
Submitted:
29 May 2025
Posted:
03 June 2025
You are already at the latest version
Abstract
Calcareous soils, characterized by high pH and calcium carbonate content, often limit the availability of essential nutrients for crops such as rice (Oryza sativa L.), reducing yield and nutritional quality. In this study, we evaluated the effect of the halotolerant yeast Debaryomyces hansenii (Dh) on the growth, nutrient uptake, and phosphorus acquisition mechanisms of rice plants cultivated in calcareous soil under controlled greenhouse conditions. Plants inoculated with Dh, particularly via root immersion, exhibited significantly higher SPAD chlorophyll index, plant height, dry matter content, and grain yield compared to controls. Foliar concentrations of Cu, Fe, Zn, and Mn also increased under certain conditions, suggesting improved micronutrient acquisition. Although phosphorus levels in leaves were not significantly enhanced, Dh stimulated acid phosphatase activity in both P-sufficient and P-deficient treatments. At the molecular level, Dh upregulated the expression of acid phosphatase genes (OsPAP3, OsPAP9) and a phosphate transporter gene (OsPTH1;6), confirming its influence on P-related physiological responses. These findings demonstrate that D. hansenii functions as a plant growth-promoting yeast (PGPY) and may serve as a promising biofertilizer for improving rice productivity and nutrient efficiency in calcareous soils, contributing to sustainable agricultural practices in challenging edaphic environments.
Keywords:
Debaryomyces hansenii
; calcareous soil
; rice
; growth-promotion
; P deficiency
; mineral nutrition
1. Introduction
Iron (Fe) and phosphorus (P) deficiencies are among the most critical agronomic challenges for global crop production, especially in calcareous soils. These soils, which occupy approximately 30% of the world’s agricultural land, are particularly prevalent in countries like Spain, where large areas in regions such as Aragón, Castilla-La Mancha, Andalucía, and the Basque Country are affected [1]. Characterized by a high calcium carbonate (CaCO3) content and alkaline pH levels above 7.5, calcareous soils limit the bioavailability of essential macro- and micronutrients including Fe, P, Zn, and Cu [2,3,4,5,6,7].
In these soils, nutrients such as Fe and P exist in forms that are largely inaccessible to plants. Phosphorus tends to precipitate with Ca2+ or Mg2+ into insoluble forms or become adsorbed onto soil particles and iron oxides, drastically reducing its mobility and availability to roots [8,9]. Similarly, Fe, particularly in its oxidized ferric (Fe3+) state, precipitates as hydroxides under high pH, resulting in chlorosis and growth inhibition in susceptible crops like rice [10]. Moreover, the availability of Zn and Cu is also severely restricted in calcareous soils due to pH-induced changes in solubility and competition with other cations such as Ca2+ [11,12].
Rice (Oryza sativa L.) is particularly vulnerable to nutrient imbalances in such soil conditions. Although rice belongs to the graminaceous group and partially relies on Strategy II Fe acquisition via phytosiderophore secretion, it secretes insufficient amounts of these chelators, making it highly susceptible to Fe deficiency [13]. Additionally, high soil pH conditions further exacerbate the problem by reducing the efficacy of Fe3+ chelation and uptake [14,15].
In this context, the use of plant growth-promoting yeasts (PGPY) such as Debaryomyces hansenii has emerged as a sustainable alternative to alleviate nutrient deficiencies. These microorganisms are known to improve plant nutrition by enhancing solubilization of mineral elements, modifying rhizospheric pH, and stimulating plant hormonal pathways [16,17]. Specifically, D. hansenii has been reported to induce Fe deficiency responses in cucumber [6] and to alleviate arsenic toxicity in rice, thereby promoting growth and nutrient status [18]. The potential of yeasts to solubilize phosphate through acidification of the medium, typically via secretion of organic acids such as citric acid, has been well documented for species like Yarrowia lipolytica, Rhodotorula sp., and Candida tropicalis [19,20]. Similarly, certain strains are capable of releasing Zn and K from insoluble mineral forms [21,22], supporting root development and biomass accumulation [23]. In addition to nutrient solubilization, PGPY strains including D. hansenii have demonstrated the capacity to synthesize phytohormones such as indole-3-acetic acid (IAA), gibberellins, cytokinins, and abscisic acid (ABA), which are integral to root proliferation, nutrient uptake, and stress tolerance [24,25].
The objective of this study is to evaluate the role of D. hansenii (strain CBS767) in promoting growth, development, and yield of rice cultivated in limestone soil. The study further investigates its capacity to modulate rhizospheric pH, activate acid phosphatase activity, and induce the expression of genes involved in phosphate acquisition. These insights may support the use of D. hansenii as a biofertilizer to enhance rice production under challenging calcareous soil conditions.
2. Material and Methods
2.1. Biological Material
Experiments were conducted using rice plants (Oryza sativa L. var. ‘Puntal’). Seeds were sterilized following the methodology described by Aparicio et al. [26]. They were sown on a layer of moist perlite at the bottom of a tray, to which 20 mL of a 5 mM CaCl2 solution was added. The seeds were covered with another layer of moist perlite, and the tray was sealed with a plastic bag to prevent desiccation. Germination was carried out in the dark at 27 °C for 4 days. After germination, the seedlings were transferred to a growth chamber maintained at 25 °C during the day and 22 °C at night, with 70% relative humidity and a 14 h photoperiod at an irradiance of 300 μmol m−2 s−1 for 7 days. Subsequently, seedlings were moved to either a hydroponic system or calcareous soil.
To transfer seedlings to the hydroponic system, they were removed from the perlite tray and thoroughly cleaned to remove root residues. The nutrient solution used was R&M [27], and aeration was continuously provided to avoid anoxia. Plants were kept under these conditions for 22 to 25 days, after which the treatments were applied.
For yeast inoculation, the wild-type genotype CBS767 of D. hansenii, obtained from the Dutch “Central Bureau von Schimmelcultures” (https://wi.knaw.nl/) and supplied by the Microbiology group at the University of Córdoba, was used. Yeast cultures were grown in YPD medium consisting of 2% D-glucose, 1% yeast extract, and 2% peptone [28].
2.2. Inoculation and Experimental Setup in Calcareous Soil
To evaluate the effects of D. hansenii on rice plants under field-like conditions, experiments were conducted in 2 L pots filled with calcareous soil. Seedlings previously grown in perlite trays were transplanted into the pots, and two inoculation methods were tested.
The soil used to fill the pots was obtained from Santa Cruz (Córdoba; 37◦47’03’’N 4◦36’35’’W) and sterilized or not at 121 °C for 50 min twice. The physical-chemical properties and phosphorus and iron availability for the plant in the sampled soil are shown in (Table 1).
In the pot experiments with calcareous soil, the following treatments were applied:
- Plants grown in previously sterilized calcareous soil.
- Plants grown in non-sterilized calcareous soil.
- Plants inoculated by root immersion and grown in sterilized calcareous soil.
- Plants inoculated by root immersion and grown in non-sterilized calcareous soil.
- Plants inoculated by surface irrigation and grown in sterilized calcareous soil.
- Plants inoculated by surface irrigation and grown in non-sterilized calcareous soil.
2.2.1. Root Inoculation
Some plants were inoculated by immersing their roots in a solution containing the desired inoculum concentration before transplanting. Roots were submerged in 1.5 L of a 107 cells/mL yeast suspension in deionized water under constant agitation for 30 min, ensuring effective contact between the yeast and the roots [29].
2.2.2. Irrigation Inoculation
For this method, pots were irrigated with the inoculum suspension (107 cells/mL in deionized water) until field capacity was reached.
2.3. Inoculation in Hydroponic System
The experiments were conducted using rice plants (Oryza sativa L. var. ‘Puntal’). The seeds were surface-sterilized as described by Aparicio et al. [26]. Subsequently, the seedlings were grouped in sets of eight and transferred to a hydroponic system. Each group of eight seedlings was placed in plastic lids and held in holes of a thin polyurethane sheet floating on an aerated nutrient solution R&M [27] containing 2 mM Ca(NO3)2, 0.75 mM K2SO4, 0.65 mM MgSO4, 0.5 mM KH2PO4, 50 μM KCl, 10 μM H3BO3, 1 μM MnSO4, 0.5 μM CuSO4, 0.5 μM ZnSO4, 0.05 μM (NH4)6Mo7O24, and 45 μM Fe-EDTA for Plants grown in complete nutrient solution. For plants grown in P-deficient nutrient solution 0.5 mM KH2PO4 was remplace by 0.5 mM KOH.
For hydroponic experiments, the yeast suspension (107 cells/mL) was directly added to the nutrient solution with and without phosphorus (P). Control treatments without inoculation were included. Plants were sampled at three time points (7, 9 and 11 days post-treatment) to assess acid phosphatase activity and gene expression. Six replicates were performed for each treatment.
The following treatments were applied:
- Plants grown in complete nutrient solution.
- Plants grown in complete nutrient solution plus inoculum.
- Plants grown in P-deficient nutrient solution.
- Plants grown in P-deficient nutrient solution plus inoculum.
2.4. Chlorophyll Content (SPAD)
Chlorophyll content was measured using a Minolta SPAD-502 (Konica Minolta, Tokyo [Japan]) portable device. Four readings were taken from the youngest fully extended leaf per plant, averaging the values for representation.
2.5. Growth Promotion and Yield Production
At the end of the growth cycle, rice plants from all treatments were harvested to measure fresh shoot weight. Samples were then dried at 75 °C for 3 days to obtain dry weight and calculate the dry matter percentage:
Rice grain yield was also determined, accounting for moisture percentage uniformity across treatments.
2.6. Elemental Analysis of Leaves
Dried leaves were homogenized using a grinder. Samples were digested with 3 mL of 65% HNO3 and incubated at room temperature for 16 h, followed by heating at 85 °C for 1.5 h. When vapor began to appear, 1 mL of 60% HClO4 was added, and heating continued until white vapor indicated complete digestion. Samples were diluted to 10 mL with deionized water. Elemental analysis (Zn, Fe, Cu, Mn) was performed using flame atomic absorption spectrometry. Phosphorus was determined using the molybdovanadate method.
2.7. Acid Phosphatase Determination
Acid phosphatase activity was evaluated using the BCIP substrate, which turns blue upon enzymatic dephosphorylation. Roots were incubated in 0.01% BCIP solution for 2 h to assess enzyme activity visually [30].
2.8. Gene Expression Analysis by qRT-PCR
RNA extraction was carried out using Tri Reagent following the manufacturer’s protocol. RNA concentration was measured at 260 nm. Complementary DNA (cDNA) synthesis was performed using M-MLV reverse transcriptase and random hexamers. Gene expression was analysed via qRT-PCR, with 40 amplification cycles and SYBR Green PCR Master Mix and using the primers listed in Table 2.
2.9. Statistical Analysis
Data normality and variance homogeneity were verified. ANOVA and post hoc Tukey’s or Dunnett’s tests (p < 0.05) were applied to compare treatments. All statistical analyses were conducted using GraphPad Prism v9, and the graphs were generated using Microsoft Excel.
3. Results
3.1. Effect of Debaryomyces hansenii Inoculation on SPAD Index
SPAD measurements were taken every 15 days throughout the rice crop cycle. As shown in Figure 1a, SPAD index levels followed a bell-shaped curve, with a peak observed around 85 days after sowing. At this stage, all treatments reached their maximum SPAD values. Statistical analysis revealed significant differences between treatments. Notably, plants grown in previously sterilized calcareous soil and inoculated with D. hansenii, either via surface irrigation or root immersion, exhibited significantly higher SPAD index values compared to the control plants (Figure 1b).
3.2. Effect of Debaryomyces hansenii on Dry Matter Content, Plant Height, and Grain Yield
The inoculation of rice plants with D. hansenii significantly affected key agronomic parameters, particularly when applied via root immersion in non-sterilized calcareous soil. As illustrated in Figure 2a, the treatment NSS-Dh Inmersion led to the highest dry matter percentage, showing statistically significant differences compared to NSS-Control and all other inoculation methods, regardless of soil sterilization. This increase in biomass may reflect enhanced nutrient uptake and photosynthetic efficiency, likely associated with improved root architecture or beneficial interactions at the rhizosphere level. This trend was also observed in grain yield per plant (Figure 2b), where NSS-Dh Inmersion resulted in a significant increase, exceeding 2.0 g on average and clearly outperforming all other treatments. In contrast, plants inoculated via irrigation (NSS-Dh Irrigation and SS-Dh Irrigation) produced lower grain yields than their respective controls, highlighting the importance of the application method in determining the effectiveness of the yeast. Interestingly, no significant differences in dry matter content or grain production were detected between the control treatments grown in sterilized (SS-Control) and non-sterilized (NSS-Control) soils, suggesting that the native soil microbiota alone was not responsible for the observed improvements. Instead, the data support the hypothesis of a synergistic interaction between D. hansenii and the native microbial community, particularly under conditions that favor root colonization. Overall, these findings confirm that root immersion is a more efficient strategy for delivering D. hansenii, likely due to improved establishment at the root-soil interface, and reinforce its potential as a microbial bioinoculant to enhance crop productivity under stress-prone conditions such as calcareous soils.
In line with these findings, visual assessment of plant height at 90 days after sowing (Figure 3) revealed that the tallest plants corresponded to the Dh Inmersion treatment, both in sterilized and non-sterilized calcareous soils. These plants clearly outperformed those from all other treatments in terms of shoot length. This enhanced shoot elongation further supports the plant growth-promoting effect of D. hansenii when applied via root immersion, reinforcing its potential role in stimulating vegetative development under calcareous soil conditions.
3.3. Effect of Debaryomyces hansenii on Leaf Content of Cu, Fe, Zn, Mn, and P in Rice Plants
The nutrient content in plant foliage is essential for photosynthesis, growth and development, grain yield and quality, as well as for overall plant health and stress tolerance (Marschner, 2012). As shown in Figure 4, the effect of D. hansenii inoculation on the foliar content of Cu, Fe, Zn, Mn, and P in rice plants grown in calcareous soil varied depending on the element and treatment condition.
In the case of copper (Cu), the highest concentrations were found in control plants grown in non-sterilized soil, whereas lower values were detected in plants cultivated in sterilized soil. Interestingly, D. hansenii inoculation, regardless of the method, only resulted in significantly higher Cu content compared to the control when plants were grown in sterilized soil (Figure 4).
Regarding iron (Fe), the treatment NSS-Dh Inmersion (non-sterilized soil, inoculated via root immersion) resulted in the highest foliar Fe levels, suggesting that this application mode may enhance Fe uptake. A similar trend was observed for zinc (Zn), with the highest Zn content also recorded in NSS-Dh Inmersion. These results are closely mirrored in the manganese (Mn) data, where the same treatment led to the most elevated Mn levels.
Phosphorus (P) content, shown in Figure 4, was consistently higher in all treatments involving non-sterilized soil, regardless of D. hansenii inoculation. However, no significant increases in P content were detected in yeast-inoculated plants compared to their respective controls. Conversely, the lowest P concentrations were observed in sterilized soil treatments inoculated with D. hansenii, suggesting a possible interaction between the yeast and the native microbial community that may influence phosphorus dynamics.
3.4. Effect of Debaryomyces hansenii on the Physiological Mechanism of Acid Phosphatase Activity in Rice Plants
As extensively described in the literature, plants can only assimilate phosphorus in its inorganic form (Pi). Therefore, acid phosphatases play a critical role by hydrolyzing organic phosphorus compounds in the soil into Pi, which can be absorbed by plant roots. The degree of acid phosphatase induction is visually estimated by the intensity of blue staining in the roots: the more intense the coloration, the greater the enzymatic activity.
Three sampling points were conducted at 7, 9, and 11 days after treatment application (data). As shown in Figure 5, plants subjected to phosphorus deficiency (-P) exhibited stronger blue staining than those cultivated under phosphorus-sufficient conditions (+P), confirming a higher induction of acid phosphatase activity in response to nutrient stress. On the other hand, the effect of D. hansenii under sufficient phosphorus conditions (+P+Dh) was not clearly visible during the first two sampling points (7 and 9 data). However, by 11 (data), a more pronounced induction of acid phosphatase activity was observed in this treatment, indicating a delayed but positive effect of the yeast.
Under phosphorus-deficient conditions (-P), the effect of D. hansenii on acid phosphatase activity became apparent as early as the first sampling point. The highest level of induction compared to the corresponding control was observed at 9 (data) and remained elevated at 11 (data), suggesting a sustained stimulatory effect of the yeast under nutrient-limiting conditions.
3.5. Effect of Debaryomyces hansenii on the Expression of Genes Related to Acid Phosphatase Activity and Phosphorus Transport
Gene expression analysis of acid phosphatases and the Pi transporter, shown in Figure 6, confirms the inductive role of D. hansenii at the molecular level. The PAP gene family is closely associated with phosphorus acquisition in plants, as these genes encode acid phosphatases, enzymes whose activity was previously demonstrated in Figure 5.
The gene OsPAP9 showed a significant increase in expression at 7 and 9 days after treatment application (data) in plants grown under phosphorus-sufficient conditions inoculated with D. hansenii (Figure 6a). Under phosphorus-deficient conditions, expression of OsPAP9 was also enhanced, with the most significant differences compared to the control observed at 9 and 11 (data) (Figure 6b). Similarly, OsPAP3 expression was significantly upregulated at 7 and 9 (data) under phosphorus sufficiency (Figure 6c), and at 11 (data) under phosphorus deficiency (Figure 6d), suggesting a time- and condition-dependent response to the presence of the yeast.
As for the gene encoding the phosphorus transporter analyzed in this study, OsPTH1;6, its relative expression level increased significantly compared to the control at 9 (data), both under phosphorus-sufficient and deficient conditions (Figure 6e, f). These data indicate that D. hansenii not only enhances enzymatic activity related to Pi acquisition but also stimulates molecular pathways involved in phosphorus uptake and transport.
4. Discussion
Rice plants cultivated in calcareous soil often face multiple nutrient deficiencies due to the high pH and the specific physicochemical characteristics of these soils. Typically, calcareous soils have a pH above 7.5 and contain significant amounts of calcium carbonate (CaCO3). The elevated pH can limit the availability of certain nutrients, while high calcium content may interfere with the uptake of others. Among the nutrients whose availability is commonly restricted under these conditions are iron (Fe), copper (Cu), manganese (Mn) [11], zinc (Zn) [31], and phosphorus (P) [32], among others. These deficiencies often result in stunted growth, reduced tillering, and lower grain yields [33]. Additionally, they can impair grain filling, leading to poorer grain quality and reduced market value [34]. Nutrient-deficient plants are also more susceptible to pest and disease attacks [35], which translates into economic losses for farmers [36].
Numerous studies have shown that beneficial soil microorganisms provide promising strategies to address nutrient deficiencies, especially in problematic soils such as calcareous soils. Arbuscular mycorrhizal fungi (AMF) are among the most widely used, as they enhance nutrient uptake by forming extensive hyphal networks that improve the absorption of phosphorus and several micronutrients that are often poorly available in high-pH soils [37]. Plant growth-promoting bacteria (PGPB), such as Pseudomonas spp., produce organic acids and siderophores that chelate micronutrients, thereby increasing their availability in alkaline environments [38]. Other microbes can release organic acids that slightly lower the soil pH, improving nutrient solubility [39]. Additionally, microorganisms that trigger induced systemic resistance (ISR) have been shown to enhance iron acquisition by activating deficiency-response mechanisms [40,41].
Plant growth-promoting yeasts (PGPY) have gained interest in recent years due to their ability to colonize plant tissues and produce phytohormones, thereby enhancing nutrient availability and soil fertility [42]. This chapter aimed to explore the potential effects of the yeast D. hansenii (Dh) on rice plant development in calcareous soil. This yeast is considered halotolerant, capable of growing in environments with high salt concentrations (5–15%) and also under low salinity (< 0.1 M) [43,44]. These features make it a strong candidate for application in calcareous soil systems. If its ability to enhance rice development is confirmed, it could be used as a biofertilizer.
Indeed, our results demonstrated that D. hansenii promoted rice plant growth by increasing chlorophyll content (SPAD index), plant height, dry matter accumulation, and grain production (Figure 1, Figure 2 and Figure 3). To our knowledge, no previous studies have directly reported the effect of D. hansenii on chlorophyll content in rice. However, environmental factors such as light intensity, temperature, and nutrient availability, which can be influenced by microbial inoculants, play key roles in chlorophyll biosynthesis and, consequently, plant development and productivity.
Moreover, increases in foliar concentrations of Cu, Fe, Zn, and Mn were observed in plants inoculated with D. hansenii under specific cultivation conditions (Figure 4). Kaur et al. [18] reported improved growth and nutritional status in rice plants inoculated with D. hansenii. Similar results have been obtained with other microorganisms in different crops. For example, Pseudomonas aeruginosa and Enterobacter spp. promoted the growth of alfalfa under alkaline conditions [45], while Agrobacterium, Bacillus, and Alcaligenes strains enhanced growth and mineral nutrition in strawberry plants [46].
Although no clear increase in phosphorus content was detected in inoculated plants (Figure 4, D. hansenii significantly induced acid phosphatase activity under both phosphorus-sufficient and -deficient conditions (Figure 5). It is well established that arbuscular mycorrhizal fungi enhance P uptake by increasing acid phosphatase activity in the root zone, thereby aiding in the solubilization of organic phosphorus compounds [47]. Certain endophytic strains such as Pantoea and Colletotrichum have also been shown to produce acid phosphatases and improve phosphorus uptake in plants [48].
The activation of physiological responses to phosphorus deficiency—such as increased acid phosphatase activity—is regulated at the transcriptional level by specific genes. According to Zhang et al. [49], only ten PAP genes in rice are induced under P-deficient conditions. In this study, we examined two of these genes, which showed higher expression levels in plants inoculated with D. hansenii (Figures 6a–d). Arbuscular mycorrhizal fungi are known to upregulate PAP genes in symbiosis with rice roots [50], and phosphate-solubilizing bacteria have also been shown to influence PAP gene expression in rice [51]. Additionally, the gene encoding the phosphate transporter analyzed in this study, OsPTH1;6, was upregulated under both phosphorus-sufficient and deficient conditions (Figures 6), suggesting that this yeast may contribute to enhanced P acquisition in rice.
5. Conclusions
In conclusion, the yeast D. hansenii demonstrated a clear capacity to promote rice plant growth in calcareous soils, particularly when applied via root immersion. This treatment resulted in significant improvements in physiological parameters such as chlorophyll content (SPAD index), plant height, dry matter accumulation, and grain yield. These benefits were especially evident under non-sterilized soil conditions, suggesting that Dh may act synergistically with the native soil microbiota to enhance plant performance.
In addition to promoting general growth, D. hansenii contributed to an increase in the foliar content of essential micronutrients such as copper, iron, zinc, and manganese—nutrients that are commonly limited in availability in calcareous soils due to their high pH and calcium content. Although the inoculation did not produce a marked increase in phosphorus content in leaf tissue, D. hansenii did significantly enhance the activity of acid phosphatases, a key enzymatic mechanism in phosphorus mobilization. This induction was observed under both phosphorus-sufficient and phosphorus-deficient conditions, indicating that D. hansenii may play a role in improving P availability in the rhizosphere. At the molecular level, Dh upregulated the expression of genes associated with phosphorus acquisition (OsPAP3 and OsPAP9) and phosphorus transport (OsPTH1;6), highlighting its potential influence on nutrient-related gene regulatory networks. These findings collectively support the role of D. hansenii as a plant growth-promoting yeast (PGPY) with significant biotechnological potential. Its ability to thrive under saline and alkaline conditions, along with its demonstrated benefits on plant growth, nutrient uptake, and gene expression, make it a strong candidate for development as a biofertilizer in sustainable rice cultivation strategies.
References
- Moreno-Jiménez, E.; Plaza, C.; Saiz, H.; Manzano, R.; Flagmeier, M.; Maestre, F.T. Aridity and Reduced Soil Micronutrient Availability in Global Drylands. Nat. Sustain. 2019, 2, 371–377. [CrossRef]
- Abel, S.; Ticconi, C.A.; Delatorre, C.A. Phosphate Sensing in Higher Plants. Physiologia Plantarum 2002, 115, 1–8. [CrossRef]
- López-Garrido, R.; Parras-Acántara, L.; Lozano-Garcia, B.; Giráldez, J. Los suelos calizos en la península ibérica. Avances en la investigación sobre suelos y desarrollo sostenible. Universidad de Granada, 2014, 49-70.
- García-Ruiz, R.; Morugan-Coronado, A.; Lavee, H. Suelos calizos en ambientes semiáridos y áridos. Suelos y Desarrollo Sostenible. Universidad de Granada 2014, 327-354.
- Lucena, C.; Porras, R.; Romera, F.J.; Alcántara, E.; García, M.J.; Pérez-Vicente, R. Similarities and Differences in the Acquisition of Fe and P by Dicot Plants. Agronomy 2018, 8, 148. [CrossRef]
- Lucena, C.; Alcalá-Jiménez, M.T.; Romera, F.J.; Ramos, J. Several Yeast Species Induce Iron Deficiency Responses in Cucumber Plants (Cucumis Sativus L.). Microorganisms 2021, 9, 2603. [CrossRef]
- Sevillano-Cano, J.; García, M.J.; Córdoba-Galván, C.; Luque-Cruz, C.; Agustí-Brisach, C.; Lucena, C.; Ramos, J.; Pérez-Vicente, R.; Romera, F.J. Exploring the Role of Debaryomyces hansenii as Biofertilizer in Iron-Deficient Environments to Enhance Plant Nutrition and Crop Production Sustainability. Int. J. Mol. Sci. 2024, 25, 5729. [CrossRef]
- Matar, A.; Torrent, J.; Ryan, J. Soil and Fertilizer Phosphorus and Crop Responses in the Dryland Mediterranean Zone. In Soil Restoration; Lal, R., Stewart, B.A., Eds.; Advances in Soil Science; Springer New York: New York, NY, 1992; Vol. 17, pp. 81–146 ISBN 978-1-4612-7684-5.
- Hirsch, J.; Marin, E.; Floriani, M.; Chiarenza, S.; Richaud, P.; Nussaume, L.; Thibaud, M.C. Phosphate Deficiency Promotes Modification of Iron Distribution in Arabidopsis Plants. Biochimie 2006, 88, 1767–1771. [CrossRef]
- Mori, S. Iron Acquisition by Plants. Curr. Opin. Plant Biol. 1999, 2, 250–253. [CrossRef]
- Marschner, H. Mineral Nutrition of Higher Plants; 2nd edition.; Academic Press: London San Diego, 1995; ISBN 978-0-12-473542-2.
- Kabata-Pendias, A.; Mukherjee, A.B. Trace Elements from Soil to Human; Springer: Berlin New York, 2007; ISBN 978-3-540-32714-1.
- Mori, S.; Nishizawa, N.; Hayashi, H.; Chino, M.; Yoshimura, E.; Ishihara, J. Why Are Young Rice Plants Highly Susceptible to Iron Deficiency? In Iron Nutrition and Interactions in Plants; Chen, Y., Hadar, Y., Eds.; Springer Netherlands: Dordrecht, 1991; pp. 175–188 ISBN 978-94-010-5455-3.
- Takagi, S. Naturally Occurring Iron-Chelating Compounds in Oat- and Rice-Root Washings: I. Activity Measurement and Preliminary Characterization. Soil Sci. Plant Nutr. 1976, 22, 423–433. [CrossRef]
- Marschner, H.; Romheld, V.; Kissel, M. Different Strategies in Higher Plants in Mobilization and Uptake of Iron. J. Plant Nutr. 1986, 9, 695–713. [CrossRef]
- Nimsi, K.A.; Manjusha, K.; Kathiresan, K.; Arya, H. Plant Growth-Promoting Yeasts (PGPY), the Latest Entrant for Use in Sustainable Agriculture: A Review. Journal of Applied Microbiology 2023, 134, lxac088. [CrossRef]
- Mukherjee, S.; Sen, S.K. Exploration of Novel Rhizospheric Yeast Isolate as Fertilizing Soil Inoculant for Improvement of Maize Cultivation. J. Sci. Food Agric. 2015, 95, 1491–1499. [CrossRef]
- Kaur, J.; Anand, V.; Srivastava, S.; Bist, V.; Tripathi, P.; Naseem, M.; Nand, S.; Anshu; Khare, P.; Srivastava, P.K.; et al. Yeast Strain Debaryomyces Hansenii for Amelioration of Arsenic Stress in Rice. Ecotoxicol. Environ. Saf. 2020, 195, 110480. [CrossRef]
- Mundra, S.; Arora, R.; Stobdan, T. Solubilization of Insoluble Inorganic Phosphates by a Novel Temperature-, pH-, and Salt-Tolerant Yeast, Rhodotorula Sp. PS4, Isolated from Seabuckthorn Rhizosphere, Growing in Cold Desert of Ladakh, India. World J. Microbiol. Biotechnol. 2011, 27, 2387–2396. [CrossRef]
- Hesham, A.E.-L. Molecular Genetic Identification of Yeast Strains Isolated from Egyptian Soils for Solubilization of Inorganic Phosphates and Growth Promotion of Corn Plants. J. Microbiol. Biotechnol. 2011, 21, 55–61. [CrossRef]
- Fu, S.-F.; Sun, P.-F.; Lu, H.-Y.; Wei, J.-Y.; Xiao, H.-S.; Fang, W.-T.; Cheng, B.-Y.; Chou, J.-Y. Plant Growth-Promoting Traits of Yeasts Isolated from the Phyllosphere and Rhizosphere of Drosera Spatulata Lab. Fungal Biology 2016, 120, 433–448. [CrossRef]
- Rosa-Magri, M.M.; Avansini, S.H.; Lopes-Assad, M.L.; Tauk-Tornisielo, S.M.; Ceccato-Antonini, S.R. Release of Potassium from Rock Powder by the Yeast Torulaspora Globosa. Braz. arch. biol. technol. 2012, 55, 577–582. [CrossRef]
- Mohamed, H.M.; El-Homosy, R.F.; Abd-Ellatef, A.-E.H.; Salh, F.M.; Hussein, M.Y. Identification of Yeast Strains Isolated from Agricultural Soils for Releasing Potassium-Bearing Minerals. Geomicrobiology Journal 2017, 34, 261–266. [CrossRef]
- Nutaratat, P.; Amsri, W.; Srisuk, N.; Arunrattiyakorn, P.; Limtong, S. Indole-3-Acetic Acid Production by Newly Isolated Red Yeast Rhodosporidium Paludigenum. J. Gen. Appl. Microbiol. 2015, 61, 1–9. [CrossRef]
- Arnesen, J.A.; Kildegaard, K.R.; Cernuda Pastor, M.; Jayachandran, S.; Kristensen, M.; Borodina, I. Yarrowia Lipolytica Strains Engineered for the Production of Terpenoids. Front. Bioeng. Biotechnol. 2020, 8, 945. [CrossRef]
- Aparicio, M.A.; Lucena, C.; García, M.J.; Ruiz-Castilla, F.J.; Jiménez-Adrián, P.; López-Berges, M.S.; Prieto, P.; Alcántara, E.; Pérez-Vicente, R.; Ramos, J.; et al. The Nonpathogenic Strain of Fusarium Oxysporum FO12 Induces Fe Deficiency Responses in Cucumber (Cucumis Sativus L.) Plants. Planta 2023, 257, 50. [CrossRef]
- Römheld, V.; Marschner, H. Iron Deficiency Stress Induced Morphological and Physiological Changes in Root Tips of Sunflower. Physiologia Plantarum 1981, 53, 354–360. [CrossRef]
- Sherman, F. Getting Started with Yeast. In Methods in Enzymology; Elsevier, 1991; Vol. 194, pp. 3–21 ISBN 978-0-12-182095-4.
- Navarro-Velasco, G.Y.; Prados-Rosales, R.C.; Ortíz-Urquiza, A.; Quesada-Moraga, E.; Di Pietro, A. Galleria Mellonella as Model Host for the Trans-Kingdom Pathogen Fusarium Oxysporum. Fungal Genet. Biol. 2011, 48, 1124–1129. [CrossRef]
- Zakhleniuk, O.V.; Raines, C.A.; Lloyd, J.C. Pho3 : A Phosphorus-Deficient Mutant of Arabidopsis Thaliana (L.) Heynh. Planta 2001, 212, 529–534. [CrossRef]
- Alloway, B.J. Zinc in Soils and Crop Nutrition; International Zinc Association (IZA), 2008.
- Brady, N.C.; Weil, R.R. The Nature and Properties of Soils; 14th ed.; Pearson Prentice Hall, 2008.
- Yoshida, S. Fundamentals of Rice Crop Science; Int. Rice Res. Inst., 1981.
- Juliano, B.O. Criteria and Tests for Rice Grain Qualities. In: Rice Chemistry and Technology, 2nd Edition, American Association of Cereal Chemists, 1985, pp. 443-524.
- Savary, S. Direct and Indirect Effects of Nitrogen Supply and Disease Source Structure on Rice Sheath Blight Spread. Phytopathology 1995, 85, 959. [CrossRef]
- Pingali, P.L. From Subsistence to Commercial Production Systems: The Transformation of Asian Agriculture. American J. Agri. Economics 1997, 79, 628–634. [CrossRef]
- Smith, S.E.; Read, D. Mycorrhizal Symbiosis; Elsevier, 2008; ISBN 978-0-12-370526-6.
- Sharma, A.; Johri, B.N.; Sharma, A.K.; Glick, B.R. Plant Growth-Promoting Bacterium Pseudomonas Sp. Strain GRP3 Influences Iron Acquisition in Mung Bean (Vigna Radiata L. Wilzeck). Soil Biol. Bioch. 2003, 35, 887–894. [CrossRef]
- Rousk, J.; Bååth, E.; Brookes, P.C.; Lauber, C.L.; Lozupone, C.; Caporaso, J.G.; Knight, R.; Fierer, N. Soil Bacterial and Fungal Communities across a pH Gradient in an Arable Soil. The ISME Journal 2010, 4, 1340–1351. [CrossRef]
- Zamioudis, C.; Korteland, J.; Van Pelt, J.A.; Van Hamersveld, M.; Dombrowski, N.; Bai, Y.; Hanson, J.; Van Verk, M.C.; Ling, H.; Schulze-Lefert, P.; et al. Rhizobacterial Volatiles and Photosynthesis-related Signals Coordinate MYB 72 Expression in Arabidopsis Roots during Onset of Induced Systemic Resistance and Iron-deficiency Responses. The Plant Journal 2015, 84, 309–322. [CrossRef]
- Romera, F.J.; García, M.J.; Lucena, C.; Martínez-Medina, A.; Aparicio, M.A.; Ramos, J.; Alcántara, E.; Angulo, M.; Pérez-Vicente, R. Induced Systemic Resistance (ISR) and Fe Deficiency Responses in Dicot Plants. Front. Plant Sci. 2019, 10, 287. [CrossRef]
- Streletskii, R.A.; Kachalkin, A.V.; Glushakova, A.M.; Yurkov, A.M.; Demin, V.V. Yeasts Producing Zeatin. PeerJ 2019, 7, e6474. [CrossRef]
- Kushner, D. Microbial Life in Extreme Environments; Acad. Pr: London, 1978; ISBN 978-0-12-430250-1.
- Prista, C.; Michán, C.; Miranda, I.M.; Ramos, J. The Halotolerant Debaryomyces Hansenii , the Cinderella of Non-conventional Yeasts. Yeast 2016, 33, 523–533. [CrossRef]
- Liu, J.; Tang, L.; Gao, H.; Zhang, M.; Guo, C. Enhancement of Alfalfa Yield and Quality by Plant Growth-promoting Rhizobacteria under Saline-alkali Conditions. J. Sci. Food Agric. 2019, 99, 281–289. [CrossRef]
- Ipek, M.; Pirlak, L.; Esitken, A.; Figen Dönmez, M.; Turan, M.; Sahin, F. Plant Growth-Promoting Rhizobacteria (Pgpr) Increase Yield, Growth And Nutrition Of Strawberry Under High-Calcareous Soil Conditions. J. Plant Nutr. 2014, 37, 990–1001. [CrossRef]
- Tarafdar, J.C.; Marschner, H. Phosphatase Activity in the Rhizosphere and Hyphosphere of VA Mycorrhizal Wheat Supplied with Inorganic and Organic Phosphorus. Soil Biol. Bioch. 1994, 26, 387–395. [CrossRef]
- Khan, A.L.; Waqas, M.; Kang, S.-M.; Al-Harrasi, A.; Hussain, J.; Al-Rawahi, A.; Al-Khiziri, S.; Ullah, I.; Ali, L.; Jung, H.-Y.; et al. Bacterial Endophyte Sphingomonas Sp. LK11 Produces Gibberellins and IAA and Promotes Tomato Plant Growth. J Microbiol. 2014, 52, 689–695. [CrossRef]
- Zhang, Q.; Wang, C.; Tian, J.; Li, K.; Shou, H. Identification of Rice Purple Acid Phosphatases Related to Posphate Starvation Signalling. Plant Biol. 2011, 13, 7–15. [CrossRef]
- Yang, S.-Y.; Grønlund, M.; Jakobsen, I.; Grotemeyer, M.S.; Rentsch, D.; Miyao, A.; Hirochika, H.; Kumar, C.S.; Sundaresan, V.; Salamin, N.; et al. Nonredundant Regulation of Rice Arbuscular Mycorrhizal Symbiosis by Two Members of the PHOSPHATE TRANSPORTER1 Gene Family. Plant Cell 2012, 24, 4236–4251. [CrossRef]
- Sharma, S.B.; Sayyed, R.Z.; Trivedi, M.H.; Gobi, T.A. Phosphate Solubilizing Microbes: Sustainable Approach for Managing Phosphorus Deficiency in Agricultural Soils. SpringerPlus 2013, 2, 587. [CrossRef]
Figure 1.
Effect of D. hansenii (Dh) on SPAD index in rice plants grown in calcareous soil under controlled greenhouse conditions. Plants were initially maintained in perlite for 15 days, then transferred to 2 L pots containing calcareous soil. The pots were placed in trays with a constant water layer throughout the crop cycle (120 days). SPAD index was recorded every 15 days. a) Temporal evolution of SPAD index during the phenological cycle of rice plants. b) Statistical analysis based on repeated-measures ANOVA and post-hoc Tukey HSD test at p < 0.05. Values are means ± SE (n = 80). Different letters indicate statistically significant differences among treatments.
Figure 1.
Effect of D. hansenii (Dh) on SPAD index in rice plants grown in calcareous soil under controlled greenhouse conditions. Plants were initially maintained in perlite for 15 days, then transferred to 2 L pots containing calcareous soil. The pots were placed in trays with a constant water layer throughout the crop cycle (120 days). SPAD index was recorded every 15 days. a) Temporal evolution of SPAD index during the phenological cycle of rice plants. b) Statistical analysis based on repeated-measures ANOVA and post-hoc Tukey HSD test at p < 0.05. Values are means ± SE (n = 80). Different letters indicate statistically significant differences among treatments.
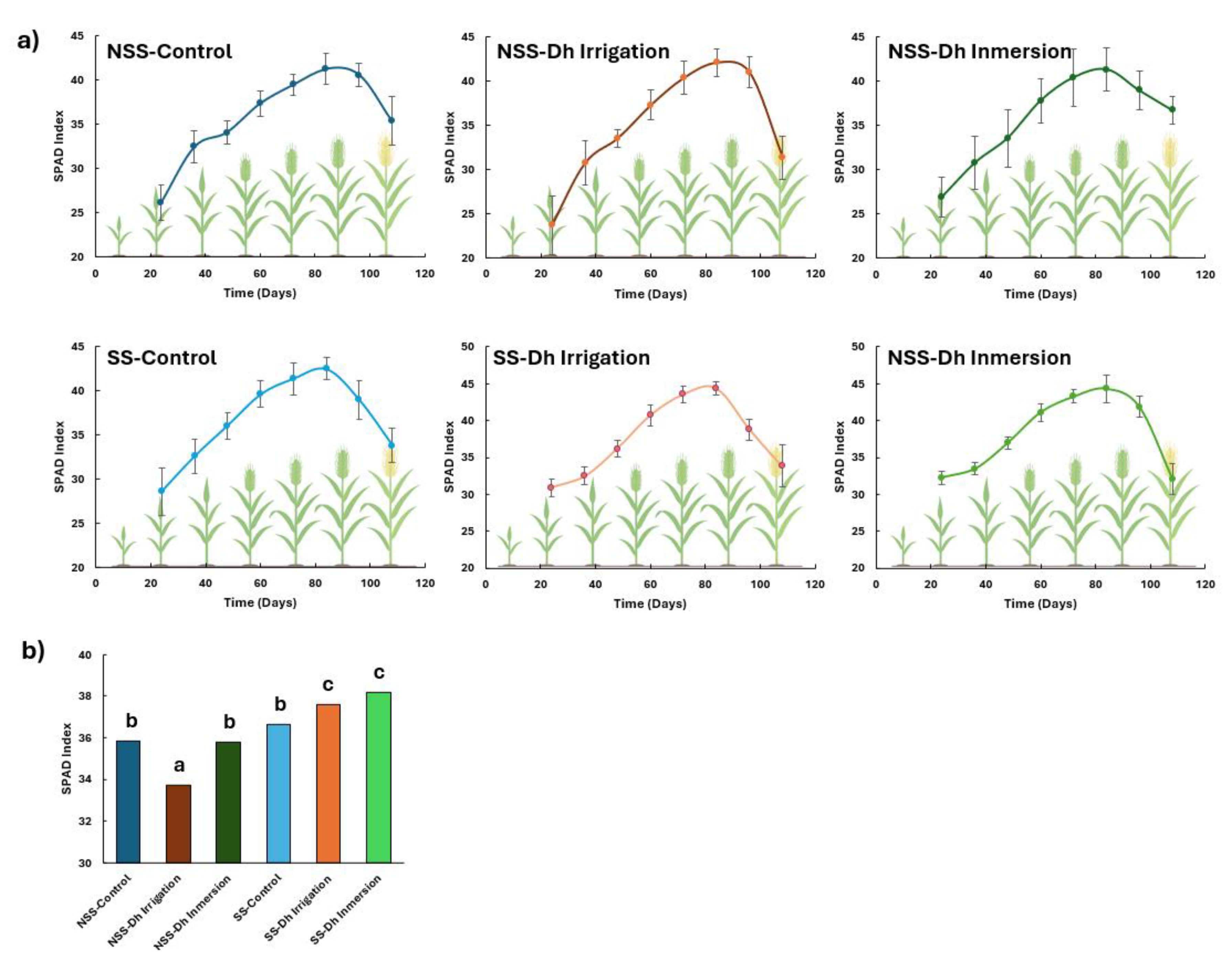
Figure 2.
Effect of D. hansenii (Dh) on (a) dry matter percentage and (b) grain yield per plant (g) in rice cultivated in calcareous soil under controlled greenhouse conditions. Experiments were carried out using both sterilized and non-sterilized soils. Treatments included: NSS-Control (non-sterilized soil, control), NSS-Dh Irrigation (non-sterilized soil inoculated with D. hansenii via irrigation), NSS-Dh Inmersion (non-sterilized soil inoculated with D. hansenii via root immersion), SS-Control (sterilized soil, control), SS-Dh Irrigation (sterilized soil inoculated with D. hansenii via irrigation), and SS-Dh Inmersion (sterilized soil inoculated with D. hansenii via root immersion). Different letters indicate statistically significant differences among treatments according to Tukey’s HSD test at p < 0.05. Values are expressed as means ± standard error (n = 10).
Figure 2.
Effect of D. hansenii (Dh) on (a) dry matter percentage and (b) grain yield per plant (g) in rice cultivated in calcareous soil under controlled greenhouse conditions. Experiments were carried out using both sterilized and non-sterilized soils. Treatments included: NSS-Control (non-sterilized soil, control), NSS-Dh Irrigation (non-sterilized soil inoculated with D. hansenii via irrigation), NSS-Dh Inmersion (non-sterilized soil inoculated with D. hansenii via root immersion), SS-Control (sterilized soil, control), SS-Dh Irrigation (sterilized soil inoculated with D. hansenii via irrigation), and SS-Dh Inmersion (sterilized soil inoculated with D. hansenii via root immersion). Different letters indicate statistically significant differences among treatments according to Tukey’s HSD test at p < 0.05. Values are expressed as means ± standard error (n = 10).
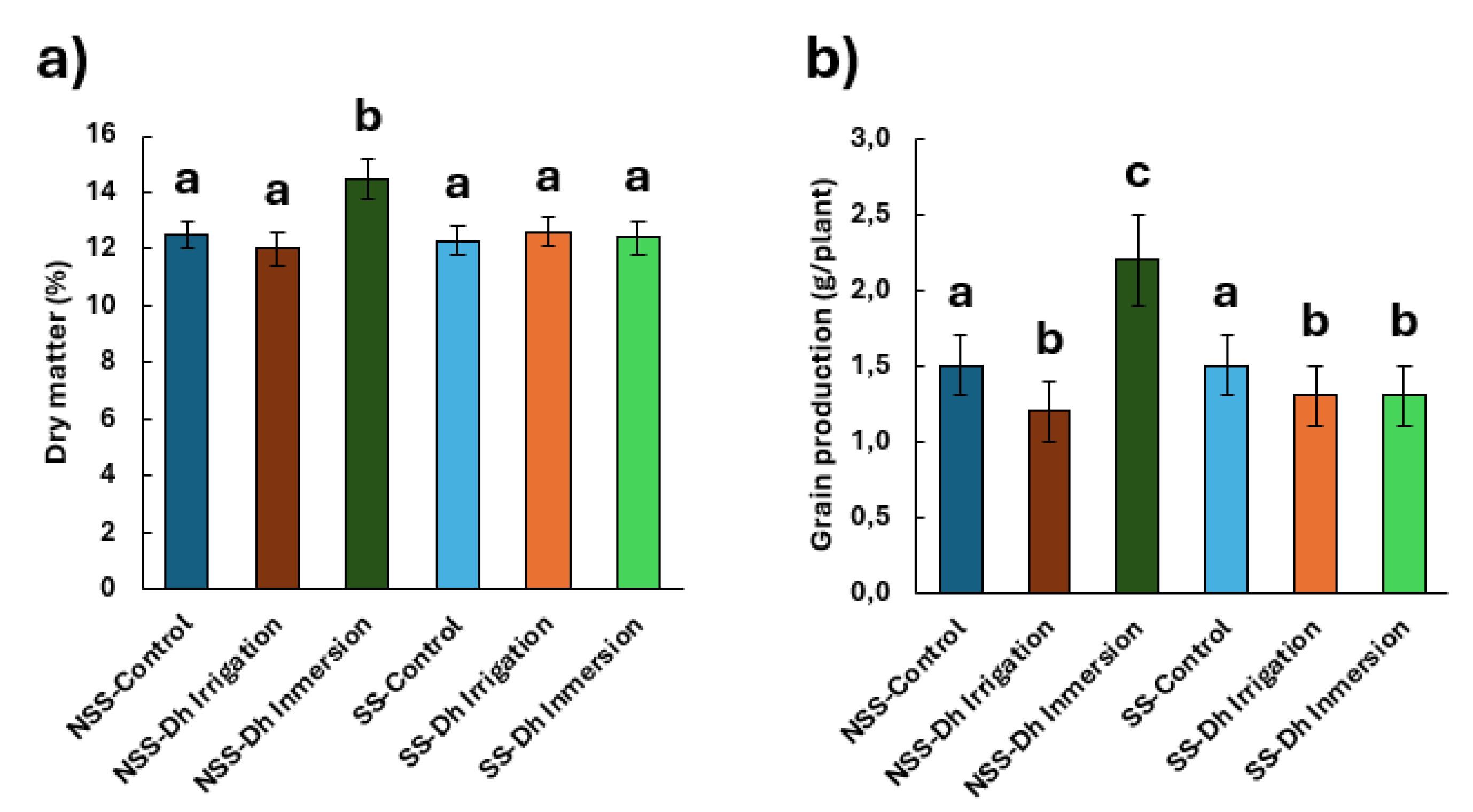
Figure 3.
Effect of D. hansenii (Dh) on plant height (cm) at 90 days after sowing and treatment application. Experiments were conducted using both sterilized (SS) and non-sterilized calcareous soil (NSS). From left to right, the treatments shown are: Control (C), D. hansenii inoculation via irrigation (Dh Irrigation), and D. hansenii inoculation via root immersion (Dh Inmersion).
Figure 3.
Effect of D. hansenii (Dh) on plant height (cm) at 90 days after sowing and treatment application. Experiments were conducted using both sterilized (SS) and non-sterilized calcareous soil (NSS). From left to right, the treatments shown are: Control (C), D. hansenii inoculation via irrigation (Dh Irrigation), and D. hansenii inoculation via root immersion (Dh Inmersion).

Figure 4.
Micronutrient content of Cu, Fe, Zn, Mn and P in the leaves of rice plants cultivated in calcareous soil and inoculated with the yeast D. hansenii (Dh). Experiments were conducted using both sterilized and non-sterilized calcareous soil. The letters on the X-axis of each graph indicate the different treatments: NSS-Control (non-sterilized soil, control), NSS-Dh Irrigation (non-sterilized soil inoculated with D. hansenii via irrigation), NSS-Dh Inmersion (non-sterilized soil inoculated with D. hansenii via root immersion), SS-Control (sterilized soil, control), SS-Dh Irrigation (sterilized soil inoculated with D. hansenii via irrigation), and SS-Dh Inmersion (sterilized soil inoculated with D. hansenii via root immersion). Different letters above the bars indicate statistically significant differences among treatments according to Tukey’s HSD test at p < 0.05. Values are presented as mean ± SE (n = 5).
Figure 4.
Micronutrient content of Cu, Fe, Zn, Mn and P in the leaves of rice plants cultivated in calcareous soil and inoculated with the yeast D. hansenii (Dh). Experiments were conducted using both sterilized and non-sterilized calcareous soil. The letters on the X-axis of each graph indicate the different treatments: NSS-Control (non-sterilized soil, control), NSS-Dh Irrigation (non-sterilized soil inoculated with D. hansenii via irrigation), NSS-Dh Inmersion (non-sterilized soil inoculated with D. hansenii via root immersion), SS-Control (sterilized soil, control), SS-Dh Irrigation (sterilized soil inoculated with D. hansenii via irrigation), and SS-Dh Inmersion (sterilized soil inoculated with D. hansenii via root immersion). Different letters above the bars indicate statistically significant differences among treatments according to Tukey’s HSD test at p < 0.05. Values are presented as mean ± SE (n = 5).
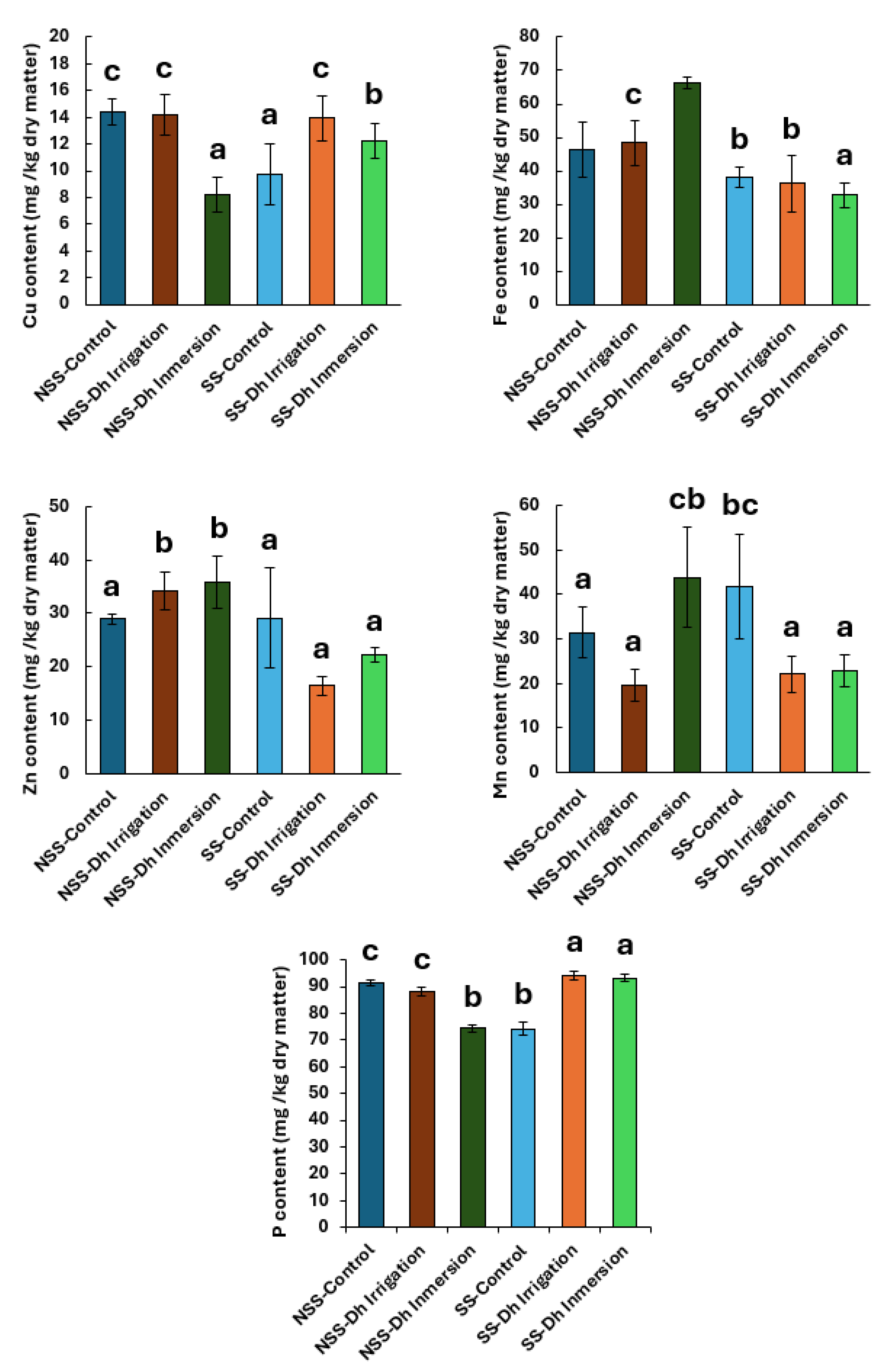
Figure 5.
Effect of the yeast D. hansenii (Dh) on acid phosphatase activity in rice plants. The assays were conducted under hydroponic conditions in a growth chamber. Treatments: +P = phosphorus-sufficient nutrient solution; +P+Dh = phosphorus-sufficient nutrient solution plus D. hansenii inoculation; –P = phosphorus-deficient nutrient solution; –P+Dh = phosphorus-deficient nutrient solution plus D. hansenii inoculation. All treatments were applied on the same day. Determinations were carried out at 7, 9, and 11 days after treatment application (data). Six plants were used for acid phosphatase activity assays; however, only two representative plants per treatment are shown to avoid overcrowding the image.
Figure 5.
Effect of the yeast D. hansenii (Dh) on acid phosphatase activity in rice plants. The assays were conducted under hydroponic conditions in a growth chamber. Treatments: +P = phosphorus-sufficient nutrient solution; +P+Dh = phosphorus-sufficient nutrient solution plus D. hansenii inoculation; –P = phosphorus-deficient nutrient solution; –P+Dh = phosphorus-deficient nutrient solution plus D. hansenii inoculation. All treatments were applied on the same day. Determinations were carried out at 7, 9, and 11 days after treatment application (data). Six plants were used for acid phosphatase activity assays; however, only two representative plants per treatment are shown to avoid overcrowding the image.

Figure 6.
Effect of the yeast D. hansenii (Dh) on the relative expression of genes associated with acid phosphatase activity for phosphorus acquisition (OsPAP9 and OsPAP3) and phosphorus transport (OsPTH1;6) in rice plant roots. Experiments were conducted under hydroponic conditions in a growth chamber. Determinations were performed at 7, 9, and 11 days after treatment application (data). Treatments: Red = phosphorus-sufficient nutrient solution, Dark Blue = phosphorus-deficient solution. Data represent mean ± SE of three independent biological replicates and two technical replicates. Asterisks (***) indicate statistically significant differences compared to the control within each time point according to Dunnett’s test (p < 0.001).
Figure 6.
Effect of the yeast D. hansenii (Dh) on the relative expression of genes associated with acid phosphatase activity for phosphorus acquisition (OsPAP9 and OsPAP3) and phosphorus transport (OsPTH1;6) in rice plant roots. Experiments were conducted under hydroponic conditions in a growth chamber. Determinations were performed at 7, 9, and 11 days after treatment application (data). Treatments: Red = phosphorus-sufficient nutrient solution, Dark Blue = phosphorus-deficient solution. Data represent mean ± SE of three independent biological replicates and two technical replicates. Asterisks (***) indicate statistically significant differences compared to the control within each time point according to Dunnett’s test (p < 0.001).

Table 1.
Physical-chemical properties and phosphorus and iron availability for the plant (average) in the calcareous soil used.
Table 1.
Physical-chemical properties and phosphorus and iron availability for the plant (average) in the calcareous soil used.
| Clay g kg−1 | Organic Carbon g kg−1 | CaCO3 g kg−1 | pH1:2.5 | EC1:5 dS m−1 | CEC cmol kg−1 | POlsen mg kg−1 | FeDTPA mg kg−1 |
|---|---|---|---|---|---|---|---|
| 370 | 9.3 | 338 | 7.9 | 1.5 | 31.3 | 13.4 | 4.3 |
CaCO3: Carbonates content. pH1:2.5: Soil pH in the extract 1:2.5 (soil: deionized water). EC1:5: Electrical conductivity in the extract 1:5 (soil: deionized water). CEC: Cation exchange capacity. POlsen: P available on soil. FeDTPA: Labile Fe in the soil.
Table 2.
Primers used in this study.
| Gene | Forward (5′-3′) | Reverse (5′-3′) |
|---|---|---|
| OsPHT1;6 | CCGCCGCCTCACAAACTGTA | GAACTGGGCGGTTTTCCTGA |
| OsPAP9 | ACCTACGTAGAGACAACATCAGGC | CATATACGTGTTGCCGGTAGTGA |
| OsPAP3 | TCATACCATGAGGAGTGAGTGATG | GTCTTCGTTTTGTGAAAATGGC |
| OsACTIN | TGCATGTAGTACAGTGC CATCCAG | AATGAGTAACCACGCTCCGTCA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



