Submitted:
22 May 2023
Posted:
23 May 2023
You are already at the latest version
Abstract
In order to understand the genetic diversity of germplasm resources of kumquat in Guangxi, 14 kumquat germplasm resources in Guangxi and 12 accessions from other provinces were analyzed by using SRAP markers. In total, 19 primer pairs with high stability, good reproducibility, and high polymorphism were chosen for analysis of all the 26 kumquat genotypes. Among the 101 amplified bands, 87 (86.14%) were polymorphic. SRAP markers were analyzed by employing Principal Coordinate Analysis, Population Structure Analysis and Hierarchical Cluster Analysis (UPGMA). The classification results showed that 26 kumquat germplasm could be divided into 5 groups, including cultivated kumquat, intergeneric hybrid, wild kumquat from other provinces, wild kumquat from Guangxi and hybrid kumquat from Guangxi. Guangxi kumquat germplasm occurred high genetic diversity, which were clearly divided into 3 groups like cultivated kumquat, wild kumquat and kumquat hybrid. And the 8 cultivated kumquat varieties in Guangxi were further divided into two subgroups. Wild kumquat in Guangxi and wild kumquat in other provinces belong to different groups, meanwhile the hybrids of Guangxi kumquat form independent groups, thus indicated that Guangxi wild kumquat and kumquat hybrid possessed certain specificity, or they maybe belonged to different species. Among the tested 26 kumquat accessions, 23 unique genotype-specific SRAP markers were detected for 14 kumquat genotypes, which made it possible to surely identify them. For the remaining 12 accessions without genotype-specific markers, they were distinguished by various combinations of markers. These results may have certain importance for kumquat genetic research and cultivar selection.
Keywords:
Guangxi kumquat
; Germplasm resources
; Diversity
; SRAP markers
; Genetic analysis
1. Introduction
Guangxi located in the Southwest of China with warm subtropical monsoon climate and diverse topography, which make it a suitable area for growing citrus since ancient times [1]. With its abundant citrus genetic resources and long cultivation history, Guangxi occupies number one position in citrus production in China. Kumquat (Fortunella Swing) is a genus close to Citrus but with distinctive citrus fruit characteristics. it serves as a key industry in Yangshuo and Rongan Counties in Guangxi. The primary cultivar is ‘Rongan’ kumquat, which has given rise to a series of new varieties. Moreover, Guangxi possesses F. hindsii and some natural hybrids. Li et al. conducted ploidy analysis and SSR identification of ‘Gui Shanjingan’ (F. hindsii from Guangxi). The results indicated that there was a significant genetic difference between ‘Gui Shanjingan’ and the F. hindsii genotypes from other provinces, and it seemed an unique wild kumquat[2]. Additionally, Huang et al. collected Shanju (a kumquat genotype in Guangxi) resources at the border of China and Vietnam and the research genetic analysis results suggested that Shanju could be a new kumquat variety or species. Seeds of Shanju are mono-embryonic and its seedlings have short juvenile period, thus could be utilized as an effective breeding model plant for kumquat [3].
The origin of certain kumquat genotypes were unclear in the past. It was obscure about the evolutionary relationship between cultivated and wild kumquats and whether F. crassifolia is a pure species or a hybrid. However, with the development of molecular markers, better classification of kumquat resources becomes possible. Zhu et al. demonstrated through phylogenetic analysis that the genus Fortunella exhibited a genetic structure of Cultivated group (F. margarita, F. japonica, and F. crassifolia) – and Wild one (Hongkong kumquat) [4].
One of the most widely utilized systems in genetic analysis is the sequence-related amplified polymorphism (SRAP), which employs PCR to detect polymorphisms in the lengths of introns, promoters, or spacers among different individuals and species. Due to its simplicity and effectiveness, SRAP has found extensive application in analysis of genetic diversity, construction of genetic maps, mapping of crucial traits, and cloning of related genes in several plant species, including grape, Camellia oleifera, plum, and mangoes [5,6,7,8,9]. In this study, we employed the SRAP molecular marker technology to assess the genetic diversity of Guangxi kumquat germplasm resources, aiming to provide a foundation for selection of new kumquat varieties through germplasm innovation and breeding.
2. Materials and Methods
2.1. Material
Twenty-six kumquat genotypes (Table 1) were collected from Guangxi Citrus Germplasm Repository at Guangxi Academy of Specialty Crops and the National Citrus Germplasm Repository at Citrus Research Institute, Southwest University (Chongqing). Young leaves were sampled from each genotype for SRAP analysis.
2.2. DNA isolation
The genomic DNA from the leaves of 26 kumquat genotypes was isolated by using an improved CTAB protocol [10]. The concentration and quality of the DNA were assessed using an all-band microplate reader and 1% agarose gel electrophoresis.
2.3. PCR amplification for SRAP markers
Referring to the primer combinations reported by Zhang et al. [11], 4 kumquat varieties (Daguojindou, NB luofu, RA jingan, and HP jingan) were used for primer screening. In total, 19 primer pairs (Table 2) with high stability, good reproducibility, and high polymorphism were chosen for further analysis of all the 26 kumquat genotypes.
The SRAP-PCR reaction was carried out as described by Xu et al. [12] with slight modification. Specifically, 0.12 mM dNTP, 0.2 μM primers, 5 U Taq DNA polymerase, 2 μL 10× Taq Buffer (containing 1.6 mM Mg2+), and 50 ng of template DNA were added to a 20 μL SRAP-PCR reaction system. The PCR amplification procedure consisted of an initial denaturation step at 94℃ for 5 min, followed by five cycles of 94℃ for 60 s, 35℃ for 60 s, and 72℃ for 2 min, and then 35 cycles of 94℃ for 60 s, 55℃ for 60 s, and 72℃ for 60 s. A final extension was performed at 72℃ for 8 min. PCR products were stored at 4℃.
2.4. Agarose gel electrophoresis
The SRAP-PCR products were separated on a 2% agarose by electrophoresis for 1.5 to 2 h, subsequently stained with 4S Green Nucleic Acid Stain (Perfemiker), and then photographed using an imager[12].
2.5. Parameters used for analysis of SRAP markers
Bands with identical mobility among 26 kumquat genotypes, amplified with SRAP primers, were scored as "0" (absence of SRAP) and those polymorphic as "1" (presence of SRAP), resulting in the construction of a binary sequence matrix of "0, 1"[12]. The principal coordinate analysis (PCoA) was used to construct the biplot using PAST 3.11 software. Jaccard similarity coefficients were used to examine data from SRAP markers[13]. The population Structure analysis was performed using the Structure 2.3.4 software and the optimal K value was determined using the ΔK values in the Structure Harvester analysis method[16,17]. The Unweighted Pair-Group Method with Arithmetic Mean Algorithm (UPGMA) in PAST 3.11 software was used to create the phylogenetic trees[14,15]. Genotype-specific markers were searched through all the SRAP markers, which were used to identify kumquat accessions.
3. Results and analysis
3.1. Polymorphism analysis using SRAP markers
Out of the 420 pairs of primers screened as SRAP markers, 19 were selected as they yielded clear and bright bands. These primers were used to genotype 26 kumquat germplasms, and 102 bands were amplified in total (Figure 1), with an average of 5.37 bands per primer pair. Among these bands, 88 (86.27%) were polymorphic, and the average number of polymorphic bands per primer pair was 4.63 (Table 3).
3.2. Principal Coordinate Analysis
Principal Coordinates Analysis (PCoA) was performed on the data generated by the amplification of kumquat genomic DNAs using 19 SRAP primer combinations. Coord. 1 represented 34.80% of the genetic variation in these samples, and Coord. 2 covered 19.25% of the genetic variation. The obtained eigenvalues indicated that the first two coordinates provided a good summary of the data, as they explained 54.05% of the total variability (Table 4).
The biplot of PC1 and PC2 showed the 26 kumquats’ grouping (Figure 2). On the PC1 26 Kumquat accessions were divided into two main categories. The first group included Daguojindou, Dajindou, Shanjingan, FC-1, FC-2, FC-3, FC-4, FC-5 and Jinganzazhong, which belonged to wild kumquat germplasm. The second group combined the interspecies (Wenzhouju, Sijiju and Shouxingju) and all cultivated kumquat varieties. On the PC2, the two groups could be further divided into subgroups. The first group was classified into three subgroups, namely Hunan wild kumquat (Daguojindou, Dajindou and Shanjingan), Guangxi wild kumquat (FC-1, FC-2, FC-3, FC-4 and FC-5) as well as Jinganzazhong. The second one was covered the subgroup of intergeneric hybrids (Wenzhouju, Sijiju and Shouxingju) and that of cultivated kumquat varieties. However, the cultivar NB luofu remained certain distance with others.
According to the combination of PC1 and PC2, 26 kumquat genotypes could be divided into 5 groups. The first group was wild kumquat (F. hindisii), comprised Daguojindou, Dajindou and Shanjingan from Hunan province. The second one occupied by Guangxi wild kumquats FC-1, FC-2, FC-3, FC-4 and FC-5. The third one contained 14 kumquat cultivars collected from different locations. The fourth one was the intergeneric hybrids (Wenzhouju, Sijiju and Shouxingju). And the fifth one was Jinganzazhong, the wild hybrid from Gupo Mountain in Hezhou, Guangxi.
3.3. Population Structure Analysis
The admixture simulation model was used to assess the kumquat clustering types by screening nineteen SRAP primer combinations on the 26 genotypes. The cluster range was evaluated From K=1 to K=10. The output results showed a sharp peak with no ambiguity, indicating the highest delta K value at K = 2. There was a second sharp peak at K = 5 (Figure 3). Furthermore, the Bayesian bar graph was used to construct the graph for the admixture model. The accessions were grouped in subgroup clusters with >70% probability of membership fractions.
At K = 2, 11 out of 26 kumquats formed subpopulation I (red color, representing 42.3% of the total number of accessions), and 13 went into subpopulation II (green color, representing 50.0%) (Figure 4). Group I mainly contained wild kumquats; Group II included mainly cultivated accessions.
At K = 5, 13 kumquat genotypes, all cultivated varieties, gathered in subpopulation I (red color). The Guangxi wild hybrid Jinganzazhong solely occupied subpopulation II (green color). The 3 intergeneric hybrids (Wenzhouju, Sijiju and Shouxingju) entered in sub-population III (blue color). The 3 wild kumquats (F. hindisii) from Hunan (Daguojindou, Dajindou and Shanjingan) were grouped in subpopulation IV (yellow color). Finally, the 5 wild kumquat genotypes from Guanxi (FC-1, FC-2, FC-3, FC-4 and FC-5) formed the subpopulation V (purple). Strangerly, NB luofu did not follow in any group but had the mixed four groups’ genetic background (Figure 5).
3.4. Hierarchical Cluster Analysis (HCA)
A cluster analysis was carried out using the Jaccard coefficient by UPGMA method based on the Genetic similarity coefficients (Table 5). In the dendrogram, the kumquat genotypes were clustered into 5 groups (Figure 6). Group one (G1) was the largest, covering 14 kumquat cultivars collected from different locations. This group was further subdivided into 4 subgroups. Subgroup one contained 5 cultivars (RA jingan, FY jingan,Guijingan1,Guijingan2,YS jingan), cultivated in Guangxi. HP jingan, a mutant of RA jingan, together with its mutants F15-1 and CM jingan also from Guangxi formed Subgroup two. While the cultivars from Zhejing (NB jindan, WZ luofu, WZ jingdan, NB luowen) and that from Hunan (LY jingan) gathered in Subgroup three; And NB luofu from Zhejiang occupied a single subgroup. Group two (G2) comprised Wenzhouju, Sijiju and Shouxingju, which are all intergeneric hybrids. Group three (G3) contained FC-1, FC-2, FC-3, FC-4 and FC-5; which were wild kumquats collected from Guangxi. Group four involved Daguojindou, Dajindou and Shanjingan, which are wild kumquat (F. hindisii) from Hunan province. The single genotype, Jinganzazhong from Gupo Mountain in Hezhou, Guangxi, clustered in the last group.
3.5. Screening of genotype-specific markers and identification od kumquat accessions
Looking through the SRAP markers, some unique kumquat genotype-specific markers (a band was present/absent only in one genotype but not in others) were detected. Among the tested 26 kumquat accessions, 14 genotypes presented 23 unique specific markers. Jinganzazhong, the kumquat hybrid from Guanxi, had 4 unique markers, NB luofu from Zhejiang had 3, NB jindan, WZ jingdan, Dajindou and Dajindou each had 2, and the remaining had one for each (Table 6). NB jindan and WZ jingdan were the only cultivated varieties that had unique specific markers.
By using the unique specific SRAP markers, the 14 kumquat genotypes could be surely distinguished. For the remaining 12 accessions without unique specific markers, it needed marker combinations to identify them. FC-3, did not have unique specific marker, however, it shared the specific marker M3E17(180+) with FC-2, FC-4, FC-5, and the later 3 accessions had unique specific markers for each. Obviously FC-3 was able to be distinguished from them by the combination of M3E17(180+) with theunique specific markers of FC-2, FC-4 and FC-5. Meanwhile, FC-3 had also the specific marker M1E23(500+) with Daguojindou, thus it was possible to be identified by the combination of M1E23(500+) with unique Daguojindou-specific marker M1E23(400-). Shouxingju is a kumquat hybrid without unique specific marker, but it had specific marker M2E21(400+) with Sijiju and M20E2(700+) with Wenzhouju. The combinations of M2E21(400+) with M4E12(1300-), unique marker for Sijiju, or with M17E2(530-), unique marker for Wenzhouju, made Shouxingju distinguishable (Table 7).
There existed another pair of genotypes, WZ luofu and NB luowen originated in Zhejiang, which remained indistinguishable. NB luowen and WZ Luofu had the commune marker M10E7(850-) with NB luofu and Wenzhouju. Nevertheless, NB luofu possessed 3 unique markers [M2E21(400-), M3E17(450+) and M11E21(250+)], and Wenzhouju owned the M17E2(530-) unique marker, which were able to discriminate them from NB luowen and WZ Luofu. In addition, WZ Luofu had specific marker M10E7(320-) with NB luofu, Wenzhouju and Shanjingan but not with NB luowen, making the discrimination of the tow genotypes. WZ Luofu was easy to be separated from other marker shared genotypes by their unique markers [NB luofu M2E21(400-), M3E17(450+), M11E21(250+); Wenzhouju M17E2(530-); and Shanjingan M1E22(300-)] (Table 7).
There were still 8 accessions that were not able to be distinguished. These are cultivars and their bud mutants cultivated mainly in Guangxi including RA jingan, HP jingan, CM jingan, FY jingan, F15-1, YS jingan, Guijingan1 and Guijingan2 (named Guangxi cultivar group) (Table 6). Carefully searching through all the SRAP markers, Guangxi cultivar group possessed two group-specific markers [M1E23(800-) and M7E4(1050+)] with NB jindan, WZ jingdan, LY jingan, WZ luofu and NB luofu (Table 8). As the later 5 genotypes were able to be distinguished from Guangxi cultivar group by their unique specific markers, therefore, the members of Guangxi cultivar formed a special group separated from all the other kumquat accessions (Table 8).
Successively, genotype-specific markers within Guangxi cultivar group were checked through. Guijingan1, HP jingan and FY jingan had genotype - specific marker for each [M1E22(740-), M10E7(710-) and M3E17(90-), respectively], permitting easily being discriminated from other group members. For others, bi- or tri-markers were detected. CM jingan and HP jingan had a bi-specific marker M2E21(480-), making CM jingan distinguishable from HP jingan by its specific marker M10E7(710-). YS jingan and F15-1 shared another bi-specific marker M14E12(760-), while Guijingan2, YS jingan and HP jingan possessed a tri-specific marker M14E12(700-). By the two combinations of markers, HP jingan was first discriminated by its single marker, YS jingan occupied both markers thus distinguishable, thereafter F15-1 only presented M14E12(760-) and Guijingan2 solely M14E12(700-). The last member was RA jingan in the group which did not have any specific marker, so as to be identified from other 7 members (Table 9).
4. Discussion
Genetic diversity of plant species is the basis of their survival and evolution, and genetic research is an effective method to evaluate and quantify genetic variation [18]. With the development of DNA fingerprinting technology, molecular markers have been widely used in molecular taxonomy, variety identification and marker-assisted selection in different plants [19,20]. SRAP is a PCR-based technique, which has been widely used in plant germplasm diversity, variety identification, genetic mapping and gene cloning in recent years [21] in various crops, including coffee [22], grape [23], lychee [24]. In this study, 19 combinations of SRAP primers were used to determine the genetic diversity of 26 kumquat accessions. Out of the 102bands amplified, 88 (86.27%) were polymorphic, which made it possible to analyze the genetic diversity and to identify all the 26 kumquat accessions. These results indicated that SRAP markers were suitable for kumquat genetic diversity analysis and genotype identification.
In the studies on germplasm diversity, principal coordinate analysis, structural analysis and UPGMA cluster analysis are often utilized to carry out data analysis [4,15]. In present work, the SRAP data of 26 kumquat genotypes were analyzed by using these three methods. In the principal coordinate analysis, though PC1 and PC2 only contained 54.05% of all the information, PC1 analysis results played an important role in the classification of wild and cultivated kumquat, which were further subdivided into 5 subgroups by PC2. The results of structural analysis showed that 26 kumquat germplasm were first divided into two groups, wild kumquat and cultivated kumquat, and then into five groups. However, could not be classified into any of the groups as it had genetic background components of four groups. it might suggest that the NB Luofu could be of hybrid origin. UPGMA cluster analysis showed that 26 kumquat genotypes were also divided into 5 groups. Surprisingly, all the 26 kumquat accessions were classified into 5 identical groups with the 3 data processing methods, which might indicate that the SRAP markers were stable and reliable. Here the results might reflect the genetic differences between the kumquat accessions. First, In the principal coordinate analysis and structural analysis at K=2, the wild and cultivated kumquat genotypes were first divided indicating that wild and cultivated kumquat possessed main genetic differences.
Other than the genetic origin, geographic regions affects also the kumquat biodiversity. Among the 26 tested kumquat accessions, 14 were from Guangxi and 12 were from other provinces. As mentioned above, genetic background classified the 14 Guangxi kumquat germplasm into three groups according to their origins, i.e. wild kumquat, hybrid kumquat and cultivated varieties. The results indicated the rich genetic diversity of k Guangxi umquat germplasm. However, in comparison with the kumquat germplasm from other provinces, the Guangxi kumquat germplasm resulted in different groups even belonging to the same cultivated or wild types. The Guangxi wild kumquat (FC-1, FC-2, FC-3, FC-4 and FC-5) stayed in different groups with the wild kumquat from Hunan (Shanjingan, Dajindou and Dagujindou). The Hunan wild kumquats belong to the F. hindisii, whose trees are dwarf with small leaves, fruits like bean in size, and long thorns. While the Guangxi wild kumquat have big trees tall over 5 m, small leaves same as those of F. hindisii in size, fruit much bigger than that of F. hindisii and little small than that of F. crassifolia. The morphological characteristic differences seemed that Guangxi wild kumquat might be a new Fortunella species, obviously such suggestion needs further investigation for comfirmation.
Jinganzazhong, a Guangxi kumquat hybrid, was collected from Gupu Mountain in Hezhou, where was remote and it was hardly introduced from outsides. In fact, its SRAP profiles were distinct from those of the well-known intergeneric hybrid (Shouxingju, Sijiju and Wenzhouju). They might have different parentage in origin, and successive identification is necessary.
Though the cultivated kumquat varieties usually formed in one group in the classification indicating a close genetic relationship, 8 cultivars from Guangxi and 6 from Hunan and Zhejiang were clustered into 2 subgroups. It suggested that the cultivated kumquat in Guangxi also has certain genetic diversity and specificity with those from other provinces. Whether this differentiation is due to their genetic origin or geographic evolution effects. It is necessary to ascertain by thorough studies.
Zhu et al. suggested that the Fortunella genus consisted of two populations: cultivated kumquat and Hongkang (wild) kumquat[4]. The results in this studyrevealed that the kumquat germplasm was divided into wild and cultivated kumquat groups in Principal Coordinate Analysis (PC1) and Population Structure Analysis (at K=2). Hereby, it seemed that Fortunella may be roughly divided into wild and cultivated genotypes.
Some researchers intended to put kumquat into Citrus (Citrus japonica Thunb) [25]. Wang et al. supported the incorporation of kumquat into Citrus, but the traditional kumquat should have three species:F. venosa,F. hindsii and F. japonica [26]. In the results of this study, the germplasm of kumquat outside hybrid can be divided into wild kumquat group, golden bean group and cultivated kumquat group. To a certain extent, these results support the research results of Wang that the wild kumquat (F. hindsii) should be divided into F. venosa and F. hindsii[26]. In the present study, no sufficient data to make point on such classification idea, but the wild kumquat (F. hindsii) had very close relationship and did not appear to be able to divide into two species.
The cultivated kumquat includes three species: Luowen (F. japonica), Luofu (F. margarita) and Jindan (F. crassifolia) [27]. Zhu et al. found that there was a clear genetic structure of "F. margarita -- F. crassifolia" in cultivated kumquat. The Luowen may be originated from a cross or backcross between Luofu and Jindan, but all three cultivated species deserved the status of "species"[4]. After RAPD and CAPs molecular marker analysis of chloroplasts, Yasuda et al. suggested that the three cultivated kumquat species should be combined into one species (F. margarita complex) [28]. In this study, the cultivars derived from the Luowen, Luofu and Jindan could not be clearly divided into three species, and it could imagine that the cultivated kumquat might not enable to be divided into three species: Luowen, Luofu and Jindan, at least there is insufficient genetic information actually.
Genotype-specific marker is an efficient tool to identify germplasm resources. SRAP markers have been successfully applied to the variety identification of fruit trees such as apple [29], kiwi [30] and grape [31]. In this study, unique genotype-specific SRAP markers were detected for 14 kumquat genotypes, which made it possible to surely identify them. For the remaining 12 accessions without genotype-specific markers, they were distinguished by various combinations of markers. Kumquat is a perennial woody plant with complex genetic background. Most of the cultivated varieties are originated from bud mutation with narrow genetic background, which usually leads to the difficulty to be distinguished by molecular markers. Therefore, the present results with SRAP markers for the identification of kumquat accessions have certain importance for kumquat genetic research.
References
- Liu Binghao; Deng Chongling; Chen Chuanwu; Deng Guangzhou; Ding Ping; Niu Ying; Tang Yan; Fu Huimin. ISSR analysis of local citrus resources in Guangxi. Journal of Fruit Science. 2015, 32, 1001–1006. [Google Scholar]
- Li Guoguo; Liu Yaoxin; Chai Lijun; Ye Junli; Mai Caisheng; Ou Zhitao; Chen Xiangling. Ploidy Analysis and SSR Molecular Identification of Gui Wild Shanjingan. Southwest China Journal of Agricultural Sciences. 2017, 30, 1872–1876. [Google Scholar]
- Huang Guixiang; Guo Liying; Zhang Shuwei; He Xinhua; Zhou Ruiyang; Chen Hu; Yang Chunjiang. Genetic relationship analysis of Fortunella germplasm resources from China and Vietnam by ISSR markers. Journal of Fruit Science. 2011, 28, 563–567. [Google Scholar]
- Chenqiao Zhu; Peng Chen; Junli Ye; Hang Li; Yue Huang; Xiaoming Yang; Chuanwu Chen; Chenglei Zhang; Yuantao Xu; Xiaoli Wang; Xiang Yan; Guangzhou Deng; Xiaolin Jiang; Nan Wang; Hongxing Wang; Quan Sun; Yun Liu; Di Feng; Min Yu; Xietian Song; Zongzhou Xie; Yunliu Zeng; Lijun Chai; Qiang Xu; Chongling Deng; Yunjiang Cheng; Xiuxin Deng. New insights into the phylogeny and speciation of kumquat (Fortunella spp.) based on chloroplast SNP, nuclear SSR and whole-genome sequencing. Front. Agr. Sci. Eng. 2022, 9, 627–641. [Google Scholar] [CrossRef]
- Zhang Zhonghai; Zhang Anshi; Gao Dengtao; Wei Zhifeng. Genetic diversity analysis and rapid identification of Grapevine Jufeng Cultivars by MCID method. Journal of Northeast Agricultural Sciences. 2022, 4, 38–42. [Google Scholar] [CrossRef]
- Yan Heqin; Zheng Wei; Dai Jiani; Yan Wuping; Yu Jing; Wu Yougen. Based on camellia species genetic diversity analysis of a molecular marker hainan. Journal of molecular plant breeding. 2022, 20, 1901–1908. [Google Scholar] [CrossRef]
- Zhang Yu; Huang Guodi; Mo Yonglong; Luo Shixing; Zhao Ying; Tang Yujuan; Lu Zushuang; Shan Bin; Rong Tao. CDDP combination of a tag analysis Mang fruit species genetic diversity. South China Fruits. 2022, 2, 57–63. [Google Scholar] [CrossRef]
- Sun Lili; Peng Lina; Li Zheng; Hou Ruining; Mou Yunhui. Using ISSR and mark build li genetic linkage map. Journal of guangdong agricultural sciences. 2022, 49, 40–48. [Google Scholar] [CrossRef]
- Jiang Wu; Li Yaping; Chen Jiadong; Tao Zhengming. Study on Genetic diversity of Polygonum sativum based on ISSR and SRAP markers [J/OL]. Chinese Traditional and Herbal Drugs 2022, 1–9. Available online: http:// http://fffg208e51c2dd88406685526280e50de659hn95vov6v6nc96vfw.fgfy.www.gxstd.com/kcms/detail/12.1108.R.20220929.1912.006. /.
- Wen Zhang; Wei Hu; Xin-Yu Zhang; Min Zhou; Qiao-Qiao Jiang; Zn-Niu Deng; Da-Zhi Li. Identification of the hybrid progeny of Shatian Pomelo × citron by embryo rescue technique and its SRAP detection. Journal of Fruit Science. 2013, 30, 386–389. [Google Scholar]
- Zhang Lihua; Han Haozhang; Wang Xiaoli; Li Suhua; Wang Fang; Dong Rong; Liu Yu. Screening of molecular markers for SRAP of Cinnamomum camphora. Anhui Agricultural Science Bulletin. 2019, 25, 25–27. [Google Scholar] [CrossRef]
- Jing Xu; Limei Tan; Hongyan Fu; Zhimei Zhu; Libing Long; Zhe Hu; Xianfeng Ma; Ziniu Deng. Molecular markers were used to analyze the diversity of 14 citron accessions. Molecular plant breeding. http://kns.cnki.net/kcms/detail/46.1068.S.20200401.1548.004.
- Jaccard, P. Nouvelles recherches sur la distribution florale. Bull. Soc. Vaud. Sci. Nat. 1908, 44, 223–270. [Google Scholar]
- Hammer, Ø.; Harper, D.A.; Ryan, P.D. Past: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Al-Ghamedi, K.; Alaraidh, I.; Afzal, M.; Mahdhi, M.; Al-Faifi, Z.; Oteef, M.D.Y.; Tounekti, T.; Alghamdi, S.S.; Khemira, H. Assessment of Genetic Diversity of Local Coffee Populations in Southwestern Saudi Arabia Using SRAP Markers. Agronomy 2023, 13, 302. [Google Scholar] [CrossRef]
- Pritchard J K, Stephens M J, Donnelly P J. Inference of Population Structure Using Multilocus Genotype Data[J]. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef]
- Earl D A, Vonholdt B M. STRUCTURE HARVESTER: a website and program for visualizing STRUCTURE output and implementing the Evanno method[J]. Conservation Genetics Resources 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Chown, S.L.; Hodgins, K.A.; Griffin, P.C.; Oakeshott, J.G.; Byrne, M.; Hoffmann, A.A. Biological invasions, climate change, and genomics. In Crop Breeding: Bioinformatics and Preparing for Climate Change; Santosh, K., Ed.; Apple Academic Press: Waretown, NJ, USA, 2016. [Google Scholar]
- Al-Murish, T.M.; Elshafei, A.A.; Al-Doss, A.A.; Barakat, M.N. Genetic diversity of coffee (Coffea arabica L.)in Yemen via SRAP, TRAP and SSR markers. J. Food Agric. Environ. 2013, 11, 411–416. [Google Scholar]
- Hasan, N.; Choudhary, S.; Naaz, N.; Sharma, N.; Laskar, R.A. Recent advancements in molecular marker-assisted selection and applications in plant breeding programmes. J. Genet. Eng. Biotechnol. 2021, 19, 128. [Google Scholar] [CrossRef] [PubMed]
- Nadeem, M.A.; Nawaz, M.A.; Shahid, M.Q.; Do ˘gan, Y.; Comertpay, G.; Yıldız, M.; Hatipo ˘glu, R.; Ahmad, F.; Alsaleh, A.; Labhane, N. DNA molecular markers in plant breeding: Current status and recent advancements in genomic selection and genome editing. Biotechnol. Biotechnol. Equip. 2018, 32, 261–285. [Google Scholar] [CrossRef]
- Al-Ghamedi, K.; Alaraidh, I.; Afzal, M.; Mahdhi, M.; Al-Faifi, Z.; Oteef, M.D.Y.; Tounekti, T.; Alghamdi, S.S.; Khemira, H. Assessment of Genetic Diversity ofLocal Coffee Populations in Southwestern Saudi Arabia Using SRAP Markers. Agronomy 2023, 13, 302. [Google Scholar] [CrossRef]
- Zhang Kun, Zhou Yuanjie, Li Yao, Liu Xinling, Guo Yuqi, Xia Hui, Liang Dong. Genetic Diversity Analysis of Kiwifruit Germplasm based on SRAP and SCoT markers [J]. Journal of Fruit Science 2021, 38, 2059–2071. [Google Scholar] [CrossRef]
- Fuchu Hu, Xiaobo Wu, Zhe Chen, Fengzhi Wu, Wenjing Zhou, Xuejie Feng, Hongyan Fan, Ruiyun Zhou, Xianghe Wang. Genetic diversity Analysis of Litchi germplasm resources based on SRAP molecular markers [J]. Journal of Tropical Crops 2021, 42, 920–926. [Google Scholar]
- Editorial Committee of Flora of China, Chinese Academy of Sciences. Flora of China, Kumquat [M]. Beijing: Science Press, 2004. http://www.iplant.cn/frps/cname/J.
- Wang Ting,Chen LingLing,Shu HuiJuan,You Fang,Liang XiaoLi,Li Jun,Ren Jing,Wanga Vincent Okelo,Mutie Fredrick Munyao,Cai XiuZhen,Liu KeMing,Hu GuangWan. Fortunella venosa (Champ. ex Benth.) C. C. Huang and F. hindsii (Champ. ex Benth.) Swingle as Independent Species: Evidence From Morphology and Molecular Systematics and Taxonomic Revision of Fortunella(Rutaceae)[J]. Front Plant Sci 2022, 13, 867659. [Google Scholar] [CrossRef] [PubMed]
- Zhou Kailong, Ye Yinmin. Chinese fruit trees citrus Roll [M]. Beijing: Beijing Forestry Publishing House,2010:126-130,427-433.
- Yasuda, K., M. Yahata, H. Komatsu and H. Kunitake. Phylogeny and classification of Fortunella (Aurantioideae) inferred from DNA polymorphisms. Bul. Fac. Agr. Univ. Miyazaki 2010, 56, l03–110. [Google Scholar]
- Li Huifeng, Ran Kun, Wang Tao. Construction of fingerprint of apple resources in Shandong Province by using SRAP markers [J]. Journal of Shenyang Agricultural University 2020, 51, 470–475. [Google Scholar]
- Anshi Zhang, Qingliang Si, Xiujuan Qi, Zhonghai Zhang. Genetic diversity Analysis and fingerprint Construction of Germplasm resources of Kiwifruit [J]. Jiangsu Agricultural Journal 2018, 34, 138–144. [Google Scholar]
- Shang Xiaoxing, Zhang Anshi, Liu Ying, Gao Dengtao. Genetic diversity Analysis and fingerprint Construction of Grapevine Germplasm resources based on SRAP [J]. Molecular plant breeding 2020, 18, 1916–1922. [Google Scholar] [CrossRef]
Figure 1.
PCR amplification products with SRAP primers. Note: PCR with SRAP primersME20+EM2; M: DNA Marker; 1-26: kumquat genotypes listed in Table 1.
Figure 1.
PCR amplification products with SRAP primers. Note: PCR with SRAP primersME20+EM2; M: DNA Marker; 1-26: kumquat genotypes listed in Table 1.
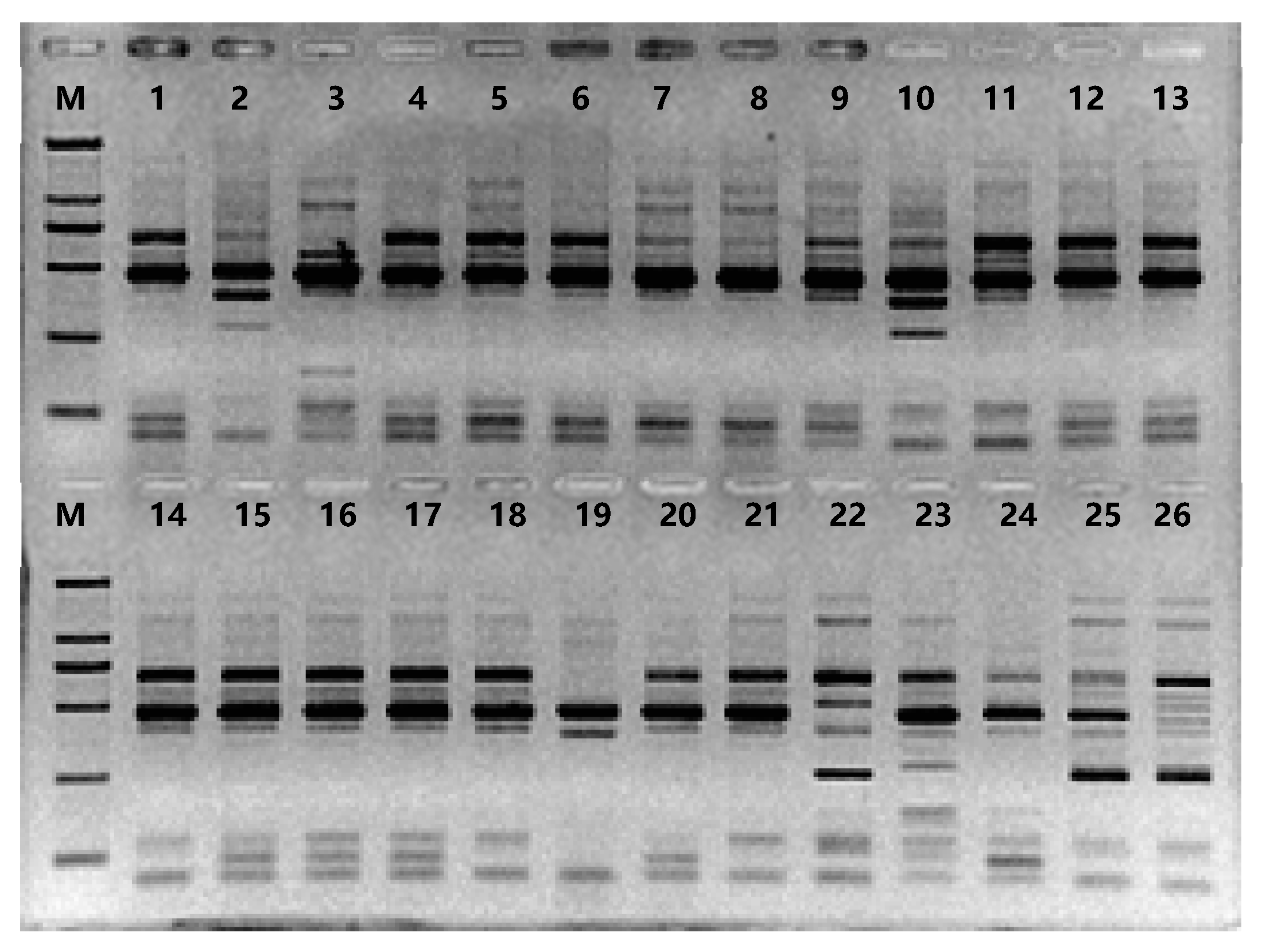
Figure 2.
Biplot analysis of kumquats’ diversity as inferred from SRAP marker data. Note: The genotype code listed in Table 1.
Figure 2.
Biplot analysis of kumquats’ diversity as inferred from SRAP marker data. Note: The genotype code listed in Table 1.
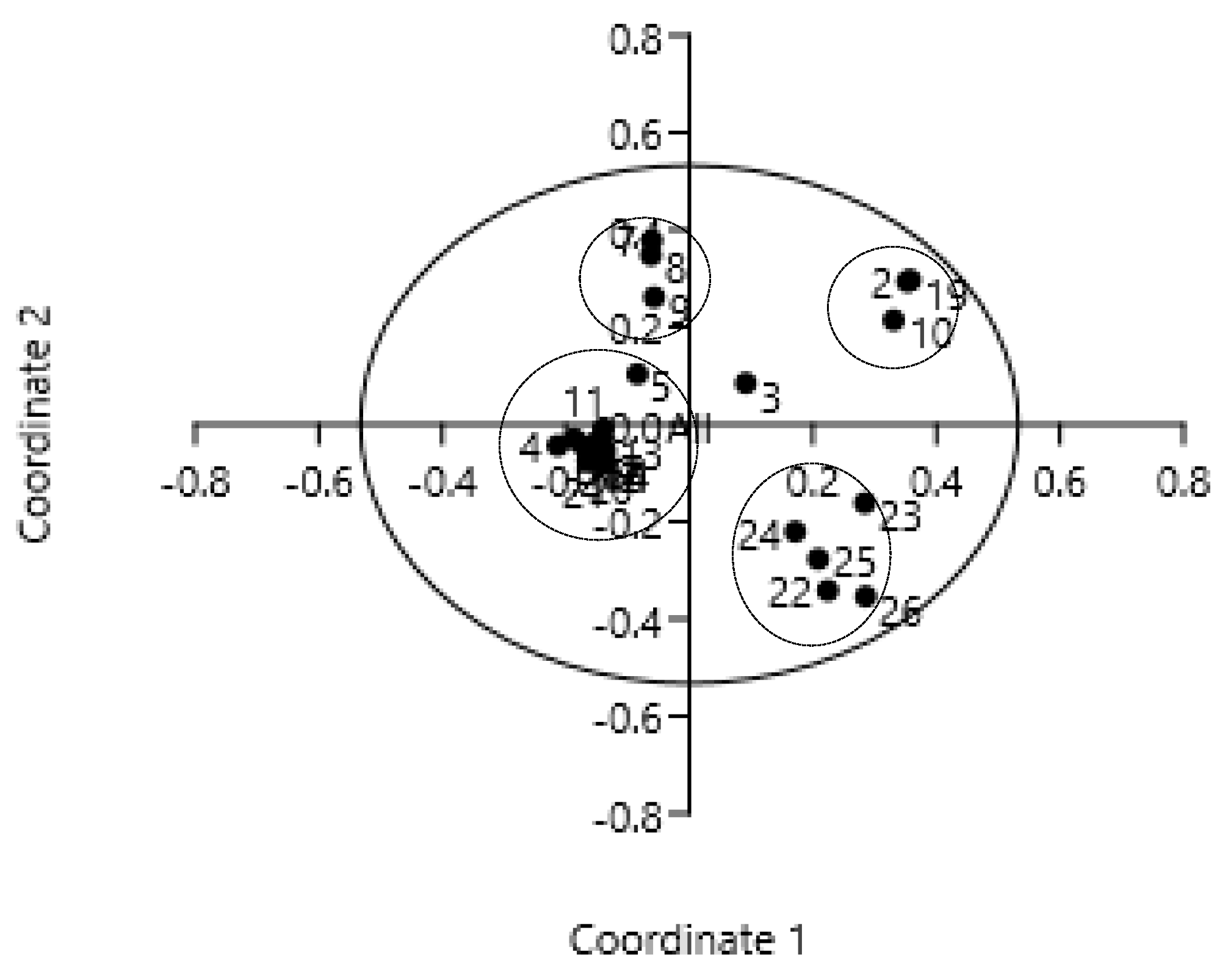
Figure 3.
The number of K clusters (1-10) generated from ninteen SRAP primer combinations.
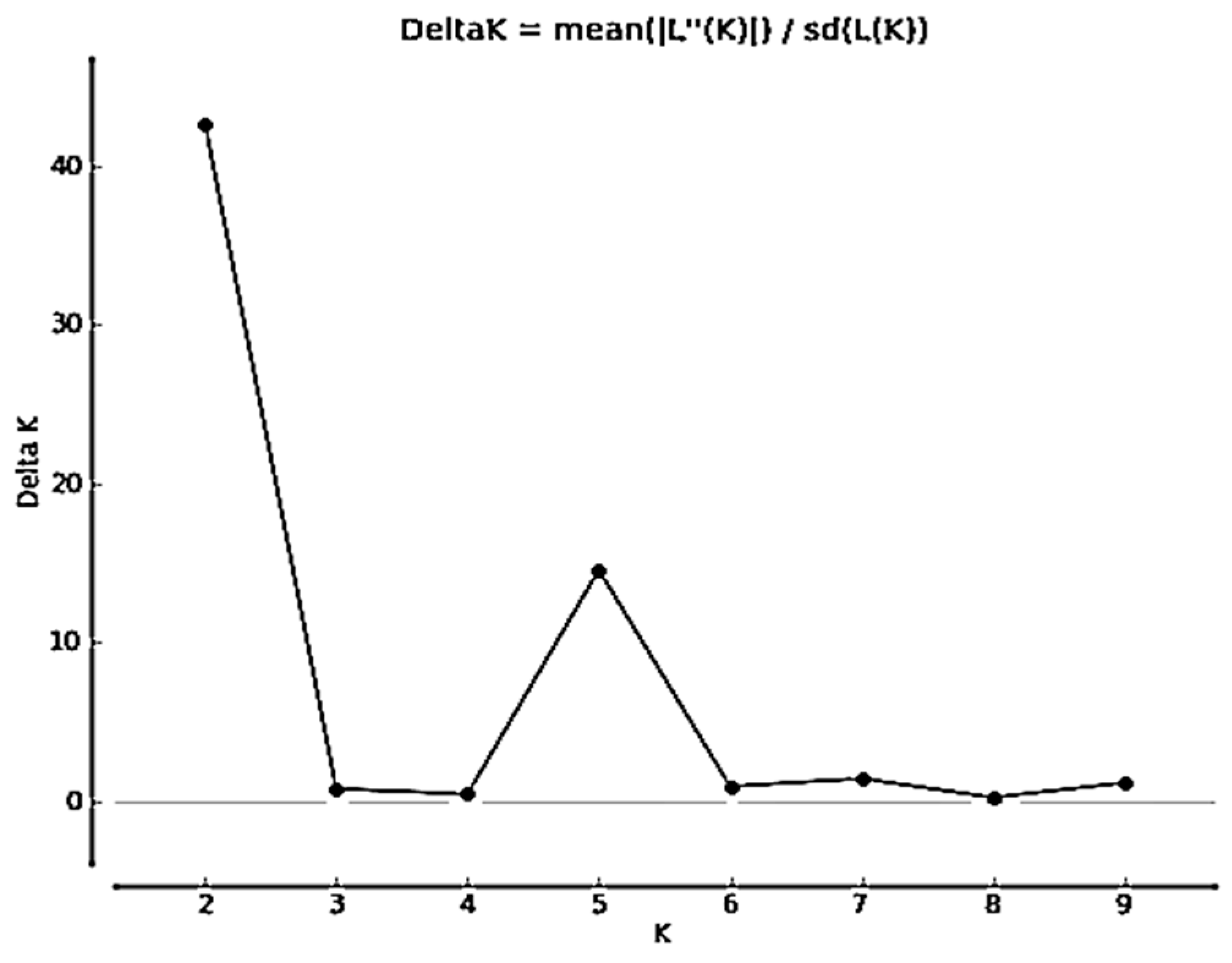
Figure 4.
Population structure analysis at K=2 using SRAP markers’ data of 26 kumquat genotypes. Note: The genotype code listed in Table 1.
Figure 4.
Population structure analysis at K=2 using SRAP markers’ data of 26 kumquat genotypes. Note: The genotype code listed in Table 1.
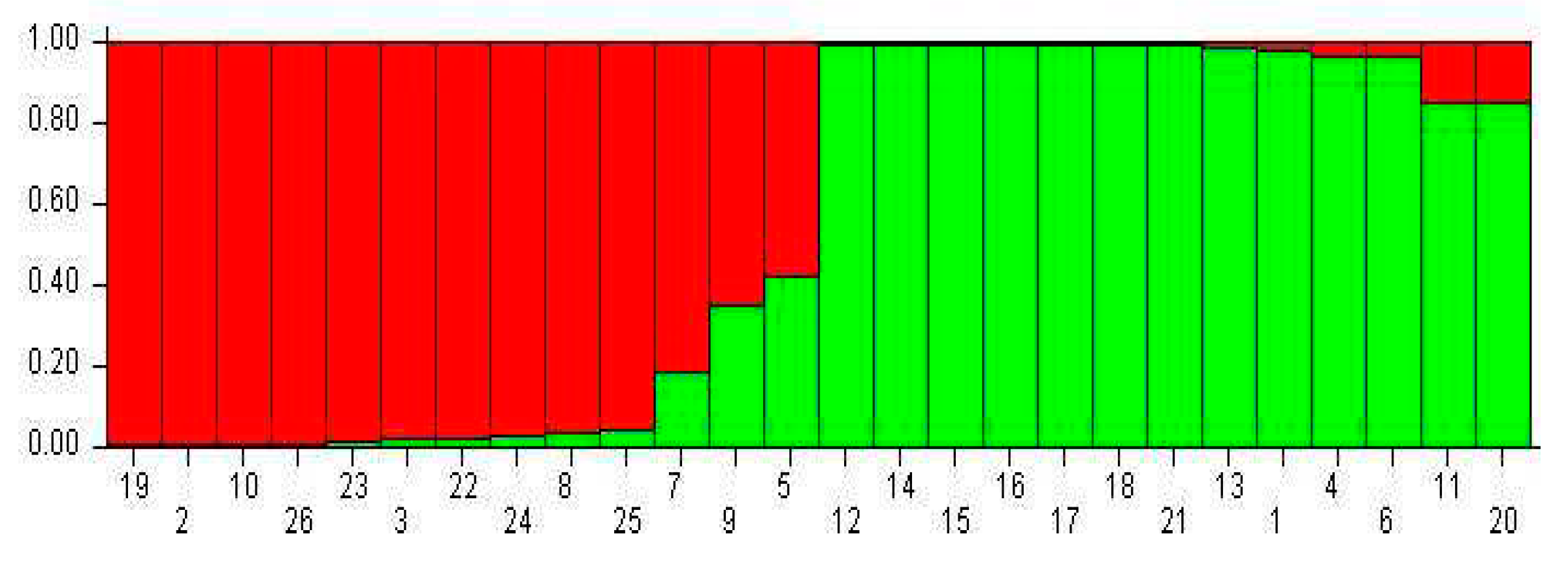
Figure 5.
Structure analysis at K=5 using SRAP markers’ data on 26 kumquat genotypes. Note: The genotype code listed in Table 1.
Figure 5.
Structure analysis at K=5 using SRAP markers’ data on 26 kumquat genotypes. Note: The genotype code listed in Table 1.
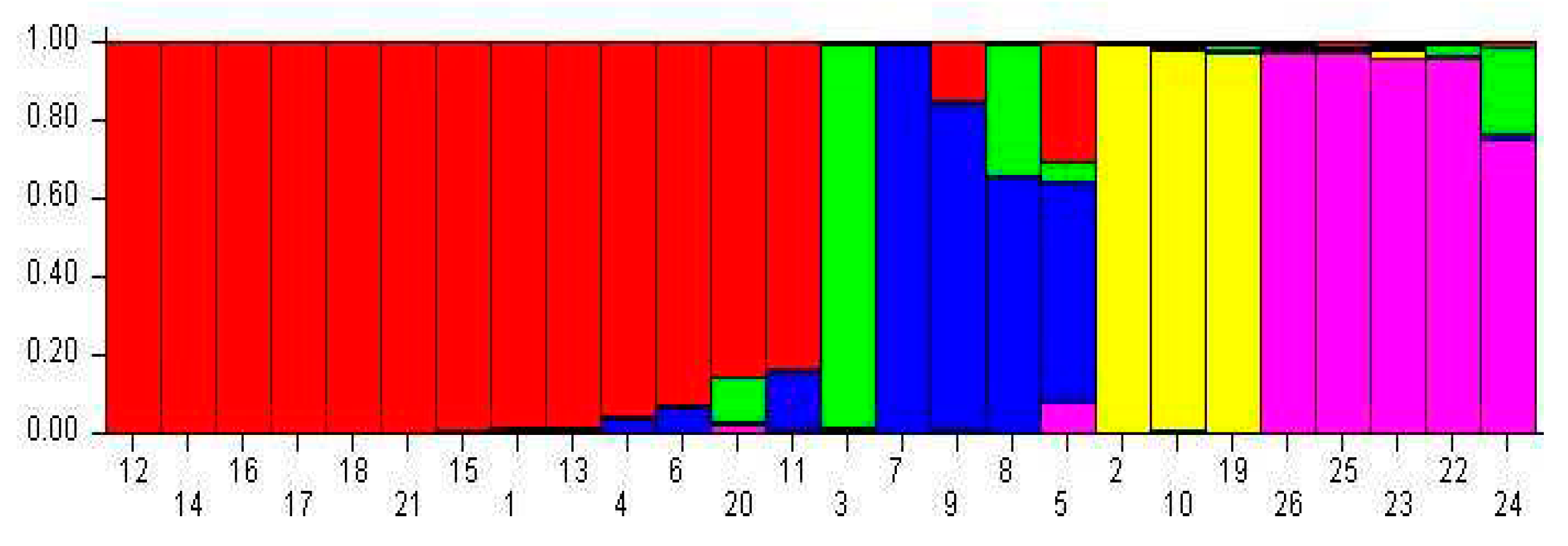
Figure 6.
Dendrogram 26 kumquats generated by Jaccard coefficient and UPGMA clustering method based on SRAP molecular markers. Note: The genotype code listed in Table 1; The numbers in front of the branches are bootstrap values.
Figure 6.
Dendrogram 26 kumquats generated by Jaccard coefficient and UPGMA clustering method based on SRAP molecular markers. Note: The genotype code listed in Table 1; The numbers in front of the branches are bootstrap values.
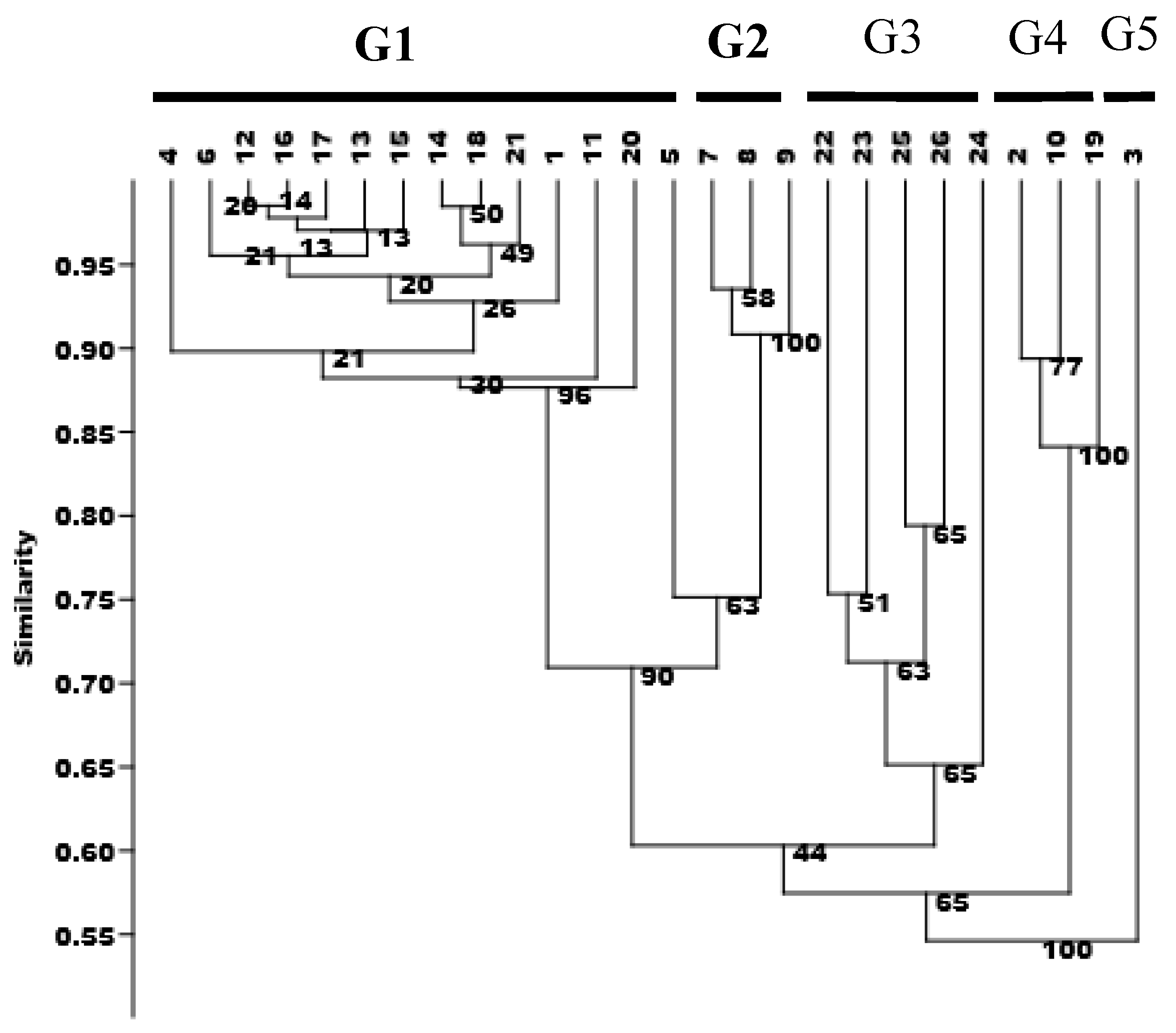

Table 1.
The tested kumquat genotypes.
| Code | Abbreviation | Genotype Name | Scientific name | Possible origin |
|---|---|---|---|---|
| 1 | NB jindan | Ningbo jindan | F. crassifolia | Ningbo, Zhejiang |
| 2 | Daguojindou | Daguojindou | F. hindsii | Citrus Research Institute, SWU/CAAS |
| 3 | Jinganzazhong | Guangxi natural kumquat hybrid | Citrus x Fortunella | Hezhou, Guangxi |
| 4 | WZ luofu | Wenzhou luofu | F. margarita | Wenzhou, Zhejiang |
| 5 | NB luofu | Ningbo luofu | F. margarita | Ningbo, Zhejiang |
| 6 | WZ jingdan | Wenzhou jingdan | F. crassifolia | Wenzhou, Zhejiang |
| 7 | Sijiju | Sijiju | Citrus x Fortunella | Citrus Research Institute, SWU/CAAS |
| 8 | Wenzhouju | Wenzhouju (kumquat hybrid) | Citrus x Fortunella | Wenzhou, Zhejiang |
| 9 | Shouxingju | Shouxingju (kumquat hybrid) | Citrus x Fortunella | Citrus Research Institute, SWU/CAAS |
| 10 | Dajindou | Dajindou | F. hindsii | Citrus Research Institute, SWU/CAAS |
| 11 | NB luowen | Ningbo luowen | F. japonica | Ningbo, Zhejiang |
| 12 | RA jingan | Rongan jingan | F. crassifolia | Liuzhou, Guangxi |
| 13 | FY jingan | Fuyuan jingan | F. crassifolia | Liuzhou, Guangxi |
| 14 | CM jingan | Cuimi jingan | F. crassifolia | Liuzhou, Guangxi |
| 15 | Guijingan1 | Guijingan No.1 | F. crassifolia | Yangshuo, Guangxi |
| 16 | Guijingan2 | Guijingan No.2 | F. crassifolia | Yangshuo, Guangxi |
| 17 | YS jingan | Yangshuo jingan | F. crassifolia | Yangshuo, Guangxi |
| 18 | F15-1 | F15-1 | F. crassifolia | Liuzhou, Guangxi |
| 19 | Shanjingan | Hunan Shanjingan | F. hindsii | Changsha, Hunan |
| 20 | LY jingan | Liuyang jingan | F. crassifolia | Changsha, Hunan |
| 21 | HP jingan | Huapi jingan | F. crassifolia | Liuzhou, Guangxi |
| 22 | FC-1 | Guangxi wild kumquat FC-1 | Fortunella sp. | Fangchenggang, Guangxi |
| 23 | FC-2 | Guangxi wild kumquat FC-2 | Fortunella sp. | Fangchenggang, Guangxi |
| 24 | FC-3 | Guangxi wild kumquat FC-3 | Fortunella sp. | Fangchenggang, Guangxi |
| 25 | FC-4 | Guangxi wild kumquat FC-4 | Fortunella sp. | Fangchenggang, Guangxi |
| 26 | FC-5 | Guangxi wild kumquat FC-5 | Fortunella sp. | Fangchenggang, Guangxi |
Note: SWU: Southwest University; CAAS: Chinese Academy of Agricultural Sciences.
Table 2.
SRAP primers used in this study.
| Primer name | Primer sequence (5’-3’) | Primer name | Primer sequence (5’-3’) |
|---|---|---|---|
| ME1 | TGAGTCCAAACCGGAAA | EM1 | GACTGCGTACGAATTAAC |
| ME2 | TGAGTCCAAACCGGAAC | EM2 | GACTGCGTACGAATTAAT |
| ME3 | TGAGTCCAAACCGGAAG | EM3 | GACTGCGTACGAATTACG |
| ME4 | TGAGTCCAAACCGGAAT | EM4 | GACTGCGTACGAATTAGC |
| ME5 | TGAGTCCAAACCGGACA | EM5 | GACTGCGTACGAATTATG |
| ME6 | TGAGTCCAAACCGGACC | EM6 | GACTGCGTACGAATTCAA |
| ME7 | TGAGTCCAAACCGGACG | EM7 | GACTGCGTACGAATTCAC |
| ME8 | TGAGTCCAAACCGGACT | EM8 | GACTGCGTACGAATTCAG |
| ME9 | TGAGTCCAAACCGGAGA | EM9 | GACTGCGTACGAATTCAT |
| ME10 | TGAGTCCAAACCGGAGC | EM10 | GACTGCGTACGAATTCCA |
| ME11 | TGAGTCCAAACCGGAGG | EM11 | GACTGCGTACGAATTCGA |
| ME12 | TGAGTCCAAACCGGATA | EM12 | GACTGCGTACGAATTCGG |
| ME13 | TGAGTCCAAACCGGTAA | EM13 | GACTGCGTACGAATTCTA |
| ME14 | TGAGTCCAAACCGGTAG | EM14 | GACTGCGTACGAATTCTC |
| ME15 | TGAGTCCAAACCGGTCA | EM15 | GACTGCGTACGAATTCTG |
| ME16 | TGAGTCCAAACCGGTCC | EM16 | GACTGCGTACGAATTCTT |
| ME17 | TGAGTCCAAACCGGTGC | EM17 | GACTGCGTACGAATTGAT |
| ME18 | TGAGTCCAAACCGGTGT | EM18 | GACTGCGTACGAATTGCA |
| ME19 | TGAGTCCAAACCGGTTA | EM19 | GACTGCGTACGAATTGGT |
| ME20 | TGAGTCCAAACCGGTTG | EM20 | GACTGCGTACGAATTGTC |
| EM21 | GACTGCGTACGAATTTAG | ||
| EM22 | GACTGCGTACGAATTTCG | ||
| EM23 | GACTGCGTACGAATTTGA | ||
| EM24 | GACTGCGTACGAATTTGC |
Table 3.
Analysis of the polymorphism detected using SRAP primers.
| No. | Primer | Amplified bands | Polymorphic bands | Polymorphic rate (%) |
|---|---|---|---|---|
| 1 | Me1Em15 | 6 | 4 | 66.67 |
| 2 | Me1Em22 | 4 | 4 | 100.00 |
| 3 | Me1Em23 | 6 | 6 | 100.00 |
| 4 | Me2Em17 | 6 | 6 | 100.00 |
| 5 | Me9Em23 | 7 | 4 | 57.14 |
| 6 | Me2Em21 | 6 | 5 | 83.33 |
| 7 | Me10Em7 | 10 | 8 | 80.00 |
| 8 | Me4Em7 | 1 | 1 | 100.00 |
| 9 | Me4Em12 | 10 | 8 | 80.00 |
| 10 | Me3Em17 | 4 | 4 | 100.00 |
| 11 | Me4Em17 | 4 | 4 | 100.00 |
| 12 | Me11Em21 | 2 | 2 | 100.00 |
| 13 | Me10Em13 | 4 | 4 | 100.00 |
| 14 | Me14Em12 | 6 | 6 | 100.00 |
| 15 | Me16Em19 | 4 | 2 | 50.00 |
| 16 | Me7Em4 | 10 | 8 | 80.00 |
| 17 | Me20Em2 | 5 | 5 | 100.00 |
| 18 | Me17Em2 | 2 | 2 | 100.00 |
| 19 | Me18Em22 | 5 | 5 | 100.00 |
| Sum/Average | 102/5.37 | 88/4.63 | 86.27 | |
Table 4.
Eigenvalues of Principal Coordinates Analysis (PCoA).
| Axis | Eigenvalue | Cumulative Eigenvalue | Percent (%) | Cumulative (%) |
|---|---|---|---|---|
| 1 | 0.53 | 0.53 | 34.80 | 34.80 |
| 2 | 0.29 | 0.83 | 19.25 | 54.05 |
| 3 | 0.19 | 1.02 | 12.74 | 66.78 |
| 4 | 0.17 | 1.19 | 10.90 | 77.68 |
| 5 | 0.09 | 1.28 | 5.67 | 83.35 |
| 6 | 0.08 | 1.36 | 5.38 | 88.72 |
| 7 | 0.03 | 1.39 | 2.10 | 90.82 |
| 8 | 0.02 | 1.41 | 1.61 | 92.43 |
| 9 | 0.02 | 1.43 | 1.34 | 93.76 |
| 10 | 0.02 | 1.45 | 1.01 | 94.78 |
| 11 | 0.01 | 1.46 | 0.78 | 95.56 |
| 12 | 0.01 | 1.47 | 0.52 | 96.08 |
| 13 | 0.00 | 1.47 | 0.24 | 96.32 |
| 14 | 0.00 | 1.48 | 0.11 | 96.42 |
| 15 | 0.00 | 1.48 | 0.00 | 96.43 |
| 16 | 0.00 | 1.48 | 0.00 | 96.43 |
Table 5.
Genetic similarity coefficients based on the SRAP markers of all the tested genotypes.
| No. | Germplasm | 1.NB jindan 2.Daguojindou 3.Jinganzazhong 4.WZ luofu5.NB luofu 6.WZ jingdan 7.Sijiju 8.Wenzhouju 9.Shouxingju 10.Dajindou 11.NB luowen 12.RA jingan 13.FY jingan 14.CM jingan 15.Guijingan116.Guijingan2 17.YS jingan18.F15-1 19.Shanjingan 20.LY jingan 21.HP jingan 22.FC14-1 23.FC14-2 24.FC14-3 25.FC14-4 26.FC14-5 |
|---|---|---|
| 1 | NB jindan | 1.000 |
| 2 | Daguojindou | 0.634 1.000 |
| 3 | Jinganzazhong | 0.634 0.604 1.00 |
| 4 | WZ luofu | 0.941 0.634 0.594 1.000 |
| 5 | NB luofu | 0.812 0.6436 0.584 0.851 1.000 |
| 6 | WZ jingdan | 0.941 0.634 0.653 0.921 0.812 1.000 |
| 7 | Sijiju | 0.762 0.673 0.634 0.762 0.851 0.782 1.000 |
| 8 | Wenzhouju | 0.703 0.594 0.584 0.703 0.772 0.703 0.871 1.000 |
| 9 | Shouxingju | 0.822 0.673 0.673 0.782 0.831 0.822 0.941 0.851 1.000 |
| 10 | Dajindou | 0.663 0.931 0.594 0.644 0.673 0.663 0.663 0.584 0.703 1.000 |
| 11 | NB luowen | 0.911 0.663 0.644 0.911 0.822 0.911 0.772 0.723 0.832 0.693 1.000 |
| 12 | RA jingan | 0.960 0.653 0.653 0.941 0.812 0.980 0.762 0.703 0.822 0.683 0.931 1.000 |
| 13 | FY jingan | 0.950 0.663 0.663 0.931 0.802 0.970 0.752 0.693 0.812 0.693 0.921 0.990 1.000 |
| 14 | CM jingan | 0.941 0.673 0.653 0.921 0.792 0.960 0.743 0.683 0.802 0.703 0.931 0.980 0.970 1.000 |
| 15 | Guijingan1 | 0.950 0.644 0.644 0.950 0.822 0.970 0.772 0.693 0.812 0.673 0.921 0.990 0.980 0.970 1.000 |
| 16 | Guijingan2 | 0.970 0.644 0.663 0.931 0.802 0.970 0.772 0.712 0.832 0.673 0.921 0.990 0.980 0.970 0.980 1.000 |
| 17 | YS jingan | 0.960 0.653 0.653 0.941 0.792 0.960 0.782 0.723 0.822 0.663 0.911 0.980 0.970 0.960 0.970 0.990 1.000 |
| 18 | F15-1 | 0.931 0.683 0.644 0.931 0.782 0.950 0.752 0.693 0.792 0.693 0.921 0.970 0.960 0.990 0.960 0.960 0.970 1.000 |
| 19 | Shanjingan | 0.644 0.911 0.594 0.644 0.653 0.624 0.683 0.614 0.683 0.881 0.653 0.644 0.653 0.663 0.634 0.653 0.663 0.673 1.000 |
| 20 | LY jingan | 0.941 0.634 0.634 0.921 0.772 0.901 0.743 0.703 0.782 0.644 0.871 0.921 0.911 0.901 0.911 0.931 0.941 0.911 0.644 1.000 |
| 21 | HP jingan | 0.960 0.653 0.653 0.921 0.792 0.941 0.743 0.693 0.802 0.683 0.931 0.960 0.950 0.980 0.950 0.970 0.960 0.970 0.663 0.921 1.000 |
| 22 | FC14-1 | 0.683 0.634 0.574 0.644 0.634 0.683 0.584 0.525 0.644 0.663 0.693 0.703 0.713 0.703 0.693 0.713 0.703 0.693 0.644 0.6434 0.703 1.000 |
| 23 | FC14-2 | 0.703 0.713 0.554 0.663 0.693 0.683 0.683 0.614 0.723 0.743 0.713 0.703 0.713 0.703 0.693 0.713 0.703 0.693 0.723 0.663 0.703 0.822 1.000 |
| 24 | FC14-3 | 0.693 0.644 0.683 0.673 0.743 0.713 0.653 0.604 0.693 0.653 0.723 0.713 0.723 0.713 0.703 0.703 0.693 0.703 0.634 0.673 0.713 0.713 0.772 1.000 |
| 25 | FC14-4 | 0.762 0.693 0.614 0.723 0.733 0.723 0.663 0.614 0.683 0.723 0.733 0.743 0.733 0.743 0.733 0.752 0.743 0.733 0.703 0.762 0.762 0.802 0.822 0.772 1.000 |
| 26 | FC14-5 | 0.683 0.653 0.594 0.663 0.673 0.644 0.604 0.545 0.624 0.683 0.653 0.663 0.673 0.663 0.653 0.673 0.683 0.673 0.663 0.703 0.683 0.762 0.782 0.733 0.861 1.000 |
Table 6.
The unique kumquat genotype-specific markers.
| Code | Genotypes | Unique specific markers |
|---|---|---|
| 1 | NB jindan | M1E15(1800-), M3E17(250-) |
| 2 | Daguojindou | M1E23(400-) |
| 3 | Jinganzazhong | M1E22(250+), M4E17(500+), M9E23(700-), M10E13(200+) |
| 4 | WZ luofu | None |
| 5 | NB luofu | M2E21(400-), M3E17(450+), M11E21(250+) |
| 6 | WZ jingdan | M2E21(450+), M16E9(350-) |
| 7 | Sijiju | M4E12(1300-) |
| 8 | Wenzhouju | M17E2(530-) |
| 9 | Shouxingju | None |
| 10 | Dajindou | M3E17(100+), M4E17(250-) |
| 11 | NB luowen | none |
| 12 | RA jingan | None |
| 13 | FY jingan | None |
| 14 | CM jingan | None |
| 15 | Guijingan1 | None |
| 16 | Guijingan2 | None |
| 17 | YS jingan | None |
| 18 | F15-1 | None |
| 19 | Shanjingan | M1E22(300-) |
| 20 | LY jingan | M7E4(500+) |
| 21 | HP jingan | None |
| 22 | FC-1 | M2E17(500+), M2E17(1000+) |
| 23 | FC-2 | M4E7(500-) |
| 24 | FC-3 | None |
| 25 | FC-4 | M4E17(300+) |
| 26 | FC-5 | M14E12(1300+) |
Note: in parentheses + meant the presence of the band, - absence of the band and the number indicated band bp.
Table 7.
The combinations of specific markers for kumquat identification.
| Genotypes | Common specific markers | Combination of unique markers |
|---|---|---|
| FC-3 | M3E17(180+) FC-2; FC-4; FC-5 | FC-2 M4E7(500-); FC-4 M4E17(300+); FC-5 M14E12(1300+) |
| M1E23(500+) Daguojindou | Daguojindou M1E23(400-) | |
| Shouxingju | M2E21(400+) Sijiju | Sijiju M4E12(1300-) |
| M20E2(700+) Wenzhouju | Wenzhouju M17E2(530-) | |
| NB luowen | M10E7(850-) WZ Luofu; NB luofu; Wenzhouju | WZ Luofu M10E7(320-); NB luofu M2E21(400-), M3E17(450+), M11E21(250+); Wenzhouju M17E2(530-) |
| WZ Luofu | M10E7(320-) NB luofu; Wenzhouju; Shanjingan | NB luofu M2E21(400-), M3E17(450+), M11E21(250+); Wenzhouju M17E2(530-); Shanjingan M1E22(300-) |
Note: in parentheses + meant the presence of the band, - absence of the band and the number indicated band bp.
Table 8.
Guangxi cultivars group specific markers for identification.
| Group markers | Shared genotypes with | Unique markers for discrimination |
|---|---|---|
| M1E23(800-) | NB jindan, WZ luofu, WZ jingdan LY jingan |
NB jindan M1E15(1800-), M3E17(250-) WZ Luofu M10E7(320-) WZ jingdan M2E21(450+), M16E9(350-) LY jingan M7E4(500+) |
| M7E4(1050+) | NB jindan NB luofu WZ jingdan LY jingan |
NB jindan M1E15(1800-), M3E17(250-); NB Luofu M2E21(400-), M3E17(450+), M11E21(250+) WZ jingdan M2E21(450+), M16E9(350-) LY jingan M7E4(500+) |
Table 9.
Genotype identification within Guangxi cultivars group.
| Marker types | Genotypes | Markers for discrimination |
|---|---|---|
| Single marker | Guijingan1 | M1E22(740-) |
| HP jingan | M10E7(710-) | |
| FY jingan | M3E17(90-) | |
| Bi/Tri- markers | CM jingan + HP jingan | M2E21(480-) |
| YS jingan + F15-1 | M14E12(760-) | |
| Guijingan2 + YS jingan+ HP jingan | M14E12(700-) | |
| No specific marker | RA jingan |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



