
Submitted:
11 June 2025
Posted:
16 June 2025
You are already at the latest version
Abstract
Scrub typhus, caused by Orientia tsutsugamushi (Ot), is a re‐emerging public health concern across Southeast Asia. Although multiple Ot strains have been identified in endemic regions, its genetic diversity in India remains limited. We analysed Ot strains from humans by targeting the conserved GroEL and variable 56‐kDa genes. A total of 105 serum samples were subjected to PCR amplification and phylogenetic analysis for the GroEL gene, of which 33 (31.4%) were positive. Phylogenetic reconstruction revealed four major clades: Karp, Kato, Ot‐TJTN, and Gilliam‐related. Among the 33‐PCR positive samples, 11 sequences formed a distinct monophyletic clade within the Gilliam‐related group, diverging significantly from classical Gilliam strains. The overall mean nucleotide diversity (π) was 0.02 (2%), while the divergence between these 11 sequences and Gilliam‐ related strains was 0.039 (3.9%). This level of divergence supports the identification of a novel Gilliam‐descendant lineage, designated INDv‐Gilliam. Further analysis of the 56‐kDa gene from the 11 INDv‐Gilliam samples revealed genotypic incongruence, indicating antigenic reassortment involving three clades: Karp‐like (n = 7), Ot‐TJTN‐like (n = 3), and Gilliam (n = 1). Similarity plot and recombination analyses, using 56‐kDa Ot‐TJTN and Karp‐like clades as queries against Gilliam‐type references, provided evidence of recombination involving TA716‐Kato and Ikeda‐Karp strains. These findings highlight the role of recombination and antigenic shift in driving the evolutionary dynamics and genetic diversity of Ot in the region. Accordingly, the isolates (11 nos.) were classified as mixed recombinant genotypes: Indian variant Gilliam (INDv‐Gilliam), INDv‐Gilliam/Karp‐like, INDv‐ Gilliam/Ot‐TJTN, and INDv‐Gilliam/Gilliam‐type.
Keywords:
scrub typhus
; O. tsutsugamushi
; genetic diversity
; Gilliam
; genotype
; Karp
; Kato
; Ot‐TJTN
1. Introduction
Scrub typhus, once a neglected febrile illness, has re-emerged as a significant public health threat across Asia and beyond [1]. It is caused by an obligate intracellular bacterium, O. tsutsugamushi and is transmitted by the larval stage (chiggers) of trombiculid mites, especially Leptotrombidium deliense, which acts as both vector and reservoir [2]. Rodents act as natural hosts, while humans are dead-end hosts [3]. Initially, O. tsutsugamushi was considered geographically restricted to the "Tsutsugamushi Triangle"; subsequently, studies have given a broader global distribution of scrub typhus, along with the discovery of new Orientia species Candidatus. O. chuto sp. nov. and Candidatus. O. chiloensis. [3,4]. Nearly 2 billion people are at risk, and an estimated 1 million cases occur annually within this region [1]. Scrub typhus can have a mortality rate ranging from 6% to 30% if left untreated and can significantly drop to 1.4% with prompt antibiotic treatment [5]. Underreporting of scrub typhus is common, as the characteristic clinical feature, an eschar at the bite site, is frequently absent or unnoticed. The absence of this feature and fever symptoms that overlap with other febrile illnesses make accurate diagnosis of scrub typhus difficult. Moreover, insensitive diagnostic methods often delay identification [6].
Mite vectors adapt to local environmental conditions, which plays a key role in the regional distribution of O. tsutsugamushi. Within these mite populations, genetic recombination can produce new strains, increasing the pathogen’s antigenic variability [3,7]. This is further enhanced by the bacterium’s genome, which contains many repetitive sequences, including multiple copies of genes related to the type IV secretion system (T4SS) and numerous transposase genes. These features promote intragenomic rearrangements and lateral gene transfer [7] and result in the emergence of many unique, region-specific genotypes, thus complicating efforts for disease surveillance and control [8]. Generally, the strain classification was based on prototype strains like Karp, Gilliam, and Kato. However, more recent studies using multilocus sequence typing and whole-genome analysis have led to a much broader diversity, and new genotypes such as Kawasaki, Boryong, TA763, and Kuroki have been identified [9,10]. This extensive diversity poses significant challenges to serodiagnosis and vaccine development, as immunity is often strain-specific and reinfections with different strains are common [11].
In India, scrub typhus is endemic, with a high incidence in the rural and semi-urban regions. However, molecular data on circulating O. tsutsugamushi genotypes remain regionally limited [6]. While over 40 distinct genotypes have been reported globally [10], genotypic information from India is largely restricted to certain northern, northeastern, and southern regions [12]. A cohort study in south India reported that approximately one in 20 participants were affected by O. tsutsugamushi infection [13]. Recent clinical cases have identified O. tsutsugamushi as an emerging cause of acute encephalitis syndrome (AES), previously attributed primarily to viral infections such as Japanese encephalitis [14]. Further investigations have revealed a wide range of circulating genotypes with varying pathogenicity, strain-dependent mortality rates of up to 50%, and antibiotic susceptibilities, causing significant diagnostic challenges [15]. A recent molecular study based on clinical samples from Thanjavur Medical College Hospital in Tamil Nadu identified a novel genotype designated as Ot-TJTN along with Karp-like and Gilliam-related strains through sequence analysis of the 56-kDa type-specific antigen (TSA) and GroEL genes [16]. This newly detected variant signifies the unrecognized genetic diversity of O. tsutsugamushi in southern India and emphasizes the need for continued molecular surveillance to monitor emerging genotypes and regional strain evolution.
Nevertheless, while type-specific antigen genes such as TSA 47-kDa, SCA, and 22-kDa are used for genotyping O. tsutsugamushi, the 56-kDa gene remains the most commonly used marker due to its antigenic variability. This immunodominant nature makes it susceptible to host-driven selective pressures, which can obscure true evolutionary relationships [17]. Hence, its use as a phylogenetic marker has limitations [18]. The TSA 56-kDa variable domain I-III (VDI-III) represents higher hypervariability levels than the entire gene [19]. In addition, numerous mobile genetic elements in the O. tsutsugamushi genome facilitate lateral gene transfer, contributing to genetic diversity. In contrast, the GroEL gene, which encodes the 60-kDa heat shock protein (HSP), shows conserved but informative polymorphisms, making it a more reliable marker for phylogenetic analysis and strain differentiation [18]. In this study, to gain a more comprehensive understanding of the genetic diversity of O. tsutsugamushi, both markers were analysed in clinical samples collected from human cases in the Theni and Dindigul districts of Tamil Nadu. These regions, located at the foothills of the Western Ghats, are ecologically diverse and have reported cases of scrub typhus [20,21], making them important areas for studying the molecular characteristics of circulating O. tsutsugamushi strains.
2. Materials and Methods
2.1. Study Area
Theni and Dindigul districts are situated along the foothills of the Western Ghats in southwestern Tamil Nadu (9°45′–10°30′ N, 77°15′–78°00′ E) and are known endemic zones for scrub typhus [21]. These regions are characterized by a tropical climate with seasonal monsoons, moderate to high humidity, and a diverse landscape with forest fringes, agricultural fields, and both rural and urban communities, conditions favourable to support infestation in both chigger mite vectors and their small mammal hosts.
2.2. Sample Collection
As part of an active surveillance study on scrub typhus, blood samples were collected from patients with acute febrile illness attending government healthcare facilities in Theni and Dindigul districts, Tamil Nadu. Patients suspected of having scrub typhus were screened using a commercial IgM ELISA kit (Scrub Typhus Detect™ IgM ELISA, InBios International, Inc., Seattle, WA, USA). Only those who tested positive for scrub typhus in the ELISA were selected for molecular analysis, resulting in 105 ELISA-positive samples. Blood samples were collected and transported under cold chain conditions to the ICMR-Vector Control Research Centre (VCRC), Field Station, Madurai. Samples were stored at −20°C until processing. Genomic DNA was extracted and analysed using nested PCR assays targeting the GroEL, 56 kDa type-specific antigen genes of O. tsutsugamushi for genotypic characterization. Ethical approval for the study was obtained from the Institutional Human Ethics Committee of ICMR-VCRC (Approval No: TMC/IHEC: 1118/2023; 27 July 2023).
2.3. Molecular Detection and Genotyping of O. tsutsugamushi
2.3.1. Genomic DNA Extraction and GroEL Gene Amplification
Genomic DNA was extracted from ELISA-positive human serum samples using the QIAamp DNA Blood Mini Kit (Qiagen, Hilden, Germany; Cat. No. 51104), following the manufacturer’s instructions. The DNA concentration and purity were assessed, and aliquots were stored at −20 °C until further analysis. Nested PCR targeting the GroEL gene was performed based on the protocol described by Li et al. [22]. The first round utilized the outer primers Gro-1 (5′-AAGAAGGACGTGATAAC-3′) and Gro-2 (5′-ACTTCACGTAGCACC-3′), and the second round employed the inner primers TF1 (5′-ATATATCACAGTACTTTGCAAC-3′) and TR2 (5′-GTTCCTAACTTAGATGTATCAT-3′) for amplification of a 365 bp product. PCR was carried out in a 25 µL reaction volume, with 2 µL of template DNA in the first round and 1 µL of PCR product in the nested round. PCR amplicons were resolved on a 2% agarose gel alongside a 100 bp molecular ladder. Negative (no-template DNA) controls were included in each run. The PCR products of the expected size were subsequently purified and subjected to Sanger sequencing in both forward and reverse directions.
2.3.2. Amplification of the 56 kDa Type-Specific Antigen Gene
All the samples positive for the GroEL gene were further analysed by nested PCR targeting the variable domains I-III (VDI–III) of the 56-kDa type-specific antigen gene. The outer primers used were JG-OtF584 (5′-CAA TGT CTG CGT TGT CGT TGC-3′) and RTS9 (5′-ACAGAT GCA CTA TTA GGC AA-3′), followed by the nested primers F (5′-AGC GCTAGG TTT ATT AGC AT-3′) and RTS8 (5′-AGG ATT AGA GTG TGG TCCTT-3′), as per the method of Ruang-areerate et al [17]. Amplification was confirmed by electrophoresis on 1% agarose gel. Positive PCR products were purified and subjected to Sanger sequencing.
2.3.3. Sequence Editing, BLAST Analysis, and GenBank Submission
Forward and reverse sequence reads were manually reviewed and assembled into consensus sequences. The resulting nucleotide sequences were compared against reference sequences in the NCBI GenBank database using the BLASTn tool to determine genetic similarity and strain identity. A total of 33 GroEL and eleven 56-kDa sequences corresponding to the Gilliam clade were generated and submitted to GenBank (Accession Nos for GroEL and 56-kDa are given in Table 1&3).
2.3.4. Phylogenetic Analysis
Phylogenetic trees were constructed using the neighbour-joining method in MEGA version 11 [23], applying the Kimura 2-parameter substitution model. Bootstrap analysis with 1000 replicates was performed to assess the robustness of the tree topology. For a comparative study, reference sequences representing prototype O. tsutsugamushi strains (Kato, Kuroki, Kawasaki, Ikeda, Hualien-1, Gilliam, Boryong, Hwasung, Karp, and TA716) were included. The overall nucleotide diversity (d) and mean distance between the genotypes and the INDv-Gilliam (11 nos.) vs Gilliam-related genotypes were calculated.
2.3.5. Similarity and Bootscan Analysis of 56 kDa Sequences
To examine sequence similarity and detect possible recombination events, SimPlot++ version 3.5.1 was used [24]. Among the three clades within 11 sequences of 56-kDa, the Karp-like and Ot-TJTN clades were independently analysed for similarity and bootscan analysis. One study sequence (PV233813) aligned closely with the Gillam prototype was excluded from the Simplot++ analysis. Bootscan was performed using the Kimura-2-parameter model, and similarity was assessed using Hamming distance with a window size of 200 bp and a step size of 20 bp. Similarly, recombinant distance plots were generated using the 11 INDv-Gilliam study sequences as a query to detect potential recombination breakpoints.
3. Results
3.1. Molecular Detection and GroEL Gene Sequence Analysis
A total of 105 clinical samples collected from Theni (n=51) and Dindigul (n=54) districts were screened for O. tsutsugamushi using nested PCR targeting the conserved GroEL gene. Among these, 33 samples (31.4%) yielded high-quality sequences that were subjected to BLASTn analysis, revealing ≥96% nucleotide identity with previously reported O. tsutsugamushi strains in the NCBI GenBank database (Table 1). Phylogenetic analysis of the GroEL 365 bp fragment, using the neighbour-joining method, resolved four major genotypic clusters: a novel Gilliam-related lineage, Ot-TJTN-like clade, Kato-like clade and Karp-like group (Figure 1). (GroEL-based Ot-TJTN, Kato-like and Karp-related data unpublished). Eleven of the 33 sequences (33.3%) formed a distinct monophyletic clade deviating from classical Gilliam-type strains (Figure 1 and Figure 2).
The estimation of overall mean nucleotide diversity (π) was 0.02 (2%) (SE:0.00), and between the eleven INDv-Gilliam/Gilliam was 0.039 (3.9%) (SE: 0.009). The nucleotide divergence between the INDv-Gilliam and five Gilliam-related reference genotypes showed a distinct divergence of >3%, with a new variant detected in this study (Table 2).
3.2. Sequence Analysis of the TSA 56 kDa Antigen Gene
Eleven DNA samples, previously clustered within a novel Gilliam-related clade based on the GroEL gene, were further analysed through nested PCR amplification targeting variable domains I–III (VDI-III) of the 56-kDa gene (Table 3). High-quality sequences were obtained from all eleven samples without background noise, enabling robust comparative phylogenetic analysis. Notably, 56-kDA sequence analysis with the prototype reference sequences revealed a marked divergence in lineage classification compared with conserved GroEL gene analysis.
Seven of the eleven identified as INDv-Gilliam in the GroEL gene were clustered to the Karp-like lineage in the 56 kDa phylogenetic tree. These sequences exhibited 100% nucleotide identity with strain OT/India/0809aTw/2008 (MW495817) and 98.8% similarity with isolates OT/AIIMS/4012/2021 (ON087065) from India and UT395 from Thailand (EF213094). This reclassification indicates that these samples, although closely related to Gilliam-like strains based on GroEL, share antigenic properties consistent with Karp-related genotypes in the more variable TSA 56 kDa region. Three of the remaining samples formed a well-supported cluster within the recently reported Ot-TJTN-like genotype, displaying 100% identity with TJ49 (PQ381701) and strain JJOtsu2C (PQ059255) from South India. Only one sample retained Gilliam-like identity, aligning most closely with strain TT0711a (GQ332755) from Taiwan (95.1% identity), strain Hualien-1 (AY243357) from Taiwan (93.93%), and isolate CH01117 from China (MT258819) (Figure 3). Based on the result of 56 kDa, the 11 Gilliam variants were classified as mixed recombinant genotypes: INDv-Gilliam/Karp-like, INDv-Gilliam/Ot-TJTN, and INDv-Gilliam/Gilliam-type. (Table 4).
3.3. Similarity Plot and Recombination Analysis
SimPlot analysis of the Ot-TJTN isolate revealed maximum sequence similarity to the TA716 strain, followed by the Kato strain. Moderate similarity was observed with Ikeda and Hualien-1 strains, whereas the other references displayed lower similarity profiles, supporting the classification of Ot-TJTN within the TA716-related group. On the other hand, analysis of the Karp clade isolates showed the highest sequence similarity to the Karp, Kuroki and Boryong reference strain across the 56-kDa gene. All other strains exhibited significantly lower similarity, confirming the association of these isolates with the Karp lineage (Figure 4). The two clades in 56-kDa were further investigated for potential recombination breakpoints by using the BootScan analysis. The Ot-TJTN isolate displayed an event of recombination with the TA716 and Kato strains. On the other hand, the bootscan analysis of the 56 kDa Karp clade revealed the occurrence of recombination between the Ikeda and Karp genotypes (Figure 5).
4. Discussion
Land use and movement of host and vector increase the zoonotic epidemiology of scrub typhus [25]. Based on the diversity in the DNA sequences, different genotypes have been reported in O. tsutsugamushi [26]. In general, due to the vertical transmission mode of Ot, the event of genetic recombination within the same and between the strains in the chigger mite result in the formation of a new small-size population with substantial genetic diversity [27]. The emergence of a new variant carries several epidemiological implications, leading to increased virulence, underreporting or misdiagnosing and complicating the vaccine development [28].
Indeed, the functional need of a gene determines the occurrence of mutation in the particular gene [29]. The GroEL gene code for the heat shock protein (HSP) is essential for the survival of the bacteria. Mutation in the HSP will destroy its function and eventually lead to cell death. Hence, this gene is relatively conserved across different strain species. This conserved nature helps identify different strains with 99% accuracy [30]. In contrast, the 56 kDa type-specific antigen (TSA) gene is required to attach and invade the host cell. Since this gene is exposed to evade the host immune system, which undergoes mutation over time, resulting in the presence of many different antigenic types in Ot, which helps to differentiate and monitor the emerging multiple genotypes in Ot [29,31]. Since the 56 kDa immunodominant gene exhibits different genotype patterns, understanding its genetic nature is essential for developing a region-specific vaccine.
Based on this genetic foundation, our study focuses on the molecular epidemiology of O. tsutsugamushi strains circulating among human scrub typhus cases in the Theni and Dindigul districts of Tamil Nadu, India. We targeted two key genetic markers, the relatively conserved groEL gene and the highly variable 56-kDa type-specific antigen (TSA) gene. We identified a distinct Gilliam-related clade, INDv-Gilliam, and detected evidence of antigenic reassortment among circulating strains. These findings broaden the known genotypic diversity of O. tsutsugamushi in India and align with emerging reports of novel variants from other endemic regions in Asia [16]. Gilliam is one of the classical prototype strains, and it is known for its genetic variability across Southeast Asia. For instance, studies conducted in Thailand from 2004 to 2007 based on the 56-kDa gene revealed several unique Gilliam-related variants circulating in the country [17]. Similarly, a mosaic strain, TGv, has been reported in Taiwan, derived from Gilliam, Ikeda, and Kato lineages [32]. Also, in India, a recent study from Karnataka documented the circulation of Japanese Gilliam (JG) and Japanese Gilliam-variant (JG-v) strains among scrub typhus cases, highlighting the ongoing diversification and geographic spread of Gilliam-related genotypes within the country [33].
Phylogenetic analysis based on the GroEL gene revealed four principal clusters: a novel Gilliam-related group, an Ot-TJTN-like clade, a Kato-like clade and a Karp-like lineage. Among the four major clades, one monophyletic clade was distinct from classical Gilliam strains, which showed 100% identity with human clinical isolates from Theni and Thanjavur (GenBank: ON156004, OR887445) and 99.12% similarity with a rodent-derived isolate from Uttar Pradesh (PP355737), suggesting active zoonotic circulation with epidemiological significance. The mean nucleotide diversity of 3.9% between INDv-Gilliam and classical Gilliam strain supports the classification of INDv-Gilliam as a new and genetically distinct variant [34]. Furthermore, the observed nucleotide divergence of over >3% between INDv-Gilliam and other Gilliam-type strains suggests that INDv-Gilliam represents a novel molecular type circulating in the human and rodent populations. [3,35]
In addition to identifying a genetically distinct genotype within the Gilliam group, another core observation of this study was the discordant phylogenetic arrangements of isolates based on GroEL and TSA 56-kDa gene sequences. While GroEL-based analysis grouped the isolates as Gilliam-related, TSA 56-kDa analysis reassigned seven isolates to a Karp-like group, three to an Ot-TJTN-like lineage, and one to the Gilliam group. This discovery is consistent with the recognized function of the TSA 56-kDa gene in driving antigenic variation influenced by immune selection. In contrast, the GroEL housekeeping gene remains relatively conserved. Such antigenic shifts complicate lineage classification when relying only on immunodominant markers, stressing the need for multilocus typing in O. tsutsugamushi epidemiology [18].
SimPlot and BootScan analyses of the 56-kDa gene sequences provided further evidence of close genetic relationships and potential recombination events among circulating strains. The results of neighbour-joining phylogenetic analysis and SimPlot were concordant for the 56-kDa Ot-TJTN clade, indicating a close genetic relationship, as shown by a flat curve overlap pattern and a recombination breakpoint involving TA716 and Kato genotypes. Similarly, analysis of the Karp-like genotype revealed a close genetic relationship, with the highest similarity to the Karp, Ikeda and Kuroki reference strains and evidence of recombination involving Karp and Ikeda genotypes. These findings are consistent with previous reports suggesting that recombination, likely facilitated by co-infections within chigger mite populations, contributes significantly to genetic heterogeneity in O tsutsugamushi. Notably, studies have demonstrated high rates of homologous recombination in O. tsutsugamushi, with evidence indicating that multiple strains can co-infect individual mites, providing possibilities for genetic exchange [7]. Additionally, research has highlighted the potential for horizontal transmission of O. tsutsugamushi among co-feeding mites, further supporting the role of mite co-infections in facilitating genetic recombination [36]. The ecological diversity of the Theni and Dindigul districts, including abundant populations of L. deliense and other trombiculid mite species (unpublished), likely promotes such genetic exchanges. Surveys conducted in southern Tamil Nadu have documented the presence of various chigger mite species, with L. deliense identified as a predominant vector in these regions [37]. The coexistence of multiple mite species in these ecologically diverse areas may enhance the likelihood of co-infections and subsequent genetic recombination events in O. tsutsugamushi.
Identification of the INDv_Gilliam variant, with a genetic divergence of 3.9% from classical Gilliam strains, reinforces its classification as a novel genotype of O. tsutsugamushi. Similar comparable levels of divergence have been associated previously with the emergence of the Kawasaki, Kuroki, and Boryong strains, indicating ongoing regional diversification [34,38]. Current serological assays, which rely primarily on classical antigens (Karp, Kato, Gilliam), may fail to detect such divergent variants, leading to underdiagnosis in endemic regions. The strain-specific nature of immunity further complicates disease control, as previous infection does not ensure protection against emerging genotypes [39]. Genotyping based solely on the groEL gene may mislead phylogenetic relationships due to its conserved nature; therefore, incorporating the more variable 56-kDa TSA gene improves accuracy in strain classification and epidemiological tracking [34]. The emergence of regional variants like INDv_Gilliam reflects localized microevolution within the Gilliam lineage. These findings emphasize the need for updated diagnostic tools and molecular surveillance strategies for evolving genotypic diversity.
5. Conclusions
The high incidence of scrub typhus across Indian states urges the need for increased awareness of the pathogen diversity and the emergence of new strains for improved diagnosis and surveillance. This study highlights previously unrecognized dimensions of O. tsutsugamushi diversity in southern India, notably the emergence of a novel Gilliam-variant (INDv-Gilliam) and evidence of ongoing antigenic reassortment. The discordance between phylogenetic signals from conserved and variable genetic markers underscores the complex evolutionary mechanisms shaping strain diversity in endemic regions. The emergence of such variants carries significant epidemiological implications, including genetic complexity, potential for drug resistance, challenges in accurate diagnosis, and barriers to developing protective vaccines.
Author Contributions
Conceptualization, S.V.J., K.N., P.R., G. R and A.M; methodology, K.N., S.V.J., R.R., R. K. M and R.G.; validation and analysis, K.N., S.V.J., P. R., R.S and S.K.; writing original draft preparation, S.V.J., K.N. and P.R.; writing review and editing, S.V.J., K.N., S.T.G., P.R and M.R.; supervision, P.R., G.R and M.R.; project administration, P.R. and M.R. All authors have read and agreed to the published version of the manuscript.
Funding
This research was carried out with limited intramural funding.
Institutional Review Board Statement
This study has been approved by the institutional Human Ethical Committee, No. TMC/IHEC:1118/2023, 27 July 2023.
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The original contributions presented in the study are included in the article; further inquiries can be directed to the corresponding authors.
Acknowledgments
The authors sincerely acknowledge the laboratory staff of the Department of Microbiology, Government Medical College and Hospital, Theni and Dindigul, Tamil Nadu, for their invaluable support and assistance during this study. The authors also extend their gratitude to Mr. T Balaji and the supporting staff of the Unit of Molecular Biology and Diagnosis, ICMR-Vector Control Research Centre, Field Station, Madurai, for their dedicated efforts in the processing and analysing samples.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Xu, G.; Walker, D.H.; Jupiter, D.; Melby, P.C.; Arcari, C.M. A Review of the Global Epidemiology of Scrub Typhus. PLoS Negl Trop Dis 2017, 11, e0006062. [Google Scholar] [CrossRef] [PubMed]
- Lerdthusnee, K.; Khlaimanee, N.; Monkanna, T.; Sangjun, N.; Mungviriya, S.; Linthicum, K.J.; Frances, S.P.; Kollars, T.M.; Coleman, R.E. Efficiency of Leptotrombidium Chiggers (Acari: Trombiculidae) at Transmitting Orientia Tsutsugamushi to Laboratory Mice. J Med Entomol 2002, 39, 521–525. [Google Scholar] [CrossRef] [PubMed]
- Izzard, L.; Fuller, A.; Blacksell, S.D.; Paris, D.H.; Richards, A.L.; Aukkanit, N.; Nguyen, C.; Jiang, J.; Fenwick, S.; Day, N.P.J.; et al. Isolation of a Novel Orientia Species ( O. Chuto Sp. Nov.) from a Patient Infected in Dubai. J Clin Microbiol 2010, 48, 4404–4409. [Google Scholar] [CrossRef] [PubMed]
- Abarca, K.; Martínez-Valdebenito, C.; Angulo, J.; Jiang, J.; Farris, C.M.; Richards, A.L.; Acosta-Jamett, G.; Weitzel, T. Molecular Description of a Novel Orientia Species Causing Scrub Typhus in Chile. Emerg Infect Dis 2020, 26, 2148–2156. [Google Scholar] [CrossRef]
- Taylor, A.J.; Paris, D.H.; Newton, P.N. A Systematic Review of Mortality from Untreated Scrub Typhus (Orientia Tsutsugamushi). PLoS Negl Trop Dis 2015, 9, e0003971. [Google Scholar] [CrossRef]
- Varghese, G.M.; Trowbridge, P.; Janardhanan, J.; Thomas, K.; Peter, J.V.; Mathews, P.; Abraham, O.C.; Kavitha, M.L. Clinical Profile and Improving Mortality Trend of Scrub Typhus in South India. International Journal of Infectious Diseases 2014, 23, 39–43. [Google Scholar] [CrossRef]
- Sonthayanon, P.; Peacock, S.J.; Chierakul, W.; Wuthiekanun, V.; Blacksell, S.D.; Holden, M.T.G.; Bentley, S.D.; Feil, E.J.; Day, N.P.J. High Rates of Homologous Recombination in the Mite Endosymbiont and Opportunistic Human Pathogen Orientia Tsutsugamushi. PLoS Negl Trop Dis 2010, 4, e752. [Google Scholar] [CrossRef]
- Paris, D.H.; Shelite, T.R.; Day, N.P.; Walker, D.H. Unresolved Problems Related to Scrub Typhus: A Seriously Neglected Life-Threatening Disease. The American Society of Tropical Medicine and Hygiene 2013, 89, 301–307. [Google Scholar] [CrossRef]
- Batty, E.M.; Chaemchuen, S.; Blacksell, S.; Richards, A.L.; Paris, D.; Bowden, R.; Chan, C.; Lachumanan, R.; Day, N.; Donnelly, P.; et al. Long-Read Whole Genome Sequencing and Comparative Analysis of Six Strains of the Human Pathogen Orientia Tsutsugamushi. PLoS Negl Trop Dis 2018, 12, e0006566. [Google Scholar] [CrossRef]
- Kelly, D.J.; Fuerst, P.A.; Ching, W.; Richards, A.L. Scrub Typhus: The Geographic Distribution of Phenotypic and Genotypic Variants of Orientia Tsutsugamushi. Clinical Infectious Diseases 2009, 48, S203–S230. [Google Scholar] [CrossRef]
- Valbuena, G.; Walker, D.H. Approaches to Vaccines against Orientia Tsutsugamushi. Front Cell Infect Microbiol 2013, 2. [Google Scholar] [CrossRef] [PubMed]
- Varghese, G.M.; Janardhanan, J.; Mahajan, S.K.; Tariang, D.; Trowbridge, P.; Prakash, J.A.J.; David, T.; Sathendra, S.; Abraham, O.C. Molecular Epidemiology and Genetic Diversity of Orientia Tsutsugamushi from Patients with Scrub Typhus in 3 Regions of India. Emerg Infect Dis 2015, 21, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Devamani, C.S.; Prakash, J.A.J.; Alexander, N.; Stenos, J.; Schmidt, W.-P. The Incidence of Orientia Tsutsugamushi Infection in Rural South India. Epidemiol Infect 2022, 150, e132. [Google Scholar] [CrossRef]
- Khan, S.A.; Bora, T.; Laskar, B.; Khan, A.M.; Dutta, P. Scrub Typhus Leading to Acute Encephalitis Syndrome, Assam, India. Emerg Infect Dis 2017, 23, 148–150. [Google Scholar] [CrossRef]
- Bonell, A.; Lubell, Y.; Newton, P.N.; Crump, J.A.; Paris, D.H. Estimating the Burden of Scrub Typhus: A Systematic Review. PLoS Negl Trop Dis 2017, 11, e0005838. [Google Scholar] [CrossRef] [PubMed]
- Nallan, K.; Kalidoss, B.C.; Jacob, E.S.; Mahadevan, S.K.; Joseph, S.; Ramalingam, R.; Renu, G.; Thirupathi, B.; Ramasamy, B.; Gupta, B.; et al. A Novel Genotype of Orientia Tsutsugamushi in Human Cases of Scrub Typhus from Southeastern India. Microorganisms 2025, 13, 333. [Google Scholar] [CrossRef]
- Ruang-areerate, T.; Jeamwattanalert, P.; Rodkvamtook, W.; Richards, A.L.; Sunyakumthorn, P.; Gaywee, J. Genotype Diversity and Distribution of Orientia Tsutsugamushi Causing Scrub Typhus in Thailand. J Clin Microbiol 2011, 49, 2584–2589. [Google Scholar] [CrossRef]
- Arai, S.; Tabara, K.; Yamamoto, N.; Fujita, H.; Itagaki, A.; Kon, M.; Satoh, H.; Araki, K.; Tanaka-Taya, K.; Takada, N.; et al. Molecular Phylogenetic Analysis of Orientia Tsutsugamushi Based on the GroES and GroEL Genes. Vector-Borne and Zoonotic Diseases 2013, 13, 825–829. [Google Scholar] [CrossRef]
- Ramaiah, A.; Koralur, M.C.; Dasch, G.A. Complexity of Type-Specific 56 KDa Antigen CD4 T-Cell Epitopes of Orientia Tsutsugamushi Strains Causing Scrub Typhus in India. PLoS One 2018, 13, e0196240. [Google Scholar] [CrossRef]
- Nallan, K.; Rajan, G.; Sivathanu, L.; Devaraju, P.; Thiruppathi, B.; Kumar, A.; Rajaiah, P. Molecular Detection of Multiple Genotypes of Orientia Tsutsugamushi Causing Scrub Typhus in Febrile Patients from Theni District, South India. Trop Med Infect Dis 2023, 8, 174. [Google Scholar] [CrossRef]
- Trend Of Scrub Typhus In Tamil Nadu-2021-2023 (Based On Ihip Data).
- Li, W.; Dou, X.; Zhang, L.; Lyu, Y.; Du, Z.; Tian, L.; Zhang, X.; Sun, Y.; Guan, Z.; Chen, L.; et al. Laboratory Diagnosis and Genotype Identification of Scrub Typhus from Pinggu District, Beijing, 2008 and 2010. Am J Trop Med Hyg 2013, 89, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol Biol Evol 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Samson, S.; Lord, É.; Makarenkov, V. SimPlot++: A Python Application for Representing Sequence Similarity and Detecting Recombination. Bioinformatics 2022, 38, 3118–3120. [Google Scholar] [CrossRef]
- Phetsouvanh, R.; Sonthayanon, P.; Pukrittayakamee, S.; Paris, D.H.; Newton, P.N.; Feil, E.J.; Day, N.P.J. The Diversity and Geographical Structure of Orientia Tsutsugamushi Strains from Scrub Typhus Patients in Laos. PLoS Negl Trop Dis 2015, 9, e0004024. [Google Scholar] [CrossRef] [PubMed]
- Duong, V.; Blassdell, K.; May, T.T.X.; Sreyrath, L.; Gavotte, L.; Morand, S.; Frutos, R.; Buchy, P. Diversity of Orientia Tsutsugamushi Clinical Isolates in Cambodia Reveals Active Selection and Recombination Process. Infection, Genetics and Evolution 2013, 15, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Lin, P.R.; Tsai, H.P.; Tsui, P.Y.; Weng, M.H.; Kuo, M.D.; Lin, H.C.; Chen, K.C.; Ji, D.D.; Chu, D.M.; Liu, W.T. Genetic Typing, Based on the 56-Kilodalton Type-Specific Antigen Gene, of Orientia Tsutsugamushi Strains Isolated from Chiggers Collected from Wild-Caught Rodents in Taiwan. Appl Environ Microbiol 2011, 77, 3398–3405. [Google Scholar] [CrossRef]
- Kelly, D.J.; Fuerst, P.A.; Richards, A.L. Origins, Importance and Genetic Stability of the Prototype Strains Gilliam, Karp and Kato of Orientia Tsutsugamushi. Trop Med Infect Dis 2019, 4, 75. [Google Scholar] [CrossRef]
- Zhang, J.; Yang, J.-R. Determinants of the Rate of Protein Sequence Evolution. Nat Rev Genet 2015, 16, 409–420. [Google Scholar] [CrossRef]
- Paris, D.H.; Aukkanit, N.; Jenjaroen, K.; Blacksell, S.D.; Day, N.P.J. A Highly Sensitive Quantitative Real-Time PCR Assay Based on the GroEL Gene of Contemporary Thai Strains of Orientia Tsutsugamushi. Clinical Microbiology and Infection 2009, 15, 488–495. [Google Scholar] [CrossRef]
- Ohashi, N.; Nashimoto, H.; Ikeda, H.; Tamura, A. Diversity of Immunodominant 56-KDa Type-Specific Antigen (TSA) of Rickettsia Tsutsugamushi. Sequence and Comparative Analyses of the Genes Encoding TSA Homologues from Four Antigenic Variants. Journal of Biological Chemistry 1992, 267, 12728–12735. [Google Scholar] [CrossRef]
- Yang, H.-H.; Huang, I.-T.; Lin, C.-H.; Chen, T.-Y.; Chen, L.-K. New Genotypes of Orientia Tsutsugamushi Isolated from Humans in Eastern Taiwan. PLoS One 2012, 7, e46997. [Google Scholar] [CrossRef] [PubMed]
- Chunduru, K.; A. R, M.; Poornima, S.; Hande H, M.; Devaki, R.; Varghese, G.M.; Saravu, K. Clinical, Laboratory Profile and Molecular Characterization of Orientia Tsutsugamushi among Fatal Scrub Typhus Patients from Karnataka, India. Infect Dis 2024, 56, 220–229. [Google Scholar] [CrossRef] [PubMed]
- Kelly, D.J.; Fuerst, P.A.; Ching, W.; Richards, A.L. Scrub Typhus: The Geographic Distribution of Phenotypic and Genotypic Variants of Orientia Tsutsugamushi. Clinical Infectious Diseases 2009, 48, S203–S230. [Google Scholar] [CrossRef] [PubMed]
- Izzard, L.; Fuller, A.; Blacksell, S.D.; Paris, D.H.; Richards, A.L.; Aukkanit, N.; Nguyen, C.; Jiang, J.; Fenwick, S.; Day, N.P.J.; et al. Isolation of a Novel Orientia Species ( O. Chuto Sp. Nov.) from a Patient Infected in Dubai. J Clin Microbiol 2010, 48, 4404–4409. [Google Scholar] [CrossRef]
- Frances, S.P.; Watcharapichat, P.; Phulsuksombati, D.; Tanskul, P. Transmission of Orientia Tsutsugamushi, the Aetiological Agent for Scrub Typhus, to Co-Feeding Mites. Parasitology 2000, 120, 601–607. [Google Scholar] [CrossRef]
- Govindarajan, R.; Rajamannar, V.; Krishnamoorthi, R.; Kumar, A.; Samuel, P.P. Distribution Pattern of Chigger Mites in South Tamil Nadu, India. Entomon 2021, 46, 247–254. [Google Scholar] [CrossRef]
- Park, S.-W.; Lee, C.K.; Kwak, Y.G.; Moon, C.; Kim, B.-N.; Kim, E.S.; Kang, J.M.; Lee, C.-S. Antigenic Drift of Orientia Tsutsugamushi in South Korea as Identified by the Sequence Analysis of a 56-KDa Protein-Encoding Gene. The American Society of Tropical Medicine and Hygiene 2010, 83, 930–935. [Google Scholar] [CrossRef]
- Kim, G.; Ha, N.-Y.; Min, C.-K.; Kim, H.-I.; Yen, N.T.H.; Lee, K.-H.; Oh, I.; Kang, J.-S.; Choi, M.-S.; Kim, I.-S.; et al. Diversification of Orientia Tsutsugamushi Genotypes by Intragenic Recombination and Their Potential Expansion in Endemic Areas. PLoS Negl Trop Dis 2017, 11, e0005408. [Google Scholar] [CrossRef]
Figure 1.
Neighbour-joining (NJ) tree of the GroEL gene using the K2P model with 1000 bootstrap replicates. The tree includes four main clades: a novel Gilliam-related lineage (INDv), Ot-TJTN-like, Kato-like, and Karp-like groups. Sequences from this study are shown in blue, and the novel INDv-Gilliam cluster (INDv) is in the red bullet. The tree is rooted using a reference sequence as an outgroup (JX188965).
Figure 1.
Neighbour-joining (NJ) tree of the GroEL gene using the K2P model with 1000 bootstrap replicates. The tree includes four main clades: a novel Gilliam-related lineage (INDv), Ot-TJTN-like, Kato-like, and Karp-like groups. Sequences from this study are shown in blue, and the novel INDv-Gilliam cluster (INDv) is in the red bullet. The tree is rooted using a reference sequence as an outgroup (JX188965).
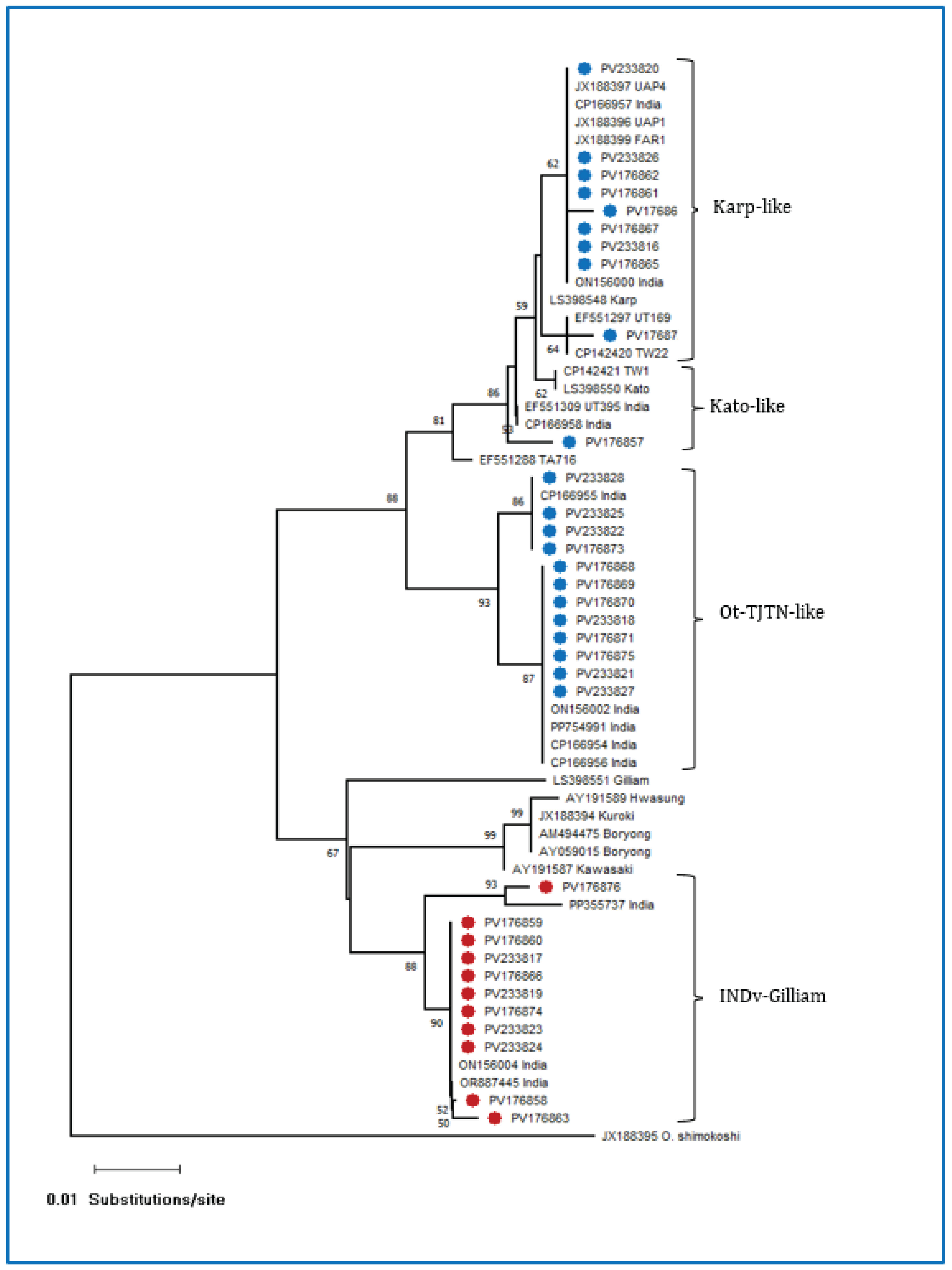
Figure 2.
Radiation distance phylogenetic tree of GroEL gene sequences. The tree reveals two distinct clusters: the newly designated INDv-Gilliam lineage (Red outline) and Gilliam reference strains (Cyan blue).
Figure 2.
Radiation distance phylogenetic tree of GroEL gene sequences. The tree reveals two distinct clusters: the newly designated INDv-Gilliam lineage (Red outline) and Gilliam reference strains (Cyan blue).
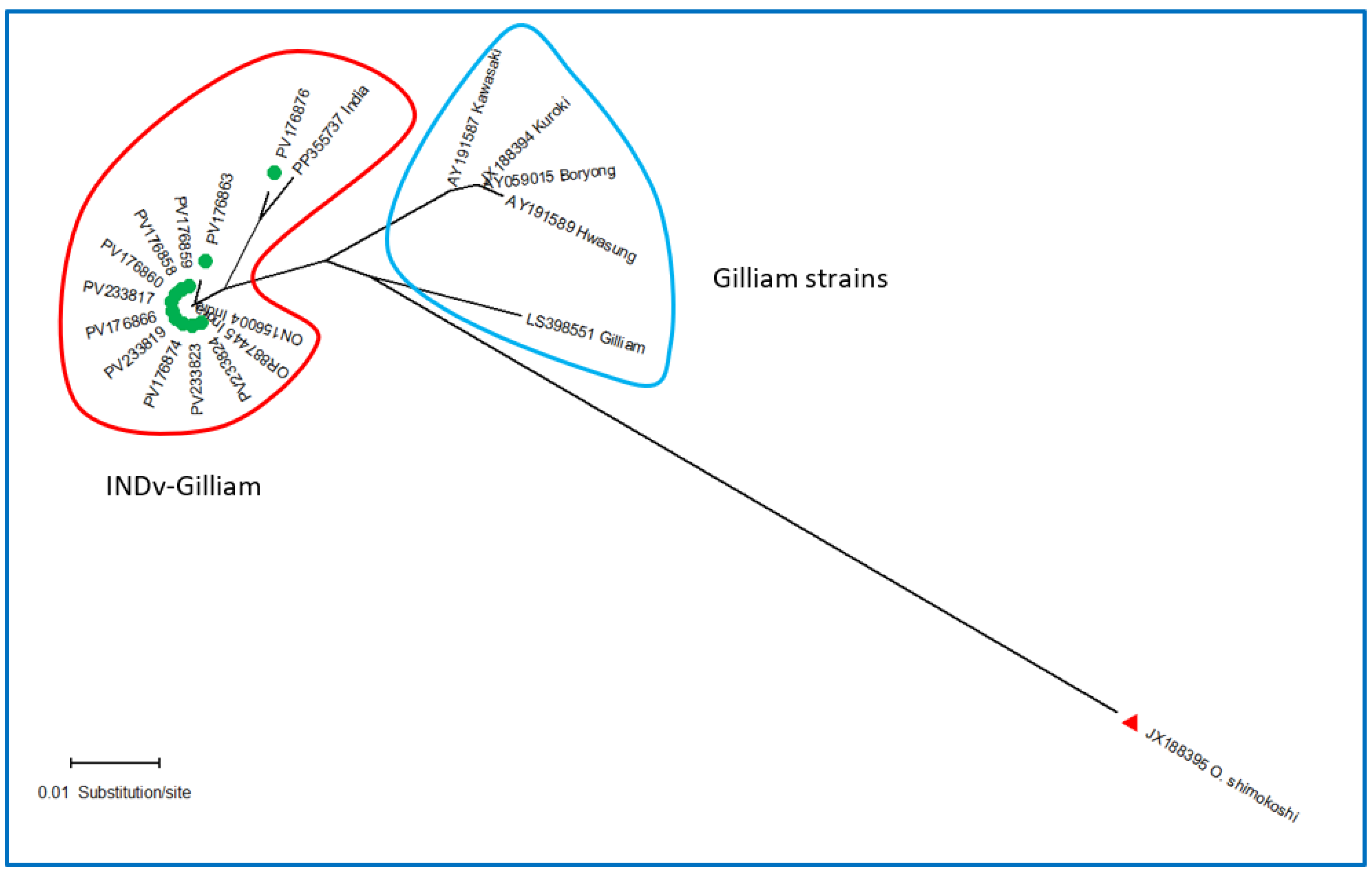
Figure 3.
Neighbor-joining (NJ) tree of the 56-kDa gene constructed using the K2P model with 1000 bootstrap replicates. The tree shows three major clades: Karp-like, Ot-TJTN-like, and Gilliam-like. Sequences from this study are indicated with red bullets. Two reference sequences (AF173042 & M63381) were used as outgroups to root the tree.
Figure 3.
Neighbor-joining (NJ) tree of the 56-kDa gene constructed using the K2P model with 1000 bootstrap replicates. The tree shows three major clades: Karp-like, Ot-TJTN-like, and Gilliam-like. Sequences from this study are indicated with red bullets. Two reference sequences (AF173042 & M63381) were used as outgroups to root the tree.
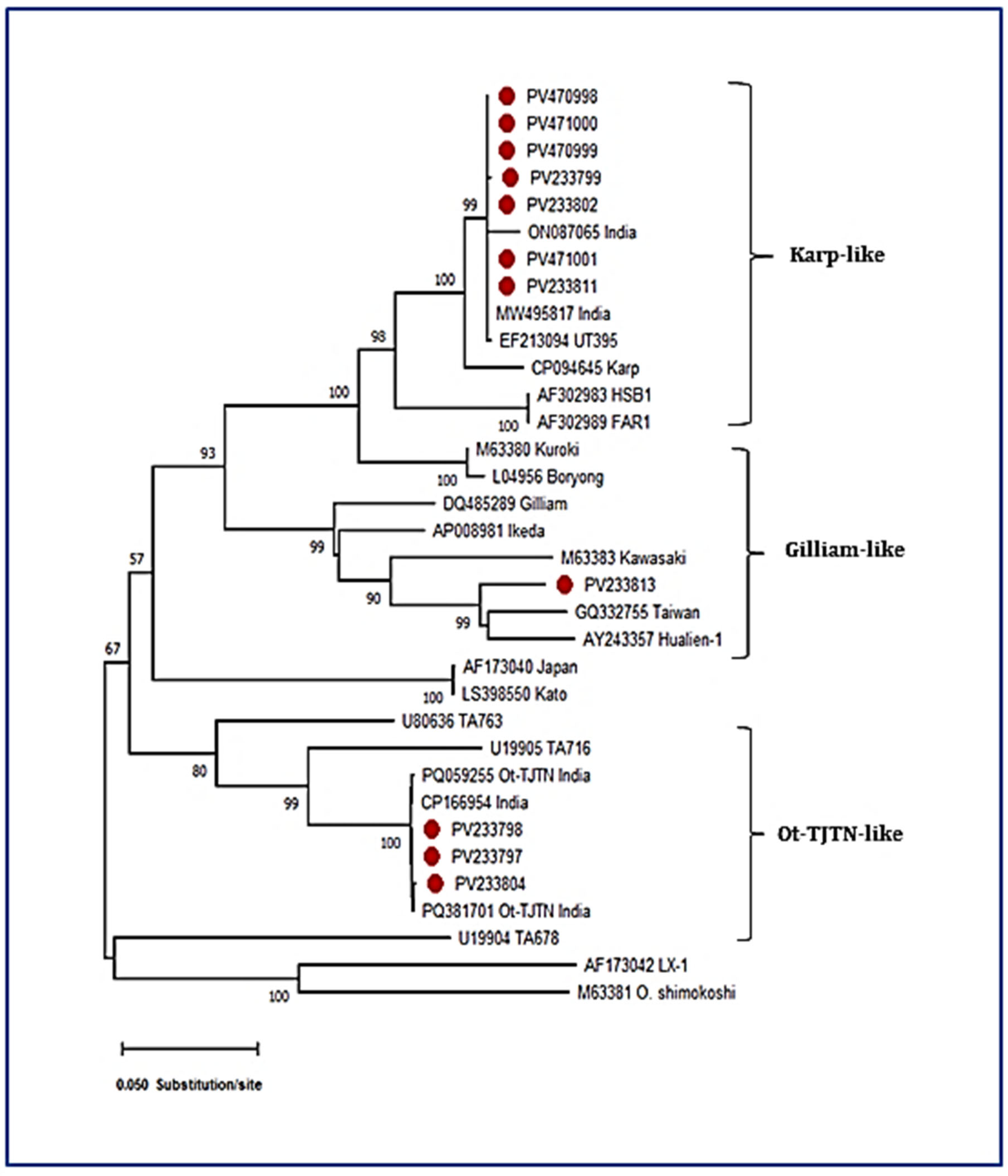
Figure 4.
(a) Similarity plot (SimPlot) analysis of the Ot-TJTN isolate against reference strains, highest sequence similarity to TA716 and Kato. (b). SimPlot analysis of the Karp clade revealed the highest similarity to the Karp, Kuroki and Boryong reference strains, supporting their classification within the Karp lineage. A stable flat curve across the genome suggests a consistent relationship to the reference throughout the region.
Figure 4.
(a) Similarity plot (SimPlot) analysis of the Ot-TJTN isolate against reference strains, highest sequence similarity to TA716 and Kato. (b). SimPlot analysis of the Karp clade revealed the highest similarity to the Karp, Kuroki and Boryong reference strains, supporting their classification within the Karp lineage. A stable flat curve across the genome suggests a consistent relationship to the reference throughout the region.
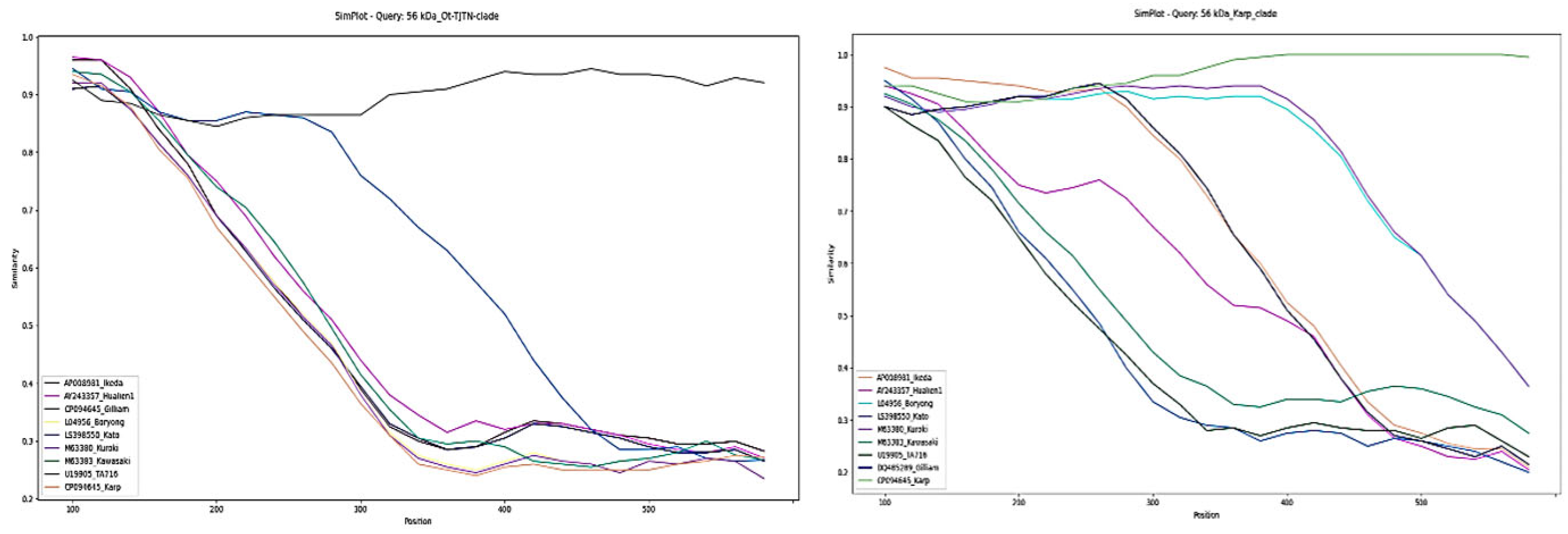
Figure 5.
(a) BootScan analysis of the Ot-TJTN isolate demonstrated recombination events involving TA716 and Kato strains (b) BootScan analysis of Karp clade isolates indicated recombination between Karp and Ikeda genotypes.
Figure 5.
(a) BootScan analysis of the Ot-TJTN isolate demonstrated recombination events involving TA716 and Kato strains (b) BootScan analysis of Karp clade isolates indicated recombination between Karp and Ikeda genotypes.
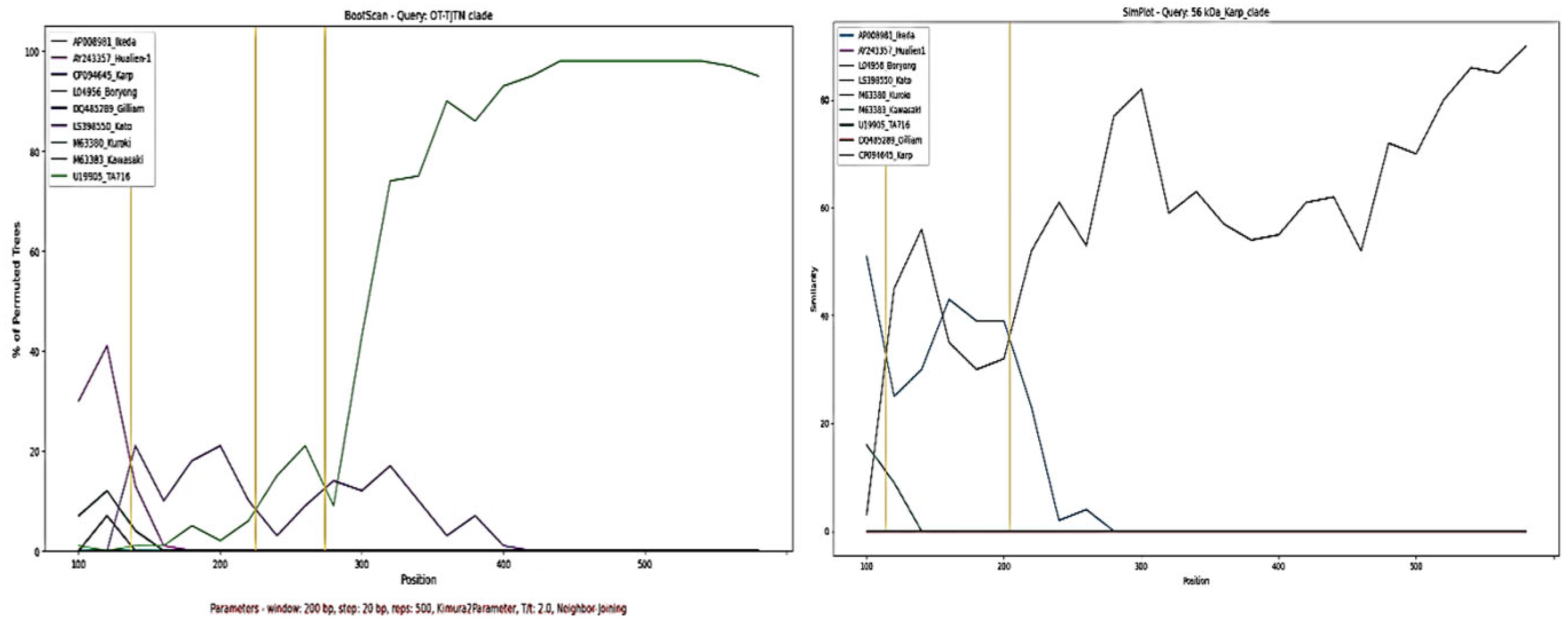

Table 1.
Details of samples, GenBank accession numbers of GroEL gene sequences, and their percentage identity to query sequences.
Table 1.
Details of samples, GenBank accession numbers of GroEL gene sequences, and their percentage identity to query sequences.
| Sl No. | DNA Code | GenBank Acc. Nos. (Present Study) | Percentage Identity | Source | Country | Closest GenBank Acc. No. |
|---|---|---|---|---|---|---|
| 1 | TH 05 | PV176858 | 100 100 98.11 96.96 |
Human Human Rodent Human |
India India India Korea |
ON156004 OR887445 PP355737 AY191587 |
| 2 | TH 10 | PV176859 | 100 100 96.98 96.98 |
Human Human Human Human |
India India Korea Korea |
ON156004 OR887445 AM494475 AY191587 |
| 3 | TH 16 | PV176860 | 100 96.9 96.9 96.9 |
Human Human Human Human |
India Korea India Korea |
ON156004 AY059015 CP166955 AY191587 |
| 4 | TH 28 | PV176863 | 99.73 96.7 96.7 |
Human Human Human |
India Korea Korea |
ON156004 AM494475 AY191587 |
| 5 | TH 45 | PV233817 | 100 98.11 96.98 96.98 |
Human Rodent Human Human |
India India Korea Korea |
ON156004 PP355737 AM494475 AY191587 |
| 6 | DM 03 |
PV176866 | 100 100 96.7 97 |
Human Human Human Human |
India India Korea Korea |
ON156004 OR887445 AM494475 AY191587 |
| 7 | DM 14 | PV233819 | 100 100 96.98 96.98 |
Human Human Human Human |
India India Korea Korea |
ON156004 |
| OR887445 | ||||||
| AM494475 | ||||||
| AY191587 | ||||||
| 8 | DM 30 | PV176874 | 100 | Human | India | ON156004 |
| 100 | Human | India | OR887445 | |||
| 96.99 | Human | Korea | AM494475 | |||
| 96.99 | Human | Korea | AY191587 | |||
| 9 | DM 33 | PV176876 | 98.63 | Human | India | ON156004 |
| 99.12 | Rodent | India | PP355737 | |||
| 96.16 | Human | Korea | AM494475 | |||
| 96.16 | Human | Korea | AY191587 | |||
| 10 | DM 43 | PV233823 | 100 | Human | India | ON156004 |
| 100 | Human | India | OR887445 | |||
| 96.98 | Human | Japan | JX188394 | |||
| 96.98 | Human | Korea | AY191587 | |||
| 96.98 | Human | Korea | AM494475 | |||
| 11 | DM 44 | PV233824 | 100 | Human | India | ON156004 |
| 100 | Human | India | OR887445 | |||
| 96.98 | Human | Japan | JX188394 | |||
| 96.98 | Human | Korea | AY191587 | |||
| 96.98 | Human | Korea | AM494475 |
Table 2.
Estimates of nucleotide diversity over sequence pairs between groups.
| Genotype | INDv-Gilliam | Hwasung | Kawasaki | Gilliam | Kuroki | Boryong |
|---|---|---|---|---|---|---|
| INDv-Gilliam | — | |||||
| Hwasung | 0.039 | — | ||||
| Kawasaki | 0.032 | 0.006 | — | |||
| Gilliam | 0.039 | 0.047 | 0.041 | — | ||
| Kuroki | 0.035 | 0.003 | 0.003 | 0.044 | — | |
| Boryong | 0.035 | 0.003 | 0.003 | 0.044 | 0.001 | — |
Table 3.
Details of samples, GenBank accession numbers of 56-kDa gene sequences, and their percentage identity to query sequences.
Table 3.
Details of samples, GenBank accession numbers of 56-kDa gene sequences, and their percentage identity to query sequences.
| Sl No. | DNA Code | GenBank Acc. Nos. (Present Study) | Percentage Identity | Source | Country | Closest GenBank Acc. No. |
|---|---|---|---|---|---|---|
| 1 | TH 05 | PV233797 | 100 100 100 |
Human Human Human |
India India India |
PQ381701 PQ059255 CP166954 |
| 2 | TH 10 | PV233798 | 100 100 100 |
Human Human Human |
India India India |
PQ381701 PQ059255 CP166954 |
| 3 | DM 3 | PV233804 | 100 100 100 |
Human Human Human |
India India India |
PQ381701 PQ059255 CP166954 |
| 4 | TH 16 | PV233799 | 100 98.84 98.15 |
Human Human Human |
India India Thailand |
MW495817 ON087065 EF213094 |
| 5 | TH 28 | PV233802 | 100 98.84 98.15 |
Human Human Human |
India India Thailand |
MW495817 ON087065 EF213094 |
| 6 | TH 45 | PV470998 | 100 98.84 98.15 |
Human Human Human |
India India Thailand |
MW495817 ON087065 EF213094 |
| 7 | DM 43 | PV471000 | 100 98.84 98.15 |
Human Human Human |
India India Thailand |
MW495817 ON087065 EF213094 |
| 8 | DM 44 | PV471001 | 100 98.84 98.15 |
Human Human Human |
India India Thailand |
MW495817 ON087065 EF213094 |
| 9 | DM 14 | PV470999 | 100 98.84 98.15 |
Human Human Human |
India India Thailand |
MW495817 ON087065 EF213094 |
| 10 | DM 30 | PV233811 | 100 98.84 98.15 |
Human Human Human |
India India Thailand |
MW495817 ON087065 EF213094 |
| 11 | DM 33 | PV233813 | 95.14 93.93 93.93 93.93 92.03 |
Human Human Human Human Human |
Taiwan Taiwan China Taiwan Japan |
GQ332755 AY243357 MT258819 GQ332754 AP008981 |
Table 4.
Mixed genotypic classification of isolates based on the GroEl and 56-kDa gene.
| S. No | GenBank Accession. Nos. (GroEL) |
GroEL Genotype | GenBank Acc. Nos. (56 kDa) | 56kDa Genotype | Dual Genotype |
|---|---|---|---|---|---|
| 1. | PV176860 | INDv_Gilliam | PV233799 | Karp-like | INDv-Gilliam/Karp-like |
| 2. | PV176863 | INDv_Gilliam | PV233802 | Karp-like | INDv-Gilliam/Karp-like |
| 3. | PV233817 | INDv_Gilliam | PV470998 | Karp-like | INDv-Gilliam/Karp-like |
| 4. | PV233819 | INDv_Gilliam | PV470999 | Karp-like | INDv-Gilliam/Karp-like |
| 5. | PV176874 | INDv_Gilliam | PV233811 | Karp-like | INDv-Gilliam/Karp-like |
| 6. | PV233823 | INDv_Gilliam | PV471000 | Karp-like | INDv-Gilliam/Karp-like |
| 7. | PV233824 | INDv_Gilliam | PV471001 | Karp-like | INDv-Gilliam/Karp-like |
| 8. | PV176858 | INDv_Gilliam | PV233797 | Ot-TJTN | INDv_Gilliam/ Ot-TJTN |
| 9. | PV176859 | INDv_Gilliam | PV233798 | Ot-TJTN | INDv_Gilliam/ Ot-TJTN |
| 10. | PV176866 | INDv_Gilliam | PV233804 | Ot-TJTN | INDv_Gilliam/ Ot-TJTN |
| 11. | PV176876 | INDv_Gilliam | PV233813 | Gilliam-type | INDv_Gilliam /Gilliam |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



