
Submitted:
28 May 2025
Posted:
29 May 2025
You are already at the latest version
Abstract
Chemical pest control is effective and widely used in agriculture because it can quickly reduce pest populations. However, it has significant environmental and agronomic impacts. This study assessed the “in vitro” compatibility of the entomopathogenic fungus Metarhizium anisopliae (Metchnikoff) Sorokin with deltamethrin and neem oil insecticides. We used three fungal isolates obtained from the commercial product Metarril® WPE9. Bioassays measured the effect of the insecticides on vegetative and reproductive parameters. The results showed that deltamethrin stimulated the vegetative growth of the three isolates tested and was compatible with the fungus at all the doses evaluated. However, neem oil reduced the growth and germination of the isolates at the highest concentration, being classified as moderately toxic. Deltamethrin showed potential for use in addition to M. anisopliae in integrated pest management (IPM) programs due to its compatibility. Neem oil showed compatibility at commercial concentrations but toxicity at the highest concentration, indicating that its combined use with M. anisopliae should be carefully evaluated. This study allowed us to understand the interactions between entomopathogenic fungi and insecticides, providing valuable information for adopting biological control strategies in agricultural systems.
Keywords:
Integrated Pest Management
; deltamethrin
; biocontrol
; biological control
1. Introduction
Pest control reduces the population of insects that cause economic damage in productive areas, with chemical control being one of the most widely used methods. The main insect pests affecting Eucalyptus production in Brazil are the defoliator caterpillars, especially Thyrinteina arnobia [1].
A wide range of formulations available on the market are easy to apply and effective for controlling defoliator caterpillars [2]. On the other hand, this form of control has negative implications, such as reducing the populations of natural enemies, increasing pest resistance and production costs, and especially environmental contamination [3,4].
Brazilian legislation has recently approved the use of tank mixtures recommended by field technicians, allowing for the simultaneous combination of chemical and biological products for pest control [5]. This approach aims to enhance integrated pest management (IPM) strategies by enabling targeted and multi-faceted control of harmful insect populations. Such practices offer a more sustainable system, as combining products can reduce selective pressure on pests and decrease the volume of chemical pesticides needed.
Pressure from international public policies is aimed at making production systems more sustainable. For this reason, IPM is a key strategy, combining two or more complementary control strategies considering economic, ecological and social factors [6,7]. Among the methods, those that preserve natural enemies and increase the mortality rate of pests to below the level of economic damage stand out, which allows for the ecological balance of the agroecosystem [3,8].
Chemical insecticides such as lufenuron, which acts as a chitin synthesis inhibitor, affecting the formation of the cuticle [9] and tebufenozide, an ecdysone agonist, which mimics the moulting hormone in insects, inducing early maturity and death by molting [10,11] are highly effective in the different stages of the insect life cycle. However, these molecules’ high persistence and toxicity severely impact the environment, so these insecticides have been banned in Brazil. This has contributed to formulating new molecules with a lower environmental impact [2].
Pyrethroids have been registered for the control of various pests in agriculture and forestry, especially in eucalyptus cultivation, as they have shown positive results in the control of defoliating lepidopterans [2,12]. Deltamethrin is a pyrethroid widely used in monocultures due to its broad spectrum of action and low persistence in the soil [13]. In addition, the degradation of deltamethrin in the soil can contribute to the bioaugmentation process, i.e., this pesticide can be efficiently biodegraded by bacteria such as those of the Serratia Bizio (Proteobacteria) genus, further minimizing its environmental impacts [14].
Botanical insecticides have advantages in generally having low persistence and environmental toxicity. Neem oil, Azadirachta indica (Meliaceae), has an active limonoid compound called azadirachtin and is found in practically all parts of the plant [15,16]. The action of this compound on insects causes food inhibition, behavioral and morphological changes, physiological disorders, inhibition of growth and the reproductive process [17,18]. There are reports of azadirachtin’s efficiency on several orders of insect pests, such as Coleoptera, Hemiptera and Lepidoptera [19,20,21].
Biological control has become a trend, as it contributes to pest control, improves productivity, reduces costs and applies chemical insecticides to the environment [22,23]. Biological control is a positive practice for the forestry sector to obtain certification seals, such as the Forest Stewardship Council - FSC, which follow export requirements and add value to the product. This control method releases, increases, regulates and conserves natural enemies such as predators, parasitoids, and microorganisms [24].
Commercializing entomopathogenic fungi as biocontrol agents is a reality on the international market. Brazil’s main microorganisms used in biocontrol products are Beauveria bassiana (Balsamo-Crivelli) Vuillemin, Isaria fumosorosea Wise, M. anisopliae (Metschnikoff) Sorokin and Isaria fumosoroseus (Wize) Brown & Smith [25]. They act by contact, involving the stages of adhesion, germination, differentiation, penetration of the cuticle and internal dissemination throughout the host’s body [26,27,28]. They can also cause infection at any stage of host development and have a high environmental dispersal capacity and multiplication capacity, making their mass production low-cost [29,30].
The species M. anisopliae (Metchnikoff) Sorokin was the first entomopathogenic fungus to be used in pest control programs in Brazil [31], because it is a generalist in pest control, has high efficiency and has advantages over chemical products. However, there are some concerns due to the broad spectrum of action that can include natural enemies [28,30,32].
Combining chemical and botanical insecticides and entomopathogenic fungi can make pest control more efficient and help reduce pesticide applications in the field [33,34]. IPM should be based on the degree of compatibility of the strategies adopted, as their inappropriate use results in deleterious effects on the development and reproduction of the entomopathogen [35,36].
Optimizing production management by combining the use of products used to control insect pests with sustainable production is considered. This study aimed to evaluate the mixing potential of the synthetic insecticides deltamethrin and botanical neem oil with the entomopathogenic fungus M. anisopliae by means of “in vitro” compatibility.
2. Material and Methods
2.1. Experiment Location
The work was carried out at the Phytopathology Laboratory - LF of the Universidade Federal dos Vales do Jequitinhonha e Mucuri - UFVJM, in Diamantina, Minas Gerais, Brazil.
2.2. Obtaining and Cultivating Isolates of Metarhizium Anisopliae
To carry out the bioassays on the effect of insecticides on the growth, production of conidia and germination of M. anisopliae “in vitro”, three isolates were used (ISO 01, ISO 02 and ISO 03) of the entomopathogenic fungus Metarhizium anisopliae strain WP E9, from the commercial product Metarril® WPE9, marketed by the company Koppert Biological Systems. For this work, three isolates were used in order to increase genetic variation.
The isolates were reactivated by depositing spores in the center of Petri dishes (100 mm) containing Sabouraud Dextrose Agar (SDA) culture medium and incubated in a BOD at 25 °C, RH 70±10%, with 12-hour photophase, for 15 days. From these cultures, 5 mm diameter disks were taken to carry out the bioassays.
2.3. Insecticides Used and Concentrations Tested
The insecticides used were botanical neem oil, commercial product Sempre Verde Killer Neem®, 6000 ppm and the synthetic insecticide Decis 25 CE® (deltamethrin, 25 g/l CE).
Two bioassays were carried out in a completely randomized design, consisting of four treatments for each insecticide and seven replicates for each of the three isolates of M. anisopliae, with one replicate being considered a Petri dish.
The first bioassay compared the isolates of the entomopathogenic fungus with three different concentrations of the chemical insecticide deltamethrin 0.75, 1.50 and 3.00%. The second bioassay compared the entomopathogenic isolates with different concentrations of the botanical insecticide neem oil and 16.67, 33.33 and 66.66 mg L-1 of deltamethrin. These concentrations were established based on the commercial concentration recommended by the manufacturers of each product for pest control in eucalyptus cultivation (1.50% for neem and 33.3 mg L-1 for deltamethrin). The lower and upper doses were determined considering half of the ideal dose and more than 50% for the upper dose. A pure SDA medium, without the addition of insecticides, was used as a control in each bioassay.
2.4. Preparing Culture Media with Insecticides
The insecticides were sterilized by filtering them through a 0.22 µm Millipore filter, and the pre-determined quantities of each treatment were added to the still liquid SDA medium at a temperature between 45 and 50 ºC to avoid possible alterations to its properties and stirred for 2 minutes to homogenize the added compounds. 20 mL of the medium was poured into Petri dishes (100 mm). After solidifying, a 5 mm diameter disc, taken from the edges of the colonies grown for 15 days, was inserted into the center of each dish with the mycelium facing downwards. The plates were incubated in a BOD at 25 °C, RH 70 ± 10%, with 12-hour photophase, for 18 days.
2.5. Evaluation of Mycelial Growth
The M. anisopliae isolates were examined after eight days, using the radial growth of each repetition. Evaluations were carried out periodically with an interval of 72 hours between measurements. Two perpendicular diameters previously marked on the outside of the bottom of the Petri dish were measured with a 30 cm ruler, and then the average of the two measurements was calculated.
2.6. Evaluation of Conidia Production
The production of conidia was assessed at 18 days of incubation, considered to be the final period for vegetative growth. Five representative dishes from each treatment were randomly selected and three discs were removed from the edges of the colonies using a previously flamed 5 mm diameter perforator and transferred to an eppendorf with 1 mL of sterile 0.01% Tween® 80 solution and homogenized by shaking for 1 minute in a vortex. Next, 100 µL of the conidia suspension was placed in a Neubauer chamber and a direct count was made under an optical microscope to quantify conidia production [37].
2.7. Evaluation of Conidia Germination
To assess germination, a 5 mm disc was removed from the edges of colonies grown for 18 days, transferred to an eppendorf with 1 mL of sterile 0.01% Tween® 80 solution and homogenized by shaking for 1 minute in a vortex. The conidia were suspended in this aqueous solution, remaining for one hour to hydrate [38]. Next, 30 µL of the suspension of each treatment were placed on 9 mm agar medium discs held on the top of sterilized microscope slides; three discs were used for each treatment. Subsequently, the slides were individualized, placed in a humid chamber of Petri dishes lined with moistened filter paper, and incubated for 15 hours in a BOD at 25 °C, RH 70 ± 10%, with 12-hour photophase. The conidia were observed under an optical microscope at 40× magnification. Germination was assessed by counting 100 to 150 conidia on each disc. Conidia with a germ tube larger than its diameter were considered germinated [39].
2.8. Biological Index (BI) - Fungicide Compatibility
The compatibility of the insecticides with the entomopathogenic fungus was calculated using the Biological Index (BI) formula:
Where: CV is the percentage of vegetative growth of the fungal colony relative to the control; ESP is the percentage of sporulation of colonies relative to the control; GER is the percentage of germination of conidia relative to the control.
According to the BI values obtained, the insecticides were classified as compatible (BI > 66), moderately toxic (42 ≤ BI ≤ 66) and toxic (BI < 42) [40].
2.9. Statistical Analysis
The data were analyzed using Analysis of Variance (ANOVA) and the means were compared using the Tukey test at a probability of 5%, using the SAEG (UFV) statistical analysis program [41]. The means of the diameter of the M. anisopliae mycelium as a function of incubation time were presented by regression.
3. Results
3.1. Effect of Insecticides on Germination
The presence of the chemical insecticide deltamethrin in the culture medium reduced the germination rates of isolates ISO 01 and ISO 03 (Table 1). The highest concentrations of deltamethrin had a significant effect, with a 19.40% reduction in germination for isolate ISO 01 at a dose of 66.66 mg L⁻¹, compared to the control. For ISO 03, germination decreased by 51.78% and 52.02% when exposed to 33.33 and 66.66 mg L⁻¹, respectively. These results indicate that ISO 03 is the most sensitive to deltamethrin, showing a significant drop in germination even at lower concentrations.
Neem oil did not affect the germination of the isolates when applied at the commercial concentration of 1.50%. However, isolate ISO 01 exhibited a germination reduction of 49.18% and 56.78% compared to the control when exposed to 0.75% and 3.00%, respectively. Isolate ISO 03 was the most sensitive, with a 72.53% reduction at the highest neem concentration (Table 1). These findings indicate that, although neem oil did not impact germination at its recommended field concentration, higher doses may negatively affect fungal germination, especially for the more sensitive isolates.
3.2. Effect of Insecticides on Vegetative Growth
The isolates of M. anisopliae exposed to deltamethrin and neem oil at different concentrations exhibited significant differences in mycelial growth in SDA culture medium.
Deltamethrin stimulated mycelial growth, particularly for ISO 01, where an increasing dosage correlated with greater growth, with the highest dose (66.66 mg L⁻¹) resulting in a 55.34% increase compared to the control. However, ISO 02 and ISO 03 showed peak growth at 16.67 mg L⁻¹ and 33.33 mg L⁻¹, respectively. The highest dose (66.66 mg L⁻¹) had a toxic effect on ISO 02 and ISO 03, indicating a dose-dependent sensitivity among isolates (Figure 1).
Conversely, neem oil reduced the growth of all three isolates, with increasing concentrations leading to greater inhibition (Figure 2). The growth curves of the control diverged progressively as neem concentration increased, demonstrating isolate sensitivity to the botanical insecticide. Statistical analysis showed that ISO 02 and ISO 03 followed a negative linear pattern, where higher neem concentrations resulted in lower mycelial growth. However, ISO 01 exhibited better tolerance, as its growth was not significantly impaired even at the highest neem concentration (Table 1). The evaluation of the mycelial growth of the M. anisopliae isolates subjected to treatments with deltamethrin and neem oil at different concentrations can be seen in Figure 3 and Figure 4, during the development of the isolates in Petri dishes over the incubation period at 8, 12, 15 and 18 days.
3.3. Effect of Insecticides on Sporulation
There was no difference in conidia production among treatments with either insecticide. However, ISO 02 was the most sensitive to deltamethrin at 66.66 mg L⁻¹, showing a lower conidia count (9.92 × 10⁶ conidia mL⁻¹) compared to the other isolates (Table 1).
ISO 02 exhibited also the highest tolerance and synergistic response to neem oil, producing 28.16 × 10⁶ conidia mL⁻¹ at the highest concentration (3.00%), surpassing all other isolates and treatments. In contrast, ISO 01 and ISO 03 showed lower tolerance, with reduced sporulation rates of 5.36 × 10⁶ and 6.48 × 10⁶ conidia mL⁻¹, respectively, at the same neem concentration (Table 1).
These findings indicate that deltamethrin negatively affected sporulation at high concentrations, especially for ISO 02, whereas neem oil had a variable impact, favoring sporulation in ISO 02 but inhibiting it in ISO 01 and ISO 03.
3.4. Compatibility Based on the Biological Index (BI)
The biological index (BI) was used to assess the overall compatibility of the insecticides with M. anisopliae.
Deltamethrin produced the highest BI values across all three isolates, classifying it as compatible at all tested doses (Table 2). This suggests that, despite its effects on germination and sporulation at higher concentrations, deltamethrin does not significantly hinder fungal development and may be used in integrated pest management programs in conjunction with M. anisopliae.
4. Discussion
IPM has become a common strategy in the forestry sector to control insect attacks that cause damage to production. This measure uses different control techniques in an integrated way, which are considered to have less impact on the environment since the aim is to keep the population density of pest insects below the level of economic damage [8,42]. Entomopathogenic fungi are an excellent option to be used in IPM programs, as they are described as agents that cause high levels of epizootics in nature, in a general way, and are the most versatile and environmentally safe biological control agents [43].
The effectiveness of IPM strategies in pest control can be compromised by pesticide application, as it directly impacts vegetative growth, conidial germination, and sporulation of beneficial fungi. Additionally, conidia, which are already vulnerable to environmental factors, may experience even greater reductions in survival due to the indiscriminate use of chemical pesticides, further weakening their role in biological control [44]. In this context, our results on the preliminary evaluation of the compatibility of entomopathogenic fungi with chemical and botanical insecticides “in vitro” prove the potential of the strategy adopted and the efficiency of combining these control strategies in the use of IPM.
The increase in vegetative growth and the absence of a negative effect on the reproductive parameters of the M. anisopliae isolates, in SDA medium contaminated with doses of deltamethrin, demonstrates the compatibility of this insecticide with this entomopathogenic fungus. This result corroborates the synergistic effect of M. anisopliae with chemical insecticides described in the literature, this positive compatibility increased the potential for insect toxicity and mortality, even when the dosage of the chemical formulation caused a reduction in the vegetative growth of the fungal isolate [45]. Band, Kabre and Dhurve [46] carried out a compatibility test between M. anisopliae exposed to 10 chemical insecticides, but, unlike the previous work, the results were negative, in which deltamethrin caused a reduction in the fungus’ vegetative growth, being considered the second most aggressive chemical formulation for this biological control agent. This is likely due to the specific characteristics of the isolates used in this test.
Khun et al. [47] proved that the compatibility and synergistic effect do not depend solely on the dosages of chemical insecticides but rather on the compatible performance of each fungal isolate individually when subjected to the different doses applied, so much so that in their work they presented results with positive (compatibility) and negative (incompatibility) effects, reinforcing the idea that when considering the application of entomopathogenic fungi and chemical insecticides in IPM programs there must be a need to understand the dynamics of each component through the degree of compatibility since the inappropriate use of any means of biological control can affect the development and reproduction of the entomopathogen [5,36].
The results obtained in this study showed that deltamethrin, regardless of the dosage, promoted the greatest increase in the vegetative growth of M. anisopliae. This same result was obtained when the chemical insecticide Flonicamide was used in different concentrations, sublethal - 0.03%, lethal - 0.06% and superlethal - 0.12%, which showed positive effects when compared to the other chemical insecticides applied to M. anisopliae [45]. Likewise, for the MA-7 isolate of M. anisopliae which, when exposed to concentrations of 40 and 200 ppm (representing 0.004% and 0.02%) of imidacloprid, had an increase in vegetative growth [48]. These stimuli can be explained by the presence of adjuvants in the commercial formulation, which promote greater adhesion and stability of formulations containing entomopathogenic fungi [49]. Adjuvants, when used correctly, can optimize the performance of biological agents, such as M. anisopliae and B. bassiana, in pest control by increasing the durability of conidia in the field and their infectivity against target insects [50,51]. The mild abrasive action of the adjuvants breaks up conidial agglomerations, which increase the number of propagules, promoting greater growth [52].
The entomopathogenic fungus may have the ability to degrade the molecules of the active ingredients of insecticides through physiological mechanisms of resistance, which enable the microorganism to metabolize these compounds and use them as a source of nutrients, especially N and C [53]. This may explain the differences in mycelial growth between isolates due to the variation in the degradation rates of insecticide molecules and the different ways in which these resources are used later [51,54,55,56]. Thus, the stimulation of microbial growth in the presence of deltamethrin could be compensation for the entomopathogenic fungus when facing a stressful (toxic) environment, which in response, increases vegetative growth [45,47,48,51,56,57].
Compatibility in the vegetative growth and sporulation of M. anisopliae was obtained in the lower dosages of 0.75% (45 ppm) and 1.5% (90 ppm - the recommended dose) of neem oil, considered within the range of the manufacturer’s technical recommendation. These positive effects confirm and strengthen the potential use of the botanical insecticide in IPM programs. Parjane et al. [43] obtained similar results and applying Azadirachtin 3000 ppm at a concentration of 4% (120 ppm) promoted the greatest increases in compatibility tests composed of 19 different types of insecticides. Only the 3.00% concentration (180 ppm) of the botanical insecticide neem in this study reduced the growth and germination of M. anisopliae, demonstrating that this insecticide is incompatible with these isolates, at this concentration, classified as moderately toxic to M. anisopliae.
This work proves that the botanical insecticide neem can have a fungicidal effect at a concentration of 3%, which can be considered a super-lethal dose, twice the amount recommended by the manufacturer (1.5%). This effect was also observed for the entomopathogenic fungi M. anisopliae and B. bassiana when the commercial neem formulation was applied at a concentration of 2% [58]. Azadirachtin obtained from aqueous neem seed extract at a concentration of 1% reduced the growth of B. bassiana [59], while for M. anisopliae the concentrations that caused the inhibitory effect were 0.04 to 5% [60] and 50% of the concentration recommended by the manufacturer [57]. The reduction in growth under different neem concentration scales may be associated with extracting azadirachtin from the products used. Also, the high variability in the amount of azadirachtin present in the commercial product or attributed to inter- and intra-specific genetic variations of the isolates [48,61].
The reduction in conidia production caused by deltamethrin may be related to the susceptibility of the entomopathogenic fungus to the product [34]. The stimulation of output caused by neem oil, unlike that previously observed with deltamethrin, can be explained as a reproductive response to generate offspring and ensure the survival of the species in the contaminated environment, which is why there may be reductions in mycelial growth and consequently increases in conidia production [53]. The production of conidia is linked to the ability of the entomopathogenic fungus to complete the entire cycle within the host and cause secondary infestations in the field [37]. This survival mechanism can present different forms of reaction depending on the different insecticide formulations and even the metabolic variation of the isolates of the same fungus. Therefore, selecting isolates with greater potential for producing conidia compatible with insecticides increases the chances of the fungus remaining in the field and extends its residual effect, reducing the need for new applications.
The variation in the impact of insecticides on the germination of M. anisopliae isolates with the insecticide and the dose/concentration used may be related to the great genetic variability that exists in entomopathogenic fungi [35,40]. The dose-response effect observed with the increase in doses/concentrations and the reduction in germination of isolates ISO 01 and ISO 03 shows greater sensitivity of the isolates when exposed to neem. On the other hand, the lower sensitivity of the isolates exposed to deltamethrin indicates a low impact on the germination of M. anisopliae. This is a promising result, given that the germination of conidia is an important biological aspect of the infection process of the pathogen, as it corresponds to the initial stage of epizooty, and successful infection of the host depends on germination [37,38,62]. Similar results were obtained in a study with the MA-K isolate of M. anisopliae, which had little effect on germination when exposed to permethrin at a concentration of 200 ppm, representing 0.02% of this insecticide [48]. This may be an indication that pyrethroids do not cause significant effects on the germination of M. anisopliae, making it possible to use them together in pest management without damaging the virulence of the pathogen.
Neem at the commercial concentration (1.5%) did not harm the germination of the M. anisopliae isolates, similar to studies with the same entomopathogenic fungus, where there was no adverse effect on germination when exposed to this concentration [48,62]. Thus, the recommended dose of the botanical insecticide neem can be combined with M. anisopliae in the field, maintaining adequate germination rates, colonization and promoting pathology of the insect pest.
The reduction in germination of the isolates exposed to the highest concentration of neem (3.00%) can be attributed to the toxicity of the dosage and the product’s composition. In this context, the product tested would not be suitable for use together with the M. anisopliae ISO 01 and ISO 03 isolates because with lower conidia germination in the field, the potential for the fungus to infect insects will be reduced [63]. In a study with entomopathogenic fungi M. anisopliae, B. bassiana (Bals.) Vuill. and Paecilomyces fumosoroseus (Wise) Brown and Smith, exposed to neem at a concentration of 5%, germination was not reduced [60]. This may be attributed to the quality and/or form of extraction used to obtain the botanical insecticide Neem, which has a different degree of aggressiveness than the product used in this work.
The compatibility of the insecticide deltamethrin with the isolates of M. anisopliae demonstrates their ability to be used as a joint strategy in IPM. Similar results were obtained in a study with the insecticides deltamethrin, fenpropathrin (Meothrin 300), betacyflutrin and permethrin, all belonging to the pyrethroid chemical group, which showed compatibility with the entomopathogenic fungi B. bassiana and M. anisopliae and Pyriproxifen in M. anisopliae [48,53,55]. This compatibility is essential for the preservation of microbial agents. It can also contribute to improving the fungus’s potential as a control agent, since some synthetic substances can cause stress in the insect, facilitate fungal infection and accelerate the proliferation of the disease [37,64].
The genetic variability of the isolates may have been the determining factor for the different compatibility ratings with neem at the highest concentration [35], this variation may have occurred due to the mutation of the fungi through the successive repicking process. The incompatibility of neem with isolates of entomopathogenic fungi M. anisopliae and B. bassiana was also found in other studies with neem at 2% of the recommended dose [58], emulsifiable neem oil at concentrations of 0.5; 1.0 and 1.5% [59] and 0.03% [53] and with the commercial product Neemseto® at a concentration of 0.5% [65]. However, at concentrations of 0.00, 0.75 and 1.50%, the BI values found were classified as compatible with all the isolates, which corroborates the results obtained with isolates of M. anisoplie exposed to the product NeemAzal at concentrations of 0.32, 1.6, 8.0, 40 and 200 ppm, representing 3.2 × 105, 1.6 × 10-4, 8 × 10-4, 0.004 and 0.02% of this botanical insecticide [48]. Thus, neem-based insecticides should be used with restrictions, as they can affect the reproductive parameters of entomopathogenic fungi and present incompatibility, which will depend on the concentration and product used.
The preservation of entomopathogenic fungi in the field is essential for balancing the population of insect pests. “In vitro” studies allow high exposure of these microbial agents to the action of insecticides and, when the product is proven to be safe in the laboratory, a lower effect is expected in the field due to the various factors that hinder the exposure of the microorganism [34]. Thus, choosing isolates compatible with insecticides in “in vitro” trials can guarantee greater success in the joint strategy when submitted to field conditions.
5. Conclusions
The association of the insecticide deltamethrin with M. anisopliae stimulated the growth and sporulation of the isolates. This insecticide was compatible with the entomopathogenic fungus at all the doses evaluated. This makes it possible to simultaneously use this agent and deltamethrin in IPM programs.
The production of M. anisopliae conidia was mostly unaffected by the insecticides deltamethrin (chemical) and neem (botanical). The highest concentration of neem oil interfered with the development and reproduction of M. anisopliae, with moderate toxicity. However, the isolates of this fungus showed potential for association with botanical insecticide at the commercial dose.
Author Contributions
Planning - C. - SS; I. - TO; M.- AS. Execution - C. - SS; Z. - VC. Writing - C. - SS; P. - HRM; M. - PRR; I. - TO; R. - SS; G. - LDL; M. - AS. Review - C. - SS; P. - HRM; M. - PRR; I. - TO; R.- SS; G.- LDL; M.- AS. Corresponding author - P. - HRM; Project Administration - M. - AS; Resource Acquisition - M. - AS.
Funding
This work received external funding from Conselho Nacional de Desenvolvimento Científico e Tecnológico -CNPq, Coordenação de Aperfeiçoamento de Pessoal de Nível Superior - Capes and Fundação de Amparo à Pesquisa do Estado de Minas Gerais - Fapemig for materials and equipment, as well as research grants.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Datasets are available on request.
Acknowledgments
The authors would like to thank CNPq, CAPES and Fapemig for funding the project and the scholarships awarded during the course of the work.
Conflicts of Interest
The author(s) declare no conflict of interest.
References
- Nanini:, F.; Souza, P.G.C.; Soliman, E.P.; Zauza, E.A.V.; Domingues, M.M.; Santos, F.A.; Wilcken, C.F.; da Silva, R.S.; Corrêa, A.S. Genetic Diversity, Population Structure and Ecological Niche Modeling of Thyrinteina Arnobia (Lepidoptera: Geometridae), a Native Eucalyptus Pest in Brazil. Sci. Rep. 2024, 14, 20963. [Google Scholar] [CrossRef] [PubMed]
- Rezende-Teixeira, P.; Dusi, R.G.; Jimenez, P.C.; Espindola, L.S.; Costa-Lotufo, L.V. What Can We Learn from Commercial Insecticides: Efficacy, Toxicity, Environmental Impacts, and Future Developments. Environ. Pollut. 2022, 300, 118983. [Google Scholar] [CrossRef]
- Camilo, S. da S.; Soares, M.A.; Zanuncio, J.C.; Leite, G.L.D.; Pires, E.M.; Cruz, M. do C.M. da Host Plants of Thyrinteina arnobia (Lepidoptera: Geometridae) affect the development of parasitoids Palmistichus elaeisis (Humenoptera: Eulophidae). Rev. Árvore 2015, 39, 159–166. [Google Scholar] [CrossRef]
- Fernandes, F.O.; Abreu, J.A.; Christ, L.M.; Rosa, A.P.S.A. Efficacy of Insecticides against Spodoptera Frugiperda (Smith, 1797). 2019, 11, 494–503. 11.
- Gandini, E.M.M.; Costa, E.S.P.; Dos Santos, J.B.; Soares, M.A.; Barroso, G.M.; Corrêa, J.M.; Carvalho, A.G.; Zanuncio, J.C. Compatibility of Pesticides and/or Fertilizers in Tank Mixtures. J. Clean. Prod. 2020, 268, 122152. [Google Scholar] [CrossRef]
- Kogan, M. Integrated Pest Management: Historical Perspectives and Contemporary Developments. Annu. Rev. Entomol. 1998, 43, 243–270. [Google Scholar] [CrossRef]
- Souto, A.L.; Sylvestre, M.; Tölke, E.D.; Tavares, J.F.; Barbosa-Filho, J.M.; Cebrián-Torrejón, G. Plant-Derived Pesticides as an Alternative to Pest Management and Sustainable Agricultural Production: Prospects, Applications and Challenges. Molecules 2021, 26, 4835. [Google Scholar] [CrossRef]
- Bortolotto, O.C.; Pomari-Fernandes, A.; Bueno, R.C.O.D.F.; Bueno, A.D.F.; Cruz, Y.K.S.D.; Sanzovo, A.; Ferreira, R.B. The use of Soybean Integrated Pest Management in Brazil: A Review. Agron. Sci. Biotechnol. 2015, 1, 25. [Google Scholar] [CrossRef]
- Nascimento, A.R.B.D.; Farias, J.R.; Bernardi, D.; Horikoshi, R.J.; Omoto, C. Genetic Basis of Spodoptera Frugiperda (Lepidoptera: Noctuidae) Resistance to the Chitin Synthesis Inhibitor Lufenuron. Pest Manag. Sci. 2016, 72, 810–815. [Google Scholar] [CrossRef]
- 2022.
- Suarez-Lopez, Y.A.; Aldebis, H.K.; Hatem, A.E.-S.; Vargas-Osuna, E. Interactions of Entomopathogens with Insect Growth Regulators for the Control of Spodoptera littoralis (Lepidoptera: Noctuidae). Biol. Control 2022, 170, 104910. [Google Scholar] [CrossRef]
- Zanuncio, J.C.; Guedes, R.N.C.; Cruz, A.P. da; Moreira, A.M. Eficiência de Bacillus thuringiensis e de Deltametrina, em aplicação aérea, para o controle de Thyrinteina arnobia STOLL, 1782 (Lepidoptera: Geometridae) em Eucaliptal no Pará. Acta Amaz. 1992, 22, 485–492. [Google Scholar] [CrossRef]
- Velíšek, J.; Jurčíková, J.; Dobšíková, R.; Svobodová, Z.; Piačková, V.; Máchová, J.; Novotný, L. Effects of Deltamethrin on Rainbow Trout (Oncorhynchus Mykiss). Environ. Toxicol. Pharmacol. 2007, 23, 297–301. [Google Scholar] [CrossRef] [PubMed]
- Cycoń, M.; Żmijowska, A.; Piotrowska-Seget, Z. Enhancement of Deltamethrin Degradation by Soil Bioaugmentation with Two Different Strains of Serratia marcescens. Int. J. Environ. Sci. Technol. 2014, 11, 1305–1316. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, X.; Wang, J.; Xun, H.; Sun, J.; Tang, F. Comparative Analysis of the Terpenoid Biosynthesis Pathway in Azadirachta indica and Melia azedarach by RNA-Seq. SpringerPlus 2016, 5, 819. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, S. Progress on Azadirachta indica Based Biopesticides in Replacing Synthetic Toxic Pesticides. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef]
- Zanuncio, J.C.; Mourão, S.A.; Martínez, L.C.; Wilcken, C.F.; Ramalho, F.S.; Plata-Rueda, A.; Soares, M.A.; Serrão, J.E. Toxic Effects of the Neem Oil (Azadirachta indica) Formulation on the Stink Bug Predator, Podisus nigrispinus (Heteroptera: Pentatomidae). Sci. Rep. 2016, 6, 30261. [Google Scholar] [CrossRef]
- Francesena, N.; Schneider, M.I. Selectivity Assessment of Two Biorational Insecticides, Azadirachtin and Pyriproxyfen, in Comparison to a Neonicotinoid, Acetamiprid, on Pupae and Adults of a Neotropical Strain Eretmocerus mundus Mercet. Chemosphere 2018, 206, 349–358. [Google Scholar] [CrossRef]
- Shannag, H.K.; Capinera, J.L.; Freihat, N.M. Effects of Neem-Based Insecticides on Consumption and Utilization of Food in Larvae of Spodoptera Eridania (Lepidoptera: Noctuidae). J. Insect Sci. 2015, 15, 152. [Google Scholar] [CrossRef]
- Ali, K. Impact of extracts of Azadirachta indica and Datura inoxia on the esterases and phosphatases of three stored grains insect pests of economic importance. Pak. J. Agric. Sci. 2017, 54, 71–81. [Google Scholar] [CrossRef]
- Alim, Md.A.; Song, J.; Lim, U.T.; Choi, J.J.; Hossain, Md.A. Bioassay of Plant Extracts Against Aleurodicus Dispersus (Hemiptera: Aleyrodidae). Fla. Entomol. 2017, 100, 350–357. [Google Scholar] [CrossRef]
- Sá, L.A.N.D.; Pessoa, M.C.P.Y.; Moraes, G.J.D.; Marinho-Prado, J.S.; Prado, S.D.S.; Vasconcelos, R.M.D. Quarantine Facilities and Legal Issues of the Use of Biocontrol Agents in Brazil. Pesqui. Agropecuária Bras. 2016, 51, 502–509. [Google Scholar] [CrossRef]
- Lopes, S.R.; Paixão, M.A.S.D.; Cruz, I. Viabilidade Econômica de Biofábrica de Trichogramma pretiosum Para Uso Contra Pragas Agrícolas Da Ordem Lepidoptera. Rev. IPecege 2018, 4, 44–50. [Google Scholar] [CrossRef]
- Francis, F.; Jacquemyn, H.; Delvigne, F.; Lievens, B. From Diverse Origins to Specific Targets: Role of Microorganisms in Indirect Pest Biological Control. Insects 2020, 11, 533. [Google Scholar] [CrossRef]
- Bettiol, W. Pesquisa, desenvolvimento e inovação com bioinsumos. In Bioinsumo na Cultura da Soja; Embrapa: Brasília - DF - Brazil, 2022; ISBN 978-65-87380-96-4. [Google Scholar]
- Shahid, A.; Rao, Q.; Bakhsh, A.; Husnain, T. Entomopathogenic Fungi as Biological Controllers: New Insights into Their Virulence and Pathogenicity. Arch. Biol. Sci. 2012, 64, 21–42. [Google Scholar] [CrossRef]
- Shin, T.Y.; Bae, S.M.; Woo, S.D. Screening and Characterization of Antimicrobial Substances Originated from Entomopathogenic Fungi. J. Asia-Pac. Entomol. 2016, 19, 1053–1059. [Google Scholar] [CrossRef]
- Mubeen, N.; Khalid, A.; Ullah, M.I.; Altaf, N.; Arshad, M.; Amin, L.; Talat, Q.; Sadaf, A.; Farwa Effect of Metarhizium anisopliae on the Nutritional Physiology of the Fall Armyworm, Spodoptera frugiperda (J. E. Smith) (Lepidoptera: Noctuidae). Egypt. J. Biol. Pest Control 2022, 32, 73. [Google Scholar] [CrossRef]
- Lacey, L.A.; Grzywacz, D.; Shapiro-Ilan, D.I.; Frutos, R.; Brownbridge, M.; Goettel, M.S. Insect Pathogens as Biological Control Agents: Back to the Future. J. Invertebr. Pathol. 2015, 132, 1–41. [Google Scholar] [CrossRef] [PubMed]
- Deka, B.; Baruah, C.; Babu, A. Entomopathogenic Microorganisms: Their Role in Insect Pest Management. Egypt. J. Biol. Pest Control 2021, 31, 121. [Google Scholar] [CrossRef]
- Biological Control in Latin America and the Caribbean: Its Rich History and Bright Future; Bueno, V. H.P., Luna, M.G., Colmenarez, Y.C., Eds.; CABI: Wallingford, 2020; ISBN 978-1-78924-243-0. [Google Scholar]
- Potrich, M.; Alves, L.F.A.; Haas, J.; Silva, E.R.L.D.; Daros, A.; Pietrowski, V.; Neves, P.M.O.J. Seletividade de Beauveria bassiana e Metarhizium anisopliae a Trichogramma pretiosum Riley (Hymenoptera: Trichogrammatidae). Neotrop. Entomol. 2009, 38, 822–826. [Google Scholar] [CrossRef]
- Abidin, A.F.; Ekowati, N.; Ratnaningtyas, N.I. Compatibility of insecticides with entomopathogenic fungi Beauveria bassiana and Metarhizium anisopliae. Scr. Biol. 2017, 4, 273. [Google Scholar] [CrossRef]
- Oliveira, R.P.; Pessoa, L.G.A.; Loureiro, E.D.S.; Oliveira, M.P. Compatibility of insecticides used for the whitefly control on soybean with Beauveria bassiana. Rev. Agric. NEOTROPICAL 2018, 5, 88–93. [Google Scholar] [CrossRef]
- Celestino, F.N.; Pratissoli, D.; Santos Junior, H.J.G.; Costa, A.V.; Bestete, L.R.; Borges Filho, R.C. Compatibilidade in Vitro Entre Beauveria bassiana e o Óleo de Mamona. Rev. Bras. Ciênc. Agrár. - Braz. J. Agric. Sci. 2018, 13, 1–9. [Google Scholar] [CrossRef]
- Halder, J.; Kushwaha, D.; Rai, A.B.; Singh, B. Interaction Effects between Entomopathogenic Fungi and Neonicotinoid Insecticides against Lipaphis erysimi in Vegetable Ecosystem. Indian J. Agric. Sci. 2019, 89. [Google Scholar] [CrossRef]
- Alves, S.B. Controle Microbiano de Insetos; 2nd, *!!! REPLACE !!!* (Eds.) P: Sérgio Batista Alves: FEALQ, 1998.
- Neves, P.M.O.J.; Hirose, E.; Tchujo, P.T.; Moino Jr, A. Compatibility of Entomopathogenic Fungi with Neonicotinoid Insecticides. Neotrop. Entomol. 2001, 30, 263–268. [Google Scholar] [CrossRef]
- Luz, C.; Fargues, J. Temperature and Moisture Requirements for Conidial Germination of an Isolate of Beauveria bassiana, Pathogenic to Rhodnius Prolixus. Mycopathologia 1997, 138, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Rossi-Zalaf, L.; Alves, S.; Lopes, R.; Silveira Neto, S.; Tanzini, M. Interação de Microrganismos Com Outros Agentes de Controle de Pragas e Doenças. Controle Microbiano Pragas Na América Lat. Avanços E Desafios 2008, 2, 270–302. [Google Scholar]
- SAEG Sistema Para Análises Estatísticas 2007.
- Shang, H.; He, D.; Li, B.; Chen, X.; Luo, K.; Li, G. Environmentally Friendly and Effective Alternative Approaches to Pest Management: Recent Advances and Challenges. Agronomy 2024, 14, 1807. [Google Scholar] [CrossRef]
- Parjane, N.; Kabre, G.; Mahale, A.; Shejale, B.; Nirgude, S. Compatibility of Pesticides with Metarhizium anisopliae. J Entomol Zool Stud 2020, 8, 633–636. [Google Scholar]
- Atrchian, H.; Mahdian, K. Compatibility of Metarhizium anisopliae (Ascomycota: Hypocreales) with Selective Insecticides against Plutella xylostella (Lepidoptera: Plutellidae). Int. J. Trop. Insect Sci. 2022, 42, 3009–3015. [Google Scholar] [CrossRef]
- Nawaz, A.; Razzaq, F.; Razzaq, A.; Gogi, M.D.; Fernández-Grandon, G.M.; Tayib, M.; Ayub, M.A.; Sufyan, M.; Shahid, M.R.; Qayyum, M.A.; et al. Compatibility and Synergistic Interactions of Fungi, Metarhizium anisopliae, and Insecticide Combinations against the Cotton aphid, Aphis gossypii Glover (Hemiptera: Aphididae). Sci. Rep. 2022, 12, 4843. [Google Scholar] [CrossRef]
- Band, S.; Kabre, G.; Dhurve, N. Effect of Chemical Pesticides on Growth Characteristics of Metarhizium anisopliae (Metschnikoff) Sorokin. Int. J. Adv. Biochem. Res. 2024, 8, 526–529. [Google Scholar] [CrossRef]
- Khun, K.K.; Ash, G.J.; Stevens, M.M.; Huwer, R.K.; Wilson, B.A. Compatibility of Metarhizium Anisopliae and Beauveria Bassiana with Insecticides and Fungicides Used in Macadamia Production in Australia. Pest Manag. Sci. 2021, 77, 709–718. [Google Scholar] [CrossRef]
- Schumacher, V.; Poehling, H.-M. In Vitro Effect of Pesticides on the Germination, Vegetative Growth, and Conidial Production of Two Strains of Metarhizium anisopliae. Fungal Biol. 2012, 116, 121–132. [Google Scholar] [CrossRef]
- Holka, M.; Kowalska, J. The Potential of Adjuvants Used with Microbiological Control of Insect Pests with Emphasis on Organic Farming. Agriculture 2023, 13, 1659. [Google Scholar] [CrossRef]
- Mascarin, G.M.; Jaronski, S.T. The Production and Uses of Beauveria bassiana as a Microbial Insecticide. World J. Microbiol. Biotechnol. 2016, 32, 177. [Google Scholar] [CrossRef]
- Mantzoukas, S.; Kitsiou, F.; Natsiopoulos, D.; Eliopoulos, P.A. Entomopathogenic Fungi: Interactions and Applications. Encyclopedia 2022, 2, 646–656. [Google Scholar] [CrossRef]
- Mohan, M.C.; Reddy, N.P.; Devi, U.K.; Kongara, R.; Sharma, H.C. Growth and Insect Assays of Beauveria bassiana with Neem to Test Their Compatibility and Synergism. Biocontrol Sci. Technol. 2007, 17, 1059–1069. [Google Scholar] [CrossRef]
- Sain, S.K.; Monga, D.; Kumar, R.; Nagrale, D.T.; Hiremani, N.S.; Kranthi, S. Compatibility of Entomopathogenic Fungi with Insecticides and Their Efficacy for IPM of Bemisia Tabaci in Cotton. J. Pestic. Sci. 2019, 44, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Tixier, C.; Bogaerts, P.; Sancelme, M.; Bonnemoy, F.; Twagilimana, L.; Cuer, A.; Bohatier, J.; Veschambre, H. Fungal Biodegradation of a Phenylurea Herbicide, Diuron: Structure and Toxicity of Metabolites. Pest Manag. Sci. 2000, 56, 455–462. [Google Scholar] [CrossRef]
- Tamai, M.A.; Alves, S.B.; Lopes, R.B.; Faion, M.; Padulla, L.F.L. Toxicity of pesticides against Baeuveria bassiana (BALS.) VUILL. Arq. Inst. Biológico 2002, 69, 89–96. [Google Scholar] [CrossRef]
- Yada, G.M.; Shiraishi, I.S.; Dekker, R.F.H.; Schirmann, J.G.; Barbosa-Dekker, A.M.; De Araujo, I.C.; Abreu, L.M.; Daniel, J.F.S. Soil and Entomopathogenic Fungi with Potential for Biodegradation of Insecticides: Degradation of Flubendiamide in Vivo by Fungi and in Vitro by Laccase. Ann. Microbiol. 2019, 69, 1517–1529. [Google Scholar] [CrossRef]
- Santos, Á.B. da; Brandão da Silva, T.F.; Coimbra dos Santos, A.; Mesquita Paiva, L.; Luna-Alves Lima, E.Á. Fungitoxic effect of neem oil on Metarhizium anisopliae var. acridum and Metarhizium anisopliae var. anisopliae. 2009, 22, 17–22. [Google Scholar]
- Hirose, E.; Neves, P.M.O.J.; Zequi, J.A.C.; Martins, L.H.; Peralta, C.H.; Moino Jr., A. Effect of Biofertilizers and Neem Oil on the Entomopathogenic Fungi Beauveria bassiana (Bals.) Vuill. and Metarhizium anisopliae (Metsch.) Sorok. Braz. Arch. Biol. Technol. 2001, 44, 419–423. [Google Scholar] [CrossRef]
- Depieri, R.A.; Martinez, S.S.; Menezes Jr., A. O. Compatibility of the Fungus Beauveria bassiana (Bals.) Vuill. (Deuteromycetes) with Extracts of Neem Seeds and Leaves and the Emulsible Oil. Neotrop. Entomol. 2005, 34, 601–606. [Google Scholar] [CrossRef]
- Marques, R.P.; Monteiro, A.C.; Pereira, G.T. Crescimento, Esporulação e Viabilidade de Fungos Entomopatogênicos Em Meios Contendo Diferentes Concentrações Do Óleo de Nim (Azadirachta indica). Ciênc. Rural 2004, 34, 1675–1680. [Google Scholar] [CrossRef]
- Fregonesi, A.F.; Mochi, D.A.; Monteiro, A.C. Compatibilidade de Isolados de Beauveria bassiana a Inseticidas, Herbicidas e Maturadores Em Condições de Laboratório. Arq. Inst. Biológico 2016, 83. [Google Scholar] [CrossRef]
- Pires, L.M.; Marques, E.J.; Oliveira, J.V.D.; Alves, S.B. Selection of isolates of entomopathogenic fungi for controlling Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae) and their compatibility with insecticides used in tomato crop. Neotrop. Entomol. 2010, 39, 977–984. [Google Scholar] [CrossRef]
- Silva, R.A.D.; Quintela, E.D.; Mascarin, G.M.; Barrigossi, J.A.F.; Lião, L.M. Compatibility of Conventional Agrochemicals Used in Rice Crops with the Entomopathogenic Fungus Metarhizium anisopliae. Sci. Agric. 2013, 70, 152–160. [Google Scholar] [CrossRef]
- Damin, S.; Vilani, A.; De Freitas, D.; Krasburg, C.; Alves De Queiroz, J.; Yumi Kagimura, F.; Becker Onofre, S. Ação de Fungicidas Sobre o Crescimento Do Fungo Entomopatogênico Metarhizium Sp. Rev. Acadêmica Ciênc. Anim. 2011, 9, 41. [Google Scholar] [CrossRef]
- Araujo Jr, J.M.D.; Marques, E.J.; Oliveira, J.V.D. Potencial de Isolados de Metarhizium anisopliae e Beauveria bassiana e Do Óleo de Nim No Controle Do Pulgão Lipaphis erysimi (Kalt.) (Hemiptera: Aphididae). Neotrop. Entomol. 2009, 38, 520–525. [Google Scholar] [CrossRef]
Figure 1.
Vegetative growth of ISO 01 (a), ISO 02 (b) and ISO 03 (c) isolates of Metarhizium anisopliae in culture medium containing various doses of deltamethrin.
Figure 1.
Vegetative growth of ISO 01 (a), ISO 02 (b) and ISO 03 (c) isolates of Metarhizium anisopliae in culture medium containing various doses of deltamethrin.
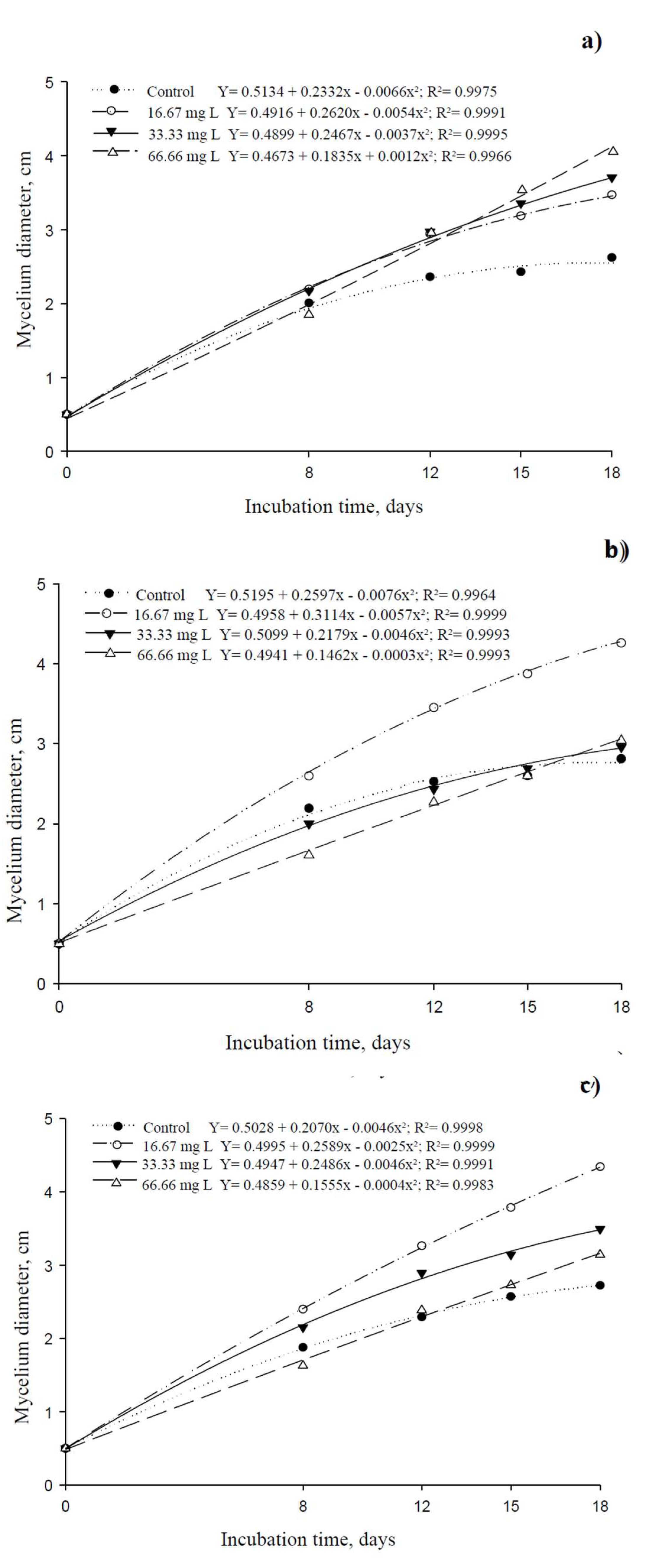
Figure 2.
Vegetative growth of ISO 01 (a), ISO 02 (b) and ISO 03 (c) isolates of Metarhizium anisopliae in culture medium containing various concentrations of neem.
Figure 2.
Vegetative growth of ISO 01 (a), ISO 02 (b) and ISO 03 (c) isolates of Metarhizium anisopliae in culture medium containing various concentrations of neem.
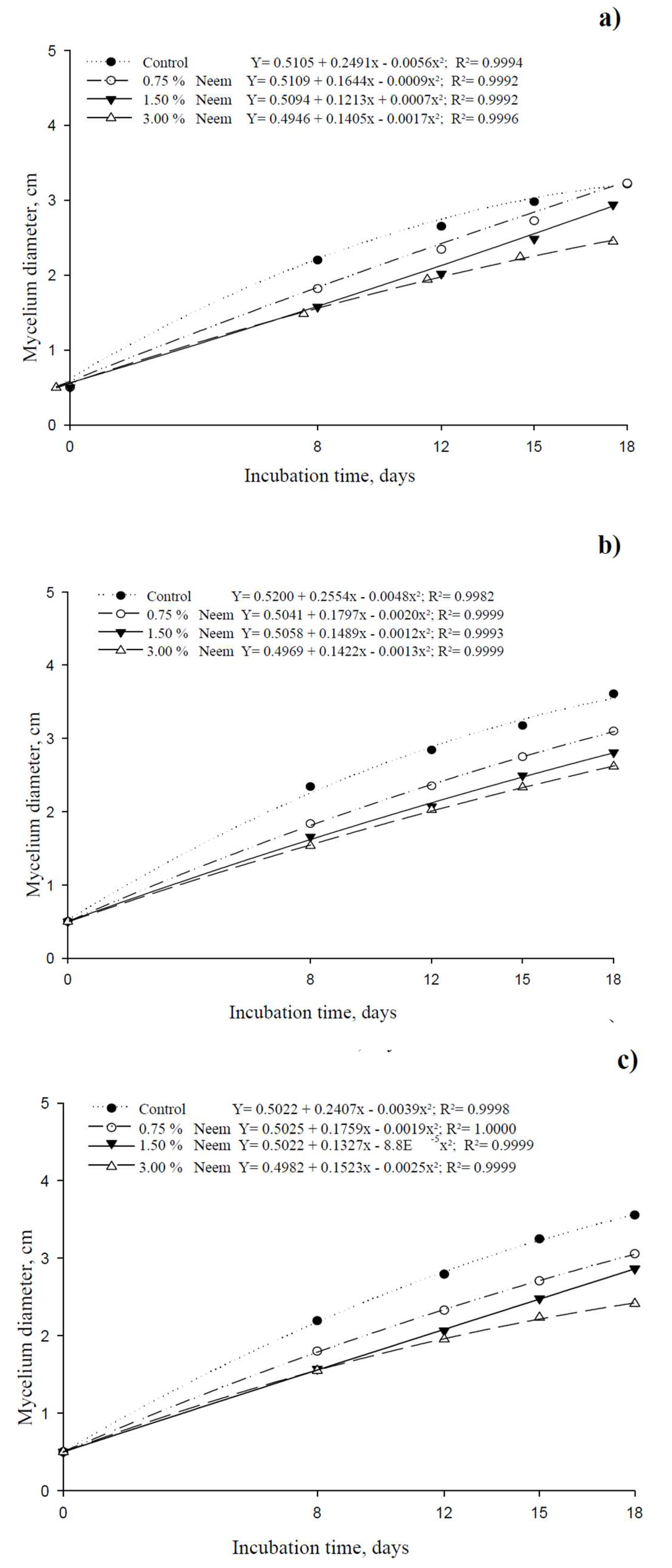
Figure 3.
Mycelial growth of Metarhizium anisopliae isolates in Sabouraud Dextrose Agricultural (SDA) medium contaminated with chemical (deltamethrin at doses of 0.00; 16.67; 33.33) insecticides assessed at 8 days (a), 12 days (b), 15 days (c) and 18 days (d).
Figure 3.
Mycelial growth of Metarhizium anisopliae isolates in Sabouraud Dextrose Agricultural (SDA) medium contaminated with chemical (deltamethrin at doses of 0.00; 16.67; 33.33) insecticides assessed at 8 days (a), 12 days (b), 15 days (c) and 18 days (d).
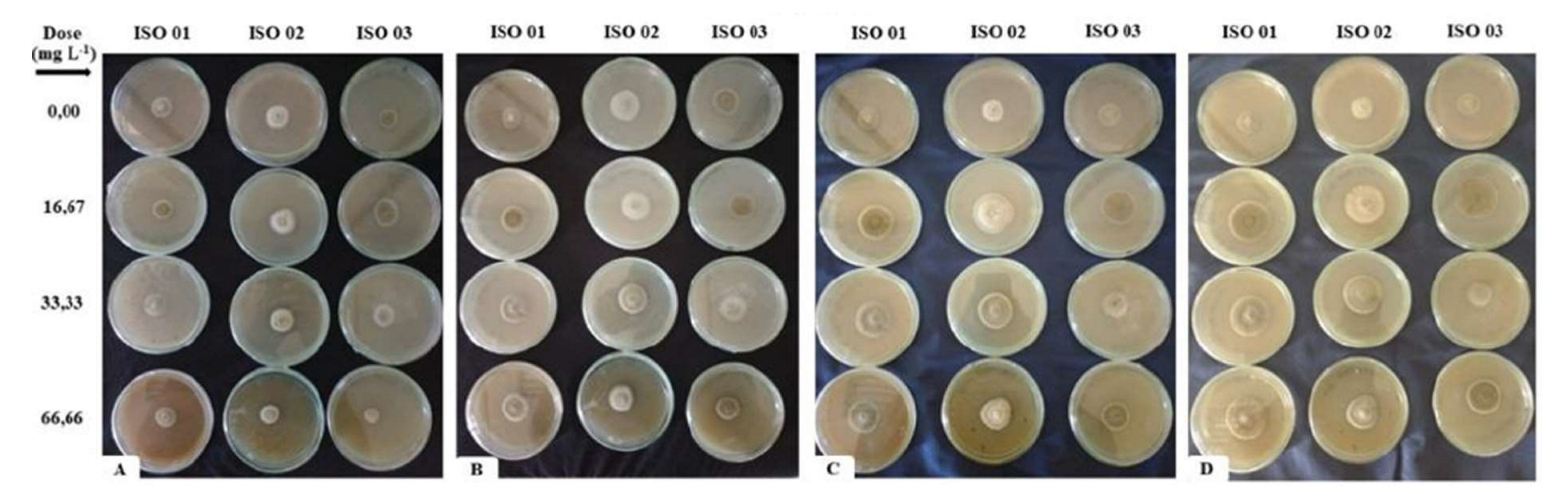
Figure 4.
Mycelial growth of Metarhizium anisopliae isolates in Sabouraud Dextrose Agricultural (SDA) medium contaminated with botanical (neem oil at concentrations of 0.00; 0.75; 1.50 and 3.00%) insecticides assessed at 8 days (a), 12 days (b), 15 days (c) and 18 days (d).
Figure 4.
Mycelial growth of Metarhizium anisopliae isolates in Sabouraud Dextrose Agricultural (SDA) medium contaminated with botanical (neem oil at concentrations of 0.00; 0.75; 1.50 and 3.00%) insecticides assessed at 8 days (a), 12 days (b), 15 days (c) and 18 days (d).


Table 1.
- Micellial growth (cresc.), sporulation and germination (germ.) of isolates of the entomopathogenic fungus Metarhizium anisopliae grown in culture medium contaminated with various doses/concentrations of the insecticides deltamethrin and neem.
Table 1.
- Micellial growth (cresc.), sporulation and germination (germ.) of isolates of the entomopathogenic fungus Metarhizium anisopliae grown in culture medium contaminated with various doses/concentrations of the insecticides deltamethrin and neem.
| Insecticide |
Dose/ Conc. |
ISO 01 | ISO 02 | ISO 03 | ||||||
| Vegetative growth (cm) |
Sporulation 106 cnds mL |
Germination (%) |
Vegetative growth (cm) |
Sporulation 106 cnds mL |
Germination (%) |
Vegetative growth (cm) |
Sporulation 106 cnds mL |
Germination (%) |
||
|
Deltamethrin (mg L-1) |
Control | 2.62bA | 18.24aA | 83.35abA | 2.81bA | 1.76aB | 72.44aA | 2.73bA | 21.92aA | 86.87aA |
| 16.67 | 3.47abA | 28.00aA | 88.62aA | 4.26aA | 3.60aB | 78.45aA | 4.34aA | 23.52aA | 73.51aA | |
| 33.33 | 3.71abA | 16.16aA | 85.12abA | 2.96bA | 8.00aA | 86.07aA | 3.49abA | 18.48aA | 41.89bB | |
| 66.66 | 4.07aA | 29.12aA | 67.18bA | 3.04bB | 9.92aB | 56.59aA | 3.14bB | 25.68aA | 41.68bA | |
| Neem (%) | Control | 3.21aA | 18.32aA | 83.35aA | 3.61aA | 6.48aA | 72.44aA | 3.56aA | 12.24aA | 86.87aA |
| 0.75 | 3.23aA | 15.04aA | 42.36bA | 3.10abA | 6.96aA | 52.05aA | 3.06abA | 13.44aA | 33.9bcA | |
| 1.5 | 2.94aA | 27.04aA | 71.67aA | 2.81abA | 12.88aA | 45.58aA | 2.86abA | 28.48aA | 56.8abA | |
| 3 | 2.45aA | 5.36aB | 36.02bA | 2.62bA | 28.16aA | 27.68aA | 2.41bA | 6.48aB | 23.86cA | |
Means between concentrations followed by equal lowercase letters in the column do not differ from each other using the Tukey test at 5% significance level and means between fungal isolates followed by equal uppercase letters in the row do not differ from each other using the Tukey test at 5% significance level within each variable tested; Conc.: Concentration; cnds mL: Conidia per mL.
Table 2.
Biological index values and classification of deltamethrin and neem insecticides as toxicity to isolates of the fungus Metarhizium anisopliae according to the doses used.
Table 2.
Biological index values and classification of deltamethrin and neem insecticides as toxicity to isolates of the fungus Metarhizium anisopliae according to the doses used.
| Insecticide | Conc. | ISO 01 | ISO 02 | ISO 03 | |||
|---|---|---|---|---|---|---|---|
| BI | Classif. | BI | Classif. | BI | Classif. | ||
|
Deltamethrin (mg L-1) |
Control | 100 | - | 100 | - | 100 | - |
| 16.67 | 139 | C | 170 | C | 129 | C | |
| 33.33 | 115 | C | 257 | C | 101 | C | |
| 66.66 | 150 | C | 301 | C | 109 | C | |
| Neem (%) | Control | 100 | - | 100 | - | 100 | - |
| 0.75 | 88 | C | 94 | C | 92 | C | |
| 1.5 | 115 | C | 128 | C | 144 | C | |
| 3 | 53 | MT | 225 | C | 57 | MT | |
Conc.: Concentration; IB: Biological index (%); Classif.: Classification; C: compatible; MT: moderately toxic; T: toxic.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



