
Submitted:
14 March 2025
Posted:
17 March 2025
You are already at the latest version
Abstract
Frankenia is a morphologically complex genus with some species exhibiting few diagnostic characters and significant morphological variability. This led to misidentification or synonymisation of many names, based on one or a few diagnostic traits. This phenomenon affects the annual sea-heath, F. pulverulenta, a Eurasian-Mediterranean herb that has become subcosmopolitan, in which several entities were included due to shared features, namely the annual lifespan or the flattened leaves. However, this fact also extends to shrubby species, such as the Madeiran F. cespitosa. Here, integrative taxonomic studies were undertaken, encompassing detailed morphological descriptions of macro- and microcharacters, along with molecular phylogenetic analyses of both nuclear ribosomal (ITS1-5.8S-ITS2 region) and plastid (matK gene) DNA sequence data, and biogeographic data. The research resulted in the most complete phylogenetic trees of Frankenia to date, leading to the reinstatement of two African species broadly differing from F. pulverulenta. Firstly, F. florida L.Chevall., a name applied to a species occurring in the Saharan regions of Algeria, Morocco, Mali and Mauritania, is often accepted as a variety or subspecies of the annual sea-heath. In contrast, F. densa Pohnert, a species endemic to Namibia and northern South Africa, is synonymised with F. pulverulenta. However, since those two names were later homonyms of two Chilean and Australian plants, they were deemed illegitimate upon publication. Consequently, two new names are proposed for them: F. sahariensis and F. dinteri, respectively. The substantiation of both entities as independent species is provided by data on morphology, distribution, ecology and molecular phylogenetics, which demonstrate their distinctiveness from F. pulverulenta. Nomenclatural synonymy and types are also presented for all concerned names, including the designation of two new lectotypes.
Keywords:
African flora
; Frankenia
; Frankeniaceae
; ITS
; matK
; new names
; nomenclature
; phylogenetic analyses
; taxonomy
1. Introduction
The genus Frankenia L. is the sole member currently recognised within Frankeniaceae Desv., nom. cons., when expanded to include Anthobryum Phil., Beatsonia Roxb., Hypericopsis Boiss. and Niederleinia Hieron. [1,2,3,4], and references therein. So broadly treated about 80−90 species are often accepted that are distributed in temperate or tropical regions, commonly in Mediterranean-type areas around the world, but with ca. 40 species found in Australia [5,6]. Most members of Frankenia exhibit a woody or suffruticose habit, erect to prostrate, sometimes densely pulvinate or with subspinescent branches, but some others are prostrate to ascendant annual herbs [7,8]. With very few exceptions, most species usually grow on saline soils, including coastal and inland salt marshes and gypsum- or calcium-rich outcrops [9], often enduring semi-arid to arid climates that mostly show prevalent winter rainfall [10]. One such exception is the South African narrow endemic F. fruticosa J.C.Manning & Helme, which occurs on quartz soils [11]. Because most taxa grow under strongly stressing conditions, many endemics are found which are restricted to small areas or archipelagos [12], whereas others such as the annual sea-heath, F. pulverulenta L., have become subcosmopolitan [13,14]. These peculiar lifestyles have led to acquisition of specialised structures, such as salt-secreting multicellular glands that often generate whitish salt crusts on leaves and stems [8]. This characteristic is shared with representatives of some related families in Caryophyllales, such as Plumbaginaceae Juss., nom. cons., or Tamaricaceae Link, nom. cons. [15], though in each case they exhibit distinct structural features [16].
From a taxonomic perspective, the first comprehensive treatment of the genus was undertaken by Niedenzu [7], who accepted Frankenia, Anthobryum, Beatsonia, Hypericopsis and Niederleinia as distinct genera. Just a few years later, Summerhayes [5] published a revision of the Australian taxa and described most of the species currently accepted. More recently, Whalen [17] conducted a taxonomic study on the American Frankeniaceae, where a detailed morphological revision was presented along with a discussion on the status of the South American genera Anthobryum and Niederleinia, which she synonymised to Frankenia. Consequently, though being a good starting point for a treatment of the whole genus, Niedenzu’s [7] account is incomplete and requires a thorough update. Among the partial monographs, different authors have presented taxonomic arrangements for the genus in large regions of Eurasia and Africa. After the revisions for Flora of the USSR [18], Flora europaea [19] and Flora iranica [20], with some additions by Lomonosova [21], up to seven species plus twelve infraspecific taxa (subspecies, varieties, and forms) are widely accepted.
For Africa, the revision of the Moroccan taxa by Nègre [22] stands out, in which the author studied in detail the circumscription and synonymy of taxa previously summarised by Jahandiez and Maire [23] and Emberger and Maire [24] for Morocco and neighbouring areas. He accepted there four species plus fourteen subspecific and varietal taxa. More recently, Crespo et al. [12] have clarified the circumscription of some neglected names concerning southern African plants and have recognised seven species, most of them endemics often misidentified with other widespread relatives. However, the diverse taxonomic criteria adopted within the last decades by authors dealing with Frankenia in diverse territories have generated quite heterogeneous treatments that much difficult a global understanding of the genus for most taxonomists.
Furthermore, molecular studies including species of Frankenia are also scarce. In a work focused on relationships between Frankeniaceae and Tamaricaceae based on nrDNA sequences, Gaskin et al. [3] included eight Frankenia species from North America, Australia, and Eurasia, such as the outstanding F. persica (Boiss.) Jaub. & Spach (≡ Hypericopsis persica Boiss.). The most recent phylogenetic studies on the genus were published by Crespo et al. [12], who presented a preliminary tree based on ITS sequences including 16 species from Eurasia and Africa, which served as a basis to recover or describe some neglected species from South Africa often misidentified with F. pulverulenta.
Very few species in this genus are annual or biennial. In particular, the annual sea-heath, Frankenia pulverulenta, is a Eurasian-Mediterranean ephemeral herb that today is broadly distributed around the world [14], occurring in saline soils disturbed by human activities [13,14,15,16,17]. According to Crespo et al. [12], it is typically a prostrate plant with many-branched, non-rooting stems, densely and minutely pubescent on one side; leaves flat, broadly obovate-cuneate, subconcolorous, glabrous on the adaxial surface and variably hairy on the abaxial surface, rounded and emarginate at the apex and cuneate at the base; flowers pentamerous, small, axillary, and solitary, scattered; calyx up to 5 mm long; corolla up to 5 mm, pinkish; capsule 3-carpelar; placentas 3, with 12–20 ovules each; seeds numerous, small (up to 0.5−0.7 mm long), ellipsoid, monosulcate, with testa weakly and irregularly ornamented, very sparsely covered with small-sized papillae (4–17 µm), homogeneous (conical-obtuse) in shape.
Recent investigations conducted in southern Africa, through both fieldwork and herbarium studies [12], revealed the existence of peculiar populations showing some superficial resemblance to F. pulverulenta, mostly on account of their flattened roundish to broadly ovate-oblong leaves. However, those authors demonstrated that several distinct biological entities had been overlooked under that designation within the last decades. On the one hand, the name F. nodiflora Lam., often synonymised to F. pulverulenta, was restored for littoral populations of creeping prostrate shrublets, with concolorous glabrous leaves and dense glomerular inflorescences, occurring in saltmarshes and salt pans of Cape Flats (Western Cape Province). On the other, F. nummularia M.B.Crespo, M.Á.Alonso, Mart.-Azorín, J.L.Villar & Mucina was newly described for populations of creeping prostrate shrublets, with discolorous glabrous leaves and loosely dichasial, usually widely branched inflorescences, from inland salt-pans and saline river beds in the southwestern part of the Karoo Region (Northern and Western Cape Provinces). These had usually been misidentified as the annual sea-heath, F. pulverulenta.
Similar research on other African populations of annual or short-lived perennial plants usually related to F. pulverulenta also unveiled surprising data. Firstly, populations of a North African plant, occurring in saline substrates of the western Sahara regions, were described as F. florida L.Chevall., a later homonym of the Chilean F. florida Phil. and hence an illegitimate name according to the Shenzhen Code (hereafter ICN, [25]). That North African name has often been treated as F. pulverulenta var. florida Maire or subsp. florida (Maire) Maire [4,26], or also as merely a synonym of the annual sea-heath. Secondly, populations of a remarkable species growing on saline soils of inland Namibia were described as F. densa Pohnert, a later homonym of the Australian F. densa Summerh., which is hence illegitimate and unavailable for use. However, despite several characteristics allow separation from the morphologically close F. pulverulenta, Pohnert’s name is currently reduced to synonymy of the latter [4].
In the present contribution, as part of our ongoing integrative taxonomic studies on the genus Frankenia, the position and phylogenetic relationships of both F. florida L.Chevall. and F. densa Pohnert are studied. Morphological, distributional, ecological, and molecular phylogenetic data are shown that support recognition of both entities as independent species, not related to F. pulverulenta, for which the new names F. sahariensis and F. dinteri are respectively proposed. Nomenclatural synonyms and types are also reported for all concerned names, including designation of two new lectotypes.
2. Materials and Methods
2.1. Morphological and Habitat Studies
Detailed morphological observations were conducted on both living plants from wild populations and dried herbarium specimens sourced from the herbaria ABH, BM, BOL, G-DC, K, LINN, MA, MPU, NBG, P, and W (acronyms according to Thiers [27]), using an OLYMPUS SZX7 binocular microscope. Digital sheets from the herbaria GH, GZU, HBG, JE, LY, M, MO, PRE, RSA, US, WAG, and Z were reviewed as well. A list of the herbarium material examined, including nomenclatural types, is reported for each species, with barcode numbers are provided after each herbarium acronym when available.
Scanning Electron Microscope (SEM) micrographs of seeds were captured using a JEOL JSM-IT500HR operating at 15 kV. No special treatment was needed for the material before observation. A minimum of 10 mature seeds from different individuals of each studied species (Table 1) were carefully examined. The samples were affixed to metallic stubs and coated with 10 nm of platinum in a QUORUM Q150T ES Plus sputter coater. Measurements on the SEM micrographs were conducted using ImageJ v.1.8.0 [28].
The authors of the taxa cited follow the guidelines of IPNI [29]. Nomenclatural issues accord with the Shenzhen Code (ICN; [25]). For the new species from southern Africa, orthography of geographical names and grid-number system follow Leistner and Morris [30], and the grid-number system accord with the National Geospatial Information [31]. The geographic coding of the collection sites (e.g., 2816BB or -BB, always linked to 4-digit code) refers to the centre point for a 15’ latitude × 15’ longitude sub-tile as defined by the South African topographical map sheet referencing system [31]. This system is broadly known as ‘quarter degree square’ (QDS) grid, since each topographic map sheet is considered a tile (coded, for instance, as 2816), divided into four sub-tiles (coded, for instance, as -BB).
The bioclimatic and biogeographic classification of southern Africa is based on Mucina and Rutherford [32]. For North African species, the biogeographical characterisations of Quézel [33] and Takhtajan [34] were also considered. Distribution maps were created using cartography tools from Google Earth.
2.2. Molecular analyses
Silica gel-dried materials and herbarium vouchers were utilised for total DNA extraction following a modified 2× cetyltrimethylammonium bromide (CTAB) protocol [35]. When herbarium material was unavailable for sampling, silica-gel dried material from wild populations (one sample per population) of each taxon was used, since the phylogenetic trees remained unchanged after addition of new samples from the same population. Total DNA was then purified using MOBIO minicolumns and typically stored in 0.1× TE buffer [10 mM Tris-HCl, 1 mM ethylenediaminetetraacetic acid (EDTA), pH 8.0]. The internal transcribed spacer (ITS) region (comprising the ITS1 spacer, the 5.8S gene, and the ITS2 spacer) of nuclear ribosomal DNA (nrDNA) was amplified using the ITS5 and ITS4 primers [36], while the Maturase K (matK) gene of plastid DNA (cpDNA) was amplified using the XF (TAATTTACGATCAATTCATTC) and 5R (GTTCTAGCACAAGAAAGTCG) primers [37]. Amplifications were conducted in a total reaction volume of 25.5 μl, which included 12.5 μl of DreamTaq PCR Master Mix (2×), 10 μl of distilled water (Thermo Scientific, Waltham, MA, USA), 0.5 μl of 0.4% bovine serum albumin (BSA), 0.5 μl of dimethyl sulfoxide (DMSO), 0.5 μl of each primer (10 pmol/μl), and 1 μl of template DNA, all processed on a 9700 GeneAmp thermocycler (Applied Biosystems). The PCR program for ITS involved an initial step of 2 min at 95 °C, followed by 35 cycles of 95 °C for 1 min, 53 °C for 1 min, and 72 °C for 2 min, concluding with a final extension at 72 °C for 5 min. The PCR program for matK included an initial step of 2 min at 94 °C, followed by 32 cycles of 94 °C for 1 min, 53 °C for 1 min, and 72 °C for 1 min and 30 seconds, finishing with a final extension at 72 °C for 4 min.
Sequencer 4.1 (Gene Codes Corp., Ann Arbor, MI, USA) was utilised to assemble complementary strands and verify software base-calling. Sequence alignment was conducted using MUSCLE [38] within MEGA X v.10.2.6 [39], supplemented by minor manual adjustments to create the final aligned matrix. The analyses incorporated fifty-six samples representing twenty species of Frankenia, encompassing the primary Eurasian and African groups within the genus, and including Myricaria germanica L., Reaumuria alternifolia (Labill.) Britten, R. songarica (Pall.) Maxim., and Tamarix gallica L. as outgroups.
Two datasets were constructed: One for the ITS region (including 54 Frankenia sequences plus 4 outgroup sequences, totalling 720 positions), and another for the matK gene (comprising 38 Frankenia sequences plus 3 outgroup sequences, with a total of 829 positions). Specifically for this study, a total of 38 matK plus 28 ITS sequences were generated (Table 2), while the remaining ones were sourced from various plant databases according to GenBank (https://www.ncbi.nlm.nih.gov/genbank/) availability (see [12]). As a result, only a few accessions in the ITS and matK datasets originate from different plant sources. Accessions listed in GenBank as “Reaumuria hypericoides Willd.” are represented in our trees as R. alternifolia, given that the former name is a superfluous synonym of the latter and, as such, has priority. Likewise, the ITS accession OR183467, previously filed as “Frankenia hirsuta L.” (voucher ABH-45933!), indeed corresponds to the closely related species F. salsuginea Adıgüzel & Aytaç (Table 2).
Phylogenetic analyses of both the ITS region and the matK gene were conducted using Maximum Parsimony (MP), Maximum Likelihood (ML), and Neighbour Joining (NJ) methods. The MP analysis was performed using both PAUP with heuristic search options based on the Nearest Neighbour Interchange (NNI) strategy, and MEGA with heuristic search options applying the Subtree-Pruning-Regrafting (SPR) method with search level 1 [40] for comparative results, with 10,000 replicates. ML [41] and NJ [42] analyses were also executed in MEGA, alongside the selection of the optimal model of DNA substitutions for each method using the Akaike Information Criterion (AIC, [43]); the models with the lowest Bayesian Information Criterion (BIC) scores were deemed most suitable for describing the substitution patterns in the ML and NJ analyses. Additionally, jModelTest 2.1.10 [44] was utilised to identify the best model of DNA substitutions for the Bayesian Inference (BI) analyses, again applying the AIC. Phylogenetic reconstructions for Maximum Likelihood (ML) and evolutionary distances for the Neighbor-Joining (NJ) method were estimated using the Kimura 2-parameter (K2) model [45] for the ITS matrix. This model allowed for some sites to be evolutionarily invariable, with 26.46% of sites fitting this criterion (+I). In contrast, the matK matrix utilised the Tamura 3-parameter (T92) model [46], coupled with a discrete Gamma distribution (G = 1.0742) to model rate variation among sites. For these analyses, all sites in the matrices were considered. To ensure comparability, ambiguous positions were removed for each sequence pair using the pairwise deletion option. This removal did not result in significant differences in the obtained phylogenies, although it affected support percentages for a few branches. Support for all methods was assessed using bootstrapping [47] with 10,000 replicates, retaining only 10 trees per replicate. Clades with bootstrap percentage (BP) values of 50–74% were considered weakly supported, 75–89% moderately supported, and 90–100% strongly supported.
Furthermore, Bayesian inference (BI) analyses were conducted using MrBayes 3.2 [48], where the Markov Chain Monte Carlo (MCMC) algorithm was executed for 10 million generations, with samples taken every 1000 generations. Two independent runs were carried out. The analyses employed the General Time Reversible (GTR) + proportion of invariant sites (I) + Gamma distribution (G) model, as suggested by the results from jModelTest based on the Akaike Information Criterion (AIC). The initial 25% of generations were excluded (burninfrac = 0.25), and the remaining trees were utilised to compile a posterior probability (PP) distribution through a 50% majority-rule consensus. Clade support was again evaluated using Bootstrap analysis [47], with 10,000 replicates while retaining only 10 trees per replicate. Clades with PP≥0.95 were regarded as strongly supported.

Table 2.
List of outgroups and Frankenia accessions used for the phylogenetic analyses, with data on provenance and source plus Genbank codes of the ITS and matK accessions utilised.
Table 2.
List of outgroups and Frankenia accessions used for the phylogenetic analyses, with data on provenance and source plus Genbank codes of the ITS and matK accessions utilised.
| Taxon | Provenance (herbarium voucher) | Source | ITS | Source | matK |
| Tamarix gallica L. | France: Saintes Maries de la Mer (ABH-57865) |
Villar et al. [49] | MH626294 | − | − |
| Italy: Sardinia | Meimberg et al. [50] | − | − | AF204861 | |
| Myricaria germanica L. | Kazakhstan: Zajsanskaya depression (LE) | Zhang et al. [51] | KJ808607 | − | − |
| China: Unspecified (CPG-11863) |
Chen et al. [52] | − | − | KX526795 | |
| Reaumuria alternifolia (Labill.) Britten1 | Azerbaijan: Caucasus (MW) | Zhang et al. [53] | KJ729627 | − | − |
| Reaumuria songarica (Pall.) Maxim. | China: Xinjiang (M) | Zhang et al. [51] | OQ617495 | − | − |
| China: Xinjiang (TURP) | Song et al. [54] | − | − | MT918094 | |
| Frankenia anneliseae M.B.Crespo & al. | South Africa: Klipfontein (ABH-76891) | Crespo et al. [12] | OR183455 | − | − |
| South Africa: Skoverfontein (ABH-83196) | Crespo et al. [12] | OR183456 | This paper | PV258653 | |
| South Africa: Klipfontein (ABH-76872) | − | − | This paper | PV258654 | |
| F. boissieri Reut. ex Boiss. | Spain: Huelva, Lepe, El Terrón (ABH-83544) | This paper | PV241633 | This paper | PV258655 |
| Portugal: Algarve, Vale de Parra (ABH-73553) | This paper | PV241634 | − | − | |
| F. cespitosa Lowe | Portugal: Madeira Is., Porto Santo, Morenos (MA-902612) | This paper | PV241635 | − | − |
| F. capitata Webb & Berthel. | Spain: Canary Is., Las Palmas de Gran Canaria, Isleta (ABH-83612) | Crespo et al. [12] | OR183458 | This paper | PV258656 |
| Spain: Canary Is., Lanzarote, Teguise (ABH-83884) | This paper | PV241636 | − | − | |
| Spain: Canary Is., Lanzarote, Yaiza (ABH-83881) | This paper | PV241637 | This paper | PV258658 | |
| Spain: Canary Is., Lanzarote, Yaiza (ABH-83880) | − | − | This paper | PV258657 | |
| F. composita Pau & Font Quer | Morocco: Al Hoceïma, Cala Iris (ABH-81590) | Crespo et al. [12] | OR183459 | This paper | PV258659 |
| Spain: Murcia, El Carmolí (ABH-84195) | This paper | PV241638 | This paper | PV258660 | |
| F. corymbosa Desf. | Spain: Alicante, Santa Pola (ABH-79956) | Crespo et al. [12] | OR183462 | This paper | PV258663 |
| Morocco: Nador, Punta Charrana (ABH-54294) | Crespo et al. [12] | OR183461 | This paper | PV258662 | |
| Morocco: Al-Hoceïma (ABH-54526) | Crespo et al. [12] | OR183460 | − | − | |
| Spain: Murcia, Cabo Cope (ABH-83531) | Crespo et al. [12] | OR183463 | This paper | PV258661 | |
| F. dinteri M.Á.Alonso & al., nom. nov. | Namibia: Goageb (ABH-76804) | This paper | PV241642 | This paper | PV258666 |
| South Africa: Onseepkans (ABH-83234) | This paper | PV241639 | This paper | PV258667 | |
| South Africa: Daberas Farm (ABH-83264) | This paper | PV241640 | This paper | PV258665 | |
| South Africa: Augrabies (ABH-83265) | This paper | PV241641 | This paper | PV258664 | |
| F. ericifolia C.Sm. ex DC., nom. cons. prop. | Spain: Canary Isl., Tenerife (ABH-79975) | Crespo et al. [12] | OR183464 | This paper | PV258668 |
| Spain: Canary Isl., Tenerife, Güímar (ABH-83613) | Crespo et al. [12] | OR183465 | − | − | |
| Spain: Canary Isl., Tenerife, Granadilla de Abona (ABH-83873) | This paper | PV241646 | This paper | PV258669 | |
| Spain: Canary Isl., Tenerife, Malpaso, IA3069 (MA) | This paper | PV241643 | This paper | PV258670 | |
| Spain: Canary Isl., Lanzarote, Caleta del Mojón Blanco (ABH-83889) | This paper | PV241644 | This paper | PV258671 | |
| Spain: Canary Isl., Lanzarote, Risco de Famara (ABH-83893) | This paper | PV241645 | This paper | PV258672 | |
| F. fruticosa J.C.Mannig & Helme | South Africa: Moedverloren (ABH-76898) | Crespo et al. [12] | OR183466 | − | − |
| F. hirsuta L. | Italy: Puglia, Bari (ABH-84234) | This paper | PV241648 | This paper | PV258673 |
| Cyprus: Akamas (MA-526424) | This paper | PV241647 | − | − | |
| F. ifniensis Caball. | Morocco: Sidi Ifni to Oued Noun (MA-758515) | Crespo et al. [12] | OR183468 | This paper | PV258674 |
| Morocco: El Farsia (MA-712824) | This paper | PV241649 | − | − | |
| Morocco: Gelmim (MA-902279) | This paper | PV241650 | − | − | |
| F. laevis L. | Libya: Cyrenaica, Jbel Akhdar (MA-826355) | This paper | PV241652 | − | − |
| France: Aude, Étang de La Palme (ABH-70584) | Crespo et al. [12] | OR183469 | This paper | PV258676 | |
| Spain: Mallorca, Conejera (ABH-57810) | This paper | PV241651 | This paper | PV258675 | |
| F. nummularia M.B.Crespo & al. | South Africa: Kookfontein River (ABH-83290) | Crespo et al. [12] | OR183471 | − | − |
| South Africa: Tankwa Karoo (ABH-83295) | Crespo et al. [12] | OR183472 | This paper | PV258677 | |
| F. pseudoericifolia Rivas Mart. & al. | Portugal: Cape Verde, São Antão (MA-0906845) | Crespo et al. [12] | OR183473 | − | − |
| F. pulverulenta L. | South Africa: Redelinghuis (ABH-77205) | Crespo et al. [12] | OR183474 | − | − |
| South Africa: Skoverfontein (ABH-83195) | This paper | PV241657 | − | − | |
| Spain: Teruel, Alcañiz (ABH-73564) | Crespo et al. [12] | OR183475 | − | − | |
| Spain: Alicante, Cabo de las Huertas (ABH-74763) | This paper | PV241653 | This paper | PV258678 | |
| Spain: Albacete, Fuentealbilla (ABH-40820) | This paper | PV241654 | − | − | |
| Morocco: Melga-el-Ouidane (ABH-59986) | This paper | PV241655 | This paper | PV258679 | |
| Spain: Canary Isl., Tenerife, Puerto de la Cruz (ABH-79974) | Crespo et al. [12] | OR183477 | This paper | PV258680 | |
| Italy: Puglia, Torre Spechiola (ABH-84244) | This paper | PV241656 | This paper | PV258681 | |
| Spain: Alicante, Cabo de las Huertas (ABH-41853) | Crespo et al. [12] | OR183476 | − | − | |
| F. repens (P.J.Bergius) Fourc. | South Africa: S of Hondeklipbaai (ABH-76862) | Crespo et al. [12] | OR183479 | − | − |
| South Africa: Velddrift (ABH-76849) | − | − | This paper | PV258682 | |
| South Africa: S of Groenrivier (ABH-76868) | Crespo et al. [12] | OR183478 | This paper | PV258683 | |
| F. sahariensis M.Á.Alonso & al., nom. nov. | Morocco: Guelmim to Tan Tan (MA-786164) | This paper | PV241658 | This paper | PV258684 |
| Morocco: Sidi Ifni (MA-913227) | This paper | PV241659 | This paper | PV258685 | |
| F. salsuginea Adıgüzel & Aytaç2 | Turkyie: Tuz Gölii, salty lagoon (ABH-45933) | Crespo et al. [12] | OR183467 | This paper | PV258687 |
| Turkyie: Dörtyol (MA-561567) | This paper | PV241660 | This paper | PV258686 | |
| F. thymifolia Desf. | Algeria: Bougtob, Chott Cherguí (ABH-59344) | Crespo et al. [12] | OR183481 | This paper | PV258688 |
| Spain: Zaragoza: Bujaraloz (ABH-75454) | Crespo et al. [12] | OR183480 | This paper | PV258689 | |
| F. velutina Brouss. ex DC. | Morocco: Essaouira (ABH-79929) | Crespo et al. [12] | OR183482 | This paper | PV258690 |
1Filed in GenBank as the synonym name R. hypericoides Willd. (nom. illeg.); 2 Filed in GenBank as F. hirsuta (see Crespo et al. [12]).
3. Results
3.1. Taxonomic treatment and description of new taxa
3.1.1. Frankenia sahariensis M.Á.Alonso, M.B.Crespo, Abad-Brotons, Mart.-Azorín & J.L.Villar, nom. nov.
- ≡
- Frankenia florida L.Chevall. in Bull. Herb. Boissier ser. 2, 3(9): 768. 1903 [replaced synonym], nom. illeg. [non Phil. in Anales Univ. Chile 41: 676. 1872] ≡ F. pulverulenta var. florida Maire in Bull. Soc. Hist. Nat. Afrique N. 27: 210. 1936 ≡ F. pulverulenta subsp. florida (Maire) Maire, Cat. Pl. Maroc 4: 1071. 1941. Type: Algeria. [El Menia Province:] El Goléa [currently, El Menia], in arenosis salsis. April 1902, L. Chevallier 404 (lectotype, designated here: P-06618528!, Figure 1; isolectotypes: P-05145114!, P-06618529!, P-06618525!, MPU-007119!, MPU-007120!, US-00679979 [digital image!], GZU-000269792 [digital image!], JE-00003246, JE-00003247 [digital image!], LY-0084391 [digital image!], WAG-0249595 [digital image!], MO-357730 [digital image!]).
- =
- F. intermedia var. annua Caball. in Trab. Mus. Ci. Nat., Ser. Bot. 30: 30. 1935. Type: Morocco. [Western Sahara]: In collibus arenosis insolatis prope Sidi-Ifni, 13 June 1934, A. Caballero (lectotype, designated here: MA-78660!; isolectotype: MPU-300233!).
- −
- F. pulverulenta subsp. floribunda sensu Quézel & Santa, Nouv. Fl. Algérie: 685. 1963 [sphalm.]. Note: The subspecific epithet, “floribunda”, is most likely a mistaken desinence for “florida”, not a formally proposed name.
Description: Herbs annual or short-lived perennial, weakly lignified at base, loosely branched, tap-rooted, and mostly glabrous to sparsely puberulous. Stems non-rooting, usually prostrate to decumbent, 5–30 cm long, often with divaricate branches, with internodes up to 2.5 cm long; young branchlets yellowish to reddish, mostly with scattered whitish depositions, glabrous or shortly and densely puberulous, with minute curled or hooked trichomes (0.1–0.2 mm long) only on one side and near the nodes. Leaves opposite, patent to erect-patent, green or sometimes reddish, and mostly with scattered glands bearing whitish depositions; petiole 0.5–0.8 × 0.2–0.3 mm, flattened, and tapering distally; sheath extending along petiole margins to blade, loosely ciliate to subglabrous, with (2–)4–5 pairs of lateral cilia, unequal in length (0.2–0.5 mm long), whitish, cylindric to flattened, and obtuse or acute at the apex; leaf blade (1.8–)2.5–4 × 0.5–0.7 mm, triangular-ovate to oblong-ovate, mostly subfalcate upwards, with obtuse apex and cordate to rounded base, often strongly revolute on margins or flattened in the lower third, somewhat fleshy and glaucescent, concolorous or sometimes slightly paler abaxially, glabrous on the upper side, loosely hairy beneath with short straight trichomes 0.1–0.2 mm long; midrib notably thickened, tapering slightly towards the apex, continuous with the petiole and raised abaxially, and extending for most of the blade length. Flowers pentamerous, perfect, sessile, solitary on dichotomies or borne in axillary or terminal dichasial groups, usually condensed and glomerular; floral bracts 2, leaf-like but smaller, 2–2.5 × 0.5–0.7 mm, erect to erect-patent, connate at base and shortly covering the calyx base for ca. 0.5 mm, about half the calyx length; bracteoles 2, bract-like but smaller, 0.5–1.5 × 0.4–0.6 mm, alternating with bracts, erect, strongly revolute on margins, about half the calyx length, with petiole ca. 0.5 mm long, adnate to the calyx base. Calyx 2.5–4.2 × 0.6–1 mm, tubular at anthesis to gradually fusiform later, often twisted, straight, indurate, sessile, reddish or rarely yellowish, prominently 5-ribbed, entirely glabrous or densely papillate (with a heterogeneous whitish indumentum of long flattened papillae 0.2–0.3 mm long, globose-claviform papillae ca. 0.1 mm long, and minute globose vesicles), only between the glabrous thickened ribs, and sometimes with scattered whitish depositions; teeth 5, 0.5–1 mm long, erect, triangular, acute, cucullate, shortly mucronate at apex (mucro ca. 0.1–0.2 mm long), narrowly membranous and shortly papillate on margins. Petals 5, 4–6 × 1–1.5 mm, long exceeding the calyx, obovate-cuneate, pinkish-mauve but whitish below, slightly overlapping laterally, exceeding about half to two-thirds the length of calyx; claw 2–2.5 × 0.3–0.4 mm, narrowly cuneate, tapering to blade, yellowish, and partly hidden into the calyx tube; ligule 1.5–2 × 0.2–0.3 mm, narrowly oblanceolate, longitudinally adnate to the claw, the free apex ca. 0.2–0.4 mm long, ovate-oblong, obtuse to subacute, and entire; blade 2.3–3.5 × 1–1.5 mm, broadly obovate-cuneate, rounded at apex, irregularly erose-denticulate. Stamens 6, in two unequal whorls, overtopping ca. 1.5 mm the calyx teeth at anthesis; filaments 3.5–5.5 mm long, whitish, expanded ca. 0.5 mm wide in the lower part but gradually tapering and filiform in the distal part; anthers 0.6–0.8 mm long, yellowish, ellipsoid, versatile. Ovary ca. 1.5 mm long, ellipsoid, subtrigonous, 3-carpelar; placentas 3, parietal-basal, extending up to the lower half to two-thirds of carpel wall length, with ventral traces moderately to profusely branched; ovules 9–12 per placenta, attached along most of the placenta by erect funiculi ca. 0.1 mm long. Style 3–4 mm long, terete, whitish, with 3 stylar branches filiform, 0.5–0.8 mm long, whitish. Capsule 1.4–2 × 0.8–1 mm, ovoid-ellipsoid, subtrigonous, hidden in the calyx tube, early dehiscent. Seeds 24–30 per capsule, 0.5–0.8 × 0.2–0.3 mm, sulcate on one side, ellipsoid, brown, darker at the funicular part, and developing rapidly even before the flower has completely withered; testa thin, not mucilaginous, with a surface weakly and irregularly ornamented with a subrectangular-reticulate pattern, finely striate, covered with small-sized papillae 8–17 µm long, homogeneous in shape, globose to conical-obtuse, sessile, more densely disposed on the distal part.
Figure 1.
Lectotype of Frankenia sahariensis from El Goléa (currently El Menia), Algeria, L. Chevallier 404 (P-06618528). Scale bar = 1 cm (on the small upper label). © Muséum National d’Histoire Naturelle, herbarium collections, Paris (reproduced with permission).
Figure 1.
Lectotype of Frankenia sahariensis from El Goléa (currently El Menia), Algeria, L. Chevallier 404 (P-06618528). Scale bar = 1 cm (on the small upper label). © Muséum National d’Histoire Naturelle, herbarium collections, Paris (reproduced with permission).

Etymology: The specific epithet (sahariensis, –e) refers to the Sahara Desert, the native area of the species. The name replaces the illegitimate “F. florida L.Chevall.”, a later homonym of the Chilean “F. florida Phil.” (see below). The type of the former was collected in 1902 and distributed by Louis P.D. Chevallier with number 404 of his exsiccata Plantae Saharae Algeriensis.
Phenology: Flowering in early January–early June (occasionally in August–September), fruiting in February–Juny (occasionally in September–October).
Habitat and distribution: Frankenia sahariensis is found in mostly sandy soils, in subsaline depressions often along ravines and wadies, from 0–1300 m elevation. It is endemic to the western parts of the Sahara Desert [55], south to the High Atlas Range, spreading from the Atlantic coast of northern Mauritania and southern Morocco to central and eastern Algeria and northwestern Mali (Figure 2). Biogeographically, and according to the current data, it is endemic to the western parts of the Saharan province of the Saharo-Arabian region of Takhtajan [34]. In this vast territory, F. sahariensis occurs in halophilous vegetation types within the Oceanic Saharan, Western Saharan, Northwestern Saharan, and Steppe-Northern African domains of Quézel [33].
Nomenclatural remarks: Chevallier [56] first described this remarkable plant after his collections near El Goléa (currently El Menia), in the Saharan region of southern Algeria. It was named “F. florida”, most probably due to the outstanding showy flowers it exhibits at anthesis. However, the earlier homonym F. florida Phil. had already been used for a Chilean congener [57], a fact that made Chevallier’s name illegitimate (Art. 53.1 of the ICN) and unavailable for use. Nonetheless, since the Saharan plant somewhat resembles F. pulverulenta (see below), it has often been subordinated to the latter at different infraspecific ranks. First, Maire [58] accepted it as a variety, and indirectly validated the name F. pulverulenta var. florida Maire (Art. 58.1 of the ICN). Later, this basionym was treated by Maire [24] at subspecies rank, a currently broadly accepted combination (see [4,59]). Nonetheless, when treated at specific rank no previous valid names are available for the Saharan plant, and therefore a replacing name, F. sahariensis nom. nov. (Art. 6.11 of the ICN), is proposed here for the illegitimate F. florida L.Chevall.
Other studied material: Algeria (Dza). Béni Abbès Province: Sahara, Moyenne vallée de la Saoura, June 1941, Volkonsky (MPU-300246!). Sahara: Embouchure de la Saoura, May 1926, Balachowsky (MPU-300247!). Djanet Province: Sahara Central, Tassili N’Ajjer, lieux humides à Ahrème, en aval d’Ihérir, 7 January 1966, ? (P-05145224!). El Menia Province: Loc d’El Goléa [currently, El Menia], vases salées, April 1954, P. Quézel (MPU-300243!). Naâma Province: Sfissifa, 3 June 1887, A. Bousquet (MPU-048475!). Aïn-Sefra, 21 April 1888, E. Bonnet & P. Maury (P-06801050!). Tindouf Province: Sahara Occidentale: Oued Jehach entre Tindouf et l’Oued Drâa, lieux sableux, printemps 1938, Ollivier 187 (MPU-048470!). Sahara Occidentale, Tindouf, assez abondant, March 1928, Dr. Estival (MPU-048483!). Sahara Occidentale, Region de Chenachane, Chenachane, ? 1923, Dr. Tripeau (MPU-300238!). Sahara Occidentale, Nkheila, jardin, 29 March 1995, Monod 19676 (P-05038753!). Ibidem, zone d’épandage, Monod 19678 (P-05038755!). Ibidem, Monod 19679 (P-05038747!). Sahara Occidentale, Entre Nkheila et Rabouni, 30 March 1995, Monod 19703 (P-05038750!). Mali. Tombouctou Region: El Mzereb (El Hank), dans un terrain sale, 20 April 1954, J. Sougy 392 (P-00799803!). Mauritania. Dakhlet Nouadhibou Province: Ténaloul (Ten Alloui), sebkhas du littorale, 22 April 1938, Murat 2390 (P-05038740!, MPU-300262!). Iouik [Iwik], côte de Mauritania, 9 April 1982, Th. Monod 18336 (P-05038748!). Tiris Zemmour Province: Soudan Français, Chegga-Guelta, 3 January 19[38], [O. de] Puygaudeau (P-05038749!). Sahara Occidental, Falaise du Hank, ravin d’Aïn-Chegga, Janvier 1939, Dr. Rolland 106 (MPU-048484!). Morocco. Dakhla-Oued Ed Dahab Region: Dakhla, río de Oro, 28 May 1977, Th. Monod 16192 (P-05144446!). Drâa-Tafilalet Region: Daoura, dans une dava, sur la Hammada de la Daoura, 2 hours vers la vallée vers Hachi-Chamba, 9 April ?, Dr. Le Carbout (MPU-300240!). Daoura, dans la vallée de la Daoura vers Achi-Chamba, 17 May ?, Dr. Le Carbout (MPU-300241!). Entre Ouarzazate et Skoura, 25 March 1932, F. Peltier (MPU-048495!). Drâa: graviers de l’Oued Imini à Ouarzazate, 14 February 1936, Gattefossé (MPU-967102!). Guelmim-Oued Noun Region: Oued Noun, Ollivier 14 (MPU-300264). In aridis subsalsis inter Labyar et Notfia, 1 April 1937, R. Maire (MPU-300242!). Anti-Altas, Goulimine (Guelmin), 23−28 August 1936, M. Langueron (MPU-300234!). Prov. Guelmin 53 km from Guelmin on road to Tan-Tan, near café, 7 February 2007, S. Jury & T.M. Upson 20492 (MA-786164!). Prov. Guelmime 14 km WSW von Guelmime an der Sttraβe (P41) Nach Tan-Tan, ca. 200 m, 28º56′N 10º08′W, 15 April 1997, D. Podlech 53647 (W-0365982!, RSA-0548173 [digital image!]). Entre Goulimine (Guelmin) et El Aioun du Drâa, 6 September 1941, J. de Lepiney, Ch. Runge & Ch. Sauvage 2035 (MPU-759265!). Laâyoune-Sakia El Hamra Region: Sahara Occidental: El Aaiun (Seguia el-Hamra), Izik [Mision d’Études de la Biologie des Acridiens 2482] (MPU-300239!). Sahara Occidentale : Zemmour [Gueltat-Zemmour], dans les regs près de la Guelta, March-April 1934, Luthereau (MPU-300237!). Souss-Massa Region: In planitie at septentr. urbis Tiznit, 13 April 1934, R. Maire & E. Wilczek (MPU-300236!). Prov. Tiznit, beach West of Gourizime, near the road from Tiznit to Sidi-Ifni, 20 m, 29º37′N 10º02′W, 14 April 1997, D. Podlech 53623 (W-0365981!). Prov. Tiznit, Oued Massa ca. 40 km Tiznit, 50 m, 3 April 1994, A. Tribsch (W-0365983!). Souss-Massa-Drâa, Tiznit, Ifni Mesti, sur del puerto de Sidi Ifni, 24 March 2015, D. Gutiérrez et al. DG503 (MA-00913227!). Prov. Agadir, 10 km NW Agadir, at the coast, 120 m, 30º29′26”N 9º40′17”W, 19 June 1996, A. Achhal et al. 96-0511 (W-0365985!). In depressis subsalsis ad Tigert, prope Herculis Promontorium (Cap Ghir), 4 April 1937, R. Maire (MPU-300235!). Sud-Ouest du Maroc, Imeoghguemmi [sic], ? 1875, Mardochée (MPU-750398!). In arvis prope Tasila, ad ostium fluminis Massa, 3 April 1937, R. Maire (MPU-300251!). In arenosis ditionis Tazeroualt, in faucibus Sidi-el-Ghiat, 400 m, 12 Abril 1934, R. Maire & E. Wilczek (MPU-300263!).
3.1.2. Frankenia dinteri M.Á.Alonso, M.B.Crespo, Abad-Brotons, Mart.-Azorín & J.L.Villar, nom. nov.
- ≡
- Frankenia densa Pohnert in Mitt. Bot. Staatssamml. München 1(9-10): 447. 1954. [replaced synonym], nom. illeg. [non Summerh. in J. Linn. Soc., Bot. 48: 373 (1930)]. Type (see Pohnert 1954: 448): Namibia. [Otjozondjupa Region:] Grootfontein-Süd, 22 November 1934, K. Dinter 8059 (holotype: M-0104483 [digital image!, available at https://plants.jstor.org/stable/viewer/10.5555/al.ap.specimen.m0104483]; isotypes: M-0104484 [digital image!], PRE-0293175-0 [digital image!], PRE-526699-0 [digital image!] HBG-516900 [digital image!], BOL-136934!, K!). Paratypes: Namibia. [Karas Region:] Bethanien [Bethany], 24 December 1934, K. Dinter 8270 (BOL-136934!, HBG-516901 [digital image!], K!). Figure 3.
Description: Herbs annual or short-lived perennial, often robust and weakly lignified at base, profusely and densely branched, tap-rooted, mostly glabrous to sparsely puberulous. Stems non-rooting, usually erect to ascendant, 5–30 cm long, often with divaricate branches, with internodes up to 2.5 cm long; young branchlets yellowish-brown to reddish, mostly with scattered whitish depositions, glabrous or shortly and loosely puberulous, with minute curled to hooked trichomes (0.1–0.2 mm long), only on one side. Leaves opposite, patent to erect-patent, green or sometimes reddish, and mostly with scattered glands bearing whitish depositions; petiole 0.8–1.2 × 0.5–0.6 mm, flattened, and tapering distally, sometimes reddish; sheath extending along petiole margins to blade, ciliate with 4–6 pairs of lateral cilia, unequal in length (up to 0.4–0.5 mm long), whitish, cylindric to flattened, and obtuse or acute at apex; leaf blade (2.5–)3–4.5(–6) × 0.5–2 mm, narrowly elliptic to oblong, mostly subfalcate upwards, with obtuse apex and ± rounded base, often strongly revolute on margins or flattened in the lower third, somewhat fleshy and glaucescent, concolorous or occasionally somewhat paler abaxially, glabrous on the upper side with scattered whitish depositions and waxes, and glabrous or loosely hairy beneath with scattered short straight trichomes 0.1–0.2 mm long; midrib narrow, linear, tapering slightly towards apex, continuous with petiole below, somewhat raised abaxially, and extending for most of the blade length. Flowers pentamerous, perfect, sessile, often crowded in dense axillary or terminal dichasial groups, usually densely condensed and glomerular, many-flowered; floral bracts 2, leaf-like, 3–4 × 0.6–0.7 mm, erect to erect-patent, connate at base and shortly covering the calyx base for ca. 0.5 mm, about equalling or slightly exceeding the calyx length; bracteoles 2, bract-like but smaller, 1–2 × 0.4–1 mm, alternating with bracts, erect, strongly revolute on margins, about half to two-thirds the calyx length, with petiole ca. 1 mm long, adnate to calyx base. Calyx 3.3–4.7 × 0.6–1 mm, tubular at the anthesis to gradually fusiform later, non-twisted, straight, indurate, often reddish, prominently 5-ribbed, entirely glabrous or loosely puberulous (with a homogeneous whitish indumentum of minute straight papillae ca. 0.1 mm long) between the thickened glabrous ribs, often with scattered whitish depositions; teeth 5, ca. 0.5 mm long, erect, triangular, acute, cucullate, shortly mucronate at apex (mucro ca. 0.4 mm long), narrowly membranous and shortly papillate on margins. Petals 5, 3.5–5 × 0.5–0.8 mm, exceeding the calyx, obovate-cuneate, pinkish-mauve but whitish below, often broadly overlapping laterally, exceeding about one-third to half the calyx length; claw 2–2.5 × 0.3–0.4 mm, narrowly cuneate, tapering to blade, yellowish, and entirely hidden into the calyx tube; ligule 2–2.5 × 0.4–0.5 mm, narrowly oblanceolate, longitudinally adnate to the claw, the free apex ca. 1 × 0.5 mm, ovate-oblong, broader than the claw at base, obtuse to subacute, and entire; blade 2–2.5 × 0.5–0.8 mm, obovate, rounded at apex, irregularly erose-denticulate. Stamens 6, in two unequal whorls, hidden in calyx teeth at anthesis; filaments 2.3–2.5 mm long, whitish, expanded ca. 0.5 mm wide in the lower part but gradually tapering and filiform in the distal part; anthers 0.6–0.8 mm long, yellowish, ellipsoid, versatile. Ovary ca. 2 mm long, elliptic-ovoid, subtrigonous, 3-carpelar; placentas 3, parietal-basal, extending up to the lower half to two thirds of carpel wall length, with ventral traces moderately to profusely branched; ovules 10–12 per placenta, attached along most of the placenta by erect funiculi ca. 0.1 mm long. Style 1.7–2 mm long, terete, whitish, with 3 stylar branches filiform, 0.5–0.6 mm long, whitish. Capsule 3–4.5 × 1.4–2 mm, ovoid-ellipsoid, subtrigonous, hidden in the calyx tube, early dehiscent. Seeds 25–30 per capsule, 0.5–0.6 × 0.2–0.3 mm, sulcate on one side, ellipsoid, light brown, darker at the funicular part, and developing rapidly even before the flower has completely withered; testa thin, not mucilaginous, with a surface weakly and irregularly ornamented with a subrectangular-reticulate pattern, finely striate, covered with scattered small-sized papillae 7–12 µm long, homogeneous in shape, conical-obtuse, sessile, more densely disposed on the distal part.
Eponymy: The specific epithet honours Moritz Kurt Dinter (1868–1945), a renowned German botanist who extensively collected plants in south-western Africa, mostly in Namibia, and particularly he gathered in 1934 the specimens upon which Pohnert [60] first described his “Frankenia densa”, an illegitimate later homonym of the Australian F. densa Summerh. (see below).
Phenology: Flowering in late April–early October (occasionally in November–January), fruiting in May–November (occasionally in December–February).
Habitat and distribution: Frankenia dinteri occurs on saline, sandy to clayish soils in wetlands, ravines, or close to lagoons, at elevations of 300–1225 m. The known distribution of the new species extends from Grootfontein (Otjozondjupa Region, Northern Namibia), the type locality, to Goageb (Karas Region, southern Namibia), but spreads southwards into northwestern South Africa to Salt River, north of Vanrhynsdorp (Western Cape Province) and far eastwards to Bloemhof (North West Province) (Figure 4). Biogeographically, and according to the current data, it is endemic to the Namib, Namaland, and Karro Provinces of the Karoo-Namib Region (sensu Takhtajan [34]). In that large territory, F. dinteri is found in halophilous vegetation-types occurring in the Nama-Karoo (NK), Succulent Karoo (SK), Desert (D), and Savanna (SV) Biomes [32].
Nomenclatural remarks: This outstanding southern African sea-heath was first described by Pohnert [60] as “Frankenia densa” from materials collected in inland Namibia which included robust, profusely branched plants with many-flowered and densely crowded inflorescences. However, the name F. densa had already been published by Summerhayes [34] for a sea-heath from central and southern Australia, which made Pohnert’s name illegitimate (Art. 53.1 of the ICN). Apparently, no valid name exists at specific rank for the southern African plant, and hence a replacing name, F. dinteri nom. nov. (Art. 6.11 of the ICN), is proposed here for the illegitimate F. densa Pohnert.
Other studied material: Namibia. Karas Region: Goageb, near bridge on B4 along dry river bed, 26º44′54”S 17º13′13”E, 922 m elevation, 10 August 2016, M. Mart.-Azorín et al. (ABH-76804!). Warmbad, im rivierbett, 22 May 1972, W. Giess & M. Müller 12120 (WAG-0249597 [digital image!]). Seeheim, 17 November 1922, K. Dinter 4214 (GH-02434737 [digital image!]). Reg. Gross-Namaland, Ganas, 12 December 1884, H. Schinz 267 (Z-000067508 [digital image!]. Southwest Africa: Kahanstal, 2 December 1934, K. Dinter 8154 (BOL!, K!). South Africa. Northern Cape Province: 2816 (Oranjemund): Sendelingsdrif, stagnant pool along Orange River near Octha Mine (-BB), 25 August 1982, W. Metelerkamp 400 (BOL!); Little Namaqualand: Alties River bed, 1 December 1910, N.S. Pillans 5381 (BOL!); Little Namaqualand: Dry sand near stream Kuboos (-BD), October 1926, N.S. Pillans 5409 (BOL!); Little Namaqualand: Mouth of Orange River (-CB), on dry mud, lagoon, October 1926, N.S. Pillans 5576 (BOL!); Little Namaqualand: Mouth of Orange River (-CB), sandy banks, October 1926, N.S. Pillans 5583 (BOL!); Namaqualand, Richtersveld Nat. Park, S. Avenants Grove, January 1995, G. & W. Williamson 5590 (NBG-153968!). 2818 (Warmbad) Regionis occidentalis: Henkries, Orange River (-CC), 30 November 1897, M. Schlechter 22 (BOL!). 2819 (Ariamsvlei): Daberas Farm (-BD), 28º29′32”S 19º56′38”E, 573 m, 23 August 2022, M. Mart.-Azorín, M.B. Crespo, M.Á. Alonso, J.L. Villar & M. Pinter (ABH-83264!); Onseepkans, near Orange River (-CD), 28º46′37”S 19º15′47”E, 359 m, 21 August 2022, M. Mart.-Azorín, M.B. Crespo, M.Á. Alonso, J.L. Villar & M. Pinter (ABH-83234!); Kenhardt Distr.: Farm Skroef, bank of Orange River (-DA), 26 September 1987, E.V. Hoepen 1887 (BOL!, PRE!). 2820 (Kakamas): Augrabies, (falls) near Park (-CB), 28º39′02”S 20º21′55”E, 616 m, 23 August 2022 M. Mart.-Azorín, M.B. Crespo, M.Á. Alonso, J.L. Villar & M. Pinter (ABH-83265!). 2917 (Springbok): Namabeep, dry river bed (-DB), 26 March 1964, D.S. Hardy 1682 (K!). 3020 (Brandvlei): Between Brandvlei and Williston, Karreekop farm, sandy plains with scattered rocks (-DC), 30º58′59”S 20º39′18”E, 987 m, 24 August 2022, M. Mart.-Azorín, M.B. Crespo, M.Á. Alonso, J.L. Villar & M. Pinter (ABH-83275!). North West Province: 2725 (Bloemhof): [Between Christiana and Bloemhof], Vaal River Salt works, saltpan (-CB), 27º44′S 25º20′E, 29 April 1973, B. Straschil (W-0365984). Western Cape Province: 3118 (Vanrhynsdorp): 18 miles N of Vanrhynsdorp, Salt River, 22 January 1930, C.E. Moss 17983 (BM!).
3.2. Phylogenetic relationships
The aligned ITS dataset was 720 bp, 249 of which (34.58%) were potentially parsimony informative. The phylogenetic relationships of taxa are shown in Figure 5 as recovered in our ML tree. The percentage of trees in which the associated taxa clustered together (after 10,000 bootstrap replicates) in the ML analysis are shown above branches, while PP values from the BI analysis are shown below branches. Trees obtained with MP and NJ methods (Figure S1A,B) yielded similar topologies to the ML tree. In the MP analysis, eight most parsimonious trees were obtained with a tree length (TL) of 581 steps, a consistency index (CI) of 0.796, and a retention index (RI) of 0.913.
The general topology of the obtained ITS tree (Figure 5) is almost identical to that reported by Crespo et al. [12], which was based on a more reduced set of species and samples, though it also yields a strongly monophyletic genus Frankenia (99BP, 1.00PP), when compared with outgroups of the sister family Tamaricaceae (Tamarix, Myricaria, and Reaumuria). The addition of samples of F. sahariensis and F. dinteri, as well as other congeners from Eurasia and Africa (including Macaronesia), revealed new internal relationships (both within and between clades).
On the one hand, a strongly supported (95BP, 1.00PP) broad aggregate is found that comprises three unresolved major lineages (clades A+B+C) matching those of Crespo et al. [12]. Firstly, Clade A is composed in the ML analysis by two groups of taxa. The first group encompasses all samples of F. dinteri from Namibia and South Africa, forming a strongly supported (99BP, 1.00PP) clade that is weakly supported (53BP) as sister of the remaining groups within Clade A, but is placed in the BI analysis in an unresolved position together with the broad aggregate of clades [A+[B+C]]. The second group is formed by a well-supported (70BP, 1.00PP) broad lineage including the South African samples of the strongly supported (96BP, 1.00PP) “F. repens group” (sensu Crespo et al. [12]) with F. anneliseae M.B.Crespo & al., F. repens (P.J.Bergius) Fourc. and F. nummularia M.B.Crespo & al., to which clades of two North African-Iberian species, F. boissieri Reuter and F. thymifolia Desf., are strongly supported successive sisters. Secondly, samples of the SW Mediterranean F. corymbosa Desf. (Clade C) are arranged in a strongly supported (99BP, 1.00PP) compact group, which is nested only in the BI analysis as a weakly supported (0.74PP) sister to Clade B. Thirdly, Clade B is formed by two major, moderately supported lineages. On one side, Subclade B1 (75BP, 0.78PP) mostly comprises members of the major Macaronesian archipelagos (i.e., the Madeiran F. cespitosa Lowe, the Canarian “F. ericifolia group”, and the Cape Verdean F. pseudoericifolia Rivas Mart. & al.) plus two western Saharan species (i.e., F. ifiniensis and the newly named F. sahariensis), which are arranged into three unresolved lineages. In one of those lineages, all samples of “F. ericifolia group” are strongly supported (93BP, 0.99PP) as sister to F. sahariensis samples. On the other side, Subclade B2 (99BP, 1.00PP) merges an apparently heterogeneous aggregate of species with western Mediterranean-Atlantic (F. laevis), North African-Atlantic (F. composita and F. velutina Brouss. ex DC.), or currently subcosmopolitan (F. pulverulenta) distributions. Among them, all samples of F. pulverulenta form in the ML analysis a weakly supported (53BP) clade.
On the other hand, the “F. hirsuta group” (sensu Crespo et al. [12]) consolidates as Clade D, a strongly supported (99BP, 1.00PP) group formed by all samples of F. hirsuta s.str. (E Mediterranean) plus F. salsuginea (central Anatolia), which is sister to the aggregate of clades A-C with a strong support (95BP, 1.00PP).
Finally, the South African narrow endemic F. fruticosa remains as sister (59BP, 0.99PP) to all other studied members of Frankenia.
The aligned matK dataset was 829 bp, 124 of which (14,95%) were potentially parsimony informative. The phylogenetic relationships of taxa are shown in Figure 6 as recovered in our ML tree. As said before, the percentage of trees in which the associated taxa clustered together (after 10,000 bootstrap replicates) in the ML analysis are shown above branches, whereas PP values from the BI analysis are shown below branches. Trees obtained with MP and NJ methods (Figure S1C,D) yielded similar topologies to ML tree. In the MP analysis, nine most parsimonious trees were obtained with a tree length (TL) of 226 steps, a consistency index (CI) of 0.848, and a retention index (RI) of 0.929.
In the obtained matK tree (Figure 6), Frankenia is also recovered as a strongly monophyletic (100BP, 1.00PP) clade, with two main lineages corresponding to clades [[A+D]+[B+C]] of the ITS tree, though displaying a different scenario of interspecific relationships. The recovered main clades are partly coincident with those shown in the ITS tree, and their codes are retained in the matK tree for parallelism. However, some of those major clades show lower internal resolution than in the ITS tree. No useful sequences were generated for the South African F. fruticosa.
On the one hand, the first lineage is strongly supported (97BP, 0.99PP) and encompasses Clade A (89BP, 0.91PP), with the South African members of the “F. repens group” not full resolved internally, plus Clade D (92BP, 1.00PP), with the “F. hirsuta group” including the eastern Mediterranean (F. hirsuta) and Anatolian (F. saslsuginea).
On the other hand, clades B+C form a lineage that is sister to Clade A+D (78BP, 0.97PP) and is fully unresolved internally. Firstly, a heterogeneous moderately supported (77BP, 1.00PP) aggregate (Clade B1+B2) is found that merges species from Clades A, B1, and B2 of the ITS tree. In this group, the northwestern African coastal species F. velutina is found, only in the ML analysis, as weakly supported (55BP) sister of the remaining members. Provenances of those members are diverse, such as F. ericifolia (Canary Islands), F. sahariensis (western Sahara Desert), F. laevis and F. composita (Mediterranean basin and Atlantic coasts of Northern Africa and Europe), and F. dinteri (southern Africa). Although internal relationships among those taxa are not well resolved, three subclades arise in Clade B1+B2 that respectively include all samples of F. sahariensis (64BP, 98PP), all samples of F. laevis (85BP, 1.00PP), and some samples of F. ericifolia (86BP, 1.00PP). Samples of F. dinteri, which formed in the ITS tree a strongly supported lineage in Clade A, are unresolved along with members of Clades B1 and B2. Secondly, the Canarian F. capitata, the western Saharan F. ifniensis Caball., and the SW Mediterranean and N African F. thymifolia, constitute Clade B1 (64BP, 0.99PP), whose internal relationships are not well resolved. However, an internally unresolved subclade with F. capitata plus F. ifniensis is found in both analyses conducted that is sister (59BP, 0.96PP) to samples of F. thymifolia that in contrast nested with members of Clade A in the ITS tree. Thirdly, Clade B2 shows a moderate support (62BP, 1.00PP) and encompasses all samples of F. pulverulenta (Eurasian-Mediterranean species that became subcosmopolitan) plus the sample of F. boissieri (SW Mediterranean and African-Atlantic species), without full internal resolution. In contrast, the BI analysis recovered all samples of F. pulverulenta in a strongly supported (1.00PP) clade. Members of Clade B2 were included in the homonymous one in the ITS tree, excepting F. boissieri that was recovered in the ITS analysis as member of Clade A. Finally, the SW Mediterranean F. corymbosa is nested in a strongly supported (96BP, 1.00PP) distinct Clade C, which only in the BI analysis is recovered as sister (0.75PP) to the former aggregate.
4. Discussion
The findings presented here are largely consistent with those reported by Crespo et al. [12] and form part of a broader study evaluating the generic and specific relationships within Frankeniaceae, with a particular focus on the Eurasian and African taxa.
Both obtained ITS and matK trees (Figure 5 and Figure 6) represent the most comprehensive phylogenetic trees of Frankenia to date. While still incomplete, they provide a solid foundation for a preliminary phylogenetic survey on the genus in Eurasia and Africa, which will be expanded in the coming years. When combined with morphological data and field observations, these molecular trees provide a clear picture, allowing for a more accurate interpretation of certain African taxa that are often synonymised or subordinated to the annual sea-heath, F. pulverulenta (Table 3). This is exemplified by the two species that have been newly named here, F. sahariensis and F. dinteri, which are the principal focus of the present work.
Frankenia pulverulenta is native to Eurasia (including the Mediterranean basin) and has become Subcosmopolitan after introduction worldwide [14]. The species is currently distributed in temperate and subtropical regions, ranging from sea level to nearly 2000 m elevation, which thrives in both very cold regions and hot deserts. According to the taxonomy currently adopted in some general databases (e.g. [4,26,59]), it is the only annual species in the genus, which occurs mostly in regions with low precipitation worldwide and grows optimally in soils with a certain concentration of salts (i.e., chlorides, sulphates, or nitrogen compounds), often in frequented and disturbed areas [61]. For that reason, it is usually regarded as a halo-nitrophilous or gypsum-loving herb [8], with a high degree of morphological plasticity. This lifestyle might be connected to the broad variation it apparently exhibits in terms of body size and indumentum [62]. In addition to its annual cycle, F. pulverulenta shows distinctive morphological characteristics that facilitate identification, including nearly flattened leaves that are cuneate at the base and notched at the apex, solitary scattered flowers, and capsules containing numerous small seeds (Table 3). That presumed morphological polymorphism has led some authors to synonymise F. pulverulenta to other congeners with broad, flattened leaves but strongly differing on many other respects. Recently, Crespo et al. [12] have studied two South African species, F. nodiflora and F. nummularia, long mistaken within the broadly circumscribed F. pulverulenta, which widely parallel the present case.
On the one hand, samples of F. sahariensis collected in the western Saharan regions are nested far apart from those of the typical F. pulverulenta in all analyses conducted. The position in the ITS tree (Figure 5) of F. sahariensis as a strongly supported sister of the Canarian F. ericifolia clade is consistent with the geographic distribution of both species, occurring about the same latitude in the Atlantic regions of NW Africa and the eastern Canary Islands, not far from each other. In contrast, the matK tree (Figure 6) yields both latter species in a wide clade with poor internal resolution, which also comprises several morphologically diverse species. Among them, only samples of F. sahariensis, F. laevis and partly F. ericifolia nest individually in well supported clades, whereas positions of the remaining (F. composita, F. velutina, and F. dinteri) are not resolved. All those entities exhibit morphological differences that allow easy distinction at different taxonomic ranks [22]. Conversely, all samples of F. pulverulenta (coming from diverse distant places, such as the Mediterranean basin, the Canary Islands, and South Africa) are found together in a heterogeneous weakly supported clade in the ITS tree, which is close to F. laevis, F. velutina and F. composita, a group of perennial mostly Atlantic and southwestern Mediterranean taxa sometimes accepted at subspecific rank in a broadly circumscribed F. laevis [22,26]. Furthermore, in the matK tree F. pulverulenta nests together with F. boissieri, but without full internal resolution. Nevertheless, all those phylogenetic relationships are apparently not correlated to sound morphological patterns and can be best interpreted on biogeographical grounds, as discussed below.
In the protologue of F. sahariensis (as F. florida L.Chevall., non Phil.), Chevallier [56] (1903) related his new species to the highly variable subcosmopolitan F. pulverulenta. In fact, it is often subordinated to the latter as a variety [58] or subspecies [24]. However, sound morphological characteristics allow confident distinction of both species (Table 3):
- i)
- blade of leaves triangular-ovate to oblong-ovate, with cordate to rounded base and a prominent thickened midrib on the underside of blade in F. sahariensis vs. broadly obovate blade with cuneate base and thin midrib not remarkably prominent in F. pulverulenta;
- ii)
- calyx indument heterogeneous, with flattened trichomes between ribs intermingled with small globose-claviform trichomes and minute papillae in F. sahariensis vs. calyx with homogeneous indument of small curled trichomes also present on the underside of leaves in F. pulverulenta;
- iii)
- petals much exceeding about half to two-thirds the calyx length (vs. petals exceeding only about one-third to half the calyx length in F. pulverulenta); and
- iv)
- seeds morphologically close in both species (Figure 7), though less abundant in F. sahariensis (24–30 per capsule) than in F. pulverulenta (up to 45 per capsule).
All these features make individuals of F. sahariensis quite distinctive during anthesis in the field. Both species exhibit significant overlap in their distributions and co-occur at least in the coastal areas, such as the Ifni region of western Sahara. From this area, Caballero [63] described F. intermedia var. annua Caball. (MA-78660!), which is now considered a synonym of F. sahariensis, as well as F. pulverulenta var. grandifolia Caball. (MA-78705!), a plant with larger leaves and longer petioles that is indistinguishable from and should be synonymised to the typical F. pulverulenta, as suggested by Maire [24] and Nègre [22].
Chevallier [56] also noted the annual or short-lived perennial habit of F. sahariensis (as F. florida), stressing its resemblance to F. laevis and F. intermedia (F. laevis var. intermedia (DC.) Bonnet), two entities predominantly occurring along the Mediterranean and Atlantic coasts of Europe and North Africa [19]. However, both latter taxa are woody perennial, shrubby species with prostrate to ascendant stems with denser indumentum all around; leaves are strongly revolute, not cordate at base, often densely pubescent to tomentose beneath [22]; and the calyx indument in both species is always lacking globose-claviform trichomes between ribs. Furthermore, calyx ribs in F. intermedia exhibit long cilia [64]. Morphological relationships are weaker to other relatives occurring in northwestern Africa, such as F. velutina, F. boissieri or F. composita (see Nègre [22]). Nevertheless, some connections can be argued with the Canarian F. ericifolia, based on the shared prostrate habit, leaves sometimes flattened and geographical proximity, which might support the phylogenetic closeness unveiled by the obtained trees (Figure 5 and Figure 6). Despite those similarities, F. ericifolia differs significantly from F. sahariensis by its shrubby woody habit, the mostly revolute leaves covered with short hairs all over, never cordate at the base, with a weaker non-thickened midrib; and the homogeneous calyx indument with short conical trichomes. For all those reasons, F. sahariensis can be readily distinguished from F. pulverulenta and other relatives, therefore deserving recognition at species rank as previously suggested by Ozenda [55, as F. florida L.Chevall.].
On the other hand, all samples of F. dinteri collected from a substantial territory encompassing southern Namibia and northwestern South Africa are distinctly separated from those of the typical F. pulverulenta in all analyses conducted. Regarding the ITS tree (Figure 5), F. dinteri forms a strongly supported group in Clade A that is weakly related to the remaining South African studied species of the “F. repens group”. In contrast, the matK tree (Figure 6) recovers all samples of F. dinteri not coalescent into a single clade but rather dispersed across Clade D, in which several subshrubby species with diverse origins (F. sahariensis, F. laevis, F. ericifolia, F. composita, and F. velutina) nest together without complete internal resolution. Once again, phylogenetic relationships of F. dinteri appear to respond best to biogeographical patterns than to morphology (see below).
Figure 7.
Seed morphology (left) and testa details (right) of: (A, B) Frankenia sahariensis (P-05038748); (C, D) F. dinteri (ABH-76891); (E, F) F. pulverulenta (ABH-7952). Scale bars = 100 µm (A, C, E), 10 µm (B, D, F).
Figure 7.
Seed morphology (left) and testa details (right) of: (A, B) Frankenia sahariensis (P-05038748); (C, D) F. dinteri (ABH-76891); (E, F) F. pulverulenta (ABH-7952). Scale bars = 100 µm (A, C, E), 10 µm (B, D, F).

In the protologue of F. dinteri, Pohnert [60, as F. densa] hypothesised that his newly described species exhibited a close resemblance to F. laevis, although also displayed intermediate characteristics with F. pulverulenta. However, F. laevis is not known to occur in sub-Saharan Africa, and shows quite different morphological features, such as the prostrate shrubby stems (vs. annual herbaceous or slightly suffrutescent at base in F. dinteri) with leaves non-fleshy, strongly revolute and densely pubescent to tomentose beneath (vs. leaves fleshy, often flattened at base and glabrous to subglabrous beneath in F. dinteri); flowers axillary and scattered along branches (vs. flowers often densely crowded in terminal cymes in F. dinteri); and bracteoles about equalling the calyx length (vs. bracteoles about half the calyx length in F. dinteri). Furthermore, the original description also presented F. densa as a shrublet, though a detailed observation demonstrated that it is a robust annual or short-living perennial herb, only superficially resembling F. pulverulenta. On that basis, both names were likely synonymised by Obermeyer [62], a treatment which was adopted by subsequent authors so far (cf. [65,66]). However, sound morphological differences exist apart from the habit that allow easy separation (Table 3) of both latter entities:
- i)
- flowers often densely crowded in many-flowered cymes in F. dinteri vs. axillary, solitary or in lax cymes in F. pulverulenta) (Figure 3A,B);
- ii)
- floral bracts about half to two-thirds the calyx length vs. equalling to slightly exceeding the calyx length in F. pulverulenta;
- iii)
- bracteoles 1–2 mm long vs. 2.5–4 mm in F. pulverulenta (Figure 3C,D);
- iv)
- calyx teeth mucro ca. 0.4 mm long vs. ca. 0.1 mm in F. pulverulenta (Figure 3D);
- v)
- anthers 0.6–0.8 mm long vs. 0.2–0.4 mm long;
- vi)
- capsule 3–4.5 mm long vs. 2–3 mm in F. pulverulenta (Table 3); and
- vii)
- seeds morphologically close in both species (Figure 7), though less abundant in F. dinteri (25–30 per capsule) than in F. pulverulenta (up to 45 per capsule).
As mentioned before, and according to our data, both species overlap their distributions and are sympatric at least in the south of its distribution area (i.e. the Karro region of South Africa). However, the morphological patterns separating both entities remain invariable and warrant confident distinction, which is congruent with their remote position in all our trees (Figure 5 and Figure 6). Resemblances to other congeners are weak and probably due to convergence.
Finally, in this context it is worth mentioning the case of F. cespitosa Lowe, a name that is sometimes synonymised with F. pulverulenta [4] or also with F. laevis L. [26]. Frankenia cespitosa was harvested in the islands of Madeira (at Ponta S. Lourenço) and Porto Santo, both from the Archipelago of Madeira (Portugal), and was described by Lowe [67] as a morphologically close relative of F. ericifolia and F. corymbosa. A thorough examination of the original material (K!, BM!, P!) reveals that the Madeiran F. cespitosa is unmistakably a woody perennial species, with leaves linear, strongly revolute and densely covered with whitish salt crusts (as observed on the stems), flowers in dense terminal crowded inflorescences, and calyx strongly twisted, among other features. All these characters separate the Madeiran plant from both F. pulverulenta and F. laevis, the latter producing leaves lacking extensive salt crusts, flowers axillary and solitary along branchlets, and calyx untwisted. In contrast, the morphological features of F. cespitosa closely resemble those of certain species from Macaronesia and northwestern Africa, as suggested in the protologue. In fact, the obtained ITS tree confirms such inferred morphological connections, placing the Madeiran F. cespitosa far apart from F. laevis, as a strongly supported sister to an unresolved clade comprising F. capitata Webb & Berthel. (Canary Islands) plus F. ifniensis Caball. (Atlantic coast of northwestern Africa), occasionally treated as F. corymbosa var. ifniensis (Caball.) Maire, with which it shares its gross morphology as well. Furthermore, they all are connected to the F. ericifolia-F. sahariensis clade (Canary Islands-northwestern Africa) plus F. pseudoericifolia (Cape Verde Archipelago), highlighting also biogeographical relationships between Macaronesia and northwestern Africa. Unfortunately, no matK data were obtained to date that illustrate about the position of F. cespitosa in the plastid phylogeny.
5. Conclusions
As currently circumscribed, many species of Frankenia are accepted in such a broad sense that they encompass various biological entities superficially similar but quite diverse when studied in detail. Although appearances can be deceptive, the application of integrative taxonomy, combining morphological, phylogenetic, ecological and distributional data, reveals the existence of distinct biological entities that need to be identified in due measure and that should be accepted in proper taxonomic ranks.
According to the morphological and molecular phylogenetic data obtained in the present work, F. sahariensis and F. dinteri show differences enough to be accepted as specifically distinct from F. pulverulenta and other Eurasian and African members of the genus. Further research is going on to generate sequences of new plastid regions that will contribute to complete the picture on phylogenetic relationships of Eurasian and African aggregates of Frankenia.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: Phylogenetic trees of Frankenia accessions after: (A) Maximum Parsimony (MP) analysis of ITS nuclear sequences; (B) Neighbour Joining (NJ) analysis of ITS nuclear sequences; (C) Maximum Parsimony (MP) analysis of matK plastid sequences; (D) Neighbour Joining (NJ) analysis of matK plastid sequence.
Author Contributions
Conceptualization, M.Á.A., M.B.C., J.A.B., M.M.A. and J.L.V.; methodology, M.Á.A., M.B.C., J.A.B., M.M.A. and J.L.V.; formal analyses, M.B.C. and J.A.B.; writing—original draft preparation, M.Á.A. and M.B.C.; writing—review and editing, M.Á.A., M.B.C. and J.A.B.; collecting permit acquisition, M.M.A.; field work, M.Á.A., M.B.C., M.M.A. and J.L.V. All authors have contributed substantially to the general discussion of results and taxonomic treatment, and have also read and agreed to the published version of the manuscript.
Funding
This work was partly supported by H2020 Research and Innovation Staff Exchange Programme of the European Commission, project 645636, “Insect-plant relationships: insights into biodiversity and new applications” (FlyHigh), and by the grants UADIF21-29, UADIF22-28, UADIF23-32, UAUSTI21-04, UAUSTI22-05, UAUSTI23-12, VIGROB21-166, VIGROB22-166, VIGROB23-166, ACIE21-01, ACIE22-01, ACIE23-02 and ACIE24-01 from diverse programmes of Universidad de Alicante.
Data Availability Statement
DNA sequences generated in the present research are available at GenBank (https://www.ncbi.nlm.nih.gov/genbank/). Other information related to this study will be provided discretionally upon request to the corresponding author.
Acknowledgments
We thank the curators and staff of the herbaria ABH, BM, BOL, G-DC, K, LINN, MA, MPU, NBG, P, and W for facilitating consultation or providing information on their collections of Frankenia. Especially, Andreas Berger (W), María Álvarez and Saba Rokni (K), Cornelia Klak (BOL), Caroline Loup (MPU), Anthony Magee (NBG) and Erik Van Wick (PRE), for their help during our visit to their corresponding herbaria. Further, Vanessa Rodríguez Invernón and Germinal Rouhan (P), and Alberto Herrero and Leopoldo Medina (MA) provided with images of some types as well as samples from some vouchers for DNA extraction and/or seed analysis. Specimens from southern Africa were harvested under permission provided by the Department of Environment and Nature Conservation of Northern Cape Province (permits FLORA0057/2017, FLORA0058/2017) and CapeNature of Western Cape Province of South Africa (permits AAA005-00080-0028, AAA008-00031-0028, 0027-AAA008-00699, 0028-AAA008-00203).
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Cuénoud, P.; Savolainen, V.; Chatrou, L.W.; Powell, M.; Grayer, R.J.; Chase, M.W. Molecular phylogenetics of Caryophyllales based on nuclear 18S rDNA and plastid rbcL, atpB, and matK DNA sequences. Amer. J. Bot. 2002, 89, 132–144. [Google Scholar] [CrossRef]
- Olson, M.E.; Gaskin, J.F.; Ghahremani-Nejad, F. Stem anatomy is congruent with molecular phylogenies placing Hypericopsis persica in Frankenia (Frankeniaceae): comments on vasicentric tracheids. Taxon 2003, 52, 525–532. [Google Scholar] [CrossRef]
- Gaskin, J.F.; Ghahremani-Nejad, F.; Zhang, D.-Y.; Londo, J.P. A systematic overview of Frankeniaceae and Tamaricaceae from nuclear rDNA and plastid sequence data. Ann. Missouri Bot. Gard. 2004, 91, 401–409. [Google Scholar]
- POWO. Plants of the World Online (continuously updated). Available online: https://powo.science.kew.org/ (accessed on 31 January 2025).
- Summerhayes, V.S. A revision of the Australian species of Frankenia. Bot. J. Linn. Soc. 1930, 48, 337–387. [Google Scholar] [CrossRef]
- Heywood, V.H.; Brummitt, R.K.; Culham, A.; Seberg, O. Flowering plant families of the world; Royal Botanic Gardens: Kew, UK, 2007; pp. 150–151. [Google Scholar]
- Niedenzu, F. Frankeniaceae. In Die Natürlichen Pflanzenfamilien, 2nd ed.; Engler, H.G., Prantl, K.A., Eds.; W. Engelmann: Leipzig, Germany, 1925; Volume 21, pp. 276–281. [Google Scholar]
- Kubitzki, K. Frankeniaceae. In The families and genera of vascular plants 5. Malvales, Capparales and non-betalain Caryophyllales; Kubitzki, K., Bayer, C., Eds.; Springer: Berlin, Germany, 2003; Volume 5, pp. 209–212. [Google Scholar]
- Mucina, L.; Bültmann, H.; Dierßen, K.; Theurillat, J.-P.; Raus, T.; et al. Vegetation of Europe: hierarchical floristic classification system of vascular plant, bryophyte, lichen, and algal communities. Appl. Veg. Sci. 2016, 19 (Suppl. 1), 3–264. [Google Scholar] [CrossRef]
- Jäger, E.J. Die Verbreitung von Frankenia in der Mongolei, in Westeurasien und im Weltmaβstab. Flora (Jena) 1992, 186, 177–186. [Google Scholar] [CrossRef]
- Manning, J.C.; Helme, N.A. Frankenia fruticosa (Frankeniaceae), a new dwarf shrub from the Knersvlakte, Western Cape. S. African J. Bot. 2014, 91, 84–87. [Google Scholar] [CrossRef]
- Crespo, M.B.; Alonso, M.Á.; Martínez-Azorín, M.; Villar, J.L.; Mucina, L. What is wrong with Frankenia nodiflora Lam. (Frankeniaceae)? New insights into the South African sea-heaths. Plants 2023, 12, 2630. [Google Scholar] [CrossRef]
- Santos, A. Frankenia. In Flora Iberica, 3. Plumbaginaceae (Partim)-Capparaceae; Castroviejo, S., Aedo, C., Cirujano, S., Laínz, M., Montserrat, P., Morales, R., Muñoz Garmendia, F., Navarro, C., Paiva, J., Soriano, I., Eds.; Real Jardín Botánico-CSIC: Madrid, Spain, 1993; Volume 3, pp. 446–453. [Google Scholar]
- Whalen, M.A. Frankeniaceae. In Flora of North America north of Mexico, 6. Magnoliophyta: Cucurbitaceae to Droseraceae; Flora of North America Editorial Committee, Ed.; Oxford University Press: New York, USA, 2015; Volume 6, pp. 409–412. [Google Scholar]
- Grigore, M.N.; Toma, C. Salt Secretion. In Anatomical Adaptations of Halophytes: A Review of Classic Literature and Recent Findings; Grigore, M.N., Toma, C., Eds.; Springer: Cham, Switzerland, 2017; pp. 147–239. [Google Scholar]
- Pérez Cuadra, V.; Verolo, M.; Cambi, V. Morphological and anatomical traits of halophytes. Adaptive versus phylogenetic value. In Handbook of Halophytes. From Molecules to Ecosystems towards biosaline agriculture; Grigore, M.N., Ed.; Springer: Cham, Switzerland, 2021; pp. 1329–1347. [Google Scholar] [CrossRef]
- Whalen, M.A. Systematics of Frankenia (Frankeniaceae) in North and South America. Syst. Bot. Monogr. 1987, 17, 1–93. [Google Scholar] [CrossRef]
- Gorshkova, S.G. Frankenia L. In Flora of the USSR; Shishkin, B.K., Ed.; Komarov Botanical Institute, Academy of Sciences of the USSR: Moscow-Leningrad, Russia, 1949; Volume 15, pp. 271–275. (in Russian) [Google Scholar]
- Chater, A.O. Frankenia L. In Flora europaea; Tutin, T.G., Heywood, V.H., Burges, N.A., Moore, D.M., Valentine, D.H., Walters, S.M., Webb, D.A., Eds.; Cambridge University Press: Cambridge, UK, 1968; Volume 2, pp. 294–295. [Google Scholar]
- Chrtek, J. Frankeniaceae. In Flora iranica; Rechinger, K.H., Ed.; Akademische Druck und Verlagsanstalt: Graz, Austria, 1972; Fascicle 99; pp. 1–5. [Google Scholar]
- Lomonosova, M.N. A new species of the genus Frankenia (Frankeniaceae) from Tuva ASSR. Bot. Zhurn. (Moscow & Leningrad) 1984, 99, 548–549. [Google Scholar]
- Nègre, R. Les Frankenia du Maroc. Trav. Inst. Sci. Chérifien, Sér. Bot. 1957, 12, 5–56. [Google Scholar]
- Jahandiez, E.; Maire, R. Catalogue des Plantes du Maroc (Spermatophytes et Ptéridophytes); Imprimerie Minerva: Alger, Algeria, 1932; Volume 2, 557p. [Google Scholar]
- Emberger, L.; Maire, R. Catalogue des Plantes du Maroc (Spermatophytes et Ptéridophytes), Supplément aux volumes I, II et III; Imprimerie Minerva: Alger, Algeria, 1941; Volume 4, 1181p. [Google Scholar]
- Turland, N.J.; Wiersema, J.H.; Barrie, F.R.; Greuter, W.; Hawksworth, D.L.; Herendeen, P.S.; Knapp, S.; Kusber, W.-H.; Li, D.-Z.; Marhold, K.; May, T.W.; McNeill, J.; Monro, A.M.; Prado, J.; Price, M.J.; Smith G.F., Eds.; International Code of Nomenclature for algae, fungi, and plants (Shenzhen Code) adopted by the Nineteenth International Botanical Congress Shenzhen, China, July 2017. − Koeltz Scientific Books: Königstein, Germany, 2018; [Regnum Veg. 159]. [CrossRef]
- EURO+MED. Euro+Med PlantBase – The information resource for Euro-Mediterranean plant diversity (continuously updated). Available online: http://www.europlusmed.org (accessed on 31 January 2025).
- Thiers, B. Index Herbariorum: a global directory of public herbaria and associated staff (continuously updated). Available online: http://sweetgum.nybg.org/ih/ (accessed on 31 January 2025).
- Rasband, W.S. ImageJ. U.S. National Institutes of Health: Bethesda, Maryland, USA. Available online: https://imagej.nih.gov/ij/ (accessed on 15 January 2025).
- IPNI. The International Plant Names Index (continuously updated). The Royal Botanic Gardens, Kew, Harvard University Herbaria & Libraries, and Australian National Herbarium. Available online: http://www.ipni.org (accessed on 31 January 2025).
- Leistner, O.A.; Morris, J.W. Southern African place names. Ann. Cape Prov. Mus. Nat. Hist. 1976, 12, 1–565. [Google Scholar]
- National Geospatial Information. SA Mapsheet Referencing. Available online: http://www.ngi.gov.za/indexphp/what-we-do/maps-and-geospatial-information/41-sa-mapsheet-referencing (accessed on 15 January 2025).
- Mucina, L.; Rutherford, M.C., Eds. The vegetation of South Africa, Lesotho and Swaziland. South African National Biodiversity Institute: Pretoria, South Africa, 2006. [Strelitzia 19].
- Quézel, P. Analysis of the flora of Mediterranean and Saharan Africa. Ann. Missouri Bot. Gard. 1978, 65, 479–534. [Google Scholar] [CrossRef]
- Takhtajan, A. Floristic regions of the world; University of California Press: London, UK, 1986; 522p. [Google Scholar]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- White, T.J.; Bruns, T.D.; Lee, S.B.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR protocols: a guide to methods and applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, USA, 1990; pp. 315–322. [Google Scholar]
- Ford, C.S.; Ayres, K.L.; Toomey, N.; Hayder, N.; et al. Selection of candidate coding DNA barcoding regions for use on land plants. Botanical Journal of the Linnean Society 2009, 159, 1–11. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucl. Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Nei, M.; Kumar, S. Molecular evolution and phylogenetics; Oxford University Press: New York, USA, 2000. [Google Scholar]
- Felsenstein, J. Evolutionary trees from DNA sequences: a maximum likelihood approach. J. Mol. Evol. 1981, 17, 368–376. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbour-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef]
- Akaike, H. A new look at the statistical model identification. IEEE Trans. Autom. Control. 1874, 19, 716–723. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: more models, new heuristics and parallel computing. Nat. Method. 2012, 9, 772. [Google Scholar] [CrossRef]
- Kimura, M. A simple method for estimating evolutionary rate of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K. Estimation of the number of nucleotide substitutions when there are strong transition-transversion and G + C-content biases. Mol. Biol. Evol. 1992, 9, 678–687. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence limits on phylogenies: an approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Hohna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Villar, J.L.; Alonso, M.Á.; Juan, A.; Gaskin, J.; Crespo, M.B. Out of the Middle East: new phylogenetic insights in the genus Tamarix (Tamaricaceae). J. Syst. Evol. 2019, 57, 488–507. [Google Scholar] [CrossRef]
- Meimberg, M.; Dittrich, P.; Bringmann, G.; Schlauer, J.; Heubl, G. Molecular phylogeny of Caryophyllidae s.l. based on matK sequences with special emphasis on carnivorous taxa. Plant Biol. (Stuttgart) 2000, 2, 218–128. [Google Scholar] [CrossRef]
- Zhang, M.-L.; Meng, H.-H.; Zhang, H.-X.; Vyacheslav, B.V.; Sanderson, S.C. Himalayan origin and evolution of Myricaria (Tamaricaeae) in the Neogene. PLoS One 2014, 9, e97582. [Google Scholar] [CrossRef]
- Chen, Z.D.; Yang, T.; Lin, L.; Lu, L.M.; et al. Tree of life for the genera of Chinese vascular plants. J. Syst. Evol. 2016, 4, 277–306. [Google Scholar] [CrossRef]
- Zhang, M.-L.; Hao, X.-L.; Sanderson, S.C.; Vyacheslav, B.V.; Sukhorukov, A.P.; Zhang, H.-X. Spatiotemporal evolution of Reaumuria (Tamaricaceae) in Central Asia: insights from molecular biogeography. Phytotaxa 2014, 167, 89–103. [Google Scholar] [CrossRef]
- Song, F.; Li, T.; Yan, H.F.; Feng, Y.; Jin, L.; Burgess, K.S.; Ge, X.J. Plant DNA barcode library for native flowering plants in the arid region of northwestern China. Mol. Ecol. Res. 2023, 23, 1389–1402. [Google Scholar] [CrossRef] [PubMed]
- Ozenda, P. Flora of the Sahara, 2nd ed.; CNRS: Paris, France, 1977. [Google Scholar]
- Chevallier, L. Deuxième note sur la Flore du Sahara (suite et fin). Bull. Herb. Boissier, sér. 2 1903, 3, 757–779. [Google Scholar]
- Philippi, R.A. Descripción de las plantas nuevas incorporadas últimamente en el herbario chileno. Anales Univ. Chile 1872, 41, 663–746. [Google Scholar] [CrossRef]
- Maire, R. Contribution à l’étude de la Flore de l’Afrique du Nord, Fascicule 24. Bull. Soc. Hist. Nat. Afrique Nord 1936, 27, 203–238. [Google Scholar]
- WFO. World Flora Online (continuously updated). Available online: http://www.worldfloraonline.org (accessed on 31 January 2025).
- Pohnert, H. Neuen Arten aus Südwest-Afrika. Frankenia densa Pohnert. Mitt. Bot. Staatssamml. München 1954, 9–10, 447–448. Available online: https://www.biodiversitylibrary.org/page/15187743.
- Rivas Martínez, S.; Díaz, T.E.; Fernández González, F.; Izco, J.; Loidi, J.; Lousã, M.; Penas, Á. Vascular plant communities of Spain and Portugal: Addenda to the syntaxonomical checklist of 2001. Part II. Itinera Geobot. 2002, 15, 433–922. [Google Scholar]
- Obermeyer, A.A. Frankeniaceae. Flora of southern Africa; Ross, J.H, Ed.; Botanical Research Institute: Pretoria, South Africa, 1976; Volume 22, pp. 32–36. [Google Scholar]
- Caballero, A. Datos botánicos del territorio de Ifni (2.ª parte). Trab. Mus. Nac. Ci. Nat., Ser. Bot. 1935, 30, 3–33. [Google Scholar]
- Candolle, A.P. de. Prodromus systematis naturalis regni vegetabilis. Treuttel et Würtz: Paris, France, 1824; Volume 1, pp. 349–350. Available online: https://www.biodiversitylibrary.org/page/153953.
- Goldblatt, P.; Manning, J.C. Cape Plants: A conspectus of the Cape Flora of South Africa. [Strelitzia 9]; National Botanical Institute: Cape Town, South Africa; Missouri Botanical Garden: St Louis, MO, USA, 2000; 743p. [Google Scholar]
- Raimondo, D.; von Staden, L.; Foden, W.; Victor, J.E.; Helme, N.A.; Turner, R.C.; Kamundi, D.A.; Manyama, P.A. (Eds.) Red List of South African Plants. [Strelitzia 25]; South African National Biodiversity Institute: Pretoria, South Africa, 2009. [Google Scholar]
- Lowe, R.T. Novitiae Florae Maderensis: or Notes and Gleanings of Maderan Botany. Trans. Cambridge Philos. Soc. 1838, 6, 523–551. [Google Scholar]
Figure 2.
Frankenia sahariensis. Distribution map of the studied material.
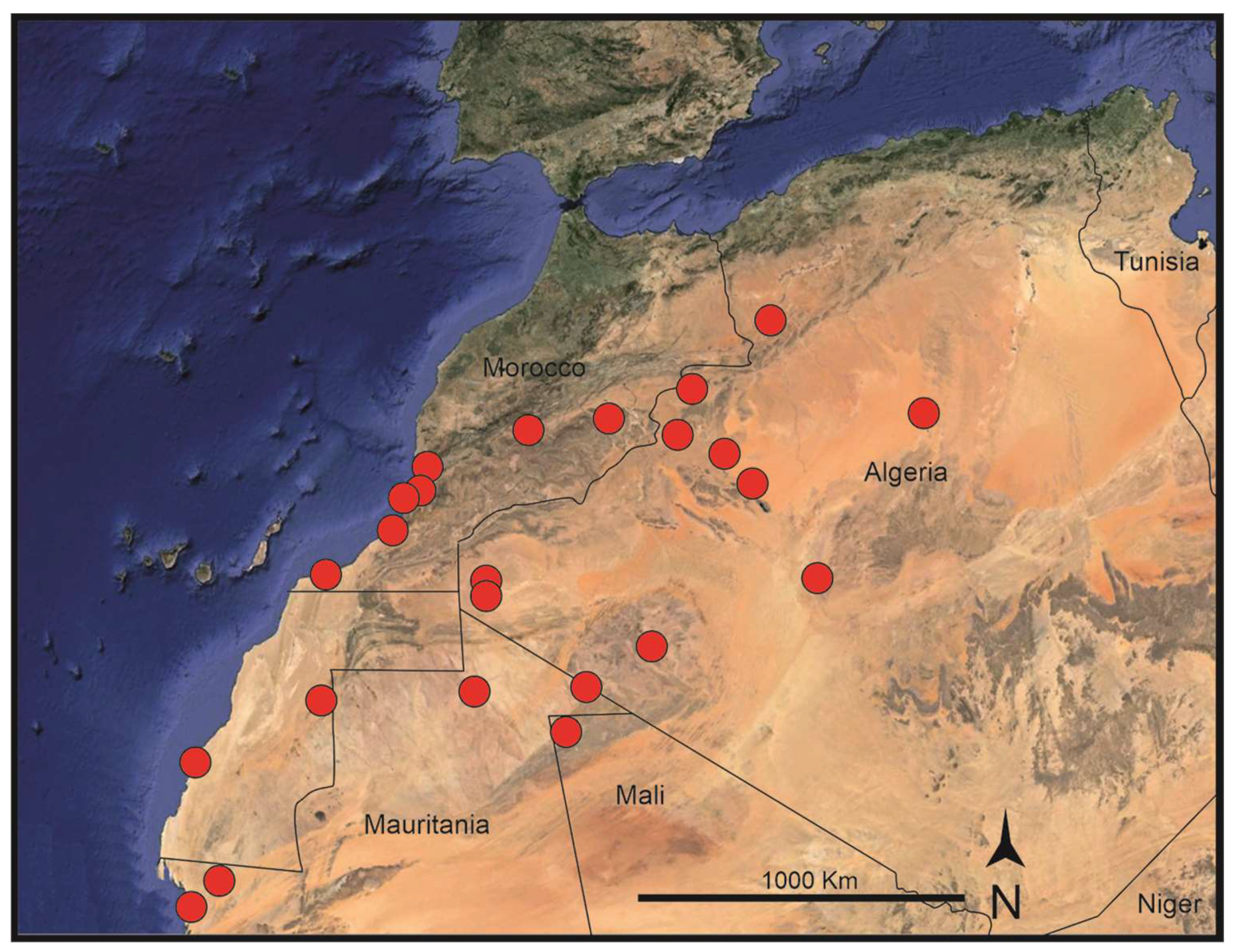
Figure 3.
Frankenia dinteri. (A) Habit in habitat; (B) Detail of many-flowered inflorescences; (C) Flowering branchlets at anthesis; (D) Detail of calyx after anthesis, with acuminate teeth (arrow). Photographs by Mario Martínez-Azorín: (A, B) Goageb, Namibia; (C, D) Onseepkans, South Africa.
Figure 3.
Frankenia dinteri. (A) Habit in habitat; (B) Detail of many-flowered inflorescences; (C) Flowering branchlets at anthesis; (D) Detail of calyx after anthesis, with acuminate teeth (arrow). Photographs by Mario Martínez-Azorín: (A, B) Goageb, Namibia; (C, D) Onseepkans, South Africa.

Figure 4.
Frankenia dinteri. Distribution map of the studied material.
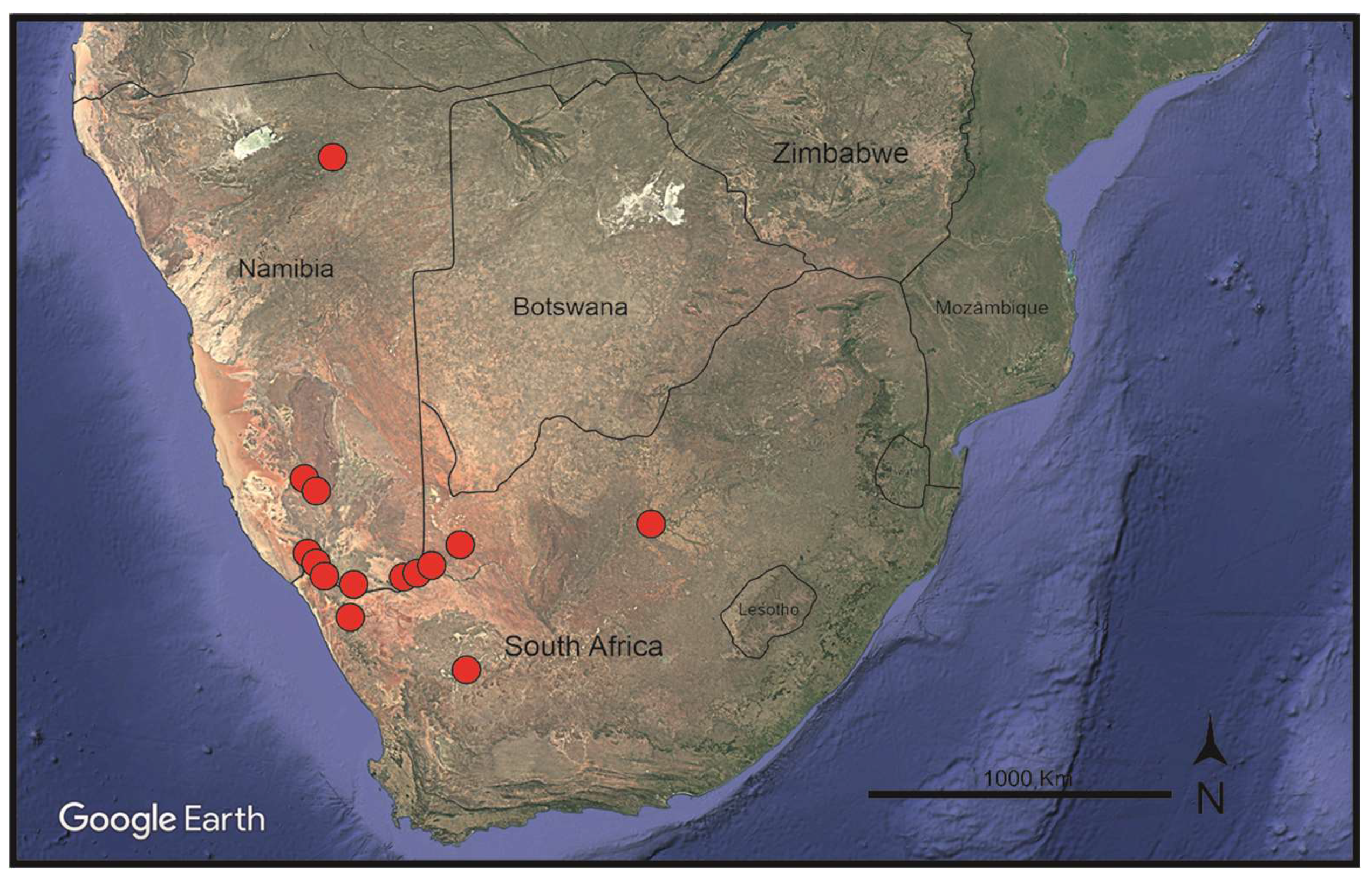
Figure 5.
Maximum Likelihood (ML) phylogenetic tree of Frankenia accessions from ITS nrDNA sequences as obtained with MEGA. The three main clades recovered in the analyses are marked A–D on the tree, following Crespo et al. [12], plus two additional subclades (B1 and B2). Samples of F. sahariensis, F. pulverulenta and F. dinteri are highlighted in their corresponding clades or subclades for easier comparison of their relative position. Numbers above branches indicate the bootstrap percentage (BP) values after 10,000 replicates from the ML analysis, whereas numbers below branches indicate the posterior probabilities (PP) from the Bayesian Inference analysis. GenBank codes are shown after each taxon/accession name.
Figure 5.
Maximum Likelihood (ML) phylogenetic tree of Frankenia accessions from ITS nrDNA sequences as obtained with MEGA. The three main clades recovered in the analyses are marked A–D on the tree, following Crespo et al. [12], plus two additional subclades (B1 and B2). Samples of F. sahariensis, F. pulverulenta and F. dinteri are highlighted in their corresponding clades or subclades for easier comparison of their relative position. Numbers above branches indicate the bootstrap percentage (BP) values after 10,000 replicates from the ML analysis, whereas numbers below branches indicate the posterior probabilities (PP) from the Bayesian Inference analysis. GenBank codes are shown after each taxon/accession name.
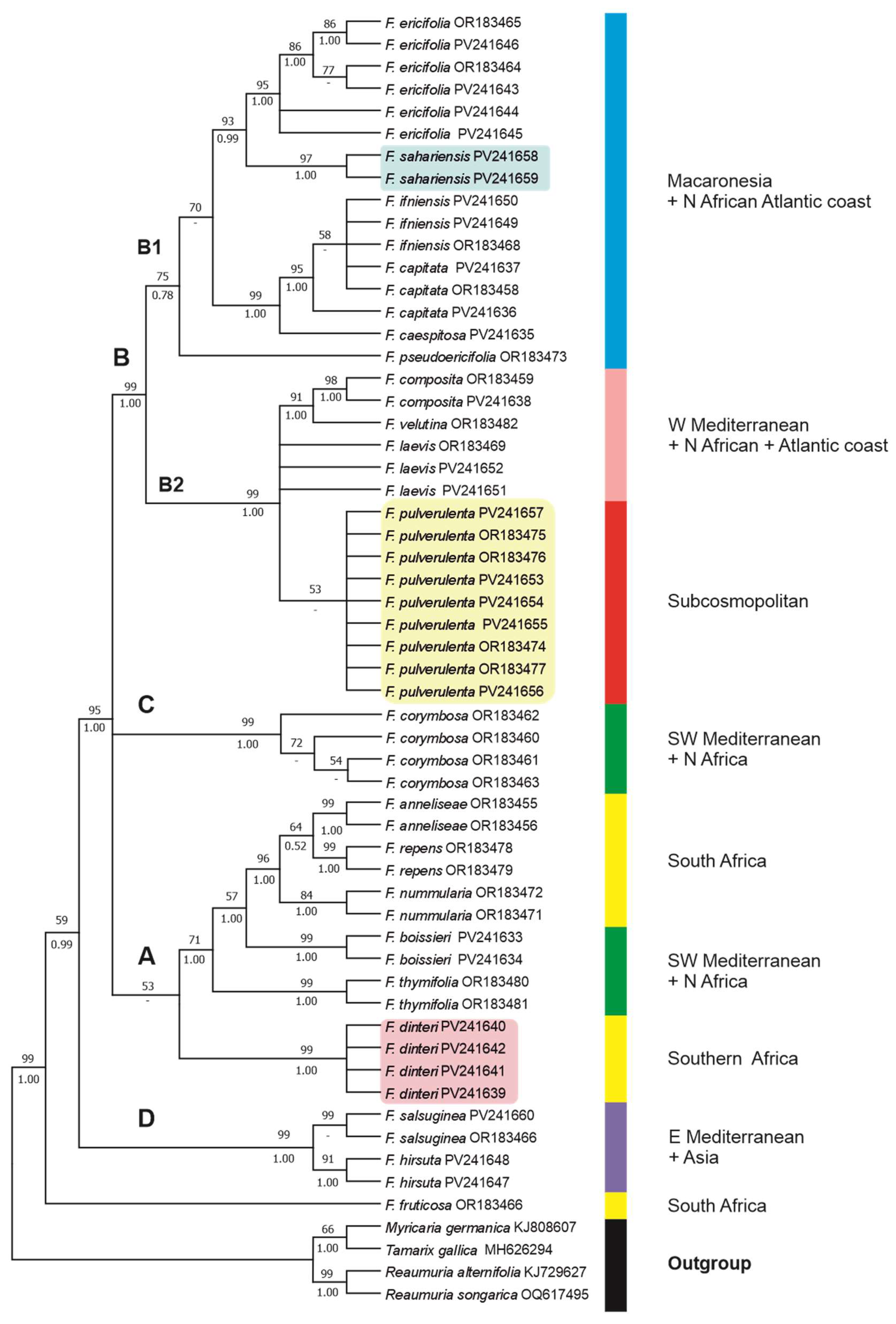
Figure 6.
Maximum Likelihood (ML) phylogenetic trees of Frankenia accessions from matK cpDNA sequences as obtained with MEGA. The four main clades and subclades recovered in the analyses are marked A–D on the tree, paralleling the ITS tree. Similarly, accessions of F. sahariensis, F. dinteri and F. pulverulenta are highlighted to easily compare their relative positions. Numbers above branches indicate the bootstrap percentage (BP) values after 10,000 replicates from the ML analysis, whereas numbers below branches indicate the posterior probabilities (PP) from the Bayesian Inference analysis. GenBank codes are shown after each taxon/accession name.
Figure 6.
Maximum Likelihood (ML) phylogenetic trees of Frankenia accessions from matK cpDNA sequences as obtained with MEGA. The four main clades and subclades recovered in the analyses are marked A–D on the tree, paralleling the ITS tree. Similarly, accessions of F. sahariensis, F. dinteri and F. pulverulenta are highlighted to easily compare their relative positions. Numbers above branches indicate the bootstrap percentage (BP) values after 10,000 replicates from the ML analysis, whereas numbers below branches indicate the posterior probabilities (PP) from the Bayesian Inference analysis. GenBank codes are shown after each taxon/accession name.
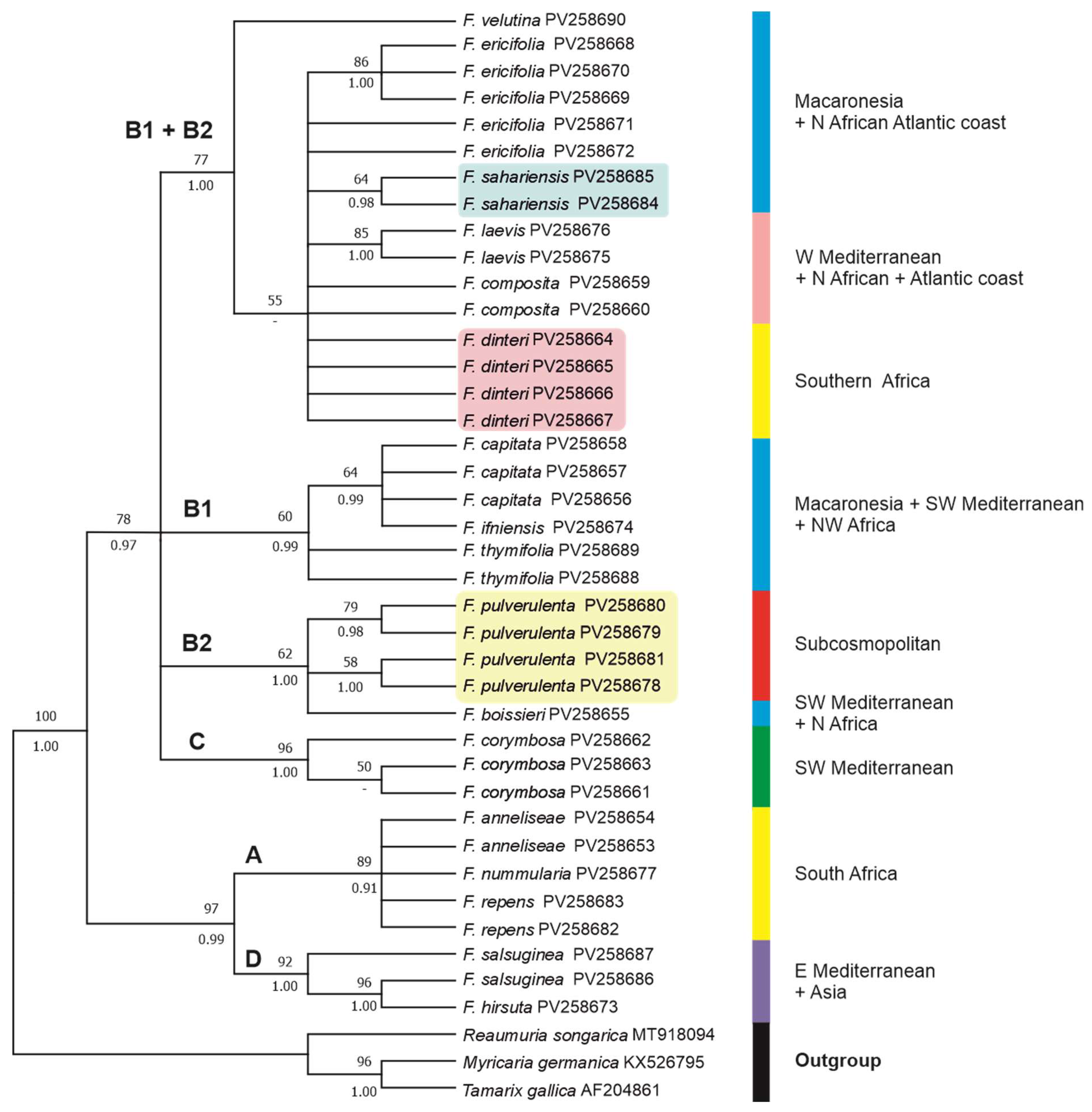
Table 1.
Seed samples of Frankenia for SEM studies, with provenance and herbarium vouchers.
| Taxon | Locality | Herbarium voucher |
| F. sahariensis | Mauritania: Iouik | P-05038748 |
| F. dinteri | South Africa: Klipfontein | ABH-76891 |
| F. pulverulenta | Spain: Alicante, Orxeta | ABH-7952 |
Table 3.
Comparison of morphological characters of the newly named African species, Frankenia sahariensis and F. dinteri, to the annual sea-heath F. pulverulenta.
Table 3.
Comparison of morphological characters of the newly named African species, Frankenia sahariensis and F. dinteri, to the annual sea-heath F. pulverulenta.
| F. saharienis | F. dinteri | F. pulverulenta | |
| General habit | Annual or perennial herbaceous and weakly lignified at base | Annual or perennial herbaceous and weakly lignified at base | Annual or rarely short-lived perennial |
| Stem features | Prostrate to decumbent, non-rooting | Erect to ascendant, non-rooting | Prostrate to ascending, non-rooting |
| Branchlet indumentum | ± densely puberulous on one side and near nodes | ± densely pubescent on one side | ± densely pubescent on one side |
| Branch trichomes: types and length (mm) | minute, ca. 0.1–0.2, curled or hooked | minute, ca. 0.1–0.2, curled or hooked | minute, ca. 0.1–0.2, curled |
| Petiole length × width (mm) | 0.5−0.8 × 0.2−0.3, | 0.8−1.2 × 0.5−0.6 | 0.5–1.5× 0.2–0.3 |
| Petiole sheath | (2−)4−5 pairs of cilia | 4−6 pairs of cilia | 2–6 pairs of cilia |
| Sheath cilia length (mm) and colour | 0.2−0.5 mm, unequal, whitish | 0.4−0.5 mm, unequal, whitish | 0.3–0.6, unequal, whitish |
| Leaf blade length × width (mm) | (1.8−)2.5−4 × 0.5−0.7 | (2.5−)3−4.5 (6) × 0.5−2 | 2–5 × 1.5–3 |
| Leaf blade outline and colour | Triangular-ovate to oblong-ovate, ± concolorous, midrib notably thickened | Elliptic to oblong, ± concolorous, midrib narrow, not thickened | Obovate-cuneate, ± concolorous, midrib narrow, not thickened |
| Leaf blade shape and margins | Obtuse apex and cordate to rounded base; often strongly revolute on margins or flattened in the lower third | Obtuse apex and rounded base; often strongly revolute on margins or flattened in the lower third | Rounded, slightly emarginate apex and cuneate base; flattened or slightly revolute on margins |
| Leaf blade indumentum | Glabrous on the upper side, glabrous to loosely hairy beneath with scattered short straight trichomes 0.1–0.2 mm long | Glabrous on the upper side, and glabrous to loosely hairy beneath with scattered short straight trichomes 0.1–0.2 mm long | Glabrous on the upper side, ± densely pubescent beneath with scattered short straight trichomes 0.1–0.2 mm long |
| Inflorescence | Flowers often in axillary or terminal dichasial groups, usually condensed and glomerular | Flowers in dense axillary or terminal dichasial groups, usually densely condensed and glomerular | Flowers solitary and scattered along branch dichotomies in loose groups |
| Bracteole length × width (mm) | 0.5–1.5 × 0.4–0.6, about half the calyx length | 1–2 × 0.4–1 mm, about half the calyx length | 2.5–4, as long as or longer than calyx |
| Calyx length × width (mm), shape and torsion | 2–4 × 0.6–1, fusiform-tubular to fusiform, often twisted | 3–4.7 × 0.6−1, fusiform-tubular to fusiform, non-twisted | 2.5–4(–5) × 0.8–1.5, fusiform-tubular to fusiform, non-twisted |
| Calyx indument (appearance and length) | Entirely glabrous or densely papillate between the glabrous ribs, with heterogeneous trichomes (flattened papillae 0.2–0.3 mm, globose-claviform papillae ca. 0.1 mm, and minute globose vesicles) | Entirely glabrous or puberulous between the glabrous ribs, with homogeneous papillae ca. 0.1 mm long | Entirely glabrous or puberulous between the glabrous ribs, with homogeneous papillae up to 0.2 mm |
| Calyx teeth length (mm) and shape | 0.5–1, acute, mucronate (mucro ca. 0.1–0.2 mm), cucullate | ca. 0.5, acute, long mucronate (mucro ca. 0.4 mm), cucullate. | 0.4–0.8, acute to mucronate (mucro ca. 0.1 mm), cucullate |
| Petal size (mm) and balde colour | 5–6 × 1–1.5, pinkish-mauve but whitish below | 4–4.5 × 0.5–0.8 pinkish-mauve but whitish below | 3.5–5 × 0.6–0.9, whitish-pink to pinkish-mauve, whitish below |
| Petal blade size (mm), and shape | 2.3–3.5 × 1–1.5, obovate-cuneate, rounded and erose-denticulate at apex | 2–2.5 × 0.5–0.8, obovate, rounded to truncate and irregularly erose-denticulate at apex | 2–2.5 × 0.5–0.8, narrowly cuneate to obovate, truncate and erose-denticulate at apex |
| Petal claw (mm) | 2–2.5 × 0.3–0.4, narrowly cuneate | 2–2.5 × 0.3–0.4, narrowly cuneate | 2–2.5 × 0.3–0.5, cuneate |
| Petal ligule (mm) | 2–2.5 × 0.3–0.4 mm,free apex ca. 0.2–0.4 × 0.2–0.3 mm, ovate- ovate-oblong, obtuse to subacute, entire | 2–2.5 × 0.4–0.5, free apex ca. 1 × 0.5 mm, ovate-oblong, obtuse to subacute, entire | 1–2 × 0.2–0.3, free apex ca. 0.4 mm, triangular, acute, entire |
| Stamen filament length (mm) and morphology | 3.5–5.5, expanded ca. 0.5 mm in the lower part, but tapering and filiform in the distal part | 2.3–2.5, expanded ca. 0.5 mm in the lower half, but tapering and filiform in the distal part | 4–6, expanded ca. 0.2 mm in the lower half, but tapering and filiform in the distal part |
| Anther length (mm), shape and colour | 0.6–0.8, ellipsoid, yellowish | 0.6−0.8, ellipsoid, yellowish | 0.2–0.4, oblong–ellipsoid, yellowish |
| Ovules per placenta | 9–12 | 10–12 | 12–20 |
| Capsule size (mm) | 1.4–2 × 0.8–1 | 3–4.5 × 1.4–2 | 2–3 × 0.5–1 |
| Seed number and size (mm) | 24–30, 0.4–0.5 × 0.2–0.25 |
25–30, 0.5−0.6 × 0.2–0.3 |
up to 45, 0.5–0.7 × 0.2–0.3 |
| Testa papillae length (µm) | 10–14 | 7–12 | 4–17 |
| Papillae morphology and distribution | Homogeneous, globose to conical-obtuse, denser on distal part | Homogeneous, conical-obtuse, denser on distal part | Homogeneous, conical-obtuse, denser on distal part |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



