
Submitted:
31 May 2025
Posted:
02 June 2025
You are already at the latest version
Abstract
Background:
Medicinal herbs are increasingly used as functional foods and therapeutics, yet their biological effects and potential toxicity remain incompletely understood. Using Caenorhabditis elegans, we previously identified several herbal extracts with potent cytotoxic effects.
Methods:
We focused on three herbal extracts—Mentha longifolia, Scrophularia orientalis, and Echium biebersteinii—to investigate their impact on germline development and fertility. Phenotypic analyses, apoptosis quantification, qRT-PCR, and LC-MS profiling were performed to assess cytotoxicity, meiotic defects, DNA damage responses, and chemical composition.
Results:
All three extracts significantly reduced worm survival, induced larval arrest, and increased the high incidence of males phenotype, suggesting chromosomal mis-segregation. Germline defects included disorganized nuclei, altered meiotic progression, and reduced bivalent formation. These were accompanied by upregulation of DNA damage checkpoint genes, increased pCHK-1 foci, and elevated apoptosis in the pachytene region. We identified 21 major compounds in three herb extracts, with four shared across all extracts. Among them, thymol and carvyl acetate activated checkpoint responses and apoptosis, while thymol and luteolin-7-O-rutinoside exhibited antioxidant activity.
Conclusion:
These findings emphasize the value of dissecting complex herbal mixtures to uncover specific bioactive compounds, which is essential for evaluating both the therapeutic potential and safety risks of medicinal plants. Also, this study underscores the need to analyze individual phytochemicals within herbal mixtures to understand their distinct biological effects. Such insight is essential for evaluating both the therapeutic potential and safety risks of medicinal plants used in food and supplements.
Keywords:
Mentha longifolia
; Scrophularia orientalis
; and Echium biebersteinii
; DNA repair
; meiosis
; germline development
; medicinal plants
Introduction
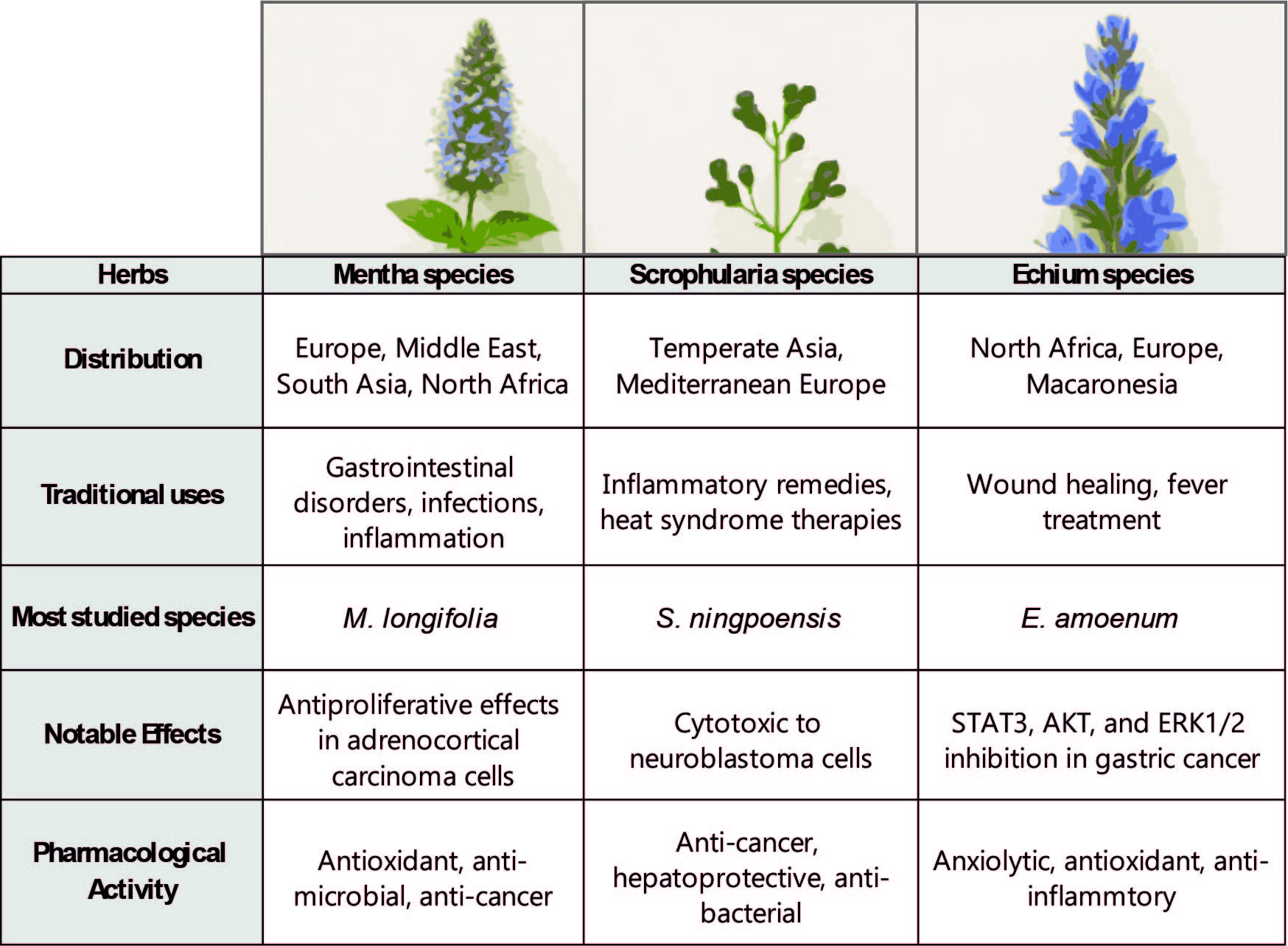
While medicinal plants have long been used in traditional remedies, their molecular effects—particularly on genome stability and reproductive health—remain poorly understood. This gap is especially significant given the rising use of herbal formulations with uncharacterized toxicological profiles. To address this gap, we investigated three herbal species native to Armenia—Scrophularia orientalis (S. orientalis), Mentha longifolia (M. longifolia), and Echium biebersteinii (E. biebersteinii). All three are known for their strong bioactivity (Figure 1 and Table 1), but their potential roles in DNA damage repair and apoptosis have not been fully validated. We hypothesized that extracts from these herbs may interfere with DNA repair pathways and germline development due to their bioactive components.
1. Scrophularia Orientalis
General Overview
The genus Scrophularia, belonging to the family Scrophulariaceae, comprises approximately 200–300 species distributed primarily across temperate regions of Asia, Europe (particularly the Mediterranean), and North America [1,2]. These plants are characterized by quadrangular, sometimes winged stems and highly variable, undivided to 3-pinnatisect, generally opposite leaves. The fruit is a septicidal capsule, globose to subconical in shape, containing numerous small seeds [3].
Medicinal Properties and Uses
Various Scrophularia species have played a longstanding role in traditional medicine due to their demonstrated antioxidant, anti-tumor, anti-cancer, anti-protozoal, and anti-inflammatory activities [2,4,5]. In particular, S. orientalis has exhibited notable anticancer properties. A dichloromethane extract of this species significantly reduced the viability of neuroblastoma (NB) cells [6]. Similarly, filtered leaf extracts of S. striata have demonstrated anti-proliferative effects on a human astrocytoma cell line [7]. Moreover, extracts from S. floribunda and S. lucida have been shown to induce both necrosis and apoptosis in cancer cells, with S. lucida notably inhibiting tumor cell intravasation into lymph endothelial cells through suppression of NF-κB activity [8].
2. Echium Biebersteinii
General Overview
Echium, a genus in the Boraginaceae family, comprises approximately 60 species native to North Africa, Europe, and the Macaronesian islands [9,10,11]. This genus is closely related to Lobostemon (including Echiostachys), which is endemic to the southwestern Cape region. Although their pollen structures are highly similar—suggesting possible taxonomic convergence—distinct morphological differences separate the two genera. For example, Echium possesses bilobed styles and a unique annulus inside the corolla tube formed by a small collar or tiny hairy lobules, whereas Lobostemon exhibits undivided styles and hairs or scales at the base of the filaments [12].
Although E. biebersteinii has not been extensively studied, other species in the genus, notably E. amoenum, have received considerable pharmacological attention.
Medicinal Properties and Uses
Species of Echium are known for their sedative, anti-inflammatory, antioxidant, and anxiolytic effects. Consequently, they are widely used in traditional medicine to treat respiratory conditions, ulcers, mental health disorders, and for wound healing [10,13,14,15,16,17]. Among these, E. amoenum is the most extensively studied and has been traditionally used since the third century BC. Its applications include managing respiratory ailments, fevers, colds, and mood disturbances. Preparations such as boiling the plant or combining its leaves with wine were historically believed to enhance mood and relieve fever [10].
Although E. biebersteinii is considered a subspecies of E. italicum and remains underexplored, E. italicum has been traditionally employed in Turkey for wound healing, blister treatment, and relief of rheumatic pain through topical applications and infusions [10,18]. This species has also demonstrated strong anti-inflammatory, antioxidant, and anxiolytic activities [10,13,14,15,16,17]..
Recent studies have highlighted E. amoenum’s potential in cancer therapy. Its active compound, rosmarinic acid, has been shown to inhibit the growth and metastasis of gastric cancer cells through the inactivation of STAT3, AKT, and ERK1/2 signaling pathways [19].
3. Mentha Longifolia
General Overview
The genus Mentha (mint; Lamiaceae) includes between 18 and 30 species [20]. M. longifolia (L.) L. is an aromatic perennial herb widely distributed across Northern Pakistan, Europe, Nepal, India, western China, Germany, the United Kingdom, Egypt, Nigeria, and Turkey [21]. Members of the Mentha genus are herbaceous plants with spreading stolons both above and below ground [22]. Their branched stems are square in cross-section, and their aromatic leaves are oppositely arranged with either serrated or entire margins, typically oblong-elliptical to lanceolate in shape [23].
Medicinal Properties and Uses
Traditionally, M. longifolia has been used to treat gastrointestinal ailments, respiratory infections, and inflammatory diseases. Its essential oil, particularly menthol, exhibits antimicrobial properties against bacteria such as Staphylococcus aureus and Streptococcus mutans, as well as antifungal activity against Candida albicans [24]. Additionally, its flavonoid content has been suggested to possess anti-HIV potential [25].
Antioxidant activity has also been reported, with significant DPPH radical-scavenging and inhibition of linoleic acid oxidation observed in various assays [26]. Furthermore, crude extracts of M. longifolia have demonstrated antiproliferative effects in adrenocortical carcinoma SW13 cells, supporting its potential as an anti-cancer agent [27].
Taken together, the Scrophularia, Echium and Mentha species illustrate the remarkable pharmacological diversity found across different botanical families. Despite their phylogenetic divergence, these plants share a spectrum of bioactivities, including antioxidant, anti-inflammatory, cytotoxic, anti-proliferative, pro-apoptotic, and antimicrobial effects. These overlapping properties suggest that common molecular mechanisms may underpin the phenotypes observed in our study.
Despite the documented bioactivities of these herbs, their effects on DNA damage and reproductive function remain largely unexplored. To explore this hypothesis, we investigated the biological activity of the three herbal species using the Caenorhabditis elegans (C. elegans) model system. This model provides a powerful in vivo platform for studying the effects of bioactive compounds on development, reproduction, and genomic stability.
Plant-specific solvents were used to prepare the extracts—E. biebersteinii with butanol, M. longifolia with DMSO, and S. orientalis with water. All extracts were subsequently resuspended in a standardized DMSO–water mixture to ensure consistency in treatment conditions.
Comparative analysis revealed that exposure to each of the three herbal extracts significantly reduced worm survival compared to untreated controls. Treated worms exhibited larval arrest or lethality, suggesting that impaired survival may be linked to disruptions in mitotic cell division during larval development. Notably, all three extracts induced a high incidence of male progeny (HIM phenotype), implying disruption of sex chromosome segregation and potential interference with meiotic processes.
Further analysis revealed a reduced number of DAPI-stained bodies and abnormal meiotic progression in the germline of treated worms, providing additional evidence for impaired meiotic development. Consistently, treatment with any of the three extracts activated the DNA damage checkpoint response via the ATM/ATR and CHK-1 pathways. This response was accompanied by defective germline development, indicating that the extracts interfere with DNA damage repair mechanisms and ultimately lead to fertility defects.
To elucidate the molecular basis of these phenotypes, we performed LC-MS analysis of the herbal extracts. Several shared components—luteolin-7-O-rutinoside, thymol, carvyl acetate, and menthyl acetate—were identified, each has been previously associated with oxidative stress regulation, apoptosis induction, or genotoxic effects. These compounds are likely contributors to the observed disruptions in worm development and reproduction.
Interestingly, S. orientalis and E. biebersteinii shared 79% of their major compounds, indicating a high degree of chemical similarity. In contrast, M. longifolia shared only 42% of its compounds with the other two and possessed seven unique compounds (58%), reflecting a more distinct chemical profile. These differences may underlie the variable biological responses observed in the C. elegans assays and suggest plant-specific mechanisms of action.
This study reveals that extracts from Mentha longifolia, Scrophularia orientalis, and Echium biebersteinii induce reproductive defects in C. elegans by activating DNA damage checkpoints and apoptotic pathways. High-resolution imaging of germline architecture linked structural abnormalities—such as disorganized nuclei, impaired meiotic progression, and reduced bivalent formation—to molecular stress responses. All three extracts significantly decreased survival, caused larval arrest, and increased the high incidence of males (HIM) phenotype, indicating chromosomal mis-segregation.
We identified 21 major compounds, including four shared across the extracts. Among them, thymol and carvyl acetate were associated with pro-apoptotic activity, while thymol and luteolin-7-O-rutinoside exhibited antioxidant effects. These findings highlight both conserved and compound-specific mechanisms of herbal reproductive toxicity and support the use of C. elegans as a model for functional toxicological screening of traditional remedies. Also, this study underscores the need to analyze individual phytochemicals within herbal mixtures to understand their distinct biological effects.
Materials and Methods
Strains and Alleles
C. elegans strains were cultured at 20°C under standard laboratory conditions, following established protocols [28]. The N2 Bristol strain, used as the wild-type control, was obtained from the Caenorhabditis Genetics Center (CGC).
Herb Extraction
Herbal materials were sourced from Armenia and processed as previously reported [29,30]. In summary, plant samples were washed, air-dried, and coarsely ground before undergoing methanol extraction. The resulting methanolic extract was concentrated, reconstituted in 90% aqueous methanol, and partitioned with n-hexane. The hydroalcoholic residue was further fractionated sequentially using dichloromethane and n-butanol, yielding hexane-, butanol-, and water-soluble fractions. Solvent selection was tailored to each plant species: E. biebersteinii was extracted using butanol, M. longifolia with DMSO, and S. orientalis with water. All hexane-based extracts were redissolved in DMSO and standardized to 1 mg/mL before being diluted in M9 buffer to a working concentration of 0.03 µg/mL for most assays, unless otherwise noted.
Survival, Larval Arrest/Lethality, and High Incidence of Males (HIM) Assay
Synchronized L1 larvae were prepared by collecting gravid hermaphrodites from NGM plates, using the method described by Kim and Colaiacovo [31,32]. The larvae were then exposed to 180 µL of herbal extract solution in 96-well plates. Following brief agitation, the plates were incubated at 20°C for 24 hours, with phenotypic observations extending up to 48 hours. Worm survival was determined based on movement after 24 hours of treatment. Brood size was calculated by counting the total number of eggs laid per worm over a 4–5 day period following the L4 stage. Larval arrest or lethality was expressed as the percentage of hatched larvae that failed to reach adulthood. The High Incidence of Males (HIM) phenotype was assessed by calculating the percentage of males among the adult population. Differences among genotypes were analyzed using the two-tailed Mann–Whitney test, applying a 95% confidence interval (C.I.). Each experiment was independently replicated three times to ensure consistency. This procedure was modified from the protocol established by Kim and Colaiacovo [31].
LC–MS/MS Analysis
Liquid chromatography–tandem mass spectrometry (LC–MS/MS) was carried out according to established protocols [29,30]. Briefly, the analysis was conducted using a Shimadzu LC-30A system equipped with a C18 column, with all procedures performed by YanBo Times (Beijing, China). Compound identification was verified through comparison with a standardized reference database. All detected compounds were authenticated through this stringent methodology. The English names in the LC–MS output were translated from the original Chinese names supplied by YanBo Times.
Immunofluorescence Assay
Whole-mount gonads were stained for immunofluorescence following previously described methods [32,33,34]. The primary antibody used was rabbit anti-phospho-CHK-1 (Ser345) at a 1:250 dilution (Cell Signaling Technology), followed by Cy3-conjugated anti-rabbit secondary antibody at a 1:300 dilution (Jackson). Fluorescent images were captured using a Nikon Eclipse Ti2-E inverted microscope paired with a DS-Qi2 camera. Imaging was conducted at 0.2 μm Z-steps using a 60x objective lens with an additional 1.5x magnification. Image processing and deconvolution were performed using Nikon NIS Elements software. Figures display either full or partial nuclear projections.
pCHK-1 Foci Quantification
Assessment of Germline Apoptosis
Germline apoptosis was evaluated via acridine orange staining in synchronized animals, 20 hours after reaching the L4 stage, as previously described [35]. Between 20 and 30 gonads were scored per condition using a Nikon Ti2-E fluorescence microscope. Statistical comparisons were conducted using the two-tailed Mann–Whitney test, with significance set at a 95% confidence level.
qRT-PCR
Total RNA was isolated from young adult hermaphrodites and reverse-transcribed into cDNA using the ABscript II First Strand Synthesis Kit (ABclonal, RK20400). qRT-PCR was carried out using ABclonal 2X SYBR Green Fast Mix (RK21200) on the LineGene 4800 system (BIOER, FQD48A). Thermal cycling conditions included an initial denaturation at 95°C for 2 minutes, followed by 40 amplification cycles at 95°C for 15 seconds and 60°C for 20 seconds, with extension. A melting curve analysis (60°C–95°C) was performed to confirm product specificity. The tba-1 gene, which encodes tubulin, was used as an internal reference, based on previously published C. elegans microarray data. All PCR reactions were repeated at least twice to ensure reproducibility.
Results
All three plant extracts exhibited potent nematocidal activity after 48 hours of treatment at 20 °C, with survival rates ranging from 24% to 38%, compared to 89.4% in the DMSO-treated control group (Figure 2A). In addition to reduced survivability, extract-treated worms exhibited a larval arrest or lethality (93% vs. 38% for DMSO and M.l., p = 0.0002; 93% vs. 29% for S.o., p < 0.0001; 93% vs. 40% for E.b., p < 0.0001; two-tailed t-test), suggesting that decreased viability is likely linked to mitotic growth defects.
Also, all three extracts also significantly increased the incidence of the High Incidence of Males (HIM) phenotype, indicative of potential sex chromosome mis-segregation and aberrant meiotic development (0.29% vs. 3.12% for DMSO and M.l., p = 0.002; 3.47% for S.o., p = 0.0015; 9.82% for E.b., p = 0.0044; two-tailed t-test; [36]).
Since C. elegans feed on E. coli, we tested whether the observed nematocidal effects might result from indirect toxicity due to impaired bacterial growth. However, bacterial growth curves showed no significant changes following treatment with any of the three extracts at 0.03 μg/mL—the same concentration that induced phenotypes in C. elegans—indicating minimal impact on bacterial proliferation (Figure 2B). After 24 hours of incubation, the OD600 values were comparable across groups: 0.12 for DMSO + E. coli, 0.13 with M.l., 0.13 with S.o., and 0.14 with E.b.
In C. elegans, germline nuclei are organized in a well-defined spatial and temporal pattern during germline development. Actively dividing mitotic nuclei are located at the distal end within the premeiotic tip (PMT), and as cells move proximally, they enter meiotic prophase, beginning at the transition zone (TZ), where nuclei display a characteristic crescent-shaped morphology [36]. To assess effects on germline architecture, adult hermaphrodites were dissected, DAPI-stained, and analyzed. In controls, germline nuclei maintained orderly progression from the premeiotic tip (PMT) through the transition zone (TZ) to the pachytene region (Figure 3A, 3B). However, S.o. and E.b. treatments caused increased nuclear gaps, especially in the pachytene region, while S.o. additionally affected the PMT. In contrast, M.l. had no visible impact on nuclear organization.
Crescent-shaped nuclei, normally restricted to the transition zone (TZ) in controls, appeared ectopically in both the pre-meiotic tip (PMT) and pachytene regions of extract-treated worms (Figure 3C). While control animals showed proper localization of these nuclei to the TZ, all three herbal extracts induced their mislocalization into adjacent germline regions. This mislocalization increased significantly: M.l. (1.2 vs. 1.9, 1.58-fold, p = 0.0103), S.o. (1.2 vs. 2.6, 2.17-fold, p = 0.0649), and E.b. (1.2 vs. 2.0, 1.67-fold, p = 0.0088). These findings suggest premature entry into meiosis and disrupted developmental timing.
At the diakinesis stage, control worms showed the expected six DAPI-stained bivalents, whereas S.o.-treated animals showed five bivalents in 3.8% of cases, indicating potential homologous recombination or synapsis defects (Figure 3D; [37]). No abnormal bivalent numbers were detected in M.l. or E.b. groups.
Proper spatial organization of germline nuclei reflects normal developmental progression, and its disruption is often associated with reduced germline size. A significant decrease in germline length was observed only in worms treated with S. orientalis extract. The TZ and pachytene region lengths decreased from 45 μm to 29 μm (Figure 3E, p = 0.0006) and from 280 μm to 247 μm (p = 0.0175), respectively. No significant changes were observed in the PMT length (60 μm vs. 42 μm, p = 0.0519).
These developmental defects correlated with reduced fertility, as evidenced by a decrease in brood size over four days. The most notable reduction occurred on day 3. S.o.-treated worms showed a 3.08-fold decline in brood size (Figure 3F, 148 to 48, p = 0.0022), while M.l. and E.b. led to 1.59-fold (to 93, p = 0.0022) and 1.44-fold (to 103, p = 0.0050) reductions, respectively. These results suggest that impaired germline development ultimately leads to reduced fertility.
We hypothesized that impaired germline progression would activate the DNA damage checkpoint and initiate DNA repair mechanisms. To determine whether germline disruption was associated with activation of the DNA damage response, we assessed expression of DNA damage checkpoint genes. All three extracts significantly upregulated atm-1 and atl-1 mRNA, two key DNA damage checkpoint kinases: M.l. (Figure 4A, 1.52- and 1.51-fold), S.o. (2.26- and 2.24-fold), and E.b. (1.93- and 1.41-fold); p = 0.0007 for all (Mann–Whitney test).
Consistent with the upregulation of key DNA damage checkpoint genes, an increase in pCHK-1 foci was observed in the pachytene region following treatment with M.l. (1.6 vs. 4.9, p = 0.0049), S.o. (2.6, p = 0.0076), and E.b. (5.9, p = 0.0049) (Figure 4B). Additional pCHK-1 foci appeared in the PMT for M.l. (1.7 vs. 2.3, p = 0.0263) and E.b. (1.7 vs. 6.5, p < 0.0001), but not significantly for S.o. (1.7 vs. 2.0, p = 0.0981).
Activation of DNA damage checkpoint along with meiotic defects would lead to DNA damage mediated Cell death in pachytene stage of germline in C. elegans. In line with this idea, Apoptosis in the pachytene region increased significantly in S.o. (Figure 4C, 1 vs. 2.3, p = 0.0008) and E.b. (1 vs. 2.1, p = 0.0024) treated groups. M.l. induced a mild, non-significant increase (1 vs. 1.7, p = 0.0776). This apoptotic response was especially pronounced in worms treated with Stachys orientalis and Euphorbia biebersteinii, underscoring their stronger detrimental effects on germline integrity.
Among the three, S.o. induced the most pronounced phenotypes—altered nuclear organization, reduced bivalents, shortened germline regions, decreased brood size, and elevated expression of DNA damage markers—prompting further analysis of DNA repair. To further investigate this, we analyzed RAD-51 foci, which mark sites of double-strand break (DSB) repair [32,34]. RAD-51 foci were significantly increased in S.o.-treated worms at both the PMT (Figure 4D, 0.04 vs. 0.12, p = 0.023) and late pachytene stages (0.71 vs. 2.14, p = 0.0028), suggesting impaired double-strand break (DSB) repair (Figure 4D). Although RAD-51 foci levels were mildly increased in the transition zone, early pachytene, mid pachytene, and diplotene stages (0.12 vs. 0.11 in TZ, P = 0.7430; 1.36 vs. 1.64 in early pachytene, P = 0.1443; 4.14 vs. 4.54 in mid pachytene, P = 0.0752; 0.05 vs. 0.04 in diplotene, P = 0.7317), these differences were not statistically significant.
To explore the molecular basis of the distinct phenotypic effects observed in C. elegans, we conducted LC-MS analysis on each of the three herbal extracts, as detailed in our previous report [29,30,38]. This analysis identified 21 major compounds across the extracts (Figure 5), with four—luteolin-7-O-rutinoside, thymol, carvyl acetate, and menthyl acetate—common to all. Mentha longifolia contained the highest number of unique compounds, including caryophyllene, genistein, and ursolic acid, totaling seven unique constituents. Scrophularia orientalis featured one exclusive compound, resveratrol, while Echium biebersteinii uniquely contained vitexin-4'-rhamnoside. These findings highlight both common and unique chemical profiles that may explain the distinct biological activities of the extracts (Table 2).
Since all three herbs produced common phenotypes—upregulation of DNA damage checkpoint regulators and elevated germline apoptosis—we next investigated whether the four shared compounds could contribute to these effects. Specifically, we examined the expression levels of key DNA damage checkpoint genes following treatment with each compound.
Thymol and carvyl acetate significantly upregulated atm-1 and atl-1 (Figure 6A, thymol: 2.0- and 1.7-fold; carvyl acetate: 1.58- and 1.8-fold; P = 0.0005 for all), whereas luteolin-7-O-rutinoside and menthyl acetate had no significant effect (luteolin-7-O-rutinoside: P = 0.5396 for atm-1, P = 0.1870 for atl-1; menthyl acetate: P = 0.6029 for atm-1, P = 0.1459 for atl-1).
To determine whether these compounds also influence germline apoptosis, we quantified DNA damage-induced apoptosis. Consistent with their effects on checkpoint gene expression, thymol and carvyl acetate promoted germline apoptosis (Figure 6B, thymol: 1.33 to 2.47, p < 0.0001; carvyl acetate: 1.3 to 1.7, p = 0.0359). In contrast, luteolin-7-O-rutinoside induced only a marginal, non-significant change (1.05-fold, P = 0.7281), and menthyl acetate showed no effect (P = 0.9797). Thus, thymol and carvyl acetate may mediate the pro-apoptotic effects of the extracts. These findings suggest that among the common constituents, thymol and carvyl acetate may play an active role in DNA damage signaling and apoptosis, thereby contributing to the biological activities of the herb extracts.
Given antioxidant properties associated with these herbs (Figure 1, Figure 5, Supplemental Table 1 and 2), we next assessed their antioxidant capacity using the DPPH radical scavenging assay. All three herb extracts exhibited dose-dependent antioxidant activity, with M. longifolia showing the strongest effect, followed by S. orientalis and E. biebersteinii (Figure 6C). At 3 µg/ml, inhibition percentages were 39.15% for E.b. (P < 0.0001), 30.95% for S.o. (P < 0.0001), and 23.05% for M.l. (P < 0.0001).
To further dissect the contribution of individual compounds, we assessed the antioxidant activity of the four common constituents. Among them, luteolin-7-O-rutinoside and thymol displayed measurable radical-scavenging activity. Luteolin-7-O-rutinoside produced a modest but significant dose-dependent inhibition (Figure 6D, 2.15% at 20 µM, P < 0.0001; 3.17% at 30 µM, P < 0.0001). In contrast, thymol exhibited a much stronger antioxidant effect, reaching 18.05% inhibition at 2 mM and 18.08% at 3 mM (P < 0.0001 for both). Meanwhile, carvyl acetate and menthyl acetate did not show significant antioxidant activity at tested concentrations (carvyl acetate: max 0.37%, P > 0.87; menthyl acetate: max 0.55%, P > 0.13), indicating they are unlikely to contribute to the antioxidant effects of the extracts.
Discussion
Herbal Extracts Induce Germline-Specific DNA Damage Checkpoint Activation and Meiotic Defects in C. elegans
All three herbal extracts—Mentha longifolia (M.l.), Stachys orientalis (S.o.), and Euphorbia biebersteinii (E.b.)—exhibited strong nematocidal activity, reducing viability and inducing developmental arrest in C. elegans. These phenotypes were accompanied by a significant increase in the High Incidence of Males (HIM) phenotype, indicative of X chromosome nondisjunction and activation of DNA damage checkpoint and defective DNA repair (Figure 2, Figure 3 and Figure 4). These effects were not attributable to indirect E. coli-mediated toxicity, as bacterial growth remained unaffected by extract treatment.
Our multi-layered analysis—linking organism-level phenotypes to cellular, genetic, and molecular markers—demonstrates that these herbal extracts induce germline-specific defects through activation of conserved DNA damage checkpoint pathways. This systems-level approach offers a comprehensive view of the reproductive toxicity caused by botanical mixtures.
Herbal Extracts Lead to Defective Mitotic and Meiotic Progression, Impaired DNA Repair, and DNA Damage Checkpoint Activation, Resulting in Germline Apoptosis
DAPI staining of dissected gonads revealed that S.o. and E.b. disrupted the spatial organization of germline nuclei. The presence of crescent-shaped nuclei beyond the transition zone, as well as increased nuclear gaps, suggest premature meiotic entry and impaired control of the mitosis-to-meiosis switch. S.o. treatment additionally disrupted the premeiotic tip (PMT), pointing to broader developmental dysregulation. These morphological disruptions correlate with reduced germline length and decreased fertility.
All three extracts induced transcriptional upregulation of key DNA damage checkpoint regulators—atm-1 and atl-1—with accompanying increases in pCHK-1 foci and germline apoptosis. These effects were particularly pronounced in S.o. and E.b.-treated animals. This suggests that the extracts induce genotoxic stress or replication challenges sufficient to activate the DNA damage response, leading to checkpoint-mediated apoptotic removal of compromised germ cells.
Although M.l. showed milder phenotypes, it still significantly elevated checkpoint gene expression and pCHK-1 foci, indicating that even low-grade germline stress is sufficient to engage surveillance pathways.
Among the three extracts, S.o. produced the most severe phenotypes, including a reduction in diakinesis-stage bivalents, indicative of defective homolog pairing or recombination. Furthermore, RAD-51 foci were significantly elevated in the PMT and pachytene stages following S.o. treatment, suggesting impaired double-strand break (DSB) repair or persistent recombination intermediates. These disruptions likely compound DNA damage signaling, culminating in heightened apoptosis.
Phytochemical Composition Underlies the Biological Activities of Herbal Extracts: Four Common Compounds Identified—Thymol, Carvyl Acetate, Luteolin-7-O-Rutinoside, and Menthyl Acetate.
LC-MS profiling revealed both shared and species-specific compounds across the three extracts. Notably, four compounds—thymol, carvyl acetate, luteolin-7-O-rutinoside, and menthyl acetate—were common to all extractS. of these, thymol and carvyl acetate significantly upregulated atm-1 and atl-1 and increased germline apoptosis, effectively recapitulating the effects of the full extracts. In contrast, luteolin-7-O-rutinoside and menthyl acetate showed no such activity, underscoring the functional specificity of individual phytochemicals. This finding suggests that a subset of shared compounds may mediate the core genotoxic effects observed across all extracts, while species-specific compounds and the combination of compounds may modulate their severity.
Phytochemical Overlap Explains Parallel DNA Damage Responses Induced by S. orientalis and E. biebersteinii Extracts
We next asked whether the phytochemical similarities between extracts could explain their shared phenotypic profiles. Interestingly, S.o. and E.b. exhibited the most phenotypic similarity among the three extracts—manifesting nearly indistinguishable effects on germline disorganization, apoptosis, and checkpoint activation. This similarity is supported by their phytochemical profiles: 12 out of 13 major compounds in S.o. were also found in E.b., suggesting a shared chemical basis for their biological effects.
In addition to the shared compounds, S.o. and E.b. both contain misoprostol and aucubin, which have been linked to modulation of DNA damage and repair pathways. Misoprostol has demonstrated radioprotective effects in mammalian models by mitigating DNA damage-induced apoptosis [39], while aucubin has been implicated in topoisomerase-mediated DNA repair regulation and has shown therapeutic relevance in cancer settings [40]. These compounds may enhance or synergize with the shared DDR-active constituents to produce stronger germline toxicity.
Moreover, resveratrol, uniquely present in S.o., is a well-known polyphenol with multiple pharmacologic activities including promotion of IR-mediated apoptosis [41,42]. Resveratrol has been shown to sensitize tumor cells to radiation and enhance DNA damage-induced apoptosis, and may contribute to the severity of phenotypes seen in S.o.-treated animals.
Uncoupling Antioxidant Activity from Germline Toxicity in Herbal Extracts
While genotoxicity emerged as a major effect of the extracts, we also considered whether antioxidant properties might modulate or counterbalance these effects. All three extracts showed dose-dependent antioxidant activity in DPPH assays, with M.l. being the most potent. However, the genotoxic and apoptotic effects did not correlate with antioxidant capacity. For example, M.l. had the highest antioxidant activity but the mildest phenotypes, while S.o. and E.b. exhibited stronger toxicity despite moderate antioxidant profiles. Among shared compounds, thymol contributed to both antioxidant and pro-apoptotic activity, whereas carvyl acetate induced apoptosis without radical-scavenging effects.
These findings indicate that the biological effects of the extracts cannot be explained solely by oxidative stress modulation. Instead, distinct compounds within each extract exert functionally divergent effects—some activating protective antioxidant pathways, others engaging pro-apoptotic DNA damage signaling.
Our findings reveal that the germline phenotypes and fertility defects observed in C. elegans upon treatment with Mentha longifolia, Stachys orientalis, and Euphorbia biebersteinii extracts are the result of both shared and species-specific phytochemicals. Among the four compounds common to all three extracts, thymol and carvyl acetate specifically induced DNA damage checkpoint activation and pachytene-stage apoptosis, while thymol and luteolin-7-O-rutinoside contributed to antioxidant activity. The identification of carvyl acetate as a potent apoptosis inducer without antioxidant activity highlights its distinct and potentially toxic function. Meanwhile, species-specific constituents—such as ursolic acid and caryophyllene in M. longifolia, or resveratrol in S. orientalis—may contribute additional, non-overlapping biological effects.
Importantly, this study illustrates how the interaction between shared and unique compounds drives the complex and divergent biological outcomes of each herbal extract. By establishing a clear correlation between LC-MS-derived chemical profiles and in vivo physiological effects, we provide a mechanistic framework for understanding how multi-component herbal formulations act in biological systems. Moreover, our results experimentally demonstrate the potential of these herbs to induce reproductive toxicity and DNA damage responses, emphasizing the need for careful evaluation of herbal products, especially those consumed as food or supplements.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Methodology: Q.M. and H-M.K.; Validation: A.H. and Q.M.; Investigation: A.H., Q.M. and H-M.K.; Reference collection and verification: A.H.; Resources: R.P.B. and H-M.K.; Writing—original draft: H-M.K. and A.H.; Writing—review and editing: R.P.B. and H-M.K.; Proofreading: A.H., Q.M., R.P.B., and H-M.K.; Supervision: H-M.K.; Project administration: H-M.K.; Funding acquisition: H-M.K. All authors have reviewed and approved the final version of the manuscript.
Funding
This work was funded by the Kunshan Shuangchuang grant award (KSSC202202060).
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within the article and supplementary materials.
Acknowledgments
We thank members of the Kim laboratory for discussions and proofreading, especially Zifei Liu.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ren, D.; et al. Pharmacology, phytochemistry, and traditional uses of Scrophularia ningpoensis Hemsl. Journal of Ethnopharmacology 2021, 269, 113688. [Google Scholar] [CrossRef] [PubMed]
- de Santos Galíndez, J.; Díaz Lanza, A.M.a.; Fernández Matellano, L. Biologically Active Substances from the Genus Scrophularia. Pharmaceutical Biology 2002, 40, 45–59. [Google Scholar] [CrossRef]
- Scheunert, A.; Heubl, G. Against all odds: reconstructing the evolutionary history of Scrophularia (Scrophulariaceae) despite high levels of incongruence and reticulate evolution. Organisms Diversity & Evolution 2017, 17, 323–349. [Google Scholar]
- Al-Dalahmeh, Y.; et al. Scrophularia peyronii Post. from Jordan: Chemical Composition of Essential Oil and Phytochemical Profiling of Crude Extracts and Their In Vitro Antioxidant Activity. Life 2023, 13, 1404. [Google Scholar] [CrossRef]
- Pasdaran, A.; Hamedi, A. The genus Scrophularia: a source of iridoids and terpenoids with a diverse biological activity. Pharmaceutical Biology 2017, 55, 2211–2233. [Google Scholar] [CrossRef]
- Lange, I.; et al. Scrophularia orientalis extract induces calcium signaling and apoptosis in neuroblastoma cells. International Journal of Oncology 2016, 48, 1608–1616. [Google Scholar] [CrossRef] [PubMed]
- Ardeshiry lajimi, A.; et al. Study of Anti Cancer Property of Scrophularia striata Extract on the Human Astrocytoma Cell Line (1321). Iranian Journal of Pharmaceutical Research : IJPR 2010, 9, 403–410. [Google Scholar]
- Giessrigl, B.; et al. Effects of Scrophularia extracts on tumor cell proliferation, death and intravasation through lymphoendothelial cell barriers. International Journal of Oncology 2012, 40, 2063–2074. [Google Scholar]
- Baltisberger, M.; Widmer, A. Chromosome numbers of plant species from the Canary Islands. Botanica Helvetica 2006, 116, 9–30. [Google Scholar] [CrossRef]
- Jin, J.; et al. Antioxidant Properties and Reported Ethnomedicinal Use of the Genus Echium (Boraginaceae). Antioxidants 2020, 9, 722. [Google Scholar] [CrossRef]
- Kefi, S.; et al. Phytochemical investigation and biological activities of Echium arenarium (Guss) extracts. Microbial Pathogenesis 2018, 118, 202–210. [Google Scholar] [CrossRef]
- Retief, E.; Van Wyk, A.E. The genus Echium (Boraginaceae) in southern Africa. Bothalia 1998, 28, 167–177. [Google Scholar] [CrossRef]
- Hosseini, N.; Abolhassani, M. Immunomodulatory properties of borage (Echium amoenum) on BALB/c mice infected with Leishmania major. Journal of clinical immunology 2011, 31, 465–471. [Google Scholar] [CrossRef] [PubMed]
- Bekhradnia, S.; Ebrahimzadeh, M.A. Antioxidant activity of Echium amoenum. Rev Chim J 2016, 67, 223–226. [Google Scholar]
- Rabbani, M.; et al. Anxiolytic effects of Echium amoenum on the elevated plus-maze model of anxiety in mice. Fitoterapia 2004, 75, 457–464. [Google Scholar] [CrossRef] [PubMed]
- Potdar, V.H.; Kibile, S.J. Evaluation of antidepressant-like effect of Citrus maxima leaves in animal models of depression. Iranian journal of basic medical sciences 2011, 14, 478. [Google Scholar] [PubMed]
- Shafaghi, B.; et al. Anxiolytic effect of Echium amoenum L. in mice. in mice. 2002.
- Yeşilada, E.; et al. Traditional medicine in Turkey. V. Folk medicine in the inner Taurus Mountains. Journal of Ethnopharmacology 1995, 46, 133–152. [Google Scholar] [CrossRef]
- Ahmadi, M.; et al. Echium amoenum and Rosmarinic Acid Suppress the Growth and Metastasis of Gastric Cancer AGS Cells by Promoting Apoptosis and Inhibiting EMT. 2024(1422-0067 (Electronic)).
- Jedrzejczyk, I.; Rewers, M. Genome size and ISSR markers for Mentha L. (Lamiaceae) genetic diversity assessment and species identification. Industrial Crops and Products 2018, 120, 171–179. [Google Scholar] [CrossRef]
- Shahar, B.; Chongtham, N. Traditional uses and advances in recent research on wild aromatic plant Mentha longifolia and its pharmacological importance. Phytochemistry Reviews 2024, 23, 529–550. [Google Scholar] [CrossRef]
- Ahmad, R.S. , Herbs and Spices: New Processing Technologies. 2021: BoD–Books on Demand.
- Brahmi, F.; et al. Chemical composition and biological activities of Mentha species, in Aromatic and medicinal plants-Back to nature. 2017, IntechOpen.
- Al-Bayati, F.A. Isolation and identification of antimicrobial compound from Mentha longifolia L. leaves grown wild in Iraq. Annals of Clinical Microbiology and Antimicrobials 2009, 8, 20. [Google Scholar] [CrossRef] [PubMed]
- Amzazi, S.; et al. Human Immunodeficiency Virus Type 1 Inhibitory Activity of Mentha longifolia. Therapies 2003, 58, 531–534. [Google Scholar] [CrossRef] [PubMed]
- Gulluce, M.; et al. Antimicrobial and antioxidant properties of the essential oils and methanol extract from Mentha longifolia L. ssp. longifolia. Food Chemistry 2007, 103, 1449–1456. [Google Scholar] [CrossRef]
- Patti, F.; et al. Anticancer Effects of Wild Mountain Mentha longifolia Extract in Adrenocortical Tumor Cell Models. Frontiers in Pharmacology 2020, 10. [Google Scholar] [CrossRef]
- Brenner, S. The genetics of Caenorhabditis elegans. Genetics 1974, 77, 71–94. [Google Scholar] [CrossRef]
- Meng, Q.; Borris, R.P.; Kim, H.M. Torenia sp. Extracts Contain Multiple Potent Antitumor Compounds with Nematocidal Activity, Triggering an Activated DNA Damage Checkpoint and Defective Meiotic Progression. Pharmaceuticals (Basel) 2024, 17, 611. [Google Scholar] [CrossRef]
- Meng, Q.; et al. Exploring the Impact of Onobrychis cornuta and Veratrum lobelianum Extracts on C. elegans: Implications for MAPK Modulation, Germline Development, and Antitumor Properties. Nutrients 2023, 16, 1–22. [Google Scholar] [CrossRef]
- Kim, H.M.; Colaiacovo, M.P. DNA Damage Sensitivity Assays in Caenorhabditis elegans. Bio-Protocol 2015, 5, 1–12. [Google Scholar] [CrossRef]
- Kim, H.M.; Colaiacovo, M.P. ZTF-8 Interacts with the 9-1-1 Complex and Is Required for DNA Damage Response and Double-Strand Break Repair in the C. elegans Germline. PLoS Genet 2014, 10, e1004723. [Google Scholar] [CrossRef]
- Colaiacovo, M.P.; et al. Synaptonemal complex assembly in C. elegans is dispensable for loading strand-exchange proteins but critical for proper completion of recombination. Dev Cell 2003, 5, 463–474. [Google Scholar] [CrossRef]
- Kim, H.M.; Colaiacovo, M.P. New Insights into the Post-Translational Regulation of DNA Damage Response and Double-Strand Break Repair in Caenorhabditis elegans. Genetics 2015, 200, 495–504. [Google Scholar] [CrossRef] [PubMed]
- Kelly, K.O.; et al. Caenorhabditis elegans msh-5 is required for both normal and radiation-induced meiotic crossing over but not for completion of meiosis. Genetics 2000, 156, 617–630. [Google Scholar] [CrossRef]
- Lui, D.Y.; Colaiacovo, M.P. Meiotic development in Caenorhabditis elegans. Advances in experimental medicine and biology 2013, 757, 133–170. [Google Scholar] [PubMed]
- Girard, C.; et al. Interdependent and separable functions of Caenorhabditis elegans MRN-C complex members couple formation and repair of meiotic DSBs. Proc Natl Acad Sci U S A 2018, 115, E4443–E4452. [Google Scholar] [CrossRef] [PubMed]
- Meng, Q.; et al. Therapeutic Potential of Lappula patula Extracts on Germline Development and DNA Damage Responses in C. elegans. Pharmaceuticals (Basel) 2025, 18, 89. [Google Scholar] [CrossRef] [PubMed]
- van Buul, P.P.; et al. Differential radioprotective effects of misoprostol in DNA repair-proficient and -deficient or radiosensitive cell systems. Int J Radiat Biol 1997, 71, 259–264. [Google Scholar]
- Galvez, M.; Martin-Cordero, C.; Ayuso, M.J. Iridoids as DNA topoisomerase I poisons. J Enzyme Inhib Med Chem 2005, 20, 389–392. [Google Scholar] [CrossRef]
- Basso, E.; et al. Resveratrol affects DNA damage induced by ionizing radiation in human lymphocytes in vitro. Mutat Res Genet Toxicol Environ Mutagen 2016, 806, 40–46. [Google Scholar] [CrossRef]
- Kao, C.L.; et al. Resveratrol-induced apoptosis and increased radiosensitivity in CD133-positive cells derived from atypical teratoid/rhabdoid tumor. Int J Radiat Oncol Biol Phys 2009, 74, 219–228. [Google Scholar] [CrossRef]
- Zou, Y.; et al. Luteolin prevents THP-1 macrophage pyroptosis by suppressing ROS production via Nrf2 activation. Chemico-Biological Interactions 2021, 345, 109573. [Google Scholar] [CrossRef]
- Shao, J.; et al. Luteoloside Inhibits Proliferation and Promotes Intrinsic and Extrinsic Pathway-Mediated Apoptosis Involving MAPK and mTOR Signaling Pathways in Human Cervical Cancer Cells. International Journal of Molecular Sciences 2018, 19, 1664. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; et al. Luteoloside induces G0/G1 arrest and pro-death autophagy through the ROS-mediated AKT/mTOR/p70S6K signalling pathway in human non-small cell lung cancer cell lines. Biochemical and Biophysical Research Communications 2017, 494, 263–269. [Google Scholar] [CrossRef]
- Lin, J.; et al. Luteoloside Inhibits IL-1β-Induced Apoptosis and Catabolism in Nucleus Pulposus Cells and Ameliorates Intervertebral Disk Degeneration. Frontiers in Pharmacology 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Salehi, B.; et al. Thymol, thyme, and other plant sources: Health and potential uses. Phytotherapy research: PTR 2018, 32, 1688–1706. [Google Scholar] [CrossRef] [PubMed]
- Slamenová, D.; et al. DNA-protective effects of two components of essential plant oils carvacrol and thymol on mammalian cells cultured in vitro. Neoplasma 2007, 54, 108–112. [Google Scholar]
- Li, Y.; et al. Thymol inhibits bladder cancer cell proliferation via inducing cell cycle arrest and apoptosis. Biochemical and Biophysical Research Communications 2017, 491, 530–536. [Google Scholar] [CrossRef]
- Liang, D.; et al. Thymol Inhibits LPS-Stimulated Inflammatory Response via Down-Regulation of NF-κB and MAPK Signaling Pathways in Mouse Mammary Epithelial Cells. Inflammation 2014, 37, 214–222. [Google Scholar] [CrossRef]
- Sun, Z.; et al. Chemical Composition and Anti-Inflammatory, Cytotoxic and Antioxidant Activities of Essential Oil from Leaves of Mentha piperita Grown in China. PLOS ONE 2014, 9, e114767. [Google Scholar] [CrossRef]
- Ahmadi, S.M.; et al. Structure-Antioxidant Activity Relationships of Luteolin and Catechin. Journal of Food Science 2020, 85, 298–305. [Google Scholar] [CrossRef]
- Xiang, C.; et al. Nonhomologous end joining and homologous recombination involved in luteolin-induced DNA damage in DT40 cells. Toxicology in Vitro 2020, 65, 104825. [Google Scholar] [CrossRef]
- Chen, K.-C.; et al. Luteolin attenuates TGF-β1-induced epithelial–mesenchymal transition of lung cancer cells by interfering in the PI3K/Akt–NF-κB–Snail pathway. Life Sciences 2013, 93, 924–933. [Google Scholar] [CrossRef] [PubMed]
- Nabavi, S.F.; et al. Luteolin as an anti-inflammatory and neuroprotective agent: A brief review. Brain Research Bulletin 2015, 119 Pt A, 1–11. [Google Scholar] [CrossRef]
- Calleja, M.A.; et al. The antioxidant effect of β-caryophyllene protects rat liver from carbon tetrachloride-induced fibrosis by inhibiting hepatic stellate cell activation. British Journal of Nutrition 2013, 109, 394–401. [Google Scholar] [CrossRef] [PubMed]
- Chan, H.-W.; et al. Beta-Caryophyllene Augments Radiotherapy Efficacy in GBM by Modulating Cell Apoptosis and DNA Damage Repair via PPARγ and NF-κB Pathways. Phytotherapy research: PTR 2025, 39, 776–788. [Google Scholar] [CrossRef]
- Dahham, S.S.; et al. The Anticancer, Antioxidant and Antimicrobial Properties of the Sesquiterpene β-Caryophyllene from the Essential Oil of Aquilaria crassna. Molecules (Basel, Switzerland) 2015, 20, 11808–11829. [Google Scholar] [CrossRef] [PubMed]
- Bakır, B.; et al. Investigation of the anti-inflammatory and analgesic activities of-caryophyllene. International Journal of Essential Oil Therapeutics 2008, 2, 41–44. [Google Scholar]
- Wójciak, M.; et al. Antioxidant Potential of Diosmin and Diosmetin against Oxidative Stress in Endothelial Cells. Molecules 2022, 27, 8232. [Google Scholar] [CrossRef]
- Shen, Z.; et al. Diosmetin protects against retinal injury via reduction of DNA damage and oxidative stress. Toxicology Reports 2016, 3, 78–86. [Google Scholar] [CrossRef]
- Choi, J.; et al. Diosmetin inhibits tumor development and block tumor angiogenesis in skin cancer. Biomedicine & Pharmacotherapy 2019, 117, 109091. [Google Scholar]
- Lee, D.-h.; et al. Anti-inflammatory effects of natural flavonoid diosmetin in IL-4 and LPS-induced macrophage activation and atopic dermatitis model. International Immunopharmacology 2020, 89, 107046. [Google Scholar] [CrossRef]
- Wu, H.-J.; Chan, W.-H. Genistein protects methylglyoxal-induced oxidative DNA damage and cell injury in human mononuclear cells. Toxicology in vitro: an international journal published in association with BIBRA 2007, 21, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; et al. Genistein-induced DNA damage is repaired by nonhomologous end joining and homologous recombination in TK6 cells. Journal of Cellular Physiology 2019, 234, 2683–2692. [Google Scholar] [CrossRef] [PubMed]
- Tominaga, Y.; et al. Genistein inhibits Brca1 mutant tumor growth through activation of DNA damage checkpoints, cell cycle arrest, and mitotic catastrophe. Cell Death & Differentiation 2007, 14, 472–479. [Google Scholar]
- Hämäläinen, M.; et al. Anti-Inflammatory Effects of Flavonoids: Genistein, Kaempferol, Quercetin, and Daidzein Inhibit STAT-1 and NF-κB Activations, Whereas Flavone, Isorhamnetin, Naringenin, and Pelargonidin Inhibit only NF-κB Activation along with Their Inhibitory Effect on iNOS Expression and NO Production in Activated Macrophages. Mediators of Inflammation 2007, 2007, 45673. [Google Scholar]
- Jayachandran, M.; et al. Isoquercetin upregulates antioxidant genes, suppresses inflammatory cytokines and regulates AMPK pathway in streptozotocin-induced diabetic rats. Chemico-Biological Interactions 2019, 303, 62–69. [Google Scholar] [CrossRef]
- da Silva, D.d.C.; et al. Antitumor effect of isoquercetin on tissue vasohibin expression and colon cancer vasculature. Oncotarget 2022, 13, 307–318. [Google Scholar] [CrossRef]
- Ma, C.; et al. Isoquercetin ameliorates myocardial infarction through anti-inflammation and anti-apoptosis factor and regulating TLR4-NF-κB signal pathway. Molecular Medicine Reports 2018, 17, 6675–6680. [Google Scholar] [CrossRef]
- El-desoky, A.H.; et al. Anti-inflammatory and antioxidant activities of naringin isolated from Carissa carandas L.: In vitro and in vivo evidence. Phytomedicine 2018, 42, 126–134. [Google Scholar] [CrossRef]
- Manna, K.; et al. Naringin inhibits gamma radiation-induced oxidative DNA damage and inflammation, by modulating p53 and NF-κB signaling pathways in murine splenocytes. Free Radical Research 2015, 49, 422–439. [Google Scholar] [CrossRef]
- Zeng, L.; et al. Naringin inhibits growth and induces apoptosis by a mechanism dependent on reduced activation of NF-κB/COX-2-caspase-1 pathway in HeLa cervical cancer cells. International Journal of Oncology 2014, 45, 1929–1936. [Google Scholar] [CrossRef]
- do Nascimento, P.G.G.; et al. Antibacterial and antioxidant activities of ursolic acid and derivatives. Molecules (Basel, Switzerland) 2014, 19, 1317–1327. [Google Scholar] [CrossRef] [PubMed]
- Ramos, A.A.; Pereira-Wilson, C.; Collins, A.R. Protective effects of Ursolic acid and Luteolin against oxidative DNA damage include enhancement of DNA repair in Caco-2 cells. Mutation Research/Fundamental and Molecular Mechanisms of Mutagenesis 2010, 692, 6–11. [Google Scholar] [CrossRef] [PubMed]
- Kassi, E.; et al. Ursolic acid, a naturally occurring triterpenoid, demonstrates anticancer activity on human prostate cancer cells. Journal of Cancer Research and Clinical Oncology 2007, 133, 493–500. [Google Scholar] [CrossRef] [PubMed]
- Checker, R.; et al. Potent Anti-Inflammatory Activity of Ursolic Acid, a Triterpenoid Antioxidant, Is Mediated through Suppression of NF-κB, AP-1 and NF-AT. PLOS ONE 2012, 7, e31318. [Google Scholar] [CrossRef]
- Carrizzo, A.; et al. Antioxidant effects of resveratrol in cardiovascular, cerebral and metabolic diseases. Food and Chemical Toxicology 2013, 61, 215–226. [Google Scholar] [CrossRef]
- Sgambato, A.; et al. Resveratrol, a natural phenolic compound, inhibits cell proliferation and prevents oxidative DNA damage. Mutation Research/Genetic Toxicology and Environmental Mutagenesis 2001, 496, 171–180. [Google Scholar] [CrossRef]
- Hogg, S.J.; et al. Resveratrol, Acetyl-Resveratrol, and Polydatin Exhibit Antigrowth Activity against 3D Cell Aggregates of the SKOV-3 and OVCAR-8 Ovarian Cancer Cell Lines. Obstetrics and Gynecology International 2015, 2015, 279591. [Google Scholar] [CrossRef]
- Das, S.; Das, D.K. Anti-Inflammatory Responses of Resveratrol. Inflammation & Allergy - Drug Targets (Formerly Current Drug Targets - Inflammation & Allergy) 2007, 6, 168–173. [Google Scholar]
- Wang, B.-W.; et al. Aucubin Protects Chondrocytes Against IL-1β-Induced Apoptosis In Vitro And Inhibits Osteoarthritis In Mice Model. Drug Design, Development and Therapy 2019, 13, 3529–3538. [Google Scholar] [CrossRef]
- Shao, M.; et al. Aucubin Exerts Anticancer Activity in Breast Cancer and Regulates Intestinal Microbiota. Evidence-Based Complementary and Alternative Medicine 2022, 2022, 4534411. [Google Scholar] [CrossRef]
- Park, K.S.; Chang, I.-M. Anti-Inflammatory Activity of Aucubin by Inhibition of Tumor Necrosis Factor-α Production in RAW 264.7 Cells. Planta Medica 2004, 70, 778–779. [Google Scholar] [CrossRef] [PubMed]
- Breinholt, V.; et al. Dose-response effects of lycopene on selected drug-metabolizing and antioxidant enzymes in the rat. Cancer Letters 2000, 154, 201–210. [Google Scholar] [CrossRef]
- Jang, S.H.; et al. Lycopene inhibits Helicobacter pylori-induced ATM/ATR-dependent DNA damage response in gastric epithelial AGS cells. Free Radical Biology and Medicine 2012, 52, 607–615. [Google Scholar] [CrossRef]
- Jeong, Y.; Lim, J.W.; Kim, H. Lycopene Inhibits Reactive Oxygen Species-Mediated NF-κB Signaling and Induces Apoptosis in Pancreatic Cancer Cells. Nutrients 2019, 11, 762. [Google Scholar] [CrossRef] [PubMed]
- Hadad, N.; Levy, R. The synergistic anti-inflammatory effects of lycopene, lutein, β-carotene, and carnosic acid combinations via redox-based inhibition of NF-κB signaling. Free Radical Biology and Medicine 2012, 53, 1381–1391. [Google Scholar] [CrossRef] [PubMed]
- Beeharry N, L.J.; Hernandez, A.R.; Chambers, J.A.; Fucassi, F.; Cragg, P.J.; Green, M.H.; Green, I.C. Linoleic acid and antioxidants protect against DNA damage and apoptosis induced by palmitic acid. Mutat Res 2003, 530, 27–33. [Google Scholar] [CrossRef]
- Lauson, C.B.N.; et al. Linoleic acid potentiates CD8+ T cell metabolic fitness and antitumor immunity. Cell Metabolism 2023, 35, 633–650.e9. [Google Scholar] [CrossRef]
- Alarcon-Gil, J.; et al. Neuroprotective and Anti-Inflammatory Effects of Linoleic Acid in Models of Parkinson’s Disease: The Implication of Lipid Droplets and Lipophagy. Cells 2022, 11, 2297. [Google Scholar] [CrossRef]
- Bilgic, S.; Ozgocmen, M. The protective effect of misoprostol against doxorubicin induced liver injury. Biotechnic & Histochemistry 2019, 94, 583–591. [Google Scholar]
- Li, B.; et al. Homoplantaginin alleviates intervertebral disc degeneration by blocking the NF-κB/MAPK pathways via binding to TAK1. Biochemical Pharmacology 2024, 226, 116389. [Google Scholar] [CrossRef]
- Ying, X.-x.; et al. HPLC determination of malondialdehyde in ECV304 cell culture medium for measuring the antioxidant effect of vitexin-4″-O-glucoside. Archives of Pharmacal Research 2008, 31, 878–885. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; et al. Effects of vitexin-2″-O-rhamnoside and vitexin-4″-O-glucoside on growth and oxidative stress-induced cell apoptosis of human adipose-derived stem cells. Journal of Pharmacy and Pharmacology 2014, 66, 988–997. [Google Scholar] [CrossRef] [PubMed]
- Salehi, B.; et al. Plants of Genus Mentha: From Farm to Food Factory. Plants 2018, 7, 70. [Google Scholar] [CrossRef] [PubMed]
- Yousefian, S., F. Esmaeili, and T. Lohrasebi, A Comprehensive Review of the Key Characteristics of the Genus Mentha, Natural Compounds and Biotechnological Approaches for the Production of Secondary Metabolites. Iranian Journal of Biotechnology 2023, 21, e3605. [Google Scholar]
- Kajimoto, T.; et al. Iridoids from Scrophularia ningpoensis. Phytochemistry 1989, 28, 2701–2704. [Google Scholar] [CrossRef]
- Miyazawa, M.; et al. Suppression of SOS-inducing activity of chemical mutagens by cinnamic acid derivatives from Scrophulia ningpoensis in the Salmonella typhimurium TA1535/pSK1002 umu test. Journal of agricultural and food chemistry 1998, 46, 904–910. [Google Scholar] [CrossRef]
- Qian, J.; Hunkler, D.; Rimpler, H. Iridoid-related aglycone and its glycosides from Scrophularia ningpoensis. Phytochemistry 1992, 31, 905–911. [Google Scholar] [CrossRef]
- Fujita, T.; et al. Traditional medicine in Turkey VII. Folk medicine in middle and west Black Sea regions. Economic botany, 1995: p. 406-422.
- Amabeoku, G.J.; et al. Antipyretic and antinociceptive properties of mentha longifoliaHuds. (Lamiaceae) leaf aqueous extract in rats and mice. Methods and Findings in Experimental and Clinical Pharmacology 2009, 31, 645. [Google Scholar] [CrossRef]
- Karimian, P.; Gholamreza, K.; Amirghofran, Z. Anti-inflammatory effect of Mentha longifolia in lipopolysaccharide-stimulated macrophages: Reduction of nitric oxide production through inhibition of inducible nitric oxide synthase. Journal of Immunotoxicology 2013, 10, 393–400. [Google Scholar] [CrossRef]
- Afkar, S.; Somaghian, S.A. Determining of chemical composition, anti-pathogenic and anticancer activity of Mentha longifolia essential oil collected from Iran. Natural Product Research 2024, 0, 1–9. [Google Scholar] [CrossRef]
- Bashir, I.; Dilshad, E. A comparative study of Mentha longifolia var. asiatica and Zygophyllum arabicum ZnO nanoparticles against breast cancer targeting Rab22A gene. PLOS ONE 2024, 19, e0308982. [Google Scholar] [CrossRef] [PubMed]
- Beheshtian, N.; et al. Mentha longifolia L. Inhibits Colorectal Cancer Cell Proliferation and Induces Apoptosis via Caspase Regulation. International Journal of Translational Medicine 2023, 3, 416–425. [Google Scholar] [CrossRef]
- Elansary, H.O.; et al. Polyphenol Profile and Antimicrobial and Cytotoxic Activities of Natural Mentha × piperita and Mentha longifolia Populations in Northern Saudi Arabia. Processes 2020, 8, 479. [Google Scholar] [CrossRef]
- Esmaeili, F.; et al. Appraisals on the anticancer properties of Mentha species using bioassays and docking studies. Industrial Crops and Products 2023, 203, 117128. [Google Scholar] [CrossRef]
- Kelidari, H.R.; et al. Anticancer Effect of Solid-Lipid Nanoparticles Containing Mentha longifolia and Mentha pulegium Essential Oils: In Vitro Study on Human Melanoma and Breast Cancer Cell Lines. Biointerface Research in Applied Chemistry 2021, 12, 2128–2137. [Google Scholar]
- Li, S.; et al. Green synthesis of gold nanoparticles using aqueous extract of Mentha Longifolia leaf and investigation of its anti-human breast carcinoma properties in the in vitro condition. Arabian Journal of Chemistry 2021, 14, 102931. [Google Scholar] [CrossRef]
- Yassin, M.T.; Mostafa, A.A.; Al-Askar, A.A. Anticandidal and anti-carcinogenic activities of Mentha longifolia (Wild Mint) extracts in vitro. Journal of King Saud University - Science 2020, 32, 2046–2052. [Google Scholar] [CrossRef]
- Arvanagh, F.M.; et al. Anti-inflammatory and collagenation effects of zinc oxide-based nanocomposites biosynthesised with Mentha longifolia leaf extract. Journal of Wound Care 2023, 32, 44–54. [Google Scholar] [CrossRef]
- Asemani, Y.; et al. Modulation of in vitro proliferation and cytokine secretion of human lymphocytes by Mentha longifolia extracts. Avicenna Journal of Phytomedicine 2019, 9, 34–43. [Google Scholar]
- Bai, X.; et al. Chemical constituents and biological activities of essential oil from Mentha longifolia: effects of different extraction methods. International Journal of Food Properties 2020, 23, 1951–1960. [Google Scholar] [CrossRef]
- Dadkhah, A.; et al. Assessing the effect of Mentha longifolia essential oils on COX-2 expression in animal model of sepsis induced by caecal ligation and puncture. Pharmaceutical Biology 2018, 56, 495–504. [Google Scholar] [CrossRef] [PubMed]
- Haikal, A.; et al. Anti-asthmatic and antioxidant activity of flavonoids isolated from Mentha longifolia subspecies typhoides (Briq. ) Harley. and Mentha longifolia subspecies schimperi (Briq.) Briq. on ovalbumin-induced allergic asthma in mice: In-vivo and in-silico study. Journal of Ethnopharmacology 2025, 339, 119133. [Google Scholar] [PubMed]
- Ibrahim, A. anti-acetylcholinesterase, anti-inflammatory and antioxidant activities of Mentha longifolia for treating Alzheimer disease. Der Pharmacia Lettre 2016, 8, 34–39. [Google Scholar]
- Lal, M.; et al. Exploring the bioactive potential of Mentha longifolia from Northeast India: an inclusive study on phytochemical composition and biological activities. Journal of Essential Oil Bearing Plants 2024, 27, 1102–1120. [Google Scholar] [CrossRef]
- Pham, D.V.; et al. Anti-inflammatory activities of some Mentha essential oils in lipopolysaccharide-activated macrophages. Tạp chí Nghiên cứu Dược và Thông tin Thuốc, 2025.
- Raeisi, H.; et al. Pleiotropic effects of Mentha longifolia L. extract on the regulation of genes involved in inflammation and apoptosis induced by Clostridioides difficile ribotype 001. Frontiers in Microbiology 2023, 14. [Google Scholar] [CrossRef]
- Tourabi, M.; et al. Efficacy of various extracting solvents on phytochemical composition, and biological properties of Mentha longifolia L. leaf extracts. Scientific Reports 2023, 13, 18028. [Google Scholar] [CrossRef]
- Janifer, R.X.; et al. Determination of Total Phenols, Free Radical Scavenging and Antibacterial Activities of Mentha longifolia Linn. Hudson from the Cold Desert, Ladakh, India. Pharmacognosy Journal 2010, 2, 470–475. [Google Scholar] [CrossRef]
- Stanisavljević, D.M.; et al. Antioxidant activity, the content of total phenols and flavonoids in the ethanol extracts of Mentha longifolia (L.) Hudson dried by the use of different techniques. Chemical Industry and Chemical Engineering Quarterly 2012, 18, 411–420. [Google Scholar] [CrossRef]
- Wang, L.; et al. Green synthesis of Ag/Fe3O4 nanoparticles using Mentha longifolia flower extract: evaluation of its antioxidant and anti-lung cancer effects. Heliyon 2022, 8. [Google Scholar] [CrossRef]
- Z, G., K. Asres, and A. Mazumder, Comparison of the Essential Oil Composition, Antibacterial and Antioxidant Activities of Four Mentha Species Growing in Ethiopia. Ethiop. Pharm. J. 2007, 25, 91–102.
- Al-janabi, A.A.; Sahib, A.A.; Ali, F.J. Studying The Effect of Mentha Longifolia Plant Extract In Inhibition Growth of Some Bacteria and Inhibiting the Emergence Fourth Stage Larvae of Mosquitoes Aedes Aegypti. Indian Journal of Forensic Medicine & Toxicology 2020. [Google Scholar]
- Al-Mijalli, S.H.; et al. Phytochemical Variability, In Vitro and In Vivo Biological Investigations, and In Silico Antibacterial Mechanisms of Mentha piperita Essential Oils Collected from Two Different Regions in Morocco. Foods 2022, 11, 3466. [Google Scholar] [CrossRef]
- Faisal, S.; et al. Exploring the antibacterial, antidiabetic, and anticancer potential of Mentha arvensis extract through in-silico and in-vitro analysis. BMC Complementary Medicine and Therapies 2023, 23, 267. [Google Scholar] [CrossRef]
- Ghazyzadeh, M.; et al. Evaluation of the Antibacterial Activity of Mentha Longifolia Essential Oil against Enterococcus faecalis and its Chemical Composition. Journal of Dentistry 2025. [Google Scholar]
- Jahani, E.; Babaeekhou, L.; Ghane, M. Chemical composition and antibacterial properties of Zataria multiflora Bioss and Mentha longifolia essential oils in combination with nisin and acid acetic. Journal of Food Processing and Preservation 2021, 45, e15742. [Google Scholar] [CrossRef]
- Mancuso, M. The antibacterial activity of Mentha, in Herbs and spices. 2020, IntechOpen.
- Mohammadi-Aloucheh, R.; et al. Green synthesis of ZnO and ZnO/CuO nanocomposites in Mentha longifolia leaf extract: characterization and their application as anti-bacterial agents. Journal of Materials Science: Materials in Electronics 2018, 29, 13596–13605. [Google Scholar] [CrossRef]
- Shnawa, B.H.; et al. Scolicidal activity of biosynthesized zinc oxide nanoparticles by Mentha longifolia L. leaves against Echinococcus granulosus protoscolices. Emergent Materials 2022, 5, 683–693. [Google Scholar] [CrossRef]
- Shen, X.; et al. Effects of Scrophularia ningpoensis Hemsl. on Inhibition of Proliferation, Apoptosis Induction and NF-?B Signaling of Immortalized and Cancer Cell Lines. Pharmaceuticals 2012, 5, 189–208. [Google Scholar] [CrossRef]
- Goudarzvand, M.; et al. Immunomodulatory and anti-inflammatory effects of Scrophularia megalantha ethanol extract on an experimental model of multiple sclerosis. Research Journal of Pharmacognosy 2019, 6, 43–50. [Google Scholar]
- Li, Y.M.; et al. Fast repairing of oxidized OH radical adducts of dAMP and dGMP by phenylpropanoid glycosides from Scrophularia ningpoensis Hemsl. Acta pharmacologica Sinica 2000, 21, 1125–1128. [Google Scholar]
- Bahmani, M.; et al. A comparative study on the effect of ethanol extract of wild Scrophularia deserti and streptomycin on Brucellla melitensis. Journal of Herbmed Pharmacology 2013, 2, 17–20. [Google Scholar]
- Fernández, M.A., M. D. García, and M.T. Sáenz, Antibacterial activity of the phenolic acids fractions of Scrophularia frutescens and Scrophularia sambucifolia. Journal of Ethnopharmacology 1996, 53, 11–14. [Google Scholar] [CrossRef]
- Ayobi, H.; et al. Antibacterial Effects of Scrophularia striata Extract on Pseudomonas aeruginosa. Journal of Medicinal Plants 2014, 13, 73–80. [Google Scholar]
- Lewenhofer, V.; et al. Chemical Composition of Scrophularia lucida and the Effects on Tumor Invasiveness in Vitro. Frontiers in Pharmacology 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Musa, A.; et al. Prominent antidiabetic and anticancer investigation of Scrophularia deserti extract: Integration of experimental and computational approaches. Journal of Molecular Structure 2024, 1315, 138769. [Google Scholar] [CrossRef]
- Namvaran, A.; et al. Effects of Scrophularia oxysepala Methanolic Extract on Early Stages of Dimethylhydrazine-Induced Colon Carcinoma in Rats: Apoptosis Pathway Approach. Advanced Pharmaceutical Bulletin 2022, 12, 835–841. [Google Scholar] [CrossRef]
- Ahmed, B.; et al. Scropolioside-D2 and Harpagoside-B: Two New Iridoid Glycosides from Scrophularia deserti and Their Antidiabetic and Antiinflammatory Activity. Biological and Pharmaceutical Bulletin 2003, 26, 462–467. [Google Scholar] [CrossRef]
- Azadmehr, A.; et al. Antioxidant and Neuroprotective Effects of Scrophularia striata Extract Against Oxidative Stress-Induced Neurotoxicity. Cellular and Molecular Neurobiology 2013, 33, 1135–1141. [Google Scholar] [CrossRef]
- Byun, S.-H.; Yang, C.-H.; Kim, S.-C. Inhibitory effect of Scrophulariae Radix extract on TNF-alpha,IL-1beta, IL-6 and Nitric Oxide production in lipopolysaccharide - activated Raw 264.7 cells. The Korea Journal of Herbology 2005, 20, 7–16. [Google Scholar]
- Dı́az, A.M.a.; et al. Phenylpropanoid glycosides from Scrophularia scorodonia: In vitro anti-inflammatory activity. Life Sciences 2004, 74, 2515–2526. [Google Scholar] [CrossRef]
- Ebrahimi, A.; et al. Investigation of anti-inflammatory and analgesic effects of hydroalcoholic extract of Scrophularia striata seeds in male mice. Iranian Journal of Physiology and Pharmacology 2019, 3, 40–34. [Google Scholar]
- Fernández, M.A.; Sáenz, M.T.; García, M.D. Anti-inflammatory Activity in Rats and Mice of Phenolic Acids Isolated from Scrophularia frutescens. Journal of Pharmacy and Pharmacology 1998, 50, 1183–1186. [Google Scholar] [CrossRef] [PubMed]
- Giner, R.-M.a.; et al. Anti-inflammatory glycoterpenoids from Scrophularia auriculata. European Journal of Pharmacology 2000, 389, 243–252. [Google Scholar] [CrossRef]
- Hadi Pour Sabet, S.; Bahramikia, S.; Baghaifar, Z. Evaluation of antioxidant and anti-inflammatory properties of hydroalcoholic and methanolic extracts of Scrophularia striata: Inhibition of albumin protein denaturation and stabilization of erythrocyte membrane. Applied Biology 2023, 35, 150–162. [Google Scholar]
- Huo, Y.-F.; et al. Two new compounds from the roots of Scrophularia ningpoensis and their anti-inflammatory activities. Journal of Asian Natural Products Research 2019, 21, 1083–1089. [Google Scholar] [CrossRef]
- Mao, G.; et al. Scrodentoids H and I, a Pair of Natural Epimerides from Scrophularia dentata, Inhibit Inflammation through JNK-STAT3 Axis in THP-1 Cells. Evidence-Based Complementary and Alternative Medicine 2020, 2020, 1842347. [Google Scholar] [CrossRef]
- Pham, T.N.A.; et al. Anti-inflammatory Effects of Scrophularia buergeriana Extract Mixture Fermented with Lactic Acid Bacteria. Biotechnology and Bioprocess Engineering 2022, 27, 370–378. [Google Scholar] [CrossRef]
- Shin, N.-R.; et al. Scrophularia buergeriana attenuates allergic inflammation by reducing NF-κB activation. Phytomedicine 2020, 67, 153159. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.a.; et al. Polysaccharides of Scrophularia ningpoensis Hemsl.: Extraction, Antioxidant, and Anti-Inflammatory Evaluation. Evidence-Based Complementary and Alternative Medicine 2020, 2020, 8899762. [Google Scholar] [CrossRef]
- Zengin, G.; et al. Scrophularia lucida L. as a valuable source of bioactive compounds for pharmaceutical applications: In vitro antioxidant, anti-inflammatory, enzyme inhibitory properties, in silico studies, and HPLC profiles. Journal of Pharmaceutical and Biomedical Analysis 2019, 162, 225–233. [Google Scholar] [CrossRef]
- Dokhani, N.; et al. Determination and evaluating the antioxidant properties of ziziphus nummularia (burm. F.) wight & arn., crataegus pontica K. Koch and scrophularia striata boiss. Egyptian Journal of Veterinary Sciences 2022, 53, 423–429. [Google Scholar]
- Jeong, E.J.; et al. Cognitive-enhancing and antioxidant activities of iridoid glycosides from Scrophularia buergeriana in scopolamine-treated mice. European Journal of Pharmacology 2008, 588, 78–84. [Google Scholar] [CrossRef] [PubMed]
- Jeong, E.J.; et al. KD-501, a standardized extract of Scrophularia buergeriana has both cognitive-enhancing and antioxidant activities in mice given scopolamine. Journal of Ethnopharmacology 2009, 121, 98–105. [Google Scholar] [CrossRef]
- Lee, H.J.; et al. Neuroprotective effects of Scrophularia buergeriana extract against glutamate-induced toxicity in SH-SY5Y cells. International Journal of Molecular Medicine 2019, 43, 2144–2152. [Google Scholar] [CrossRef]
- Mahboubi, M.; Kazempour, N.; Boland Nazar, A.R. Total Phenolic, Total Flavonoids, Antioxidant and Antimicrobial Activities of Scrophularia Striata Boiss Extracts. Jundishapur Journal of Natural Pharmaceutical Products 2013, 8, 15–19. [Google Scholar] [CrossRef]
- Roya Alizadeh, S.; et al. Scrophularia striata extract mediated synthesis of gold nanoparticles; their antibacterial, antileishmanial, antioxidant, and photocatalytic activities. Inorganic Chemistry Communications 2023, 156, 111138. [Google Scholar] [CrossRef]
- Shiri, H.; et al. Evaluation of Antioxidant Potential and Free Radical Scavenging Activity of Methanol Extract from Scrophularia striata. Acta Biochimica Iranica 2023. [Google Scholar] [CrossRef]
- Zargoosh, Z.; et al. Effects of ecological factors on the antioxidant potential and total phenol content of Scrophularia striata Boiss. Scientific Reports 2019, 9, 16021. [Google Scholar] [CrossRef]
- Akşit, Z. Chemical composition and antimicrobial activity of Scrophularia catariifolia Boiss. & Heldr essential oil. Journal of Essential Oil Bearing Plants 2025, 28, 37–43. [Google Scholar]
- Ebrahimzadeh, M.A.; et al. In vitro cytotoxicity against human cancer cell lines (MCF-7 and AGS), antileishmanial and antibacterial activities of green synthesized silver nanoparticles using Scrophularia striata extract. Surfaces and Interfaces 2021, 23, 100963. [Google Scholar] [CrossRef]
- Jafari, A.A.; et al. Chemical composition and biological activities of Scrophularia striata extracts. Minerva Biotecnologica 2014, 26, 183–189. [Google Scholar]
- Li, J.; et al. Study of chemical composition and antimicrobial activity of leaves and roots of Scrophularia ningpoensis. Natural Product Research 2009, 23, 775–780. [Google Scholar] [CrossRef]
- Lin, T.; et al. The in vitro and in vivo antibacterial activities of uniflorous honey from a medicinal plant, Scrophularia ningpoensis Hemsl. , and characterization of its chemical profile with UPLC-MS/MS. Journal of Ethnopharmacology 2022, 296, 115499. [Google Scholar]
- Mameneh, R.; et al. Characterization and antibacterial activity of plant mediated silver nanoparticles biosynthesized using Scrophularia striata flower extract. Russian Journal of Applied Chemistry 2015, 88, 538–546. [Google Scholar] [CrossRef]
- Pasdaran, A.; et al. Chemical Composition, and Antibacterial (Against Staphylococcus aureus) and Free-Radical-Scavenging Activities of the Essential Oil of Scrophularia amplexicaulis Benth. Rec. Nat. Prod. 2012. [Google Scholar]
- Renda, G.; et al. Chemical Composition and Antimicrobial Activity of the Essential Oils of Five Scrophularia L. Species from Turkey. Records of Natural Products 2017, 11, 521–531. [Google Scholar] [CrossRef]
- Sharafati-chaleshtori, R.; Rafieian-kopaei, M. Screening of antibacterial effect of the Scrophularia Striata against E. coli in vitro. Journal of Herbmed Pharmacology 2014, 3, 31–34. [Google Scholar]
- Stavri, M.; Mathew, K.T.; Gibbons, S. Antimicrobial constituents of Scrophularia deserti. Phytochemistry 2006, 67, 1530–1533. [Google Scholar] [CrossRef]
- Tavarideh, F.; Pourahmad, F.; Nemati, M. Diversity and antibacterial activity of endophytic bacteria associated with medicinal plant, Scrophularia striata. Veterinary Research Forum 2022, 13, 409–415. [Google Scholar]
- Yook, K.-D. Antimicrobial activity and cytotoxicity test of Scrophularia ningpoensis hemsl extracts against Klebsiella pneumoniae. Journal of the Korea Society of Computer and Information 2016, 21, 135–139. [Google Scholar] [CrossRef]
- Zangeneh, M.M.; et al. Assessment of In Vitro Antibacterial Properties of the Hydroalcoholic Extract of Scrophularia striata Against Staphylococcus aureus (ATCC No. 25923). International Journal of Pharmacognosy and Phytochemical Research 2017, 9. [Google Scholar] [CrossRef]
- Naseri, N.; Kalantar, K.; Amirghofran, Z. Anti-inflammatory activity of Echium amoenum extract on macrophages mediated by inhibition of inflammatory mediators and cytokines expression. Research in Pharmaceutical Sciences 2018, 13, 73. [Google Scholar] [PubMed]
- Asghari, B.; et al. Therapeutic target enzymes inhibitory potential, antioxidant activity, and rosmarinic acid content of Echium amoenum. South African Journal of Botany 2019, 120, 191–197. [Google Scholar] [CrossRef]
- Farahani, M. Farahani, M., Q. Branch, and I. Azad, Antiviral effect assay of aqueous extract of Echium amoenum-L against HSV-1. Zahedan J Res Med Sci 2013, 15, 46–48. [Google Scholar]
- Abolhassani, M. Clinical research Antiviral activity of borage (Echium amoenum). Archives of Medical Science 2010, 6, 366–369. [Google Scholar] [CrossRef]
- Alamholo, M. Antiradical and antibacterial activity of Echium altissimum extracts on human infective bacteria and chemical composition analysis. Microbiology, Metabolites and Biotechnology 2020, 3, 19–27. [Google Scholar]
- El-Tantawy, H.M., A.R. Hassan, and H.E. Taha, Antioxidant potential and LC/MS metabolic profile of anticancer fractions from Echium angustifolium Mill. aerial parts. Journal of Applied Pharmaceutical Science 2021, 11, 200–208.
- El-Tantawy, H.M.; Hassan, A.R.; Taha, H.E. Anticancer mechanism of the non-polar extract from Echium angustifolium Mill. aerial parts in relation to its chemical content. Egyptian Journal of Chemistry 2022, 65, 17–26. [Google Scholar] [CrossRef]
- Fazeli, M. The effect of Echium amoenum hydro-methanolic extract on MCF-7 and MDA-MB468 breast cancer cell lines and increasing the cytotoxicity of doxorubicin. Iranian Journal of Physiology and Pharmacology 2023, 7, 84–94. [Google Scholar]
- Wang, W.; et al. Comparative analysis of the main medicinal substances and applications of Echium vulgare L. and Echium plantagineum L.: A review. Journal of Ethnopharmacology 2022, 285, 114894. [Google Scholar] [CrossRef]
- Abed, A.; et al. Effect of Echium amoenum Fisch. et Mey a Traditional Iranian Herbal Remedy in an Experimental Model of Acute Pancreatitis. International Scholarly Research Notices 2012, 2012, 141548. [Google Scholar] [CrossRef] [PubMed]
- Benlakhdar, H.; et al. Chemical Composition and Anti-inflammatory Activity of the Essential Oil of Echium humile (Boraginaceae) in vivo from South-West of Algeria. Jordan Journal of Biological Sciences 2021, 14. [Google Scholar]
- Eruygur, N.; Yilmaz, G.; Üstün, O. Analgesic and antioxidant activity of some Echium species wild growing in Turkey. FABAD Journal of Pharmaceutical Sciences 2012, 37, 151. [Google Scholar]
- Heidari, M.R.; Azad, E.M.; Mehrabani, M. Evaluation of the analgesic effect of Echium amoenum Fisch & C.A. Mey. extract in mice: Possible mechanism involved. Journal of Ethnopharmacology 2006, 103, 345–349. [Google Scholar] [PubMed]
- Kitessa, S.M., P.D. Nichols, and M. Abeywardena, Purple Viper's Bugloss (Echium plantagineum) Seed Oil in Human Health, in Nuts and Seeds in Health and Disease Prevention, V.R. Preedy, R.R. Watson, and V.B. Patel, Editors. 2011, Academic Press: San Diego. p. 951-958.
- Mir, M. Echium oil: A valuable source of n-3 and n-6 fatty acids. Oléagineux, Corps gras, Lipides 2008, 15, 252–256. [Google Scholar] [CrossRef]
- Moita, E.; et al. Integrated Analysis of COX-2 and iNOS Derived Inflammatory Mediators in LPS-Stimulated RAW Macrophages Pre-Exposed to Echium plantagineum L. Bee Pollen Extract. PLOS ONE 2013, 8, e59131. [Google Scholar] [CrossRef]
- Moreira, R.; et al. Echium plantagineum L. honey: Search of pyrrolizidine alkaloids and polyphenols, anti-inflammatory potential and cytotoxicity. Food Chemistry 2020, 328, 127169. [Google Scholar] [CrossRef]
- Abbasi Larki, R.; et al. Protective Effects of Echium amoenum on Oxidative Stress and Gene Expression Induced by Permethrin in Wistar Rats. Hepatitis Monthly 2020, 20. [Google Scholar] [CrossRef]
- Abbaszadeh, S.; Rajabian, T.; Taghizadeh, M. Antioxidant Activity, Phenolic and Flavonoid Contents of Echium Species from Different Geographical Locations of Iran. Journal of Medicinal plants and By-Products 2013, 2, 23–31. [Google Scholar]
- Abdol, N.; Setorki, M. Evaluation of Antidepressant, Antianxiolytic, and Antioxidant Effects of Echium amoenum L. Extract on Social Isolation Stress of Male Mice. Iranian Red Crescent Medical Journal 2020, 22. [Google Scholar] [CrossRef]
- Adel Pilerood, S.; Prakash, J. Evaluation of nutritional composition and antioxidant activity of Borage (Echium amoenum) and Valerian (Valerian officinalis). Journal of Food Science and Technology 2014, 51, 845–854. [Google Scholar] [CrossRef] [PubMed]
- Al-Rimawi, F.; et al. Free radicals and enzymes inhibitory potentials of the traditional medicinal plant Echium angustifolium. European Journal of Integrative Medicine 2020, 38, 101196. [Google Scholar] [CrossRef]
- Alsanie, W.; et al. Viper's Bugloss (Echium Vulgare L) Extract as A Natural Antioxidant and Its Effect on Hyperlipidemia. 2018. Volume 8: p. 81-89.
- Aouadi, K.; et al. HPLC/MS Phytochemical Profiling with Antioxidant Activities of Echium humile Desf. Extracts: ADMET Prediction and Computational Study Targeting Human Peroxiredoxin 5 Receptor. Agronomy 2021, 11, 2165. [Google Scholar] [CrossRef]
- Benbrinis, S. In vitro and In vivo anti-inflammatory and hepatoprotector effects of Algerian Echium plantagineum and Brassica rapa extracts. 2024.
- Mehran, M., S. Masoum, and M. Memarzadeh, Improvement of thermal stability and antioxidant activity of anthocyanins of Echium amoenum petal using maltodextrin/modified starch combination as wall material. International Journal of Biological Macromolecules 2020, 148, 768–776. [CrossRef]
- Safaeian, L.; et al. Cytoprotective and antioxidant effects of Echium amoenum anthocyanin-rich extract in human endothelial cells (HUVECs). Avicenna Journal of Phytomedicine 2015, 5, 157–166. [Google Scholar] [PubMed]
- Uysal, İ.; et al. Antioxidant and Oxidant status of medicinal plant Echium italicum collected from different regions. Turkish Journal of Agriculture - Food Science and Technology 2021, 9, 1902–1904. [Google Scholar] [CrossRef]
- Abolhassani, M. Antibacterial effect of borage (Echium amoenum) on Staphylococcus aureus. Brazilian Journal of Infectious Diseases 2004, 8, 382–385. [Google Scholar] [CrossRef]
- Atessahin, D.A.; et al. Evaluation of cytotoxic, antimicrobial, and antioxidant activities of Echium italicum L. in MCF-7 and HepG2 cell lines. International Journal of Plant Based Pharmaceuticals 2025, 5, 25–32. [Google Scholar] [CrossRef]
- Kaïss, A.; et al. Phytochemical Profiling, Antimicrobial and α-Glucosidase Inhibitory Potential of Phenolic-Enriched Extracts of the Aerial Parts from Echium humile Desf.: In Vitro Combined with In Silico Approach. Plants 2022, 11, 1131. [Google Scholar]
- Kavehei, M.; Sefidroo, S.H. Investigating the Antimicrobial Activity of Different Extracts of Echium on Selected Gram-Positive and Gram-Negative Bacteria. Tabari Biomedical Student Research Journal 2023, 5, 18–24. [Google Scholar] [CrossRef]
- Morteza-Semnani, K.; Saeedi, M.; Akbarzadeh, M. Chemical Composition and Antimicrobial Activity of Essential Oil of Echium italicum L. Journal of Essential Oil Bearing Plants, 2009. Journal of Essential Oil Bearing Plants 2009. [Google Scholar]
- Nabipour*, F.; et al. Evaluation of the Antifungal effects of various Extracts of the Aerial part and Root of Echium italicum on Candida albicans compared with two common antibiotics. Yafteh 2019, 21. [Google Scholar]
- Sabour, M.; et al. Evaluation of the antibacterial effect of Echium amoenum Fisch. et Mey. against multidrug resistant Acinetobacter baumannii strains isolated from burn wound infection. Novelty in Biomedicine 2015, 3, 38–42. [Google Scholar]
- Shariatifar, N.; Fathabad, A.E.; Madihi, S. Antibacterial activity of aqueous and ethanolic extracts of Echium amoenum on food-borne pathogens. Journal of Food Safety and Hygiene 2016, 2, 63–63. [Google Scholar]
- Tabata, M.; Tsukada, M.; Fukui, H. Antimicrobial Activity of Quinone Derivatives from Echium lycopsis Callus Cultures. Planta Medica 1982, 44, 234–236. [Google Scholar] [CrossRef]
- Beeharry, N.; et al. Linoleic acid and antioxidants protect against DNA damage and apoptosis induced by palmitic acid. Mutation Research/Fundamental and Molecular Mechanisms of Mutagenesis 2003, 530, 27–33. [Google Scholar] [CrossRef]
Figure 1.
Venn diagram summarizing the reported biological activities of the genera Scrophularia, Mentha, and Echium based on published literature. This diagram emphasizes the shared biological properties among the three genera from which the herb extracts were derived. Notably, all three have been consistently reported to exhibit antioxidant, pro-apoptotic, anti-inflammatory, cytotoxic, anti-proliferative, and antimicrobial activities. Non-overlapping regions represent additional, genus-specific effects reported in the literature. Detailed information can be found in Table 1, Supplementary Table 1, and Supplementary Table 2.
Figure 1.
Venn diagram summarizing the reported biological activities of the genera Scrophularia, Mentha, and Echium based on published literature. This diagram emphasizes the shared biological properties among the three genera from which the herb extracts were derived. Notably, all three have been consistently reported to exhibit antioxidant, pro-apoptotic, anti-inflammatory, cytotoxic, anti-proliferative, and antimicrobial activities. Non-overlapping regions represent additional, genus-specific effects reported in the literature. Detailed information can be found in Table 1, Supplementary Table 1, and Supplementary Table 2.
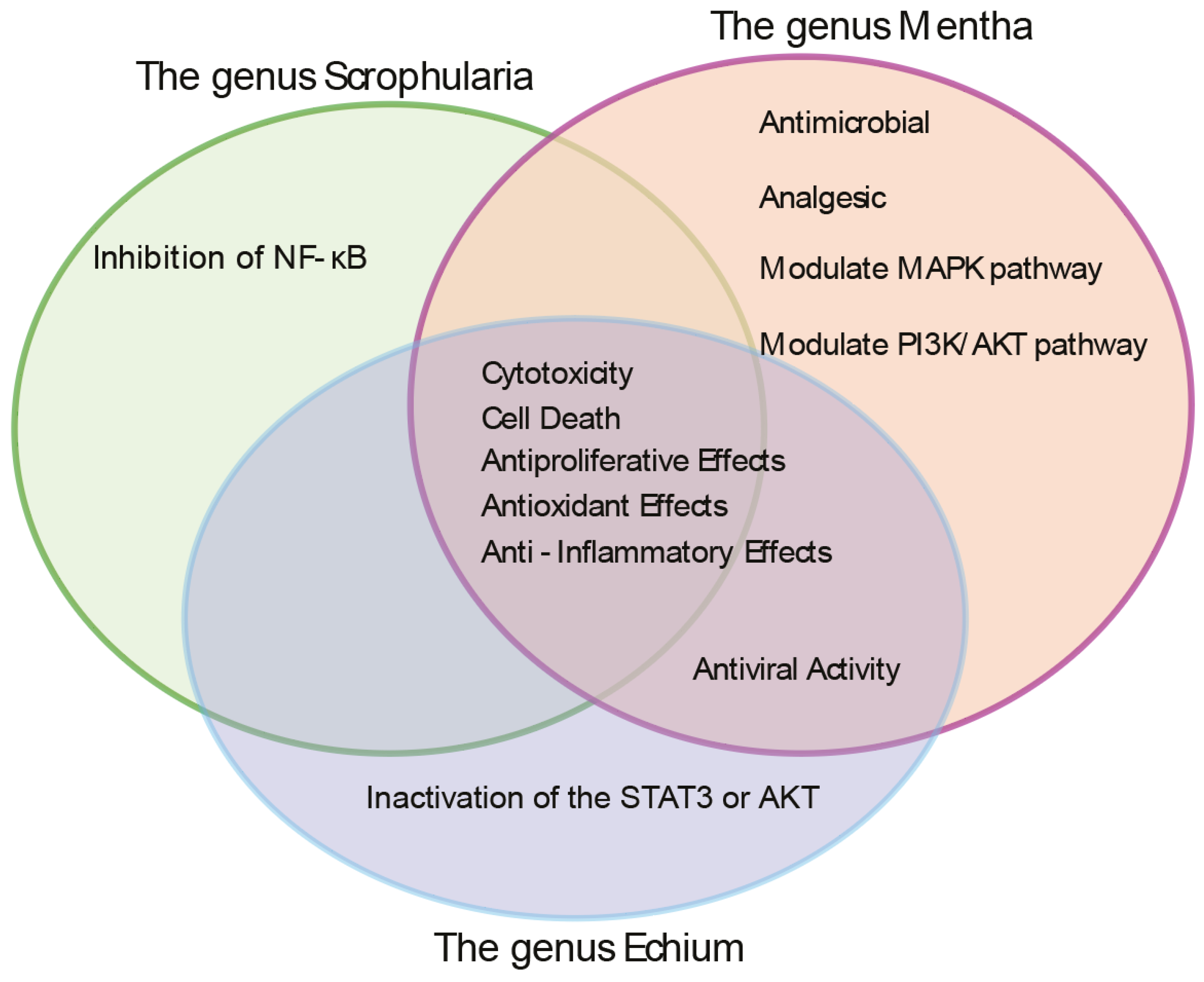
Figure 2.
Extracts obtained from Mentha longifolia (M.l), Scrophularia orientalis (S.o), and Echium biebersteinii (E.b) exhibit marked nematocidal, larval arrest/lethality and HIM phenotype of C. elegans, without exerting discernible impact on bacterial growth. A) M.l, S.o, and E.b extracts significantly reduced the survival and larval development and high incidence of male (HIM) of C. elegans. The effect of herb extracts were evaluated by treating worms with different extracts of M.l, S.o, and E.b (indicated by brown, orange, and gray colors) and monitoring their survival, adult formation, and male (HIM) phenotype over a 48 h period. Statistical significance was assessed using a two-tailed t-test, with ** p < 0.01; *** p < 0.001; and **** p < 0.0001, comparing the control (+DMSO) with the treated samples. Nocodazole is a positive control. (B) Assessment of bacterial growth in the presence of herbal extracts. E. coli OP50 was incubated with 0.03 μg/mL of M.l, S.o, and E.b extracts—the same concentration used in C. elegans assays—for 24 hours. No significant inhibition of bacterial growth was observed at Absorbance (600nm), indicating that the extracts' nematocidal effects are unlikely to result from compromised bacterial food source (P=0.100 for all three herb at 24 hour of incubation).
Figure 2.
Extracts obtained from Mentha longifolia (M.l), Scrophularia orientalis (S.o), and Echium biebersteinii (E.b) exhibit marked nematocidal, larval arrest/lethality and HIM phenotype of C. elegans, without exerting discernible impact on bacterial growth. A) M.l, S.o, and E.b extracts significantly reduced the survival and larval development and high incidence of male (HIM) of C. elegans. The effect of herb extracts were evaluated by treating worms with different extracts of M.l, S.o, and E.b (indicated by brown, orange, and gray colors) and monitoring their survival, adult formation, and male (HIM) phenotype over a 48 h period. Statistical significance was assessed using a two-tailed t-test, with ** p < 0.01; *** p < 0.001; and **** p < 0.0001, comparing the control (+DMSO) with the treated samples. Nocodazole is a positive control. (B) Assessment of bacterial growth in the presence of herbal extracts. E. coli OP50 was incubated with 0.03 μg/mL of M.l, S.o, and E.b extracts—the same concentration used in C. elegans assays—for 24 hours. No significant inhibition of bacterial growth was observed at Absorbance (600nm), indicating that the extracts' nematocidal effects are unlikely to result from compromised bacterial food source (P=0.100 for all three herb at 24 hour of incubation).
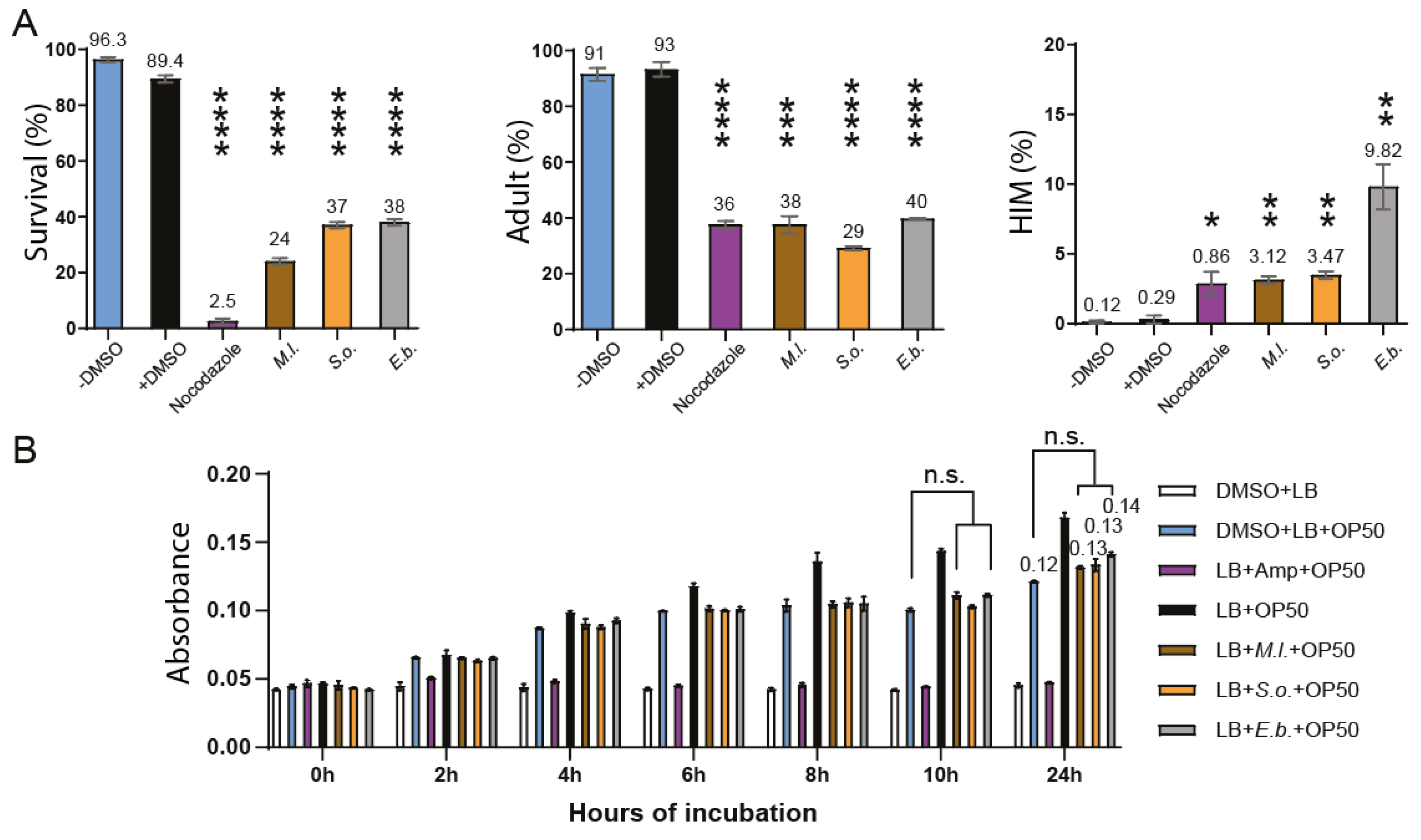
Figure 3.
Effects of herbal extracts on nuclear organization, germline development, and fertility defective outcomes in C. elegans. S.o and E.b herb extracts induced increased spacing between nuclei within the pachytene region. In contrast, the M.l extract did not produce any discernible changes in nuclear spacing or organization. (A) DAPI-stained nuclei during germline development of 24 hours post L4 hermaphrodite with or without treatment of three herb extracts. Yellow Arrows indicate crescent shape nuclei positioned at pachytene. White arrow indicates chromatin bridge. (B) Quantification of the increased nuclear spacing in the PMT and pachytene stages shown in the panel. (C) Quantification of crescent -shaped nuclei in per gonad arm is indicated. Asterisks indicate statistically significant differences compared to the control group. (D) Quantification of DAPI-stained bivalents in the germline. The percentage of bivalent at -1 position oocyte is indicated. Numbers in the bracket indicate the number of bivalents. (E) Germline length was measured in three regions: the premeiotic tip (PMT), transition zone (TZ), and pachytene. S.o. extract shortens specific TZ and pachytene stage. (F) Brood size of herb-exposed C. elegans hermaphrodites. The number of offspring produced by individual hermaphrodite worms was monitored daily over a four-day reproductive period following treatment with herbal extracts. Data are presented as mean ± SEM. Statistical significance was assessed using a two-tailed T-test. Asterisks indicate statistically significant differences compared to the control group.
Figure 3.
Effects of herbal extracts on nuclear organization, germline development, and fertility defective outcomes in C. elegans. S.o and E.b herb extracts induced increased spacing between nuclei within the pachytene region. In contrast, the M.l extract did not produce any discernible changes in nuclear spacing or organization. (A) DAPI-stained nuclei during germline development of 24 hours post L4 hermaphrodite with or without treatment of three herb extracts. Yellow Arrows indicate crescent shape nuclei positioned at pachytene. White arrow indicates chromatin bridge. (B) Quantification of the increased nuclear spacing in the PMT and pachytene stages shown in the panel. (C) Quantification of crescent -shaped nuclei in per gonad arm is indicated. Asterisks indicate statistically significant differences compared to the control group. (D) Quantification of DAPI-stained bivalents in the germline. The percentage of bivalent at -1 position oocyte is indicated. Numbers in the bracket indicate the number of bivalents. (E) Germline length was measured in three regions: the premeiotic tip (PMT), transition zone (TZ), and pachytene. S.o. extract shortens specific TZ and pachytene stage. (F) Brood size of herb-exposed C. elegans hermaphrodites. The number of offspring produced by individual hermaphrodite worms was monitored daily over a four-day reproductive period following treatment with herbal extracts. Data are presented as mean ± SEM. Statistical significance was assessed using a two-tailed T-test. Asterisks indicate statistically significant differences compared to the control group.
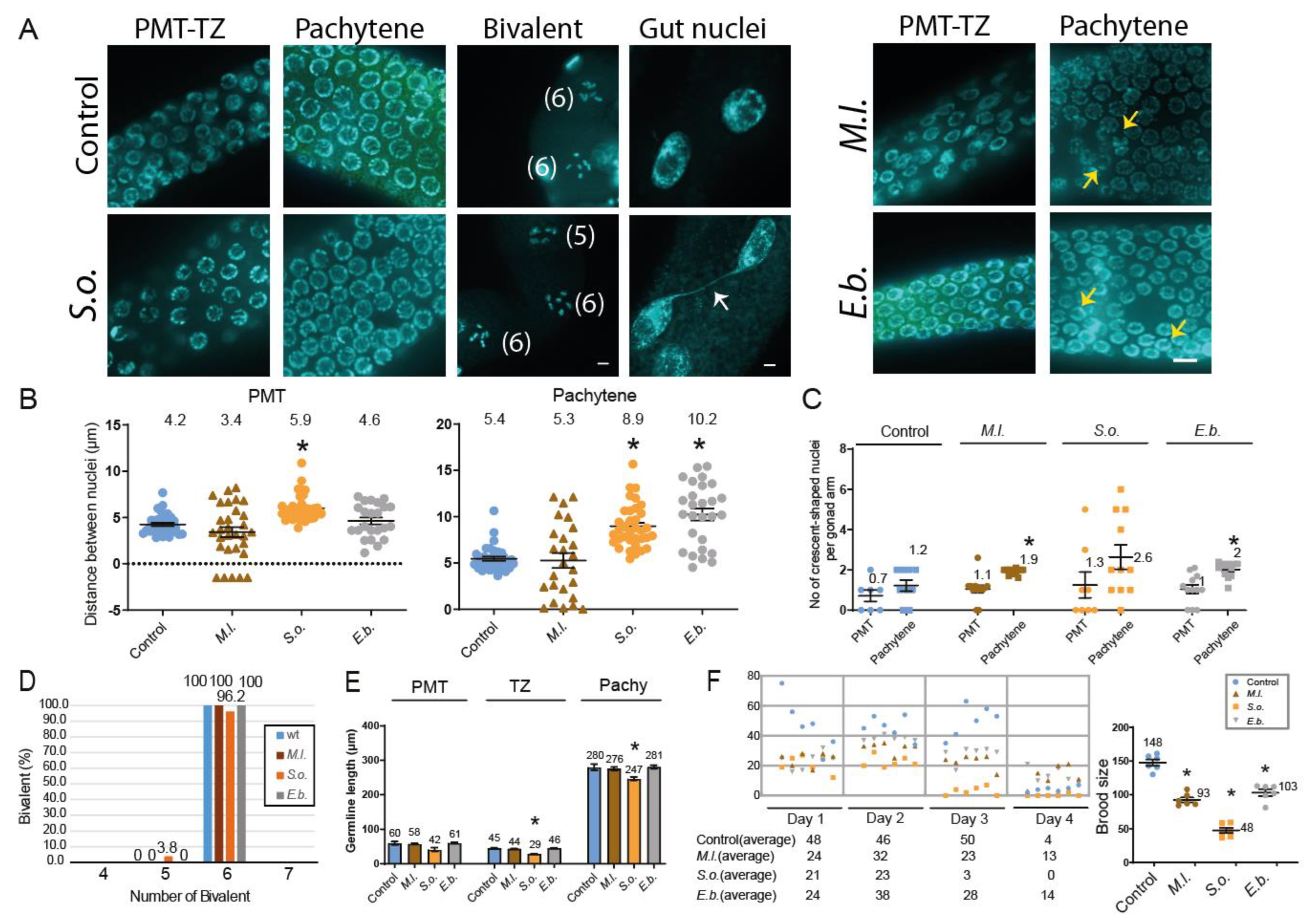
Figure 4.
The three herb extract exposure activates the DNA damage checkpoint pathway and apoptosis. S.o. extract leads to defective DSB repair in the germline. (A) Quantitative PCR analysis of DNA damage checkpoint gene expression in whole worms treated with herbal extracts. Transcript levels of atm-1 and atl-1 were normalized to tba-1 (tubulin) and compared to untreated controls. (B) Quantification of pCHK-1 foci, a downstream marker of ATM/ATR checkpoint activation, in the premeiotic tip (PMT) and pachytene region. All three herb treatments significantly increased pCHK-1 foci in the pachytene stage. Arrows indicate pCHK-1 foci adjacent to chromatin. (C) Quantification of germline apoptosis using acridine orange staining. Apoptotic nuclei were significantly elevated in the pachytene region following S.o. and E.b. treatments, while M.l. treatment caused a mild, non-significant increase compared to the control. (D) RAD-51 foci quantification to assess double-strand break (DSB) repair. S.o. treatment led to significantly increased RAD-51 foci in both the PMT and late pachytene, indicating impaired DSB repair during both mitotic and meiotic stages. All statistical analyses were performed using two-tailed Mann–Whitney tests. Data are presented as mean ± SEM from biological replicates. Asterisks indicate statistically significant differences compared to the control group.
Figure 4.
The three herb extract exposure activates the DNA damage checkpoint pathway and apoptosis. S.o. extract leads to defective DSB repair in the germline. (A) Quantitative PCR analysis of DNA damage checkpoint gene expression in whole worms treated with herbal extracts. Transcript levels of atm-1 and atl-1 were normalized to tba-1 (tubulin) and compared to untreated controls. (B) Quantification of pCHK-1 foci, a downstream marker of ATM/ATR checkpoint activation, in the premeiotic tip (PMT) and pachytene region. All three herb treatments significantly increased pCHK-1 foci in the pachytene stage. Arrows indicate pCHK-1 foci adjacent to chromatin. (C) Quantification of germline apoptosis using acridine orange staining. Apoptotic nuclei were significantly elevated in the pachytene region following S.o. and E.b. treatments, while M.l. treatment caused a mild, non-significant increase compared to the control. (D) RAD-51 foci quantification to assess double-strand break (DSB) repair. S.o. treatment led to significantly increased RAD-51 foci in both the PMT and late pachytene, indicating impaired DSB repair during both mitotic and meiotic stages. All statistical analyses were performed using two-tailed Mann–Whitney tests. Data are presented as mean ± SEM from biological replicates. Asterisks indicate statistically significant differences compared to the control group.
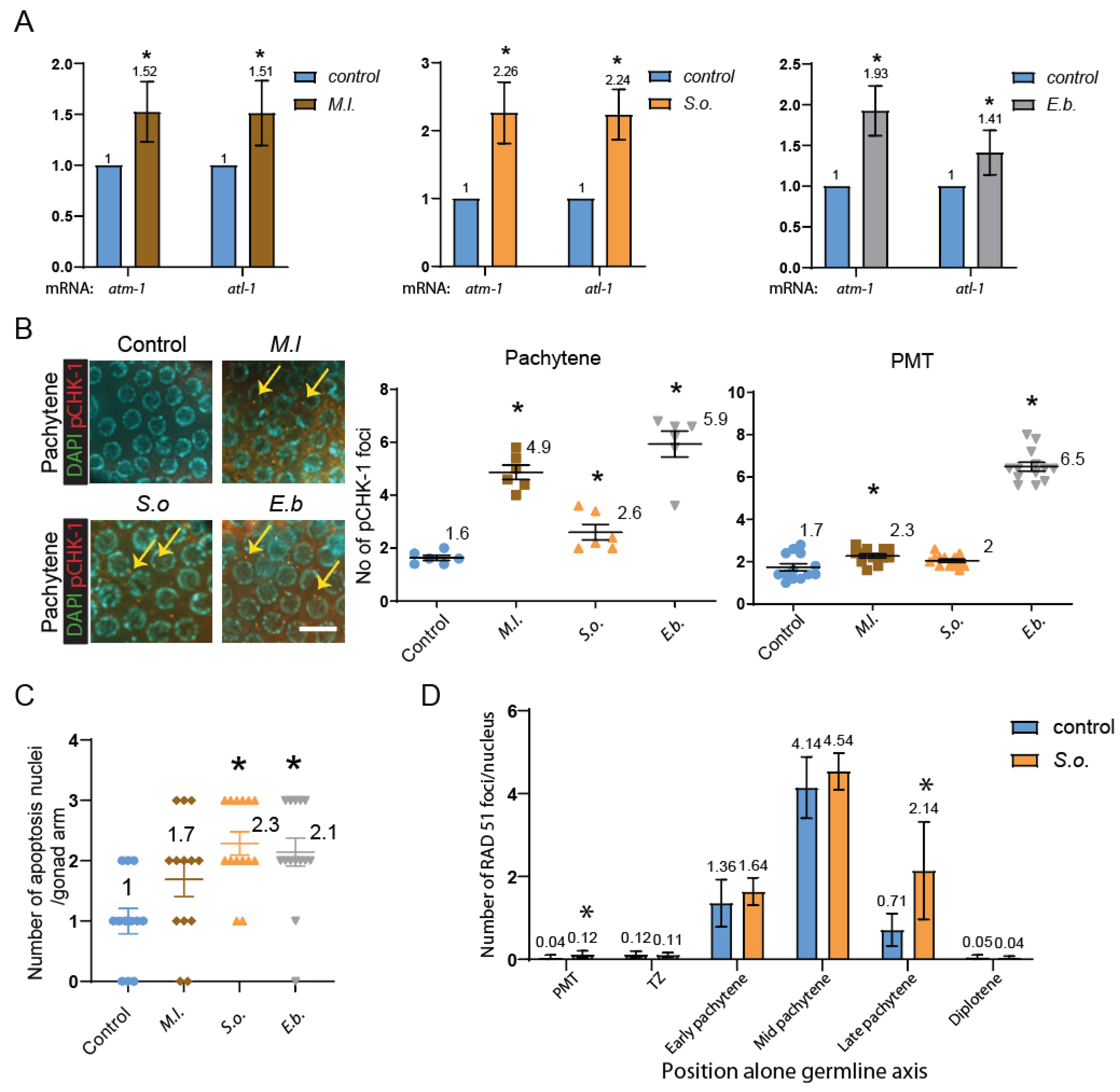
Figure 5.
Comparative LC-MS analysis of major compounds in three herbal extracts. Venn diagram and heat map summarizing the 21 major compounds identified across Mentha longifolia, Scrophularia orientalis, and Echium biebersteinii. Four compounds—luteolin-7-O-rutinoside, thymol, carvyl acetate, and menthyl acetate—were common to all three extracts. M. longifolia contained seven unique major compounds, including caryophyllene, genistein, and ursolic acid. S. orientalis had one unique compound, resveratrol, while E. biebersteinii uniquely possessed vitexin-4'-rhamnoside. Compound identification was performed based on methods previously described (see Materials and Methods; [29,30,38]).
Figure 5.
Comparative LC-MS analysis of major compounds in three herbal extracts. Venn diagram and heat map summarizing the 21 major compounds identified across Mentha longifolia, Scrophularia orientalis, and Echium biebersteinii. Four compounds—luteolin-7-O-rutinoside, thymol, carvyl acetate, and menthyl acetate—were common to all three extracts. M. longifolia contained seven unique major compounds, including caryophyllene, genistein, and ursolic acid. S. orientalis had one unique compound, resveratrol, while E. biebersteinii uniquely possessed vitexin-4'-rhamnoside. Compound identification was performed based on methods previously described (see Materials and Methods; [29,30,38]).
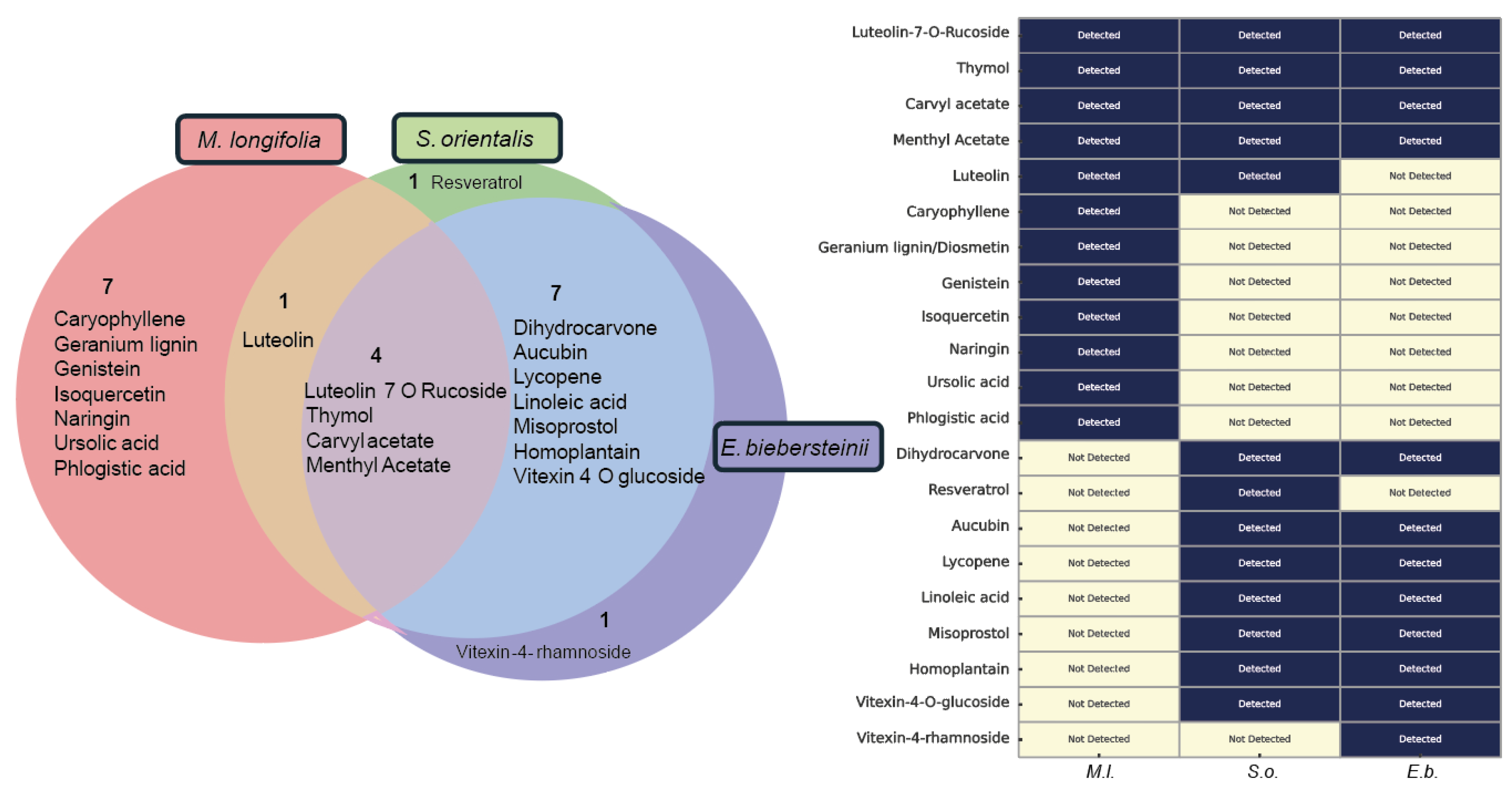
Figure 6.
Functional characterization of shared herbal compounds reveals their roles in DNA damage response, apoptosis, and antioxidant activity. (A) Expression levels of DNA damage checkpoint genes (atm-1 and atl-1) in response to treatment with four common herbal compounds. Young adult hermaphrodites were treated with luteolin-7-O-rutinoside (30 µM), thymol (3 mM), carvyl acetate (8 µg/ml), or menthyl acetate (10 µg/ml), and qRT-PCR was performed to assess expression of atm-1 and atl-1. Thymol and carvyl acetate significantly upregulated both genes, whereas luteolin-7-O-rutinoside and menthyl acetate showed no significant effect. Data are presented as fold change relative to control (mean ± SEM, n ≥ 20 animals per group). (B) Quantification of germline apoptosis in the pachytene region following compound treatment. Germline apoptosis was measured in wild-type C. elegans treated with the four shared compounds. Thymol and carvyl acetate significantly increased apoptotic cell counts, consistent with their induction of atm-1 and atl-1 checkpoint gene expression. Luteolin-7-O-rutinoside and menthyl acetate showed no significant effects. Data are presented as mean apoptotic nuclei per gonad. Mean ± SEM, n ≥ 20 animals per group. (C) DPPH radical scavenging activity of M.l, S.o, and E.b herb extracts at increasing concentrations. All three extracts exhibited dose-dependent antioxidant activity, with E.b showing the strongest effect (39.15% inhibition at 3 µg/ml, P < 0.0001), followed by S.o (30.95%, P < 0.0001) and M.l (23.05%, P < 0.0001). (D) Antioxidant activity of four common constituents found in the herb extracts at three different doses: luteolin-7-O-rutinoside (10–30 µM), thymol (1–3 mM), carvyl acetate (1–8 µg/ml), and menthyl acetate (1–10 µg/ml). Luteolin-7-O-rutinoside and thymol showed dose-dependent radical scavenging activity, with thymol demonstrating stronger inhibition (up to 18.08% at 3 mM). In contrast, carvyl acetate and menthyl acetate showed negligible activity at all tested concentrations. Data are presented as mean ± SEM. Statistical significance was calculated using two-tailed Mann-Whitney test.
Figure 6.
Functional characterization of shared herbal compounds reveals their roles in DNA damage response, apoptosis, and antioxidant activity. (A) Expression levels of DNA damage checkpoint genes (atm-1 and atl-1) in response to treatment with four common herbal compounds. Young adult hermaphrodites were treated with luteolin-7-O-rutinoside (30 µM), thymol (3 mM), carvyl acetate (8 µg/ml), or menthyl acetate (10 µg/ml), and qRT-PCR was performed to assess expression of atm-1 and atl-1. Thymol and carvyl acetate significantly upregulated both genes, whereas luteolin-7-O-rutinoside and menthyl acetate showed no significant effect. Data are presented as fold change relative to control (mean ± SEM, n ≥ 20 animals per group). (B) Quantification of germline apoptosis in the pachytene region following compound treatment. Germline apoptosis was measured in wild-type C. elegans treated with the four shared compounds. Thymol and carvyl acetate significantly increased apoptotic cell counts, consistent with their induction of atm-1 and atl-1 checkpoint gene expression. Luteolin-7-O-rutinoside and menthyl acetate showed no significant effects. Data are presented as mean apoptotic nuclei per gonad. Mean ± SEM, n ≥ 20 animals per group. (C) DPPH radical scavenging activity of M.l, S.o, and E.b herb extracts at increasing concentrations. All three extracts exhibited dose-dependent antioxidant activity, with E.b showing the strongest effect (39.15% inhibition at 3 µg/ml, P < 0.0001), followed by S.o (30.95%, P < 0.0001) and M.l (23.05%, P < 0.0001). (D) Antioxidant activity of four common constituents found in the herb extracts at three different doses: luteolin-7-O-rutinoside (10–30 µM), thymol (1–3 mM), carvyl acetate (1–8 µg/ml), and menthyl acetate (1–10 µg/ml). Luteolin-7-O-rutinoside and thymol showed dose-dependent radical scavenging activity, with thymol demonstrating stronger inhibition (up to 18.08% at 3 mM). In contrast, carvyl acetate and menthyl acetate showed negligible activity at all tested concentrations. Data are presented as mean ± SEM. Statistical significance was calculated using two-tailed Mann-Whitney test.
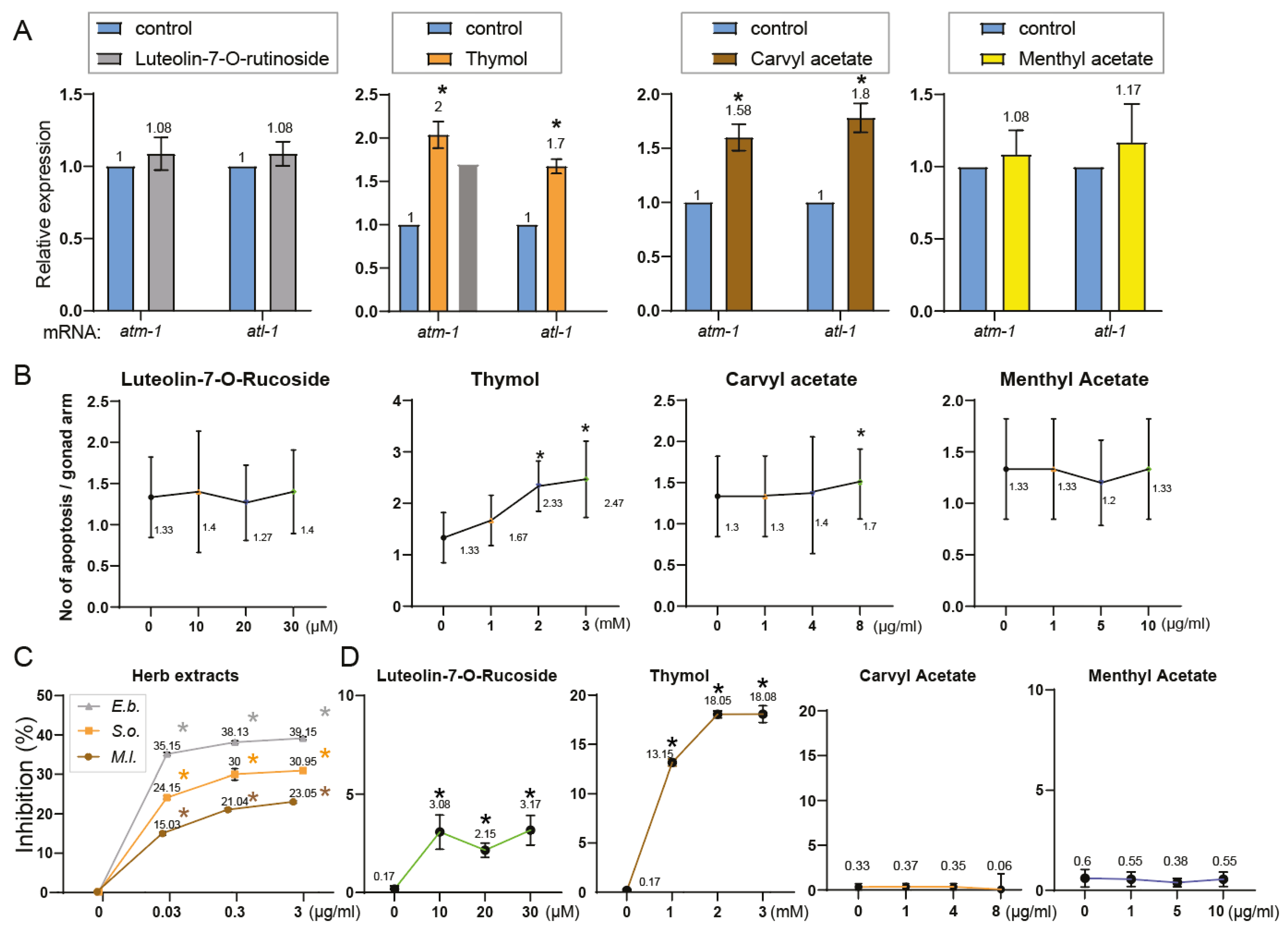

Table 1.
Taxonomy, Characteristics, and Distribution of Three Medicinal Herbs.
| Genus | Taxonomy | Characteristics | Distribution | Sample Used in this study |
|---|---|---|---|---|
| Mentha | Defined as 18–30 species across five sections: Mentha, Preslia, Audibertia, Eriodontes, Pulegium. Includes M. spicata, M. aquatica, M. arvensis, M. longifolia [20,21,96,97] | Aromatic, herbaceous perennials with extensive stolons [97] | Widely distributed: Northern Pakistan, Europe, Nepal, India, Western China, Germany, UK, Egypt, Nigeria, Turkey [21] | Mentha longifolia |
| Scrophularia | Genus Scrophularia (Scrophulariaceae); ~300 species [2] | Mostly herbaceous perennials; also subshrubs, biennials, or annuals [3] | Temperate Asia, Mediterranean Europe, North America [1] | Scrophularia orientalis |
| Echium | Genus Echium (Boraginaceae); ~60 species, 30 in Canary Islands, 24 endemic [9] | Annual, biennial, or perennial flowering plants [9,10] | Native to North Africa, Europe, Macaronesia (Azores, Madeira, Canaries, Cape Verde) [9,10] | Echium biebersteinii |
Table 2.
Reported biological functions of major compounds in three herbal extracts. Each compound—such as antioxidant, DNA damage response/repair, anti-tumor, and anti-inflammatory functions—are based on previous reports. However, many compounds remain insufficiently characterized and require further investigation.
Table 2.
Reported biological functions of major compounds in three herbal extracts. Each compound—such as antioxidant, DNA damage response/repair, anti-tumor, and anti-inflammatory functions—are based on previous reports. However, many compounds remain insufficiently characterized and require further investigation.
| No | Compounds | Antioxidant | DNA damage response/repair | Anti-tumor | Anti-inflammatory |
|---|---|---|---|---|---|
| 1 | Luteolin-7-O-Rucoside | [43] | [44] | [45] | [46] |
| 2 | Thymol | [47] | [48] | [49] | [50] |
| 3 | Carvyl acetate | ||||
| 4 | Menthyl Acetate | [51] | |||
| 5 | Luteolin | [52] | [53] | [54] | [55] |
| 6 | Caryophyllene | [56] | [57] | [58] | [59] |
| 7 | Geranium lignin/Diosmetin | [60] | [61] | [62] | [63] |
| 8 | Genistein | [64] | [65] | [66] | [67] |
| 9 | Isoquercetin | [68] | [69] | [70] | |
| 10 | Naringin | [71] | [72] | [73] | |
| 11 | Ursolic acid | [74] | [75] | [76] | [77] |
| 12 | Phlogistic acid | ||||
| 13 | Dihydrocarvone | ||||
| 14 | Resveratrol | [78] | [79] | [80] | [81] |
| 15 | Aucubin | [82] | [83] | [84] | |
| 16 | Lycopene | [85] | [86] | [87] | [88] |
| 17 | Linoleic acid | [215] | [90] | [91] | |
| 18 | Misoprostol | [92] | |||
| 19 | Homoplantain | [93] | |||
| 20 | Vitexin-4-O-glucoside | [94] | |||
| 21 | Vitexin-4-rhamnoside | [95] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



