
Submitted:
27 May 2025
Posted:
28 May 2025
You are already at the latest version
Abstract
Neurosteroids are important regulators of numerous central nervous system (CNS) functions, playing essential roles in protecting the brain from insults such as neurodegeneration or ischemia. The choroid plexus (CP) epithelial cells establish an important physical interface between the peripheral blood and the cerebrospinal fluid (CSF), the Blood-CSF barrier (BCSFB), contributing to CNS homeostasis. There is evidence that the BCSFB is a target of sex steroid hormones, which regulate the expression of neuroprotective proteins. In contrast to other brain regions, little is known regarding the production of neurosteroids in the CP. Previously, a cDNA microarray analysis of the CP transcriptome showed the expression of several enzymes involved in steroidogenesis. We, therefore, investigated whether the key steroidogenic enzymes P450scc, P450aro, 17βHSD3, 17βHSD8, 5α-reductase 1, and 5α-reductase 2 are present in the rat and human CP. The present study demonstrates mRNA and protein expression of these enzymes in rat and human CP. Collectively the present data suggest that neurosteroids can be synthesised at the BCSFB and released into the CSF contributing to brain neuroprotection.
Keywords:
Choroid plexus
; Blood-cerebrospinal fluid barrier
; neurosteroidogenesis
1. Introduction
The choroid plexus (CP), located in the ventricles of the vertebrate brain, are branched and highly vascularized structures, consisting of a secretory epithelial cell monolayer joined by connective tissue [1]. These cuboidal epithelial cells are characterized by a large spherical central or basal nucleus, abundant cytoplasm, and numerous mitochondria [2,3]. The apical membrane of the choroid epithelial cells is in contact with the cerebrospinal fluid (CSF) and the basolateral membrane located beneath the basal lamina.
CP are involved in various functions of the central nervous system (CNS) due to its unique architecture and location, constituting the interface between blood and CSF [2,4]. Therefore, it allows the transport of nutrients and other molecules from the blood into the CSF, and in contrast, CP is also involved in the reabsorption and elimination of sub-products derived from cerebral catabolism and toxic products, such as amyloid beta in the context of Alzheimer’s disease, from the CSF to the bloodstream [5,6]. The best known role of CP is the production and secretion of 70-90% of the total CSF [7], highlighting the implication of CP in brain homeostasis [8]. In addition, the CP epithelial cells have several transporters and receptors for molecules, such as neurotransmitters, cytokines, and sex steroid hormones. Importantly, several of these receptors signal downstream cascades that ultimately influence the CP transcriptome and secretome [9,10]. CP epithelium is not only a target but is also a source of a wide variety of bioactive peptides [6,11], contributing to brain recovery processes and neurogenesis [12]. These regenerative functions led to the recent idea that transplanting CP epithelium into ectopic sites might minimize or prevent neural degeneration and its functional consequences [13].
Steroid hormones play an important role in the development, growth, maturation, and differentiation of the CNS [14]. It is now well documented that these actions are not only attributed to the sex hormones synthesized in the steroidogenic glands, which reach the brain via the blood circulation but also to the local synthesis of estrogens and androgens in the brain [15]. A wide variety of steroids and steroidogenic enzymes have been found in various regions of the rodent brain; there is no recognized single center that is steroidogenic so steroids appear to be widely produced for local action [16]. The first step in the biosynthesis of steroid hormones is the conversion of cholesterol into pregnenolone within the mitochondria by the cleavage of the cholesterol side chain catalyzed by the CYP11A1 (P450scc) enzyme. In the subsequent steps, pregnenolone is metabolized into different steroids, including progesterone (P4), testosterone, and estradiol (E2) [17]. For example, the mRNA or protein expression of various key enzymes of the biosynthetic pathways of steroid hormones, such as CYP11A1, CYP19A1 (or aromatase), hydroxysteroid 17-β dehydrogenase (HSD17b) 3, and type 1 and 2 5α-reductases (SRD5A1 and SRD5A2), was shown in several brain loci like the hippocampus, cerebellum, hypothalamus, and brain cortex [18,19,20,21,22,23,24,25,26]. However, certain less-studied enzymes also involved in the metabolism of steroid hormones, such as HSD17b8, which can convert estradiol into estrone [27], have not been identified as being expressed in the brain. The mentioned enzymes play important roles in the synthesis of steroid hormones, where CYP19A1 is responsible for converting androstenedione to estrone and testosterone to E2, HSD17b3 catalyzes the conversion of androstenedione to testosterone, and 5α-reductase can convert testosterone to dihydrotestosterone (DHT) [17,28].
One of the steroid pathways that occurs in the rodent brain leads to the synthesis of testosterone. Androstenedione may be converted to testosterone by 17βHSD type 3 or 5 enzymes, which are found throughout the rat brain [29]. Testosterone exerts analgesic and anxiolytic properties, affects mood and cognition, and promotes synaptic plasticity in rats. Testosterone also prevents neuronal death, and decreased levels in plasma may represent a risk factor for the development of neurodegenerative diseases [30]. The brain is also a source of estrogens that, like androgens, can regulate and influence several physiological properties of the nervous system. For example, estrogens modulate brain functions such as neurotransmission and synaptic plasticity [31,32], can have neuroprotective functions [33], and influence social behaviors like cognitive function and sexual behavior [34,35].
Previous analysis of rat CP transcriptome by cDNA microarrays performed by our group [36] allowed the identification of several steroidogenic enzymes. A more recent report from another laboratory also detected the gene expression of a few steroidogenic enzymes in rat CP, where 17βHSD1 was detected at the protein level by immunohistochemistry [37]. In addition, the gene expression of 17βHSD3, 17βHSD5, and 5α-Reductase 1 was also observed in human CP [38].
In this paper, we analyzed the presence of steroidogenic enzymes in rat and human CP, providing a rationale for subsequent investigation to determine the functional significance of CP as a local site of sex hormones synthesis and its contribution to brain neuroprotection.
2. Materials and Methods
2.1. Cell Culture
For in vitro studies, primary rat CP epithelial cells (CPEC) and human epithelial CP papilloma (HIBCPP) cell line were used. Rat CPEC were prepared as previously described [39]. In brief, CP from lateral ventricles were dissected from newborn rats (P3-P5), previously anesthetized on ice, and digested in PBS containing 0.2% pronase (Fluka, Seelze, Germany) at 37ºC for 5 min. Dissociated cells were washed in Dulbecco’s modified Eagle medium (Sigma-Aldrich, Merck, Portugal) with 10% foetal bovine serum (FBS, Biochrom AG, Berlin), and 100 units/ml of penicillin/ streptomycin (P/S) (Sigma-Aldrich, Merck, Portugal). Cells were seeded into 12 mm culture wells, and cultured in a humidified incubator in 95% air–5% CO2 at 37ºC. Normal DMEM growth medium, supplemented with 100 units/mL P/S, 10% FBS, 10 ng/ml epidermal growth factor (Sigma-Aldrich, Merck, Portugal), 5 µg/ml insulin (Sigma-Aldrich, Merck, Portugal) and 20 µM cytosine arabinoside (Sigma-Aldrich, Merck, Portugal) was replaced 1 day after the initial seeding. A confluent monolayer was obtained within 4–5 days after seeding.
The HIBCPP cell line derived from a human malignant CP papilloma [40] was also used in this work. HIBCPP cells were cultured in DMEM/F12 (Sigma-Aldrich, Merck, Portugal) supplemented with 5 µg/mL insulin (Sigma-Aldrich, Merck, Portugal), 100 units/mL P/S and 10% FBS, and cultured in a humidified incubator in 95% air–5% CO2 at 37ºC.
2.2. In Vivo Assays
The present study used adult Wistar rats, aged 2 months±2 weeks old. All animals were housed in appropriate cages at constant room temperature in a 12h light/12h dark photoperiod, and given standard laboratory chow and water ad libitum. All animal experiments were performed in compliance with the NIH guidelines, and the National and European Union rules for the care and handling of laboratory animals (Directive 2010/63/EU). Animals were deeply anesthetized (i.p. administration of ketamine/ xylazine) before decapitation and brain removal. CPs were obtained from both the lateral ventricles.
2.3. Reverse Transcriptase-Polymerase Chain Reaction (RT-PCR)
Total RNA was extracted from CPs dissected from rat and HIBCPP cells, upon homogenization in TRIzol Reagent (Ambion, Applied Biosystems, USA) according to the manufacturer’s instructions. The purity and yield of the RNA were determined by a spectrophotometer (NanoPhotometer™, Implen, Munich, Germany), and the integrity was assessed by agarose gel electrophoresis.
The RNA (500 ng of total RNA) was reverse transcribed using an M-MLV Reverse Transcriptase (NZYTech, Ltd., Portugal) according to the protocol supplied by the manufacturer. For the RT-PCR, cDNA was amplified by Taq polymerase (Invitrogen) in the presence of specific primers (Table 1) in a final volume of 25µL. Every set of PCR included a control without cDNA. The PCR protocols consisted of 40 cycles of a three-step reaction: 45 s denaturation at 95°C, a 30 s annealing at 58-60ºC, and a 45 s extension at 72°C. The annealing temperature for each gene tested is described in Table 1. PCR products were separated by electrophoresis on 1.5% agarose gels, visualized by GreenSafe (NZYTech, Ltd., Portugal) staining, and detected using UVITEC transilluminator (UVitec Cambridge). The PCR products sequence was confirmed by DNA Sanger sequencing (Stabvida, Portugal).
2.4. Immunocytochemistry
Protein expression and cellular localization of steroidogenic enzymes were analyzed in CPEC and HIBCPP cells by immunocytochemistry. Rat CPEC and HIBCPP were cultured in 12 well plates with glass coverslips for 4-5 days. Then, rat CPEC and HIBCPP cells were washed with phosphate-buffered saline (PBS) and fixed with 4% paraformaldehyde (PFA) at room temperature for 10 min. Cells were washed with PBS, permeabilized and blocked for 1 h, at room temperature, with PBS containing 0.2% Triton X-100 and 3% bovine serum albumin (BSA). Then, rat CPEC and HIBCPP cells were incubated with the primary antibodies diluted in blocking solution, rabbit anti-rat 17β-HSD (1:50, Santa Cruz Biotechnology, Inc.), aromatase (1:50, Abcam), and 5α-redutase (1:50, Santa Cruz Biotechnology, Inc.), overnight at 4ºC. Cells were washed and incubated with Alexa Fluor 488 goat anti-rabbit (1:1000, RRID AB_143165; A11008, Molecular Probes) in blocking solution at room temperature for 1 h. After several washes, nuclei were stained with Hoechst 33342 (1:1000, I34406, Molecular Probes) for 10 min at room temperature, washed once and then the coverslips were placed on glass slides using fluorescence mounting medium (Dako). Cell preparations were examined on a AxioImager Z2 microscope (Carl Zeiss, Germany) or a LSM710 confocal laser scanning microscope (Carl Zeiss, Germany) using a magnification of 40x or 63x.
2.5. Immunohistochemistry
Protein expression of 17β-HSD, P450 aromatase, and 5α-reductase was analyzed in both rat and human (men and women) CP paraffin-embedded slices. Human post-mortem CP tissue was obtained from the Institute of Neuropathology Brain Bank (HUB-ICO-IDIBELL Biobank, Barcelona, Spain) following the ethical guidelines of the Real Decreto de Biobancos 1716/2011 (BOE 2011/18919), and the approval of the Ethics Committee of the Hospital Universitari de Bellvitge (HUB). Slices were deparaffinized, rehydrated, and unmasked by pretreatment with Trilogy™ (Cell Marque™, Sigma-Aldrich, Merck, Portugal) according to the manufacturer’s instructions. Endogenous peroxidase activity was blocked with H2O2 (3%) for 10 min. Slices were washed twice with TBS-T and incubated with the selected primary monoclonal antibody: rabbit 17β-HSD (1:100, Santa Cruz Biotechnology, Inc.), P450 aromatase (1:100, Abcam), or 5α-redutase (1:100, Santa Cruz Biotechnology, Inc.) for 1h at RT. Then, slices were washed twice with TBS-T and treated with HiDef Detection™ HRP Polymer System (Cell Marque™, Sigma-Aldrich, Merck, Portugal). After washing with TBS-T, immunoreactivity was detected with diaminobenzidine peroxidase (DAB) for 10 min. Following DAB reaction, slices were washed twice with TBS-T and counterstained with Hematoxylin for 3 min to allow nuclei visualization. Negative control slices were treated under strictly similar conditions without primary antibody. After dehydration slides were mounted and coverslipped and images were acquired using a AxioImager Z2 microscope (Carl Zeiss, Germany) using a magnification of 20x.
3. Results
3.1. Gene and Protein Expression of Key Enzymes Involved in the Androgenic Pathway Are Found in Rat Choroid Plexus
The mRNA expression of enzymes involved in steroid hormones biosynthetic pathway in rat CP: P450scc (Cyp11A1), aromatase (Cyp19A1), 17βHSD3 (HSD17B3), 17βHSD8 (HSD17B8), 5α-Reductase 1 (Srd5a1), and 5α-Reductase 2 (Srd5a2) was investigated by RT-PCR analyses using specific primers. Transcripts of all the steroid hormones Cyp11A1 (125 bps), Cyp19A1 (150 bps), HSD17B3 (278 bps), HSD17B8 (167 bps), Srd5a1 (194 bps), and Srd5a2 (192 bps) were obtained as shown in Figure 1A. Sanger sequencing was used to confirm the transcripts’ identity.
The detection of protein is also reported in Figure 1B. The expression and localization of 17βHSD, Aromatase, and 5α-Reductase were evaluated in primary rat CPEC by immunocytochemistry. 17βHSD showed a cytoplasmatic distribution (Figure 1B). The same pattern of expression and localization was identified for 5α-Reductase (Figure 1B). For aromatase, a perinuclear staining was observed.
The rat CP was positively immunostained for 17βHSD, Aromatase, and 5α-Reductase (Figure 1C). Aromatase was expressed in the cytoplasm and a very few nuclei of the CP epithelial cells (Figure 1C). Although we did not perform a quantitative analysis of Aromatase expression we observed a very low cytoplasmic staining intensity compared with the other steroidogenic enzymes examined. 17βHSD and 5α-Reductase (Figure 1C) enzymes also showed a predominantly positive cytoplasmic expression. In both enzymes, the staining was relatively consistent and of moderate intensity. There were a few labeled nuclei but with very intense staining. Figure 1C also shows the negative control without the primary antibody confirming the signal specificity for the enzymes detected.
3.2. Gene and Protein Expression of Key Enzymes Involved in the Androgenic Pathway Are Found in the Human CP
The expression of steroidogenic enzymes of the androgenic pathway was also analyzed in the human CP cell line HIBCPP by RT-PCR. Figure 2A shows the mRNA transcripts corresponding to HSD17B3 (125 bps), HSD17B8 (119 bps), and SRD5A1 (139 bps).
Evaluation of the protein expression and localization of steroidogenic enzymes were also performed in HIBCPP cells. Expression of 17βHSD, aromatase, and SRD5A1 was observed in the cells cytoplasm and also on the cell membrane for aromatase and SRD5A1 (Figure 2B). Protein expression of these three steroidogenic enzymes was also studied in human CP samples from both men and women (Figure 2C). A positive immunostaining for 17βHSD, Aromatase, and 5α-Reductase was obtained in the CP samples from men and women with results showing a distribution of the steroidogenic enzymes across the cytoplasm and nuclei of CPEC. For 17βHSD more intense staining is observed in comparison to aromatase and SRD5A1, with the latter showing the lower intensity staining (Figure 2C).
4. Discussion
At the interface between the periphery and the CNS, the CP is particularly well positioned to monitor and respond to changes in the CNS extracellular fluid, and in the peripheral circulation, controlling the biochemistry of the brain, under both normal and pathological conditions [6,9]. The CP is also a source of polypeptides to the CNS [9], playing an integral role in autocrine/paracrine and distal/endocrine effects on target cells in the brain [6]. The CP is a target organ for steroid hormones that expresses androgen, estrogen and progesterone receptors [41,42] and we gathered evidences that many peptides produced in the CP implicated in neuroprotection and neurodegeneration can be altered in response to steroid hormones [43,44,45]. Brain sex steroids are derived from both peripheral and local (neurosteroids) sources and are crucial for normal brain function [46]. The CP, being a robust producer of compounds that are vital for CNS, the presence of key elements of the steroid hormone biosynthetic pathway is an essential step to enhance our understanding of how CP controls CNS changes. To our knowledge this study confirms and provides evidence of steroidogenic activity in the CP, alike what is observed in several brain regions well-equipped with enzymes necessary for steroid hormone biosynthesis where extensive steroid metabolism occurs [15]. The expression of the steroidogenic enzymes in rat brains is cell-type specific and developmentally regulated, indicating a complex synthesis pathway [47]. For instance, P450scc gene is expressed at particularly high levels in the cerebral cortex and, to a lesser extent, in the amygdala, hippocampus, and midbrain of both female and male rats [14,48], whereas 17βHSD is widely distributed in ependymocytes and astrocytes of the hippocampus, cerebral cortex, thalamus, and hypothalamus [48]. In the rat brain, 5α-reductase immunoreactivity has been observed in astrocytes, ependymocytes, and tanycytes within various brain regions including the hypothalamus, thalamus, hippocampus, cerebral cortex, and circumventricular organs [14,49], and significant levels of aromatase have been shown in amygdaloid structures and supraoptic nucleus [50].
Previous works have shown the expression of P450scc, aromatase, 5α-reductase 1, 17βHSD3 [37], and 17βHSD8 [48] in rat CP. Additionally, the expression of P450c17(CYP17A1), 17βHSD3, and 3βHSD1 (HSD3B1) was also detected in the CP of rodents CP [37]. 17βHSD3 and 5α-Reductase 1 mRNA were identified in human CP samples where the researchers found that the gene expression of 3βHSD2, aromatase, CYP11B1, and AKR1D1 were negligible or undetectable [38,51].
In the present study mRNAs encoding the key enzymes that are locally involved in the synthesis of neurosteroids, namely P450scc (Cyp11A1), P450aro (Cyp19A1), 17βHSD8 (HSD17B8), 17βHSD3 (HSD17B3), 5α-Reductase 1 (Srd5A1), and 5α-Reductase 2 (Srd5A2), were detected in rat CP, supporting the hypothesis of the novo production of neurosteroids in a small and complex organ.
The localization of steroidogenic enzymes in rat CP has also been examined using immunohistochemistry (IHC) and immunocytochemistry (ICC). Aromatase, which catalyzes the conversion of the C19 androgens, androstenedione, and testosterone, to the C18 estrogens, estrone, and estradiol, respectively, is localized in the endoplasmic reticulum [52]. Our data from rat CPEC may corroborate the presence of P450aro in the endoplasmic reticulum, since we observed a perinuclear localization, suggesting that CP might be able to convert estradiol from its precursors. Many of the regions where P450aro is expressed are also areas in which ER isoforms are expressed; for example, ERα and β are expressed in the hypothalamus and hippocampus [53]. Immunohistochemical studies have demonstrated the presence of ER α and β in CP epithelial cells in both cytosolic and nuclear compartments [41]. In steroid metabolism, 17betaHSD catalyzes the final steps in androgen and estrogen biosynthesis and is bound to the endoplasmic reticulum [52]. Indeed, our study demonstrated that in CPEC most of the immunoreactive material is present in the nucleus and cytoplasm but, particularly in rat primary CP epithelial cells, a more intense staining in the perinuclear region was observed in a few cells which may suggest an association with endoplasmic reticulum. These observations are in agreement with the results from Pelletier et al., who demonstrated the presence of the 17betaHSD in the cytoplasm of glial cells [54]. This non-P450 enzyme is also known to bind to the endoplasmic reticulum [47]. Moreover, it was also demonstrated that in human placentas 17betaHSD immunoreactivity appears in both cytoplasm and nucleus, making the staining more intense in nuclei [55]. Testosterone is converted to the more potent androgen DHT by the enzyme 5α-reductase [52]. At the ultrastructural level, 5α-reductase appeared to be diffusely distributed throughout the cytoplasm of the glial and ependymal cells [48]. Indeed, our results also demonstrated a diffuse distribution of 5α-reductase in the cytoplasm of CPEC, with a particular more intense staining around the nucleus in primary rat CPEC that may be consistent with endoplasmic reticulum distribution. In addition, we also observed an expression in the nucleus of these cells, both in IHC and ICC. The same distinct localization was recently reported in human prostate cancer, showing a nuclear and cytoplasmic expression of 5α-reductase 1 [56]. These observations are in agreement with subcellular fractionation studies which had shown that 5α-reductase activity was mostly found in the microsomal fraction with some activity in the nuclear fraction [57]. It thus appears that the enzyme activity in CP could be restricted to specific organelles, such as endoplasmic reticulum, and that the neurosteroids synthesized in the CP might have free access to the plasma membrane to be released into the CSF.
In summary, in the present study, we have shown the expression of steroidogenic enzymes needed for the synthesis of neurosteroids in rat and human CP. These findings are important in the way that they provide evidence that the CP is a possible site of sex hormone synthesis.
5. Conclusions
A decrease in brain androgen availability has been associated with several senescent effects, as demonstrated by disturbances in mood, cognition, and libido in aging males [58]. As androgens, estrogens may regulate several functions affected by aging, like mood, sleep, and cognition [59].
Although the biosynthesis of neurosteroids had already been demonstrated in the brain, our data confirms and corroborates the presence of local machinery for the synthesis of neurosteroids in rat and human CP. Understanding the contribution of CP neurosteroid biosynthesis to brain homeostasis may become a stimulus to develop new targets or drugs that act through the actions of neurosteroids and could be implicated in brain neuroprotection.
Author Contributions
Conceptualization, CS, TQ, and ACD; methodology, TQ, ACD, HI, CS, HS, IF, and CS; formal analysis, CS.; investigation, TQ, ACD, FP, TA, CLP, and CS; resources, CS.; writing—original draft preparation, TQ, ACD, and RM; writing—review and editing, CS, HI, CS, HS, IF, and IG; supervision, CS; funding acquisition, EC and CS. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Portuguese Foundation for Science and Technology (FCT, Portugal—http://www.fct.pt) project grants (PTDC/SAU-NEU/114800/2009; 10.54499/UIDB/00709/2020; 10.54499/UIDP/00709/2020), COMPETE (PEst-C/ SAU/UI0709/2011). Telma Quintela received a FCT fellowship (SFRH/BPD/70781/2010).
Institutional Review Board Statement
All animal experiments were performed in compliance with the NIH guidelines, and the National and European Union rules for the care and handling of laboratory animals (Directive 2010/63/EU). Human post-mortem CP tissue was obtained from the Institute of Neuropathology Brain Bank (HUB-ICO-IDIBELL Biobank, Barcelona, Spain) following the ethical guidelines of the Real Decreto de Biobancos 1716/2011 (BOE 2011/18919), and the approval of the Ethics Committee of the Hospital Universitari de Bellvitge (HUB).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Redzic, Z.B.; Segal, M.B. The Structure of the Choroid Plexus and the Physiology of the Choroid Plexus Epithelium. Adv Drug Deliv Rev 2004, 56, 1695–1716. [Google Scholar] [CrossRef] [PubMed]
- Santos, C.R.A.; Duarte, A.C.; Quintela, T.; Tomás, J.; Albuquerque, T.; Marques, F.; Palha, J.A.; Gonçalves, I. The Choroid Plexus as a Sex Hormone Target: Functional Implications. Front Neuroendocrinol 2017, 44, 103–121. [Google Scholar] [CrossRef]
- Praetorius, J.; Damkier, H.H. Transport across the Choroid Plexus Epithelium. Am J Physiol Cell Physiol 2017, 312, C673–C686. [Google Scholar] [CrossRef] [PubMed]
- Spector, R.; Robert Snodgrass, S.; Johanson, C.E. A Balanced View of the Cerebrospinal Fluid Composition and Functions: Focus on Adult Humans. Exp Neurol 2015, 273, 57–68. [Google Scholar] [CrossRef] [PubMed]
- Vargas, T.; Ugalde, C.; Spuch, C.; Antequera, D.; Morán, M.J.; Martín, M.A.; Ferrer, I.; Bermejo-Pareja, F.; Carro, E. Aβ Accumulation in Choroid Plexus Is Associated with Mitochondrial-Induced Apoptosis. Neurobiol Aging 2010, 31, 1569–1581. [Google Scholar] [CrossRef]
- Emerich, D.F.; Skinner, S.J.M.; Borlongan, C. V.; Vasconcellos, A. V.; Thanos, C.G. The Choroid Plexus in the Rise, Fall and Repair of the Brain. BioEssays 2005, 27, 262–274. [Google Scholar] [CrossRef]
- Speake, T.; Whitwell, C.; Kajita, H.; Majid, A.; Brown, P.D. Mechanisms of CSF Secretion by the Choroid Plexus. Microsc Res Tech 2001, 52, 49–59. [Google Scholar] [CrossRef]
- Skipor, J.; Thiery, J. The Choroid Plexus Cerebrospinal Fluid System.Pdf. Acta Neurobiol Exp (Wars) 2008, 414–428. [Google Scholar] [CrossRef]
- Falcão, A.M.; Marques, F.; Novais, A.; Sousa, N.; Palha, J.A.; Sousa, J.C. The Path from the Choroid Plexus to the Subventricular Zone: Go with the Flow! Front Cell Neurosci 2012, 6, 1–8. [Google Scholar] [CrossRef]
- Johanson, C.; Stopa, E.; Mcmillan, P.; Roth, D.; Funk, J.; Krinke, G. The Distributional Nexus of Choroid Plexus to Cerebrospinal Fluid, Ependyma and Brain:Toxicologic/Pathologic Phenomena, Periventricular Destabilization, and Lesion Spread. Toxicol Pathol 2011, 39, 186–212. [Google Scholar] [CrossRef]
- Chodobski, A.; Szmydynger-Chodobska, J. Choroid Plexus: Target for Polypeptides and Site of Their Synthesis. Microsc Res Tech 2001, 52, 65–82. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Chen, J.; Chopp, M. Cell Proliferation and Differentiation from Ependymal, Subependymal and Choroid Plexus Cells in Response to Stroke in Rats. J Neurol Sci 2002, 193, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Emerich, D.F.; Borlongan, C. V. Potential of Choroid Plexus Epithelial Cell Grafts for Neuroprotection in Huntington’s Disease: What Remains before Considering Clinical Trials. Neurotox Res 2009, 15, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Mensah-Nyagan, A.G.; Do-Rego, J.-L.; Beaujean, D.; Luu-The, V.; Pelletier, G.; Vaudry, H. Neurosteroids: Expression of Steroidogenic Enzymes and Regulation of Steroid Biosynthesis in the Central Nervous System. Pharmacol Rev 1999, 51, 63–81. [Google Scholar] [CrossRef]
- Stoffel-Wagner, B. Neurosteroid Metabolism in the Human Brain. Eur J Endocrinol 2001, 145, 669–679. [Google Scholar] [CrossRef]
- Miller, W.L.; Auchus, R.J. The Molecular Biology, Biochemistry, and Physiology of Human Steroidogenesis and Its Disorders. Endocr Rev 2011, 32, 81–151. [Google Scholar] [CrossRef]
- Chakraborty, S.; Pramanik, J.; Mahata, B. Revisiting Steroidogenesis and Its Role in Immune Regulation with the Advanced Tools and Technologies. Genes Immun 2021, 22. [Google Scholar] [CrossRef]
- Andreoli, M.F.; Stoker, C.; Rossetti, M.F.; Lazzarino, G.P.; Luque, E.H.; Ramos, J.G. Dietary Withdrawal of Phytoestrogens Resulted in Higher Gene Expression of 3-Beta-HSD and ARO but Lower 5-Alpha-R-1 in Male Rats. Nutrition Research 2016, 36. [Google Scholar] [CrossRef]
- Azcoitia, I.; Mendez, P.; Garcia-Segura, L.M. Aromatase in the Human Brain. Androgens 2021, 2. [Google Scholar] [CrossRef]
- Castelli, M.P.; Casti, A.; Casu, A.; Frau, R.; Bortolato, M.; Spiga, S.; Ennas, M.G. Regional Distribution of 5α-Reductase Type 2 in the Adult Rat Brain: An Immunohistochemical Analysis. Psychoneuroendocrinology 2013, 38. [Google Scholar] [CrossRef]
- Giatti, S.; Diviccaro, S.; Garcia-Segura, L.M.; Melcangi, R.C. Sex Differences in the Brain Expression of Steroidogenic Molecules under Basal Conditions and after Gonadectomy. J Neuroendocrinol 2019, 31. [Google Scholar] [CrossRef] [PubMed]
- Hojo, Y.; Murakami, G.; Mukai, H.; Higo, S.; Hatanaka, Y.; Ogiue-Ikeda, M.; Ishii, H.; Kimoto, T.; Kawato, S. Estrogen Synthesis in the Brain-Role in Synaptic Plasticity and Memory. Mol Cell Endocrinol 2008, 290. [Google Scholar] [CrossRef] [PubMed]
- Kimoto, T.; Ishii, H.; Higo, S.; Hojo, Y.; Kawato, S. Semicomprehensive Analysis of the Postnatal Age-Related Changes in the MRNA Expression of Sex Steroidogenic Enzymes and Sex Steroid Receptors in the Male Rat Hippocampus. Endocrinology 2010, 151. [Google Scholar] [CrossRef]
- Kiyokage, E.; Toida, K.; Suzuki-Yamamoto, T.; Ishimura, K. Cellular Localization of 5α-Reductase in the Rat Cerebellum. J Chem Neuroanat 2014, 59–60, 59–60. [Google Scholar] [CrossRef]
- Leicaj, M.L.; Pasquini, L.A.; Lima, A.; Gonzalez Deniselle, M.C.; Pasquini, J.M.; De Nicola, A.F.; Garay, L.I. Changes in Neurosteroidogenesis during Demyelination and Remyelination in Cuprizone-Treated Mice. J Neuroendocrinol 2018, 30. [Google Scholar] [CrossRef]
- Mukai, H.; Tsurugizawa, T.; Ogiue-Ikeda, M.; Murakami, G.; Hojo, Y.; Ishii, H.; Kimoto, T.; Kawato, S. Local Neurosteroid Production in the Hippocampus: Influence on Synaptic Plasticity of Memory. In Proceedings of the Neuroendocrinology, 2007; 84. [Google Scholar]
- Ohno, S.; Nishikawa, K.; Honda, Y.; Nakajin, S. Expression in E. Coli and Tissue Distribution of the Human Homologue of the Mouse Ke 6 Gene, 17β-Hydroxysteroid Dehydrogenase Type 8. Mol Cell Biochem 2008, 309, 209–215. [Google Scholar] [CrossRef]
- Schiffer, L.; Barnard, L.; Baranowski, E.S.; Gilligan, L.C.; Taylor, A.E.; Arlt, W.; Shackleton, C.H.L.; Storbeck, K.H. Human Steroid Biosynthesis, Metabolism and Excretion Are Differentially Reflected by Serum and Urine Steroid Metabolomes: A Comprehensive Review. Journal of Steroid Biochemistry and Molecular Biology 2019, 194. [Google Scholar] [CrossRef]
- Tsai-Morris, C.H.; Khanum, A.; Tang, P.-Z.; Dufau, M.L. The Rat 17β-Hydroxysteroid Dehydrogenase Type III: Molecular Cloning and Gonadotropin Regulation. Endocrinology 1999, 140, 3534–3542. [Google Scholar] [CrossRef]
- Moffat, S.D. Effects of Testosterone on Cognitive and Brain Aging in Elderly Men. Ann N Y Acad Sci 2005, 1055, 80–92. [Google Scholar] [CrossRef]
- Brann, D.W.; Lu, Y.; Wang, J.; Zhang, Q.; Thakkar, R.; Sareddy, G.R.; Pratap, U.P.; Tekmal, R.R.; Vadlamudi, R.K. Brain-Derived Estrogen and Neural Function. Neurosci Biobehav Rev 2022, 132. [Google Scholar] [CrossRef]
- Krolick, K.N.; Zhu, Q.; Shi, H. Effects of Estrogens on Central Nervous System Neurotransmission: Implications for Sex Differences in Mental Disorders. Progress in Molecular Biology and Translational Science; 2018; 160. [Google Scholar]
- Bustamante-Barrientos, F.A.; Méndez-Ruette, M.; Ortloff, A.; Luz-Crawford, P.; Rivera, F.J.; Figueroa, C.D.; Molina, L.; Bátiz, L.F. The Impact of Estrogen and Estrogen-Like Molecules in Neurogenesis and Neurodegeneration: Beneficial or Harmful? Front Cell Neurosci 2021, 15. [Google Scholar] [CrossRef] [PubMed]
- Aspesi, D.; Cornil, C.A. Role of Neuroestrogens in the Regulation of Social Behaviors—From Social Recognition to Mating. Neurosci Biobehav Rev 2024, 161. [Google Scholar] [CrossRef] [PubMed]
- Luine, V.; Frankfurt, M. Estrogenic Regulation of Memory: The First 50 Years. Horm Behav 2020, 121. [Google Scholar] [CrossRef] [PubMed]
- Quintela, T.; Gonçalves, I.; Carreto, L.C.; Santos, M.A.S.; Marcelino, H.; Patriarca, F.M.; Santos, C.R.A. Analysis of the Effects of Sex Hormone Background on the Rat Choroid Plexus Transcriptome by CDNA Microarrays. PLoS ONE 2013, 8. [Google Scholar] [CrossRef]
- Ono, K.; Yano, A.; Ishimura, K. Effect of Peripherally Derived Steroid Hormones on the Expression of Steroidogenic Enzymes in the Rat Choroid Plexus. The Journal of Medical Investigation 2021, 68, 238–243. [Google Scholar] [CrossRef]
- O’Reilly, M.W.; Westgate, C.S.J.; Hornby, C.; Botfield, H.; Taylor, A.E.; Markey, K.; Mitchell, J.L.; Scotton, W.J.; Mollan, S.P.; Yiangou, A.; et al. A Unique Androgen Excess Signature in Idiopathic Intracranial Hypertension Is Linked to Cerebrospinal Fluid Dynamics. JCI Insight 2019, 4. [Google Scholar] [CrossRef]
- Gonçalves, I.; Quintela, T.; Duarte, A.C.; Hubbard, P.; Schwerk, C.; Belin, A.C.; Toma, J.; Santos, R.A. Experimental Tools to Study the Regulation and Function of the Choroid Plexus. In Blood-Brain Barrier. Neuromethods; Barichello, T., Ed.; Humana Press: New York, NY, USA, 2018; Volume 142, ISBN 9781493989461. [Google Scholar]
- Ishiwata, I.; Ishiwata, C.; Ishiwata, E.; Sato, Y.; Kiguchi, K.; Tachibana, T.; Hashimoto, H.; Ishikawa, H. Establishment and Characterization of a Human Malignant Choroids Plexus Papilloma Cell Line (HIBCPP). Hum Cell 2005, 18, 67–72. [Google Scholar] [CrossRef]
- Hong-Goka, B.C.; Chang, F.-L.F. Estrogen Receptors α and β in Choroid Plexus Epithelial Cells in Alzheimer’s Disease. Neurosci Lett 2004, 360, 113–116. [Google Scholar] [CrossRef]
- Alves, C.H.; Gonçalves, I.; Socorro, S.; Baltazar, G.; Quintela, T.; Santos, C.R.A. Androgen Receptor Is Expressed in Murine Choroid Plexus and Downregulated by 5α-Dihydrotestosterone in Male and Female Mice. Journal of Molecular Neuroscience 2009, 38, 41–49. [Google Scholar] [CrossRef]
- Quintela, T.; Alves, C.H.; Goncalves, I.; Baltazar, G.; Saraiva, M.J.; Santos, C.R. 5Alpha-Dihydrotestosterone up-Regulates Transthyretin Levels in Mice and Rat Choroid Plexus via an Androgen Receptor Independent Pathway. Brain Res 2008, 1229, 18–26. [Google Scholar] [CrossRef]
- Quintela, T.; Goncalves, I.; Baltazar, G.; Alves, C.H.; Saraiva, M.J.; Santos, C.R. 17beta-Estradiol Induces Transthyretin Expression in Murine Choroid Plexus via an Oestrogen Receptor Dependent Pathway. Cell Mol Neurobiol 2009, 29, 475–483. [Google Scholar] [CrossRef] [PubMed]
- Quintela, T.; Goncalves, I.; Martinho, A.; Alves, C.H.; Saraiva, M.J.; Rocha, P.; Santos, C.R. Progesterone Enhances Transthyretin Expression in the Rat Choroid Plexus in Vitro and in Vivo via Progesterone Receptor. J Mol Neurosci 2011, 44, 152–158. [Google Scholar] [CrossRef] [PubMed]
- Bates, K.A.; Harvey, A.R.; Carruthers, M.; Martins, R.N. Androgens, Andropause and Neurodegeneration: Exploring the Link between Steroidogenesis, Androgens and Alzheimer’s Disease. Cellular and Molecular Life Sciences 2005, 62, 281–292. [Google Scholar] [CrossRef] [PubMed]
- Mellon, S.H.; Vaudry, H.B.T.-I.R. of N. Biosynthesis of Neurosteroids and Regulation of Their Sysnthesis. In Neurosteroids and Brain Function; Academic Press, 2001; Vol. 46, pp. 33–78 ISBN 0074-7742.
- Pelletier, G. Chapter 11—Steroidogenic Enzymes in the Brain: Morphological Aspects. In Neuroendocrinology: The Normal Neuroendocrine System; Martini, L.B.T.-P., Ed.; Elsevier, 2010; Volume 181, pp. 193–207. ISBN 0079-6123. [Google Scholar]
- Pelletier, G.; Luu-The, V.; Labrie, F. Immunocytochemical Localization of 5α-Reductase in Rat Brain. Molecular and Cellular Neuroscience 1994, 5, 394–399. [Google Scholar] [CrossRef]
- Hojo, Y.; Hattori, T.; Enami, T.; Furukawa, A.; Suzuki, K.; Ishii, H.; Mukai, H.; Morrison, J.H.; Janssen, W.G.M.; Kominami, S.; et al. Adult Male Rat Hippocampus Synthesizes Estradiol from Pregnenolone by Cytochromes P45017α and P450 Aromatase Localized in Neurons. Proceedings of the National Academy of Sciences 2004, 101, 865–870. [Google Scholar] [CrossRef]
- O’Reilly, M.W.; Westgate, C.S.J.; Hornby, C.; Botfield, H.; Taylor, A.E.; Markey, K.; Mitchell, J.L.; Scotton, W.J.; Mollan, S.P.; Yiangou, A.; et al. A Unique Androgen Excess Signature in Idiopathic Intracranial Hypertension Is Linked to Cerebrospinal Fluid Dynamics. JCI Insight 2019, 4. [Google Scholar] [CrossRef]
- Compagnone, N.A.; Mellon, S.H. Neurosteroids: Biosynthesis and Function of These Novel Neuromodulators. Front Neuroendocrinol 2000, 21, 1–56. [Google Scholar] [CrossRef]
- Simpson, E.R.; Clyne, C.; Rubin, G.; Wah, C.B.; Robertson, K.; Britt, K.; Speed, C.; Jones, M. Aromatase—A Brief Overview. Annu Rev Physiol 2002, 64, 93–127. [Google Scholar] [CrossRef]
- Pelletier, G.; Luu-The, V.; Labrie, F. Immunocytochemical Localization of Type I 17β-Hydroxysteroid Dehydrogenase in the Rat Brain. Brain Res 1995, 704, 233–239. [Google Scholar] [CrossRef]
- Dupont, E; Labrie, F; Luu-The, V; Pelletier, G Localization of 17 Beta-Hydroxysteroid Dehydrogenase throughout Gestation in Human Placenta. Journal of Histochemistry & Cytochemistry 1991, 39, 1403–1407. [CrossRef]
- Das, K.; Lorena, P.D.N.; Ng, L.K.; Lim, D.; Shen, L.; Siow, W.Y.; Teh, M.; Reichardt, J.K. V; Salto-Tellez, M. Differential Expression of Steroid 5α-Reductase Isozymes and Association with Disease Severity and Angiogenic Genes Predict Their Biological Role in Prostate Cancer. Endocr Relat Cancer 2010, 17, 757–770. [Google Scholar] [CrossRef]
- Lephart, E.D. Brain 5α-Reductase: Cellular, Enzymatic, and Molecular Perspectives and Implications for Biological Function. Molecular and Cellular Neuroscience 1993, 4, 473–484. [Google Scholar] [CrossRef] [PubMed]
- Pike, C.J.; Nguyen, T.-V. V; Ramsden, M.; Yao, M.; Murphy, M.P.; Rosario, E.R. Androgen Cell Signaling Pathways Involved in Neuroprotective Actions. Horm Behav 2008, 53, 693–705. [Google Scholar] [CrossRef] [PubMed]
- Russell, J.K.; Jones, C.K.; Newhouse, P.A. The Role of Estrogen in Brain and Cognitive Aging. Neurotherapeutics 2019, 16. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Analysis of the expression of steroidogenic enzymes required for androgen biosynthesis in rat CP. A. RT-PCR analysis of mRNA expression of steroidogenic enzymes. Transcripts were found for steroidogenic enzymes Cyp11A1, HSD17b3, HSD17b8, Srd5a1, Srd5a2, and Cyp19A1. (-) sample without template DNA as negative control. B. Immunofluorescence detection of 17β-hydroxysteroid dehydrogenase (17βHSD), 5α-Reductase (Srda1), and Aromatase in primary rat CP epithelial cells (green). Nuclei were stained with Hoechst 33423 (blue). Scale bar 20 µm. C. Immunohistochemical localization of steroidogenic enzymes in rat CP. Tissue sections showing immunopositive 17βHSD, 5α-Reductase (Srda1), and Aromatase. Immunohistochemistry was performed using DAB and Hematoxylin. (-) negative control. Scale bar 26 µm.
Figure 1.
Analysis of the expression of steroidogenic enzymes required for androgen biosynthesis in rat CP. A. RT-PCR analysis of mRNA expression of steroidogenic enzymes. Transcripts were found for steroidogenic enzymes Cyp11A1, HSD17b3, HSD17b8, Srd5a1, Srd5a2, and Cyp19A1. (-) sample without template DNA as negative control. B. Immunofluorescence detection of 17β-hydroxysteroid dehydrogenase (17βHSD), 5α-Reductase (Srda1), and Aromatase in primary rat CP epithelial cells (green). Nuclei were stained with Hoechst 33423 (blue). Scale bar 20 µm. C. Immunohistochemical localization of steroidogenic enzymes in rat CP. Tissue sections showing immunopositive 17βHSD, 5α-Reductase (Srda1), and Aromatase. Immunohistochemistry was performed using DAB and Hematoxylin. (-) negative control. Scale bar 26 µm.
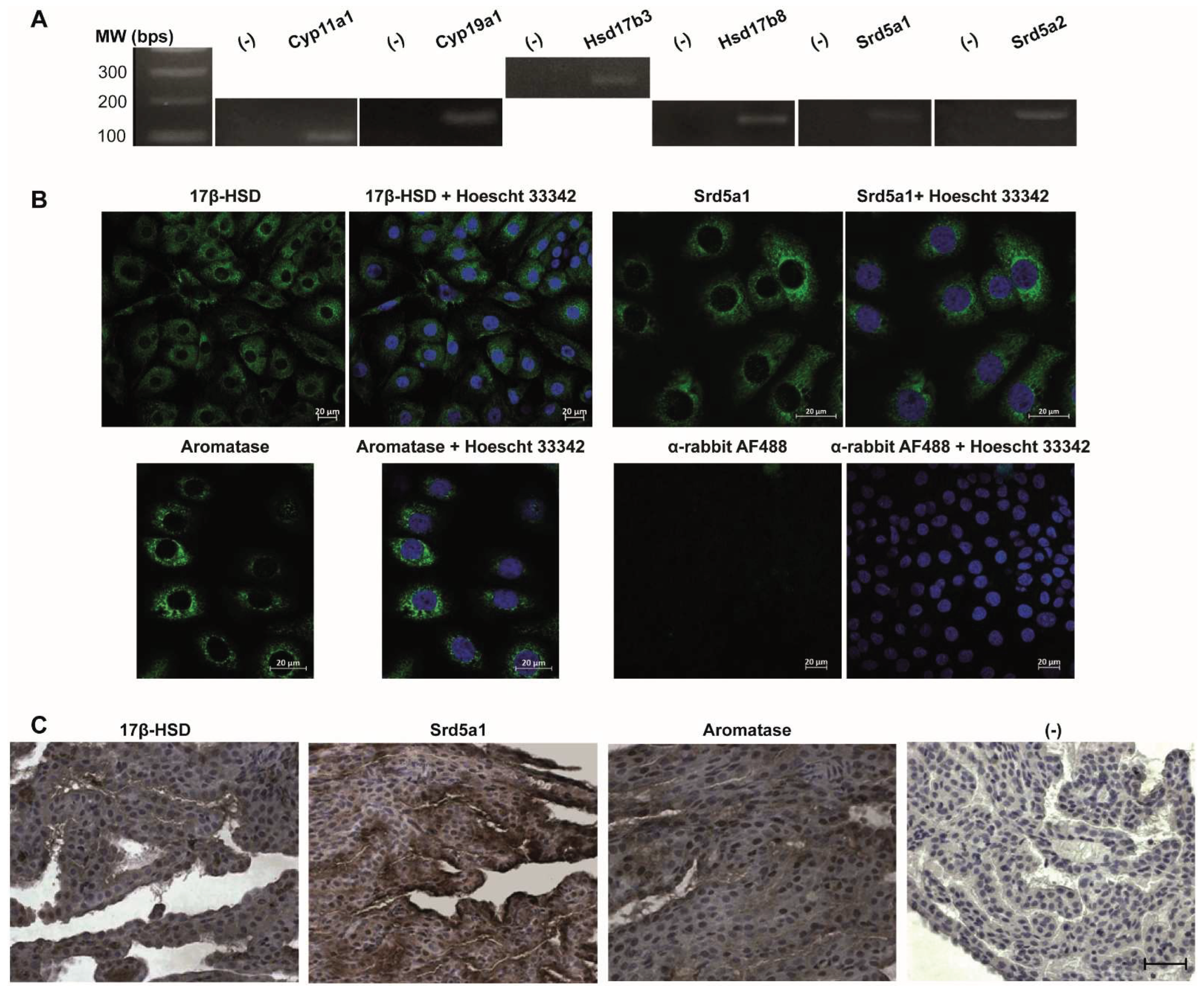
Figure 2.
Analysis of the expression of steroidogenic enzymes required for androgen biosynthesis in human CP. A. RT-PCR analysis of mRNAs from steroidogenic enzymes in HIBCPP cells. Representative RT-PCR analysis for steroidogenic enzymes HSD17b3, HSD17b8, and SRD5A1. (-) sample without template DNA as negative control. B. Immunofluorescence detection of steroidogenesis enzymes in HIBCPP cells. Microscopical images obtained with an Axio Imager Z2 of the steroidogenesis enzymes 17β-hydroxysteroid dehydrogenase (17β-HSD), 5α-Reductase (SRD5A1), and Aromatase expression in HIBCPP cells (green). Nuclei were stained with Hoechst 33423 (blue). Scale bar 20 µm. C. Representative images showing the immunolocalization of steroidogenesis enzymes in human CP slices of men and women. The enzymes 17β-hydroxysteroid dehydrogenase (17β-HSD), 5α-Reductase (SRD5A1), and Aromatase were detected in human CP. Immunohistochemistry was performed using DAB and Hematoxylin. (-) negative control. Scale bar 50 µm.
Figure 2.
Analysis of the expression of steroidogenic enzymes required for androgen biosynthesis in human CP. A. RT-PCR analysis of mRNAs from steroidogenic enzymes in HIBCPP cells. Representative RT-PCR analysis for steroidogenic enzymes HSD17b3, HSD17b8, and SRD5A1. (-) sample without template DNA as negative control. B. Immunofluorescence detection of steroidogenesis enzymes in HIBCPP cells. Microscopical images obtained with an Axio Imager Z2 of the steroidogenesis enzymes 17β-hydroxysteroid dehydrogenase (17β-HSD), 5α-Reductase (SRD5A1), and Aromatase expression in HIBCPP cells (green). Nuclei were stained with Hoechst 33423 (blue). Scale bar 20 µm. C. Representative images showing the immunolocalization of steroidogenesis enzymes in human CP slices of men and women. The enzymes 17β-hydroxysteroid dehydrogenase (17β-HSD), 5α-Reductase (SRD5A1), and Aromatase were detected in human CP. Immunohistochemistry was performed using DAB and Hematoxylin. (-) negative control. Scale bar 50 µm.
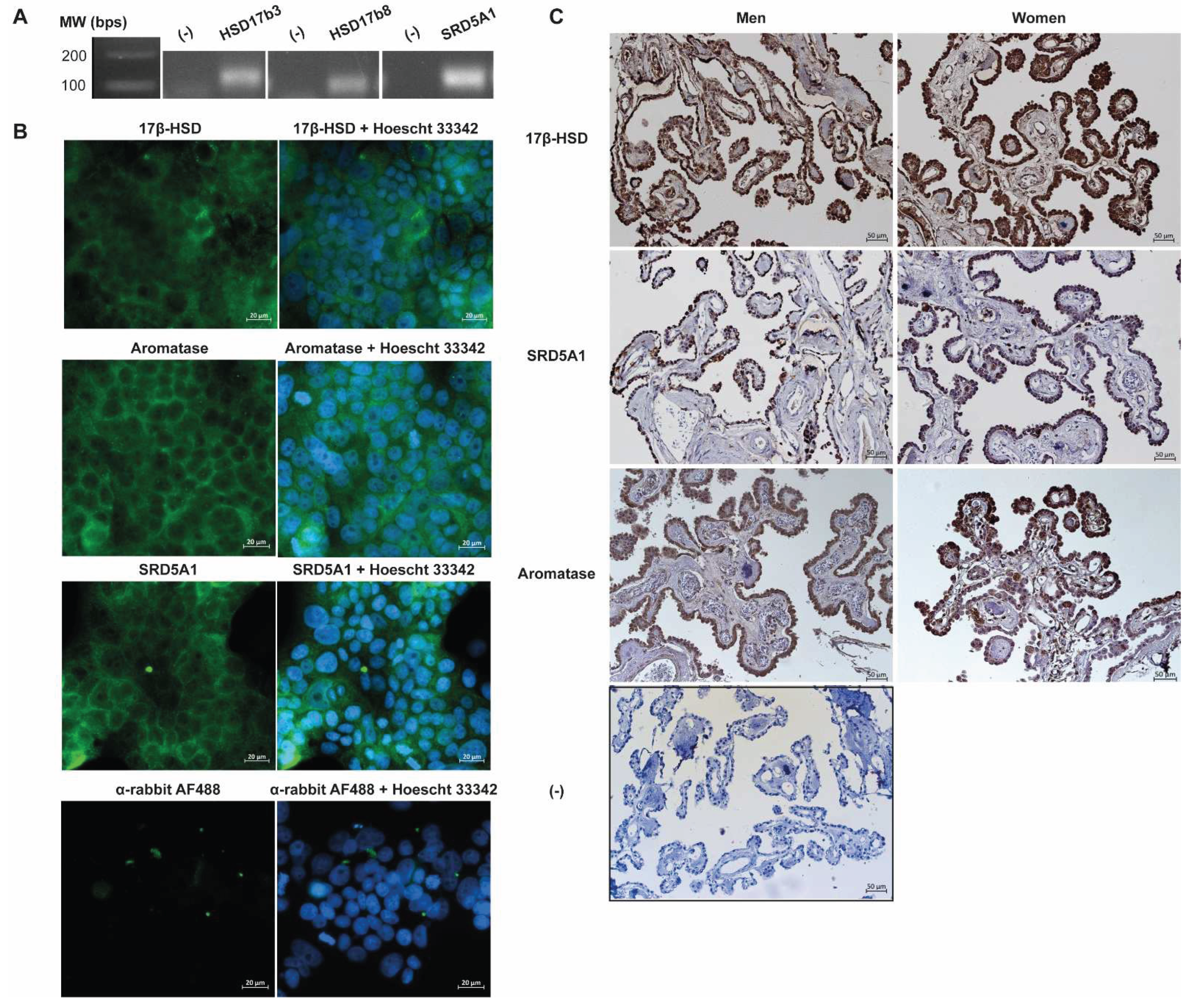

Table 1.
Primer sequences for RT-PCR.
| Gene | Primer Fw (5′-3′) | Primer Rv (5′-3′) | Amplicon size (bp) | Annealing temperature (˚C) |
|---|---|---|---|---|
| Rat | ||||
| CYP11A1 | CAAAACACCACGCACTTCC | TCAATTCTGAAGTTTTCCAGCA | 125 | 54 |
| CYP19A1 | CGTCATGTTGCTTCTCATCG | TACCGCAGGCTCTCGTTAAT | 150 | 60 |
| HSD17b3 | CCAACCTGCTCCCAAGTCAT | AGGGGTCAGCACCTGGATAA | 278 | 60 |
| HSD17b8 | TTTTTCGCCCGCCATCTGTCG | TGCAGGTGCCAGGAGCTACCA | 167 | 59 |
| Srd5a1 | TGCTCGACATGCTGGTCTAC | GGCTGCAGGACGAATGTACT | 194 | 60 |
| Srd5a2 | ATTTGTGTGGCAGAGAGAGG | TTGATTGACTGCCTGGATGG | 192 | 59 |
| Human | ||||
| CYP11A1 | CACGCTCAGTCCTGGTCAAA | GGGGATCTCATTGAAGGGGC | 134 | 58 |
| CYP19A1 | CGTCGCGACTCTAAATTGCC | AAAAAGGCCAGTGAGGAGCA | 149 | 58 |
| HSD17b3 | TTCTTGCGGTCAATGGGACA | TTTTCCAGCGTCCGGCTAAT | 125 | 58 |
| HSD17b8 | TTTTCTCGCCCACCATCTGT | AAGGTGCCCTTGAGGTTGAC | 119 | 58 |
| SRD5A1 | TACGGGCATCGGTGCTTAAT | ACACTGCACAATGGCTCAAG | 139 | 58 |
| SRD5A2 | CAGGTTCAGTGCCAGCAGAG | TCTCCGTGTGCTTCCCGTAG | 112 | 58 |
Fw-forward; Rv-reverse; P450scc (CYP11A1); aromatase (CYP19A1); type 3 and type 8 17β-hydroxysteroid dehydrogenase (HSD17b3 and HSD17b8); 1 and 2 5α-reductase (Srd5a1/SRD5a1 and Srd5a2/SRD5A2).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



