
Submitted:
03 June 2025
Posted:
03 June 2025
You are already at the latest version
Abstract
Walnut (Juglans regia L.) holds a significant position among Kazakhstan’s nut crops, both in terms of cultivation and consumption. Although genetic resources of walnut are accessible within the country, they remain underexplored, yet they represent a valuable foundation for breeding initiatives and the development of locally adapted varieties. Currently, a comprehensive genetic profile of these resources is laking, which hampers their effective utilization. The aim of this research was to evaluate the genetic diversity within a collection of walnut accessions collected from the southern and southeastern regions of Kazakhstan, including Almaty and Turkestan regions. To achieve this, eight SSR molecular markers were employed, providing insights into the genetic structure and relationships among the germplasm. All markers exhibited a high degree of polymorphism, with the WGA276 locus standing out as the most informative, displaying the greatest number of alleles and a high Shannon diversity index. The average expected heterozygosity (He) was 0.704, markedly higher than the observed heterozygosity (Ho), which was 0.547. The findings indicated substantial genetic variation and intra-population polymorphism: on average, 5.875 alleles per locus were identified, and the Shannon index was 1.444. The population from Turkestan region (population 1, Sairam-Ugam) demonstrated the highest levels of diversity. Analyses of genetic structure, conducted through STRUCTURE, PCoA, and UPGMA methods, confirmed the existence of two genetically distinct groups exhibiting considerable diversity. These findings highlight the importance of conserving and harnessing the genetic resources of Kazakhstan’s J. regia populations for future breeding efforts.
Keywords:
Juglans regia L.
; microsatellite markers
; genetic diversity
; polymorphism
; population structure
; breeding
1. Introduction
Juglans regia L. is a globally significant nut crop with considerable economic importance. Its kernels are distinguished by a rich profile of nutrients and phytochemicals that confer various health benefits [1,2,3,4,5]. These nuts serve as a valuable source of macronutrients, including high-quality proteins, healthy fatty acids, vitamins, and essential minerals such as omega-3 fatty acids [6,7,8]. Furthermore, they contain bioactive compounds such as flavonoids and polyphenols, which exhibit antioxidant properties [7,9]. The elevated protein and lipid content of walnut kernels underpin their essential role in human nutrition [10]. They are widely utilized in fresh consumption, confectionery manufacturing, and oil extraction, with fat content ranging from 65% to 70% [11,12]. The superior nutritional profile of walnuts underscores their importance in dietary practices, leading to their recognition as a priority crop by the Food and Agriculture Organization (FAO) [10].
The origin and dissemination of Juglans regia in Asia are the result of intricate interactions between biogeographical processes and human activities. The spread of walnuts across the continent was significantly influenced by trade routes, notably the Silk Road and the Persian Royal Road, which served as "green corridors" enabling the movement of nuts despite geographical obstacles. These routes facilitated the widespread exchange of walnut germplasm throughout Asia, contributing to its broad distribution [13,14]. Currently, the primary range of walnut extends from the Balkan region eastward into Asia, encompassing countries such as China, Iran, Kazakhstan, Afghanistan, and Pakistan [12,15].
Currently, China stands as the world's largest producer of walnuts, contributing approximately 50.27% of the global output. Following China, the leading producers include the United States, Iran, Turkey, Ukraine, Chile, and several countries within the European Union [11,16,17]. Over the period from 2012 to 2021, global walnut production experienced a significant upward trend, increasing from approximately 2.37 million tons to over 3.50 million tons [18]. This growth can be attributed to increasing international demand, rising market prices, advancements in cultivation techniques, and the development and adoption of new high-yielding, pest- and disease-resistant varieties.
Investigating and evaluating the genetic diversity of walnut germplasm in Central Asia is crucial for advancing breeding initiatives tailored to this region. Recently, Kazakhstan has experienced a growing demand for walnuts, particularly in areas conducive to cultivation, which has led to increased yields and expanded production. Although Kazakhstan possesses a diverse gene pool of walnut resources, there is currently no established national breeding program dedicated to their improvement. Several international collaborations are underway to develop breeding technologies, involving joint projects with Russia, the United States, Bulgaria, the Czech Republic, and Poland, aimed at harnessing and conserving the region’s genetic potential [19].
Kazakhstan's predominantly continental climate plays a crucial role in fostering a highly diverse genetic pool of walnuts, including genotypes adapted to withstand low temperatures and produce high-quality fruits. These genetic resources are of great importance for the development of targeted breeding programs focused on improving winter hardiness, enhancing resistance to pests and diseases, and increasing both yield and fruit quality. Leveraging this genetic diversity is vital for ensuring the sustainability and resilience of walnut cultivation in the region, ultimately supporting its economic and agricultural development [19].
A study was conducted on the genetic resources of walnut (Juglans regia L.) in Kazakhstan, aiming to identify and select promising genotypes for further use in breeding programs, as well as to assess the genetic diversity of this species [19,20]. As a result, the most promising genotypes were selected, which can be further utilized in breeding programs and for commercial cultivation. The region’s favourable soil and climatic conditions present significant potential for the development of walnut industry in southern Kazakhstan. An evaluation of local walnut populations allowed for the selection of genotypes best suited to the specific environmental conditions of the southern and south-eastern regions [21]. Previously, our research included mapping the distribution of Juglans regia in this area, as well as morphological characterization of walnut fruits [22]. Additional studies focused on the species’ biological traits, its adaptation to local climate, and the impact of cold stress on its morphological, physiological, and phytochemical properties [23]. Valuable germplasm from natural populations was collected, preserved in vitro, and stored in a cryobank for long-term conservation and future utilization [24,25]. Moreover, efforts were made to assess the resistance of local varieties to pests and diseases, including bacterial blight caused by Pantoea agglomerans, leading to the identification of resistant genotypes [26]. Despite extensive research and the region’s conducive natural conditions, the commercial cultivation of walnuts in Kazakhstan remains relatively underdeveloped.
Kazakhstan possesses extensive genetic resources related to the germplasm of Juglans regia; however, their identification and detailed study have yet to be conducted. Investigating the genetic diversity within these resources is essential for effective management of the gene pool and for enhancing breeding efficiency. Molecular markers serve as a vital tool in this context, enabling precise characterization of germplasm, verification of cultivar origins, and elucidation of genetic relationships among different accessions. Unlike morphological traits, molecular markers are unaffected by environmental conditions and can reveal cryptic genetic variation, thereby significantly broadening the scope for selective breeding and conservation of genetic diversity.
Various types of molecular markers have been developed to facilitate genetic studies, including RFLP (Restriction Fragment Length Polymorphism) [27], RAPD (Random Amplified Polymorphic DNA) [28], AFLP (Amplified Fragment Length Polymorphism) [29,30], SSR (Simple Sequence Repeats) [31], and SNP (Single Nucleotide Polymorphism) [32]. Among these, SSR markers are regarded as the most reliable due to their high polymorphism, with repeat motifs typically ranging from 1 to 6 base pairs [33,34,35]. Their widespread distribution across the genome and uniform coverage make SSRs a highly informative tool for various genetic analyses [36]. They are extensively employed in studies of biotic and abiotic stress responses, linkage map construction, analysis of genetic relationships among germplasm populations, and assessment of genetic diversity in Juglans regia [31,37,38,39,40,41,42,43,44,45]. Characterized by high polymorphism levels, co-dominant inheritance, and locus-specificity, SSR markers are invaluable for detailed genetic characterization.
Investigating the genetic variability of walnut using molecular techniques is a crucial step toward establishing effective breeding strategies, understanding population structure, and elucidating genetic relationships among germplasm from different regions. Despite the availability of walnut genetic resources within Kazakhstan, these resources remain largely underexplored, limiting their potential for breeding and conservation efforts. They represent a valuable foundation for developing locally adapted varieties, yet a comprehensive genetic profile of these resources is currently lacking, which hampers their optimal utilization.
The aim of this research is to evaluate the genetic diversity and population structure of Juglans regia accessions collected from the southern and southeastern regions of Kazakhstan, utilizing molecular markers to generate detailed insights into their genetic relationships and variability.
2. Materials and Methods
2.1. Plant Materials
The plant material for this study was collected from the primary walnut-growing regions of Kazakhstan, specifically from the Turkestan and Almaty regions, in 2024. A total of 75 samples were gathered through field collection across eight different sites in the southern and southeastern parts of the country, including four locations within Turkestan region and four within Almaty region. The sampling comprised 48 specimens from Turkestan and 27 from Almaty regions. Detailed information regarding the collected samples is provided in Table 1, while their sampling locations are illustrated in Figure 1.
The freshly collected leaves were placed in self-sealing bags and stored in a cooler with ice. After freezing in liquid nitrogen, the samples were kept in a freezer at a temperature of −80°C to ensure preservation until further analysis.
The plant material was collected from trees growing in the vicinity of settlements and in mountainous areas. The populations consisted of mature Juglans regia trees, all of which originated from open-pollinated seedlings derived from orchards or home gardens. From each population, between 15 and 40 individual trees were sampled, depending on availability (total N = 102 individuals), to ensure representative coverage [46]. Tree density and accessibility varied across different sites. The sampled walnut trees were grouped into six geographic populations: three in Turkestan region (Sairam-Ugam, Mashat, and Sarkyrama) and three in Almaty region (Almaty city, Baidibek Bi, and Esik) (Table 1 and Figure 1).
Figure 2 displays photographs of selected Juglans regia samples collected during field expeditions in the Turkestan and Almaty regions.
2.2. DNA Extraction and Molecular Analysis Using SSR Markers
For molecular genetic analysis in 2024, five young leaves were collected from each walnut tree during the active growth phase. Samples were immediately preserved in silica gel to prevent polyphenol oxidation and homogenized in liquid nitrogen using an optimized CTAB protocol with modern modifications [47]. DNA quality evaluation and quantification was performed by using a SmartSpecTMPlus Spectrophotometer (Bio-RAD Laboratories, Inc., USA). DNA concentration was standardized to 30 ng/µl for PCR using Tris-EDTA buffer (pH 8.0). The protocol was specifically adapted for tannin-rich tissues, as confirmed by successful SSR marker amplification in downstream experiments. To minimize fragmentation, lysis time was reduced to 30 min, and temperature was maintained at 65°C throughout the process. DNA was quantified spectrophotometrically (260 and 280 nm) and visualized on 1.0% agarose gels stained with ethidium bromide. Eight SSR molecular markers (WGA001, WGA027, WGA042, WGA118, WGA009, WGA202, WGA276, and WGA376) were chosen for the genotypic characterization of walnut cultivars based on a comprehensive review of existing scientific studies [48,49]. The selection criteria included their even distribution across the walnut genome, high polymorphism levels (informative content), and consistent amplification performance in previous research. These markers were specifically selected to ensure robust and reliable genotyping results. Detailed information regarding each primer, including sequence, annealing temperature, and expected amplicon size, is presented in Table 2.
The PCR reaction mixture (25 µL) consisted of 2.5 µL of genomic DNA (30 ng), 1 µL of each primer (1 pM/µL) (Sigma-Aldrich, St. Louis, MO, USA), 2.5 µL of a dNTP mixture (2.5 mM; dATP, dCTP, dGTP, and dTTP in aqueous solution) (ZAO Sileks, Moscow, Russia), 2.5 µL of MgCl₂ (25 mM), 0.2 µL of Taq polymerase (5 U/µL) (ZAO Sileks, Russia), 2.5 µL of 10× PCR buffer, and 12.8 µL of ddH₂O. Amplification products were separated using TBE buffer (45 mM Tris-borate, 1 mM EDTA, pH 8), with ethidium bromide added for visualization. A 100 bp DNA ladder (Fermentas, Vilnius, Lithuania) was used as a molecular size marker to estimate fragment lengths. PCR was performed on a Bio-Rad T100™ Thermal Cycler (Bio-RAD, Hercules, CA, USA) with the following conditions: an initial denaturation step at 94°C for 5 minutes; 35 cycles consisting of denaturation at 94°C for 30 seconds and extension at 72°C for 30 seconds; and a final extension step at 72°C for 5 minutes.
Amplification products were analyzed using a 2% agarose gel, with TBE buffer (45 mM Tris-borate, 1 mM EDTA, pH 8) employed for separation. Ethidium bromide was added to visualize the DNA fragments. A 100 bp DNA ladder (Fermentas, Vilnius, Lithuania) served as a molecular size reference to estimate fragment lengths. The Gel Documentation System (Gel Doc XR+, Bio-RAD, Hercules, CA, USA) was utilized to capture and analyze the results. Each sample was subjected to three independent tests to ensure reliability and reproducibility of the data.
2.3. Data Analysis
For the analysis of SSR data, GenAlEx 6.5 software was utilized [50,51]. Genetic diversity within the studied walnut populations was evaluated using several key parameters: the total number of observed alleles (NA), observed heterozygosity (Ho), expected heterozygosity (He), effective number of alleles (Ne), Shannon’s information index (I). Further genetic analysis included an Analysis of Molecular Variance (AMOVA). The resulting genetic distance matrix was used for Principal Coordinate Analysis (PCoA) and to construct a UPGMA (Unweighted Pair Group Method with Arithmetic Mean) dendrogram using PAST 4.03 software [52]. This approach enabled the identification of genetic clusters within the population and provided insights into the extent of genetic admixture among them.
3. Results
3.1. Analysis of the Genetic Diversity of Juglans Regia
Using eight pairs of SSR markers, the genetic diversity and population structure of 75 Juglans regia genotypes were assessed, along with an analysis of polymorphism at SSR loci. DNA profiles specific to the studied walnut samples were generated. The results of the genetic diversity analysis are summarized in Table 3. A total of 47 alleles were identified across all markers, with an average of 5.875 alleles per locus. The number of alleles per locus (Na) ranged from 2, observed at loci WGA027 and WGA042, to 10 at locus WGA276.
The effective number of alleles (Ne) ranged from 1.930 to 6.664, with an average of 4.219 (Table 3). Expected heterozygosity (He) varied between 0.482 to 0.850 averaging 0.704. Observed heterozygosity (Ho), in turn, ranged from 0.238 to 0.717, with an overall mean of 0.547. The highest observed heterozygosity and expected heterozygosity was detected at locus WGA276. The Shannon diversity index ranged from oт 0.675 to 2.095, with an average value of 1.444. The marker WGA027, exhibiting the lowest polymorphism, also showed the smallest Shannon index (I), whereas the most polymorphic marker, WGA276, demonstrated the highest diversity index.
The selected walnut samples from six locations, representing two geographic regions (Turkestan and Almaty regions), were compared based on several genetic parameters. These included the number of alleles (Na), the effective number of alleles (Ne), the Shannon diversity index (I), observed heterozygosity (Ho), and expected heterozygosity (He). The analysis of genetic diversity across the six populations is summarized in Table 4.
The study results indicated that the average number of alleles per locus ranged from 1,125 to 4,625. The effective number of alleles (Ne) varied between 1.075 to 3.658. Heterozygosity parameters showed that expected heterozygosity (He) ranged from 0.109 to 0.683, while observed heterozygosity (Ho) fluctuated from 0.063 to 0.697. The Shannon diversity index spanned from 0.157 дo 1.307. The lowest levels of genetic diversity were observed in population 5 (pop5) from Baidibek Bi in Almaty region, as evidenced by their low values of Na, Ne, and Shannon index. Conversely, the highest genetic diversity was recorded in population 1 (pop1) from Sairam-Ugam in Turkestan region.
3.2. Population Structure Analysis
For the subsequent genetic analysis, the population structure of 75 Juglans regia genotypes was analysed (Figure 3 and Figure 4). The STRUCTURE software evaluated the likelihood of classifying individuals into different numbers of clusters. The optimal number of groups was identified based on the ∆K statistic.
The analysis utilized a model incorporating admixture and correlated allele frequencies, with the ∆K method relying on the rate of change of the second-order likelihood function [54]. A clear peak was detected at K = 2, confirming the most probable number of genetic clusters, as shown in Figure 3.
Based on the amplification of 8 SSR markers across 75 walnut samples, a genetic distance was calculated. The genotypes were grouped into two clusters. The first cluster mainly included individuals numbered 20, 32, 41, 21, 25, 30, 48, 44, 22, 16, 7, 4, 40, 36, 15, 42, 38, 27, 34, 6, 12, 23, 29, 2, 8, 11, 9, 33, 18, 5, 35, 45, 10, 3, 19, 43, 13 and 39 from Turkestan region (populations pop 1 (S-U), pop 2 (MA), pop 3 (SA). The average genetic distance (expected heterozygosity) within this cluster was 0.486 (Figure 4). The second cluster comprised genotypes numbered 71, 73, 59, 67, 62, 52, 72, 70, 14, 28, 37, 24, 57, 26, 46, 68, 50, 49, 66, 69, 58, 53, 47, 65, 74, 75, 54, 56, 55, 51, 63, 31, 17, 1, 60, 61 and 64, collected from Almaty region and Turkestan region (populations: pop 1 (S-U), pop 2 (MA), and pop 3 (SA), pop 4 (AL), pop 5 (BA), and pop 6 (ES)). The average genetic distance (expected heterozygosity) within this cluster was 0.747 (Figure 4).
3.3. Cluster Analysis Using UPGMA and Principal Coordinates Analysis (PCoA
Genetic distances among Juglans regia groups based on origin were assessed using cluster analysis with the UPGMA (Unweighted Pair Group Method with Arithmetic Mean) and Principal Coordinates Analysis (PCoA). The list of studied samples, along with their population designations and geographical locations, is provided in Table 1 and Table 4. The dendrogram of 75 Juglans regia genotypes, generated through cluster analysis using the UPGMA method, revealed that the studied material formed subgroups of individuals with distinct differences, as illustrated in Figure 5. The analysis divided the samples into two main branches, further subdivided into six sub-branches. The first branch comprised 37 genotypes of Juglans regia (49.3%), including populations from Almaty and Turkestan region (S-U pop 1, MA pop 2, SA pop 3, AL pop 4, BA pop 5, and ES pop 6). The second branch included 38 genotypes from Turkestan region (50.7%), with 24 samples from Sairam-Ugam and 14 from Mashat and Sarkyrama. The results indicated that genotypes from the same geographic origin did not consistently cluster together (Figure 5). For example, the first sub-branch of the first main branch contained not only genotypes from Almaty but also ten samples (S-U-14, S-U-24, S-U-17, MA-37, SA-47, S-U-26, SA-46, S-U-28, MA-31, S-U-1) originating from Turkestan region.
The results of the Principal Coordinates Analysis (PCoA) confirmed the findings of the UPGMA cluster analysis. Based on PCoA, two distinct groups of samples were identified (Figure 6). The first group comprised genotypes from Almaty region and Turkestan region (pop1-pop6 S-U; MA; SA; AL; BA; ES). The second group included samples from Turkestan region (pop1-pop3, SU; MA; SA).
Therefore, the analysis of the 75 Juglans regia samples demonstrated considerable genetic variation among genotypes from different regions, highlighting a broad genetic diversity present within the studied collection.
4. Discussion
Walnut (Juglans regia) populations exhibit a high level of genetic diversity. Identifying valuable traits and understanding their genetic variability are key priorities for breeding programs. Molecular markers serve as powerful tools for analysing genome structure and genetic variation. Among these, SSR (microsatellite) markers are most frequently used in walnut diversity studies. As a result of this research, a comparative analysis of microsatellite polymorphism was conducted among local walnut populations from different geographic regions of Kazakhstan.
Our findings confirm that Juglans regia populations in Kazakhstan exhibit a high level of genetic diversity, consistent with data reported in international studies. The average number of alleles per locus (Na = 5.87) and the effective number of alleles (Ne = 4.21) align closely with results from Ebrahimi et al. (2011), who analysed 31 samples and identified an average of 5.10 alleles per locus [55]. Similar values have been observed in Turkish studies, such Orhan et al. (2020), where the mean number of alleles reached 6,43, indicating a richer gene pool within Turkish populations [56]. These differences are likely driven by historical factors, including long-term breeding activities and migration of genetic material across regions. Such variability underscores the importance of conserving diverse genetic resources to enhance breeding strategies and adapt to environmental challenges. In recent studies, Kairova et al. (2025) evaluated the genetic diversity and population structure of wild and cultivated Juglans regia in Kazakhstan. Their results revealed that wild walnut populations in Kazakhstan form distinct genetic clusters, refecting historical isolation and adaptation, while cultivars exhibit a more dispersed genetic profle, indicative of admixture and artifcial selection. They showed, that Inbreeding coefcients (F) were highest in wild Kazakh walnuts (0.17), indicating genetic bottlenecks and habitat fragmentation, whereas local genotypes showed lower inbreeding levels (0.08), suggesting a more diverse genetic background [20].
Regarding heterozygosity metrics, our results reveal a significant discrepancy between the expected heterozygosity (He = 0.70) and the observed heterozygosity (Ho = 0.54). Similar patterns of heterozygote deficiency have been reported in studies by Bernard et al. (2018), Plugatar et al. (2023), and Aradhya et al. (2010), where low level of heterozygosity was consistently observed. As proposed by Manthos et al. (2023), such disparities may indicate the influence of inbreeding, limited gene flow, or intensive selection pressures [57,58,59,60]. In the context of our study, these findings may reflect a historical isolation of populations, emphasizing the need for further investigation into their demographic history and gene flow dynamics. Understanding these factors is crucial for developing strategies to maintain genetic diversity and improve breeding programs.
The Shannon information index (I), reaching 2.095 at locus WGA276, indicates a high level of genetic variability within the studied populations. Similar values have been reported in the study by Manthos et al. (2023) and Suprun et al. (2025), confirming the broad applicability and high informativeness of the selected markers [60,61]. However, the level of polymorphism at the WGA276 locus in our studies was higher than in the study by Shamlu et al. (2018), the average number of alleles per locus was about 7.9 [62]. This suggests a more diverse gene pool in the Kazakh populations, which is a critical factor to consider in conservation strategies and breeding programs aimed at maintaining and utilizing genetic diversity effectively.
Regarding population structure, the identification of two clusters that do not correlate with geographic location aligns with findings from previous studies, such as Bernard et al (2020) and Plugatar et al. (2023), which also reported high levels of within-population differentiation and a lack of clear geographic structuring [58,63]. This pattern may be attributed to historical migration events and interregional exchanges of genetic material, highlighting the importance of incorporating analyses of migration dynamics and historical demographic processes in future research. Such insights are essential for understanding the mechanisms shaping genetic diversity and for developing effective conservation and breeding strategies.
Analysis of population structure using STRUCTURE, UPGMA, and PCoA methods identified two major genetic clusters that do not align with the geographic origins of the samples. This suggests a high level of genetic differentiation within the studied populations. The presence of these unique genetic forms offers valuable opportunities for their integration into breeding programs aimed at improving winter hardiness, pest and disease resistance, and overall agronomic traits. Harnessing this genetic diversity can facilitate the development of superior cultivars adapted to local conditions and contribute to the broadening of the genetic base of cultivated walnuts
Overall, our findings indicate that Kazakh populations of Juglans regia possess a rich genetic resource, providing a solid foundation for future breeding initiatives. However, the low observed heterozygosity highlights the need for more detailed investigations into factors that may constrain genetic variability, such as inbreeding, selection pressures, and migration dynamics. Understanding these processes is crucial for developing effective conservation strategies and optimizing the utilization of the gene pool for sustainable breeding and preservation efforts.
5. Conclusions
This study confirmed the high genetic diversity of Kazakh Juglans regia populations, highlighting their potential for use in breeding programs. The pronounced polymorphism of SSR markers, particularly at locus WGA276, underscores their effectiveness in assessing intrapopulation variation. Population structure analysis revealed two main genetic clusters primarily linked to geographic origin, although significant internal differentiation suggests long-term isolation and local adaptation. The population from Turkestan region possesses the most diverse gene pool and represents a valuable resource for developing resilient and high-yielding cultivars. These findings emphasize the importance of conserving and strategically utilizing genetic diversity to enhance the adaptive capacity of the species, forming a foundation for future conservation and gene pool expansion initiatives.
Author Contributions
MN: Conceptualization, Data curation, Formal Analysis, Investigation, Methodology, Resources, Validation, Software, Visualization, Writing – original draft, Writing – review & editing. AK: Project administration, Data curation, Formal Analysis, Funding acquisition, Validation, Writing – original draft, Writing – review & editing. MK: Data curation, Formal Analysis, Investigation, Methodology, Validation, Writing – original draft. ZHK: Data curation, Formal Analysis. KB: Investigation, Methodology. AB: Formal Analysis, Investigation, AK: Validation, Formal Analysis, KM: Data curation, Investigation. KA: Methodology, Writing – original draft.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This research has been funded by the Committee of Science of the Ministry of Science and Higher Education of the Republic of Kazakhstan (Grant No.BR21882024 "Study of biodiversity and development of techniques for ex situ conservation of genetic resources of fruit and nut plants”).
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- Halvorsen, B. L.; Carlsen, M. H.; Phillips, K. M.; Bøhn, S. K.; Holte, K.; Jacobs, D. R.; Blomhoff, R. Content of redox-active compounds (ie, antioxidants) in foods consumed in the United States. Am. J. Clin. Nutr. 2006, 84 (1), 95–135. [CrossRef]
- Kornsteiner, M.; Wagner, K.-H.; Elmadfa, I. Tocopherols and total phenolics in 10 different nut types. Food Chem. 2006, 98 (2), 381–387. [CrossRef]
- Hayes, D.; Angove, M. J.; Tucci, J.; Dennis, C. Walnuts (Juglans regia) chemical composition and research in human health. Crit. Rev. Food Sci. Nutr. 2016, 56 (8), 1231–1241. [CrossRef]
- Ros, E.; Izquierdo-Pulido, M.; Sala-Vila, A. Beneficial effects of walnut consumption on human health: Role of micronutrients. Curr. Opin. Clin. Nutr. Metab. Care 2018, 21 (6), 498–504. [CrossRef]
- Liu, X.; Guasch-Ferré, M.; Tobias, D. K.; Li, Y. Association of walnut consumption with total and cause-specific mortality and life expectancy in U.S. adults. Nutrients 2021, 13 (8), 2699. [CrossRef]
- Poggetti, L.; Ferfuia, C.; Chiabà, C.; Testolin, R.; Baldini, M. Kernel oil content and oil composition in walnut (Juglans regia L.) accessions from north-eastern Italy. J. Sci. Food Agric. 2018, 98 (3), 955–962. [CrossRef]
- Geng, S.; Ning, D.; Ma, T.; Chen, H.; Zhang, Y.; Sun, X. Comprehensive analysis of the components of walnut kernel (Juglans regia L.) in China. J. Food Qual. 2021, 2021, 9302181. [CrossRef]
- Wang, P.; Zhong, L.; Yang, H.; Zhang, J.; Hou, X.; Wu, C.; Zhang, R.; Cheng, Y. Comprehensive comparative analysis of lipid profile in dried and fresh walnut kernels by UHPLC-Q-exactive orbitrap/MS. Food Chem. 2022, 386, 132706. [CrossRef]
- Sanchez-Gonzalez, C.; Ciudad, C. J.; Noé, V.; Izquierdo-Pulido, M. Health benefits of walnut polyphenols: An exploration beyond their lipid profile. Crit. Rev. Food Sci. Nutr. 2017, 57 (15), 3373–3383. [CrossRef]
- Gandev, S. Budding and grafting of the walnut (Juglans regia L.) and their effectiveness in Bulgaria (Review). Bulgar. J. Agri. Sci. 2007, 13 (5), 683–689.
- Martinez, M. L.; Labuckas, D. O.; Lamarque, A. L.; Maestri, D. M. Walnut (Juglans regia L.): genetic resources, chemistry, by-products. J. Sci. Food Agric. 2010, 90 (12), 1959–1967. [CrossRef]
- McGranahan, G.; Leslie, C. Walnut. In Fruit Breeding; Badenes, M., Byrne, D., Eds.; Handbook of Plant Breeding; Springer: Boston, MA, USA, 2012; pp 827–846.
- Pollegioni, P.; Woeste, K. E.; Chiocchini, F.; Del Lungo, S.; Olimpieri, I.; Tortolano, V.; et al. Ancient humans influenced the current spatial genetic structure of common walnut populations in Asia. PLoS ONE 2015, 10 (7), e0135980. [CrossRef]
- Vahdati, K. Traditions and folks for walnut growing around the Silk Road. Acta Hortic. 2014, 1032, 19–24. [CrossRef]
- Zohary, D.; Hopf, M.; Weiss, E. Domestication of Plants in the Old World: The Origin and Spread of Domesticated Plants in Southwest Asia, Europe, and the Mediterranean Basin, 4th ed.; Oxford University Press: Oxford, UK, 2012; p 149.
- FAOSTAT. Food and Agriculture Organization of the United Nations. FAO Statistics Division. 2018. Available online: http://faostat.fao.org/site/567/default.aspx#ancor (accessed on [date]).
- Vahdati, K.; Arab, M. M.; Sarikhani, S.; Sadat-Hosseini, M.; Leslie, C. A.; Brown, P. J. Advances in Persian walnut (Juglans regia L.) breeding strategies. In Advances in Plant Breeding Strategies: Nut and Beverage Crops; Springer: Cham, Switzerland, 2019; pp 401–472. [CrossRef]
- FAO. World Food and Agriculture - Statistical Yearbook 2023; Rome, 2023. [CrossRef]
- Akça, Y.; Yusupov, B. Y.; Erdenov, M.; Vahdati, K. Exploring of walnut genetic resources in Kazakhstan and evaluation of promising selections. Int. J. Hortic. Sci. Technol. 2020, 7 (1), 93–102. [CrossRef]
- Kairova, G.; Taskuzhina, A.; Yanin, K.; Ismagulova, E.; Oleichenko, S.; Sarshayeva, M.; Sapakhova, Z.; Gritsenko, D. First evaluation of genetic diversity and population structure of wild and cultivated Juglans regia in Kazakhstan. Genet. Resour. Crop Evol. 2025, 72 (4), 1755–1771. [CrossRef]
- Oleichenko, S. N.; Yegizbayeva, T. K.; Apushev, A. K.; Nusipzhanov, N. S. Assessment of promising local walnut forms for the South and South-East of Kazakhstan. Rep. Natl. Acad. Sci. Repub. Kazakhstan 2020, 333, 27–34. [CrossRef]
- Bakhytuly, K.; Kokhmetova, A. M.; Umirzakova, A. T.; Kuliev, A. S.; Kumbarbayeva, M. T.; Keishilov, Z. S. Monitoring of the distribution of Juglans regia L. in the southern and south-eastern regions of Kazakhstan and morphological study of walnut fruits. Izd. Natigeler 2024, 2, 238–248. [CrossRef]
- Terletskaya, N. V.; Shadenova, E. A.; Litvinenko, Y. A.; Ashimuly, K.; Erbay, M.; Mamirova, A.; Nazarova, I.; Meduntseva, N. D.; Kudrina, N. O.; Korbozova, N. K.; et al. Influence of cold stress on physiological and phytochemical characteristics and secondary metabolite accumulation in microclones of Juglans regia L. Int. J. Mol. Sci. 2024, 25 (9), 4991. [CrossRef]
- Kushnarenko, S. V.; Rymkhanova, N. K.; Aralbayeva, M. M.; Romadanova, N. V. In vitro cold acclimation is required for successful cryopreservation of Juglans regia L. shoot tips. CryoLetters 2023, 44 (3), 240–248. [CrossRef]
- Kushnarenko, S.; Aralbayeva, M.; Rymkhanova, N.; et al. Initiation pretreatment with Plant Preservative Mixture™ increases the percentage of aseptic walnut shoots. In Vitro Cell. Dev. Biol. - Plant 2022, 58 (6), 964–971. [CrossRef]
- Kairov, G.; Ismagulova, E.; Oleichenko, S.; Suleimanova, G.; Basim, H.; Sarshabayeva, M. Resistance of walnut varieties to bacteriosis caused by Pantoea agglomerans in the southern fruit-growing zone of Kazakhstan. Izdenister Natigeler 2024, 2, 238–248. [CrossRef]
- Çarpar, H.; Sertkaya, G. First report of Crepis phyllody disease associated with phytoplasma in Crepis foetida in a walnut orchard in Turkey. J. Plant Dis. Prot. 2023, 130 (3), 177–180. [CrossRef]
- Zhao, P.; Zhao, G.-F.; Zhang, S.-X.; Zhou, H.-J.; Hu, Y.-H.; Woeste, K. E. RAPD derived markers for separating Manchurian walnut (Juglans mandshurica) and Japanese walnut (J. ailantifolia) from close congeners. J. Syst. Evol. 2014, 52 (2), 101–111. [CrossRef]
- Ma, Q.; Zhang, J.; Pei, D. Genetic analysis of walnut cultivars in China using fluorescent amplified fragment length polymorphism. J. Am. Soc. Hortic. Sci. 2011, 136 (6), 422–428. [CrossRef]
- Kafkas, S.; Ozkan, H.; Sutyemez, M. DNA polymorphism and assessment of genetic relationships in walnut genotypes based on AFLP and SAMPL markers. J. Am. Soc. Hortic. Sci. 2005, 130 (4), 585–590. [CrossRef]
- Itoo, H.; Shah, R. A.; Qurat, S.; Jeelani, A.; Khursheed, S.; Bhat, Z. A.; Mir, M. A.; Rather, G. H.; Zargar, S. M.; Shah, M. D.; et al. Genome-wide characterization and development of SSR markers for genetic diversity analysis in Northwestern Himalayas walnut (Juglans regia L.). 3 Biotech 2023, 13 (3), 136. [CrossRef]
- Arab, M. M.; Brown, P. J.; Abdollahi-Arpanahi, R.; Sohrabi, S. S.; Askari, H.; Aliniaeifard, S.; Mokhtassi-Bidgoli, A.; Mesgaran, M. B.; Leslie, C. A.; Marrano, A.; et al. Genome-wide association analysis and pathway enrichment provide insights into the genetic basis of photosynthetic responses to drought stress in Persian walnut. Hortic. Res. 2022, 9, uhac124. [CrossRef]
- Grover, A.; Aishwarya, V.; Sharma, P. C. Biased distribution of microsatellite motifs in the rice genome. Mol. Genet. Genom. 2007, 277 (5), 469–480. [CrossRef]
- Taheri, S.; Lee Abdullah, T.; Yusop, M. R.; Hanafi, M. M.; Sahebi, M.; Azizi, P.; Shamshiri, R. R. Mining and development of novel SSR markers using next generation sequencing (NGS) data in plants. Molecules 2018, 23, 399. [CrossRef]
- Nie, X.; Wang, Z.; Liu, N.; Song, L.; Yan, B.; Xing, Y.; Zhang, Q.; Fang, K.; Zhao, Y.; Chen, X.; et al. Fingerprinting 146 Chinese chestnut (Castanea mollissima Blume) accessions and selecting a core collection using SSR markers. J. Integr. Agric. 2021, 20, 1277–1286. [CrossRef]
- Kalia, R. K.; Rai, M. K.; Kalia, S.; Singh, R.; Dhawan, A. K. Microsatellite markers: An overview of the recent progress in plants. Euphytica 2011, 177, 309–334. [CrossRef]
- Ali Khan, M.; Shahid Ul, I.; Mohammad, F. Extraction of natural dye from walnut bark and its dyeing properties on wool yarn. J. Nat. Fibers 2016, 13, 458–469. [CrossRef]
- Bernard, A.; Barreneche, T.; Donkpegan, A.; Lheureux, F.; Dirlewanger, E. Comparison of structure analyses and core collections for the management of walnut genetic resources. Tree Genet. Genomes 2020, 16, 1–14. [CrossRef]
- Bernard, A.; Marrano, A.; Donkpegan, A.; Brown, P. J.; Leslie, C. A.; Neale, D. B.; Lheureux, F.; Dirlewanger, E. Association and linkage mapping to unravel genetic architecture of phenology-related traits and lateral bearing in Persian walnut (Juglans regia L.). 2019.
- Shah, U. N.; Mir, J. I.; Ahmed, N.; Fazili, K. M. Assessment of germplasm diversity and genetic relationships among walnut (Juglans regia L.) genotypes through microsatellite markers. J. Saudi Soc. Agric. Sci. 2018, 17, 339–350. [CrossRef]
- Shah, R. A.; Baksi, P.; Jasrotia, A.; Bhat, D. J. I.; Gupta, R.; Bakshi, M. Genetic diversity of walnut (Juglans regia L.) seedlings through SSR markers in north-western Himalayan region of Jammu. Bangladesh J. Bot. 2020, 49, 1003–1012. [CrossRef]
- Pollegioni, P.; Woeste, K.; Chiocchini, F.; Del Lungo, S.; Ciolfi, M.; Olimpieri, I.; Tortolano, V.; Clark, J.; Hemery, G. E.; Mapelli, S.; et al. Rethinking the history of common walnut (Juglans regia L.) in Europe: its origins and human interactions. PLoS ONE 2017, 12, e0172541. [CrossRef]
- Doğan, Y.; Kafkas, S.; Sütyemez, M.; Akça, Y.; Türemiş, N. Assessment and characterization of genetic relationships of walnut (Juglans regia L.) genotypes by three types of molecular markers. Sci. Hortic. 2014, 168, 81–87. [CrossRef]
- Nickravesh, M. H.; Vahdati, K.; Amini, F.; Di Pierro, E. A.; Amiri, R.; Woeste, K.; Arab, M. M. Reliable propagation of Persian walnut varieties using SSR marker-based true-to-type validation. HortScience 2023, 58, 64–66. [CrossRef]
- Wambulwa, M. C.; Fan, P. Z.; Milne, R.; Wu, Z. Y.; Luo, Y. H.; Wang, Y. H.; Wang, H.; Gao, L. M.; Ye, L. J.; Jin, Y. C.; et al. Genetic analysis of walnut cultivars from Southwest China: Implications for germplasm improvement. Plant Divers. 2022, 44, 530–541. [CrossRef]
- Shamlu, F. Genetic diversity of walnut (Juglans regia L.) populations in Iran based on SSR markers. J. Nuts 2018, 9, 1–12.
- Rogers, S. O.; Bendich, A. J. Extraction of DNA from milligram amounts of fresh, herbarium and mummified plant tissues. Plant Mol. Biol. 1985, 5, 69–76. [CrossRef]
- Woeste, K.; Burns, R.; Rhodes, O.; Michler, C. Thirty polymorphic nuclear microsatellite loci from black walnut. J. Hered. 2002, 93, 58–60. [CrossRef]
- Dangl, G. S.; Woeste, K.; Aradhya, M. K.; Koehmstedt, A.; Simon, C.; Potter, D.; Leslie, C. A.; McGranahan, G. Characterization of 14 microsatellite markers for genetic analysis and cultivar identification of walnut. J. Am. Soc. Hortic. Sci. 2005, 130, 348–354. [CrossRef]
- Peakall, R.; Smouse, P. E. GenAlEx 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [CrossRef]
- Peakall, R.; Smouse, P. E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research—An update. Bioinformatics 2012, 28, 2537–2539. [CrossRef]
- Hammer, Ø.; Harper, D. A. T.; Ryan, P. D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9.
- Pritchard, J. K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [CrossRef]
- Ebrahimi, A.; Fatahi, R.; Zamani, Z. Analysis of genetic diversity among some Persian walnut genotypes (Juglans regia L.) using SSR markers. Sci. Hortic. 2011, 130, 146–151. [CrossRef]
- Orhan, E.; Eyduran, S. P.; Poljuha, D.; Akin, M.; Weber, T.; Ercisli, S. Genetic diversity detection of seed-propagated walnut (Juglans regia L.) germplasm from Eastern Anatolia using SSR markers. Horticulturae 2020, 32, 67–76. [CrossRef]
- Bernard, A.; Barreneche, T.; Lheureux, F.; Dirlewanger, E. Analysis of genetic diversity and structure in a worldwide walnut (Juglans regia L.) germplasm using SSR markers. PLoS ONE 2018, 13, e0208021. [CrossRef]
- Plugatar, Y. V.; Suprun, I. I.; Khokhlov, S. Yu.; Stepanov, I. V.; Al-Nakib, E. A. Comprehensive Agrobiological Assessment and Analysis of Genetic Relationships of Promising Walnut Varieties of the Nikitsky Botanical Gardens. Vavilov J. Genet. Breed. 2023, 27, 454–462. [CrossRef]
- Aradhya, M. K.; Woeste, K.; Velasco, D. Genetic Diversity, Structure and Differentiation in Cultivated Walnut (Juglans regia L.). Acta Hortic. 2010, 861, 127–132. [CrossRef]
- Manthos, I.; Sotiropoulos, T.; Karapetsi, L.; Ganopoulos, I.; Pratsinakis, E. D.; Maloupa, E.; Madesis, P. Molecular Characterization of Local Walnut (Juglans regia) Genotypes in the North-East Parnon Mountain Region of Greece. Int. J. Mol. Sci. 2023, 24, 17230. [CrossRef]
- Suprun, I.; Stepanov, I.; Anatov, D. Analysis of Genetic Diversity and Relationships of Local Walnut Populations in the Western Caspian Region of the North Caucasus. Horticulturae 2025, 11, 65. [CrossRef]
- Samlu, F.; Rezaei, M.; Lawson, S.; Ebrahimi, A.; Biabani, A.; Khan-Ahmadi, A. Genetic Diversity of Superior Persian Walnut Genotypes in Azadshahr, Iran. Physiol. Mol. Biol. Plants 2018, 24, 939–949. [CrossRef]
- Bernard, A.; Barreneche, T.; Lheureux, F.; Dirlewanger, E. SSR Genetic Diversity Assessment of the INRAE’s Walnut (Juglans spp.) Germplasm Collection. Acta Hortic. 2020, 1297, 387–394. [CrossRef]
Figure 1.
Distribution of Juglans regia and collection sites on the map of the southern (A) and southeastern (B) regions of Kazakhstan.
Figure 1.
Distribution of Juglans regia and collection sites on the map of the southern (A) and southeastern (B) regions of Kazakhstan.
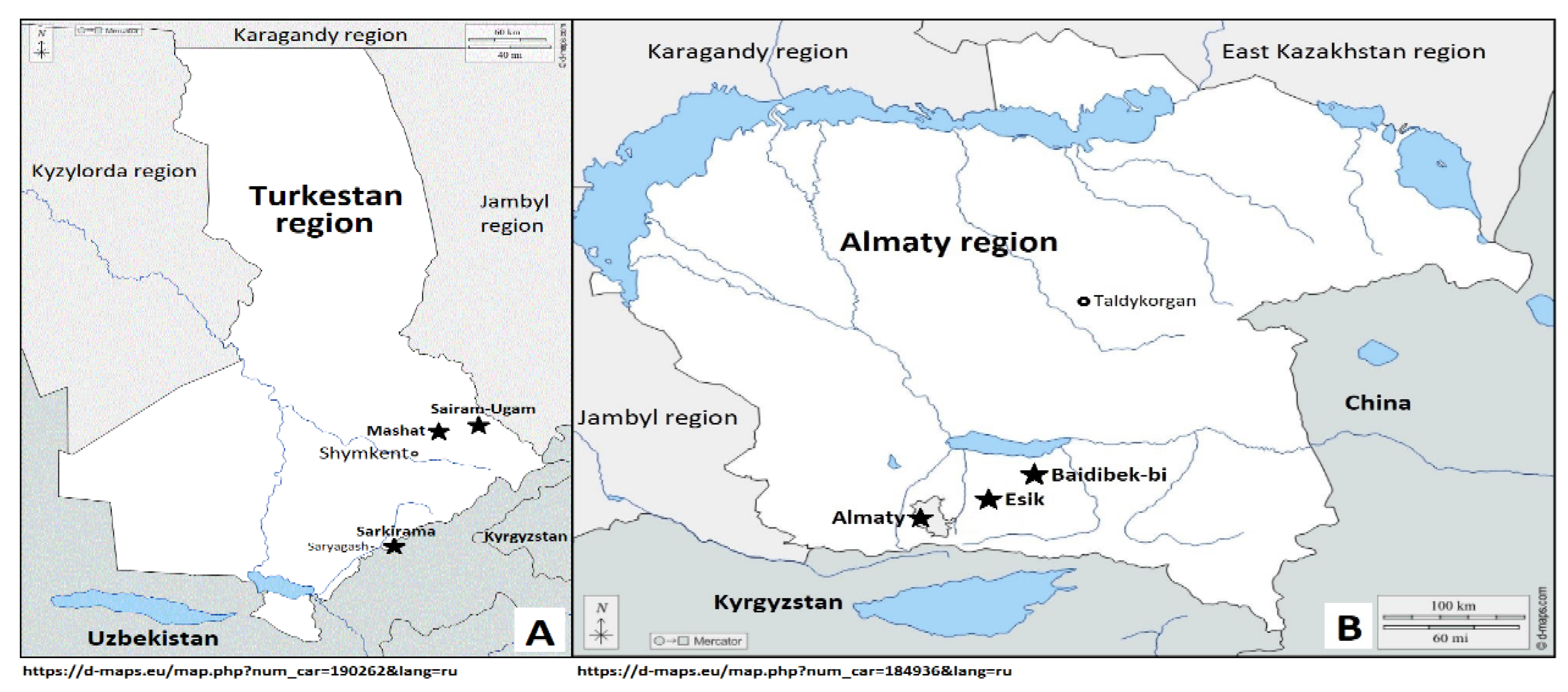
Figure 2.
Juglans regia L. trees from various populations: (a) Sairam-Ugam State National Nature Park, (b) Mashat v., (c) Sarkyrama v. in Turkestan Region; (d) Almaty city, (e) Baidibek-bi, and (f) Esik in Almaty Region.
Figure 2.
Juglans regia L. trees from various populations: (a) Sairam-Ugam State National Nature Park, (b) Mashat v., (c) Sarkyrama v. in Turkestan Region; (d) Almaty city, (e) Baidibek-bi, and (f) Esik in Almaty Region.

Figure 3.
Population structure of 75 Juglans regia genotypes based on 8 SSR markers, as determined by STRUCTURE analysis, assuming K = 2.
Figure 3.
Population structure of 75 Juglans regia genotypes based on 8 SSR markers, as determined by STRUCTURE analysis, assuming K = 2.
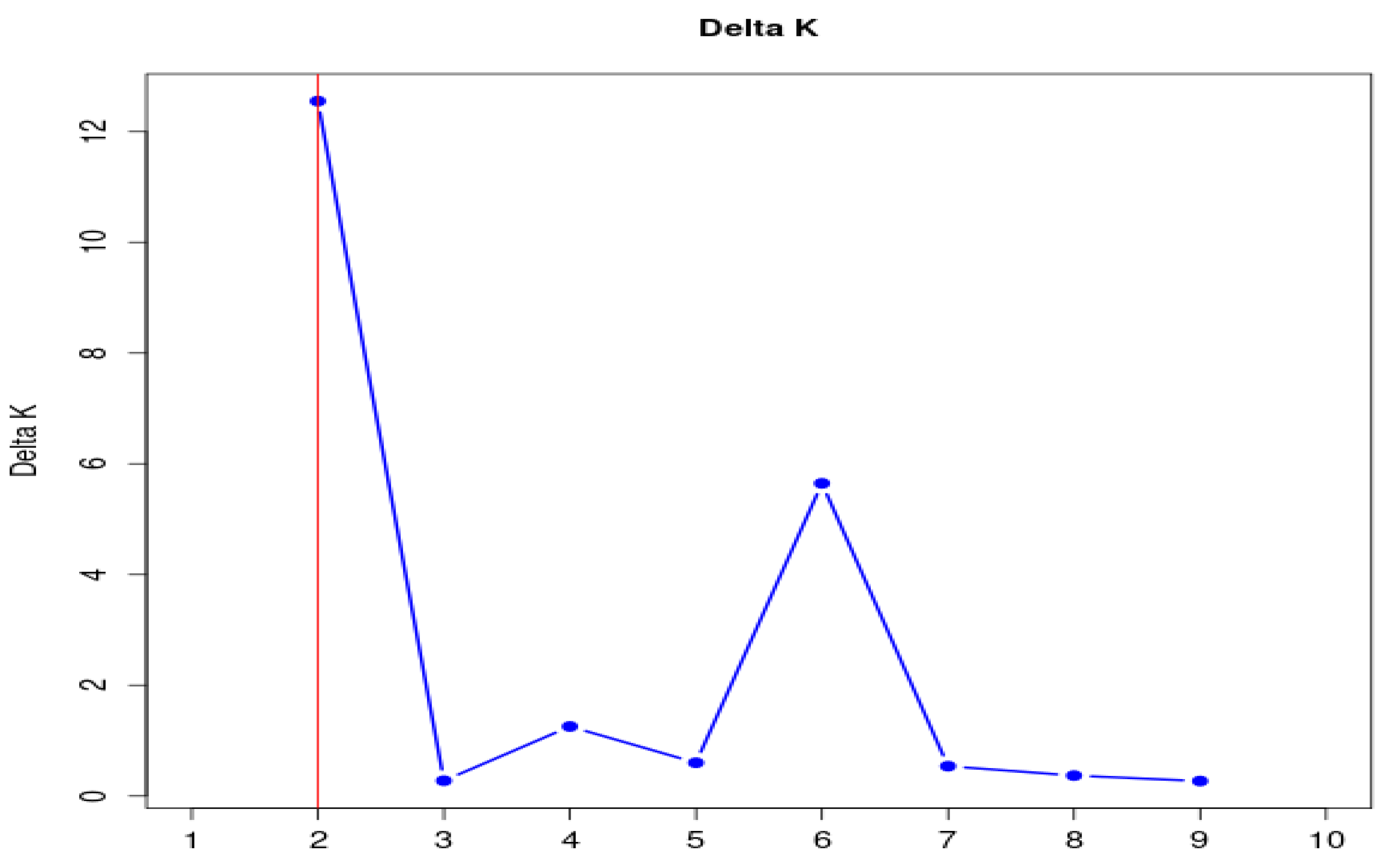
Figure 4.
Genetic structure of 75 Juglans regia genotypes analysed using 8 SSR markers. Each color (red, green) represents a different genetic cluster among the samples.
Figure 4.
Genetic structure of 75 Juglans regia genotypes analysed using 8 SSR markers. Each color (red, green) represents a different genetic cluster among the samples.
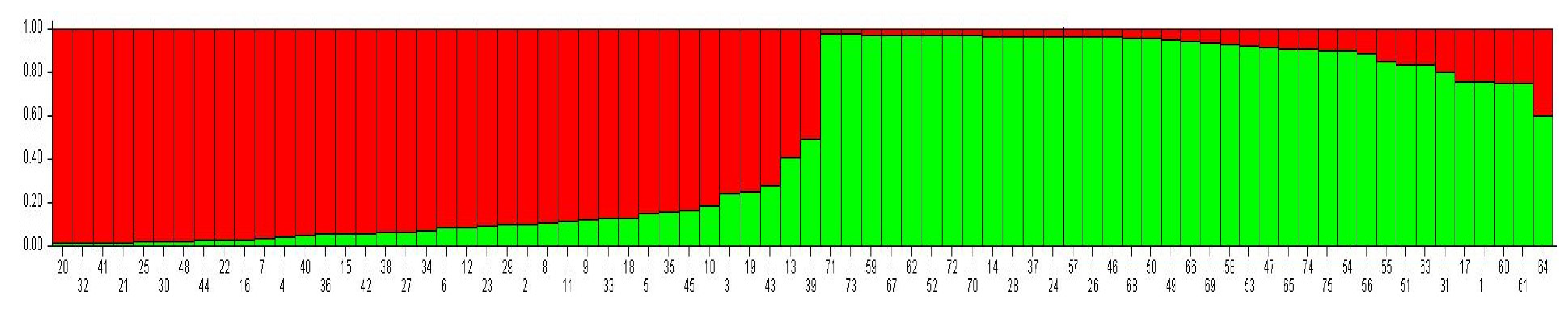
Figure 5.
Clustering analysis for 75 walnut individuals collected from the 6 locations of Kazakhstan using the UPGMA method.
Figure 5.
Clustering analysis for 75 walnut individuals collected from the 6 locations of Kazakhstan using the UPGMA method.
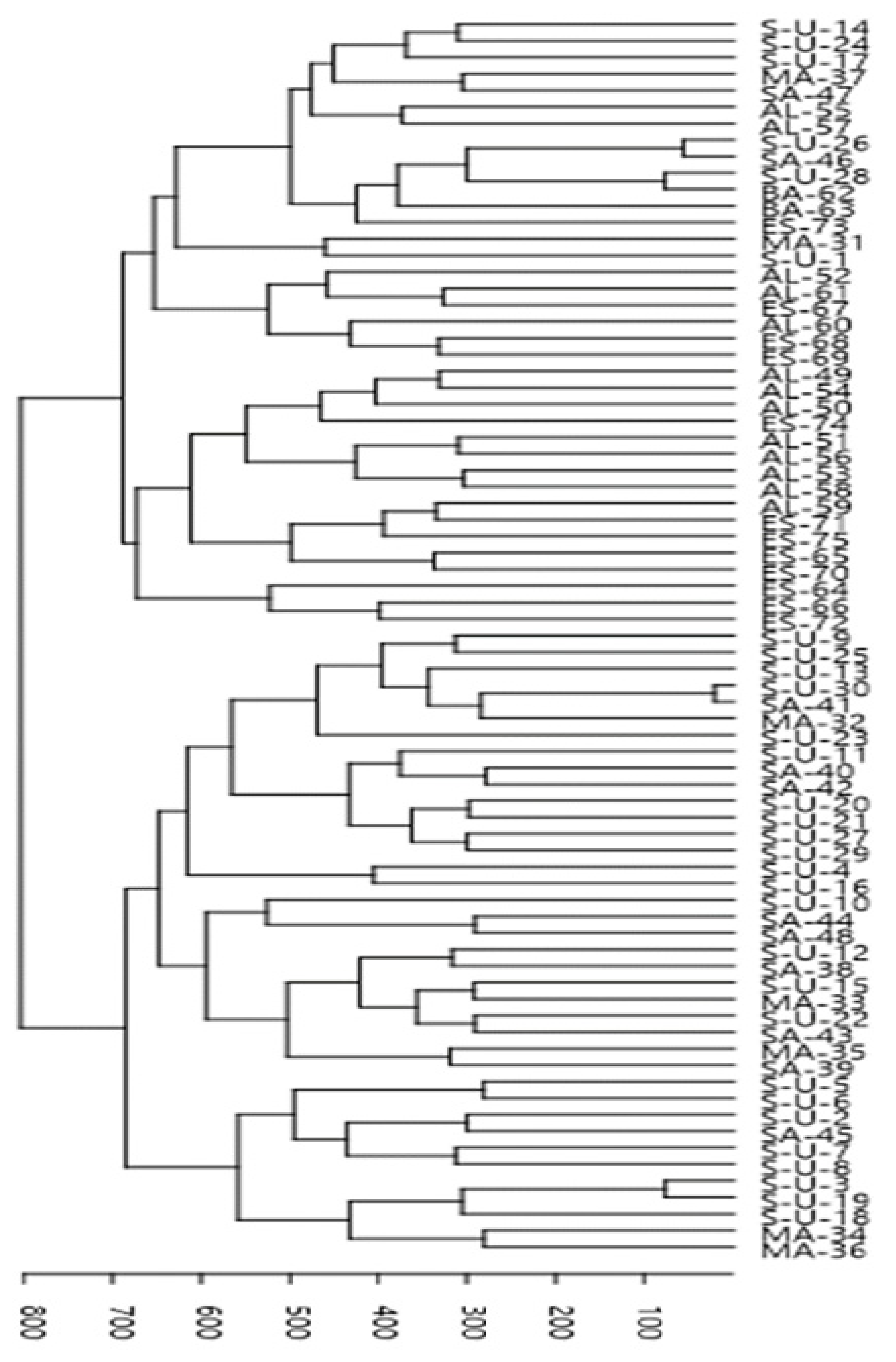
Figure 6.
Principal Coordinates Analysis (PCoA) plot of 75 Juglans regia genotypes, illustrating a clear overlap between the two clusters, which reflects low genetic distances among the studied individuals.
Figure 6.
Principal Coordinates Analysis (PCoA) plot of 75 Juglans regia genotypes, illustrating a clear overlap between the two clusters, which reflects low genetic distances among the studied individuals.
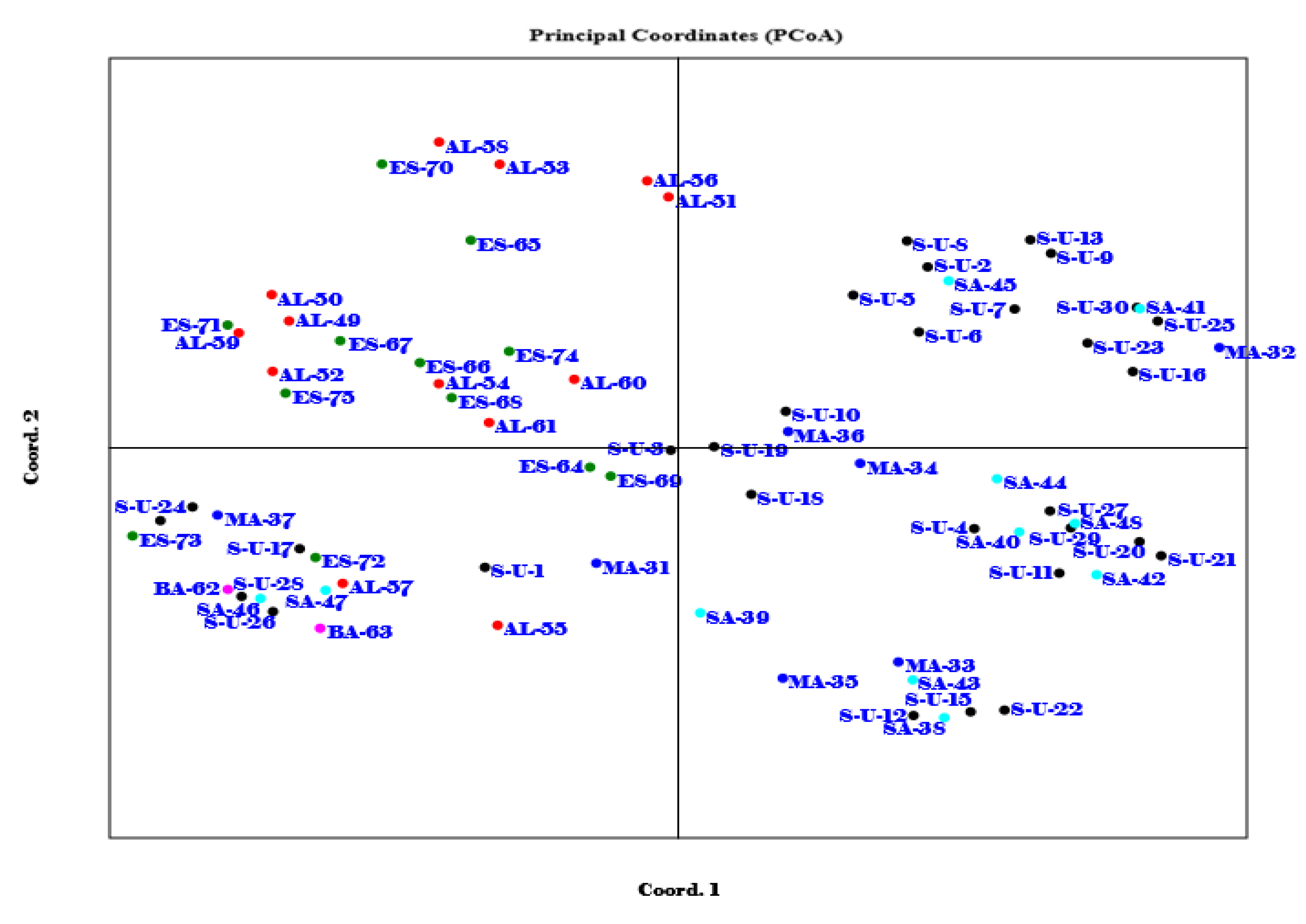

Table 1.
Collection sites: key geographic information for each sampling location in the southern and southeastern regions of Kazakhstan.
Table 1.
Collection sites: key geographic information for each sampling location in the southern and southeastern regions of Kazakhstan.
| No. | Geographic Region, Location | Population/Local Walnut Sample | Latitude | Longitude | Altitude (m) |
|---|---|---|---|---|---|
| 1 | Turkestan region, S-U | S-U-01 | N 42 39 995 | E 070 15 080 | 820 |
| 2 | Turkestan region, S-U | S-U-02 | N 42 39 995 | E 070 15 095 | 817 |
| 3 | Turkestan region, S-U | S-U-03 | N 42 39 992 | E 070 15 098 | 815 |
| 4 | Turkestan region, S-U | S-U-04 | N 42 39 991 | E 070 15 119 | 811 |
| 5 | Turkestan region, S-U | S-U-05 | N 42 39 988 | E 070 15 119 | 808 |
| 6 | Turkestan region, S-U | S-U-06 | N 42 39 982 | E 070 15 117 | 804 |
| 7 | Turkestan region, S-U | S-U-07 | N 42 39 981 | E 070 15 113 | 811 |
| 8 | Turkestan region, S-U | S-U-08 | N 42 39 980 | E 070 15 115 | 806 |
| 9 | Turkestan region, S-U | S-U-09 | N 42 39 975 | E 070 15 113 | 804 |
| 10 | Turkestan region, S-U | S-U-10 | N 42 39 986 | E 070 15 130 | 810 |
| 11 | Turkestan region, S-U | S-U-11 | N 42 39 995 | E 070 15 139 | 814 |
| 12 | Turkestan region, S-U | S-U-12 | N 42 40 001 | E 070 15 135 | 811 |
| 13 | Turkestan region, S-U | S-U-13 | N 42 40 010 | E 070 15 136 | 805 |
| 14 | Turkestan region, S-U | S-U-14 | N 42 399 95 | E 070 15 112 | 799 |
| 15 | Turkestan region, S-U | S-U-15 | N 42 39 955 | E 070 15 112 | 804 |
| 16 | Turkestan region, S-U | S-U-16 | N 42 39 985 | E 070 15 153 | 810 |
| 17 | Turkestan region, S-U | S-U-17 | N 42 39 994 | E 070 15 156 | 808 |
| 18 | Turkestan region, S-U | S-U-18 | N 42 39 999 | E 070 15 161 | 811 |
| 19 | Turkestan region, S-U | S-U-19 | N 42 40 020 | E 070 15 154 | 817 |
| 20 | Turkestan region, S-U | S-U-20 | N 42 40 021 | E 070 15 157 | 816 |
| 21 | Turkestan region, S-U | S-U-21 | N 42 40 017 | E 070 15 157 | 816 |
| 22 | Turkestan region, S-U | S-U-22 | N 42 40 039 | E 070 15 135 | 819 |
| 23 | Turkestan region, S-U | S-U-23 | N 42 40 041 | E 070 15 140 | 820 |
| 24 | Turkestan region, S-U | S-U-24 | N 42 40 051 | E 070 15 069 | 831 |
| 25 | Turkestan region, S-U | S-U-25 | N 42 40 049 | E 070 15 067 | 829 |
| 26 | Turkestan region, S-U | S-U-26 | N 42 40 041 | E 070 15 073 | 828 |
| 27 | Turkestan region, S-U | S-U-27 | N 42 39 994 | E 070 15 082 | 821 |
| 28 | Turkestan region, S-U | S-U-28 | N 42 39 987 | E 070 15 088 | 816 |
| 29 | Turkestan region, S-U | S-U-29 | N 42 39 986 | E 070 15 097 | 815 |
| 30 | Turkestan region, S-U | S-U-30 | N 42 39 984 | E 070 15 068 | 845 |
| 31 | Turkestan region, MA | MA -01 | N 42 25 665 | E 070 03 024 | 669 |
| 32 | Turkestan region, MA | MA -02 | N 42 25 670 | E 070 03 025 | 665 |
| 33 | Turkestan region, MA | MA-03 | N 42 25 672 | E 070 03 030 | 664 |
| 34 | Turkestan region, MA | MA-04 | N 42 25 675 | E 070 03 028 | 664 |
| 35 | Turkestan region, MA | MA-05 | N 42 25 672 | E 070 02 669 | 660 |
| 36 | Turkestan region, MA | MA-06 | N 42 25 671 | E 070 02 682 | 662 |
| 37 | Turkestan region, MA | MA-07 | N 42 25 670 | E 070 02 642 | 664 |
| 38 | Turkestan region, SA | SA-01 | N 41 27 773 | E 069 25 611 | 579 |
| 39 | Turkestan region, SA | SA-02 | N 41 27 794 | E 069 25 634 | 577 |
| 40 | Turkestan region, SA | SA-03 | N 41 27 796 | E 069 25 641 | 580 |
| 41 | Turkestan region, SA | SA-04 | N 41 27 802 | E 069 25 649 | 578 |
| 42 | Turkestan region, SA | SA-05 | N 41 27 813 | E 069 25 671 | 578 |
| 43 | Turkestan region, SA | SA-06 | N 41 27 860 | E 069 25 729 | 578 |
| 44 | Turkestan region, SA | SA-07 | N 41 27 874 | E 069 25 744 | 576 |
| 45 | Turkestan region, SA | SA-08 | N 41 27 958 | E 069 25 637 | 575 |
| 46 | Turkestan region, SA | SA-09 | N 41 27 955 | E 069 25 636 | 576 |
| 47 | Turkestan region, SA | SA-10 | N 41 27 692 | E 069 25 686 | 577 |
| 48 | Turkestan region, SA | SA-11 | N 41 27 693 | E 069 25 684 | 574 |
| 49 | Almaty region, AL | AL-01 | N 43 179 141 | E 076 867 784 | 898 |
| 50 | Almaty region, AL | AL-02 | N 43 197 52 | E 076 84 600 | 895 |
| 51 | Almaty region, AL | AL-03 | N 43 19 733 | E 076 84 664 | 896 |
| 52 | Almaty region, AL | AL-04 | N 43 19 756 | E 076 84 601 | 895 |
| 53 | Almaty region, AL | AL-05 | N 43 197 55 | E 076 84 602 | 894 |
| 54 | Almaty region, AL | AL-06 | N 43 20 214 | E 076 84 941 | 896 |
| 55 | Almaty region, AL | AL-07 | N 43 13 322 | E 076 55 063 | 880 |
| 56 | Almaty region, AL | AL-08 | N 43 13 331 | E 076 55 262 | 883 |
| 57 | Almaty region, AL | AL-09 | N 43 13 013 | E 076 54 955 | 896 |
| 58 | Almaty region, AL | AL-10 | N 43 13 011 | E 076 54 954 | 895 |
| 59 | Almaty region, AL | AL-11 | N 43 13 010 | E 076 54 950 | 892 |
| 60 | Almaty region, AL | AL-12 | N 43 13 012 | E 076 54 951 | 898 |
| 61 | Almaty region, AL | AL-13 | N 43 13 013 | E 076 54 956 | 891 |
| 62 | Almaty region, BA | BA-01 | N 43 51 353 | Е 077 69 19 | 720 |
| 63 | Almaty region, BA | BA-02 | N 43 51 329 | E 077 69 359 | 725 |
| 64 | Almaty region, ES | ES-01 | N 43 18 085 | Е 077 29 997 | 1286 |
| 65 | Almaty region, ES | ES-02 | N 43 18 086 | Е 077 29 993 | 1284 |
| 66 | Almaty region, ES | ES-03 | N 43 18 081 | Е 077 30 000 | 1287 |
| 67 | Almaty region, ES | ES-04 | N 43 341 342 | Е 077 471 442 | 1285 |
| 68 | Almaty region, ES | ES-05 | N 43 341 11 | Е 077 481 579 | 1286 |
| 69 | Almaty region, ES | ES-06 | N 43 341 054 | Е 077 480 984 | 1284 |
| 70 | Almaty region, ES | ES-07 | N 43 341 010 | Е 077 480 994 | 1286 |
| 71 | Almaty region, ES | ES-08 | N 43 18 164 | Е 077 29 976 | 1283 |
| 72 | Almaty region, ES | ES-09 | N 43 18 162 | Е 077 29 973 | 1282 |
| 73 | Almaty region, ES | ES-10 | N 43 18 112 | Е 077 30 004 | 1291 |
| 74 | Almaty region, ES | ES-11 | N 43 18 107 | Е 077 29 990 | 1286 |
| 75 | Almaty region, ES | ES-12 | N 43 18 104 | Е 077 29 991 | 1284 |
Note: Geographic region, location are: Turkestan region (Sairam-Ugam, S-U; Mashat, MA; Sarkyrama, SA); Almaty region (Almaty, AL; Baidibek-bi, BA and Esik, ES).
Table 2.
Details and characteristics of selected SSR markers used for analysis of walnut genetic diversity.
Table 2.
Details and characteristics of selected SSR markers used for analysis of walnut genetic diversity.
| Primer | Forward Primer (5'-3') |
Reverse Primer (5'-3') |
Annealing Temperature (°C) | Product Size Range (bp) |
|---|---|---|---|---|
| WGA001 | ATTGGAAGGGAAGGGAAATG | CGCGCACATACGTAAATCAC | 56 | 180–210 |
| WGA027 | AACCCTACAACGCCTTGATG | TGCTCAGGCTCCACTTCC | 57 | 225-260 |
| WGA042 | GTGGGTTCGACCGTGAAC | AACTTTGCACCACATCCACA | 55 | 210–260 |
| WGA118 | TGTGCTCTGATCTGCCTCC | GGGTGGGTGAAAAGTAGCAA | 60 | 186–200 |
| WGA009 | CATCAAAGCAAGCAATGGG | CCATTGCTCTGTGATTGGG | 56 | 231–245 |
| WGA202 | CCCATCTACCGTTGCACTTT | GCTGGTGGTTCTATCATGGG | 62 | 259–295 |
| WGA276 | CTCACTTTCTCGGCTCTTCC | GGTCTTATGTGGGCAGTCGT | 60 | 168–194 |
| WGA376 | GCCCTCAAAGTGATGAACGT | TCATCCATATTTACCCCTTTCG | 56 | 230–265 |
Table 3.
Polymorphism levels of SSR markers and genetic diversity parameters.
| Locus | Na | Ne | I | Ho | He |
|---|---|---|---|---|---|
| WGA001 | 6,000 | 3,882 | 1,507 | 0,612 | 0,742 |
| WGA027 | 2,000 | 1,930 | 0,675 | 0,238 | 0,482 |
| WGA042 | 2,000 | 1,943 | 0,679 | 0,341 | 0,485 |
| WGA118 | 3,000 | 2,665 | 1,039 | 0,593 | 0,625 |
| WGA009 | 6,000 | 4,592 | 1,650 | 0,625 | 0,782 |
| WGA202 | 9,000 | 5,619 | 1,916 | 0,571 | 0,822 |
| WGA276 | 10,000 | 6,664 | 2,095 | 0,717 | 0,850 |
| WGA376 | 9,000 | 6,458 | 1,995 | 0,676 | 0,845 |
| Mean | 5,875 | 4,219 | 1,444 | 0,547 | 0,704 |
Note: Na – number of detected alleles; Ne – effective number of alleles; Ho – observed heterozygosity; He – expected heterozygosity; I – Shannon information index.
Table 4.
Comparison of six Juglans regia populations from different regions of Kazakhstan based on genetic parameters.
Table 4.
Comparison of six Juglans regia populations from different regions of Kazakhstan based on genetic parameters.
| Pop | Location | Na | Ne | I | Ho | He |
|---|---|---|---|---|---|---|
| pop1 | S-U | 4,625 | 3,658 | 1,307 | 0,697 | 0,683 |
| pop2 | MA | 2,375 | 1,851 | 0,670 | 0,367 | 0,388 |
| pop3 | SA | 2,500 | 1,877 | 0,642 | 0,290 | 0,363 |
| pop4 | AL | 3,500 | 2,612 | 1,000 | 0,437 | 0,555 |
| pop5 | BA | 1,125 | 1,075 | 0,157 | 0,063 | 0,109 |
| pop6 | ES | 3,875 | 3,420 | 1,187 | 0,634 | 0,639 |
Note: pop 1- pop3 – walnut populations from the Turkestan region (Sairam-Ugam, S-U; Mashat, MA; Sarkyrama, SA); pop 4- pop6 - populations from the Almaty region (Almaty, AL; Baidibek-bi, BA and Esik, ES).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



