
Submitted:
01 June 2025
Posted:
04 June 2025
You are already at the latest version
Abstract
For their energy needs, cancer cells require large amounts of glucose, since they use the inefficient glycolytic pathway (Warburg effect), producing only two moles of lactate and two moles of adenosine triphosphate for each mole of glucose consumed. By contrast, normal cells consume glucose via glycolysis and utilize its end-product, lactate, as the substrate of the oxidative mitochondrial tricarboxylic acid cycle and it’s coupled oxidative phosphorylation. This process is 18 times more efficient in producing adenosine triphosphate than glycolysis alone. It is hypothesized here that lactate, produced by cancer cells, is preferentially used oxidatively by normal cells surrounding those cancer cells, bypassing glycolysis and sparing glucose, which is readily available for the glucose-craving cancer cells. Moreover, especially where gliomas are concerned, their ability to secrete glutamate, which excites glutamatergic neurons, drives the latter to consume even more lactate and spare more glucose. Such symbiotic exchange, especially at the early stages of malignancy, assures the budding cancer cells an ample glucose supply ahead of the development of additional vasculature. This hypothesis focuses on brain cancers but may also apply to other cancer types.
Keywords:
Glioma
; glucose
; glycolysis
; lactate
; neuron
; symbiosis
Introduction
A century ago, Otto Warburg showed that cancer cells consume large amounts of glucose and secrete excessive lactate [1]. This hallmark of malignancy, known as Warburg’s effect, has been cited over 28,700 times since 1926, and over 5,200 times in the first 5 months of 2025, according to Google Scholar search. The main impetus for this activity has always been the pursuit of possible anti-cancer therapy by understanding this phenomenon. Nevertheless, Warburg’s observation of glucose utilization without oxygen by cancerous cells occurred more than a decade prior to the revelation in 1940 of the metabolic sequence of the glycolytic pathway. The general notion that lactate is a useless end-product of fermentation that also could be, at high levels, poisonous to normal cells, has prevailed for years. Our knowledge about the relationships between cancer cells and the normal cells that surround them has grown significantly over the past half a century. However, most published studies are focused mainly on the routes by which cancerous tissue utilizes its surrounding healthy tissue to proliferate and survive [2,3,4,5]. Many studies on the interactions between these cell populations deal with brain cancers i.e., gliomas and glioblastomas. While we have a better understanding of the benefits the different brain cancer types gain from their surrounding neurons, very little is known about any possible benefits that neurons may gain from those brain cancers. Where glycolysis is concerned, significant discoveries over the past four decades have brought about a paradigm shift of this pathway [6,7,8,9,10]. Accordingly, glycolysis should not be divided into ‘aerobic’ and ‘anaerobic’ pathways with two different outcomes, but rather being described without any prefix, just ‘glycolysis,’ the cytosolic pathway that hydrolyzes glucose to lactate, where its last reaction, the conversion of pyruvate to lactate is catalyzes by cytosolic lactate dehydrogenase (cLDH), independently of the presence or absence of oxygen or mitochondria (Figure 1). Lactate, in turn, can enter the mitochondrion via a monocarboxylate transporter (MCT), where it is converted back to pyruvate via a specific mitochondrial lactate dehydrogenase (mLDH). The latter then enters the mitochondrial tricarboxylic acid (TCA) cycle via acetyl CoA [11,12,13,14]. The idea that lactate is an oxidative energy substrate for neurons has slowly gained acceptance. It is plausible that lactate is a favored mitochondrial substrate over glucose, since glucose requires an investment of two moles of adenosine triphosphate (ATP) before it is converted to lactate. By contrast, the oxidative utilization of lactate does not require any investment of ATP, and where one mole of lactate produces 17 moles of ATP through the mitochondrial TCA cycle, the electron transport chain and its coupled oxidative phosphorylation (OXPHOS). Does lactate, secreted by glioma cells, could be used as an oxidative mitochondrial substrate for ATP biosynthesis by neurons?
Glioma Neuron Symbiosis (GNS) Hypothesis: Exchanging Lactate for Glucose
It is hypothesized that neurons regularly benefit from the endless supply of glioma-produced lactate, the preferred neuronal oxidative mitochondrial energy substrate over glucose, especially at the initial stages of malignancy, just as much as glioma cells benefit from the neuronal machinery. Moreover, the neuronal preference of lactate over glucose spares the latter, making it readily available to the glucose-craving glioma cells. Such symbiotic relationships would explain the tendency of cancerous cells (gliomas) to flourish in the vicinity of the more active brain regions [5]. Contrary to the established thinking, according to which cancerous cells almost parasitically utilize normal cells to propagate and survive, the GNS hypothesis postulates that both cell types benefit from each other, i.e., they have a symbiotic relationship.
The GNS hypothesis has been culminated following a four-decade long research of brain energy metabolism. That research established lactate as the end-product of the brain glycolytic pathway, independent of the presence or absence of oxygen, demonstrating the preference of neurons, especially glutamatergic ones, to utilize it as the oxidative mitochondrial substrate. In addition, the metabolic relationship between astrocytes and neurons as laid out by the astrocyte neuron lactate shuttle (ANLS) hypothesis [15], and the possible origin of glioma cells from stem cells of the oligodendroglial type, both lend support to a postulated symbiotic relationship between glioma cells and neurons.
Evolution of the Hypothesis
The discovery in 1988 that lactate can both support survival of brain tissue and its function in vitro [7], despite the skepticism it faced for years, is now accepted universally [8,9,10,11,12,13,14]. The astrocyte neuron lactate shuttle (ANLS) hypothesis was published in 1994 [15]. According to this hypothesis, the excitatory neurotransmitter glutamate, after activating glutamatergic neurons, is taken up by astrocytes, a function that requires the participation of the Na+/K+- ATPase pump [16]. The ATP that the pump consumes for its action is produced by a glycolytic pathway dedicated to this function. The lactate thus produced is transported out of astrocytes through membranal monocarboxylate transporters (MCT1 and MCT4), and into neurons via MCT2, where it is consumed oxidatively [17,18,19,20,21,22,23]. Our own study demonstrated that glial cells are the source of lactate consumed by neurons [24]. Although the ANLS hypothesis is still being debated among its supporters and detractors, ample evidence supports its central concept: Neurons oxidatively consume lactate mitochondrially as the substrate of the TCA cycle and its coupled OXPHOS, the main source of ATP needed for neuronal function [25,26,27,28,29,30,31,32,33,34,35,36,37,38,39,40,41,42,43,44,45,46,47,48,49,50,51,52,53,54,55,56]. Neuronal preference for lactate over glucose has been documented both in vitro and in vivo [40,41,44,45,46,47]. Taking into consideration the above-cited studies, it is reasonable to extrapolate from neuronal preference for lactate as the oxidative energy substrate to the hefty amounts of lactate glioma cells produce. Moreover, neuronal glucose spared using lactate can be easily transported and made available for use by glioma cells. Much understanding has been gained over the past two decades demonstrating the gliomas’ ability to hijack neuronal mechanisms [3,56,57,58], flourish in the neuronal microenvironment [4], and especially interact with active glutamatergic neurons [5]. While the origin of glioma cells is still being deliberated, accumulated evidence points at “neural stem or precursor cells of the oligodendroglial type” [5 and refs. within], which could explain the ability of glioma cells to manipulate neurons for their growth needs, the very needs that are specifically provided by active neurons [5,59]. Clearly, in all the studies on the topic, the prevailing message is that glioma cells take over the neuronal machinery necessary for their own proliferation, a one-way relationship, where these cells take all and give nothing back. However, the probable glial origin of glioma cells could indicate that their interactions with neurons are like those demonstrated between astrocytes and neurons, where these two cell types have, in essence, a symbiotic relationship [15,17,19]. Astrocytes take up the neurotransmitter glutamate following its activation of neurons, an uptake that requires the activity of the astrocytic Na+/K+-ATPase pump, the ATP for which its supplied by its own dedicated glycolytic pathway. The glycolytic product, lactate, is transported to neurons, where it is utilized as a substrate for their oxidative mitochondrial ATP production. Therefore, like astrocytes, glioma cells could supply neurons, especially glutamatergic ones, with lactate. In return, the glucose spared by neurons due to their preference for lactate, is readily consumed by the glucose-craving glioma cells. Moreover, evidence shows that glioma cells can secrete glutamate, which excites glutamatergic neurons [59,60,61,62]. Such excitation should increase neuronal lactate consumption and spare even more glucose for consumption by glioma cells (Figure 2). While direct communications between glioma cells and neurons have been described, including the formation of synaptic connections between the two cell types, no reports exist on possible benefits neurons may gain through their interactions with glioma cells. A single review article [63] alludes to the possibility that neurons interact with astrocytoma cells, where similarly to astrocytes, they extrude the lactate produced glycolytically, making it available for neuronal use. Therefore, the present hypothesis evolved with this information in mind. It proposes that neurons regularly benefit from the endless supply of glioma-produced lactate, the preferred neuronal oxidative energy substrate over glucose. That preference spares neuronal glucose, making it readily available to the glucose-craving glioma cells. The latter ability to secrete glutamate, which excites glutamatergic pyramidal neurons, increases neuronal consumption of lactate, sparing more glucose for gliomas’ consumption. Such a symbiotic relationship, at least at the beginning of gliomas’ proliferation, benefits both cell types equally. This attraction between the two cell types is probably driven by both the glucose-hungry glioma cell and the lactate-preferred neuron. The fundamental appetite of normal and malignant cells alike for continuous supply of energy substrates could be at the basis of other types of cancer interaction with the normal tissue they invade. Recognizing the ability of certain cell types in other tissues and organs to efficiently utilize lactate, the glycolytic end-product of cancer cells (Warburg’s effect), could explain the tendency of certain cancer types to relocate to and proliferate in other locations.
Testing the Hypothesis
This GNS hypothesis could be tested in vitro, using glioma cell lines [62] alone or preferentially, when combined with neuronal cell lines, by measuring lactate and/or glucose transport through the manipulation of MCTs activity. Such manipulation can be achieved via the use of known MCT inhibitors, such as α-cyano-4-hydroxycinnamate [64,65], where the transport of lactate, extruded from cancerous cells, could be blocked. This blockade would prevent neuronal utilization of glioma lactate. It could most likely also block lactate transport into neurons and their mitochondria, consequently preventing any neuronal glucose-sparing. An alternative in vitro approach would be the exposure of glioma cell lines to the glial metabolic toxin fluorocitrate (FC) [24,66]. This toxin specifically impairs the flow of carbon through the glial TCA cycle [66]. Considering the similarity between glial cells and glioma cells, it is possible that glioma cells would respond to FC in the same manner that glia cells do. Under conditions when neurons are incubated with glioma cells, where the former mainly depend on glioma-released lactate as their oxidative energy substrate, FC should indirectly diminish neuronal survival due to the toxin-induced death of glioma cells.
Implications
The hypothesized symbiosis between glioma cells and neurons first implies that there is a facet of the interactions between these two cell types that, so far, has not been considered, let alone investigated. Establishing such a symbiotic relationship between glioma cells and neurons could explain their reciprocated attraction and possible mutual dependency. Second, if such a symbiosis exists, it could open a new direction in the development of anti-cancer treatments, especially if said symbiosis is crucial for the survival of one or both cell types.
Conclusions
The GNS hypothesis proposes that neurons regularly benefit from the endless supply of glioma-produced lactate, the preferred neuronal oxidative energy substrate over glucose. That preference spares neuronal glucose, which becomes readily available for use by the glucose-craving glioma cells. The latter’s ability to secrete glutamate, which excites glutamatergic pyramidal neurons, increases neuronal consumption of lactate, and sparing more glucose for consumption by glioma cells. Such a symbiotic relationship, at least at the beginning of a glioma’s proliferation, benefits both cell types equally. Therefore, the attraction between the two cell types could be driven by both, the glucose-demanding glioma cell and the lactate-favored neuron. The ability of glioma cells to secrete glutamate should enhance its symbiosis with neurons. Two possible in vitro experimental procedures are offered to test the hypothesis and predicted implications of the hypothesis, if confirmed, are also included here.
Funding
No specific funding was received for the work discussed in this manuscript.
Acknowledgments
The author wishes to thank Paige Hessel for her invaluable editorial critique during the preparation of this manuscript.
Conflicts of Interest
The author has no conflicts of interest to declare.
References
- Warburg, F. Wind, E. Negelein, The metabolism of tumors in the body. J. Gen. Physiol. 8 (1927) 519 –530. [CrossRef]
- V.A. Cuddapah, S. Robel, S. Watkins, H. Sontheimer, (2014). A neurocentric perspective on glioma invasion. Nature Rev. Neurosci. 15 (2014) 455-465. [CrossRef]
- V. Venkataramani, Y. Yang, M.C. Schubert, E. Reyhan, S.K. Tetzlaff, N. Wißmann, M. Botz, S.J. Soyka, C.A. Beretta, R.L. Pramatarov, L. Fankhauser, L.G.A. Freudenberg, J. Wagner, D.I. Tanev, M. Ratliff, R. Xie, T. Kessler, D.C. Hoffmann, L. Hai, Y. Dorflinger, S. Hoppe, Y.A. Yabo, A. Golebiewska, S.P. Niclou, F. Sahm, A. Lasorella, M. Slowik, L. Doring, A. Iavarone, W. Wick, T. Kuner, F. Winkler. Glioblastoma hijacks neuronal mechanisms for brain invasion. Cell 185 (2022) 2899–2917 . [CrossRef]
- C.B. Crivii, A.B. Boșca, C.S. Melincovici, A.M. Constantin, M. Mărginean, E. Dronca, R. Suflețel, D. Gonciar, M. Bungărdean, A. Șovrea, Glioblastoma microenvironment and cellular interactions. Cancers, 14 (2022) 1092. [CrossRef]
- S. Gillespie, M. Monje, M. An active role for neurons in glioma progression: making sense of Scherer’s structures. Neuro-oncology, 20 (2018) 1292-1299. [CrossRef]
- G.A. Brooks, Lactate: glycolytic end product and oxidative substrate during sustained exercise in mammals—the “lactate shuttle. Circulation, respiration, and metabolism: current comparative approaches. Berlin, Heidelberg: Springer Berlin Heidelberg, Germany, 1985, 208-218.
- Schurr, C.A. West, B.M. Rigor, B.M. Lactate-supported synaptic function in the rat hippocampal slice preparation. Science, 1988, 240, 1326–1328. [CrossRef]
- L.B. Gladden, Lactate metabolism: a new paradigm for the third millennium. J. Physiol. 558 (2004) 5-30. [CrossRef]
- L.B. Gladden, A lactatic perspective on metabolism. Medicine and science in sports and exercise, 40 (2008) 477-485. [CrossRef]
- M.M. Hall, S. Rajasekaran, T.W. Thomsen, A.R. Peterson, (2016). Lactate: friend or foe. PM&R, 8 (2016) S8-S15. [CrossRef]
- S. Passarella, L. de Bari, D. Valenti, R. Pizzuto, G. Paventi, A. Atlante, Mitochondria and L-lactate metabolism. FEBS letters, 582 (2008) 3569-3576. [CrossRef]
- Schurr, Cerebral glycolysis: a century of persistent misunderstanding and misconception. Front. Neurosci. 8 (2014) 360. [CrossRef]
- M.J. Rogatzki, B.S. Ferguson, M.L. Goodwin, L.B. Gladden, Lactate is always the end product of glycolysis. Front. Neurosci. 22 (2015) 22. [CrossRef]
- G. Van Hall, Lactate as a fuel for mitochondrial respiration. Acta Physiol. Scand. 168 (2000) 643-656. [CrossRef]
- L. Pellerin, P.J. Magistretti, Glutamate uptake into astrocytes stimulates aerobic glycolysis: a mechanism coupling neuronal activity to glucose utilization. Proc. Nat. Acad. Sci. 91 (1994) 10625-10629. [CrossRef]
- L. Pellerin, P.J. Magistretti, Excitatory amino acids stimulate aerobic glycolysis in astrocytes via an activation of the Na+/K+ ATPase. Dev. Neurosci. 18 (1996) 336–342. [CrossRef]
- L. Pellerin, Lactate as a pivotal element in neuron-glia metabolic cooperation. Neurochem. Intern. 43 (2003) 331-338. [CrossRef]
- J. Handy, Lactate—the bad boy of metabolism, or simply misunderstood? Current Anaesth. & Crit Care 17 (2006) 71-76. [CrossRef]
- L. Pellerin, P.J. Magistretti, Sweet sixteen for ANLS. J. Cereb Blood Flow Metab. 32 (2012) 1152-1166. [CrossRef]
- S. Passarella, L. de Bari, D. Valenti, R. Pizzuto, G. Paventi, A. Atlante, Mitochondria and L-lactate metabolism. FEBS letters, 582 (2008) 3569-3576. [CrossRef]
- Schurr, Cerebral glycolysis: a century of persistent misunderstanding and misconception. Front. Neurosci. 8 (2014) 360. [CrossRef]
- M.J. Rogatzki, B.S. Ferguson, M.L. Goodwin, L.B. Gladden, Lactate is always the end product of glycolysis. Front. Neurosci. 22 (2015) 22. [CrossRef]
- G. Van Hall, Lactate as a fuel for mitochondrial respiration. Acta Physiol. Scand. 168 (2000) 643-656. [CrossRef]
- M. Nalbandian, M. Takeda, Lactate as a signaling molecule that regulates exercise-induced adaptations.” Biology, 5 (2016) 38. [CrossRef]
- B.S. Ferguson, M.J. Rogatzki, M.L. Goodwin, D.A. Kane, Z. Rightmire, L.B. Gladden, Lactate metabolism: historical context, prior misinterpretations, and current understanding. Euro J. Appl. Physiol. 118 (2018) 691-728. [CrossRef]
- Schurr, R.S. Payne, J.J. Miller, B.M. Rigor, Glia are the main source of lactate utilized by neurons for recovery of function posthypoxia. Brain Res. 774 (1997) 221-224. [CrossRef]
- Schurr, Glycolysis Paradigm Shift Dictates a Reevaluation of Glucose and Oxygen Metabolic Rates of Activated Neural Tissue. Front. Neurosci. 12 (2018) 700. [CrossRef]
- Schurr, Lactate: the ultimate cerebral oxidative energy substrate? J. Cereb Blood Flow Metab. 26 (2006) 142-152. [CrossRef]
- Schurr, Lactate: a major and crucial player in normal function of both muscle and brain. J. Physiol, 586 (2008) 2665. [CrossRef]
- Schurr, From rags to riches: Lactate ascension as a pivotal metabolite in neuroenergetics. Front. Neurosci. 17 (2023) 1145358. [CrossRef]
- Schurr, How the ‘Aerobic/Anaerobic Glycolysis’ Meme Formed a ‘Habit of Mind’ Which Impedes Progress in the Field of Brain Energy Metabolism. Intern. J. Mol. Sci. 25 (2024) 1433. [CrossRef]
- P.G. Bittar, Y. Charnay, L. Pellerin, C. Bouras, P.J. Magistretti, Selective distribution of lactate dehydrogenase isoenzymes in neurons and astrocytes of human brain. J. Cereb. Blood Flow Metab. 16 (1996) 1079-1089. [CrossRef]
- L. Pellerin, G. Pellegri, P.G. Bittar, Y. Charnay, C. Bouras, J.L. Martin, N. Stella, P.J. Magistretti, Evidence supporting the existence of an activity-dependent astrocyte-neuron lactate shuttle. Dev. Neurosci. 20 (1998) 291-299. [CrossRef]
- L. Pellerin, G. Pellegri, J.L. Martin, P.J. Magistretti, Expression of monocarboxylate transporter mRNAs in mouse brain: support for a distinct role of lactate as an energy substrate for the neonatal vs. adult brain. Proc. Nat Acad Sci, 95 (1998) 3990-3995. [CrossRef]
- K. Pierre, L. Pellerin, R. Debernardi, B.M. Riederer, P.J. Magistretti, Cell-specific localization of monocarboxylate transporters, MCT1 and MCT2, in the adult mouse brain revealed by double immunohistochemical labeling and confocal microscopy. Neuroscience, 100 (2000) 617-627. [CrossRef]
- Aubert, R. Costalat, P.J. Magistretti, L. Pellerin, Brain lactate kinetics: modeling evidence for neuronal lactate uptake upon activation. Proc. Nat. Acad. Sci. 102 (2005) 16448-16453. [CrossRef]
- A.K. Bouzier-Sore, P. Voisin, P. Canioni, P.J. Magistretti, L. Pellerin, L. Lactate is a preferential oxidative energy substrate over glucose for neurons in culture. J. Cereb. Blood Flow Metab. 23 (2003) 1298-1306. [CrossRef]
- A.K. Bouzier-Sore, P. Voisin, V. Bouchaud, E. Bezancon, J. Franconi, L. Pellerin, Competition between glucose and lactate as oxidative energy substrates in both neurons and astrocytes: a comparative NMR study. European J. Neurosci. 24 (2006) 1687-1694. [CrossRef]
- L. Pellerin, A.K. Bouzier-Sore, A. Aubert, S. Serres, M. Merle, R. Costalat, P.J. Magistretti, P.J. Activity-dependent regulation of energy metabolism by astrocytes: an update. Glia, 55 (2007) 55, 1251-1262. [CrossRef]
- M.T. Wyss, R. Jolivet, A. Buck, P.J. Magistretti, B. Weber, In vivo evidence for lactate as a neuronal energy source. J. Neurosci. 31 (2011) 7477-7485. [CrossRef]
- P. Proia, C.M. Di Liegro, G. Schiera, A. Fricano, I. Di Liegro, Lactate as a metabolite and a regulator in the central nervous system. Internat. J. Mol. Sci. 17 (2016) 1450. [CrossRef]
- Y. Hu, G.S. Wilson, A temporary local energy pool coupled to neuronal activity: fluctuations of extracellular lactate levels in rat brain monitored with rapid-response enzyme-based sensor. J. Neurochem. 69 (1997) 1484-1490. [CrossRef]
- Schurr, J.J. Miller, R.S. Payne, B.M. Rigor, An increase in lactate output by brain tissue serves to meet the energy needs of glutamate-activated neurons. J. Neurosci. 19 (1999) 34-39. [CrossRef]
- G.A. Brooks, Intra-and extra-cellular lactate shuttles. Medicine and science in sports and exercise, 32 (2000) 790-799. [CrossRef]
- G.A. Brooks, Cell–cell and intracellular lactate shuttles. J. Physiol. 587 (2009) 5591-5600. [CrossRef]
- H. Qu, A. Håberg, O. Haraldseth, G. Unsgård, U. Sonnewald, U. 13C MR spectroscopy study of lactate as substrate for rat brain. Dev. Neurosci 22 (2000) 429-436. [CrossRef]
- S. Mangia, G. Garreffa, M. Bianciardi, F. Giove, F. Di Salle, B. Maraviglia, The aerobic brain: lactate decrease at the onset of neural activity. Neuroscience, 118 (2003) 7-10. [CrossRef]
- D. Smith, A. Pernet, W.A. Hallett, E. Bingham, P.K. Marsden, S.A. Amiel, Lactate: a preferred fuel for human brain metabolism in vivo. J. Cereb. Blood Flow Metab. 23 (2003) 658-664. [CrossRef]
- K.A. Kasischke, H.D. Vishwasrao, P.J. Fisher, W.R. Zipfel, W.W. Webb, Neural activity triggers neuronal oxidative metabolism followed by astrocytic glycolysis. Science, 305 (2004) 99-103. [CrossRef]
- A.S. Herard, A. Dubois, C. Escartin, K. Tanaka, T. Delzescaux, P. Hantraye, G. Bonvento, Decreased metabolic response to visual stimulation in the superior colliculus of mice lacking the glial glutamate transporter GLT-1. Eur. J. Neurosci. 22 (2005) 1807-1811. [CrossRef]
- Schurr, R.S. Payne, Lactate, not pyruvate, is neuronal aerobic glycolysis end- product: an in vitro electrophysiological study. Neuroscience, 147 (2007) 613-619. [CrossRef]
- T. Hashimoto, R. Hussien, H. Cho, D. Kaufer, G.A. Brooks, Evidence for the mitochondrial lactate oxidation complex in rat neurons: demonstration of an essential component of brain lactate shuttles. PloS one, 3 (2008) e2915. [CrossRef]
- J.S. Erlichman, A. Hewitt, T.L. Damon, M. Hart, J. Kurascz, A. Li, J.C. Leiter, Inhibition of monocarboxylate transporter 2 in the retrotrapezoid nucleus in rats: a test of the astrocyte–neuron lactate-shuttle hypothesis. J. Neurosci. 28 (2008) 4888-4896. [CrossRef]
- C.N. Gallagher, K.L. Carpenter, P. Grice, D.J. Howe, A. Mason, I. Timofeev, D.K. Menon, P.J. Kirpatrick, J.D. Pickard, G.R. Sutherland, P.J. Hutchinson, The human brain utilizes lactate via the tricarboxylic acid cycle: a 13C-labelled microdialysis and high-resolution nuclear magnetic resonance study. Brain, 132 (2009) 2839-2849. [CrossRef]
- J. Chuquet, P. Quilichini, E.A. Nimchinsky, G. Buzsáki, Predominant enhancement of glucose uptake in astrocytes versus neurons during activation of the somatosensory cortex. J. Neurosci. 30 (2010) 15298-15303. [CrossRef]
- C.R. Figley, Lactate transport and metabolism in the human brain: implications for the astrocyte-neuron lactate shuttle hypothesis. J. Neurosci 31 (2011) 4768-4770. [CrossRef]
- Dias, E. Fernandes, R.M. Barbosa, J. Laranjinha, A. Ledo, Astrocytic aerobic glycolysis provides lactate to support neuronal oxidative metabolism in the hippocampus. Biofactors, 49 (2023) 875-886. [CrossRef]
- Y. Pan, M. Monje, Neuron–Glial Interactions in Health and Brain Cancer. Adv. Biol. 6 (2022) 2200122. [CrossRef]
- E. Jung, J. Alfonso, H. Monyer, W. Wick. F. Winkler, Neuronal signatures in cancer. Intern J. Cancer, 147 (2020) 3281-3291. [CrossRef]
- H. Tianzhen, H. Shi, M. Zhu, C. Chen, Y. Su, S. Wen, X. Zhang, J. Chen, Q. Huang, H. Wang, Glioma-neuronal interactions in tumor progression: Mechanism, therapeutic strategies and perspectives. Inter. J. Oncol. 61 (2022) 104. [CrossRef]
- S.C. Buckingham, S.L. Campbell, B.R. Haas, V. Montana, S. Robel, T. Ogunrinu, H. Sontheimer, Glutamate release by primary brain tumors induces epileptic activity. Nature Med. 17 (2011) 1269-1274. [CrossRef]
- S.L. Campbell, S.C. Buckingham, H. Sontheimer. Human glioma cells induce hyperexcitability in cortical networks. Epilepsia 53 (2012) 1360-1370. [CrossRef]
- V. Venkataramani, D.I. Tanev, C. Strahle, A. Studier-Fischer, L. Fankhauser, T. Kessler, C. Körber, M. Kardorff, M. Ratliff, R. Xie, H. Horstmann, M. Messer, S.P. Paik, J. Knabbe, F. Sahm, F.T. Kurz, A.A. Acikgöz, F. Herrmannsdörfer, A. Agarwal, D.E. Bergles, A. Chalmers, H. Miletic, S. Turcan, C. Mawrin, D. Hänggi, H-K. Liu, W. Wick, F. Winkler, T. Kuner, Glutamatergic synaptic input to glioma cells drives brain tumour progression. Nature, 573 (2019) 538. [CrossRef]
- Z-C, Ye, H. Sontheime, Glioma Cells Release Excitotoxic Concentrations of Glutamate. Cancer Res, 59 (1999) 4383–4391.
- D.A. Turner, D.C. Adamson, Neuronal-astrocyte metabolic interactions: understanding the transition into abnormal astrocytoma metabolism. J. Neuropathol. Exper. Neurol. 70 (2011) 167-176. [CrossRef]
- Schurr, R.S. Payne, J.J. Miller, M.T. Tseng, B.M. Rigor, Blockade of lactate transport exacerbates delayed neuronal damage in a rat model of cerebral ischemia. Brain Res. 895 (2001) 268-272. [CrossRef]
- Schurr, R.S. Payne, M.T. Tseng, E. Gozal, D. Gozal, Excitotoxic preconditioning elicited by both glutamate and hypoxia and abolished by lactate transport inhibition in rat hippocampal slices. Neurosci. Let. 307 (2001) 151-154. [CrossRef]
- R.A. Swanson, S.H. Graham, Fluorocitrate and fluoroacetate effects on astrocyte metabolism in vitro. Brain Res. 664 (1994) 94-100. [CrossRef]
Figure 1.
The paradigm shift of glycolysis that took place over the past four decades presents the first metabolic pathway to be elucidated as a series of 11 enzymatic reactions that begin with glucose and end with lactate, not pyruvate, independently of the presence or absence of oxygen or mitochondria. As such, it also guarantees the continuous supply of reducing power in the form of nicotinamide adenine dinucleotide (NADH), preserving the cyclical nature of the pathway.
Figure 1.
The paradigm shift of glycolysis that took place over the past four decades presents the first metabolic pathway to be elucidated as a series of 11 enzymatic reactions that begin with glucose and end with lactate, not pyruvate, independently of the presence or absence of oxygen or mitochondria. As such, it also guarantees the continuous supply of reducing power in the form of nicotinamide adenine dinucleotide (NADH), preserving the cyclical nature of the pathway.
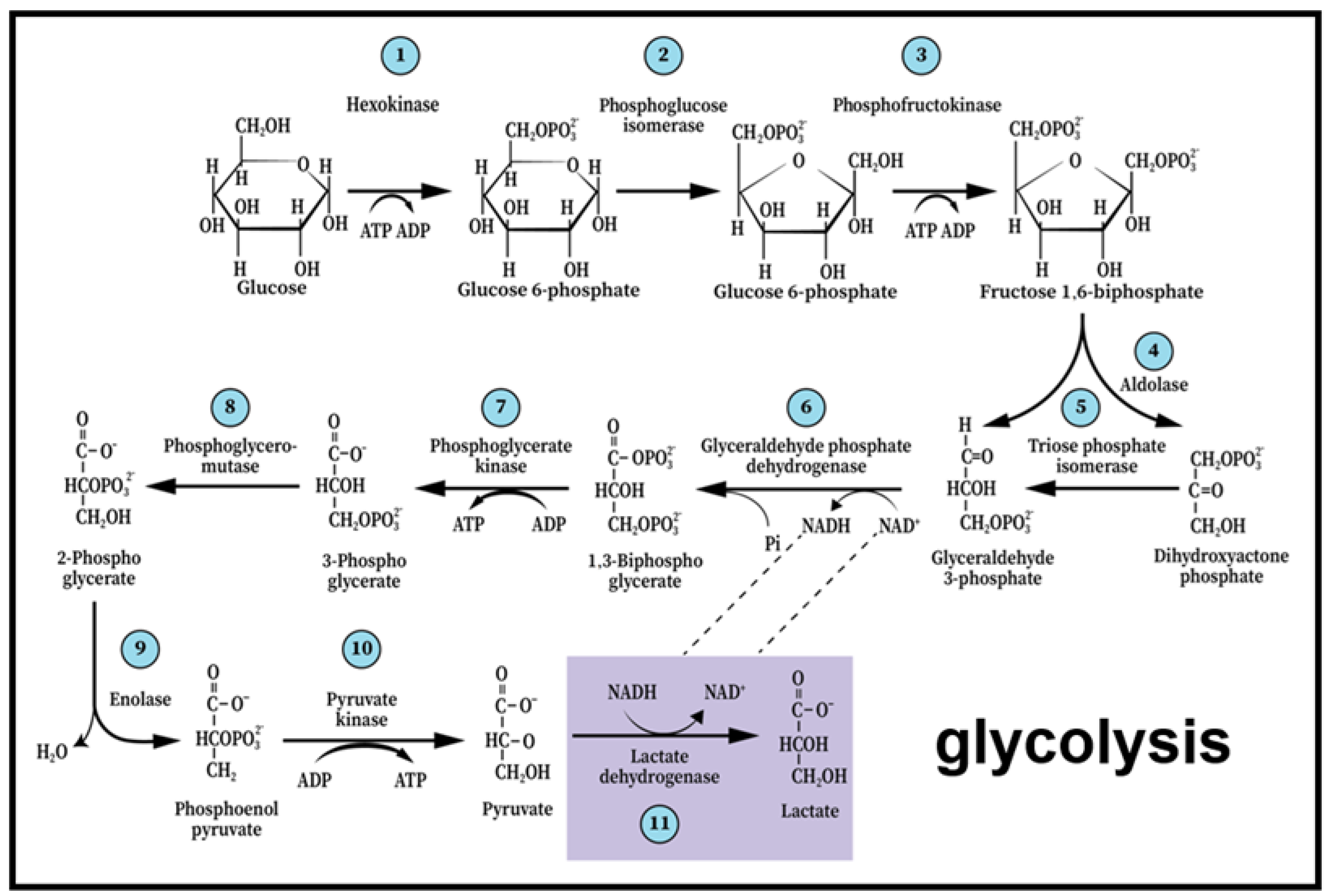
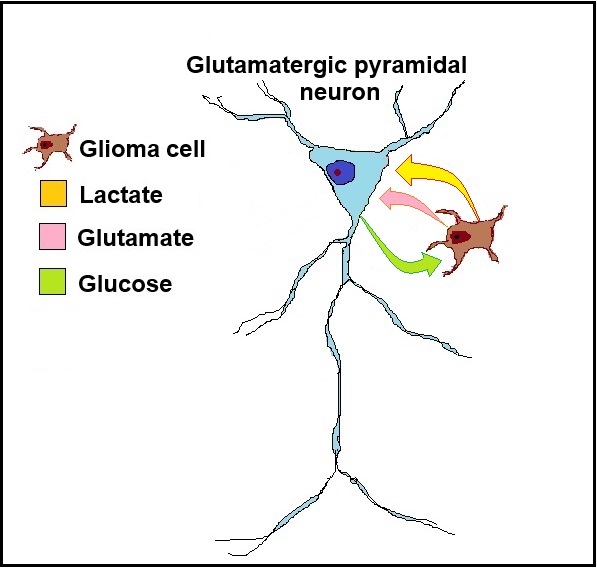
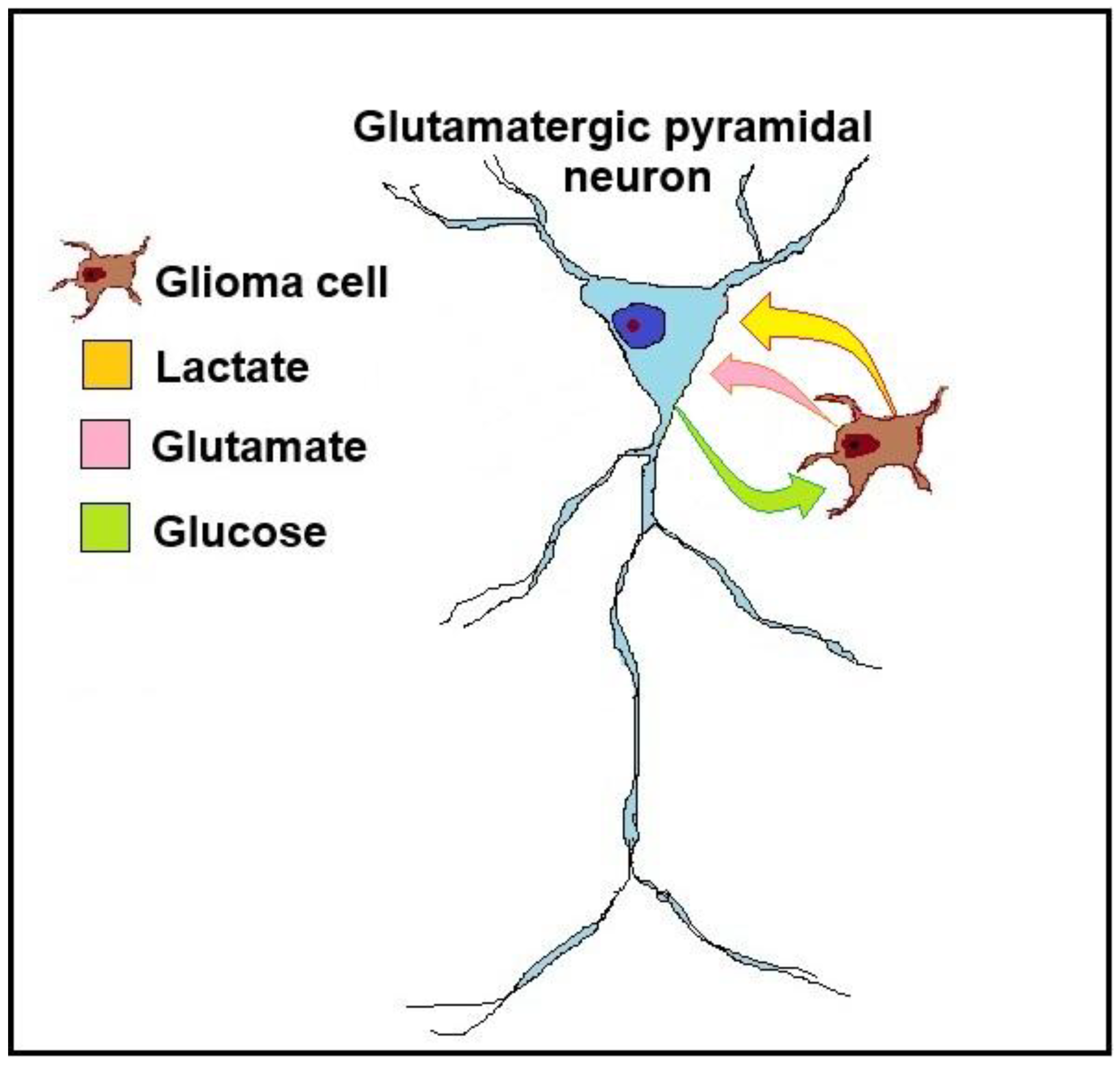
Figure 2.
A schematic illustration of the glioma neuron symbiosis (GNS) hypothesis, presented as a relationship between a glioma cell and a glutamatergic pyramidal neuron. The glioma cell secretes the neurotransmitter glutamate, which activates the neuron. As the glioma cell consumes glucose glycolytically to produce ATP, it also generates lactate, which is transported via monocarboxylate transporters (MCTs) from the glioma cell to the neuron. The neuron consumes lactate as its preferred substrate of oxidative energy metabolism, sparing its own glucose supply, making it available for the glioma cell.
Figure 2.
A schematic illustration of the glioma neuron symbiosis (GNS) hypothesis, presented as a relationship between a glioma cell and a glutamatergic pyramidal neuron. The glioma cell secretes the neurotransmitter glutamate, which activates the neuron. As the glioma cell consumes glucose glycolytically to produce ATP, it also generates lactate, which is transported via monocarboxylate transporters (MCTs) from the glioma cell to the neuron. The neuron consumes lactate as its preferred substrate of oxidative energy metabolism, sparing its own glucose supply, making it available for the glioma cell.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



