
Submitted:
05 June 2025
Posted:
05 June 2025
You are already at the latest version
Abstract
Host selectivity or preference plays a critical role in enabling parasitic plants to identify suitable hosts and influence plant community dynamics. Phosphorus (P) is known to affect the growth of root hemiparasitic plants and their interaction with single host species, but its role in shaping host selectivity across multiple hosts is unclear. In a pot experiment, we evaluated whether the root hemiparasitic plant Pedicularis kansuensis exhibits selective parasitism on legumes (Medicago sativa) versus grasses (Elymus nutans) and assessed the impact of soil P availability on this preference. The results showed that P. kansuensis inhibited the growth of both host species, but the magnitude of suppression varied with P availability. Under low P conditions, P. kansuensis preferentially parasitized the tender M. sativa, causing a greater biomass reduction in the legume. In contrast, at high P levels, P. kansuensis dropped to forage on legumes and had more chances to parasitize the dominant E. nutans, leading to stronger suppression of grass growth. Our findings demonstrate that soil P availability modulates host selectivity in P. kansuensis, emphasizing the influence of soil nutrient conditions on parasitic-host dynamics. This research provides insights into managing the impacts of parasitic plants on plant community structure through nutrient interventions.
Keywords:
host preference
; legume-grass mixture
; parasitic plant
; phosphorous
; haustorium
1. Introduction
Root hemiparasitic plants account for more than half of all parasitic plant species and are commonly found in diverse natural and semi-natural environments [1]. These plants extract essential resources from host plants, including carbon (C), nitrogen (N), phosphorus (P), and water, via specialized structures called haustoria [2,3]. This parasitism significantly inhibits host growth and reduces overall plant productivity [4,5]. Additionally, root hemiparasites influence interactions between host and non-host plants, altering plant community composition and diversity [6,7]. While most root hemiparasitic plants are generalists capable of parasitizing a wide range of neighboring plant species, they exhibit clear preferences for certain hosts [5]. Foraging for preferred hosts (host selectivity) is crucial for successful parasitism and has profound implications for plant community dynamics. However, the mechanisms driving host selectivity in root hemiparasitic plants remain poorly understood.
Host selectivity in root hemiparasitic plants generally occurs in three stages: growth toward the host root (host tropism), attachment to the host root, and haustorium formation. Although research into these processes is still emerging, there is evidence that root hemiparasites exhibit host tropism. For instance, a recent study demonstrated that seedlings of the facultative root hemiparasite Rhinanthus minus preferentially grew toward legume roots rather than grass roots in an agar plate experiment [8]. However, while this short-term (3-week) study provided direct evidence of early-stage host selectivity, it did not examine whether such tropism results in the formation of more haustoria on legume hosts or leads to greater suppression of legume biomass compared to grasses in multi-species systems. A pot experiment found that the obligate root hemiparasite Melampyrum arense had a more pronounced negative impact on the legume host Medicago sativa than on the grass Lolium perenne in grass-legume mixture pots [9]. These findings suggest that root hemiparasites may preferentially target legumes over grasses, causing uneven effects between host species. However, as obligate root hemiparasites depend entirely on their hosts for survival from germination onward [2,10], host selectivity is more critical for their survival than for facultative hemiparasites. In contrast, facultative hemiparasites can grow independently for a portion of their lifecycle and may not exhibit the same degree of host specificity [2,3]. Whether facultative hemiparasites demonstrate host selectivity and impose unequal effects on coexisting host species remains largely unexplored.
Although root hemiparasitic plants have shown a preference for suppressing legume hosts over grass hosts in aforementioned pot experiments, this phenomenon was not consistently observed in field experiments. For example, Rhinanthus species typically harmed grass biomass but had varying effects on legume biomass - positive [11,12], negative [13,14], or neutral [15]. Several reasons have been proposed to explain these contrasting effects of parasitism on legumes. Firstly, different plant species have been found to differ in their quality as hosts [16], so the species diversity of legumes in field conditions may be a factor. Secondly, the abundance of legumes in the plant community also plays a role in determining the parasite’s effect on legume biomass, as the most abundant host species are more likely to be encountered by a parasite than less abundant ones [17]. Additionally, environmental factors such as nutrient availability may significantly influence host selectivity for legumes in natural habitats.
Nitrogen (N) and phosphorus (P) are essential macronutrients that influence the growth of hemiparasitic plants and meanwhile the primary mineral nutrients the parasites extract from their hosts [18,19]. Previous studies found that soil N and P levels impacted the growth of hemiparasitic plants [20] and their interactions with host plants [21,22,23]. However, they were mainly conducted in single host-parasite pairs, limiting our understanding of how nutrients influence the interactions between hemiparasitic plants and their coexisting hosts in natural environments. While N has been a primary focus of studies examining nutrient effects on parasitic plants [24], the role of P remains underexplored [22]. Phosphorus not only affects the growth of hemiparasitic plants [25,26] but also influences grass-legume interactions [27,28] and the secretion of plant exudates, such as flavonoids and strigolactones [29,30,31], which may act as chemical signals for host tropism [8,32]. Therefore, we hypothesize that the host selectivity of root hemiparasitic plants between grass and legume hosts may be modulated by soil P levels.
Pedicularis kansuensis, a facultative root hemiparasitic plant, is widespread in subalpine regions of western China [33], including the Qinghai-Tibet Plateau and Tianshan Mountains[34]. This species has a significant ecological and economic impact, as it can reduce grassland productivity by up to 80%, thereby threatening the local livestock industry [35]. Previous research indicates that P. kansuensis parasitizes a wide range of hosts, with a preference for species in the Poaceae (grasses) and Fabaceae (legumes) [36]. However, the host selectivity of P. kansuensis between coexisting grass and legume hosts and the factors that influence its host selectivity in mixed-species systems remain poorly understood. This study investigates whether P. kansuensis selectively parasitizes a legume host (Medicago sativa) over a grass host (Elymus nutans) in a grass-legume mixture and whether soil P availability affects this selectivity. By clarifying the interactions between P. kansuensis and its hosts under varying P levels, this research aims to improve our understanding of nutrient-mediated parasitism and provide insights for managing the impacts of parasitic plants in grassland ecosystems.
2. Results
2.1. Haustorium Formation of the Hemiparasite
Pedicularis kansuensis formed more PFH on the grass host compared to the legume host, especially under 1P (p<0.05) and 2P (p<0.05) treatments (Figure 1A). However, the number of PFH per gram of dry root was higher on legume roots than grass roots under low P conditions (particularly in 1P treatment, p<0.05) but not under 2P condition (p>0.05, Figure 1B). P availability had no significant impact on the number of PFH on grass roots (p>0.05) but did affect that in the legume host (p<0.05) (Figure 1A). The total number of haustoria produced by P. kansuensis was not significantly affected by P availability (p>0.05; Figure 1C). Conversely, the total number of haustoria per gram dry root of P. kansuensis was significantly decreased under 2P treatments compared to 0P (p<0.01) and 1P (p<0.01) conditions (Figure 1D).
2.2. Growth Performance and Biomass Allocation of Hosts and the Hemiparasite
There were no significant interaction effects between P availability and parasitism on the growth performance of E. nutans and M. sativa, except on the root DW of M. sativa (Table 1). Both P levels and parasitism significantly affected the shoot DWs of E. nutans and M. sativa, as well as the root DWs of M. sativa, but had no notable impact on the root DW of E. nutans (Table 1). Infection by P. kansuensis significantly reduced the shoot and root DWs of both host species (Figure 2 and 3), and their impacts on host growth were more severe than P availability, as indicated by the larger ηP2 values in Table 1. Parasitized E. nutans exhibited a decreased shoot DW as P levels increased (Figure 2A), while the shoot and root DWs of infected M. sativa increased with higher P availability (Figure 2A,B).
P availability had no obvious effect on the root: shoot ratio of infected (p>0.05) or uninfected (p>0.05) E. nutans, despite the root: shoot ratio of infected E. nutans appeared to increase with higher levels of P (Figure 2C). The root: shoot ratios of uninfected M. sativa varied significantly among P treatments, while there were no apparent differences for infected M. sativa (p>0.05; Figure 2C). Compared to uninfected controls, the root: shoot ratio of infected M. sativa (p<0.05) and E. nutans (p<0.05) was significantly higher under 2P conditions (Figure 2C).
For P. kansuensis, its shoot DW (p<0.05), root DW (p<0.01) and root: shoot ratio (p<0.05) were significantly affected by P availability. Higher P availability (2P) significantly increased both shoot and root DWs compared to 1P, while P deficiency (0P) did not suppress its growth (Figure 2A,B). In addition, the root: shoot ratio of P. kansuensis was higher under 2P than other P levels (Figure 2C).
2.3. Host Suppression by P. kansuensis
The degree of growth suppression caused by P. kansuensis varied between E. nutans and M. sativa, which was influenced by P availability (Figure 3). At low P levels, M. sativa experienced significantly greater suppression of shoot (0P, 82.08%; 1P, 79.39%) and root DWs (0P, 77.39%; 1P, 81.38%) compared to E. nutans (0P, shoot 50.81%, root 47.20%; 1P, shoot 48.98%, root 27.02%). However, under high P conditions (2P), growth suppression of E. nutans increased substantially (shoot, 78.68%; root, 45.54%), while M. sativa (shoot, 64.41%; root, 36.65%) showed reduced inhibition (Figure 3).
2.4. Biomass Proportions of Host and Hemiparasite per Pot
The shoot and root DW proportion of E. nutans consistently exceeded that of M. sativa across all treatments (Figure 4A,C). Infection by P. kansuensis reduced the shoot DW proportions of E. nutans, which was more seriously at 2P treatment (54.63%, p<0.05) than 0P (16.98%, p<0.05) and 1P (12.41%, p<0.05) conditions. Conversely, the suppression of shoot DW proportion of M. sativa by P. kansuensis was lower at high P level (2P, 30.86%, p>0.05) than at low P treatments (0P, 69.39%, p<0.01; 1P, 66.00%, p<0.01). Infection by P. kansuensis significantly suppressed the root DW proportion of E. nutans under 2P treatment (27.80%, p<0.05), but obviously inhibited that of M. sativa under 0P (59.78%, p<0.05) and 1P (73.50%, p<0.05) treatments. The shoot and root DW proportion of P. kansuensis was highest under 2P conditions and similar between 0P and 1P treatments (Figure 4B,D). Total shoot biomass per pot decreased significantly in the presence of P. kansuensis (Table 1), and the percentage suppression of total shoot biomass by P. kansuensis increased with P availability (Figure 5).
2.5. Shoot N and P Status of Hosts and Hemiparasite
The effect of P availability and parasitism on N concentrations of E. nutans and M. sativa, as well as the P concentration of E. nutans, were analyzed. It was found that no significant single and interactive effects were observed (Table 2; Figure 6A,C). P availability and parasitism had single effects on the N contents of E. nutans and M. sativa, and parasitism (owing to larger ηP2 values) played a more critical role in host N content than P levels (Table 2). The presence of P. kansuensis significantly reduced N contents in E. nutans and M. sativa (Figure 6B). Under low P conditions (0P and 1P), the N content of M. sativa was suppressed more severely than E. nutans (80.35% vs. 50.67% at 0P and 79.42% vs. 36.36% at 1P). However, under 2P, N suppression in M. sativa decreased (68.06%), while it intensified in E. nutans (77.35%). The increase in P availability significantly reduced the N content of E. nutans, particularly when parasitized by P. kansuensis (Figure 6B). On the other hand, the N contents of infected M. sativa and P. kansuensis increased with increasing P availability (Figure 6B).
According to the results of two-way ANOVA, parasitism had a significant effect on the P content of E. nutans (Table 2), while there was no notable decrease in the P content of E. nutans between treatments with or without P. kansuensis under the same P level (0P, 51.35%, p>0.05; 1P, 27.12%, p>0.05; 2P, 83.70%, p>0.05; Figure 6D). In contrast, P availability did not have a noticeable effect on the P content of E. nutans (Table 2), but it significantly decreased with P increment when the grass host was infected by P. kansuensis (Figure 6D). The P contents of parasitized M. sativa and P. kansuensis could not be measured due to insufficient biomass.
3. Discussion
3.1. P Affected Host Selectivity of Root Hemiparasitic Plants Between Grass and Legume
To evaluate host preference of P. kansuensis while controlling “meeting chance” caused by differential root biomass, we compared the haustoria number per gram of dry weight between different host plants. The results showed a distinct foraging preference of P. kansuensis between grass and legume hosts, which was affected by P availability. Under 0P and 1P treatment, although M. sativa (legume) exhibited substantially lower root biomass than E. nutans (dominant grass) (Figure 2 and 4), P. kansuensis developed significantly more haustoria on the legume host (Figure 1B), indicated P. kansuensis preferred to forage legume under low P conditions. Under high P condition (2P), P. kansuensis dropped its preference to the legume host, forming more haustoria on roots of dominant E. nutans (Figure 1B). This study demonstrates that soil P availability influences the host selectivity of the root hemiparasitic plant P. kansuensis in a grass-legume mixture.
The modulation of host selectivity by P in P. kansuensis altered their effects on growth dynamics of grass and legume hosts, impacting their relationship in a grass-legume mixture. When P levels were low, M. sativa was stronger suppressed compared to E. nutans, whereas the opposite pattern was observed under high P conditions (Figure 3). These changes in host suppression suggest that hemiparasitic plants can alter interspecific competition in plant communities by affecting coexisting host species differently[37,38]. The suppression of dominant species like E. nutans by P. kansuensis under high P conditions (Figure 4) supports the hypothesis that parasitic plants reduce the competitive advantage of dominant species, facilitating coexistence and potentially increasing plant community diversity [1,17]. However, the overall productivity of the plant community was reduced in the presence of P. kansuensis, with the extent of suppression increasing at higher P levels (Figure 5). This suggests that the ecological benefits of hemiparasitic plants in promoting species coexistence may come at the cost of reduced ecosystem productivity.
3.2. Potential Mechanisms of P Affect Host Selectivity in Root Hemiparasitic Plants
Several studies have shown that root hemiparasitic plants can distinguish legumes from grasses[8], and they generally exhibit stronger suppression on legume in a multispecies system[9,39]. However, the mechanisms behind this phenomenon still need to be further investigated. Our study confirmed the ability of root hemiparasitic plants to differentiate between legumes and grasses, and also suggested that their host selectivity may vary with changes in P availability.
The P-mediated regulation of host-parasite interactions was initially established in a single host system, where high levels of P inhibited haustorium formation in Rhinanthus minor on Lolium perenne by causing root thickening that mechanically blocked haustoria penetration [40]. In our two-host experiment, despite not quantitatively assessing the root morphological parameters and haustoria structure, both hosts exhibited significantly suppressed growth performance (shoot DW, E. nutans 78.68%, M. sativa 64.41%; root DW, E. nutans 45.54%, M. sativa: 36.65%) and nutrient status (shoot N content, E. nutans 77.35%, M. sativa 68.06%) under high P conditions, indicating their susceptibility to parasitism by P. kansuensis. Therefore, the changes in the capacity of P. kansuensis to develop haustoria on different hosts under various P levels imply that the P-mediated host selection of the root hemiparasite operates through alternative mechanisms rather than purely physical barriers.
Recently, strigolactones (SLs) like orobanchol and 5-deoxystrigol have been found to be act as chemoattractants for host tropism (the initial step of host selectivity) in both the facultative root hemiparasite Phtheirospermum japonicum and the obligate root hemiparasite Striga hermonthica [32]. This indicated that the host preference of root hemiparasitic plants is likely driven by the production of chemical signals derived from the host. In this study, we found that P availability impacted the number of haustoria on legume roots (Figure 1A), indicating that host-derived haustorium-inducing factors, such as flavonoids, phenolics or quinones [32,40], might also play a role in determining host selectivity of parasitic plants. Previous studies have shown that P deficiency can increase the secretion of SLs [31], flavonoids [41,42,43] and phenolic acids [44] in legumes, suggesting that changes in the production of these host-derived chemical signals may drive shifts in host preference with varying P availability. Higher secretion of chemoattractants under low P conditions likely explains the preference of P. kansuensis for M. sativa, while the preference for grasses under high P conditions may be due to a decrease in chemoattractant production by legumes, allowing the dominant grass hosts to be more readily attached by P. kansuensis. Identifying the specific chemical signals and their regulation under different nutrient conditions is crucial for understanding the mechanisms behind host selectivity in root hemiparasitic plants.
3.3. Impact of Soil P Availability on Growth Performance of Hemiparasites
The growth of P. kansuensis was positively influenced by high P availability, as evidenced by increased shoot and root biomass under 2P conditions (Figure 2A,B). These findings are consistent with previous studies showing that nutrient addition enhances hemiparasite growth [24,45], either through improved host resource availability [46] or direct nutrient uptake by hemiparasites [47]. While we did not quantify the nutrient transferred from host to hemiparasite or evaluate the photosynthesis capability of the root hemiparasite in this study, we found that the root: shoot ratio of P. kansuensis was higher in the 2P treatment than in the 1P treatment (Figure 2C), implying that root hemiparasitic plants invest more energy in root growth and improve their nutrient absorption capability in high P conditions.
On the other hand, some studies suggested that fertilization had a negative effect on the growth of root hemiparasitic plants because nutrient addition facilitated the growth of host plants, leading to increased competition from host plants for light [4,48]. For example, Liu et al. [35] found that adding P fertilizer suppressed the above-ground biomass of P. kansuensis in a field study, as the host plants outcompeted the hemiparasite under high nutrient conditions. The discrepancy in the effects of P on the growth of P. kansuensis between our study and Liu’s research may be partly explained by differences in plant growth conditions, as various factors such as nutrient, water, and soil microbes can influence the interaction between host and hemiparasitic plant [21,49].
Interestingly, the growth of P. kansuensis under P-deficient conditions (0P) was not significantly lower than 1P treatment (Figure 2A,B). This resilience can be attributed to its parasitic strategy, which allows it to obtain critical nutrients from host plants. This capacity to thrive in nutrient-poor environments likely explains the prevalence of root hemiparasitic plants in habitats with low soil fertility [47].
3.4. Implications for Management of Parasitic Plants
This study highlights the importance of soil nutrient availability in influencing the dynamics of parasitic plant-host interactions. Manipulating soil nutrient levels, particularly P availability, could be a potential strategy for managing the impacts of parasitic plants like P. kansuensis on agricultural systems and natural plant communities. For example, in systems where legumes are the preferred hosts, increasing P availability could shift parasitism toward grasses, potentially reducing damage to economically important legume crops. Future research should focus on field-based experiments to validate the findings of this study and explore the broader ecological and practical implications of nutrient-mediated host selectivity in root hemiparasitic plants. Additionally, integrating chemical analyses of host root exudates could provide deeper insights into the mechanisms driving host preference and its modulation by soil nutrients.
4. Materials and Methods
4.1. Experimental Design
Elymus nutans (grass) and Medicago sativa (legume) were selected as host species due to their known preference by P. kansuensis (PK) and their common use in alpine grassland restoration efforts in China. A factorial pot experiment was conducted with two levels of parasitism (presence [+PK] or absence [-PK] of P. kansuensis) and three levels of P availability (0 mM/L Na2HPO4 [0P], 1.33 mM/L Na2HPO4 [1P], and 2.66 mM/L Na2HPO4 [2P]), based on the Long Ashton nutrient solution formula [20,50]. Each treatment combination was replicated with 6 pots for +PK and 5 pots for -PK, resulting in a total of 33 pots.
4.2. Plant Materials
Seeds of P. kansuensis and E. nutans were collected from Bayanbulak Grassland (83°42′ E, 42°52′ N, 2472 m) in Xinjiang Uygur Autonomous Region, China, in late August 2016. M. sativa seeds were sourced commercially in 2010. All seeds were stored at 4 ℃ until use. Seeds were surface-sterilized with 4.5% sodium hypochlorite for 5 minutes and rinsed 5 times with distilled water. Seeds were germinated on moistened filter paper in Petri dishes and incubated at 18/25 ℃ (day/night) with a 12 h photoperiod and a light intensity of 22.2 μmol photons m-2s-1.
4.3. Planting and Growth Conditions
Germinated E. nutans and M. sativa seedlings were planted in pairs in the center of each pot, spaced 2 cm apart. After one week, a single germinated P. kansuensis seedling was added to +PK pots, forming an equilateral triangle with the host seedlings. The growth substrate consisted of 10% soil from Kunming Botanical Garden and 90% sand. To eliminate the influence of soil microbes on the host selection of root hemiparasitic plants, we utilized autoclaved substrate (121℃ for 2 h).
Pots were maintained in a greenhouse at Kunming Institute of Botany, Chinese Academy of Sciences, under natural light conditions with temperatures ranging from 15 to 35 °C. The experiment ran from 23 Jun. to 30 Oct. 2017, 129 days. P fertilizer solutions were applied weekly (20 mL per pot) two weeks after planting, corresponding to the specified P levels for each treatment. Pots were randomized weekly to minimize positional effects.
4.4. Harvest and Sampling
Only pots where all plants survived until the end of the experiment were included in the analysis (three pots under 0P conditions with PK, five pots under 0P and 1P conditions without PK, four pots under other conditions). At harvest, shoots and roots were separated. The roots of each plant species were identified morphologically and separated before being stored in 50% alcohol for later measurement. Three replicates for each +PK treatment were randomly selected to analyze the number of haustoria (HN) produced by P. kansuensis. Haustoria were categorized into two types: (1) presumably functional haustoria (PFH) that attaches to host roots with distinct xylem bridges [51], and (2) non-functional haustoria (NFH) without distinct xylem bridges. The number of PFH attached to each host was counted separately. The total number of haustoria produced by P. kansuensis was a sum of the number of PFH attached to each host and the number of NFH. The incidence of haustorium formation was recorded as the number of haustoria per gram of dry root samples [52]. Plant shoot and root dry weights (DWs) were determined after oven drying at 75 ℃ for 48 h.
4.5. Measurement of Shoot N and P Concentrations
For nutrient analysis, shoot N and P concentrations were measured from the three replicates per treatment that were used for examining haustoria numbers. N concentration was determined using an elemental analyzer (Vario Macro Cube, Elementar, Germany), while P concentration was measured with the phosphovanado-molybdate method on a spectrophotometer (L6S, Shanghai Yidian Analytical Instrument Co., LTD, Shanghai, China). Due to limited biomass in certain pots, shoot P concentration for M. sativa and P. kansuensis was not analyzed. Shoot N or P contents were calculated by multiplying the element concentration and shoot DW.
4.6. Statistical Analyses
Two-way ANOVA was used to evaluate the effects of P availability and parasitism on shoot dry weight (DW), root DW, shoot N and P status (concentration and content) of E. nutans and M. sativa, as well as total shoot biomass per pot. One-way ANOVA followed by the Tukey HSD test assessed the influence of P availability on growth performance, nutrient status, and haustorium formation. Independent sample t-tests were used to analyze the effects of parasitism on host growth and nutrient status, as well as differences in haustorium formation by P. kansuensis between host species. Before analysis, data were assessed for normality and homogeneity. For specific datasets, data were transformed by log10 to meet assumptions of normality and homogeneity of variances. Mann-Whitney U tests were employed for datasets with non-normal distributions. Statistical analyses and visualization were conducted in R (version 4.4.2) and R-Studio using packages dplyr [53], emmeans [54], lme4 [55], car [56], ggpubr [57] and ggplot2 [58].
5. Conclusions
This study demonstrates that host selectivity of the root hemiparasitic plant P. kansuensis is influenced by soil P availability. Under low P conditions, P. kansuensis preferentially parasitized the legume host M. sativa, forming more haustoria and causing significant biomass reduction in the legume host despite much higher grass biomass in the host mixture. In contrast, under higher P levels P. kansuensis shifted parasitism toward the grass host E. nutans, leading to more severe suppression of grass growth. These findings emphasize the significance of soil nutrient status in shaping the interactions between parasitic plants and their hosts. They also have practical implications for mitigating the damage caused by parasitic weeds in agricultural and natural ecosystems via adjusting nutrient supply. Future research should explore these interactions in field settings and investigate the chemical and physiological mechanisms underlying host preference. Such studies will deepen our understanding of parasitic plant ecology and help develop strategies for managing their impact on plant communities.
Author Contributions
Conceptualization, X.L.S and A.R.L.; methodology, X.L.S.; software, X.L.S.; validation, X.L.S. and A.R.L.; formal analysis, X.L.S.; investigation, X.L.S. and R.J.X.; writing—original draft preparation, X.L.S.; writing—review and editing, X.L.S, R.J.X. and A.R.L.; visualization, X.L.S.; supervision, A.R.L.; funding acquisition, X.L.S. and A.R.L. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by Yunnan Ten Thousand Talents Program Plan Young & Elite Talents Project [YNWR-QNBJ-2020-290 for Xiaolin Sui] and National Natural Science Foundation of China [No. 31872686], as well as partly supported by Yunnan Fundamental Research Project [No. 202401AT070237 and Grant No. 202201AS070046], and Yunnan Revitalization Talent Support Program “Innovation Team” Project [202505AS350017].
Data Availability Statement
The original contributions presented in the study are included in the article, further inquiries can be directed to the first author.
Acknowledgments
We are grateful to Dr. Ning Yan and her team for their help with greenhouse management, as well as to Miss. Qiuping Chen for her advice on utilizing R software for data analysis.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Těšitel, J. Functional biology of parasitic plants: a review. Plant Ecol. Evol. 2016, 149, 5-20. [CrossRef]
- Westwood, J.H.; Yoder, J.I.; Timko, M.P.; dePamphilis, C.W. The evolution of parasitism in plants. Trends Plant Sci. 2010, 15, 227-235. [CrossRef]
- Irving, L.J.; Cameron, D.D. You are what you eat: interactions between root parasitic plants and their hosts. Adv. Bot. Res. 2009, 50, 87-138. [CrossRef]
- Matthies, D. Parasitic and competitive interactions between the hemiparasites Rhinanthus serotinus and Odontites rubra and their host Medicago sativa. J. Ecol. 1995, 83, 245-251.
- Press, M.C.; Phoenix, G.K. Impacts of parasitic plants on natural communities. New Phytol. 2005, 166, 737-751. [CrossRef]
- Fibich, P.; Leps, J.; Chytry, M.; Tesitel, J. Root hemiparasitic plants are associated with high diversity in temperate grasslands. J. Veg. Sci. 2017, 28, 184-191. [CrossRef]
- Těšitel, J.; Li, A.-R.; Knotkova, K.; McLellan, R.; Bandaranayake, P.C.G.; Watson, D.M. The bright side of parasitic plants: what are they good for? Plant Physiol. 2021, 185, 1309-1324. [CrossRef]
- Sandner, T.M.; Schoppan, L.; Matthies, D. Seedlings of a hemiparasite recognize legumes, but do not distinguish good from poor host species. Folia Geobot. 2022, 57, 117-126. [CrossRef]
- Matthies, D. Interactions between the root hemiparasite Melampyrum arvense and mixtures of host plants: Heterotrophic benefit and parasite-mediated competition. Oikos 1996, 75, 118-124. [CrossRef]
- Cechin, I.; Press, M.C. Nitrogen relations of the sorghum-Striga hermonthica host-parasite association: germination, attachment and early growth. New Phytol. 1993, 124, 681-687. [CrossRef]
- Rowntree, J.K.; Craig, H. The contrasting roles of host species diversity and parasite population genetic diversity in the infection dynamics of a keystone parasitic plant. J. Ecol. 2019, 107, 23-33. [CrossRef]
- Westbury, D.B.; Dunnett, N.P. The promotion of grassland forb abundance: A chemical or biological solution? Basic Appl. Ecol. 2008, 9, 653-662. [CrossRef]
- Ameloot, E.; Verheyen, K.; Hermy, M. Meta-analysis of standing crop reduction by Rhinanthus spp. and its effect on vegetation structure. Folia Geobot. 2005, 40, 289-310. [CrossRef]
- Fisher, J.P.; Phoenix, G.K.; Childs, D.Z.; Press, M.C.; Smith, S.W.; Pilkington, M.G.; Cameron, D.D. Parasitic plant litter input: a novel indirect mechanism influencing plant community structure. New Phytol. 2013, 198, 222-231. [CrossRef]
- Ameloot, E.; Verlinden, G.; Boeckx, P.; Verheyen, K.; Hermy, M. Impact of hemiparasitic Rhinanthus angustifolius and R.minor on nitrogen availability in grasslands. Plant Soil 2008, 311, 255-268. [CrossRef]
- Matthies, D. Interactions between a root hemiparasite and 27 different hosts: growth, biomass allocation and plant architecture. Perspect. Plant Ecol. Evol. Syst. 2017, 24, 118-137. [CrossRef]
- Chaudron, C.; Mazalova, M.; Kuras, T.; Malenovsky, I.; Mladek, J. Introducing ecosystem engineers for grassland biodiversity conservation: A review of the effects of hemiparasitic Rhinanthus species on plant and animal communities at multiple trophic levels. Perspect. Plant Ecol. Evol. Syst. 2021, 52. [CrossRef]
- Irving, L.J.; Kim, D.; Schwier, N.; Vaughan, J.K.E.; Ong, G.; Hama, T. Host nutrient supply affects the interaction between the hemiparasite Phtheirospermum japonicum and its host Medicago sativa. Environ. Exp. Bot. 2019, 162, 125-132. [CrossRef]
- Jiang, F.; Jeschke, W.D.; Hartung, W.; Cameron, D.D. Interactions between Rhinanthus minor and its hosts: a review of water, mineral nutrient and hormone flows and exchanges in the hemiparasitic association. Folia Geobot. 2010, 45, 369-385. [CrossRef]
- Li, A.R.; Li, Y.J.; Smith, S.E.; Smith, F.A.; Guan, K.Y. Nutrient requirements differ in two Pedicularis species in the absence of a host plant: implication for driving forces in the evolution of host preference of root hemiparasitic plants. Ann. Bot. 2013, 112, 1099-1106. [CrossRef]
- Těšitel, J.; Těšitelová, T.; Fisher, J.P.; Lepš, J.; Cameron, D.D. Integrating ecology and physiology of root-hemiparasitic interaction: interactive effects of abiotic resources shape the interplay between parasitism and autotrophy. New Phytol. 2015, 205, 350-360. [CrossRef]
- Cirocco, R.M.; Facelli, E.; Delean, S.; Facelli, J.M. Does phosphorus influence performance of a native hemiparasite and its impact on a native legume? Physiol. Plantarum 2021. [CrossRef]
- Cirocco, R.M.; Watling, J.R.; Facelli, J.M. The combined effects of water and nitrogen on the relationship between a native hemiparasite and its invasive host. New Phytol. 2021, 229, 1728-1739. [CrossRef]
- Korell, L.; Sandner, T.M.; Matthies, D.; Ludewig, K. Effects of drought and N level on the interactions of the root hemiparasite Rhinanthus alectorolophus with a combination of three host species. Plant Biol. 2020, 22, 84-92. [CrossRef]
- Jiang, F.; Jeschke, W.D.; Hartung, W. Solute flows from Hordeum vulgare to the hemiparasite Rhinanthus minor and the influence of infection on host and parasite nutrient relations. Funct. Plant Biol. 2004, 31, 633-643. [CrossRef]
- Seel, W.E.; Parsons, A.N.; Press, M.C. Do inorganic solutes limit growth of the facultative hemiparasite Rhinanthus minor L in the absence of a host. New Phytol. 1993, 124, 283-289. [CrossRef]
- He, Y.; Bi, Y.; Yu, H.; Zhang, Y.; Struik, P.C.; Jing, J. Positive legacy effects of grass-legume mixture leys on phosphorus uptake and yield of maize weaken over the growing season. Field Crop Res. 2024, 314. [CrossRef]
- Zhu, S.G.; Tao, H.Y.; Li, W.B.; Zhou, R.; Gui, Y.W.; Zhu, L.; Zhang, X.L.; Wang, W.; Wang, B.Z.; Mei, F.J.; et al. Phosphorus availability mediates plant-plant interaction and field productivity in maize-grass pea intercropping system: field experiment and its global validation. Agric. Syst. 2023, 205. [CrossRef]
- Tawaraya, K.; Horie, R.; Shinano, T.; Wagatsuma, T.; Saito, K.; Oikawa, A. Metabolite profiling of soybean root exudates under phosphorus deficiency. Soil Sci. Plant Nutr. 2014, 60, 679-694. [CrossRef]
- Xiong, C.; Li, X.; Wang, X.; Wang, J.; Lambers, H.; Vance, C.P.; Shen, J.; Cheng, L. Flavonoids are involved in phosphorus-deficiency-induced cluster-root formation in white lupin. Ann. Bot. 2022, 129, 101-112. [CrossRef]
- Yoneyama, K.; Yoneyama, K.; Takeuchi, Y.; Sekimoto, H. Phosphorus deficiency in red clover promotes exudation of orobanchol, the signal for mycorrhizal symbionts and germination stimulant for root parasites. Planta 2007, 225, 1031-1038. [CrossRef]
- Ogawa, S.; Cui, S.; White, A.R.F.; Nelson, D.C.; Yoshida, S.; Shirasu, K. Strigolactones are chemoattractants for host tropism in Orobanchaceae parasitic plants. Nat. Commun. 2022, 13. [CrossRef]
- Sui, X.L.; Kuss, P.; Li, W.J.; Yang, M.Q.; Guan, K.Y.; Li, A.R. Identity and distribution of weedy Pedicularis kansuensis Maxim. (Orobanchaceae) in Tianshan Mountains of Xinjiang: morphological, anatomical and molecular evidence. J. Arid Land 2016, 8, 453-461. [CrossRef]
- Li, W.J.; Sui, X.L.; Kuss, P.; Liu, Y.Y.; Li, A.R.; Guan, K.Y. Long-Distance Dispersal after the Last Glacial Maximum (LGM) led to the disjunctive distribution of Pedicularis kansuensis (Orobanchaceae) between the Qinghai-Tibetan Plateau and Tianshan Region. Plos One 2016, 11, e0165700. [CrossRef]
- Liu, Y.Y.; Taxipulati, T.; Gong, Y.M.; Sui, X.L.; Wang, X.Z.; Parent, S.E.; Hu, Y.K.; Guan, K.Y.; Li, A.R. N-P fertilization inhibits growth of root hemiparasite Pedicularis kansuensis in natural grassland. Front. Plant Sci. 2017, 8, 2088. [CrossRef]
- Bao, G.S.; Suetsugu, K.; Wang, H.S.; Yao, X.; Liu, L.; Ou, J.; Li, C.J. Effects of the hemiparasitic plant Pedicularis kansuensis on plant community structure in a degraded grassland. Ecol. Res. 2015, 30, 507-515. [CrossRef]
- Tesitel, J.; Mladek, J.; Hornik, J.; Tesitelova, T.; Adamec, V.; Tichy, L. Suppressing competitive dominants and community restoration with native parasitic plants using the hemiparasitic Rhinanthus alectorolophus and the dominant grass Calamagrostis epigejos. J. Appl. Ecol. 2017, 54, 1487-1495. [CrossRef]
- Těšitel, J.; Mladek, J.; Fajmon, K.; Blazek, P.; Mudrak, O. Reversing expansion of Calamagrostis epigejos in a grassland biodiversity hotspot: hemiparasitic Rhinanthus major does a better job than increased mowing intensity. Appl. Veg. Sci. 2018, 21, 104-112. [CrossRef]
- Sandner, T.M.; Matthies, D. Multiple choice: hemiparasite performance in multi-species mixtures. Oikos 2018, 127, 1291-1303. [CrossRef]
- Goyet, V.; Wada, S.; Cui, S.; Wakatake, T.; Shirasu, K.; Montiel, G.; Simier, P.; Yoshida, S. Haustorium inducing factors for parasitic Orobanchaceae. Front. plant sci. 2019, 10. [CrossRef]
- Luo, J.J.; Liu, Y.X.; Zhang, H.K.; Wang, J.P.; Chen, Z.J.; Luo, L.J.; Liu, G.D.; Liu, P.D. Metabolic alterations provide insights into Stylosanthes roots responding to phosphorus deficiency. Bmc. Plant Biol. 2020, 20. [CrossRef]
- Malusà, E.; Russo, M.; Mozzetti, C.; Belligno, A. Modification of secondary metabolism and flavonoid biosynthesis under phosphate deficiency in bean roots. J. Plant Nutr. 2006, 29, 245-258. [CrossRef]
- Mo, X.H.; Zhang, M.K.; Liang, C.Y.; Cai, L.Y.; Tian, J. Integration of metabolome and transcriptome analyses highlights soybean roots responding to phosphorus deficiency by modulating phosphorylated metabolite processes. Plant Physiol. Bioch. 2019, 139, 697-706. [CrossRef]
- Dakora, F.D.; Phillips, D.A. Root exudates as mediators of mineral acquisition in low-nutrient environments. Plant Soil 2002, 245, 35-47. [CrossRef]
- Borowicz, V.A.; Armstrong, J.E. Resource limitation and the role of a hemiparasite on a restored prairie. Oecologia 2012, 169, 783-792. [CrossRef]
- Hautier, Y.; Hector, A.; Vojtech, E.; Purves, D.; Turnbull, L.A. Modelling the growth of parasitic plants. J. Ecol. 2010, 98, 857-866. [CrossRef]
- Těšitel, J.; Fibich, P.; de Bello, F.; Chytry, M.; Leps, J. Habitats and ecological niches of root-hemiparasitic plants: an assessment based on a large database of vegetation plots. Preslia 2015, 87, 87-108.
- Fibich, P.; Leps, J.; Berec, L. Modelling the population dynamics of root hemiparasitic plants along a productivity gradient. Folia Geobot. 2010, 45, 425-442. [CrossRef]
- Sui, X.L.; Zhang, T.; Tian, Y.T.; Xue, R.J.; Li, A.R. A neglected alliance in battles against parasitic plants: arbuscular mycorrhizal and rhizobial symbioses alleviate damage to a legume host by root hemiparasitic Pedicularis species. New Phytol. 2019, 221, 470-481. [CrossRef]
- Hewitt, E.J. Sand and water culture methods used in the study of plant nutrition; Commonwealth Agricultural Bureaux: Farnham Royal, UK, 1966; pp. 547.
- Li, A.R.; Guan, K.Y. Arbuscular mycorrhizal fungi may serve as another nutrient strategy for some hemiparasitic species of Pedicularis (Orobanchaceae). Mycorrhiza 2008, 18, 429-436. [CrossRef]
- Li, A.R.; Smith, S.E.; Smith, F.A.; Guan, K.Y. Inoculation with arbuscular mycorrhizal fungi suppresses initiation of haustoria in the root hemiparasite Pedicularis tricolor. Ann. Bot. 2012, 109, 1075-1080. [CrossRef]
- Wickham H., François R., Henry L., Müller K., Vaughan D. dplyr: A grammar of data manipulation. R package version 1.1.4 2023. https://CRAN.R-project.org/package=dplyr. (accessed on 31 Oct. 2024).
- Lenth R. emmeans: estimated marginal means, aka least-squares means. R package version 1.10.6 2024. https://CRAN.R-project.org/package=emmeans. (accessed on 12 Dec. 2024).
- Bates D., Maechler M., Bolker B., Walker S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67: 1-48. [CrossRef]
- Fox J., Weisberg S. An R companion to applied regression, 3rd ed.; Sage: Thousand Oaks, CA. 2019.
- Kassambara A. gpubr: ‘ggplot2’ based publication ready plots. R package version 0.6.0 2023. https://CRAN.R-project.org/package=ggpubr.(accessed on 31 Oct. 2024).
- Wickham H. ggplot2: elegant graphics for data analysis. New York: Springer-Verlag, USA, 2016.
Figure 1.
Haustoria formation capability of Pedicularis kansuensis under various P levels. (A) Number of presumably functional haustoria (PFH) produced on EN and MS; (B) number of PFH per gram dry root of EN and MS; (C) total number of haustoria (H) formed by PK; and (D) total number of H per gram dry root of PK. Data are presented as mean ± 1SE. Different letters indicate statistically significant differences among P availability at p<0.05 level. Asterisks denote significant differences between EN and MS under the same P level (*p<0.05). Species: Elymus nutans (EN), Medicago sativa (MS), P. kansuensis (PK). P treatments: 0P, 0 mM/L Na2HPO4; 1P, 1.33 mM/L Na2HPO4; 2P, 2.66 mM/L Na2HPO4.
Figure 1.
Haustoria formation capability of Pedicularis kansuensis under various P levels. (A) Number of presumably functional haustoria (PFH) produced on EN and MS; (B) number of PFH per gram dry root of EN and MS; (C) total number of haustoria (H) formed by PK; and (D) total number of H per gram dry root of PK. Data are presented as mean ± 1SE. Different letters indicate statistically significant differences among P availability at p<0.05 level. Asterisks denote significant differences between EN and MS under the same P level (*p<0.05). Species: Elymus nutans (EN), Medicago sativa (MS), P. kansuensis (PK). P treatments: 0P, 0 mM/L Na2HPO4; 1P, 1.33 mM/L Na2HPO4; 2P, 2.66 mM/L Na2HPO4.
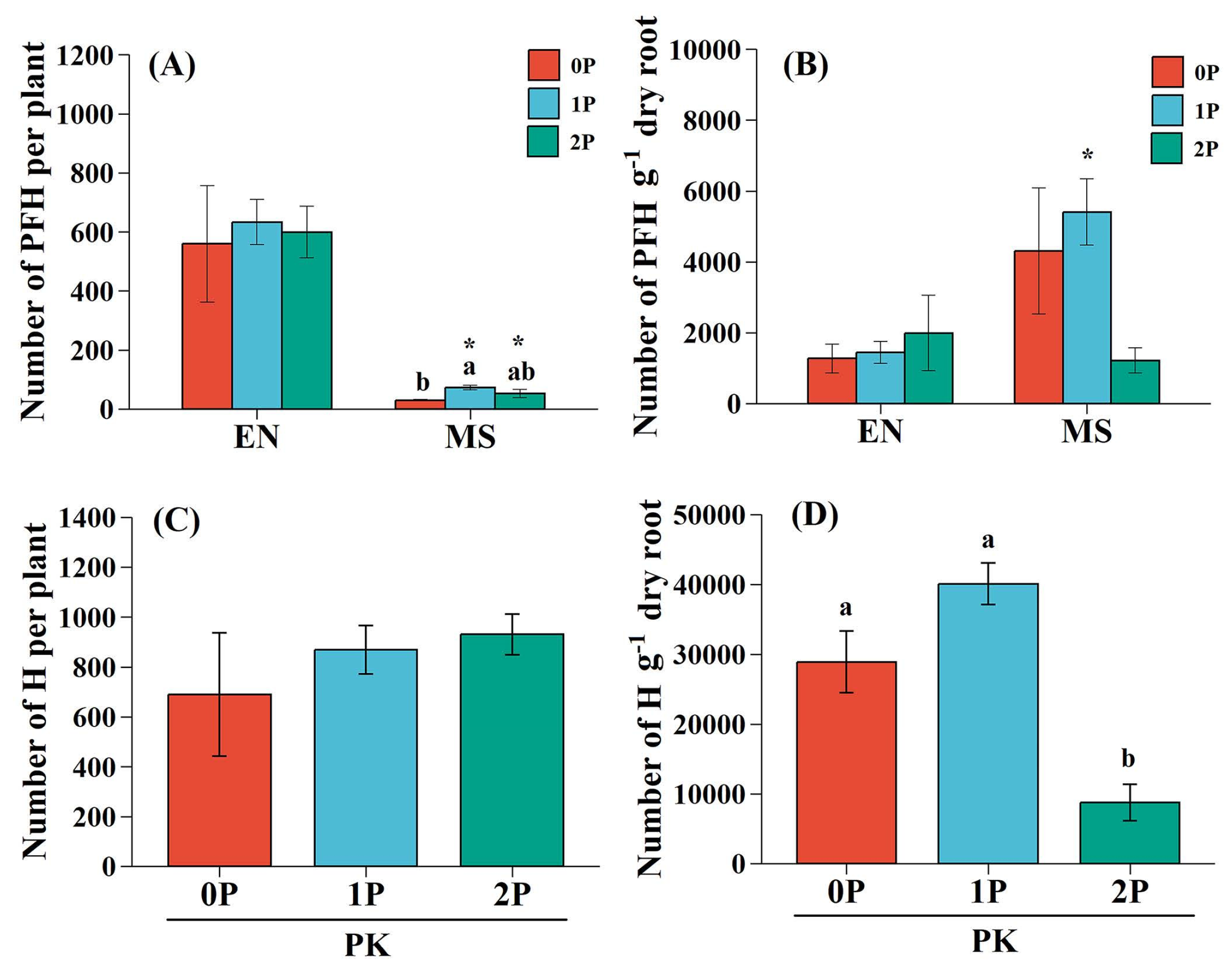
Figure 2.
Growth performance and biomass allocation of host plants and hemiparasite under different P and parasitism conditions. (A) Shoot dry weight, (B) root dry weight, and (C) root: shoot ratio. Data are mean ±1 SE. Different letters indicate significant effects of P availability on infected or uninfected host plants and hemiparasite (p<0.05). Asterisks denote significant differences between treatments with (+PK) or without PK (-PK) under the same P level (*p<0.05; **p<0.01). Species: Elymus nutans (EN), Medicago sativa (MS), P. kansuensis (PK). P treatments: 0P, 0 mM/L Na2HPO4; 1P, 1.33 mM/L Na2HPO4; 2P, 2.66 mM/L Na2HPO4.
Figure 2.
Growth performance and biomass allocation of host plants and hemiparasite under different P and parasitism conditions. (A) Shoot dry weight, (B) root dry weight, and (C) root: shoot ratio. Data are mean ±1 SE. Different letters indicate significant effects of P availability on infected or uninfected host plants and hemiparasite (p<0.05). Asterisks denote significant differences between treatments with (+PK) or without PK (-PK) under the same P level (*p<0.05; **p<0.01). Species: Elymus nutans (EN), Medicago sativa (MS), P. kansuensis (PK). P treatments: 0P, 0 mM/L Na2HPO4; 1P, 1.33 mM/L Na2HPO4; 2P, 2.66 mM/L Na2HPO4.
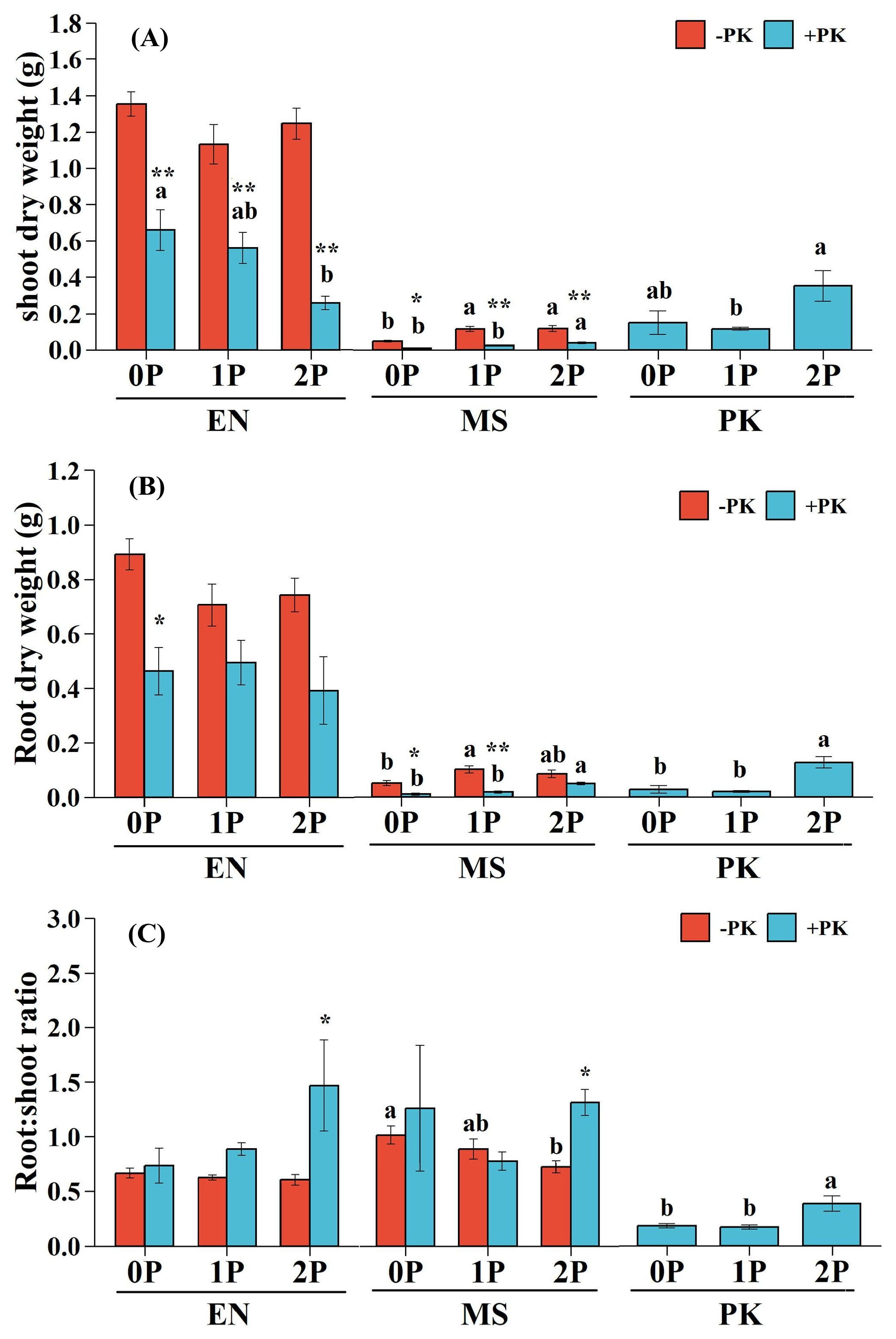
Figure 3.
The degree of suppression of host growth by Pedicularis kansuensis under different P levels. (A) The suppression of shoot dry weight (DW); (B) the suppression of root DW. Data are mean ±1 SE. Asterisks denote significant differences between species under the same P level (*p<0.05; **p<0.01; ***p<0.001). Species: Elymus nutans (EN), Medicago sativa (MS), P. kansuensis (PK). P treatments: 0P, 0 mM/L Na2HPO4; 1P, 1.33 mM/L Na2HPO4; 2P, 2.66 mM/L Na2HPO4.
Figure 3.
The degree of suppression of host growth by Pedicularis kansuensis under different P levels. (A) The suppression of shoot dry weight (DW); (B) the suppression of root DW. Data are mean ±1 SE. Asterisks denote significant differences between species under the same P level (*p<0.05; **p<0.01; ***p<0.001). Species: Elymus nutans (EN), Medicago sativa (MS), P. kansuensis (PK). P treatments: 0P, 0 mM/L Na2HPO4; 1P, 1.33 mM/L Na2HPO4; 2P, 2.66 mM/L Na2HPO4.
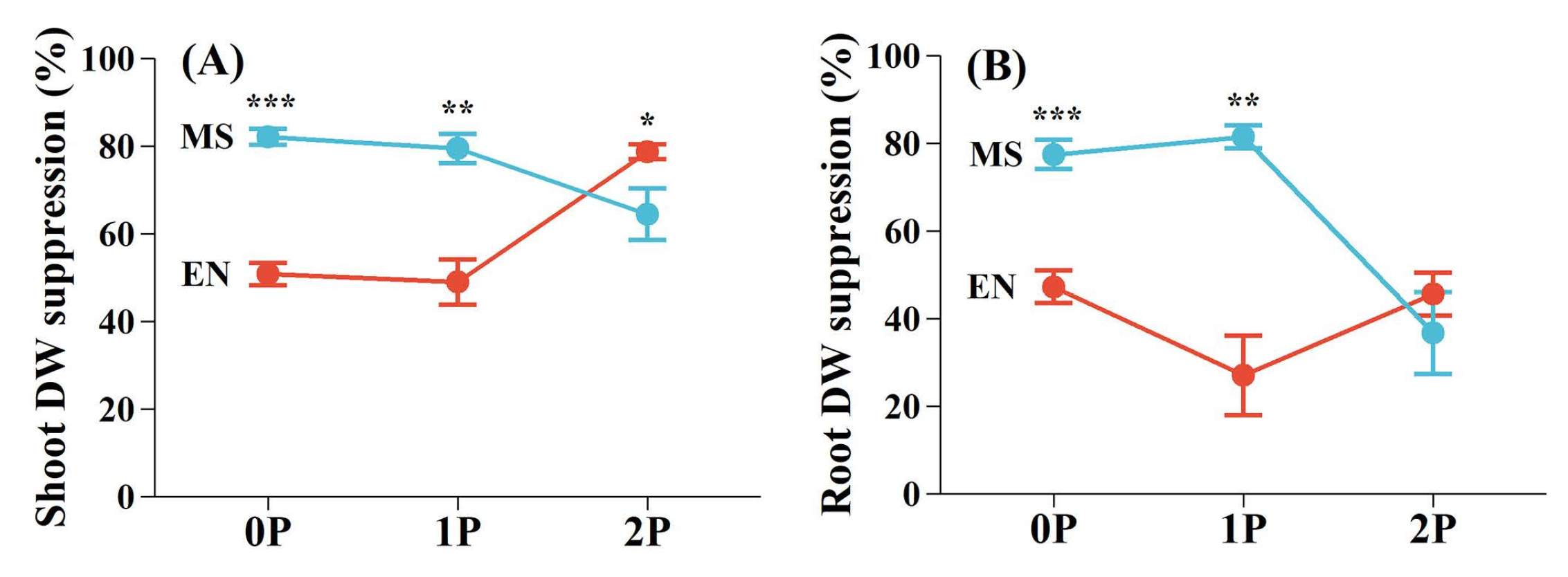
Figure 4.
The shoot and root dry weight (DW) proportion that hosts (A&C) and hemiparasite (B&D) per pot under different P conditions. Data are mean ±1 SE. Asterisks denote significant effect of parasitism on different host plants (*p<0.05; **p<0.01). Species: Elymus nutans (EN), Medicago sativa (MS), and Pedicularis kansuensis (PK). EN-PK, uninfected EN; EN+PK, infected EN; MS-PK, uninfected MS; MS+PK, infected MS. P treatment: 0P, 0 mM/L Na2HPO4; 1P, 1.33 mM/L Na2HPO4; 2P, 2.66 mM/L Na2HPO4.
Figure 4.
The shoot and root dry weight (DW) proportion that hosts (A&C) and hemiparasite (B&D) per pot under different P conditions. Data are mean ±1 SE. Asterisks denote significant effect of parasitism on different host plants (*p<0.05; **p<0.01). Species: Elymus nutans (EN), Medicago sativa (MS), and Pedicularis kansuensis (PK). EN-PK, uninfected EN; EN+PK, infected EN; MS-PK, uninfected MS; MS+PK, infected MS. P treatment: 0P, 0 mM/L Na2HPO4; 1P, 1.33 mM/L Na2HPO4; 2P, 2.66 mM/L Na2HPO4.
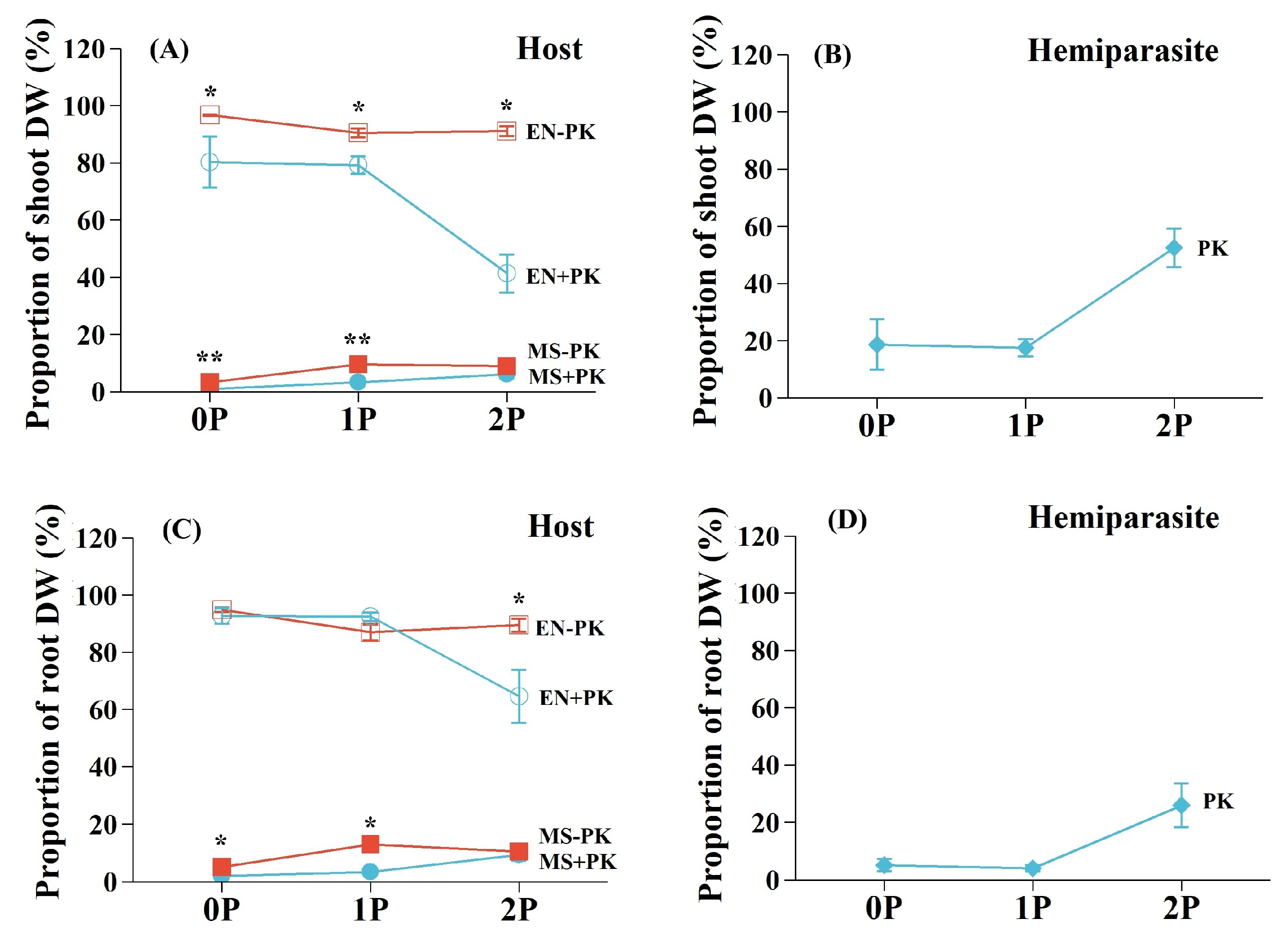
Figure 5.
Total shoot biomass in each pot with or without Pedicularis kansuensis under different P levels. Data are presented as means of replicate pots under different P and parasitism conditions, with stacked bars showing mean vales of Elymus nutans (EN), Medicago sativa (MS), and P. kansuensis (PK) separately. In addition, the percentage of total shoot biomass per pot (including PK, outside bracket) and the percentage of total shoot biomass of both host plants suppressed by PK (inside bracket) are displayed for each P treatment. Parasitism: -PK, without PK; +PK, with PK. P treatments: 0P, 0 mM/L Na2HPO4; 1P, 1.33 mM/L Na2HPO4; 2P, 2.66 mM/L Na2HPO4.
Figure 5.
Total shoot biomass in each pot with or without Pedicularis kansuensis under different P levels. Data are presented as means of replicate pots under different P and parasitism conditions, with stacked bars showing mean vales of Elymus nutans (EN), Medicago sativa (MS), and P. kansuensis (PK) separately. In addition, the percentage of total shoot biomass per pot (including PK, outside bracket) and the percentage of total shoot biomass of both host plants suppressed by PK (inside bracket) are displayed for each P treatment. Parasitism: -PK, without PK; +PK, with PK. P treatments: 0P, 0 mM/L Na2HPO4; 1P, 1.33 mM/L Na2HPO4; 2P, 2.66 mM/L Na2HPO4.
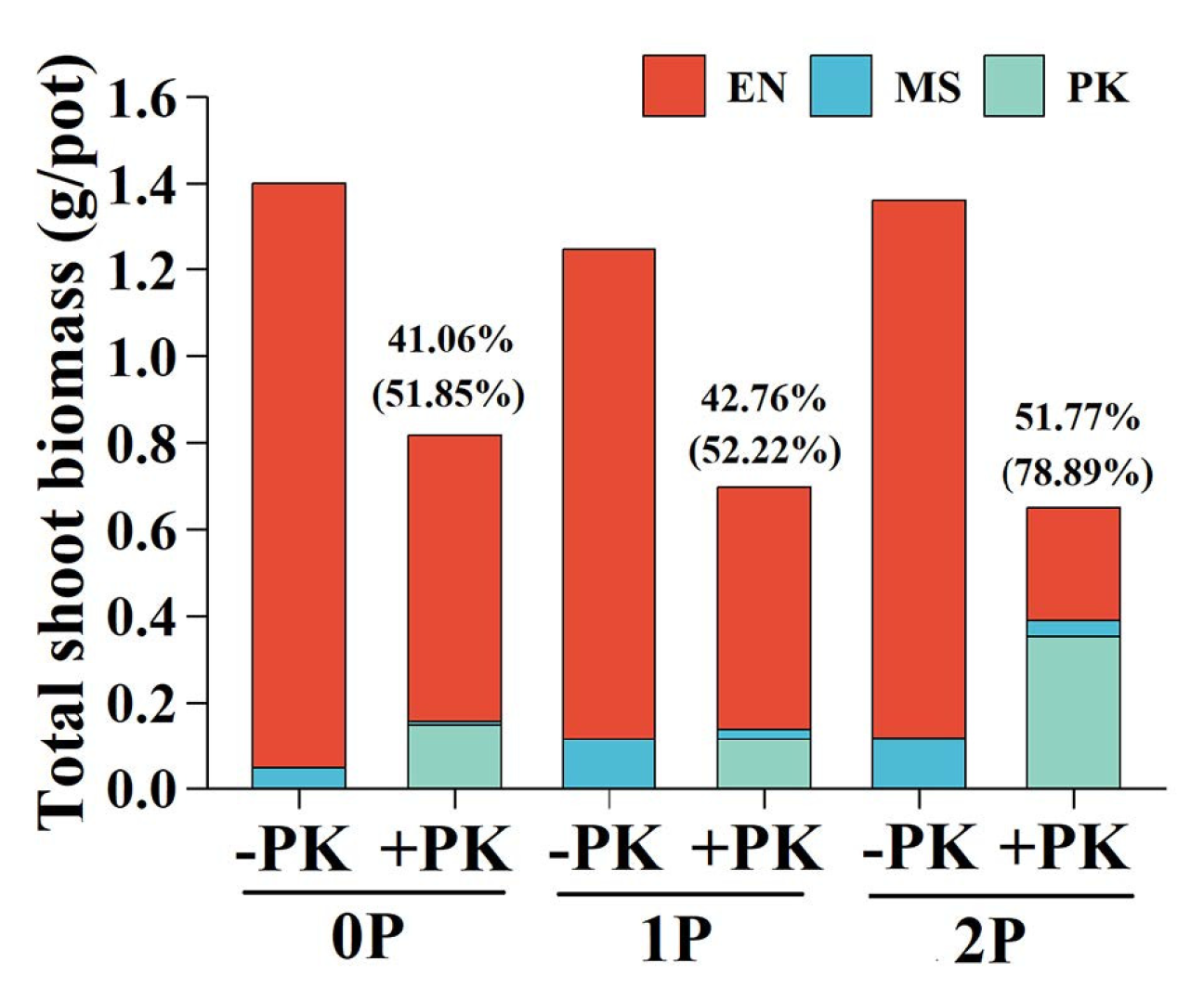
Figure 6.
Shoot N and P status of hosts and hemiparasite in pots with or without Pedicularis kansuensis under different P conditions. (A) Shoot N concentration; (B) shoot N concent; (C) shoot P concentration; (D) shoot P content. Data are mean ±1 SE. Different letters indicate significant effects of P availability on infected or uninfected host plants and hemiparasite (p<0.05). Asterisks denote significant differences between treatments with (+PK) or without PK (-PK) under the same P level (*p<0.05; **p<0.01). Species: Elymus nutans (EN), Medicago sativa (MS), P. kansuensis (PK). P treatments: 0P, 0 mM/L Na2HPO4; 1P, 1.33 mM/L Na2HPO4; 2P, 2.66 mM/L Na2HPO4.
Figure 6.
Shoot N and P status of hosts and hemiparasite in pots with or without Pedicularis kansuensis under different P conditions. (A) Shoot N concentration; (B) shoot N concent; (C) shoot P concentration; (D) shoot P content. Data are mean ±1 SE. Different letters indicate significant effects of P availability on infected or uninfected host plants and hemiparasite (p<0.05). Asterisks denote significant differences between treatments with (+PK) or without PK (-PK) under the same P level (*p<0.05; **p<0.01). Species: Elymus nutans (EN), Medicago sativa (MS), P. kansuensis (PK). P treatments: 0P, 0 mM/L Na2HPO4; 1P, 1.33 mM/L Na2HPO4; 2P, 2.66 mM/L Na2HPO4.
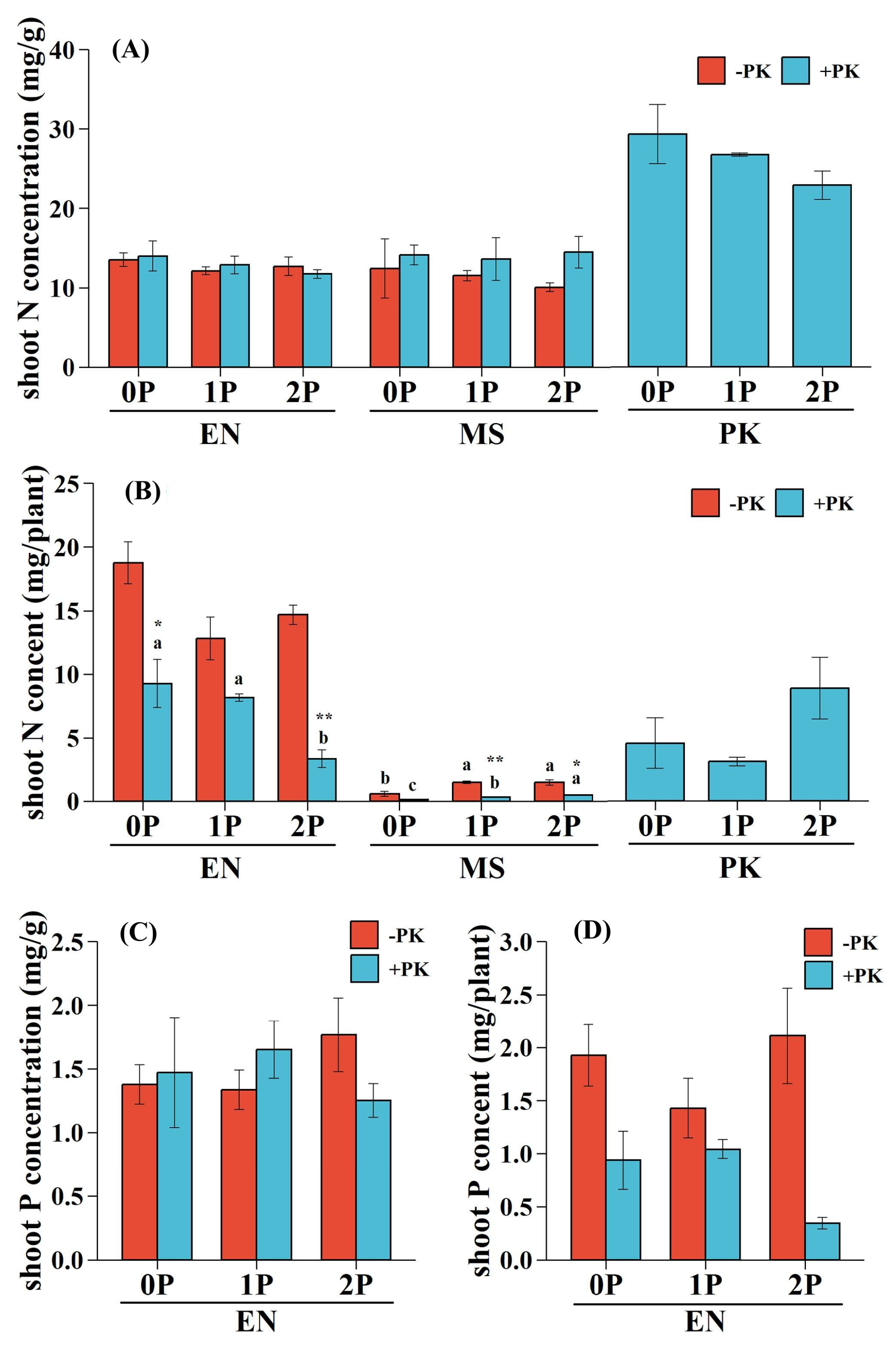

Table 1.
Two-way ANOVA results (F and ηP2 values) for the effects of phosphorus availability (P) and parasitism (Pa) on shoot and root dry weight of Elymus nutans (EN), Medicago sativa (MS) and shoot biomass per pot (Total, including PK).
Table 1.
Two-way ANOVA results (F and ηP2 values) for the effects of phosphorus availability (P) and parasitism (Pa) on shoot and root dry weight of Elymus nutans (EN), Medicago sativa (MS) and shoot biomass per pot (Total, including PK).
| Shoot dry weight | Root dry weight | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EN | MS† | Total | EN | MS† | Total | ||||||||
| df | F | ηP2 | F | ηP2 | F | ηP2 | F | ηP2 | F | ηP2 | F | ηP2 | |
| P | 2,19 | 6.81** | 0.42 | 32.02*** | 0.77 | 3.20 | 0.25 | 1.91 | 0.17 | 12.49*** | 0.77 | 0.95 | 0.09 |
| Pa | 1, 19 | 121.47*** | 0.86 | 176.11*** | 0.90 | 83.74*** | 0.82 | 24.06*** | 0.56 | 65.27*** | 0.90 | 25.40*** | 0.57 |
| P*Pa | 2,19 | 3.32 | 0.26 | 3.16 | 0.25 | 0.57 | 0.06 | 0.87 | 0.08 | 6.80** | 0.25 | 0.81 | 0.08 |
Values suggesting significant effects (p<0.05) were given in bold (**p<0.01, ***p<0.001) † indicated data were log10 transformed before analyses.
Table 2.
Two-way ANOVA results (F and ηP2 values) for the effects of phosphorus availability (P) and parasitism (Pa) on shoot N and P status of Elymus nutans (EN) and Medicago sativa (MS).
Table 2.
Two-way ANOVA results (F and ηP2 values) for the effects of phosphorus availability (P) and parasitism (Pa) on shoot N and P status of Elymus nutans (EN) and Medicago sativa (MS).
| N concentration | P concentration | N content | P content | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EN | MS | EN | MS | EN | MS† | EN | MS | ||||||||
| df | F | ηP2 | F | ηP2 | F | ηP2 | F | ηP2 | F | ηP2 | F | ηP2 | |||
| P | 2, 12 | 1.091 | 0.15 | 0.117 | 0.02 | 0.063 | 0.01 | -- | 7.717** | 0.56 | 30.153*** | 0.83 | 0.358 | 0.06 | -- |
| Pa | 1, 12 | 0.010 | 0.00 | 2.484 | 0.17 | 0.028 | 0.00 | -- | 63.166** | 0.84 | 101.813*** | 0.89 | 21.840** | 0.65 | -- |
| P*Pa | 2, 12 | 0.334 | 0.05 | 0.239 | 0.04 | 1.440 | 0.19 | -- | 3.479 | 0.37 | 1.057 | 0.15 | 3.169 | 0.35 | -- |
Values suggesting significant effects were given in bold (**p<0.01; ***p<0.001). --indicated data were unanalyzed because samples in some pots were insufficient for P measurement. † indicated data were log 10 transformed before analyses.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



