. niger. Black arrow represents pathways observed in A. niger. Boxes near an arrow represents the enzymes catalyzing the reactions. Pathways that were not observed in A. niger but in other Aspergilli are marked with gray (A. oryzae) and light blue (A. nidulans) arrows.
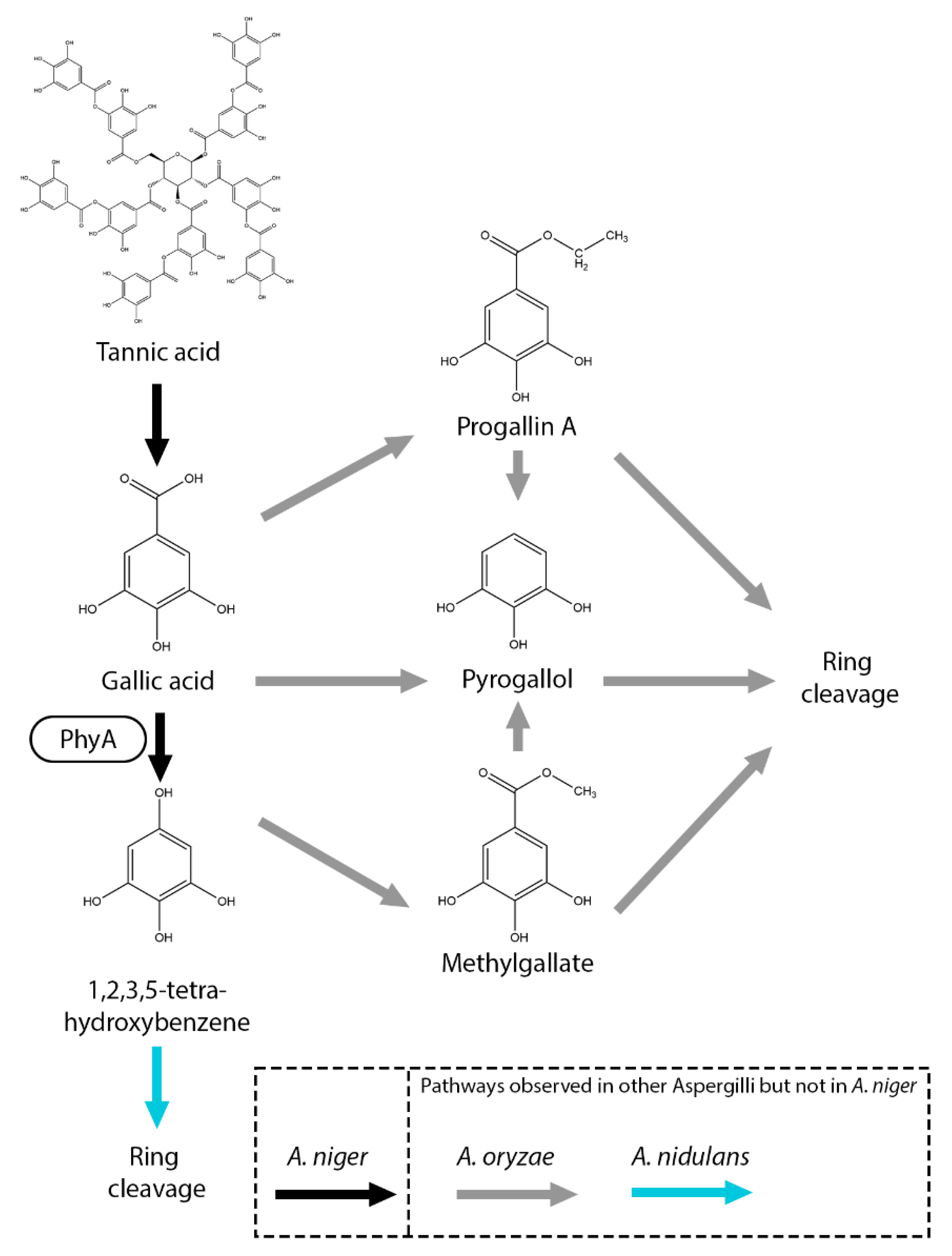
Figure 6.
Salicylic acid metabolic pathway of A. niger. Boxes near an arrow represents the enzymes catalyzing the reactions. Suggested pathways are marked with dashed arrows. Pathways that were not observed in A. niger but in other Aspergilli are marked with brown (A. terreus), orange (A. fumigatus) or light blue (A. nidulans) arrows.
Figure 6.
Salicylic acid metabolic pathway of A. niger. Boxes near an arrow represents the enzymes catalyzing the reactions. Suggested pathways are marked with dashed arrows. Pathways that were not observed in A. niger but in other Aspergilli are marked with brown (A. terreus), orange (A. fumigatus) or light blue (A. nidulans) arrows.
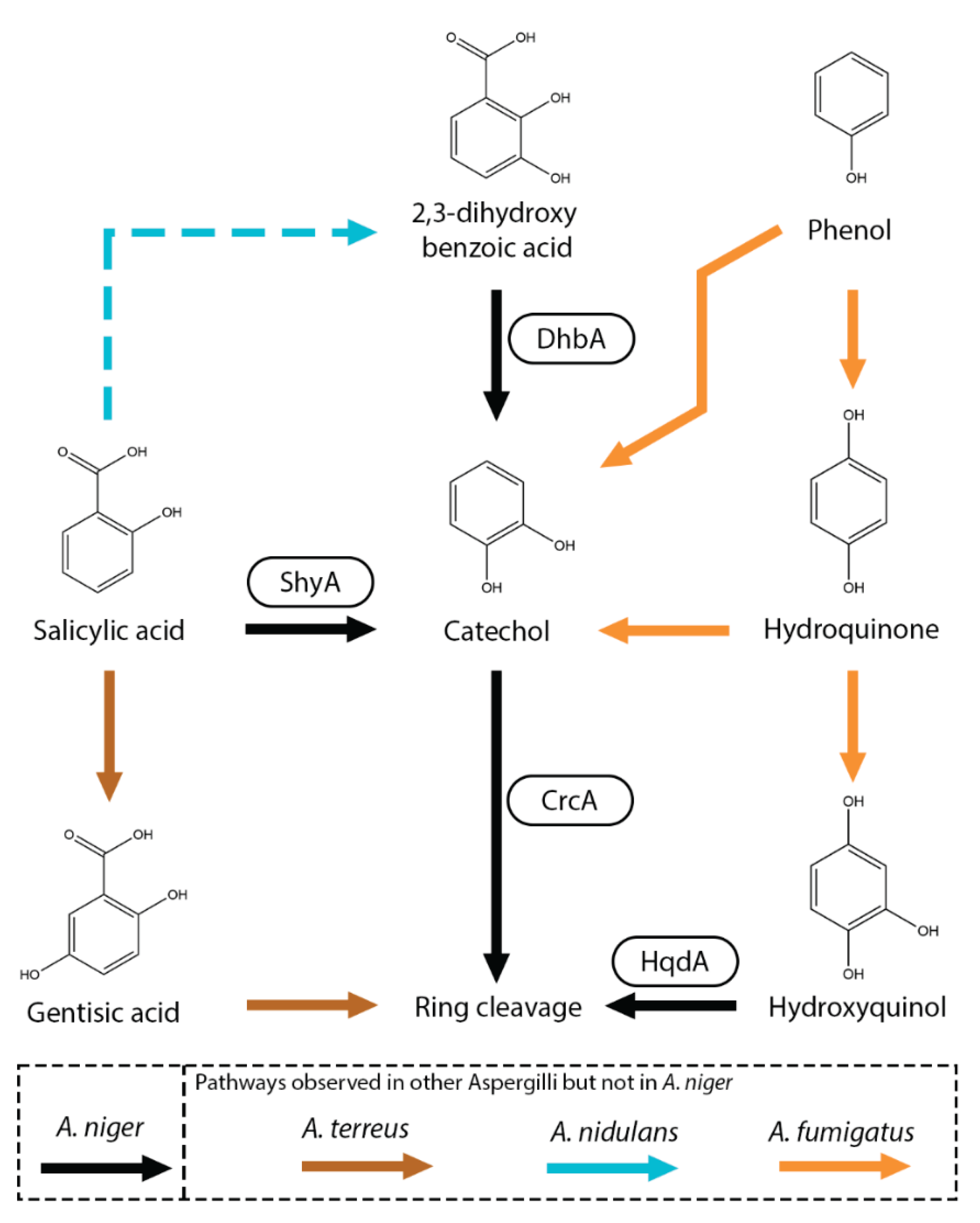

Table 1.
Phenotypes observed of A. niger dioxygenase deletion transformants on aromatic compounds.
 |
|||||||||||||
| Aromatic compound | 1 | 2 | 3 | 4 | 5 | 6 | ΔprcA [46,47,78] | ΔhqdA[46,47,78] | ΔprcA/hqdA[46,47] | ΔcrcA[78] | ΔmhdA[71] | Δ5330[78] | Δ17[83] |
| H-unit related compounds | |||||||||||||
| Cinnamic acid | CH=CHC=OOH | H | H | H | H | H | ++ | - | ++ | - | - | - | n.d. |
| p-coumaric acid | CH=CHC=OOH | H | H | OH | H | H | + | - | ++ | - | - | - | ++ |
| Caffeic acid | CH=CHC=OOH | H | OH | OH | H | H | + | - | ++ | - | - | - | ++ |
| Benzaldehyde | CH=O | H | H | H | H | H | ++ | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| Benzoic acid | C=OOH | H | H | H | H | H | ++ | - | ++ | n.d. | - | n.d. | n.d. |
| Benzyl alcohol | CH2OH | H | H | H | H | H | ++ | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| p-hydroxybenzoic acid | C=OOH | H | H | OH | H | H | ++ | - | ++ | n.d. | - | n.d. | ++ |
| p-hydroxybenzaldehyde | CH=O | H | H | OH | H | H | ++ | - | ++ | n.d. | n.d. | n.d. | n.d. |
| m-hydroxybenzoic acid | C=OOH | H | OH | H | H | H | ++ | n.d. | n.d. | n.d. | - | n.d. | n.d. |
| Protocatechuic acid | C=OOH | H | OH | OH | H | H | + | - | ++ | - | - | - | ++ |
| Protocatechuic aldehyde | CH=O | H | OH | OH | H | H | + | - | ++ | n.d. | - | n.d. | n.d. |
| p-Anisic acid | C=OOH | H | H | OCH3 | H | H | ++ | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| p-Anisyl alcohol | CH2OH | H | H | OCH3 | H | H | ++ | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| G-unit related compounds | |||||||||||||
| Coniferyl alcohol | CH=CHCH2=OH | H | OCH3 | OH | H | H | n.d. | n.d. | n.d. | n.d. | ++ | n.d. | n.d. |
| Ferulic acid | CH=CHC=OOH | H | OCH3 | OH | H | H | - | - | - | - | ++ | - | - |
| Vanillic acid | C=OOH | H | OCH3 | OH | H | H | - | - | - | n.d. | ++ | n.d. | - |
| Vanillin | CH=O | H | OCH3 | OH | H | H | n.d. | n.d. | n.d. | n.d. | ++ | n.d. | n.d. |
| Vanillyl alcohol | CH2OH | H | OCH3 | OH | H | H | n.d. | n.d. | n.d. | n.d. | ++ | n.d. | n.d. |
| Veratric acid | C=OOH | H | OCH3 | OCH3 | H | H | - | n.d. | n.d. | n.d. | + | n.d. | n.d. |
| S-unit related compounds | |||||||||||||
| Gallic acid | C=OOH | H | OH | OH | OH | H | - | - | - | n.d. | n.d. | n.d. | - |
| Others | |||||||||||||
| Salicylic acid | C=OOH | OH | H | H | H | H | - | - | n.d. | ++ | n.d. | - | ++ |
| 2,3-dihydroxybenzoic acid | C=OOH | OH | OH | H | H | H | - | - | n.d. | ++ | n.d. | - | n.d. |
| Catechol | OH | OH | H | H | H | H | - | - | - | ++ | n.d. | - | ++ |
| Gentisic acid | C=OOH | OH | H | H | OH | H | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | - |
- no phenotype; + phenotype; ++ strong phenotype; n.d. not determined.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



