
Submitted:
29 May 2025
Posted:
30 May 2025
You are already at the latest version
Abstract
Xylosalsola chiwensis (Popov) Akhani & Roalson is listed in the Red Data Book of Kazakhstan as a rare species with a limited distribution, occurring in small populations in Kazakhstan, Uzbekistan, and Turkmenistan. The aim of this study was to deepen the understanding of the ecological conditions of its habitats, the floristic composition of its associated plant communities, the species' morphological and anatomical characteristics, and its molecular phylogeny, as well as to identify the main threats to its survival. The ecological conditions of the Xy. chiwensis habitats include coastal sandy plains and the slopes of chinks and denudation plains with grey–brown desert soils and bozyngens on the Mangyshlak Peninsula and the Ustyurt Plateau at altitudes ranging from – 3 to 270 meters above sea level. The species is capable of surviving in arid conditions (less than 100 mm of annual precipitation) and under extreme temperatures (air temperatures exceeding 45 °C and soil surface temperatures above 65 °C). In Xy. chiwensis communities, we recorded 53 species of vascular plants. Anthropogenic factors associated with livestock grazing, industrial disturbances, and off-road vehicle traffic along an unregulated network of dirt roads have been identified as contributing to population decline and the potential extinction of the species under conditions of unsustainable land use. The morphometric traits of Xy. chiwensis could be used for taxonomic analysis and for identifying diagnostic morphological characteristics to distinguish between species of Xylosalsola. The most taxonomically valuable characteristics include the fruit diameter (with wings) and cone length, as they differ consistently between species and exhibit relatively low variability. Anatomical adaptations to arid conditions were observed, including a well-developed hypodermis, which is indicative of a water-conserving strategy. The moderate photosynthetic activity, reflected by a thinner palisade mesophyll layer, may be associated with reduced photosynthetic intensity, which is compensated for through structural mechanisms for water conservation. The flow cytometry analysis revealed a genome size of 2.483 ± 0.191 pg (2n/4x = 18) and the phylogenetic analysis confirmed the placement of Xy. chiwensis within the tribe Salsoleae of the subfamily Salsoloideae, supporting its taxonomic distinctness. To support the conservation of this rare species, measures are proposed to expand the area of the Ustyurt Nature Reserve through the establishment of cluster sites.
Keywords:
Xylosalsola chiwensis
; DNA molecular analysis
; Mangyshlak Peninsula
; Ustyurt Plateau
; ecological conditions
; floristic composition
; nature conservation
1. Introduction
The genus Salsola L. was established by Linnaeus in Species Plantarum in 1753 and originally comprised five species. The scope and classification of this heterogeneous and polymorphic genus have been revised multiple times [1]. Until recently, the genus Salsola included between 120 and 170 species, the vast majority of which are distributed in the desert regions of Eurasia and Africa [2,3,4,5,6]. In 2007, Akhani H. et al. [7], using molecular genetic methods, published a study in which the genus Salsola was split into several genera. One of these was Xylosalsola Tzvelev, originally described by N.N. Tzvelev in 1993. The type specimen, previously known as Salsola arbuscula Pall., was designated Xylosalsola arbuscula (Pall.) Tzvelev. This taxonomic revision gained wider recognition following the work of Akhani H. et al. in 2007. Later, in 2012, A.P. Sukhorukov noted that the previously monotypic genus Xylosalsola actually included several species.
The members of the genus Xylosalsola are subshrubs, shrubs, or low-growing trees with light-gray bark and a highly branched structure. Their leaves, bracts, and perianth segments are covered with short, stiff hairs. Their leaves are alternate, fleshy or filiform, glaucous, almost club-shaped, blunt-tipped, and slightly narrowed or broadened at the base. Their bracteoles are significantly shorter than the leaves and flowers, semicircular, with broad membranous margins and short pointed tips. The perianth segments are broadly lanceolate, narrowly membrane-margined, glabrous or scabrid, and the fruit bear wings below the middle with a loose cone converging above them [2,8,9].
According to Plants of the World Online [10], the genus Xylosalsola includes four species: Xy. arbuscula, Xy. chiwensis, Xy. richteri (Moq.) Akhani & Roalson, and Xy. paletzkiana (Litv.) Akhani & Roalson. Xy. paletzkiana, a species of the Southern Turanian region (Afghanistan, Tajikistan, Turkmenistan, and Uzbekistan), occurs on sandy soils and is not found in Kazakhstan. Xy. richteri has a Turanian and Southern Turanian distribution (Kazakhstan, Afghanistan, Iran, Pakistan, Tajikistan, Turkmenistan, and Uzbekistan) and is typically associated with sandy habitats, occurring in the Kyzylkum sands in Kazakhstan. Xy. arbuscula is the most widely distributed species in the genus, with an Irano-Turanian range. In addition to Central Asian countries (Kazakhstan, Kyrgyzstan, Tajikistan, Turkmenistan, Uzbekistan, Afghanistan, Iran, and Pakistan), it is also found in Russia (eastern and southern European parts), Mongolia, and China (Inner Mongolia, north-central China, and Xinjiang).
Xy. chiwensis has a Turanian type of range and is distributed in Kazakhstan and Turkmenistan [10], and has also been reported in Uzbekistan in multiple publications [11,12,13,14]. The species is listed in the Red Data Books of both Kazakhstan and Uzbekistan [11,15].
According to data from iNaturalist and the GBIF database [16,17], the species has been recorded in six localities in Kazakhstan (added by Osmonali B. during the 2024 expedition and by Islamgulova A. in 2025). In the Global Biodiversity Information Facility Secretariat checklist dataset [17], the species is reported in Uzbekistan (Fergana Valley). However, the checklist of the Flora of Protected Natural Areas in the Fergana Valley does not include Xy. chiwensis. This contradicts the distribution indicated in the Red Data Book [11], which states its occurrence in Karakalpakstan on the slopes of the Ustyurt Plateau and in the Kyzylkum sands (Sultan Uveysdag). These discrepancies are not supported by published data [18] and do not correspond to the species’ known ecological preferences, leading us to believe that they are errors. It is likely that mistakes were made in the GBIF database entry.
On the Plantarium website [19], the species was recorded in a single locality in 2024 by V. Kolbintsev in the Ustyurt Nature Reserve (Kazakhstan), in the border region between the Mangyshlak lowlands and the Ustyurt Plateau. The presence of the species in the flora of the reserve is supported by previous publications [20,21]. In 1996, the species was also recorded by I.N. Safronova at two sites in the Mangyshlak lowlands and one site in the southern part of the Ustyurt [22].
Herbarium specimens of Xy. chiwensis from the collections of the Institute of Botany and Phytointroduction (AA, Almaty, Kazakhstan), Lomonosov Moscow State University (MW; Moscow, Russia), and the Komarov Botanical Institute (LE; Saint Petersburg, Russia) were examined. Based on the studied herbarium collections, a table of specimens was compiled (Table S1). Excluding the records obtained from field research, the herbarium list includes 15 specimens. The herbarium specimens are dated from 1926 to 2015 and 2024.
All available data on the species’ distribution, including field surveys conducted in 2024 and 2025, herbarium specimens, published sources, and verified records from online databases containing georeferenced information, are presented in Figure 1.
Within Kazakhstan, Xy. chiwensis occurs in the territory of the Mangyshlak Peninsula and the Ustyurt Plateau in the Mangystau and Aktobe administrative regions. The area belongs to the North Turanian (southern part of the West North Turanian subprovince) and Southern Turanian (northern part of the West-Southern Turanian subprovince) botanical provinces of the Irano-Turanian subregion, Saharo-Gobi Desert region [23]. It includes the Mangystau, South Mangystau, North Ustyurt, and South Ustyurt floristic districts [24].
The western Caspian part of the Ustyurt Plateau lies within Kazakhstan, while its eastern (Aral region) part is located in Uzbekistan and Turkmenistan. To the north and northwest, the plateau borders the Caspian Lowland, and to the west it is adjacent to the flat Mangyshlak. The boundary of the plateau is sharply defined by high (150–200 m), deeply dissected cliffs known as "chinks" (a chink is a steep escarpment of the Earth's surface, up to 350 m high, of erosional, denudational, or tectonic origin), which have winding contours. In terms of relief, Ustyurt is classified as an arid denudational mesa plateau. The absolute elevations range from 100–120 m in the north to 150–300 m in the central and southern parts. The plateau’s relief is characterized as an undulating and gently rolling plain, featuring an alternation of low knolls, gently sloping weakly undulating surfaces, depressions, and hollows. The latter are widespread in the northern part of the plateau and are occupied by solonchaks and sands. Various forms of meso- and microreliefs are well developed on the plateau, especially in the northern and southern parts, including dry valleys, takyrs (a special type of soil consisting of flat, poorly dissected areas of clay in deserts, the formation of which is due to the periodic stagnation of surface waters in conditions of drainless surfaces composed of loams and clays with the participation of salinization and desalinization processes), and depressions, which contribute to the complexity of the soil vegetation cover. The Ustyurt Plateau is characterized by the predominance of gray–brown desert solonetz soils, takyrs, and solonchaks.
The Mangyshlak Plain encompasses the territory situated between the Ustyurt escarpments (chinks), the ridges of the mountainous Mangyshlak, and the coastal strip of the Caspian Lowland. Similar to Ustyurt, the Mangyshlak Plain is an arid denudation table plateau that has been lowered to absolute elevations of 50–150 m, representing the lower tier of the same massif. A distinctive feature of the landscape is the presence of extensive and deep depressions – such as Karagiye (–132 m below sea level) and Kaundy – on the gently undulating surface. These depressions are occupied by solonchaks (salt flats).
Cenopopulations 1–4 are distributed in regions dominated by gray–brown desert solonetzic soils and underdeveloped gray–brown desert soils. The fifth cenopopulation occurs in the Kenderli–Kayasan undulating plain region, characterized by gray–brown desert normal and solonetzic soils in the southern part of the Mangyshlak Plain. The sixth cenopopulation is distributed between the Karynzharyk depression and the western chink of Ustyurt, in the southeastern part of the hilly Mangyshlak, on bozyngens–convex or flat surfaces with highly gypsum-rich gray–brown desert soils [25].
Xy. chiwensis is a semishrub that is 30–60 cm tall, with strongly branched stems. Its annual shoots are glabrous, smooth, and highly branched. Its leaves are alternate, semi-terete, succulent, and glaucous. The bracteal leaves exceed the length of the flowers and bracteoles; the latter are semicircular, glaucous, nearly hemispherical, and narrowly membranous along the margins while the inflorescence is spike-like. The perianth segments are glabrous and bear wings at the fruiting stage, forming a loose, convergent cone above; the wings are yellowish and semi-transparent [26,27]. The species is a xerophytic succulent, flowering and fruiting from July to September. It reproduces through seeds (Figure 2).
Xy. chiwensis is listed in the Red Data Book of Kazakhstan under Category II. It is a rare species, occurring in small populations and within limited areas. Ecologically, it is associated with saltwort–desert complexes on eroded gray–brown desert soils of marl slopes, typical solonchaks, and solonetzic soils. The main threat to its survival is anthropogenic activity within its habitats, including the construction of industrial facilities and roads, as well as oil and gas extraction [15].
Studies of plant communities in the Mangystau and Aktobe regions conducted in 2024–2025 confirmed that Xy. chiwensis rarely occurs and has a low abundance.
The current state of Xy. chiwensis remains insufficiently studied. Although the species is listed in the Red Data Books of two countries (Kazakhstan and Uzbekistan), it is not included in the IUCN Red List [28] and does not have international conservation status as a rare species, highlighting the relevance and urgency of further research on Xy. chiwensis.
The purpose of this study was to investigate the current status of the rare species Xy. chiwensis. The research objectives were to (1) analyze the natural and climatic conditions of its habitats; (2) analyze the floristic composition of the plant communities; (3) conduct morphological and anatomical studies on the species to determine the phylogenetic position of Xy. chiwensis using molecular genetic analysis; (4) identify threats to the existence of the species; and (5) develop recommendations for the conservation of the species.
2. Results
2.1. Floristic Composition of Plant Communities
Vegetation surveys on the Mangyshlak Peninsula identified six coenopopulations involving Xy. chiwensis. Analysis of the species composition revealed 53 species of vascular plants. Our research was complemented by the floristic list of N. K. Rakhimova et al. [12] for two cenopopulations distributed on the Ustyurt Plateau (Uzbekistan), from which, six species were added (Table S2).
The coenoflora of plant communities with Xy. chiwensis on the Mangyshlak Peninsula consisted of 53 vascular plant species, belonging to 39 genera and 15 families. The dominant families were Amaranthaceae (17 species), Poaceae (6), and Asteraceae (5), together accounting for 53% of the coenoflora. The most representative genera were Anabasis and Artemisia, each comprising four species. The weed flora were represented by six species, comprising 11% of the total flora identified: Ceratocarpus arenarius L., Salsola tragus L., Alyssum desertorum Stapf, Descurainia sophia (L.) Webb ex Prantl, Alhagi pseudalhagi (M.Bieb.) Desv. ex Wangerin, and Ranunculus testiculatus Crantz. The contribution of weed species in the five studied cenopopulations did not exceed 2% of the total projected cover, while in one cenopopulation, it accounted for 11%, which indicates anthropogenic transformation of certain habitats. This included livestock grazing, traffic on an unregulated network of dirt roads, mole colonies, and locally observed recreation.
In the spectrum of life forms according to Serrebrakov’s classification [29], semishrubs (SS) predominate at 32% (17 species) and herbaceous annual (HA) forms at 28% (15 species) (Figure 3). According to the Raunkiaer classification [30], the most common group was chamaephytes (Ch) (17–32%). The chamaephytes (Ch) group consists of semishrubs, dwarf shrubs, and dwarf semishrubs with an annual dying-off of the generative shoots. The Ch group is the most characteristic group for the arid zone of Central Asia and is associated with the ability to survive in the dry summer season and droughts [31]. The most common species of this group, with considerable project coverage, were Artemisia arenaria DC., A. gurganica (Krasch.) Filatova, A. terrae-albae Krasch., Caroxylon gemmascens (Pall.) Tzvelev, Anabasis brachiata Fisch. & C.A.Mey. ex Kar. & Kir., and Bassia prostrata (L.) Beck. The next most species-rich group was therophytes (Th), comprising 15 species (28%), and represented primarily by ephemerals, including genera such as Eremopyrum, Alyssum, and Ranunculus, as well as annual saltworts like Climacoptera crassa (M.Bieb.) Botsch. and C. lanata (Pall.) Botsch.
The coenoflora of plant communities containing Xy. chiwensis in the southern part of the Ustyurt Plateau [12] consisted of 20 species belonging to 18 genera and 13 families. Weedy species accounted for 10% of the coenoflora.
An analysis of the species composition of Xy. chiwensis coenopopulations from the Mangyshlak Peninsula and the Ustyurt Plateau revealed a Sørensen similarity coefficient of 0.385, indicating a moderate level of similarity, with approximately 24% of the species shared between the two regions. The observed differences in species composition are attributed to climatic conditions and the geographical positions of the regions.
2.2. Characteristics of Plant Communities, Abiotic Factors of Habitats and the Inflence of Anthropogenic Factors
Two plant communities differing in ecological conditions were identified between the Karagie and Ashisor depressions (Figure 4). The floristic composition of the communities is presented in Table S1 (No. 1 and 2). Xy. chiwensis accounted for no more than 1% of these communities. The distance between the two coenopopulations was greater than 5 km.
The first community (No. 1) was composed of perennial saltwort–sagebrush vegetation, including Artemisia terrae-albae, Oreosalsola arbusculiformis (Drobow) Sennikov, and Anabasis brachiata. It developed on brown, underdeveloped soils of an undulating plain with gentle knolls at an elevation of 24 m above sea level. Bedrock outcrops covered approximately 20% of the surface. The total projective cover (TPC) was 25%. The vegetation showed a low degree of disturbance, primarily from grazing and vehicle traffic. A total of 17 vascular plant species were recorded in the community. The semishrubs (PC: 18%) included Artemisia terrae-albae, A. lercheana Weber ex Stechm., Anabasis brachiata, An. salsa (Ledeb.) Benth. ex Volkens, Nanophyton erinaceum (Pall.) Bunge, and Limonium suffruticosum suffruticosum (L.) Kuntze. The shrubs and dwarf shrubs (PC: 5%) included Oreosalsola arbusculiformis, Convolvulus fruticosus Pall., Ephedra distachya L., and Atraphaxis spinosa L. The herbal layer (PC: 2%) consisted of Poa bulbosa L., Stipa arabica Trin. & Rupr., Tulipa spp., Alyssum desertorum, Ranunculus testiculatus, and Eremopyrum bonaepartis (Spreng.) Nevski.
The second community (No. 2) was distributed across a sandy massif of the coastal plain with sor-type depressions, located at an elevation of 3 m below sea level. It was represented by Alhagi pseudalhagi–sagebrush (Artemisia arenaria) vegetation that developed on stabilized sandy flatlands. The TPC was 35%. A total of 12 vascular plant species were recorded, including the semishrubs (PC: 32%) Artemisia arenaria, A. lercheana, Alhagi pseudalhagi, and Limonium suffruticosum, and the shrubs and dwarf shrubs (PC: 1–2%) Ephedra distachya and Atraphaxis replicata Lam. The herbal layer (PC: 1%) contained Stipa arabica, Salsola tragus, Descurainia sophia, Echinops albicaulis Kar. & Kir., and Zygophyllum turcomanicum Fisch. ex Boiss. The vegetation cover was in a transitional stage between slight and moderate disturbance due to grazing and anthropogenic impacts (power lines, pipelines, and a railway).
The third and fourth plant communities were located approximately 1 km apart near Zhilandy Cape (Figure 5). The third community (No. 3), occupying the western slope of the chink escarpment, was represented by sparse Bassia prostrata–sagebrush (Artemisia gurganica) vegetation with a total projective cover (TPC) of up to 20%. The coenoflora comprised 15 vascular plant species, including the semishrubs Artemisia gurganica, Bassia prostrata, Limonium suffruticosum, Caroxylon orientale (S.G.Gmel.) Tzvelev, Nanophyton erinaceum, and Anabasis truncata (Schrenk) Bunge, with Xy. chiwensis occurring sporadically; these species dominated the projective cover (PC: 18%). The shrubs and dwarf shrubs (PC: 1%) were Atraphaxis spinosa and Oreosalsola arbusculiformis. The herbal layer (PC: 1%) contained Agropyron fragile (Roth) P.Candargy L., Poa bulbosa, Climacoptera crassa, and Eremopyrum distans (K.Koch) Nevski. The slopes where this vegetation occurred were relatively steep, ranging from 20° to 40°, while the negatively inclined or nearly vertical slopes (80–90°) were almost devoid of plant cover. The soils were eroded brown soils. The plant community was mildly disturbed, with evidence of grazing and dirt roads descending the gentler slopes toward the Caspian Sea.
The fourth community (No. 4) was dominated by sagebrush (Artemisia gurganica) vegetation at a TPC of 25%. It was located on a denudation plain of the Mangyshlak region and had developed on brown underdeveloped soils. A total of 11 vascular plant species were recorded. The semishrub layer (PC: 23%) consisted of Artemisia gurganica, A. terrae-albae, Anabasis aphylla L., Nanophyton erinaceum (Pall.) Bunge, and Xy. chiwensis (1%). The only shrub or dwarf shrub (PC: 1%) was Atraphaxis replicata. The herbal layer (PC: 1%) contained Ceratocarpus arenarius, Eremopyrum orientale (L.) Jaub. & Spach, Alyssum desertorum, Girgensohnia oppositiflora (Pall.) Fenzl, and Onosma staminea Ledeb. This community was moderately disturbed due to grazing and off-road vehicle traffic. A significant proportion of the sagebrush appeared dry, likely as a consequence of the drought conditions experienced in 2021.
The fifth community (No. 5) was located on the Kendirli–Kayasan Plateau, 11 km from the Kendirli Bay and over 110 km from the fourth Xy. chiwensis population (Figure 6). There was a community dominated by Caroxylon gemmascens on the gray–brown desert soils on the undulating plain. Eremopyrum orientale played a significant role, accounting for up to 5% of the total projective cover (TPC). The TPC was 35%, with 16 species recorded. The semishrubs dominated (PC: 25%) and included the following species: Caroxylon gemmascens, C. orientale, Artemisia terrae-albae, Anabasis salsa, Nanophyton erinaceum, Limonium suffruticosum, Anabasis brachiata, A. truncata, and Xy. chiwensis (less than 1%). The herbal layer (PC: 5%) consisted of Eremopyrum orientale, Climacoptera lanata, and Halothamnus subaphyllus (C.A.Mey.) Botsch. The shrubs and dwarf shrubs (PC: less than 1%) included Atraphaxis spinosa, Oreosalsola arbusculiformis, Convolvulus fruticosus, and Ephedra distachya. The vegetation cover was slightly disturbed, with evidence of grazing, vehicle tracks along dirt roads, and burrowing animal colonies.
The sixth community (No. 6) was found approximately 5 km from the boundaries of two protected areas (the Ustyurt Nature Reserve and Kendirli–Kayasan State Nature Reserve Zone) and over 125 km southeast of the fifth Xy. chiwensis population (Figure 6). Along the runoff depressions on the elongated upland bozyngens, the vegetation was represented by dwarf semishrub groupings, with a total projective cover (TPC) of 10–15%. Twenty-four species were identified, with the semishrub layer (PC: 8%) consisting of Artemisia gurganica, Caroxylon gemmascens, and Xy. chiwensis (1–2%). The shrubs and dwarf shrubs (PC: 2%) included Oreosalsola arbusculiformis, Convolvulus fruticosus, and Ephedra strobilacea Bunge. The herbal layer (PC: 1–3%) consisted of Climacoptera lanata, Arnebia decumbens (Vent.) Coss. & Kralik, and Onosma staminea, among others, including Euphorbia sclerocyathium Korovin & Popov—a species listed in the Red Book of Kazakhstan [15].
The community was slightly disturbed, with the presence of field roads. The coenopopulation of Xy. chiwensis, occurred on soils with a high gypsum content, indicating its broad ecological range.
2.3. Climate
The climatic conditions were determined based on climatic normals, with the standard period for calculating these normals being 30 years (1991–2020). The data from four meteorological stations (Akkuduk, Aktau, Beineu, and Sam) located within the range of Xy. chiwensis in Kazakhstan (Figure 1) were provided by Kazhydromet [32].
The average annual precipitation norm varied from 171 to 121 mm per year. The data from the past four years (no data available for the Akkuduk station in the registry) were variable (Figure 7), with the precipitation in a dry year (2021) only reaching 32% of the norm, while in a wet year, it exceeded the norm by 37%. The average annual air temperature norm ranged from +10.5 to +13.2 °C. Over the last four years, there has been a trend of an increase in temperature of approximately 1.5 °C on average per year, with minimum and maximum deviations of 1.2 °C and 2.2 °C, respectively.
The absolute maximum and minimum air temperatures (Figure 8), recorded from 2021 to 2024, show a maximum of 45.2 °C (WS 2022, Beineu station) and a minimum of –26.4 °C (WS 2021, Sam station). The absolute maximum and minimum soil surface temperatures reached 68 °C (WS 2021, Sam station) and –24 °C (WS 2021, Sam station), respectively.
The average monthly temperatures throughout the year, according to the climate normals, ranged from –7.4 to +30.1 °C. It ranged from –7.4 to +1.3 °C in winter, from +2.1 to +21.2 °C in spring, from +23.5 to +30.1 °C in summer, and from +20.8 to +0.8 °C in autumn (Table S3). The average monthly maximum air temperature varied from –3.5 to +4.9 °C in winter, from +7.9 to +28.6 °C in spring, from +29.3 to +37.4 °C in summer, and from +5.8 to +28.6 °C in autumn (Table S4).
The average monthly minimum air temperature ranged from –1.5 to –10.9 °C in winter, from +2.5 to +14.2 °C in spring, from +17.9 to +22.2 °C in summer, and from –3.2 to +15.5 °C in autumn (Table S5).
2.4. Flow Cytometry and Morphological-Anatomical Research
Materials for the flow cytometry, molecular genetic, morphological, and anatomical studies were collected from two populations (Figure 4 and Figure 5). Two herbarium specimens were prepared and deposited in the herbarium collection of the Institute of Botany and Phytointroduction (AA). The specimen numbers are provided in Table S1 (Figure 9).
2.4.1. Fruit Morphology
The analysis of the fruits, as the primary morphological characteristic for genus and species identification, was conducted on two species: Xy. chiwensis (the study species) and Xy. arbuscula (selected as a comparative specimen) (Figure 10).
The morphological data were compiled into a table (Table 1). The comparative analysis revealed that the mean diameter of the fruit (with wings) in Xy. chiwensis is 5.30 mm, whereas in Xy. arbuscula, it is significantly larger at 8.53 mm. The diameter of the fruit (without wings) is slightly greater in Xy. chiwensis (2.81 mm) compared to that of Xy. arbuscula (2.57 mm). The fruits of Xy. arbuscula have a longer cone (1.90 mm) compared to Xy. chiwensis (1.09 mm) and the bract leaf length in Xy. arbuscula (8.91 mm) is longer than that of Xy. chiwensis (6.07 mm). The analysis data indicated clear morphological differences between the two species, which can be used for their identification and classification.
2.4.2. Anatomical Analysis
An anatomical analysis was conducted on the leaves of Xy. chiwensis and Xy. arbuscula, as leaf structure remains the most relevant feature in current anatomical studies of the Amaranthaceae family. Both species exhibit the most common leaf anatomical type, classified as the Salsoloid type [33,34,35]. The leaves possess a single-layered palisade parenchyma, and Kranz cells are arranged circumferentially around the leaf. Peripheral vascular bundles are adjacent to the Kranz cells, while the main vascular bundle is located centrally among the water-storage cells. The hypodermis may be present or absent. It is noteworthy that the anatomical section of Xy. chiwensis was prepared from herbarium material collected during the expedition (Figure 11).
The study of the anatomical characteristics of the leaves revealed differences between the two species, reflecting their adaptations to distinct ecological conditions. The numerical data are presented in Table 2.
Xy. arbuscula exhibits a greater thickness of the epidermis, palisade mesophyll, and Kranz cells, which may indicate more effective protection against excessive water loss and higher photosynthetic activity. In contrast, Xy. chiwensis possesses a more developed hypodermis, likely associated with mechanisms for moisture retention.
The enhanced structural protection observed in Xy. arbuscula may be explained by its more arid habitat, whereas Xy. chiwensis employs a different adaptive strategy, balancing photosynthetic activity and water regulation.
These anatomical features not only confirm the ecological adaptability of these species but also serve as additional criteria for their taxonomic differentiation.
2.4.3. Flow Cytometry
The genome size analysis of the two species using flow cytometry revealed that the genome size of Xy. chiwensis was 2.483 ± 0.191 pg, while Xy. arbuscula exhibited two genome size variants: 3.250 pg (noted in one sample) and 6.723 ± 0.582 pg (Table 3).
2.4.5. Molecular genetics
Nuclear (nrITS) and chloroplast (rps16 intron) fragment sequencing was performed on the two species. The ITS sequence phylogenetic tree was constructed using the results from the studied samples, supplemented with sequences from the NCBI database. To compare and confirm the precise phylogenetic placement of Xylosalsola species, available ITS sequences from the subfamily Salsoloideae, particularly from the tribe Salsoleae, were obtained from the NCBI database (Figure 12). Species from the tribe Caroxyloneae, the closest relative to the studied genus—especially Xy. chiwensis—were used as the outgroup. Our samples are highlighted in bold font in the tree. The color coding indicates the genetic affiliation of the Xylosalsola species (Figure 13).
Both the nrITS and rps16 intron fragments of the genus Xylosalsola are well resolved in the phylogenetic tree, with clear distinctions between species. It should be noted that there are insufficient chloroplast fragment (rps16 intron) data available in public databases for the studied and closely related species to construct a more detailed phylogenetic tree.
3. Discussion
3.1. Ecological Conditions
Xy. chiwensis inhabits desert ecosystems characterized by extreme climatic conditions, including low precipitation and significant fluctuations in air and soil temperatures. It grows on brown eroded and underdeveloped soils, sometimes with a high gypsum content. In Kazakhstan, the species is distributed across coastal sandy plains, steep rocky slopes, denudation plains, and elongated uplands of the bozyngens of the Mangyshlak Peninsula and the Ustyurt Plateau. The vegetation cover in the studied areas varied from sparse formations with 10–15% projected cover to denser communities with up to 35% cover. The anthropogenic impacts were unevenly distributed. In the northwestern part of the study area, moderate to severe disturbances were recorded due to livestock grazing, vehicle traffic, and industrial infrastructure development. In the southern regions of the Mangyshlak Peninsula, the impact on vegetation was observed at low to moderate levels.
3.2. Floristic Diversity
The floristic composition of the plant communities containing Xy. chiwensis showed regional differences between the Mangyshlak Peninsula and the Ustyurt Plateau. In the studies in Mangyshlak, 53 vascular plant species belonging to 39 genera and 15 families were recorded, while in the southern part of Ustyurt, 20 species from 18 genera and 13 families were documented. The Sørensen’s similarity coefficient (0.38) indicates a moderate degree of floristic similarity, with each region characterized by specific floral elements.
The overall list of vascular plants comprises 59 species from 18 families, dominated by Amaranthaceae, Poaceae, and Asteraceae, forming a typical floristic spectrum of Central Asian desert ecosystems [31,36]. The life form analysis showed a predominance of semishrubs (30%), annuals (27%), and perennial herbaceous plants (24%), reflecting these species’ high adaptability to uneven water availability and arid conditions. Among the semishrubs, species such as Artemisia arenaria, A. gurganica, Anabasis brachiata, and Bassia prostrata stood out. The annual flora included Eremopyrum spp., Alyssum desertorum, Climacoptera crassa, and other species that are typical of disturbed habitats.
The proportion of ruderal species (10–11%) indicates anthropogenic impacts, including livestock grazing, expansion of dirt roads, and local technogenic and recreational pressures, leading to soil vegetation cover degradation and habitat deterioration. Anthropogenic activities in the Ustyurt and Mangyshlak regions contribute to the expansion of degraded habitats colonized by secondary vegetation.
3.3. Flow Cytometry and Morphological-Anatomical Research
3.3.1. Fruit Morphology
A comparative analysis of the morphometric parameters of the fruits from two species revealed pronounced differences between Xy. chiwensis and Xy. arbuscula, which can serve as diagnostic traits to distinguish between them.
The mean fruit diameter (with wings) in Xy. chiwensis was 5.30 mm, with a narrow variation range (5.19–5.37 mm) and a low standard deviation (0.06 mm), indicating high uniformity in this parameter. In contrast, Xy. arbuscula exhibited a larger fruit diameter averaging 8.53 mm, with a wide range (7.85–9.53 mm) and a high standard deviation (0.53 mm), reflecting greater fruit variability.
The fruit diameter (without wings) in Xy. chiwensis was larger than that of Xy. arbuscula (2.81 mm vs. 2.57 mm). The fruits of Xy. arbuscula showed less size variation, which may indicate greater stability of the fruit shape.
The cone length of Xy. arbuscula was almost twice that of Xy. chiwensis (1.90 mm vs. 1.09 mm), with higher variability in Xy. arbuscula. This trait is an important morphological distinction between the two species.
The length of the bracts in Xy. arbuscula was longer that of Xy. chiwensis (8.91 mm vs. 6.07 mm). However, the high degree of variation in this parameter (broad range and standard deviation of 3.59 mm) reduces its diagnostic reliability.
The key morphological differences between the two species are the fruit diameter (with wings) and cone length, which show distinct differences alongside relatively low variability. The bract length is characterized by high variability, making it a less reliable trait for taxonomic identification.
The observed differences support the morphological differentiation of Xy. chiwensis and Xy. arbuscula, which is consistent with their taxonomic status. These findings have practical value for species identification and can be used to refine the systematics of the genus Xylosalsola.
3.3.2. Anatomy
A distinctive feature of the anatomical structure of Chenopodiaceae species is the presence of Kranz cells in their leaves.
In 2003, G. Kadereit et al. analyzed the molecular structure of the leaves of Amaranthaceae and Chenopodiaceae and identified seven groups comprising 17 types of mesophyll [37]. The mesophyll type refers to the arrangement of assimilative, conductive, spongy, or water-storage tissues relative to the cross-section of the studied plant organ.
- Non-Kranz types include axilloidoid, corispermoid, austrobasidioid (similar to corispermoid but containing water-storage tissue), neokochioid (with peripheral vascular bundles), and sympegmoid (similar to neokochioid but with peripheral channels separated from chlorenchyma).
- Kranz types, such as atriplicoid, kochioid, salsoloid, and others, differ in the arrangement of Kranz cells relative to the vascular bundles [34].
There are two variants of Kranz cell arrangement: around each individual vein and around all veins of the leaf in a double concentric layer. An increase in the number of species possessing Kranz cells during ecological monitoring could be an indicator of climate aridification [33,34].
The Chenopodiaceae family has the highest number of species using C4 photosynthesis [38,39], as well as the greatest diversity of C4 leaf anatomy, including eight major structural types [40,41] and up to 16 forms [37].
C4 photosynthesis studies have been conducted on species with the Kranz anatomy characterized by two layers of chlorenchyma surrounding each vein [42]. Among C4 species, nine types of Kranz anatomy with two concentric layers of chlorenchyma surrounding all veins have been described [40,42]. Among these, the Salsoloid-type anatomy is the most widespread, indicating that C4 photosynthesis is commonly used by species in the subfamily Salsoloideae [34,43]. Both studied species exhibit the common Salsoloid leaf anatomical type.
The anatomical structure of Xy. chiwensis (i.e., its well-developed hypodermis and stable epidermal structure) reflect its adaptations to arid habitat conditions.
The epidermis of Xy. chiwensis is comparatively thinner (20.09 μm) than that of Xy. arbuscula (35.03 μm); however, its low variability (SD = 2.24 μm) indicates structural stability, which may be important for reducing water loss and protecting internal tissues.
One key distinction of Xy. chiwensis is a more developed hypodermis (20.96 μm), indicating a high capacity for water retention. In the marl slopes of inselbergs, solonchak depressions, and brown and gray–brown desert soils where this species occurs, this feature may play an essential role in survival under limited water availability conditions.
The palisade mesophyll in Xy. chiwensis is somewhat thinner (29.59 μm) compared to Xy. arbuscula (33.67 μm), possibly indicating that the less intensive photosynthesis is compensated for using water-conserving strategies.
The thickness of Kranz cells in Xy. chiwensis (17.44 μm) was lower than that of Xy. arbuscula, which may point to specific carbon fixation mechanisms and features of photosynthetic activity.
Ecological Interpretation of Anatomical Features in Xy. chiwensis
- Water-conserving strategy
The well-developed hypodermis in Xy. chiwensis effectively accumulates and retains moisture, which is crucial under conditions of low and irregular atmospheric humidity.
A thin but structurally stable epidermis helps protect tissues from evaporation and mechanical damage.
- 2.
- Habitat association
Xy. chiwensis can grow under various ecological conditions, and its anatomical traits indicate high tolerance to water deficiency and soil salinity.
- 3.
- Photosynthetic activity
A less developed palisade mesophyll suggests moderate photosynthetic activity, likely compensated for through efficient water usage.
The structure of Kranz cells in Xy. chiwensis implies a specialized photosynthetic strategy, allowing for adaptation to harsh drought and high-temperature conditions.
3.3.3. Flow Cytometry
The flow cytometry analysis showed that the genome size of Xy. chiwensis is 2.483 ± 0.191 pg. For Xy. arbuscula, two values were recorded: 3.250 pg (based on a single sample) and 6.723 ± 0.582 pg.
The difference in genome sizes observed in Xy. arbuscula indicates the presence of polyploidy, which is characteristic of this genus and family, which was confirmed by recent studies [44,45,46]. A review of the literature and genomic databases did not reveal any published data on the genome size of Xy. chiwensis, suggesting that this is the first such analysis for this species.
According to the Chromosome Counts Database (CCDB), the chromosome numbers of closely related species in the genus Xylosalsola are as follows: 2n = 18 for Xy. arbuscula [47,48], 2n = 18 for Xy. paletzkiana, and 2n = 54 for Xy. richteri [49], with a basic chromosome number of n = 9. Diploid forms have been confirmed for Xy. arbuscula and Xy. paletzkiana, while some hexaploid specimens of Xy. richteri have been found. The flow cytometry results for Xy. chiwensis presumably indicate its ploidy level.
Assuming that Xy. arbuscula with a genome size of 3.250 pg is diploid, and the value of 6.723 ± 0.582 pg corresponds to a possible tetraploid specimen (although this has not yet been confirmed karyological studies), the genome size of Xy. chiwensis (2.483 ± 0.191 pg) corresponds to a diploidy.
To further validate the accuracy of these data, information from related species of other genera was considered. According to the Plant DNA C-values Database, Salsola kali L. has 2C = 1.22 pg (2n = 18) [50], Salsola soda L. (Soda inermis Fourr.) has 2C = 2.62 ± 0.04 pg (2n = 18), and Salicornia europaea L. has 2C = 2.75 ± 0.03 pg (2n = 18) [51].
Recent studies not included in the database provided the following values:
- Salsola tragus: – 2C = 4.48–4.57 pg (2n = 36);
- Krascheninnikovia ceratoides (L.) Gueldenst.: – 2C = 2.26–2.71 pg (2n = 18) [44];
- Kalidium foliatum (Pall.) Moq.: – 2C = 2.259 ± 0.023 pg (2n = 18);
- K. caspicum (L.) Ung.-Sternb.: – 2C = 2.981 ± 0.149 pg (2n = 18) and 2C = 5.993 ± 0.139 pg (2n = 36) [52];
- Bassia prostrata s.l.: – 2C = 2.66 ± 0.12 pg (2n = 18) and 2C = 5.01 ± 0.04 pg (2n = 36) [46];
- Krascheninnikovia ceratoides s.l.: – 2C = 2.94 ± 0.15 pg (2n = 18) and 2C = 5.83 ± 0.07 pg (2n = 36) [45].
Comparing these results with previously published data, Xy. chiwensis may be a diploid species with a genome size of 2.483 ± 0.191 pg (2n/4x = 18). Further karyological studies are needed to confirm this assumption.
This study focused primarily on Xy. chiwensis, while the results for Xy. arbuscula are presented as supplementary data.
3.3.4. Phylogeny
The obtained results confirmed the reliable identification of Xy. chiwensis (Figure 12 and Figure 13). The phylogenetic analysis showed that this species belongs to the tribe Salsoleae within the subfamily Salsoloideae, and representatives of the genus Xylosalsola form a clade distinct from closely related species of the genus Salsola (Figure 12 and Figure 13).
When considering the phylogenetic tree (Figure 12), it is important to note that the number of sequences available for Xylosalsola species in the NCBI database is limited. For Xy. chiwensis, only three sequences are available besides our own samples. For tree construction, only the MT393880 sequence [53] was used; the other sequences (AF318632 and AF318642) [54] were too short (510 bp) and therefore excluded from the analysis.
3.4. Threats to Existence and Ways to Conservation
Xy. chiwensis is a rare species with a narrow distribution range in Kazakhstan, Uzbekistan, and Turkmenistan. It is listed in the Red Books of Kazakhstan and Uzbekistan. However, the latest edition of the Red Book of Turkmenistan excluded this species [55,56]. Reliable up-to-date information on the distribution of Xy. chiwensis in Turkmenistan is lacking.
Populations of Xy. chiwensis continue to face significant pressure from anthropogenic factors and climate change, which could result in a population decline and habitat fragmentation. Therefore, the development and implementation of comprehensive conservation measures aimed at preserving the species in its natural habitats is essential.
The main threats to the species’ survival are the following:
- Habitat fragmentation and degradation: The expansion of infrastructure (roads, pipelines, and industrial facilities) leads to habitat destruction and fragmentation of populations especially in the northwestern part of the study region. The construction of new unpaved roads intensifies erosion processes, worsening the conditions for natural population recovery.
- Livestock grazing: This is also commonly occurs in the northwestern part. High grazing pressure causes mechanical damage to plants, reducing their reproductive capacity and altering the species composition of plant communities, thereby limiting the potential for natural regeneration.
- Climate change: Increasing aridization, reduced precipitation, and rising average annual temperatures may negatively impact population stability, especially at early ontogenetic stages [12].
- Industrial development: In the northwestern part, oil and gas extraction and mining activities alter hydrological regimes, contaminate soils, and increase dust loads, negatively affecting the physiological condition of plants.
The following conservation and protection measures are recommended:
- Expansion of protected areas: Inclusion of new Xy. chiwensis habitats into the existing Ustyurt Nature Reserve and Kendirli–Kayasan State Nature Reserve zones and establishment of new cluster sites based on research data. For example, the proposal in [22] to designate a site near the tri-border area (Chink Kaplankyr) is considered unjustified because the chink territory is behind a border fence and not exposed to anthropogenic threats.
- Limiting anthropogenic pressures: Regulation of livestock grazing in Xy. chiwensis habitats, especially in heavily degraded sites, exerting better control over transport route construction, and develop alternative routes to minimize the impacts on natural ecosystems.
- Scientific research: Regular monitoring of populations with a focus on the ontogenetic structure and reproductive status.
- Rehabilitation activities: Cultivation of Xy. chiwensis in the Mangyshlak Experimental Botanical Garden with subsequent reintroduction into natural habitats.
- Awareness and outreach: Informing local communities about the importance of conserving Xy. chiwensis and natural ecosystems as a whole.
4. Materials and Methods
4.1. Data Collection
This study was conducted in 2024–2025 on the Mangyshlak Peninsula and the Ustyurt Plateau. Six populations of Xy. chiwensis were discovered on the Mangyshlak Peninsula, whereas the species was not found within the Kazakhstani part of the Ustyurt Plateau.
The vegetation was studied using the traditional methods of geobotanic field research [57]. The studied sites were 10 × 10 sq m. The geolocation was registered using a Global Positioning System (GPS) device, and detailed geobotanical descriptions were compiled. The coordinates, landscapes, soils, water regime, total projective coverage, layers, and degree of transformation were recorded for each plant community. The full floristic composition was given, and the phenological phases, vigor (according to a five-point scale), spread (using Bykov’s scale), morphometric parameters (height, habitus), and abundance (using the Drude scale) [58] (in percentages) of the plant species were determined. Descriptions of the vegetation were recorded using the vegetation description forms. The plant species were identified based on the identification keys of the nine-volume Flora of Kazakhstan [13,59] and the two-volume Illustrated Guide for Identification of the Plants of Kazakhstan [60]. The names of the plant species, genera, and families were recorded in accordance with the online resource Plants of the World Online [10]. To determine the similarity of the species composition of Xy. chiwensis populations on the Mangyshlak Peninsula and on the Ustyurt Plateau, the Sørensen coefficient was used [27,61].
We reviewed herbarium specimens of the species Xy. chiwensis from the Institute of Botany and Phytointroduction (AA, Almaty, Kazakhstan), Lomonosov Moscow State University (MW; Moscow, Russia), and the Komarov Botanical Institute (LE; Saint Petersburg, Russia). Morphological images of the herbarium specimens were taken using a Levenhuk DTX RC4 Remote Control Microscope (China).
4.2. Anatomic Method
The materials used for the anatomical sections were fixed in 70% ethanol. The samples were frozen and embedded in histological paraffin in special molds measuring 15 × 15 mm. Cross-sections were prepared using a MEDITE M530 rotary semi-automatic microtome. The section thickness was 40 µm. The sections were examined using a Levenhuk Zoom&Joy microscope (China) and images were captured using a Levenhuk D740T 5.1 camera with the LevenhukLite software, which was also used for biometric data collection. The mean values and standard errors for the biometric parameters were calculated using the data analysis functions in Microsoft Excel. The anatomical structure descriptions were based on recent literature on closely related species [33,34,62,63,64,65,66].
4.3. Flow Cytometry
The DNA content was determined using flow cytometry techniques with propidium iodide (PI) staining. Leaves dried with silica gel were used as samples. The samples were chopped with a sharp razor blade in LB01 buffer containing PI (50 µg/ml). RNase (50 µg/ml) [67] supplemented with 12 mM sodium thiosulfate and 1% polyvinylpyrrolidone [68] was added. The nuclear suspension was filtered through a nylon filter with a pore size 30 μm. The analyses were performed using a Cytoflex (Beckman Coulter. Inc.) cytometer. The histograms were visualized and processed using the CytExpert software (Beckman Coulter. Inc.). The descriptive statistics were calculated using XLStat (Addinsoft). The internal standards used were Pisum sativum ‘Ctirad’ (2C = 9.09 pg) [69], Petroselinum crispum ‘Moss Curled2’ (2C = 4.5 pg), and Solanum pseudocapsicum (2C = 2.835 pg) [68].
4.4. Molecular Genetics Methods
Amplification and sequencing. DNA extraction and amplification were performed using standard protocols at the Laboratory of Bioengineering, South Siberian Botanical Garden, Altai State University [70]. Total genomic DNA was isolated from the herbarium specimens using the commercial DiamondDNA extraction kit according to the manufacturer's instructions (Scientific Production Firm “Altaybiotech” LLC, Barnaul, Russia).
Two DNA regions were sequenced: the nuclear ITS1–5.8S–ITS2 region and the chloroplast rps16 intron. The ITS region was amplified using the primers ITS-for (5′-CGTAACAAGGTTTCCGTAG-3′) and ITS-rew (5′-GGAATCCTTGTAAGTTTCTTT-3′) [71]. Amplification was carried out using a MyCycler thermal cycler under the following conditions: initial denaturation at 95 °C for 60 s and then 33 cycles of primer annealing at 57 °C for 30 s and extension at 72 °C for 30 s.
The chloroplast rps16 intron was amplified using the primers rpsF (5′-GTGGTAGAAAGCAACGTGCGACTT-3′) and rpsR2 (5′-TCGGGATCGAACATCAATTGCAAC-3′) [72].
The PCR amplification was performed using BioMaster HS-Taq PCR mix (Biolabmix, Russia) in a reaction volume of 15 μL, with a final primer concentration of 400 nM. The primers were synthesized by Evrogen LLC (Russia).
Sequencing was conducted using an ABI Prism 3130xl Genetic Analyzer (Applied Biosystems, USA). The sequences from all individuals were manually edited in Chromas Lite 2.1 (Technelysium Pty Ltd., South Brisbane, QLD, Australia) and aligned using ClustalX [73]; the alignment was manually corrected using MEGA11 [74].
For the phylogenetic analyses, both datasets (nrITS and the cpDNA markers) were analyzed separately through Fitch parsimony using the heuristic search option in PAUP version 4.0 b10 [75] with MULTREES, TBR branch swapping, and 100 replicates of random addition sequence. Gaps were treated as missing data. The consistency index (CI) was calculated to estimate the amount of homoplasy in the character set [76]. The most parsimonious trees returned by the analysis were summarized in one consensus tree using the strict consensus method. Bootstrap support (BS) was performed using 1000 pseudoreplicates to assess the support for the clades [77]. Bayesian phylogenetic analyses were also performed using MrBayes 3.1.23 [78]. The sequence evolution model was chosen by following the Akaike information criterion (AIC) obtained from jModelTest2. Two independent analyses with four Markov chains were run for 10 million generations, and sampling trees were generated every 100 generations. The first 25% of the trees were discarded as burn-in. The remaining 150.000 trees were combined into a single dataset, and a majority-rule consensus tree was obtained, along with posterior probabilities (PPs).
Species from Tribus Caroxyloneae were selected as an outgroup: Caroxylon orientale and Caroxylon dzhungaricum (Iljin) Akhani & Roalson [79].
5. Conclusions
This study provides a comprehensive analysis of Xy. chiwensis, a rare and vulnerable species endemic to the arid ecosystems of Central Asia. It integrates its ecological characteristics, floristic composition, morphological and anatomical features, genome size data, and phylogenetic placement to contribute to our understanding of the species’ adaptive mechanisms and taxonomic status.
Our findings confirmed the following:
- There is moderate floristic similarity between the populations on the Mangyshlak Peninsula and the Ustyurt Plateau (Sørensen index = 0.385), reflecting the influence of regional environmental factors. These results are consistent with previously reported floristic differences in the area.
- There are pronounced morphological distinctions between Xy. chiwensis and the closely related species Xy. arbuscula, particularly in fruit size and cone length, which serve as important taxonomic characteristics.
- Xy. chiwensis shows anatomical adaptations to arid conditions, including a well-developed hypodermis, thin epidermis, and characteristic mesophyll structure, which align with general patterns of xerophytic adaptation among desert plants.
- The genome size of Xy. chiwensis (2C = 2.483 pg) is reported here for the first time and the species’ ploidy level was confirmed, which aligns with the polyploid tendencies observed within the genus Xylosalsola and the family Amaranthaceae.
- The phylogenetic placement of Xy. chiwensis was confirmed to be within the tribe Salsoleae, distinguishing it as a separate lineage within Xylosalsola based on its nrITS and rps16 intron sequences.
A review of the literature and our field observations identified the main threats to Xy. chiwensis: habitat fragmentation and degradation (particularly in the northwestern part of the study region), livestock grazing, climate change, and industrial development. In line with established approaches for the conservation of rare species, we propose the following measures: expansion of protected areas based on scientific justification of key habitat importance, regulation of anthropogenic pressures, population monitoring, rehabilitation activities, and raising public awareness regarding the species’ conservation value.
The results significantly enhance our knowledge of Xy. chiwensis and provide a scientific foundation for the development of effective conservation strategies under global change. Future research directions include assessing its genetic diversity and adaptation mechanisms to extreme arid environments, as well as implementing proposed conservation measures, with a particular emphasis on detailed studies in Turkmenistan, where data on the species' distribution and population status remain insufficient.
Supplementary Materials
The following supporting information can be downloaded at: Table S1: Herbarium data; Table S2: List of vascular plants with Xylosalsola chiwensis described in Kazakhstan and Uzbekistan; Table S3: Average monthly and annual air temperature, °C; Table S4: Average monthly maximum air temperature, °C; Table S5: Average monthly minimum air temperature, °C.
Author Contributions
Conceptualization, A.I. and B.O.; Formal analysis, A.I., B.O., S.M. and A.K.; Investigation, A.I., B.O., S.M., A.K. and Az.Im.; Project administration, A.I.; Resources, A.I.; Supervision, A.I. and B.O.; Writing—original draft, A.I., B.O., S.M. and A.K.; Writing—review and editing, A.I., V.P. and Az.Im. All authors have read and agreed to the published version of the manuscript.
Funding
This research has been funded by the Committee of Science of the Ministry of Science and Higher Education of the Republic of Kazakhstan. Grant No. BR21882122 «Sustainable Development of Natural-Industrial and Socio-Economic Systems of the West Kazakhstan Region in the Context of Green Growth: A Comprehensive Analysis, Concept, Forecast Estimates and Scenarios».
Data Availability Statement
The datasets generated during the study are available from the corre sponding authors upon reasonable request.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Sukhorukov, A.P. Carpology of the Chenopodiaceae Family in Connection with the Problems of Phylogeny, Systematics and Diagnostics of Its Representatives, Abstract of a PhD Thesis for a Doctor of Biological Sciences, Moscow, 2015.
- Bochantsev, V.P. The Genus Salsola L., a Brief History of Its Development and Dispersal. Bot. J. 1969, 54, 989–1001. [Google Scholar]
- Bochantsev, V.P. Review of Species of the Section Coccosalsola Fenzl of the Genus Salsola L. Nov. Syst. Vyssh. Rast. 1976, 13, 74–102. [Google Scholar]
- Bochantsev, V.P. Review of the Genus Halothamnus Jaub. et Spach (Chenopodiaceae). Nov. Syst. Vyssh. Rast. 1981, 18, 146–176. [Google Scholar]
- Freitag, H.; S. Rilke. Salsola [s. l.]. In Flora Iranica; Editor K.H. Rechinger, Graz: Akademische Druck- und Verlagsanstalt, 1997; Volume 17, pp. 2154–255.
- Kamelin, R.V. Ancient Xerophilous Family Chenopodiaceae in the Flora of Turan and Central Asia. Bot. J. 2011, 96, 441–464. [Google Scholar]
- Akhani, H.; Edwards, G.; Roalson, E.H. Diversification of the Old World Salsoleae s.l. (Chenopodiaceae): Molecular Phylogenetic Analysis of Nuclear and Chloroplast Data Sets and a Revised Classification. Int. J. Plant Sci. 2007, 168, 931–956. [Google Scholar] [CrossRef]
- Almerekova, S.; Yermagambetova, M.; Osmonali, B.; Vesselova, P.; Turuspekov, Y.; Abugalieva, S. Complete Plastid Genome Sequences of Four Salsoleae s.l. Species: Comparative and Phylogenetic Analyses. Biomolecules 2024, 14, 890. [Google Scholar] [CrossRef]
- Tzvelev, N.N. Notes on Chenopodiaceae of Eastern Europe. Ukrains’kij Botaničnij Žurnal 1993, 50, 78–85. [Google Scholar]
- POWO. 2025. Plants of the World Online. Royal Botanic Gardens, Kew. Accessed February 27, 2025. http://www.plantsoftheworldonline.org/.
- Khasanov, F.O. (Ed.). Red Book of the Republic of Uzbekistan. Volume 1: Plants; Chinor ENK: Tashkent, Uzbekistan, 2019.
- Rakhimova, N.K.; Rakhimova, T.; Shomurodov, Kh.F.; Sharipova, V.K. The Status of Coenopopulations of Xylosalsola chiwensis (Popov) Akhani & Roalson and Scorzonera bungei Krasch. & Lipsch. on the Ustyurt Plateau (Uzbekistan). Arid Ecosyst. 2023, 13, 189–195. [Google Scholar] [CrossRef]
- Pavlov, N.V. (Ed.) Flora of Kazakhstan; Publishing House of the Academy of Sciences of the Kazakh SSR: Alma-Ata, 1960; Volume 3.
- Rakhimova, N.K. The Current State of the Anabasis salsa Pasture Varieties in the Karakalpak Ustyurt (Uzbekistan). Probl. Bot. South Sib. Mong. 2022, 21, 144–147. [Google Scholar] [CrossRef]
- Baitulin, I.O. (Ed.). Red Data Book of Kazakhstan: Plants, 2nd ed.; ArtPrintXXI LLP: Astana, Kazakhstan, 2014.
- iNaturalist. 2024. Xylosalsola chiwensis. Accessed February 28, 2024. https://www.inaturalist.org/taxa/1021062.
- GBIF. 2024. Xylosalsola chiwensis. Accessed February 28, 2024. https://www.gbif.org/search?q=Xylosalsola%20chiwensis.
- Arifkhanova, M.M. Vegetation of the Fergana Valley. Fan Publishing House: Tashkent, 1967.
- Plantarium. 2024. Xylosalsola chiwensis Image. Accessed February 28, 2024. https://www.plantarium.ru/page/image/id/830093.html.
- Urkimbaev, Sh.U.; Nurmukhambetov, D.E. Flora of the Ustyurt Reserve. Zhanaozen, Kazakhstan, 2011.
- Smelyansky, I.E.; Pestov, M.V.; Terentyev, V.A.; Laktionov, A.P.; Nurmukhambetov, Zh.E.; Dieterich, Til; Sultanova, B.M; Mukhashov, A.T.; Barashkova, A.N. Justification and Prospects for the Creation of the 'Southern Ustyurt' Cluster Site of the Ustyurt State Nature Reserve (Kazakhstan). In Current State and Problems of Biodiversity Conservation of the Ustyurt Plateau; Astrakhan State University: Astrakhan, Russia, 2024; pp. 9–82.
- Aralbay, N.K. (Ed.). Catalogue of Rare and Endangered Plant Species of the Mangistau Region (Red Book). Aktau, Kazakhstan, 2006; 22 pp.
- Rachkovskaya, E.I.; Volkova, E.A.; Khramtsov, V.N., eds. Botanical Geography of Kazakhstan and Middle Asia (Desert Region). Boston-Spectrum: St. Petersburg, Russia, 2003.
- Aralbaev, N.K. Scheme of New Floristic Zoning of the Territory of Kazakhstan (Materials for the 2nd Edition of the Flora of Kazakhstan). Poisk, Ser. Tech. Nat. Sci. 2002, 4 (2), 66–72.
- Faizov, K.Sh. Soils of the Kazakh SSR. Soils of the Guryev Region. Volume 13. Academy of Sciences of the Kazakh SSR: Alma-Ata, 1970.
- Goloskokov, V.P.; Polyakov, P.P. Chenopodiaceae. In Flora of Kazakhstan, Volume 3; Publishing House of the Academy of Sciences of the Kazakh SSR: Alma-Ata, 1960; pp. 198–359.
- Pratov, U. Chenopodiaceae." In Plant Identifier of Plants of Central Asia, Volume 3; Fan Publishing House: Tashkent, 1972; pp. 20–98.
- IUCN. 2024. The IUCN Red List of Threatened Species: Xylosalsola chiwensis. Accessed February 28, 2024. https://www.iucnredlist.org/search?query=Xylosalsola%20chiwensis&searchType=species.
- Serebryakov, I.G. Life forms of higher plants and their significance. In Field Geobotany; Lavrenko, E.M., Korchagin, A.A., Eds.; Vol. 3; Academy of Sciences of the USSR: Moscow–Leningrad, 1964.
- Raunkiaer, C. Statistik der Lebensformen als Grundlage für die biologische Pflanzengeographie. Beihefte zum Botanischen Zentralblatt 1910, 27, 171–206. [Google Scholar]
- Korovin, E.P. Vegetation of Central Asia and Southern Kazakhstan. Publishing House of the Academy of Sciences of the UzSSR: Tashkent, 1961.
- State Climate Cadastre. Available online: http://ecodata.kz:3838/app_persona/ (accessed on 26 February 2025).
- Butnik, A.A.; Ashurmetov, O.A.; Nigmanova, R.N.; Payzieva, S.A. Ecological Anatomy of Desert Plants in Central Asia (Half–Shrubs, Shrubs); Fan Publishing House: Tashkent, Uzbekistan, 2001; p. 132. [Google Scholar]
- Butnik, A.A.; Duschanova, G.M.; Yusupova, D.M.; Abdullaeva, A.T.; Abdinazarov, S.H. Types leaf mesophyll species of Chenopodiaceae Vent. Central Asia and their role in the monitoring of desertification. J. Nov. Appl. Sci. 2017, 6, 13–21. [Google Scholar]
- Vesselova, P.V.; Alikhanova, A.A. Anatomical structure of assimilative organs in dominant species of the family Chenopodiaceae (Amaranthaceae s.l.). Experimental Biology 2024, 100, 19–32. [Google Scholar] [CrossRef]
- Lavrenko, E.M. Main Features of Botanical Geography of the Deserts of Eurasia and North Africa. Komarov Readings, USSR Academy of Sciences: Leningrad–Moscow, 1962.
- Kadereit, G.; Borsch, T.; Weising, K.; Freitag, H. Phylogeny of Amaranthaceae and Chenopodiaceae and the evolution of C4 photosynthesis. Int. J. Plant Sci. 2003, 164, 959–986. [Google Scholar] [CrossRef]
- Kadereit, G.; Freitag, H. Molecular phylogeny of Camphorosmeae (Camphorosmoideae, Chenopodiaceae): Implications for biogeography, evolution of C4-photosynthesis and taxonomy. Taxon 2011, 60, 51–78. [Google Scholar] [CrossRef]
- Sage, R.F.; Sage, T.L.; Kocacinar, F. Photorespiration and the evolution of C4 photosynthesis. Annu. Rev. Plant Biol. 2012, 63, 19–47. [Google Scholar] [CrossRef]
- Edwards, E.J.; Smith, S.A. Phylogenetic analyses reveal the shady history of C4 grasses. Proc. Natl. Acad. Sci. USA 2010, 107, 2532–2537. [Google Scholar] [CrossRef]
- Edwards, G.E.; Voznesenskaya, E.V. C4 photosynthesis: Kranz forms and single-cell C4 in terrestrial plants; C4 Photosynthesis and Related CO₂ Concentrating, Mechanisms, Raghavendra, A.S., Sage, R.F., Eds.; Springer: Dordrecht, The Netherlands, 2011; pp. 29–61. [Google Scholar] [CrossRef]
- Peter, G.; Katinas, L. A new type of Kranz anatomy in Asteraceae. Aust. J. Bot. 2003, 51, 217–226. [Google Scholar] [CrossRef]
- Osmonali, B.B.; Akhtaeva, N.Z.; Veselova, P.V.; Kudabaeva, G.M.; Kurbatova, N.V. Anatomical structure features of various species of the genus Salsola L. Probl. Bot. South Sib. Mong. 2020, 19, 146–154. [Google Scholar] [CrossRef]
- An’kova, T.V.; Lomonosova, M.N.; Voronkova, M.S.; Petruk, A.A.; Osmonali, B.; Vesselova, P.V. IAPT chromosome data 32. Taxon 2020, 69, 1126–1132. [Google Scholar] [CrossRef]
- Lomonosova, M.N.; Pankova, T.V.; Korolyuk, E.A.; Shaulo, D.N.; Osmonali, B.; Nikolin, E.G. Nuclear genome size analysis in Krascheninnikovia ceratoides s.l. (Amaranthaceae). Bot. Pac. 2024, 13, 183–187. [Google Scholar] [CrossRef]
- Pankova, T.V.; Lomonosova, M.N.; Vaulin, O.V.; Korolyuk, A.Yu.; Korolyuk, E.A.; Shaulo, D.N.; Osmonali, B. Cytogeography of the polyploid complex Bassia prostrata s.l. (Chenopodiaceae) based on genome size analysis and PCR-RFLP cpDNA. Russ. J. Genet. 2024, 60, 274–287. [Google Scholar] [CrossRef]
- Semiotroczeva, N.L. Examinatio caryosystematica specierum monnularum generum Salsola L. et Climacoptera Botsch. Not. Syst. Herb. Inst. Bot. Acad. Sci. Kazakhst. 1983, 13, 66–70. [Google Scholar]
- Lomonosova, M.N.; Krasnikov, A.A.; Krasnikova, S.A. Chromosome numbers of Chenopodiaceae family members of the Kazakhstan flora. Bot. J. 2003, 88, 134–135. [Google Scholar]
- Bochantseva, Z. P. O chislakh chromosom. In Introd. Akklim. Rasteny Akad. Nauk Uzbekist. SSR, Tashkent, 1972; pp. 45–53.
- Morgan, H.D.; Westoby, M. The relationship between nuclear DNA content and leaf strategy in seed plants. Ann. Bot. 2005, 96, 1321–1330. [Google Scholar] [CrossRef]
- Koce, J.D.; Škondrić, S.; Bačič, T.; Dermastia, M. Amounts of nuclear DNA in marine halophytes. Aquat. Bot. 2008, 89, 385–389. [Google Scholar] [CrossRef]
- Osmonali, B.B.; Vesselova, P.V.; Kudabayeva, G.M.; Skaptsov, M.V.; Shmakov, A.I.; Friesen, N. Phylogeny and flow cytometry of the genus Kalidium Moq. (Amaranthaceae s.l.) in Kazakhstan. Plants 2023, 12, 2619. [Google Scholar] [CrossRef]
- Almerekova, S.; Favarisova, N.; Turuspekov, Y.; Abugalieva, S. Cross-genera transferability of microsatellite markers and phylogenetic assessment of three Salsola species from Western Kazakhstan. Proc. Latv. Acad. Sci., Sect. B, Nat. Exact Appl. Sci. 2020, 74, 325–334. [Google Scholar] [CrossRef]
- Pyankov, V.I.; Artyusheva, E.G.; Edwards, G.E.; Black, C.C.; Soltis, P.S. Phylogenetic analysis of tribe Salsoleae (Chenopodiaceae) based on ribosomal ITS sequences: implications for the evolution of photosynthesis types. Am. J. Bot. 2001, 88, 1189–1198. [Google Scholar] [CrossRef]
- Annabayramov, B. The Red Book of Turkmenistan. Volume 1: Plants and Fungi; 3rd ed., revised and enlarged; Ylym: Ashgabat, Turkmenistan, 2011.
- Red Book of Turkmenistan. Vol. 1. Plants; Ashgabat, Turkmenistan, 2024; p. 294.
- Bykov, B. A. Geobotany (3rd ed.). Nauka: Alma-Ata, 1978.
- Drude, O. Die Ökologie der Pflanzen. Vieweg: Braunschweig, 1913.
- Pavlov, N. V. (Ed.). Flora of Kazakhstan; Vols. 1–9; Publishing House of the Academy of Sciences of the Kazakh SSR / Nauka: Alma-Ata, 1956–1966.
- Goloskokov, V. P. (Ed.). Illustrated Guide for Identification of the Plants of Kazakhstan; Vols. 1–2; Nauka: Alma-Ata, 1969–1972.
- Sörensen, T. A method of establishing groups of equal amplitude in plant sociology based on similarity of species and its application to analyses of the vegetation on Danish commons. Kongelige Danske Videnskabernes Selskab 1948, 5, 1–34. [Google Scholar]
- Pyankov, V.I.; Voznesenskaya, E.V.; Kondratschuk, A.V.; Black, C.C. A comparative anatomical and biochemical analysis in Salsola (Chenopodiaceae) species with and without a Kranz type leaf anatomy: a possible reversion of C4 to C3 photosynthesis. Am. J. Bot. 1997, 84, 597–606. [Google Scholar] [CrossRef]
- Voznesenskaya, E.V.; Koteyeva, N.; Akhani, H.; Roalson, E.; Edwards, G.E. Structural and phylogenical analyses in Salsoleae (Chenopodiaceae) indicate multiple transitions among C3, intermediate and C4 photosynthesis. J. Exp. Biol. 2013, 64, 3583–3604. [Google Scholar] [CrossRef]
- Vesselova, P.; Alikhanova, A. Anatomical structure of assimilative organs in dominant species of the family Chenopodiaceae (Amaranthaceae s.l.). Experimental Biology 2024, 100, 19–32. [Google Scholar] [CrossRef]
- Schüssler, C.; Freitag, H.; Koteyeva, N.; Schmidt, D.; Edwards, G.; Voznesenskaya, E. V.; Gudrun Kadereit. Molecular phylogeny and forms of photosynthesis in tribe Salsoleae (Chenopodiaceae). J. Exp. Biol. 2016, 68, 207–223. [CrossRef]
- Osmonali, B.B.; Ahtaeva, N.Z.; Vesselova, P.V.; Kudabayeva, G.M.; Kurbatova, N.V. Features of the anatomical structure of various species of the genus Salsola L. Probl. Bot. South Sib. Mong. 2020, 19, 146–154. [Google Scholar] [CrossRef]
- Doležel, J.; Sgorbati, S.; Lucretti, S. Comparison of three DNA fluorochromes for flow cytometric estimation of nuclear DNA content in plants. Physiol. Plant. 1992, 85, 625–631. [Google Scholar] [CrossRef]
- Skaptsov, M.V.; Kutsev, M.G.; Smirnov, S.V.; Vaganov, A.V.; Uvarova, O.V.; Shmakov, A.I. Standards in plant flow cytometry: an overview, polymorphism and linearity issues. Turczaninowia 2024, 27, 86–104. [Google Scholar]
- Doležel, J.; Greilhuber, J.; Lucretti, S.; Meister, A.; Lysák, M.; Nardi, L.; Obermayer, R. Plant genome size estimation by flow cytometry: inter-laboratory comparison. Ann. Bot. 1998, 82, 17–26. [Google Scholar] [CrossRef]
- Kutsev, M. G. Fragmentnyy analiz DNK rasteniy: RAPD, DAF, ISSR. ARTIKA: Barnaul, Russia, 2009.
- Kutsev, M. G.; Uvarova, O. V.; Sinitsyna, T. A. Set of synthetic oligonucleotides for the amplification and sequencing of ITS1-5.8S-ITS2 vascular plants. RU Patent 2528063, 2014.
- Oxelman, B.; Lidén, M.; Berglund, D. Chloroplast rps16 intron phylogeny of the tribe Silleneae (Caryophyllaceae). Plant Syst. Evol. 1997, 206, 393–410. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The Clustal X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Swofford, D.L. PAUP*: Phylogenetic Analysis Using Parsimony (* and Other Methods). Version 4. Sinauer Associates, Sunderland, MA, USA, 2002.
- Kluge, A.G.; Farris, J.S. Quantitative phyletics and the evolution of anurans. Syst. Zool. 1969, 18, 1–32. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence limits on phylogenies: an approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed]
- Almerekova, S.; Yermagambetova, M.; Osmonali, B.; Vesselova, P.; Abugalieva, S.; Turuspekov, Y. Characterization of the plastid genomes of four Caroxylon Thunb. species from Kazakhstan. Plants 2024, 13, 1332. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Research area.
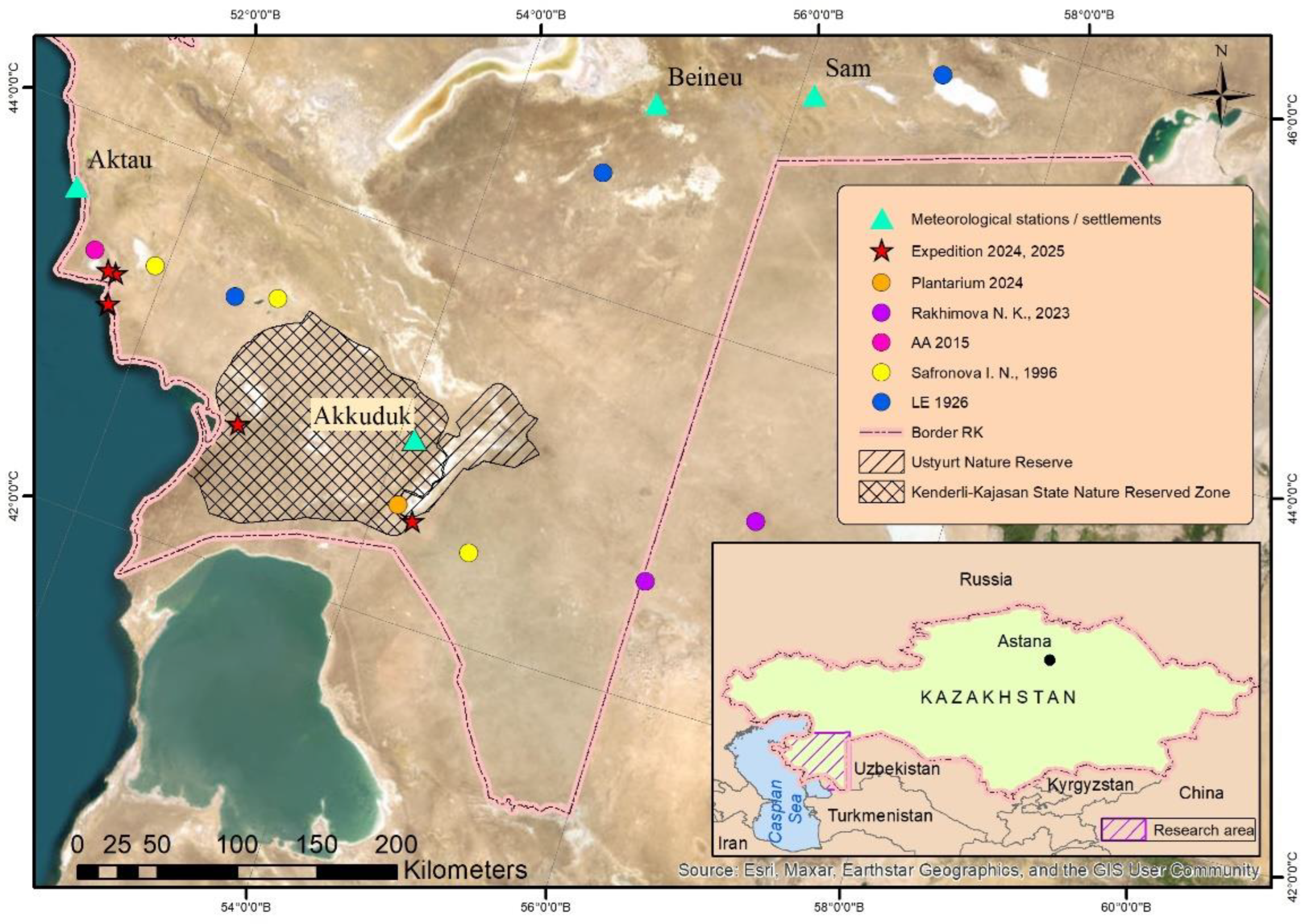
Figure 2.
The studied species Xy. chiwensis.

Figure 3.
Life forms: A – according to Raunkiaer classification: hemicryptophytes (Hc), geophytes (G), therophytes (Th), chamaephytes (Ch), phanerophytes (Ph); B – according to Serebryakov classification: Tree (Tr); Shrubs and dwarf shrubs (Sh); Semishrubs (SS); Herbaceous forms: perennials (HP), annuals (HA).
Figure 3.
Life forms: A – according to Raunkiaer classification: hemicryptophytes (Hc), geophytes (G), therophytes (Th), chamaephytes (Ch), phanerophytes (Ph); B – according to Serebryakov classification: Tree (Tr); Shrubs and dwarf shrubs (Sh); Semishrubs (SS); Herbaceous forms: perennials (HP), annuals (HA).
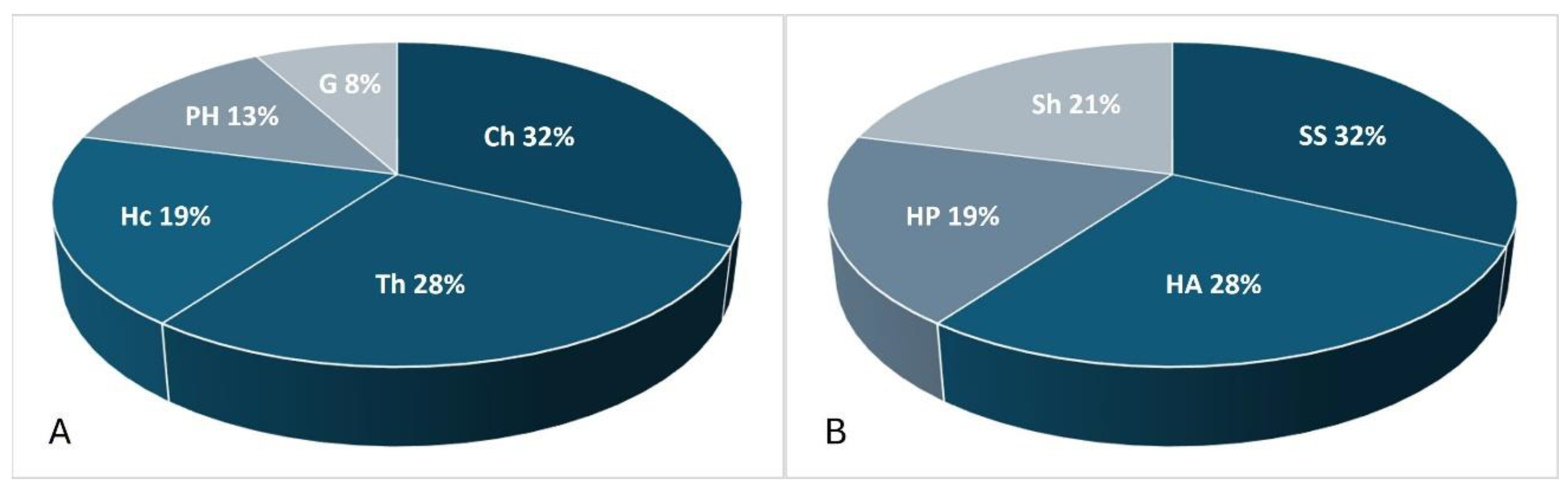
Figure 4.
Cenopopulations 1 and 2 with Xy. chiwensis.

Figure 5.
Cenopopulations 3 and 4 with Xy. chiwensis.

Figure 6.
Cenopopulations 5 and 6 with Xy. chiwensis.

Figure 7.
Average annual precipitation and air temperature.
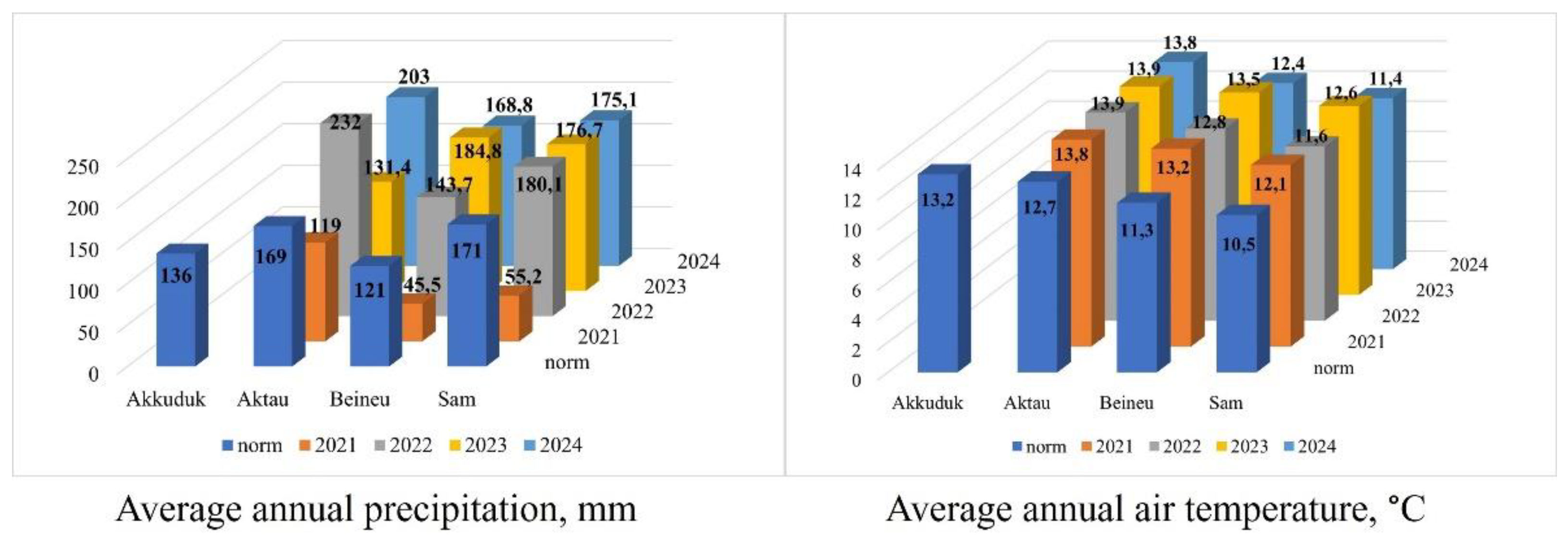
Figure 8.
Absolute maximum and minimum air and soil surface temperatures.
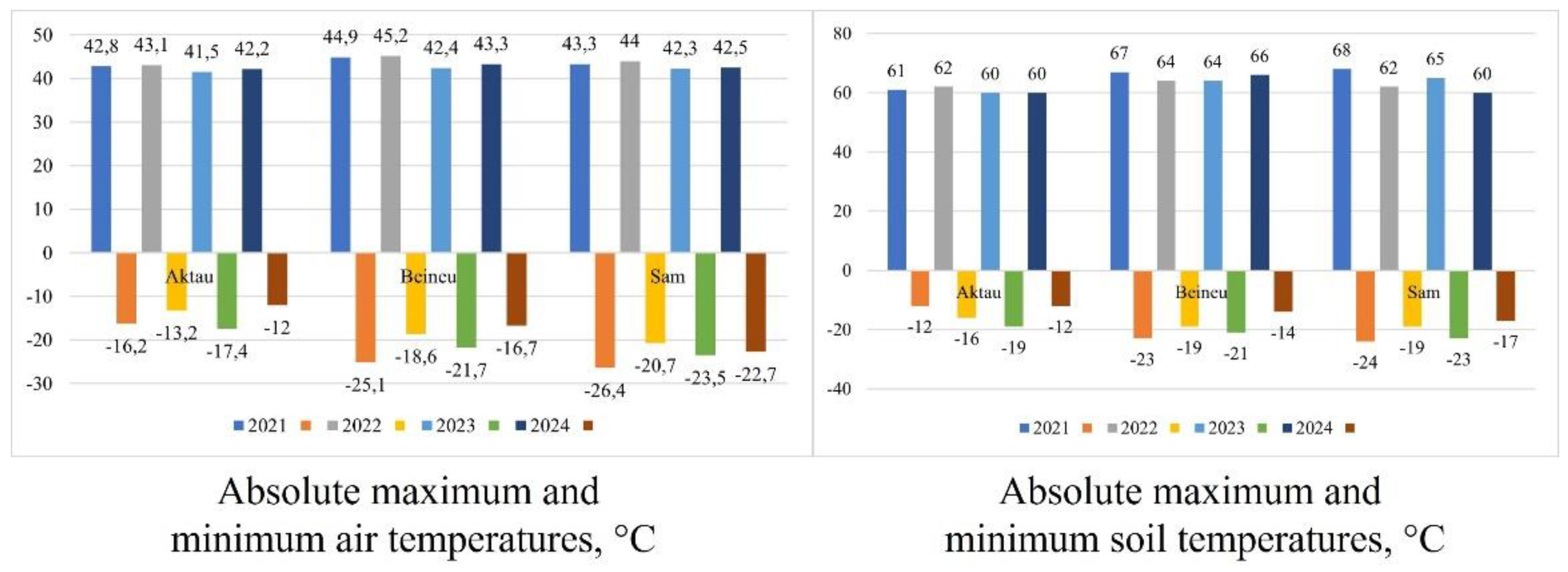
Figure 9.
A – Type specimen (AA); B, C – Our collected specimens.

Figure 10.
Fruit morphology of two species of the genus Xylosalsola. A-E – Xy. chiwensis; F-H – Xy. аrbuscula.
Figure 10.
Fruit morphology of two species of the genus Xylosalsola. A-E – Xy. chiwensis; F-H – Xy. аrbuscula.

Figure 11.
Anatomical cross-sections of the studied species: A-B – Xy. chiwensis; C-D - Xy. аrbuscula. e – epidermis, h – hypodermis, p – palisade mesophyll, kc – Kranz cells, d – druse, pvb – peripheral vascular bundle, wc – water-storage cell, vb – vascular bundle.
Figure 11.
Anatomical cross-sections of the studied species: A-B – Xy. chiwensis; C-D - Xy. аrbuscula. e – epidermis, h – hypodermis, p – palisade mesophyll, kc – Kranz cells, d – druse, pvb – peripheral vascular bundle, wc – water-storage cell, vb – vascular bundle.
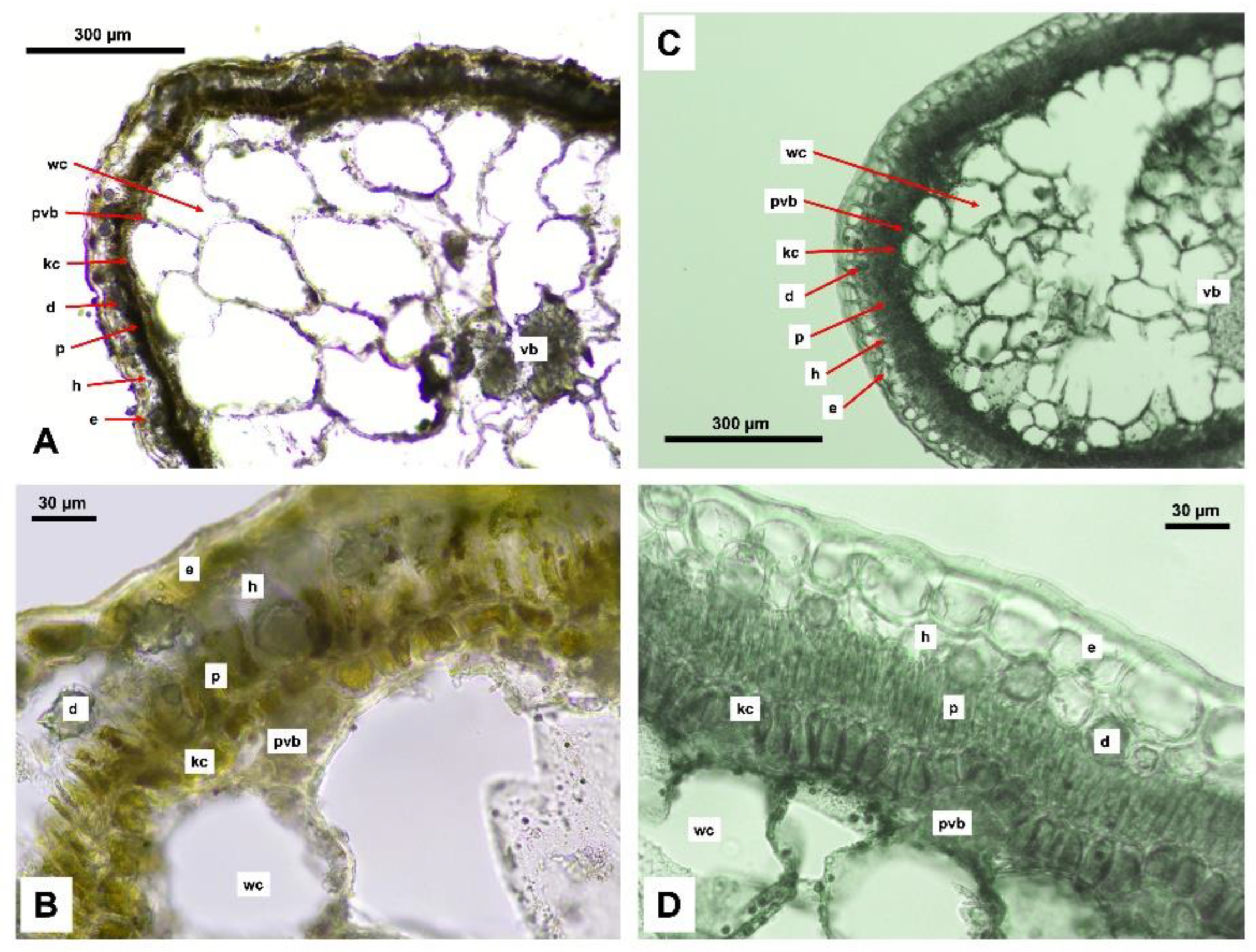
Figure 12.
Phylogenetic nrITS tree of the subfamily Salsoloideae. The joint presence of Bayesian with a probability greater than 0.98 and bootstrap support greater than 95% is indicated by a black dot. The genus under study is indicated by a short vertical line and a solid green coloration. Tribes are marked by vertical lines. The accession number of sequences from the NCBI GenBank is given for species name.
Figure 12.
Phylogenetic nrITS tree of the subfamily Salsoloideae. The joint presence of Bayesian with a probability greater than 0.98 and bootstrap support greater than 95% is indicated by a black dot. The genus under study is indicated by a short vertical line and a solid green coloration. Tribes are marked by vertical lines. The accession number of sequences from the NCBI GenBank is given for species name.
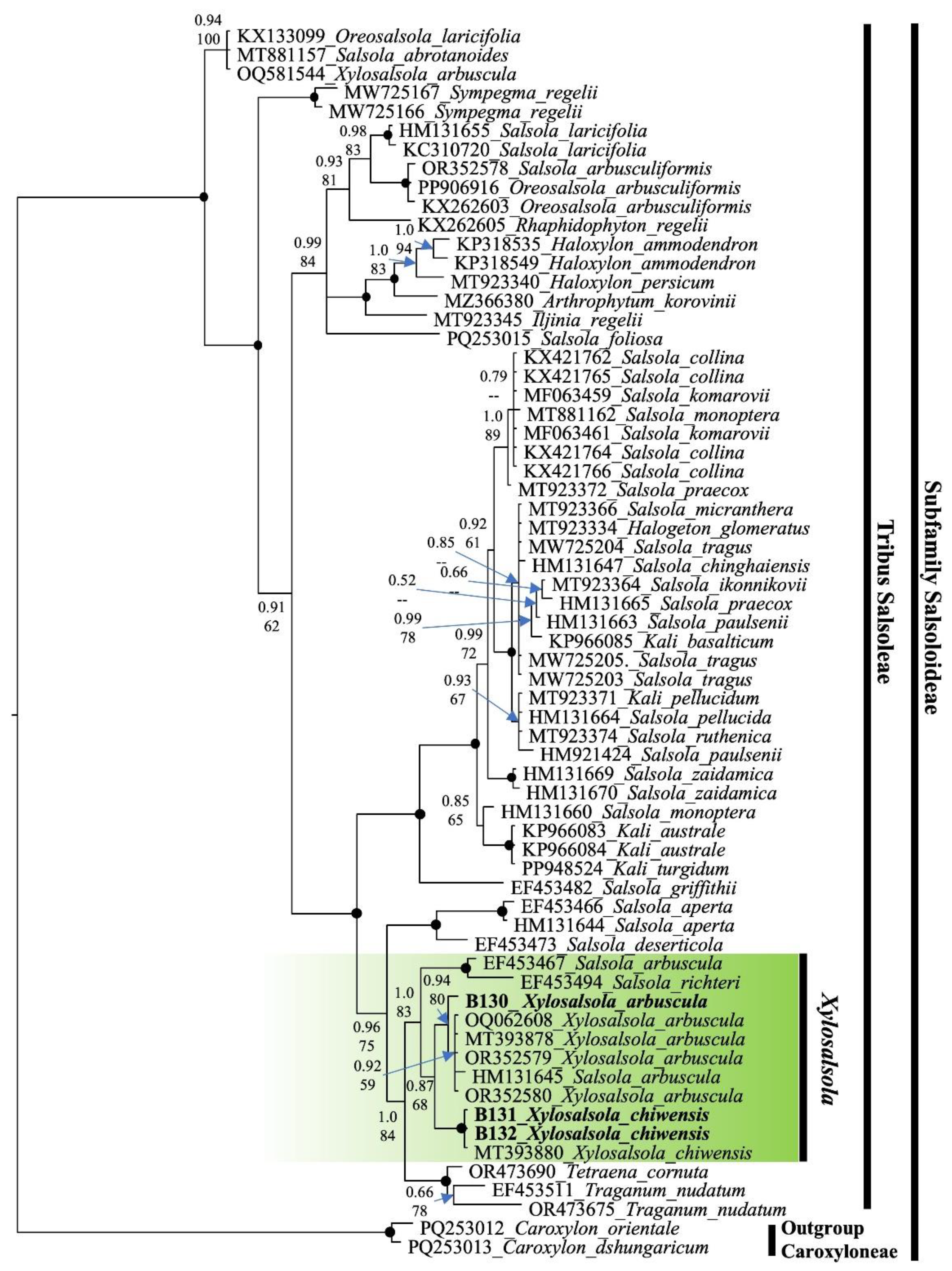
Figure 13.
Plastid Tree (rpsF-prsR2) of the subfamily Salsoloideae. The joint presence of Bayesian with a probability greater than 0.98 and bootstrap support greater than 95% is indicated by a black dot. The bold font indicates the specimens we selected. The genus under study is indicated by a short vertical line and a solid green coloration. Tribes are marked by vertical lines. The accession number of sequences from the NCBI GenBank is given for species name.
Figure 13.
Plastid Tree (rpsF-prsR2) of the subfamily Salsoloideae. The joint presence of Bayesian with a probability greater than 0.98 and bootstrap support greater than 95% is indicated by a black dot. The bold font indicates the specimens we selected. The genus under study is indicated by a short vertical line and a solid green coloration. Tribes are marked by vertical lines. The accession number of sequences from the NCBI GenBank is given for species name.
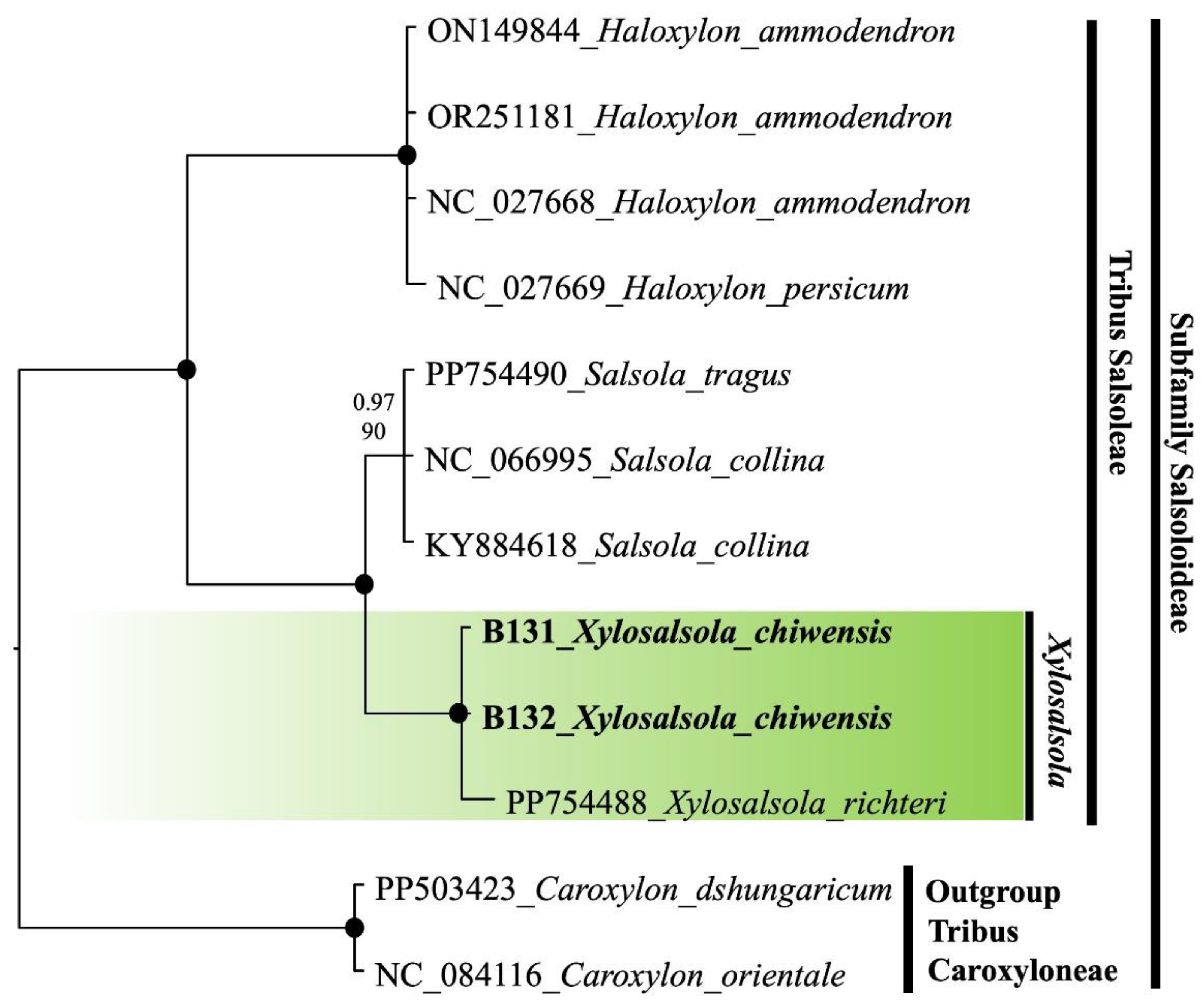

Table 1.
Morphometric data on the fruits of two species.
| Parameters | Xy. chiwensis | Xy. arbuscula |
| Fruit diameter with wings, mm | Mean: 5.30 (±0.02) Min: 5.19 / Max: 5.37 SD: 0.06 |
Mean: 8.53 (±0.17) Min: 7.85 / Max: 9.53 SD: 0.53 |
| Fruit diameter without wings, mm | Mean: 2.81 (±0.09) Min: 2.47 / Max: 3.18 SD: 0.29 |
Mean: 2.57 (±0.06) Min: 2.29 / Max: 2.75 SD: 0.19 |
| Cone length, mm | Mean: 1.09 (±0.03) Min: 0.90 / Max: 1.19 SD: 0.09 |
Mean: 1.90 (±0.05) Min: 1.60 / Max: 2.13 SD: 0.17 |
| Bract length, mm | Mean: 6.07 (±0.27) Min: 4.93 / Max: 8.03 SD: 0.87 |
Mean: 8.91 (±1.13) Min: 5.18 / Max: 12.94 SD: 3.59 |
Table 2.
Anatomical leaf structure of two species.
| Parameter (µm) | Xy. chiwensis | Xy. arbuscula |
| Epidermis (E) | 20.09 ± 2.24 (15.72 – 24.40) | 35.03 ± 3.29 (30.20 – 39.00) |
| Hypodermis (H) | 20.96 ± 2.97 (16.94 – 25.82) | 16.31 ± 2.43 (11.60 – 20.60) |
| Palisade Mesophyll (P) | 29.59 ± 2.88 (23.76 – 33.19) | 33.67 ± 2.98 (30.00 – 41.50) |
| Kranz Cells (KC) | 17.44 ± 2.97 (13.32 – 22.24) | 21.55 ± 4.28 (14.50 – 29.00) |
Notes: The mean value is given along with the standard deviation (±). The range of values (minimum – maximum) is provided in parentheses.
Table 3.
DNA content of the studied Xylosalsola samples.
| Species | Mean 2C ± SD, pg | CV |
| Xy. arbuscula | 3.250 | - |
| Xy. arbuscula | 6.723 ± 0.582 | 8.65% |
| Xy. chiwensis | 2.483 ± 0.191 | 7.68% |
Table 4.
Origin and number of sequences used for phylogenetic analyses.
| Accession | Name | Coordinates | Voucher | rITS | rps16f-rps16r2 |
| В130 | Xy. arbuscula | 44.79060083 N 63.14563274 E |
AA0003576 | PV032237 | - |
| В131 | Xy. chiwensis | 43.250147 N 51.671042 E |
AA0003564 | PV032238 | PV036952 |
| B132 | Xy. chiwensis | 43.080778 N 51.696008 E |
AA0003563 | PV032239 | PV036953 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



