
Submitted:
26 November 2024
Posted:
27 November 2024
You are already at the latest version
Abstract
Common walnut (Juglans regia L.) is a valuable nut crop widely distributed within the temperate climate zone. It has several secondary centers of origin associated with refugia formed during the LGM (Last Glacial Maximum) period, one of which was presumably located in the Caucasus region. Currently, walnut populations of the Caucasus are studied fragmentarily, especially in the northeast of the region. Our study is aimed at investigating the genetic diversity, population structure and possible processes of walnut introduction in the Caspian region of the Northeast Caucasus (Dagestan). Genotyping using 11 SSR markers was performed for 158 samples from local walnut populations. Also, to identify the facts of introduction, 42 varieties of different geo-graphical origin were added to research. The genetic diversity of SSR markers in the analysis of population varied from 4 to 20 alleles per locus, with an average value of 9.82. Analysis of the genetic structure revealed significant genetic differences between the subpopulations of the Mountain and Flatland Dagestan. Presumably, the structure of the gene pool could have been in-fluenced by terrain factors and ethnocultural characteristics. Subpopulations of Flatland Dagestan have an intermediate position between the western cultivars and the gene pool of Mountain Dagestan, the populations of which showed significant genetic isolation. In the studied samples set of population selections, the established facts of introduction were rare. We have suggested that a significant part of the gene pool of walnut from Mountain Dagestan has an autochthonous origin from plants that survived the LGM in the Caucasian refugia. The results of the work are the basis for further studies of the local gene pool and verification of our hypothesis about the presence of an indigenous germplasm preserved in this region from the period the LGM.
Keywords:
genetic structure
; Juglans regia
; North-East Caucasus
; Western Caspian
; microsatellites
; refugia
; polymorphism
1. Introduction
Juglans regia L., commonly known as the Persian or English walnut, is one of the most important nut crops [1,2]. Its nuts are rich in microelements and polyunsaturated fatty acids and have up to 70-75% fat [3,4]. They are used both for fresh consumption, in the confectionery industry, and for processing into oil [2]. Juglans regia L. is prized for its high-quality timber as well [5]. It is grown both as a grafted crop in industrial gardens and as seedlings in a large number of countries with a temperate and subtropical climate worldwide. China, USA, Türkiye and Iran are the leaders in the production of walnuts [1]. The conservation of biological diversity of the species in the world is ensured by significant natural tracts of walnut trees. They are seedlings (natural walnut forests, as well as local seed populations).
It has been argued that the most probable primary center of origin and diversity of Juglans regia L is the Central Asian region. At the same time, the last ice age had a significant impact on the walnut gene pool. It led to habitat fragmentation and refugia formation, from where the walnut subsequently spread again [6-8]. Such refugia were formed due to mountain ranges that prevented glacier spread [8]. The study of the genetic relationships of walnut germplasm is an important direction in the genetics of this crop. It allows for the effective management of genetic resources for better breeding practices. This is no less important for identifying species distribution mechanisms, local gene pool formation, as well as their relationships with germplasm from other regions.
Microsatellite markers are one of the effective tools for solving this problem; they are widely used to characterize walnuts [9]. Academic literature contains numerous studies analyzing collections of genetic resources of research centers in different countries worldwide, including Iran [10], Türkiye [11], Spain [12], France – INRAs worldwide collection [13], USA – USDA walnut germplasm repository at the University of California, Davis, USA [14], Russia [15,16], and China [17]. Moreover, a number of large-scale studies have been carried out to analyze local populations in different regions of Eurasia. They clarified and significantly supplemented scientific data on species distribution [18,6,7,8] and human impact [19]. The study of local walnut populations is of great importance as well. A number of studies were carried out in various regions of Iran [20,21], Central Asian countries [22,23], China [24-27], Pakistan [28], India [29], Türkiye [30], Italy [31], total samples of genotypes from various countries of Europe, Asia, and Africa [32].
Genetic diversity of walnuts in the North Caucasus is relatively poorly studied. A number of scientists researched local populations to some extent. The studies were mainly presented on germplasm from regions located on the southern slope of the Greater Caucasus Range or in Transcaucasia [7,8]. The gene pool of walnut representing the Caspian region of the North Caucasus has not yet been studied in terms of genetic diversity, structure and relationships between populations and germplasm from other regions of the world, as well as the influence of the geographical factors on the structure of populations within the region.
Dagestan is a mountainous region of the Russian Federation, located in the northwestern part of the Caucasus and adjacent to the Caspian Sea. It has a complex geographical structure, including plains, highlands, mountain river valleys, and the Caspian lowland. It borders Azerbaijan in the south and Georgia in the southwest and has a rich and long history, as well as a complex, ethnic structure.
Geobotanists during expeditions and descriptions of plant communities of Dagestan in the late 19th – early 20th centuries noted the presence of walnut trees in the forests of Dagestan (mainly in the valleys of the rivers of Southern Dagestan: Samur, Gyulgerichay, Rubas-chay and in mixed forest communities). In addition, it was frequently found in populated areas. Let us mention that planting walnut trees was not widespread. Since due to cultural traditions and beliefs of some local nations, this entailed negative consequences for the person who planted it. However, the nut trees that grew by self-seeding was never cut down [33,34]. This implies the predominant role of natural distribution, rather than anthropogenic factors. This also ensured the preservation of the local gene pool and a low level of introduction from other regions. At the same time, a number of ethnographers and historians describe the high level of distribution of walnuts, both trees and nuts (in markets) in some areas of Dagestan [35,36]. This indicates differences in the traditions of local peoples and the probable cultivation of this crop by part of the local population. Effective agroforestry to obtain walnuts is likely to take place as well. Such is mentioned in some sources [37].
Evidently, these facts indicate a high probability of the presence of an autochthonous gene pool of walnut in Dagestan, which could have been present here even before the Last Glacial Maximum and preserved in the refugia. This assumption is supported by the fact that there are two refugia of woody summer-green plants located between the Black and Caspian Seas – the Colchic and Hyrcanian. [38]. Palynological studies of fossil pollen also support this theory [39-41]. Aradhaya M. et al (2017) [8], based on molecular genetic studies of the gene pool from Eastern Europe, Asian regions and the South Caucasus, revealed a certain level of genetic isolation of the Caucasian walnut gene pool. It should be pointed out that the study involved samples collected in Azerbaijan (69 samples) and Georgia (15 samples).
Our objective was to study genetic diversity of walnut populations in Dagestan, taking into account limited knowledge of walnut in the North Caucasus in general and in its Caspian part in particular, the heterogeneity of climate and geography of this region, as well as the complex and rich history of socio-economic relationships.
2. Materials and Methods
2.1. Plant material
Walnut samples were selected from 16 locations in the Republic of Dagestan (Northeast of the Greater Caucasus Range): 4 sampling points are located in the flat part of Dagestan, 3 sampling points are in the north of Mountain Dagestan, 9 sampling points are in the south of Mountain Dagestan. The number of samples at one sampling point varied from 5 to 15. A total of 158 samples were collected from 16 locations. Figure 1 shows the sampling locations on the map of Dagestan.
The selection of plant material was carried out from trees growing on the household plots of settlements (flat terrain) and in the vicinity of populated areas (mountainous terrain). The figure 2 shows photos of several walnut plants found during expeditions.
When collecting samples, a distance of at least 250-300 meters was maintained between trees and samples were taken from trees with a diameter of at least 25-30 cm at a level of 1 m from the soil surface. Some samples were taken from trees with a diameter of more than 1 meter at a level of 1 m from the soil surface in a number of locations (Dyubek, Khotoch, Kakhtsug, Karchag). More detailed information about sampling points is presented in Supplementary Materials Table S1.
The general samples set included varieties from the former USSR: 5 varieties from the Republic of Crimea, 15 varieties from Krasnodar Krai (Kuban region), 6 varieties from the Republic of Moldova, 10 varieties from the Republic of Kyrgyzstan. Six varieties from France and the USA were added as well. Thus, the total volume for genotyping was 200 genotypes of population selections and walnut varieties. List of samples from the general set presented in (Supplementary Materials Table S2).
2.2. DNA extraction. PCR setup. PCR product analysis
Genomic DNA was isolated from young leaves using a modified CTAB protocol [42]. Genotyping of 200 samples was performed for 11 SSR loci (WGA001, WGA004, WGA009, WGA069, WGA072, WGA079, WGA089, WGA202, WGA276, WGA321, WGA376); the PCR was performed using primer pairs designed by Woeste et al. (2002) and Dangl et al. (2005) [43,44]. PCR conditions were selected according to the protocol of Pollegioni et al. (2014) [6]. Electrophoresis of PCR products was carried out on a Nanophor 05 genetic analyzer. Accurate assessment of the sizes of PCR products was performed using the GeneMarker V3.0.1 software.
2.3. Analysis of genetic diversity
GenAlEx 6.5 software was used to calculate the parameters of microsatellite loci (number of alleles (Na), effective number of alleles (Ne), the Shannon diversity index (I), observed heterozygosity (Ho), expected heterozygosity (He), and fixation index (F) [45]. STRUCTURE 2.3.4 software was used for the genetic structure analysis of walnut populations [46]. The following parameters were used to perform the Bayesian analysis: burn-in period of 200,000 repeats, 100,000 Markov Chain Monte Carlo (MCMC) replications. The value of K varied from 1 to 7. The most plausible number K was determined by the value of deltaK set in a web-based program STRUCTURE HARVESTER v.0.6.1 [47]. Clustering of groups by origin (population selections groups and groups of varieties) by the UPGMA method was carried out in the Popgen 1.23 program [48] Distribution of population selections groups and groups of varieties on the PCoA graph was computed in Genalex version 6.5. The distribution of the studied walnut samples on the PCoA graph was computed in the Past 2.17 program. [49] The variance between different regions was estimated using AMOVA in Genalex version 6.5 [45].
3. Results
3.1. Genetic variability of studied set of walnut genotypes
Genotyping of 158 walnut samples from the Republic of Dagestan using 11 SSR markers allowed us to identify a total of 108 alleles with an average value of 9.818 alleles per locus (Table 1). The maximum number of alleles (20) was found in marker WGA276; the minimum number of alleles (4) in markers WGA 4 and WGA 72. The number of effective alleles ranged from 0.906 (WGA089) to 2.504 (WGA276) with a mean of 4.262. The mean values of expected and observed heterozygosity were 0.715 and 0.600, respectively. The overall fixation index has a low positive value. This is due to similar values of expected and observed heterozygosity.
The selected walnut samples from 16 locations were divided into 3 geographical regions: “South of Mountain Dagestan”, “North of Mountain Dagestan” and “Flatland Dagestan”. Samples from the indicated regions were compared based on the mean values of the following microsatellite loci parameters: number of alleles (Na), effective number of alleles (Ne), ratio between the effective number of alleles and the observed number of alleles (Ne/Na), the Shannon diversity index (I), observed heterozygosity (Ho), expected heterozygosity (He), fixation index (F). Table 2 shows the results of the analysis.
The lowest genetic diversity is characteristic of "North of Mountain Dagestan". This is evidenced by the low values of such parameters as the number of alleles, the effective number of alleles, and the Shannon diversity index. The average number of observed alleles is highest in the “South of Mountain Dagestan” samples, while the number of effective alleles is higher in the “Flatland Dagestan” samples. This is due to a larger number of alleles with low frequency of occurrence in the “South of Mountain Dagestan” set of samples relative to that from the “Flatland Dagestan”, which is characterized by higher Ne/Na values. In addition, the Shannon diversity index (I) values indicate a greater genetic diversity of the “Flatland Dagestan” samples compared to those from the mountainous area. The maximum level of observed and expected heterozygosity was found in the “Flatland Dagestan” samples. All three samples are characterized by positive values of the fixation index; the lowest value is in the "Flatland Dagestan" set of samples, the highest is in the "North of Mountain Dagestan" set of samples.
3.2. Evaluation of AMOVA parameters between regional groups of Juglans regia
Similarly, samples from three regions (“South of Mountain Dagestan”, “North of Mountain Dagestan” and “Flatland Dagestan”) were compared according to AMOVA parameters (Table 3). When comparing the values of the intergroup variance (Est. Var. Among Pops), reflecting the degree of dispersion between groups, the lowest values were found between the North and South of Mountain Dagestan. The highest values of intergroup variance were noted between the North of Mountain Dagestan and Flatland Dagestan. Intragroup variance in the Table 3 is divided into intra-individual and inter-individual.
The inter-individual variance, which reflects the intra-group level of dispersion, has the highest values between the North and South of Mountain Dagestan. The lowest values are typical when comparing the groups Northern Mountain Dagestan and Flatland Dagestan. The intra-individual variance, reflecting the level of observed heterozygosity of the analyzed groups, when comparing samples did not vary significantly within the range of 3.107 - 3.423.
F-statistics (Fst, Fis, Fit, Nm) were calculated based on the variance values (Among Pops, Among Indiv, Within Indiv). Based on the values of the Fst parameter, it can be stated that the degree of genetic isolation is most pronounced between groups from Mountain and Flatland Dagestan. The smallest differences between the parameters Fis and Fit were obtained when comparing the groups North and South of Mountain Dagestan. This indicates their genetic proximity. High values of gene flow Nm=6.54 also confirm this.
3.3. Estimated genetic distances between groups by origin of Juglans regia with used UPGMA and PCoA methods
We used UPGMA and PCoA methods for estimated genetic distances between groups by origin of samples. The distribution of samples by group is presented in Supplementary Materials (Table S2). The following groups of varieties (eastern origin) were added to the population selections to establish genetic relationships of the Dagestan walnut populations with the species world gene pool: varieties from Kyrgyzstan and Kuban varieties of Central Asian origin (further Asian Kuban); western origin: varieties from Moldova, Western Europe, USA, Kuban varieties and varieties from Crimea. The varieties of Moldova, Crimea, Kuban and Kyrgyzstan are mostly selections from local populations or progeny obtained from such trees. [16, 50-52].
UPGMA dendrogram had three main clusters (Figure 3). Cluster 1 is represented by varieties from Kyrgyzstan and Kuban varieties of Asian origin. This cluster is the most genetically distant from the other studied groups. Cluster 2 is formed by groups of varieties from Moldova, Kuban, Crimea, Western Europe and USA. Cluster 3 included population selections from Dagestan. Cluster 3 included two subclusters: subcluster 3.1 (sampling points from Flatland Dagestan) and subcluster 3.2 (sampling points from Mountain Dagestan).
The PCoA analysis identified three localization areas of sample groups (Figure 4). The first area contained two groups of varieties of Asian origin (Kyrgyz and Asian Kuban). The second area included groups of Western origin varieties and two groups from Flatland Dagestan (Komsomolskij, Tatajurt). The third area was formed by groups from Mountain Dagestan. The two groups from Flatland Dagestan (Shamhal Termen, Chapaevo) occupied an intermediate position between the second and third areas on the graph. It is worth noting that of all the groups from Mountain Dagestan, the three groups from Northern Mountain Dagestan (Nizhneye Kazanishche, Gimry, Khotoch) occupied the most distant position from the groups from Flatland Dagestan.
3.4. Estimated genetic distances between individual samples Juglans regia on the PCoA graph
The distribution of individual samples on the PCoA graph reflects their geography as well (Figure 5). Thus, samples from Mountain Dagestan are located in the upper left part; samples from Flatland Dagestan occupied the lower part; varieties of Western origin are in the right part; varieties of Asian origin are located in the upper right corner. However, this approach did not allow us to separate samples from the North and South of Mountain Dagestan.
3.4. Genetic analysis of walnut samples in the program STRUCTURE
STRUCTURE software estimated the probability of samples set distribution between a given numbers of clusters. Based on the deltaK values, the optimal cluster value was determined. Two set of samples were formed for analysis. The first samples set consisted of population selections of walnuts from the Republic of Dagestan, as well as varieties of Kyrgyz, Kuban, Moldovan, Crimean, Western European and USA origin to establish genetic links between walnuts from Dagestan and varieties from other regions. The second samples set included only samples from the Republic of Dagestan and was formed to reflect the structure of the regional gene pool. The optimal cluster value of three (K=3) was determined for two sets. A list of samples from the two sets and Q values of samples obtained from STRUCTURE software analysis is presented in (Supplementary Materials Table S3 and Table S4). Analysis of the first samples set in the Structure program enabled us to divide the studied genotypes into three groups: samples from Mountain Dagestan (Cluster 1), samples from Flatland Dagestan (Cluster 3), and walnut of varietal origin (Cluster 2). The genetic distance between clusters (Net nucleotide distance) varied: the greatest distance was between Clusters 1 (Mountain Dagestan) and 2 (Walnut varieties); Cluster 3 (Flatland Dagestan) occupied an intermediate position. The analysis of the first samples set in the Structure program is presented in Figure 6.
The second set of samples is represented by population selections of walnut from Dagestan. The analyzed samples distributed between clusters as follows: samples from Mountain Dagestan were assigned to Clusters 1 and 2; samples from the plain part of Dagestan were grouped in Cluster 3. At the same time, Cluster 1 predominates in the genotypes of the Southern part of Mountain Dagestan.; in turn, Cluster 2 prevails in selections from the Northern part of Mountain Dagestan. Cluster 3 (Flatland Dagestan) is genetically the most distant from the other clusters. Cluster 1 is genetically closer to Cluster 3 than Cluster 2. Figure 7 presents the analysis of the second samples set in the STRUCTURE program and the geographic distribution of population selection locations.
4. Discussion
4.1. Polymorphism of SSR loci
Genetic evaluation of 158 walnut samples from the Republic of Dagestan using 11 SSR markers allowed identifying a total of 108 alleles, with an average value of 9.818 alleles per locus. The number of effective alleles varied from 1.972 (marker WGA089) to 9.678 (marker WGA276). Expected and observed heterozygosity ranged from 0.480–0.897 and 0.297–0.715, respectively. The average values were 0.715 for He and 0.600 (Ho). Since WGA markers are most often used to study walnut genetic diversity [9], we compared the level of polymorphism of our results only with WGA markers to obtain a more objective picture.
We conducted a comparative analysis of the polymorphism of SSR markers with studies of the germplasm of walnut local populations in different regions worldwide. The data were also compared with larger-scale studies that involved germplasm from different regions. One study similar to ours (in terms of geographic scale) examined local populations in the eastern Italian Alps. In it, on a set of 215 samples, the “number of alleles” indicator for WGA SSR markers varied from two to nine alleles per locus with an average value of 4.7. At the same time, the average value of observed heterozygosity was 0.558 [53]. The lower polymorphism levels in relation to our study (Na=9.8 alleles per locus and Ho=0.600) may be a consequence of both the distance from biodiversity centers and selection influence during species distribution in the region. The authors assume this in the article.
In China, in a study of five walnut populations in Tibet, comprising a total of 100 samples [24], the total number of alleles for 12 WGA SSRs was 107, with an average of 8.9. The values of the number of effective alleles, observed and expected heterozygosity were 3.432, 0.512 and 0.586, respectively. Note that this is lower than the values found in our study (Ne = 4.262; Ho=0.600; He=0.715). However, as in our study, the WGA 276 marker turned out to be the most polymorphic (22 alleles were identified in a total set of 100 J. regia + 109 J. sagilliata samples).
In another study, carried out on the germplasm of J. regia from Tibet, the average allele count per locus for 12 microsatellites was 9.92 with Ne=3.34 in a set of 86 samples; Ho and He were 0.504 and 0.588 [25]. Despite the relatively high mean allele count per locus, the lower Ne, Ho and He values compared to our study indicate not only a higher level of homozygosity but also a greater number of rare/unique alleles detected in populations.
Yet another example is a study relatively larger in scale by samples set volume, but more significant in geographical terms of the selection locations. The above-mentioned study of populations from 14 countries in Europe, Asia and Africa used 12 WGA SSRs to genotype 189 samples. The identified average number of alleles per locus was 11.5. The number of effective alleles was 4.2, and the expected and observed heterozygosity was 0.73 and 0.62, respectively. [32]. Slightly higher values of these quantities (except Ne) may be a consequence of a more heterogeneous gene pool, selected in geographically remote regions, rather than a slight excess of the samples number in the studied set, in comparison with our study (a total of 158 samples from Dagestan).
Comparative analysis with data on SSR polymorphism of the gene pool of Central Asian walnut makes it possible to compare our populations with those of the potential centers of origin. V. Roor et al. analyzed 520 samples using seven SSR markers. 440 samples were from Asia (Kyrgyzstan, Uzbekistan, Iran, Pakistan, and India), and the rest from three European countries (Czech Republic, Austria, and Croatia). They identified between 5 and 24 alleles per locus for a total of 88 alleles across the seven markers, corresponding to an average of 12.5 alleles per locus. Moreover, the highest rates of allelic diversity were found in populations sampled in South Asia. The lowest level of polymorphism was characteristic of the samples set from Europe – two times lower than in the samples set from South Asia [22].
Polymorphism level varied from 5 to 26 alleles per locus with an average value of 11.4 when studying genetic relationships and polymorphism of Central Asian populations (Kyrgyzstan, Uzbekistan, and Tajikistan). The set included 624 samples from 21 locations (wild populations/forest and local seed populations within settlements); 12 WGA SSR markers were used [23]. In our study (158 Dagestan samples), the average number of alleles was 9.818. The Ne value in Gaisberger H. et al (2020) ranged from 1.51 to 8.78 (with a mean of 3.97); the mean values of expected and observed heterozygosity were 0.67 and 0.57, respectively. This is somewhat lower than the values we identified (Ne=4.92; He=0.715; Ho=0.600) and is a consequence of a more uniform distribution of allele frequencies and a higher level of heterozygosity in our set of samples.
Eurasian gene pool large-scale studies confirm a fairly high level of genetic diversity in our set of samples, revealed by microsatellite genotyping. In Aradhya M. et al (2017) – about 680 samples, 19 markers (eight common markers with our study) – the average number of alleles was 12; the expected and observed heterozygosity were 0.700 and 0.501, respectively [8]. Pollegioni P. et al (2017) used 14 SSR markers and found an average allele count per locus of 14.214. The samples set included 91 populations from 16 countries in Asia and Europe (2008 samples in total); the average values of expected, observed heterozygosity, and the number of effective alleles were 0.720, 0.559, and 4.334 [7].
Summarizing the comparative assessment of the polymorphism level studies in worldwide academic literature, we conclude a relatively high level of genetic diversity in the populations of the western Caspian region of the North Caucasus. The identified high level of genetic polymorphism may indicate the absence of both pronounced effect of any limiting environmental factors and genetic erosion of the gene pool, as well as favorable conditions for maintaining the genetic diversity of local populations of J. regia at a high level.
4.2. Analysis of the genetic structure of walnut populations from Dagestan
The analysis established the genetic relationships of walnut in the region and the influence of terrain on the formation of the local population structure of the species. Varieties of different origins were also included to identify the possible impact of introduction on the local gene pool. Taking into account the possible introduction of germplasm to Dagestan from the former USSR, we included varieties from the following regions of active walnut breeding: Moldova, Crimea, Kyrgyzstan, and Kuban. Varieties of Western European and USA origin, which are widely distributed worldwide, were added as well.
The mountain ranges of Dagestan serve as a natural barrier preventing the exchange of genetic material and contribute to the formation of unique local subpopulations. These processes could contribute, on the one hand, to a decrease in the values of observed heterozygosity and, on the other hand, to an increase in the values of the fixation index [54]. This tendency is a special case of the manifestation of the Wahlund effect [55]. Comparison of Ho and F values between samples from Mountain and Flatland Dagestan allows us to estimate the influence of mountain ranges on the structure of the walnut population. The results indicate that the Flatland Dagestan set has a higher value of observed heterozygosity; in turn, the fixation index values are higher for Mountain Dagestan samples. This fact indicates the presence of genetic barriers in the mountainous terrain of the region. A similar pattern is observed when assessing the values of the parameters of the effective number of alleles and the Shannon diversity index, reflecting the allelic diversity. The highest values of Ne and I were found for Flatland Dagestan; the lowest indicator values were characteristic for the North of Mountain Dagestan. The samples set from the South of Mountain Dagestan occupied an intermediate position between the specified groups of samples in terms of Ne and I indicators.
It is worth noting that samples from the South of Mountain Dagestan and Flatland Dagestan have similar values for the average number of alleles per locus. However, the parameters effective number of alleles and the Shannon diversity depend on not only the number of alleles, but their occurrence frequency as well. Thus, making them more reliable indicators for assessing genetic diversity [56]. In general, subpopulations in the mountainous part of the region are characterized by lower allelic diversity than lowland plants. Thus, geography and terrain also influence the allelic diversity of populations.
AMOVA determined the reliability of the allocation of the three designated groups in the general set of population selections in Dagestan. Flatland Dagestan samples are allocated to a separate group with the highest degree of reliability. This is evidenced by the indicators of intergroup variance when compared with Mountain Dagestan groups, as well as their significant contribution to the overall diversity (8% and 10%). At the same time, the North Mountain Dagestan group is more isolated from Flatland Dagestan than the South Mountain Dagestan group. In turn, the differences between the groups from Mountain Dagestan are rather insignificant (0.140) (4%), which confirms their genetic proximity. Thus, there is a border dividing the areas of distribution of two significantly different gene pools of walnut between the Mountain and Flatland Dagestan. Terrain is a more significant factor influencing the local gene pool than geographic distances. It is worth noting the relatively high values of intergroup variances that we obtained in comparison with the results of previously conducted studies of local walnut germplasm using SSR markers. Thus, the analysis of the mountain and foothill population of the Italian Alps walnut revealed a low contribution of intergroup variance to the overall diversity (2.12%) [53]. On the other hand, for a larger study of the gene pool of the Italian walnut, which included selections from different regions of the country, the percentage contribution of intergroup variance was 5.37% [57]. A more significant contribution of intergroup diversity (9%) was concluded in a study of walnut subpopulations in Pakistan [28].
Consequently, the value of intergroup variance in the SSR markers studies of local walnut populations varied depending on the region. It is worth noting a regional study of the walnut gene pool in China, where the percentage contribution of intergroup variance was 29.08%. High values of intergroup variance in Chinese populations are explained by significant geographic distances between walnut groups, which contribute to genetic isolation and gene flow weakening [26]. In turn, in our study, the selection of population samples was carried out on a sufficiently limited territory to assert the role of geographic distances in the isolation of individual plant groups.
We applied STRUCTURE software for a more complete analysis of the local gene pool structure. The optimal number of clusters (K=3) corresponded to the three identified geographic groups. Cluster 3 was predominant for all Flatland Dagestan samples. Samples from Southern and Northern Mountain Dagestan were assigned to Clusters 1 and 2. Despite the similar genetic structure and likely common genetic origin, the subpopulations of Southern and Northern Mountain Dagestan differed in the type of predominant cluster. It is worth noting that Cluster 1, which is predominant in Southern Mountain Dagestan, has a smaller genetic distance from Cluster 3 (Flatland Dagestan) than Cluster 2, which is predominant in the Northern Mountain Dagestan samples. This fact is confirmed by AMOVA data: The Northern Mountain Dagestan population is more isolated from the Flatland Dagestan walnut than the Southern Dagestan population.
In connection with the above, we have established two main facts reflecting the regional structure of the walnut population: 1) a contrasting boundary between the subpopulations of mountain and flat terrain, expressed in sharp genetic differences in the selected walnut samples; 2) within Northern Mountain Dagestan there exist an area with a greater degree of genetic isolation and less diversity. We assume, that the exchange of genetic material between the flat and southern parts of Mountain Dagestan is carried out bypassing the northern mountain ranges along the western coast of the Caspian Sea, penetrating deep into the mountains in the south along the rivers Samur, Gyulgerichay, Rubas-chay valleys.
Analysis of the mechanisms of exchange of genetic material between populations sheds light on the factors that contribute to the emergence of isolation and genetic barriers.
The exchange of genetic material between walnut populations is carried out through the dissemination of pollen and seeds. Since walnuts are anemophilous, pollen can travel distances significantly greater than the natural dissemination range of most seeds. Pollen has been recorded to travel distances of up to 1.5 kilometers for the genus Juglans [58-60]. However, the contribution of closely located trees prevails in pollination [61]. It is more difficult to identify the role of humans in the distribution and exchange of genetic material. This factor largely depends on the cultural characteristics of the region and cultivating traditions. Seed material for the reproduction of walnut plantations in household plots and in general on the territory of settlements can be either selected from local populations or brought in from outside.
Studies of walnut distribution at the local level confirm both of these approaches. Thus, the Italian Alps are characterized by the absence of a significant division of walnut into subpopulations. This is explained by the practice of intensive exchange of seed material between alpine settlements [53]. On the other hand, the walnut genetic structure in the villages of Tibet corresponds to the settlements from which the samples were collected. This fact indicates that the seed exchange is mainly carried out by neighbors within settlements and/or between related families [62]. In addition, the majority of the studied walnut plantations in the settlements of Central Asia were related to local semi-wild plants growing in the vicinity of settlements. The fact of introduction of walnut from another region was recorded only once [23].
Mountain Dagestan is distinguished by the ethnic and linguistic diversity of its indigenous population, which is especially noticeable in small remote villages [63]. This regional specificity serves as a factor that complicates seed distribution between individual settlements, which leads to greater isolation of local walnut subpopulations.
We studied the possible introduction of walnut in Dagestan taking into account regional peculiarities. To assess the possible impact of this process on local gene pool were also included in the analysis Varieties from the territory of the former Soviet Union and Western Europe.
Based on the clustering results in the STRUCTURE program (Figure 3), only three samples from Dagestan were assigned with a high degree of probability to Cluster 3, characteristic of cultivated varieties. This fact indicates that these three genotypes are not of local origin. Presumably, these genotypes are seedlings from cultivated varieties. The remaining Dagestan samples were assigned to clusters corresponding to their localization (Mountain or Plain Dagestan). Therefore, walnut introduction in Dagestan is weak; most seedlings originate from local seed material. Calculation of genetic distances between individual sampling points allowed to estimate both the degree of their isolation relative to each other and from groups of cultivated varieties. The results are presented as PCoA and UPGMA graphs (Figure 1 and 2). When comparing the above methods, genotypes distribution pattern can be identified in three main areas/clusters: varieties of eastern origin, varieties of western origin, and population selections from Dagestan.
At the same time, the PCoA graph shows that samples from the two northernmost locations of Flatland Dagestan are located along with Western varieties. Samples from the two remaining locations of Flatland Dagestan occupy an intermediate position between varieties of Western origin and Mountain Dagestan. We assume that the Flatland Dagestan subpopulation was formed as a result of mixing of the local walnut gene pool, preserved in the mountainous area, and the gene pool of Western origin. In turn, the influence of Asian germplasm has not been established.
On the other hand, the gene pool of walnuts from Mountain Dagestan probably is more autochthonous for this location. The isolated gene pool of J. regia from Mountain Dagestan may originates from the autochthonous gene pool of the Caucasian refugia formed during the LGM (last glacial maximum) period. Considering this possibility, we note that Aradhaya M. et al 2017 [8] indicate the possibility of the existence of such a refugia in the Caucasus. However, in their study, the gene pool of the Caucasus was represented by regions on the southern slope of the Greater Caucasus (Georgia and Azerbaijan). Our results (samples from the North-Eastern part of the Caucasus Mountains) showed significant genetic differences between the gene pool of the mountainous region and samples from the plain area, which are genetically closer to the cultivated varieties. In addition, the studied gene pool is characterized by a relatively high level of genetic diversity. All this may indicate the preservation of autochthonous germplasm in the LGM. It is obvious that further study of local germplasm from different regions of the Caucasus and the world will allow a more complete picture.
5. Conclusions
Both under-exploration of walnut in the North Caucasus and complex geographic structure of the Caspian region determines the interest in studying the genetic diversity and population structure of this species in Dagestan. The study analyzed genetic relationships and assessed the genetic diversity indicators of the South of Mountain Dagestan, the North of Mountain Dagestan and Flatland Dagestan.
Significant genetic differences were found between populations of J. regia growing in mountainous and flat areas. They may be a consequence of both the influence of the terrain and differences in walnut cultivation traditions. Based on the obtained data on the relationships between the populations of Flatland Dagestan, the South of Mountain Dagestan and the North of Mountain Dagestan, it was suggested that the gene flow between the subpopulations of the lowland and southern parts of Mountain Dagestan passes along the western coast of the Caspian Sea, bypassing the mountainous terrain in the northern part of Mountain Dagestan. At the same time, the introduction of the gene pool and intensive exchange of germplasm is more likely for Flatland Dagestan, while for the mountainous regions, where the walnut grows in a wild and semi-wild state, isolation is characteristic. The revealed genetic isolation of Mountain Dagestan populations suggests them originating from the autochthonous J. regia gene pool, including the likelihood of this species refugia preservation during the last glacial maximum.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1: Geographical information on the sites where material was collected from natural populations of Juglans regia; Table S2: List of population selections and varieties samples; Table S3: STRUCTURE analysis of varieties and Dagestan populations; Table S4: STRUCTURE analysis of Dagestan populations.
Author Contributions
Conceptualization, Iv.S.; methodology, Iv.S., Il.S.; investigation, Iv.S., Il.S, D.A.; writing—original draft preparation, Iv.S., Il.S. D.A.; writing—review and editing, Iv.S., Il.S., D.A.; project administration, Iv.S.; funding acquisition, Iv.S.. All authors have read and agreed to the published version of the manuscript.
Funding
This work was financially supported by grant of Russian Science Foundation and Kuban Science Foundation (project № 22-16-20061, https://rscf.ru/project/22-16-20061/), Russia.
Data Availability Statement
Source data are available from the authors upon request.
Acknowledgments
We thank to Sergey Khokhlov (‘The Labor Red Banner Order Nikita Botanical Gardens - National Scientific Center of the Russian Academy of Sciences’, Yalta, Russia) for samples of Crimean walnut varieties; Davlet Mamadzhanov (National Academy of Sciences of the Kyrgyz Republic «Jalal-Abad Scientific Center», Jalal-Abad, Kyrgyzstan) for samples of Kyrgyz walnut varieties and Ramidin Kazakhmedov (Dagestan Breeding Experimental Station of Viticulture and Vegetable Growing of North Caucasian Federal Scientific Center of Horticulture, Viticulture, Wine-making, Derbent, Russia) for help with collecting of walnut samples in South of Dagestan.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Vahdati, K.; Arab, M.M.; Sarikhani, S.; Sadat-Hosseini, M.; Leslie, C. A.; Brown, P.J. Advances in Persian walnut (Juglans regia L.) breeding strategies. In Advances in Plant Breeding Strategies: Nut Beverage Crops 2019, 4, 401-472. [CrossRef]
- McGranahan, G.; Leslie, C. Walnut, in: Badenes, M., Byrne, D. (Eds.), Fruit Breeding. Handbook of Plant Breeding. Springer, Boston, MA, 2012; pp 827-846. [CrossRef]
- Rébufa, C.; Artaud J.; Dréau, Y.L. Walnut (Juglans regia L.) oil chemical composition depending on variety, locality, extraction process and storage conditions: A comprehensive review. Journal of Food Composition and Analysis 2022, 110, 104534. [CrossRef]
- Rabrenović, B.; Natić, M.; Zagorac, D.D.; Meland, M.; Akšić, M. F. Bioactive Phytochemicals from Walnut (Juglans spp.) Oil Processing By-products. In: Ramadan Hassanien, M.F. (eds) Bioactive Phytochemicals from Vegetable Oil and Oilseed Processing By-products. Reference Series in Phytochemistry. Springer, Cham. 2023, 535-557. [CrossRef]
- Najafian Ashrafi, M.; Shaabani Asrami, H.; Vosoughi Rudgar, Z.; Ghorbanian Far, M.; Heidari, A.; Rastbod, E.; Jafarzadeh, H.; Salehi, M.; Bari, E.; Ribera, J. Comparison of Physical and Mechanical Properties of Beech and Walnut Wood from Iran and Georgian Beech. Forests. 2021, 12, 801. [CrossRef]
- Pollegioni, P.; Woeste, K.; Chiocchini, F.; Olimpieri, I.; Tortolano, V.; Clark, J.; Hemery, G. E.; Mapelli, S.; Malvolti M.E. Landscape genetics of Persian walnut (Juglans regia L.) across its Asian range. Tree Genet. Genomes 2014, 10, 1027–1043. [CrossRef]
- Pollegioni, P.; Woeste, K.; Chiocchini, F.; Lungo, S.D.; Ciolfi, M.; Olimpieri, I.; Tortolano, V.; Clark, J.; Hemery, G.E.; Mapelli, S.; Malvolti, M.E. Rethinking the history of common walnut (Juglans regia L.) in Europe: Its origins and human interactions. PLoS ONE 2017, 12(3), e0172541. [CrossRef]
- Aradhya, M.; Velasco D.; Ibrahimov, Z.; Toktoraliev, B.; Maghradze, D.; Musayev, M.; Bobokashvili, Z.; Preece, J. E. Genetic and ecological insights into glacial refugia of walnut (Juglans regia L.). PLoS ONE 2017, 12(10), e0185974. [CrossRef]
- Bernard, A.; Lheureux, F.; Dirlewanger, E. Walnut: past and future of genetic improvement. Tree Genetics & Genomes 2018, 14, 1 . [CrossRef]
- Davoodi, F.; Rezaei, M.; Heidari, P.; Hokmabadi, H.; Lawson, S. Identification and DNA Fingerprinting of Some Superior Persian Walnut Genotypes in Iran. Erwerbs-Obstbau 2021, 63, 393-402. [CrossRef]
- Bozhuyuk, M.R. Determination of the genetic diversity of walnut (Juglans regia L.) cultivar candidates from northeastern Turkey using SSR markers. Mitt. Klosterneuburg 2020, 70, 269-277.
- Ruiz-Garcia, L.; Lopez-Ortega G.; Fuentes, A.; Frutos, D. Identification of a walnut (Juglans regia L.) germplasm collection and evaluation of their genetic variability by microsatellite markers. Spanish Journal of Agricultural Research 2011, 9(1), 179-192.
- Bernard, A.; Barreneche, T.; Lheureux, F.; Dirlewanger. E. Analysis of genetic diversity and structure in a worldwide walnut (Juglans regia L.) germplasm using SSR markers. PLOS ONE 2018, 13(11), e0208021. [CrossRef]
- Aradhya. M.; Woeste K.; Velasco, D. Genetic Diversity, Structure and Differentiation in Cultivated Walnut (Juglans regia L.). Acta Horticulturae 2010, 861, 127-132.
- Balapanov, I.; Suprun, I.; Stepanov, I.; Tokmakov, S.; Lugovskoy, A. Comparative analysis Crimean, Moldavian and Kuban Persian walnut collections genetic variability by SSR-markers. Sci. Hortic 2019, 253, 322-326. [CrossRef]
- Suprun, I.I.; Stepanov I.V.; Vahdati, K.; Tokmakov, S.V.; Balapanov, I.M.; Al-Nakib, E.A.; Khokhlov, S.Yu.; Sokolova, V.V. Analysis of genetic diversity in three Eastern European walnut germplasm collections. Scientia Horticulturae 2024, 334, 113275. [CrossRef]
- Chen, L.N.; Ma, Q.G.; Chen, Y.K.; Wang, B.Q.; Pei, D. Identification of major walnut cultivars grown in China based on nut phenotypes and SSR markers. Scientia Horticulturae 2014, 168, 240-248. [CrossRef]
- Khan, H.; Ullah, I.; Woeste, K.; Fiaz, S.; Zeb, U.; Ghazy, A. I.; Azizullah, A.; Shad, S.; Malvolti, M.E.; Yue, M.; et al Population genetics informs new insights into the phytogeographic history of Juglans regia L. Genet Resour Crop Evol 2023, 70, 2263–2278. [CrossRef]
- Pollegioni, P.; Woeste, K.E.; Chiocchini, F.; Lungo, S.D.; Olimpieri, I.; Tortolano, V.; Clark, J.; Hemery, G.E.; Mapelli, S.; Malvolti, M.E. Ancient Humans Influenced the Current Spatial Genetic Structure of Common Walnut Populations in Asia. PLoS ONE 2015, 10(9), e0135980. [CrossRef]
- Vahdati, K.; Mohseni Pourtaklu, S.; Karimi, R.; Barzehkar, R.; Amiri, R.; Mozaffari, M.; Woeste, K. Genetic diversity and gene flow of some Persian walnut populations in southeast of Iran revealed by SSR mrakers. Plant Syst Evol 2015, 301, 691-699.
- Shahi Shavvon, R.; Qi, H.L., Mafakheri, M.; Fan, P.-Z.; Wu, H.-Y.; Vahdati, F.B.; Al-Shmgani, H.S.; Wang, Y.-H.; Liu, J. Unravelling the genetic diversity and population structure of common walnut in the Iranian Plateau. BMC Plant Biol 2023, 23, 201. [CrossRef]
- Roor, V.; Konrad, H.; Mamadjanov, D.; Geburek, T. Population Differentiation in Common Walnut (Juglans regia L.) across Major Parts of Its Native Range-Insights from Molecular and Morphometric Data. Journal of Heredity 2017, 108(4), 391-404. [CrossRef]
- Gaisberger, H.; Legay, S.; Andre, C.; Loo, J.; Azimov, R.; Aaliev, S.; Bobokalonov, F.; Mukhsimov, N.; Kettle, C.; Vinceti, B. Diversity Under Threat: Connecting Genetic Diversity and Threat Mapping to Set Conservation Priorities for Juglans regia L. Populations in Central Asia. Front. Ecol. Evol. 2020, 8, 171. [CrossRef]
- Wang, H.; Pan, G.; Ma, Q.; Zhang, J.; Pei, D. The genetic diversity and introgression of Juglans regia and Juglans sigillata in Tibet as revealed by SSR markers. Tree Genetics & Genomes 2015, 11, 1. [CrossRef]
- Wang, H.; Wu, W.; Pan, G.; Pei, D. Analysis of genetic diversity and relationships among 86 Persian walnut (Juglans regia L.) genotypes in Tibet using morphological traits and SSR markers. The Journal of Horticultural Science and Biotechnology 2015, 90(5), 563–570. [CrossRef]
- Feng, X.; Zhou, H.; Zulfiqar, S.; Hu, Y.; Feng, L.; Malvolti, M.E.; Woeste, K.; Zhao, P. The Phytogeographic History of Common Walnut in China. Front. Plant Sci. 2018, 9, 1399. [CrossRef]
- Han, H.; Woeste, K.E.; Hu, Y.; Dang, M.; Zhang, T.; Gao, X.-X.; Zhou, H.; Feng, X.; Zhao, G.; Zhao, P. Genetic diversity and population structure of common walnut (Juglans regia) in China based on EST-SSRs and the nuclear gene phenylalanine ammonia-lyase (PAL). Tree Genetics & Genomes 2016, 12, 111. [CrossRef]
- Magige, E.A.; Fan, P.-Z.; Wambulwa, M.C. Milne, R.; Wu, Z.-Y.; Luo, Y.-H.; Khan, R.; Wu, H.-Y.; Qi, H.-L.; Zhu, G.-F.; et al. Genetic Diversity and Structure of Persian Walnut (Juglans regia L.) in Pakistan: Implications for Conservation. Plants 2022, 11, 1652. [CrossRef]
- Shah, U.N.; Mir, J.I.; Ahmed, N.; Fazili, K.M. Assessment of germplasm diversity and genetic relationships among walnut (Juglans regia L.) genotypes through microsatellite markers. Journal of the Saudi Society of Agricultural Sciences 2018, 17(4), 339-350. [CrossRef]
- Guney, M.; Kafkas, S.; Keles, H.; Zarifikhosroshahi, M.; Gundesli, M.A.; Ercisli, S.; Necas, T.; Bujdoso, G. Genetic Diversity among Some Walnut (Juglans regia L.) Genotypes by SSR Markers. Sustainability 2021, 13, 6830. [CrossRef]
- Cipriani, P.G.; Paffetti, D.; Vettori, C.; Testolin, R. Genetic Diversity of Walnut (Juglans Regia L.) in the Eastern Italian Alps. Forests 2017, 8, 81. [CrossRef]
- Ebrahimi, A.; Zarei, A.; Lawson, S.; Woeste, K.E.; Smulders, M.J.M. Genetic diversity and genetic structure of Persian walnut (Juglans regia) accessions from 14 European African, and Asian countries using SSR markers. Tree Genetics & Genomes 2016, 12, 114. [CrossRef]
- Voronov, Yu. N. Wild relatives of fruit trees and shrubs of the Caucasian region and the Near East. Proceedings on Applied Botany, genetics and breeding 1925, 14, 44.
- Ilyinsky, A.A. Walnut and other fruit trees in the forests of the Samur River delta. Proceedings of the Dagestan Agricultural Institute 1941, 3, 144-168.
- Gritsenko, N.P. Socio-economic development of the Twilight regions in the XVIII – first half of the XIX century. The works of CHINIYAL 1969, 4, 289.
- Magomedov, N.A. Regional and international transit trade in the Western Caspian Region in the XVII – XVIII centuries; Makhachkala: IIAE DNC RAS, ALEF, 2015; 184 p.
- Magaramov, S.A.; Chekulaev, N.D.; Inozemtseva, E.I. The history of the Derbent garrison of the Russian Imperial Army (1722-1735); Makhachkala: Publishing House "Lotus", 2021; 200 p.
- Ghobad-Nejhad, M. Wood-inhabiting basidiomycetes in the Caucasus region—systematics and biogeography. Publications in Botany from the University of Helsinki 2011, 40, 8-25.
- Carrión, J.S.; Sánchez-Gómez, P. Palynological data in support of the survival of walnut (Juglans regia L.) in the western Mediterranean during last glacial times. J. Biogeogr 1992, 19, 623–630. [CrossRef]
- Filipova-Marinova, M.V.; Kvavadze, E.V.; Connor, S.E.; Connor, S.E.; Sjögren, P. Estimating absolute pollen productivity for some European Tertiary-relict taxa. Veget Hist Archaeobot 2010, 19, 351–364. [CrossRef]
- Gogichaishvili, L.K. Vegetational and climatic history of the western part of the Kura River basin. In: Bintliff JL, van Zeist W (eds) Paleoclimates, paleoenvironments, and human communities in the Eastern Mediterranean region in later prehistory; B.A.R. International Series: Oxford, 1984; pp 325–341.
- Rogers, S. O.; Bendich, A. J. Extraction of DNA from milligram amounts of fresh, herbarium and mummified plant tissues. Plant Mol. Biol. 1985, 5, 69-76. [CrossRef]
- Woeste, K.; Burns, R.; Rhodes, O.; Michler, C. Thirty polymorphic nuclear microsatellite loci from black walnut. J Hered. 2002, 93(1), 58–60 . [CrossRef]
- Dangl, G.S.; Woeste, K.; Aradhya, M.K.; Pitcher, A. M. K. Characterization of 14 microsatellite markers for genetic analysis and cultivar identification of walnut. J Am Soc Hortic Sci 2005, 130, 348–354. [CrossRef]
- 59Peakall, R.; Smouse, P.E. GenAlEx 6.5: genetic analysis in Excel. Population genetic software for teaching and research--an update. Bioinformatics 2012, 28(19), 2537-9. [CrossRef]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [CrossRef]
- Earl, D.A.; von Holdt, B.M. Structure harvester: a website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. 2012, 4, 359-361. [CrossRef]
- Yeh, F.C.; Yang, R.C.; Boyle, T. POPGENE Version 1.32: Microsoft Window-Based Freeware for Population Genetics Analysis. University of Alberta, Edmonton 1999.
- Hammer, O.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontologia Electronica 2001, 4(1), 1-9.
- Mamadzhanov, D.K. Recommendations for the introduction of the best varieties and forms of walnut, Institute of Forestry and Walnut Growing of the National Academy of Sciences of the Kyrgyz Republic named after prof. P.A. Gan.-B. 2005, 157, 499-506.
- Mamadzhanov, D.; Kenzhebaev, S.A. Walnut diversity and breeding in Kyrgyzstan. BIO Web of Conferences. International Scientific Online-Conference «Bioengineering in the Organization of Processes Concerning Breeding and Reproduction of Perennial Crops» 2020, 25, 02009. [CrossRef]
- Curkan, I.P. Walnut; Kishineu, 2004, P. 156.
- Vischi, M.; Chiabà, C.; Raranciuc, S.; Poggetti, L.; Messina, R.; Ermacora, P.; Cipriani, G.; Paffetti, D.; Vettori, C.; Testolin, R. Genetic Diversity of Walnut (Juglans Regia L.) in the Eastern Italian Alps. Forests 2017, 8, 81. [CrossRef]
- Vranckx, G.; Jacquemyn, H.; Muys, B.; Honnay, O. Meta-analysis of susceptibility of woody plants to loss of genetic diversity through habitat fragmentation. Conserv. Biol 2012, 26, 228–237. [CrossRef]
- Garnier-Géré, P.; Chikhi, L. Population subdivision, Hardy–Weinberg equilibrium and the Wahlund effect; In eLS; John Wiley andSons: Hoboken, NJ, USA, 2013. [CrossRef]
- Kanaka, K.K.; Sukhija, N.; Goli, R. Ch.; Singh, S.; Ganguly, I.; Dixit, S.P.; Dash, A.; Malik, М. On the concepts and measures of diversity in the genomics era. Current Plant Biology 2023, 33, 100278. [CrossRef]
- Pollegioni, P.; Woeste, K.; Olimpieri, I.; Marandola, D.; Cannata, F.; Malvolti, M.E. Long-term human impacts on genetic structure of Italian walnut inferred by SSR markers. Tree Genetics & Genomes 2011, 7, 707–723. [CrossRef]
- Bai, W.N.; Zeng, Y.F.; Zhang D.Y. Mating patterns and pollen dispersal in a heterodichogamous tree, Juglans mandshurica (Juglandaceae). New Phytol 2007, 176, 699–707. doi.10.1111/j.1469-8137.2007.02202.x.
- Robichaud, R.L.; Glaubitz, J.C.; Rhodes, O.E.; Woeste, K. A. Robust set of black walnut microsatellites for parentage and clonal identification. New Forests 2006, 32, 179–196. [CrossRef]
- Ebrahimi, A.; Lawson, S.S.; Frank, G.S.; Coggeshall, M.V.; Woeste, K.E.; McKenna, J.R. Pollen flow and paternity in an isolated and non-isolated black walnut (Juglans nigra L.) timber seed orchard. PLoS One 2018, 13(12), e0207861. [CrossRef]
- Polito. V.S.; Pinney, K.; Weinbaum, S. et al. Walnut pollination dynamics: pollen flow in walnut orchards. In V International Walnut Symposium 2004, 705, 465–472. 10.17660/ActaHortic.2005.705.68.
- Gunn, B. F.; Aradhya, M.; Salick, J. M. Genetic variation in walnuts (Juglans regia and J. sigillata; Juglandaceae): Species distinctions, human impacts, and the conservation of agrobiodiversity in Yunnan, China. American Journal of Botany 2010, 97(4), 660–671. [CrossRef]
- Caciagli, L.; Bulayeva, K.; Bulayev, O.; Bertoncini, S.; Taglioli, L.; Pagani, L.; Paoli, G.; Tofanelli, S. The key role of patrilineal inheritance in shaping the genetic variation of Dagestan highlanders. J Hum Genet 2009, 54, 689–694. [CrossRef]
Figure 1.
Distribution of walnut sampling points on the map of Dagestan.
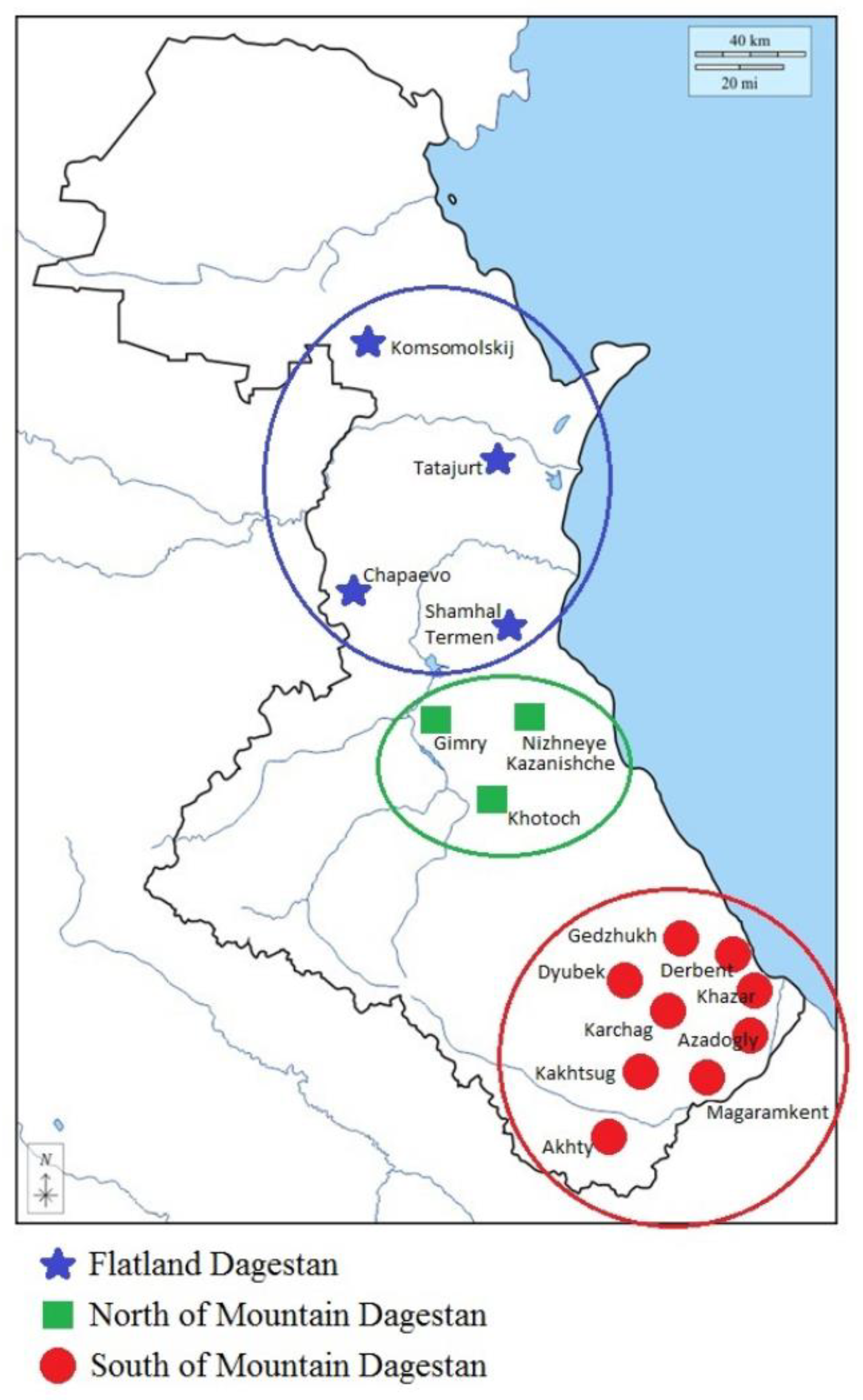
Figure 2.
Photos of some Persian walnuts grow in the Dagestan territory. А. An aged walnut tree in the vicinity of the village of Dyubek (approximate age is about 150-200 years; Аnatov Dzhalaludin is staying on right side from the tree). B. Two aged walnut trees on the pasture meadow nearby the Dyubek. C. Walnut trees on the slope along the mountain road to the village of Kakhtsug (left side from the road). D. Walnut trees on the slopes of canyon closely to Khotoch.
Figure 2.
Photos of some Persian walnuts grow in the Dagestan territory. А. An aged walnut tree in the vicinity of the village of Dyubek (approximate age is about 150-200 years; Аnatov Dzhalaludin is staying on right side from the tree). B. Two aged walnut trees on the pasture meadow nearby the Dyubek. C. Walnut trees on the slope along the mountain road to the village of Kakhtsug (left side from the road). D. Walnut trees on the slopes of canyon closely to Khotoch.

Figure 3.
Clustering of groups by origin of Juglans regia using the UPGMA method
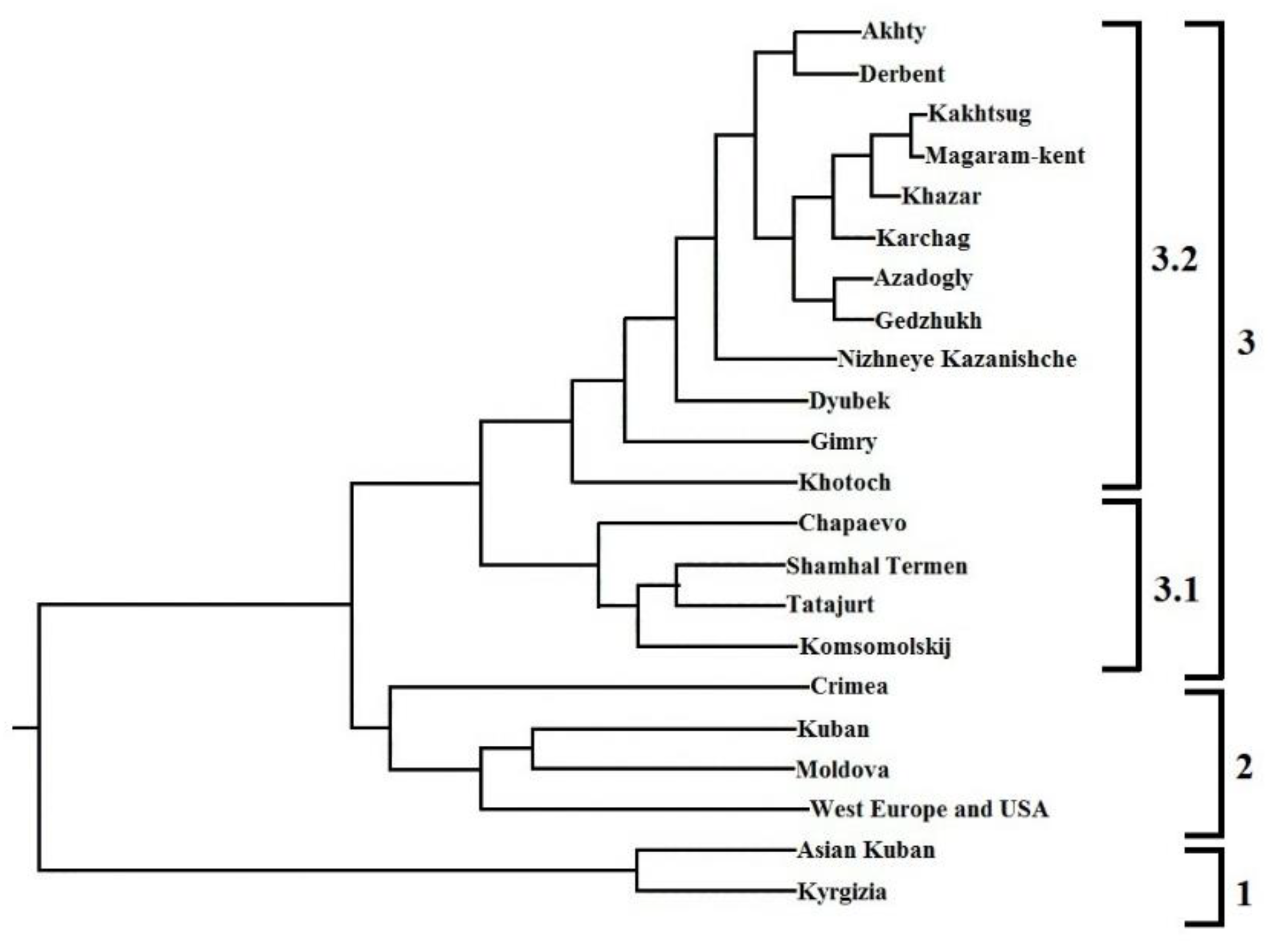
Figure 4.
Distribution of groups by origin of Juglans regia on the PCoA graph
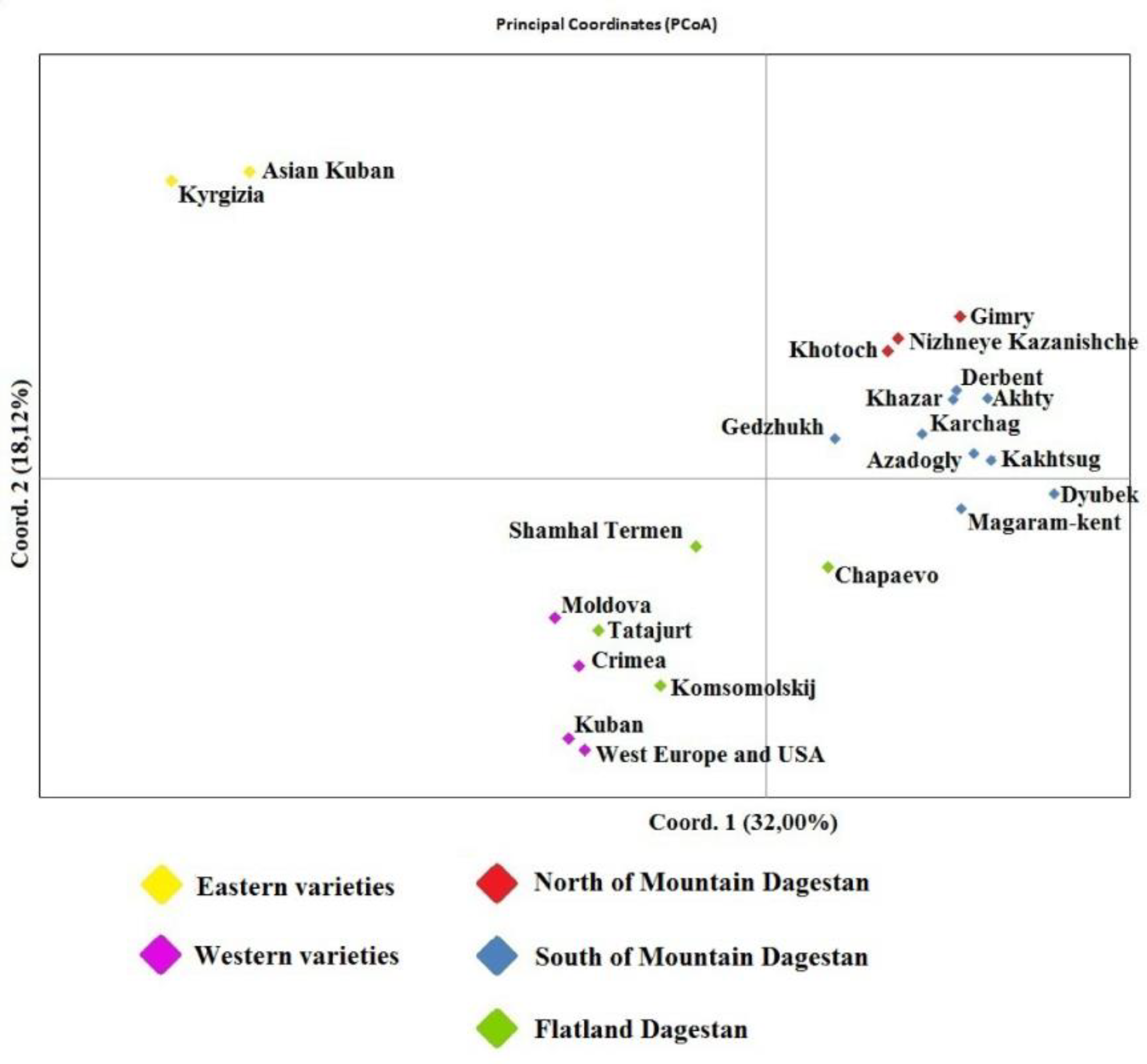
Figure 5.
Distribution of individual samples Juglans regia on the PCoA graph.
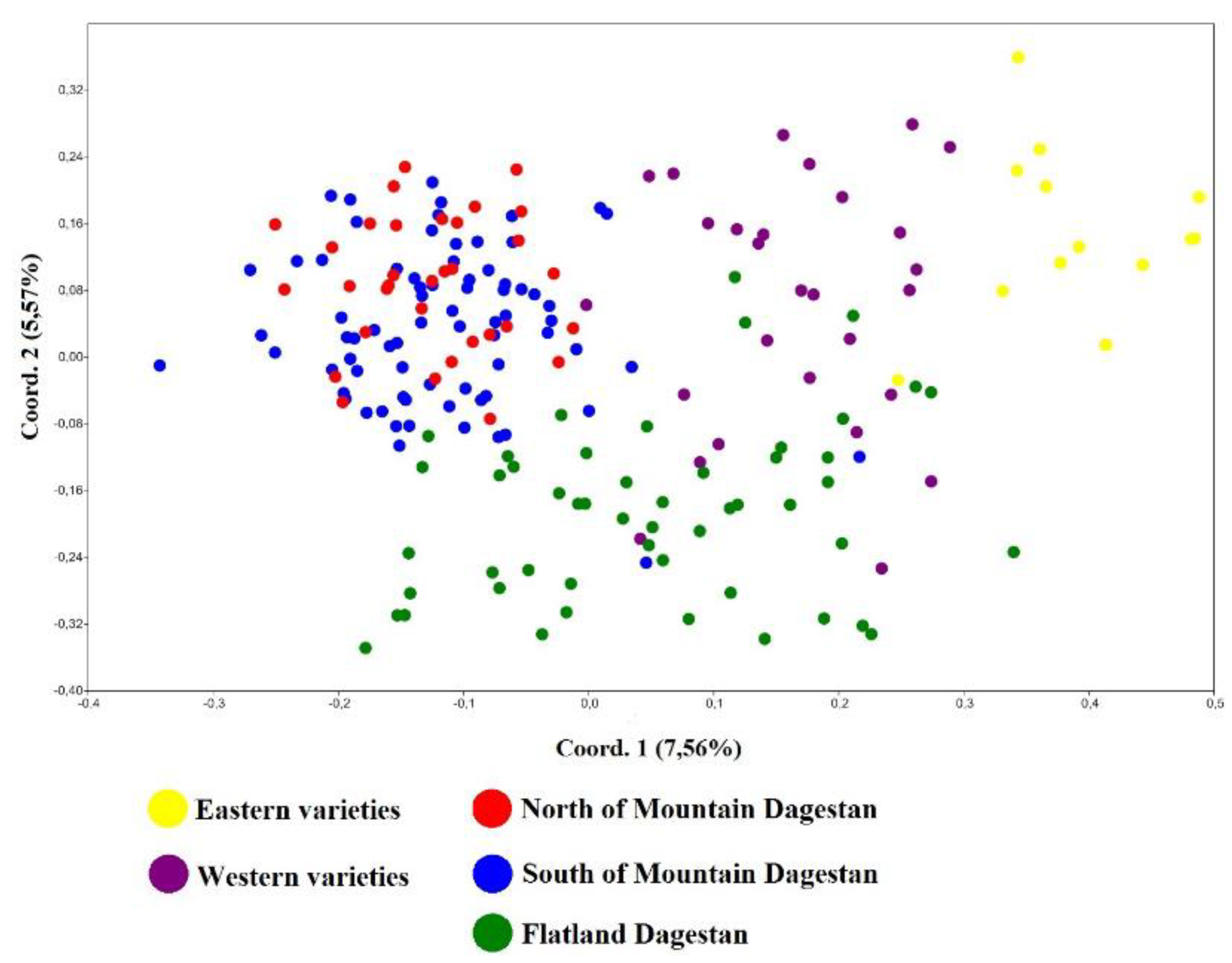
Figure 6.
Graph of analysis of walnut varieties and population samples in the STRUCTURE program. A. STRUCTURE graph: MD – Mountain Dagestan, FD – Flatland Dagestan, PWV – Persian walnut varieties. B. The NJ tree displays the genetic distances between the three clusters used in the STRUCTURE graph. The values of the genetic distances between the clusters are presented in the graph.
Figure 6.
Graph of analysis of walnut varieties and population samples in the STRUCTURE program. A. STRUCTURE graph: MD – Mountain Dagestan, FD – Flatland Dagestan, PWV – Persian walnut varieties. B. The NJ tree displays the genetic distances between the three clusters used in the STRUCTURE graph. The values of the genetic distances between the clusters are presented in the graph.
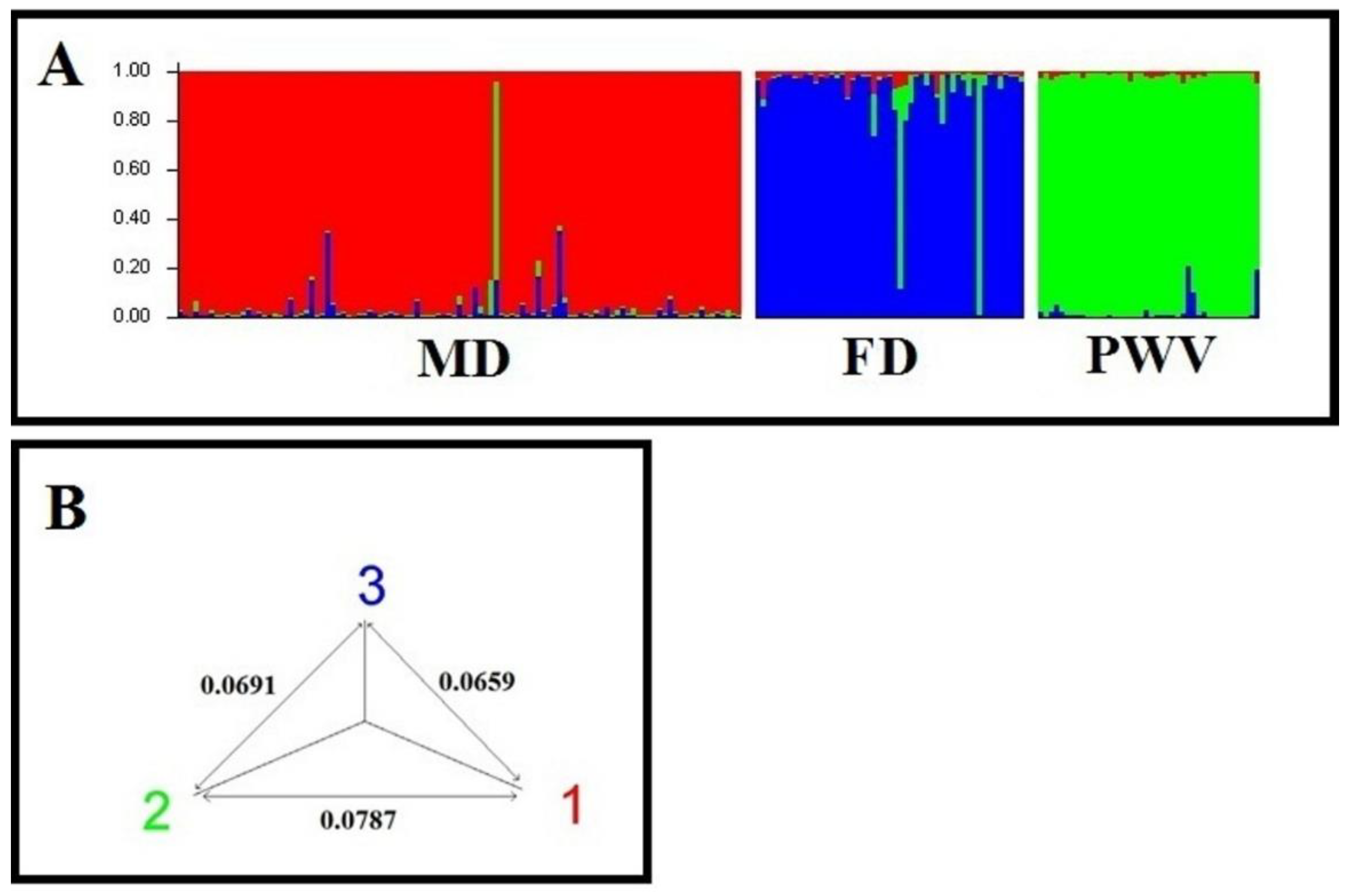
Figure 7.
Comparison of the analysis results of population selections in the STRUCTURE software with geographical location of the samples. A. Location of population sampling points of walnut on the map of Dagestan. The sampling points are presented as diagrams showing the contribution shares of the three clusters in local samples. Name of sampling points: 1 - Akhty, 2 – Kakhtsug, 3 – Magaramkent, 4- Karchag, 5 – Azadogly, 6 – Dyubek, 7 – Khazar, 8- Gedzhukh, 9 – Derbent, 10 – Khotoch, 11- Gimry, 12 - Nizhneye Kazanishche, 13 – Chapaevo, 14 - Shamhal Termen, 15 - Komsomolskij, 16 – Tatajurt. A.1. Terrain (altitude above sea level is given in meters). B. Distribution graph of samples between three clusters calculated in the STRUCTURE program. Groups highlighted on the graph: SMD – South of Mountain Dagestan, NMD – North of Mountain Dagestan, FD – Flatland Dagestan. С. The NJ tree displays the genetic distances between the three clusters used in the STRUCTURE graph. The values of the genetic distances between the clusters are presented in the graph. .
Figure 7.
Comparison of the analysis results of population selections in the STRUCTURE software with geographical location of the samples. A. Location of population sampling points of walnut on the map of Dagestan. The sampling points are presented as diagrams showing the contribution shares of the three clusters in local samples. Name of sampling points: 1 - Akhty, 2 – Kakhtsug, 3 – Magaramkent, 4- Karchag, 5 – Azadogly, 6 – Dyubek, 7 – Khazar, 8- Gedzhukh, 9 – Derbent, 10 – Khotoch, 11- Gimry, 12 - Nizhneye Kazanishche, 13 – Chapaevo, 14 - Shamhal Termen, 15 - Komsomolskij, 16 – Tatajurt. A.1. Terrain (altitude above sea level is given in meters). B. Distribution graph of samples between three clusters calculated in the STRUCTURE program. Groups highlighted on the graph: SMD – South of Mountain Dagestan, NMD – North of Mountain Dagestan, FD – Flatland Dagestan. С. The NJ tree displays the genetic distances between the three clusters used in the STRUCTURE graph. The values of the genetic distances between the clusters are presented in the graph. .
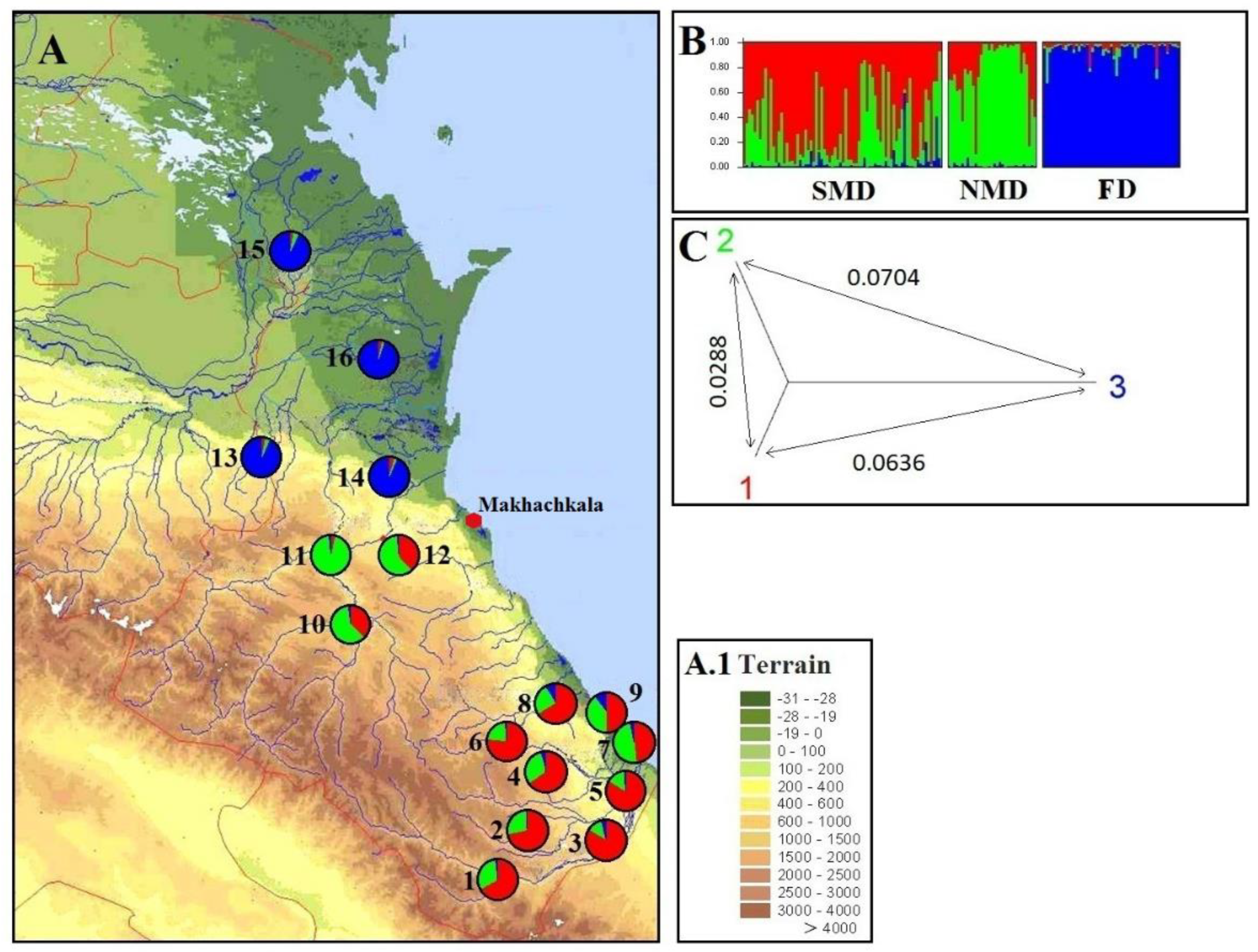

Table 1.
SSR loci polymorphism indices.
| Locus | Na | Ne | I | Ho | He | F |
| WGA001 | 8 | 3,477 | 1,466 | 0,633 | 0,712 | 0,112 |
| WGA376 | 11 | 6,052 | 2,032 | 0,709 | 0,835 | 0,151 |
| WGA069 | 14 | 4,606 | 1,814 | 0,462 | 0,783 | 0,41 |
| WGA276 | 20 | 9,678 | 2,504 | 0,759 | 0,897 | 0,153 |
| WGA009 | 7 | 2,198 | 1,1 | 0,513 | 0,545 | 0,06 |
| WGA202 | 14 | 5,286 | 1,912 | 0,741 | 0,811 | 0,087 |
| WGA089 | 7 | 1,923 | 0,906 | 0,494 | 0,48 | -0,028 |
| WGA321 | 10 | 3,981 | 1,551 | 0,69 | 0,749 | 0,079 |
| WGA 72 | 4 | 3,424 | 1,289 | 0,297 | 0,708 | 0,58 |
| WGA 79 | 9 | 3,618 | 1,455 | 0,722 | 0,724 | 0,003 |
| WGA 4 | 4 | 2,635 | 1,037 | 0,576 | 0,62 | 0,072 |
| Mean | 9,818 | 4,262 | 1,552 | 0,6 | 0,715 | 0,152 |
Table 2.
Genetic parameters of walnut sample groups in Dagestan.
| Regions | N | Na | Ne | Ne/Na | I | Ho | He | F |
|---|---|---|---|---|---|---|---|---|
| South of Mountain Dagestan | 74 | 7.545 | 3.444 | 0.456 | 1.376 | 0.574 | 0.662 | 0.114 |
| North of Mountain Dagestan | 33 | 5.273 | 2.991 | 0.567 | 1.254 | 0.545 | 0.646 | 0.146 |
| Flatland Dagestan | 51 | 7.455 | 4.024 | 0.540 | 1.491 | 0.672 | 0.715 | 0.067 |
Table 3.
Comparison of walnut regional groups from Dagestan according to AMOVA parameters.
| Source | df | SS | MS | Est. Var. | % | F-statistica |
|---|---|---|---|---|---|---|
| South and North of Mountain Dagestan | ||||||
| Among Pops | 1 | 16.559 | 16.559 | 0.140 | 4% | Fst=0.036 |
| Among Indiv | 105 | 443.090 | 4.220 | 0.556 | 15% | Fis=0.151 |
| Within Indiv | 107 | 332.500 | 3.107 | 3.107 | 82% | Fit=0.183 |
| Total | 213 | 792.150 | 3.804 | 100% | Nm=6.54 | |
| South of Mountain Dagestan and Flatland Dagestan | ||||||
| Among Pops | 1 | 44.231 | 44.231 | 0.331 | 8% | Fst=0,080 |
| Among Indiv | 123 | 517.853 | 4.210 | 0.417 | 10% | Fis=0,110 |
| Within Indiv | 125 | 422.000 | 3.376 | 3.376 | 82% | Fit=0.181 |
| Total | 249 | 984,084 | 4,124 | 100% | Nm= 2.86 | |
| North of Mountain Dagestan and Flatland Dagestan | ||||||
| Among Pops | 1 | 38.299 | 38.299 | 0.425 | 10% | Fst=0.100 |
| Among Indiv | 82 | 348.052 | 4.245 | 0.411 | 10% | Fis=0.107 |
| Within Indiv | 84 | 287.500 | 3.423 | 3.423 | 80% | Fit=0.196 |
| Total | 167 | 673.851 | 4.259 | 100% | Nm=2.26 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



