
Submitted:
23 May 2025
Posted:
26 May 2025
You are already at the latest version
Abstract
Constipation, a widespread gastrointestinal disorder, imposes significant burdens on healthcare systems and global health-related quality of life, yet current options remain suboptimal due to limited mechanistic understanding and efficacy limitations. Given the pivotal significance of the interactions between the gut microbiota and the host on governing bowel movement, we employed a multi-modal approach integrating animal experiments, ELISA, histopathology, qRT-PCR, GC-MS, and 16S rRNA metagenomics to evaluate the functional potential of Lactobacillus rhamnosus LRa05 against loperamide-induced constipation in mice. LRa05 treatment markedly alleviated constipation symptoms, as evidenced by reduced first black stool expulsion time, increased fecal moisture, and enhanced intestinal motility. Mechanistic investigations revealed that LRa05 balanced gastrointestinal regulatory peptides, and downregulated aquaporin (AQP4/AQP8) mRNA levels while activating the SCF/C-Kit signaling pathway, thereby restoring intestinal peristalsis. Furthermore, LRa05 rebalanced gut microbiota composition by enriching beneficial genera (Alloprevotella, Bacteroides, Lachnospiraceae NK4A136, and Rikenellaceae RC9), elevating short chain fatty acid (SCFA) production, and augmenting gut microecological balance. These findings demonstrated that LRa05 could mitigate constipation through a tripartite mechanism: transcriptional regulation of motility-associated genes, microbial community restructuring, and SCFA-mediated modulation of the gut-brain axis. Our study positions LRa05 as a promising multi-target probiotic candidate for constipation management.
Keywords:
Probiotic
; constipation
; gastrointestinal regulatory peptides
; gut-brain axis
; gut microbiota
1. Introduction
Constipation, a prevalent digestive disorder with a global reach, affects approximately 14% of the world's population, as reported by Grand View Research in 2024. Characterized by challenging defecation, dry stools, and infrequent bowel movements, this condition significantly diminishes the quality of life for those affected [1]. If left untreated, constipation can progress to severe complications, including various gastrointestinal disorders, an elevated risk of colorectal cancer, and other critical health issues. Chronic constipation, in the absence of effective intervention, can cause irreversible structural and functional damage to the intestinal tract, thereby increasing systemic health risks and placing a substantial burden on healthcare systems [2]. This underscores the urgent need for therapeutic strategies that target the underlying mechanisms of constipation to mitigate its cascading pathological effects.
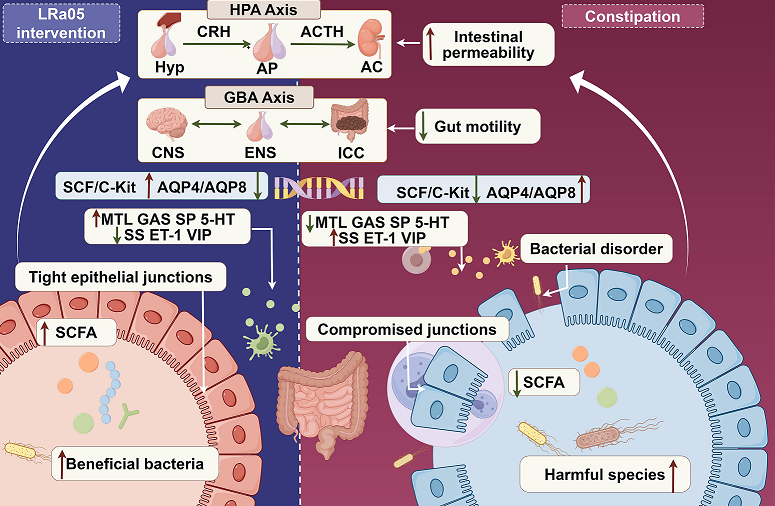
From a pathophysiological perspective, several key mechanisms contribute to the development of constipation. These include an imbalance in the intestinal microbiota, abnormal secretion of gastrointestinal hormones, dysfunction of the intestinal nervous system, and impaired intestinal motility. Disruption of the gut microbiota causes a decline in metabolite generation, which subsequently affects intestinal motility and water absorption [3]. Abnormal secretion of gastrointestinal hormones, such as motilin and substance P, weakens the contractility of the intestine [4]. Disorders within the enteric nervous system impede the regulation of intestinal movement [5]. Recent research has also revealed that abnormalities in the SCF/C-kit signaling pathway are closely associated with functional defects in interstitial cells of Cajal (ICC), which act as the "pacemaker" for intestinal motility. A decrease in the number or impaired function of ICC can significantly slow down the transit of contents through the intestine [6]. Additionally, abnormal expression of aquaporins (AQPs) in colonic epithelial cells can disrupt the transport of water in the intestine, resulting in hard stools and exacerbating constipation symptoms. Among these, AQP3, 4, 8, and 9 are most closely linked to constipation [7]. While the roles of ICC dysfunction (mediated by the SCF/C-kit signaling pathway) and aquaporin dysregulation in constipation are well-recognized, the upstream factors that trigger these dysfunctions (such as genetic predisposition and epigenetic modifications) and the downstream interactions between these pathways remain largely unclear. For example, the modulation of AQP expression in colonic epithelia by metabolites derived from the gut microbiota is still a relatively unexplored area.
Currently, the treatment of constipation predominantly relies on laxatives and gastrointestinal motility drugs. Laxatives, which typically contain chemicals that enhance the frequency of bowel peristalsis, are the most commonly prescribed medications for constipation [8]. They facilitate defecation by boosting the delivery of water and electrolytes to the intestinal mucosa and softening hardened stools [9]. However, these drugs provide only temporary relief and do not address the underlying issues such as restoring the production of short-chain fatty acids (SCFAs) or keeping the intestinal barrier intact, highlighting the necessity for multi-targeted therapeutic approaches. The limitations of conventional treatments are further emphasized by the fact that, despite laxatives dominating a market worth $4.2 billion, they are associated with a 67% recurrence rate and a 32% adverse event rate, according to statistics from the National Institutes of Health (NIH). This situation underscores the urgent need for precision nutrition-based solutions.
Recently, probiotics have arisen as a potential group of functional agents for the management of gastrointestinal disorders, particularly constipation. The global market for probiotics used in constipation treatment is increasing at an annual compound growth rate (CAGR) of 11.2%, and products with strain-specific claims command a 35-40% premium. Probiotic supplements exert their beneficial effects through multiple physiological mechanisms, including modulating the structure of the gut microbiota, promoting intestinal peristalsis, restoring the function of the mucosal barrier, and regulating the enteric nervous system [10]. Among the various probiotic species, Lactobacillus rhamnosus has gained prominence as a potent modulator of the gut microbiota, due to its excellent adhesive properties, immunomodulatory capabilities, and ability to maintain intestinal homeostasis [11]. The present study focuses on Lactobacillus rhamnosus LRa05, a strain renowned for its exceptional resistance to gastric acid and bile salts [12]. This probiotic has demonstrated significant efficacy in restructuring the gut microbial community by promoting the development of beneficial bacteria while curbing the expansion of harmful microbes [13]. Besides its microbiota-modulating effects, Lactobacillus rhamnosus LRa05 exhibits anti-inflammatory characteristics, increases SCFA levels, and modulates the expression of tight-junction proteins (ZO-1, occludin, claudin), thereby strengthening the intestinal barrier and optimizing epithelial permeability [14]. Given the vital role of the intestinal barrier in constipation pathogenesis, these findings suggest that Lactobacillus rhamnosus LRa05 holds great potential for the treatment of this condition. By integrating mechanistic understanding with commercial viability, LRa05 represents a new frontier in precision nutrition probiotics, marking a shift from merely managing symptoms to proactively intervening in the pathophysiological processes of constipation [15].
To evaluate the efficacy of Lactobacillus rhamnosus LRa05, a loperamide-induced murine model of constipation was utilized. The study comprehensively analyzed changes in intestinal motility patterns, characteristics of fecal output, and serum levels of regulatory gastrointestinal peptides. Additionally, the impact of the strain on SCFA production, the SCF/C-kit signaling pathway, and the expression of aquaporins (AQP4 and AQP8) was investigated to gain a mechanistic understanding at the molecular level in colonic tissue. Complementary 16S rRNA gene sequencing was performed to identify constipation-associated dysbiosis in the gut microbiota, providing a framework for understanding how LRa05 modulates the gut-brain axis, which acts as a foundation for its utilization in functional foods.
2. Materials and Methods
2.1. Chemicals
Lactobacillus rhamnosus LRa05 probiotic strain was supplied by Wecare Probiotics Co., Ltd. (Suzhou, China). Loperamide hydrochloride capsules (2 mg/capsule) were obtained from Xian Janssen Pharmaceutical Ltd. (Xi'an, China). Mosapride citrate tablets (5 mg/tablet) were purchased from Yabao Pharmaceutical Group Co., Ltd. (Shanghai, China). All other chemical reagents were of analytical grade (≥99% purity) unless otherwise specified.
2.2. Strain activation and bacterial suspension preparation
Lactobacillus rhamnosus LRa05 was activated and propagated in MRS broth at 37°C for 16 h. Serial subculturing was performed by inoculating 2% v/v into fresh MRS broth, with a minimum of three passages to ensure stable growth characteristics. Bacterial cells were collected by centrifugation (6,000 rpm, 12 min), washed twice with sterile saline (0.9% NaCl), and resuspended to final concentrations of 6×108 CFU/mL (low dose) and 1.2×109 CFU/mL (high dose). The bacterial suspension was prepared daily and used immediately for gavage administration during animal experiments.
2.3. Laboratory animals and intervention procedures
Male SPF-grade Balb/c mice (n=60, 20 ± 2.0 g) were sourced from SPF Biotechnology Co., Ltd. (Certification SCXK [Jing] 2019-0010) and maintained in specific pathogen-free conditions at TUST Animal Research Center (23±1℃, 12 h/12 h light cycle, ad libitum access to sterilized feed/water). Through stratified randomization after 7-day acclimatization, cohorts were established as the native control group (NC), the model group (MC), the positive control group (PC), the low-dose LRa05 (LRa05.L) group, and the high-dose LRa05 (LRa05.H) group. During the second week, constipation models were induced in all groups except NC by gavage administration of loperamide hydrochloride (10 mg/kg body weight). Starting from the third week, the MC group received daily 0.9% sterile saline (0.2 mL) via gavage, while the PC group was treated with mosapride citrate suspension at 5 mg/kg using the same administration route. For the probiotic intervention groups, LRa05.L (6×108 CFU/mL) and LRa05.H (1.2×109 CFU/mL) concentrations were calculated based on human probiotic intake guidelines (1-10×109 CFU/day). Allometric scaling principles (body surface area normalization) were applied to convert the human-equivalent doses to murine equivalents. The final formulations were adjusted to deliver 6×108 CFU (LRa05.L) and 1.2×109 CFU (LRa05.H) per 0.2 mL gavage volume for mice weighing 20-25 g, which is consistent with established rodent probiotic dosing standards [16].
The animal study protocol was approved by the Institutional Animal Care and Utilization Committee of TUST (protocol code 2023010 and date of approval March 15, 2023 ).
2.4. Sample Collection
On the day 22 (including the adaptation period), all mice were humanely euthanized. Prior to euthanasia, fresh fecal pellets were cryopreserved within 2 minutes of spontaneous defecation using liquid nitrogen immersion prior to -80°C archival (Thermo Scientific™ Forma™ 900 series). Mice underwent a 12-hour fasting period (with ad libitum water access) prior to terminal procedures. Following retro-orbital venous plexus blood collection into lithium heparin-coated vacutainers, specimens underwent coagulation at 25°C ± 0.5°C for 60 min. Serum isolation was achieved through dual-phase centrifugation (Primary: 4,000 rpm, 15 min, 4°C; Secondary: 12,000 rpm, 2 min, 4°C) using a refrigerated centrifuge (Model TDL-5-A, Xiangyi Centrifuge, Changsha, China). After blood collection, mice were euthanized by cervical dislocation under isoflurane anesthesia. Colon tissues were rapidly excised, rinsed in ice-cold PBS, and divided for histopathological evaluation (fixed in 10% neutral-buffered formalin) and RNA extraction (snap-frozen in liquid nitrogen for qPCR analysis).
2.5. Body Mass, Feeding Behavior, Fecal Transit And Intestinal Motility Assays
Body mass dynamics and ad libitum food consumption were continuously monitored using a computer-assisted monitoring system (PhenoMaster NG, TSE Systems China, Beijing, China).
On Day 21 (including the adaptation period), mice were fasted for 12 h (water ad libitum) before gavage administration of 0.2 mL activated charcoal solution (5% w/v activated charcoal+10% w/v gum arabic). Time to first black stool (min) was recorded using a digital timer. Fresh feces were collected, weighed (wet weight, W1), and dried at 105°C to constant weight (dry weight, W2) [17]. Fecal water content (%) was expressed as the following an equation:
Fecal water content (%)=[(W1-W2)/W1]×100
On Day 22 (including the adaptation period), intestinal propulsion was evaluated using a modified charcoal transit assay [18]. After 12 h fasting, mice received 0.2 mL charcoal solution. At 25 min post-gavage, animals were euthanized, and intestines excised. Total small intestine length (L1, cm) and charcoal front distance (L2, cm) were measured. Propulsion rate was expressed as the following an equation:
Propulsion rate (%)=L2/L1×100
2.6. Quantitative Profiling of Gastrointestinal Regulatory Peptides
Circulating levels of key neuroendocrine regulators, gastrin (GAS), motilin (MTL), substance P (SP), somatostatin (SS), vasoactive intestinal peptide (VIP), endothelin-1 (ET-1), and 5-hydroxytryptamine (5-HT), were quantified using high-sensitivity ELISA kits (Sigma-Aldrich(Shanghai)Trading Co.,Ltd, China). Optical density (OD) measurements were conducted at 450 nm using a SpectraMax i3x microplate reader (Molecular Devices, USA), with a reference wavelength of 630 nm to correct for nonspecific absorbance. All assays strictly adhered to the manufacturer's validated protocols, including pre-incubation of samples at 4°C overnight to optimize antigen-antibody binding. Inter-assay coefficients of variation (CVs) were maintained below 8% across all analytes, demonstrating assay reproducibility.
For quantitative analysis of enteroendocrine mediators, a standardized ELISA protocol was employed. Briefly, 50 μL serum aliquots were diluted 1:5 in assay buffer (PBS, pH 7.4) containing 0.05% Tween-20 and 1% bovine serum albumin (BSA) to minimize matrix effects. Samples were loaded in triplicate onto pre-coated 96-well plates and incubated at room temperature for 2 hours. Parallel standard curves were generated using seven-point serial dilutions of calibrators (ranging from 0.1 to 100 ng/mL), with blank controls (assay buffer alone) run in quadruplicate for background subtraction. After washing with PBS-Tween (0.05%) buffer, bound analytes were detected using HRP-conjugated secondary antibodies and developed with tetramethylbenzidine (TMB) substrate. Reactions were terminated with 2 M sulfuric acid, and OD values were immediately recorded. Data were analyzed using SoftMax Pro 7.0 software, with concentrations calculated from four-parameter logistic regression curves.
2.7. Histopathological Evaluation of Colonic Mucosa
To evaluate LRa50 probiotic's impact on intestinal barrier integrity in murine constipation models, a systematic histomorphometric analysis of distal colonic architecture was conducted. Post-euthanasia, 2 cm colonic segments underwent immediate fixation in 4% paraformaldehyde (Servicebio, Wuhan, China) for >12 h to preserve cytoarchitecture. Specimens underwent standard paraffin embedding, followed by sectioning at 4 μm (CV < 5%) using a rotary microtome (HistoCore MULTICUT, Leica Microsystems). Section flattening was achieved through controlled thermal equilibration (40-42°C water bath, 120 s) followed by adhesive mounting on pre-treated slides.
Deparaffinization was achieved through two 10-minute xylene immersions, followed by gradient ethanol rehydration (100% to 70%). Hematoxylin-eosin (H&E) staining proceeded as: 10-minute hematoxylin (Solaibao Reagent Co., Ltd) nuclear staining, 5-second differentiation with 1% HCl-ethanol, and blueing via running water. Eosin (Solaibao Reagent Co., Ltd) counterstaining (3 minutes) highlighted cytoplasmic structures. Specimens were dehydrated in graded alcohols, cleared in xylene (2 × 8 minutes), and coverslipped with a cover glass.
Slides were observed under a upright microscope (BX53, Olympus, Tokyo, Japan) at 200 × magnification. Quantitative comparisons of the mucus layer thickness between treatment and control groups were documented for statistical evaluation.
2.8. Quantitative Analysis of Fecal Short-Chain Fatty Acids (SCFAs)
Quantitative analysis of SCFAs was determined through optimized gas chromatography-mass spectrometry (GC-MS) methodology modified from established protocols [19]. Freeze-dried fecal specimens (30.0 ± 0.5 mg) underwent acidified extraction using 800 μL ultrapure water, 200 μL sulfuric acid (50% v/v, HPLC grade), and 1 mL diethyl ether (GC-MS grade). The homogenate was vortex-mixed (2500 rpm, 3 min) followed by phase separation in an ice bath (4°C, 30 min). After centrifugation (10,000 rpm, 10 min, 4°C), the organic layer was dehydrated with anhydrous sodium sulfate (0.25 g) and clarified through secondary centrifugation (10,000 rpm, 15 min). Processed extracts were transferred to amberized GC vials with PTFE-lined septa for immediate analysis on a GC-MS systerm (Thermo Scientific ISQ 7000 GC/MS, USA).
Separation was performed using a Thermo Scientific™ TG-WAXMS polar capillary column (30 m × 0.25 mm i.d., 0.25 μm film thickness). The initial oven temperature program was set at 80°C for 2 min, followed by a temperature gradient increase at 10°C/min to 240°C with a 5-minute hold. In electron ionization (EI) mode, the ionization energy was fixed at 70 eV, and full-scan mass spectrometry was conducted over the m/z range of 40–450. For short-chain fatty acid analysis, characteristic fragment ions m/z 43 (Acetate), m/z 74 (Propionate), m/z 60 (Butyrate), m/z 87 (Isobuturic), m/z 101 (Valeric and Isovaleric), were selected as quantitative ions to ensure specific detection and accurate quantification of target compounds.
2.9. Gene expression Analyzed via Quantitative Reverse Transcription PCR (qRT-PCR)
To characterize mRNA expression levels of colonic genes including aquaporin 4 (AQP4), aquaporin 8 (AQP8), stem cell factor (SCF), and C-Kit, qRT-PCR analysis was conducted using validated methodologies [20]. Total RNA was extracted from snap-frozen intestinal specimens using TRIzol™ Reagent (Invitrogen, Carlsbad, CA, USA) with mechanical homogenization (Bertin Precellys® 24). RNA integrity was verified through microfluidic electrophoresis (Agilent 2100 Bioanalyzer, RIN > 8.0). First-strand cDNA synthesis was performed using the PrimeScript™ RT Master Mix (Takara, Kusatsu, Japan) with genomic DNA elimination, following the manufacturer's protocol to synthesize cDNA.
The qPCR amplification was conducted utilizing TB Green® Premix Ex Taq™ II (Takara) on a CFX Connect™ Real-Time System (Bio-Rad) with the following thermal profile: initial denaturation at 95°C for 30 seconds, followed by 40 cycles of 95°C for 5 seconds and 60°C for 10 seconds.
Relative gene expression was calculated using the 2−ΔΔCT method with β-actin as the endogenous control. Each sample was analyzed in triplicate, and average Ct values were obtained. ΔCt was determined by subtracting the β-actin Ct value from the target gene Ct value (ΔCt = Ct(target) - Ct(β-actin)). ΔΔCt was calculated by subtracting the ΔCt value of the blank control group from that of the treatment group, following the formula:
ΔΔCt=ΔCt(treatment) - ΔCt(control)
Final relative expression levels were expressed as 2-ΔΔCt values. Gene-specific primers used in this study are listed in Table S1.
2.10. Gut Microbiota Analysis
Mouse fecal samples (200 mg) were aseptically collected and immediately stored at -80°C to preserve microbial DNA integrity. Samples were homogenized using sterile mortar and pestle to ensure uniform microbial cell distribution and improve DNA extraction efficiency. Genomic DNA was extracted using the UltraClean Fecal DNA Isolation Kit (MoBio Laboratories, Carlsbad, CA, USA), involving mechanical lysis and column-based purification.
The V3-V4 hypervariable regions were amplified and sequenced on the Illumina NovaSeq 6000 platform (2×250 bp PE, >100,000 reads/sample). The raw paired-end reads were processed through a comprehensive bioinformatics pipeline. Initially, primer sequences were removed using Cutadapt v2.10. Subsequently, the reads underwent quality filtering with a quality score threshold of Q ≥ 30 and were truncated at 250 base pairs. The processed reads were then subjected to denoising and chimera removal using DADA2 v1.24, followed by the generation of amplicon sequence variants (ASVs).
Taxonomic annotation was performed against the Greengenes database (Silva version 138.1) using QIIME2 v2020.8 (Illumina, Inc.,USA). Phylogenetic relationships were inferred via MAFFT v7.490 multiple sequence alignment. Relative abundances at taxonomic levels (phylum to species) were normalized using cumulative sum scaling.
Alpha diversity indices (Shannon, Chao1, Simpson, observed OTUs) were calculated to assess within-sample diversity. Beta diversity was evaluated using Bray-Curtis dissimilarity and visualized via non-metric multidimensional scaling (NMDS). Linear discriminant analysis effect size (LEfSe) was applied to identify differentially abundant taxa between groups (LDA score > 3.0). Spearman’s rank correlation coefficients were calculated to assess associations between dominant phyla and physiological parameters, with significance adjusted via Benjamini-Hochberg correction.
2.11. Statistical Analysis
Data are presented as mean ± standard error of the mean (SEM). Normality and homogeneity of variances were confirmed prior to analysis. One-way analysis of variance (ANOVA) followed by Duncan's multiple range test was applied to evaluate significant differences among the ten experimental groups. All statistical analyses and graph generation were performed using GraphPad Prism 10.1 (GraphPad Software, San Diego, CA, USA), with statistical significance defined as p < 0.05 (two-tailed tests).
3. Results and Discussion
3.1. LRa05-Mediated Alterations in Excretory Patterns and Intestinal Functionality
Constipation pathophysiology manifests as colonic dysmotility, diminished defecation frequency, and prolonged intestinal transit latency. When compared to the NC group, the loperamide-induced constipation model (MC group) demonstrated marked stool dehydration, exhibiting 38.2% reduction in fecal water content versus native controls (NC, 61.5 ± 1.7%; MC, 48.2 ± 2.1% [g H₂O/g stool], p < 0.001; Figure 1a). The duration until the initial occurrence of black feces was notably prolonged (NC, 91.8 ± 7.1; MC, 157.8 ± 5.1 min, p < 0.001; Fig 1a). This indicated that the mice clearly experienced defecation difficulties after the establishment of the constipation model.
Upon intragastric delivery of Lactobacillus rhamnosus LRa05, there was a significant elevation in the moisture content of mouse feces relative to the MC group (p < 0.01) reaching 52.17% and 54.83% in the low- and high-dose groups (LRa05.L and LRa05.H), respectively. Additionally, the time to initial black feces passage showed a substantial reduction (p < 0.001 vs. MC group), with decreases of 22.9% (LRa05.L, 121.7 ± 8.0 min) and 30.1% (LRa05.H, 110.3 ± 5.6 min) for the two dosage regimens.
These results implied that Lactobacillus rhamnosus LRa05 has the potential to optimize defecation-related metrics in mice suffering from constipation. Specifically, it could stimulat the defecation process and boosts the moisture content of feces. In prior investigations, it has been convincingly shown that Bifidobacterium strains are capable of substantially accelerating intestinal peristalsis, preserving the water content within feces, and reducing the time taken for defecation. Collectively, these observations underscore the crucial role that probiotics play in alleviating constipation symptoms [21].
Disrupted intestinal motility may extend fecal retention in the colon, resulting in excessive water reabsorption from intestinal contents and subsequent formation of dry, hardened stools. As depicted in Figure 1a and Figure S1, the carbon powder transport experiment demonstrated that mice in the MC group exhibited significantly reduced intestinal propulsion capacity, with an intestinal propulsion rate of 39.3 ± 3.4%, which was markedly lower than that of NC group (64.2 ± 3.0%, p < 0.001). Significantly, the intervention with LRa05 enhanced intestinal propulsion in a dose-dependent manner (LRa05.L, 44.8 ± 2.8%; LRa05.H, 48.7 ± 2.3%). The effect was especially remarkable at elevated dosages, with a marked disparity relative to the MC group (p < 0.001). These findings corroborated existing literature demonstrating that Lactobacillus rhamnosus enhanced gastrointestinal transit efficiency and ameliorates constipation by modulating intestinal motility, thereby aligning with the observed functional effects [22].
Table S2 delineated longitudinal alterations in body mass and food intake profiles across experimental cohorts. Univariate analysis of variance (ANOVA) revealed the trend of weight loss in intervention groups relative to baseline controls (NC group) during modeling phase (Δmass = 0.62-0.86 vs. 1.02 g). The administration of Lactobacillus rhamnosus LRa05 (LRa05.H, 2.4×109 CFU/d) restored adipostatic equilibrium, achieving mass accretion indices comparable to the NC group (Δmass = 0.58 vs. 0.81 g, p > 0.05), whereas model group (MC group) exhibited negligible somatic recovery (Δmass = 0.08 g).
A quantitative investigation disclosed that, during the course of treatment, the food intake of the MC group's constipated mice was diminished by 25% in comparison with the NC group. Subsequent probiotic intervention with Lactobacillus rhamnosus LRa05 restored circadian feeding rhythms, achieving 99% recovery of food intake. The findings indicated that LRa05 could alleviate the weight loss and food intake reduction caused by constipation in mice to a certain extent. This might be related to the dual-action mechanism of LRa05. For example, it enhanced ghrelin-mediated orexigenic signaling while suppressing leptin-induced appetite inhibition, thereby playing a role in improving the symptoms associated with constipation-induced decreases in food intake and body weight in mice [23].
3.2. Impact of LRa05 on the serum concentrations of gastrointestinal regulatory peptides
To investigate whether Lactobacillus rhamnosus LRa05 is capable of modulating the levels of gastrointestinal hormone peptides, serum levels of MTL, GAS, SP, SS, ET-1, VIP, and 5-HT were quantified. In this context, MTL, GAS and SP, as excitatory neurotransmitters, enhanced gastrointestinal motility, while ET-1, SS and VIP, as inhibitory ones, decreased it. In the intestine, a dynamic equilibrium exists between inhibitory peptides and excitatory peptides, which respectively regulate intestinal secretion and absorption functions. 5-HT bidirectionally regulates gastrointestinal motility. Deficiency causes constipation, while excess leads to diarrhea.
The results, as depicted in Table 1, revealed a significant reduction in the concentrations of excitatory neurotransmitters, namely SP, MTL, and GAS, when comparing the MC group with the native NC group (p < 0.001). Specifically, the concentration of MTL, GAS and SP measured 395.0 ± 8.6 pg/mL, 66.4 ± 2.2 pg/mL, and 304.1 ± 8.0 pg/mL respectively. In stark contrast, in the MC group, these levels decreased to 264.8 ± 13.5 pg/mL, 36.8 ± 2.4 pg/mL, and 176.4 ± 8.5 pg/mL, respectively. The observed neurotransmitter depletion likely reflects a vicious cycle where constipation-induced ICC/ENS damage, inflammation, and SCF/C-Kit suppression collectively impair excitatory signaling, exacerbating colonic inertia. Upon the intervention of LRa05, when compared with the concentration in the MC group, the high-dose LRa05 could significantly restore the concentration of GAS and SP (p < 0.001). Specifically, the level of GAS increased by 31.1% (from the baseline value in the MC group to 48.2 ± 1.5 pg/mL), and the level of SP increased by 26.7% (from the baseline value in the MC group to 223.5 ± 5.6 pg/mL). This suggested that Lactobacillus rhamnosus LRa05 beneficially regulates excitatory neurotransmitter levels in constipation. By restoring their reduced levels, it could enhance gastrointestinal motility. LRa05 likely promoted SP and GAS synthesis or release, perhaps by interacting with gut-related receptors or signaling pathways involved in neurotransmitter production and secretion. It could also modulate neurotransmitter-producing cell activity, raising levels and improving gut function [24].
Meanwhile, compared with the NC group, the levels of inhibitory neurotransmitters VIP, SS, and ET-1 (Table 1) in the serum of MC group were significantly elevated (p < 0.001), reaching 174.1 ± 5.7 pg/mL, 135.8 ± 5.5 pg/mL, and 216.7 ± 10.0 pg/mL respectively. This dominance of inhibitory peptides likely promotes excessive intestinal water absorption, contributing to dehydration-associated constipation. The elevated inhibitory neurotransmitters in constipated mice likely represent a maladaptive interplay between neuroimmune activation, compensatory dehydration mechanisms, and ENS-ICC cross-talk disruption. Targeting VIP/SS/ET-1 signaling (e.g., receptor antagonists) or restoring neuro-glial-ICC balance could break this cycle and alleviate dehydration-related constipation. After the intervention of LRa05, the levels of VIP, SS, and ET-1 in the high-dose group were significantly reduced by 18.06%, 36.44%, and 38.30% (from the baseline value in the MC group to 142.6 ± 7.1 pg/mL, 86.3 ± 2.5 pg/mL, 133.7 ± 7.9 pg/mL, p < 0.01), respectively. These findings suggested that LRa05 rebalanced gut neurotransmission by suppressing inhibitory pathways, likely via inhibiting enzymes/transcription factors involved in their synthesis or modulating secretory cell activity. This intervention restored the gut's excitatory-inhibitory neurotransmission balance, potentially enhancing gastrointestinal motility [25]. Through dual modulation of peptide networks, LRa05 could concurrently promote intestinal fluid secretion, alleviate luminal dehydration, and softens fecal consistency. This multi-targeted intervention addresses the upstream etiology of constipation-disruption of mucosal water-salt homeostasis-by restoring epithelial transport function and neuromodulatory signaling.
5-HT levels, which bidirectionally regulate gut motility, were severely reduced in constipated mice (NC, 5.68 ± 0.20 pg/mL; MC, 2.24 ± 0.06 pg/mL; p < 0.001; Table 1). The collapse of 5-HT levels in constipated mice reflects EC cell failure, microbiota-ENS-ICC axis disruption, and inflammatory-ischemic damage. Restoring 5-HT signaling (e.g., probiotics, 5-HT4 agonists, or tryptophan supplementation) could break the cycle of motility arrest and dehydration. Similar to moxapride applied in the PC group, partially restored 5-HT (PC, 5.07 ± 0.16 pg/mL; LRa05.L, 2.90 ± 0.08 pg/mL; LRa05.H, 3.66 ± 0.11 pg/mL; p < 0.01 or p < 0.001; Table 1). Though less effective than the drug moxapride, LRa05 likely enhanced tryptophan hydroxylase (TPH1) activity in enterochromaffin cells or modulates gut microbiota metabolites to boost 5-HT signaling [26]. Optimal 5-HT concentrations balance bidirectional signaling in the gut-brain axis (GBA) while coordinating hypothalamic-pituitary-adrenal (HPA) axis cross-talk, thereby optimizing intestinal motility and mucosal permeability. Specifically, 5-HT binds to 5-HT4 receptors on enteric neurons, stimulating acetylcholine release and enhancing smooth muscle contraction, thereby accelerating intestinal transit. Conversely, chronic 5-HT elevation (e.g., in diarrhea) desensitizes 5-HT receptors (e.g., 5-HT4), paradoxically reducing motility. Furthermore, excess 5-HT recruits mucosal immune cells (e.g., mast cells) to release inflammatory mediators (e.g., histamine), indirectly inhibiting smooth muscle activity.
Collectively, Lactobacillus rhamnosus LRa05 exerts its constipation-relieving effects through multi-faceted modulation of gut hormone peptides, with a primary focus on GBA pathway. These effects are hypothesized to be mediated via direct interactions with intestinal epithelial cells, microbial metabolites, or host signaling cascades, underscoring its potential as a functional probiotic. This observation aligns with prior studies demonstrating that divergent Lactobacillus rhamnosus strains could selectively elevate MTL and 5-HT levels via distinct molecular routes, thereby improving gastrointestinal transit in constipated murine models [27]. Gut microbiota are known to directly or indirectly synthesize and regulate host neuroactive compounds such as 5-HT and SP, thereby exerting far-reaching effects on both enteric nervous system (ENS) and central nervous system (CNS) physiology. Mechanistically, LRa05 augmented 5-HT biosynthesis through tryptophan metabolic reprogramming-manifested by TPH1 upregulation and IDO1 inhibition-and promoted SP secretion, thereby triggering vagal afferent signaling to brainstem nuclei such as the nucleus tractus solitarius. Furthermore, through suppression of pro-inflammatory mediators ET-1 and VIP, LRa05 restored gut-brain signaling integrity. Specifically, ET-1 reduction mitigated intestinal ischemia and inflammatory responses, whereas VIP downregulation diminishes its inhibitory effects on ENS excitability, rebalancing smooth muscle contractility. This multi-transmitter balancing capability positions LRa05 as a promising therapeutic candidate, potentially outperforming single-target interventions such as 5-HT agonists by concurrently addressing gastrointestinal motility disorders, visceral hypersensitivity, and central nervous system comorbidities.
The observation that the positive control group (e.g., treated with mosapride citrate) demonstrated more pronounced alterations in gastrointestinal peptide levels (e.g., MTL, VIP shown in Table1) relative to the probiotic intervention group (LRa05.), yet failed to achieve superior constipation relief (displayed by Figure1a), highlighted the multifactorial complexity of constipation regulation. This paradox underscores that gastrointestinal peptides alone do not fully govern constipation pathophysiology, and additional mechanisms-particularly those modulated by probiotics-are critical for symptom resolution.
3.3. Mediation of Colonic Histopathological Integrity by LRa05
To evaluate the restorative potential of Lactobacillus rhamnosus LRa05 on colonic integrity, systematic histopathological analyses in loperamide-induced constipated mice were performed. Hematoxylin-eosin (HE) stained colon sections were examined at 200× magnification to characterize tissue architecture (Figure 1b) and quantify indices (Figure 1c).
The NC group displayed intact colonic morphology featuring orderly intestinal villi alignment and abundant goblet cell populations within crypts. In contrast, the MC group exhibited hallmark colonic pathological alterations, including disrupted epithelial polarity, mucosal atrophy with muscularis dissociation (green arrow), focal inflammatory infiltrates (blue arrow), and significant muscularis propria thinning (red arrow) in Figure 1b. Quantitative histomorphometric analysis demonstrated severe constipation-induced colonic structural degeneration (Figure 1c). The smooth muscle layer thickness plummeted by 67.4% in the model group (MC, 45.2 ± 0.9 μm) compared to native controls (NC, 138.6 ± 6.7 μm; p < 0.001), indicating compromised contractile capacity. The crypt depth and villus height reduced 65.1% and 45.6% (NC, 93.6 ± 0.8 and 181.3 ± 2.9 μm vs. MC, 32.7 ± 1.3 and 98.6 ± 2.1 μm). These metrics collectively reflect profound mucosal-muscular dissociation and impaired barrier functionality, hallmarks of chronic constipation pathophysiology. These observations aligned with established constipation models demonstrating goblet cell disorganization, muscular hypoplasia, and leukocyte migration [28]. These pathological changes in the MC group likely arose from constipation-related physiological impairments. Slow fecal transit exerts abnormal mechanical stress on the colonic mucosa, which disrupts epithelial architecture and induces muscularis propria dissociation. Prolonged contractile hypoactivity in constipation progressively thins the muscular layer. Concurrently, compromised epithelial barrier function triggers focal inflammatory infiltrates via immune cell migration. The disorganization of goblet cells-critical for mucus production that maintains intestinal lubrication-further exacerbates mucosal dysfunction. Impaired mucus secretion culminates in dry, hardened stools, perpetuating a vicious cycle of colonic tissue damage [29].
Notably, LRa05 administration dose-dependently reversed pathological remodeling. The low-dose group (LRa05.L) restored colonic architecture to levels comparable with the positive control (PC) group, demonstrating intermediate recovery of mucosal folds (PC, 76.3 ± 12.0 μm and 72.2 ± 6.8 μm) and goblet cell density. The high-dose group (LRa05.H) achieved the reestablished goblet cell stratification and attenuated inflammatory infiltration (Figure1b), which was critical for intestinal lubrication and barrier defense. Colon wall thickness surged 1.8-fold versus the model group (MC, 45.2 ± 0.9 μm vs. LRa05.H, 81.2 ± 0.9 μm; Figure 1c), reinstating contractile vigor. The crypt depth and villus height achieved 2.7-fold recovery, and 64.1% restitution (MC, 32.7 ± 1.3 and 98.6 ± 2.1 μm vs. LRa05.H, 87.8 ± 2.0 and 161.7 ± 1.1 μm; Figure 1c), enhancing absorptive surface area. These findings demonstrated LRa05’s capacity to repair mucosa-muscularis dissociation and reinstate colonic architectural homeostasis, aligning with its prokinetic efficacy. This functional profile parallelled reports that Lactobacillus rhamnosus LRJ-1 alleviated distal colonic injury via goblet cell regeneration and muscularis reinforcement (p < 0.05).
The observed effects of Lactobacillus rhamnosus LRa05 were likely mediated by dual mechanisms, augmentation of gastrointestinal motility and biosynthesis of bioactive metabolites, which acted synergistically to mitigate constipation-induced colonic injury. Reduced fecal retention alleviates mechanical stress on the mucosa and limits exposure to cytotoxic metabolites (e.g., ammonia, secondary bile acids) and pathogenic bacteria-derived endotoxins. This mitigates epithelial barrier disruption and inflammatory cascades, preserving crypt-villus architecture and goblet cell functionality. Additionally, as a commensal probiotic, LRa05 could ferment dietary fibers to produce SCFAs (e.g., acetate, propionate, butyrate), which exert pleiotropic restorative effects [30,31]. The histomorphometric data demonstrated that Lactobacillus rhamnosus LRa05 mitigated constipation-associated colonic degeneration through mucosal barrier regeneration and anti-inflammatory reprogramminga. These structural improvements likely synergize with its prokinetic effects (Figure 1a) and serotonergic modulation (Table 1), collectively restoring gut homeostasis.
3.4. LRa05's Role in Affecting the Content of SCFAs
Short-chain fatty acids (SCFAs), the primary metabolites of gut microbial fermentation, alleviate constipation through multiple mechanisms: modulating gut microbiota composition, stimulating luminal hydration, and preserving intestinal barrier integrity [32].
As shown in Table 1, constipated mice treated with loperamide in MC group exhibited significantly reduced cecal SCFA levels relative to the NC group (p < 0.05). The levels of acetic acid (9.46 µmol/g), propionic acid (5.43 µmol/g), butyric acid (4.17 µmol/g), isobutyric acid (3.78 µmol/g), valeric acid (1.32 µmol/g), and isovaleric acid (2.10 µmol/g) were notably lower the NC group. These reductions could be attributed to constipation-induced fecal stasis disrupting the gut microbial ecosystem, leading to suppression of SCFA-producing taxa such as Bacteroides, Clostridium, and Roseburia. This systemic SCFA depletion be likely to contributed to impaired intestinal motility and stool desiccation in constipation.
Notably, Lactobacillus rhamnosus LRa05 intervention dose-dependently restored SCFA production, with the high-dose group (LRa05.H) showing superior efficacy (p < 0.01). Specifically, the high-dose LRa05 could increase the acetic acid and butyric acid contents to 38.2 µmol/g and 12.3 µmol/g, respectively, which were higher than thosein the NC group (27.1 µmol/g and 8.55 µmol/g). Acetic acid, a dominant SCFA, binds to G protein-coupled receptor 43 (GPR43) on enteroendocrine cells and enteric neurons, triggering release of motility-stimulating peptides (e.g., PYY, GLP-1), which accelerate colonic transit [33]. Butyric acid, functions as the main energy source for colonocytes, undergoing β-oxidation to generate ATP essential for maintaining epithelial integrity and tight junction function (e.g., upregulating occludin and zonula occludens-1). Additionally, butyrate acts as a histone deacetylase (HDAC) inhibitor, suppressing pro-inflammatory cytokine production (TNF-α, IL-6) while promoting regulatory T-cell (Treg) differentiation. This dual action mitigates "leaky gut"-induced inflammation and preserves ENS and ICC function critical for motility [34]. Propionate enhances vagal afferent signaling to the brainstem, thereby stimulating defecation reflexes. Valerate modulates serotonin receptor (5-HT4) sensitivity, potentiating smooth muscle contraction. As a probiotic, LRa05 competitively inhibits pathogenic bacteria (e.g., Escherichia, Enterobacter) while promoting symbiotic SCFA-producing microbiota. This microbial rebalancing reactivates dietary fiber fermentation-the primary substrate for SCFA synthesis. Mechanistically, LRa05 upregulates key microbial enzymes involved in SCFA biosynthesis, including butyryl-CoA:acetate CoA-transferase (for butyrate) and phosphate acetyltransferase (for acetate), thereby restoring SCFA metabolic pathways disrupted in constipation.
These results indicate that Lactobacillus rhamnosus LRa05 exerts constipation-relieving effects through dual mechanisms: restoring microbiota-gut crosstalk via SCFA-mediated motility enhancement, and preserving intestinal barrier homeostasis through butyrate-driven anti-inflammatory pathways. The underlying mechanisms involve intricate interactions among LRa05, microbiota modulation, and SCFA-mediated host signaling networks, which collectively target the core pathophysiology of constipation, including motility dysfunction, barrier injury, and neuroendocrine dysregulation. Compared to the MC group, the PC group (treated with mosapride citrate) demonstrated partial restoration of SCFA production, though its efficacy was significantly weaker than that of the probiotic intervention group (LRa05.L and LRa05.H). For instance, acetic acid levels in the mosapride group reached 21.6 µmol/g, compared to 38.1 µmol/g in LRa05.H and 9.46 µmol/g in MC (Table 1). This disparity suggested that mosapride citrate and LRa05 operate through divergent mechanistic pathways to alleviate constipation. The differential efficacy between mosapride citrate and LRa05 underscores the importance of microbiota-centric therapies in addressing SCFA-mediated constipation. While mosapride transiently improves motility, probiotics achieve sustained recovery by rebuilding the gut ecosystem and enhancing host-microbe crosstalk. Combining prokinetics with probiotics may synergize motility enhancement and SCFA restoration.
3.5. LRa05's Role in Affecting the Expression of Linked Genes
To delve deeper into the mechanism underlying the effects of Lactobacillus rhamnosus LRa05, the critical genes governing gastrointestinal motility and fluid homeostasis were profiled systematically (Figure 2). ICCs, the pacemakers of gastrointestinal rhythmicity, coordinate smooth muscle contractions through electrical slow waves [35, 36]. The SCF/C-Kit axis is indispensable for ICC maintenance, with SCF binding to its tyrosine kinase receptor C-Kit to drive ICC proliferation and network formation [37]. Dysregulation of this pathway disrupts slow-wave generation, precipitating motility disorders [38].
The qRT-PCR data of mRNA levels revealed constipation-induced colonic suppression of both SCF and C-Kit (4.41- and 3.8-fold downregulation, respectively) in the MC group versus the NC group (p < 0.01 or p < 0.001; Figure 2a and 2b). This was because SCF/C-Kit signaling was essential for the growth, survival, and functional maintenance of ICC-the pacemaker cells responsible for generating intestinal slow-wave potentials. Reduced SCF expression and C-Kit receptor downregulation disrupt ICC network integrity, leading to diminished slow-wave activity and impaired gastrointestinal motility. This molecular perturbation aligns with the pathophysiological hallmark of chronic constipation-the loss of ICC density and function. ICC depletion directly compromises the intrinsic myoelectric activity of the intestinal smooth muscle, resulting in aberrant peristaltic contractions and reduced propulsive motility, which are cardinal features of hypomotility disorders. The SCF and C-Kit axis dysfunction thus represents a critical molecular node linking colonic mucosal signaling to motility deficits in constipation, underscoring its role as a potential target for restoring ICC-mediated gastrointestinal rhythm.
Following treatment with various doses of LRa05 or administration of mosapride citrate tablets, all experimental groups demonstrated differential recovery responses. LRa05 intervention dose-dependently restored SCF and C-Kit expression levels (p < 0.01 for LRa05.L and p < 0.001 for LRa05.H vs. MC), showing 73.1% and 47.2% recovery with low-dose treatment, and 92.6% and 69.6% recovery with high-dose treatment. Mechanistically, SCF activation bound to C-Kit receptors on ICC progenitor cells, promoting their proliferation and differentiation. This molecular restoration correlated with the histological observations of muscularis propria thickening (Figure 1c) and enhanced intestinal propulsion (Figure 1a), suggesting LRa05 reactivates ICC-dependent pacemaking to reinstate peristaltic rhythms. This recovery likely stemmed from LRa05-mediated GBA regulation. Probiotic SCFAs boosted enterochromaffin cell activity, increasing 5-HT biosynthesis (Table 1). 5-HT activates 5-HT4 receptors on epithelial cells or ICCs, triggering cAMP/PKA signaling that enhances SCF secretion [39]. SCF, in turn, binds to C-Kit on ICCs, promoting their proliferation and restoring slow-wave generation (Figure 2b). In addition, Lactobacillus strains, including LRa05, could directly engage Toll-like receptor 2 (TLR2) on intestinal epithelial cells, activating MyD88-dependent signaling to upregulate SCF transcription [40]. SCFAs amplify this process by stabilizing epithelial barrier function, ensuring sustained SCF release [41]. The PC group (mosapride citrate-treated) exhibited only partial restoration of SCF and C-Kit expression (e.g., 53.8% and 32.4% recovery vs. MC, respectively), significantly lagging behind the probiotic intervention groups. This disparity implies that mosapride’s monotherapy primarily targets acute motility enhancement rather than addressing the multifactorial pathophysiology of constipation.This differential therapeutic effect can be mechanistically attributed to Mosapride's role as a selective 5-HT4 receptor agonist, which transiently enhances serotonin-dependent smooth muscle contractility. However, it lacks direct regulatory effects on ICC progenitor cell proliferation or SCF transcriptional activity-key processes required for sustainable ICC network regeneration. Moreover, Mosapride fails to restore intestinal microbiota diversity or SCFA concentrations (notably butyrate), which are critical for epithelial SCF secretion and ICC metabolic homeostasis.
Complementary to motor function, the levels of aquaporins (AQP4 and AQP8) governing intestinal water transport were assessed. Compared with the NC group, chronic constipation led to a significant upregulation of AQP4 and AQP8 in the MC group, as shown in Figure 2c,d. Specifically, the expression level of AQP4 was 1.68-fold higher (p < 0.05), and that of AQP8 was 1.83-fold higher (p < 0.01) than those in the NC group. This aligned with prior evidence that AQP overexpression exacerbates pathological fluid retention and stool desiccation [42]. LRa05 administration reversed this dysregulation, with high-dose treatment reducing AQP4 and AQP8 expression to 47.3% and 38.1% of MC levels (p < 0.01), effectively rebalancing luminal hydration. Overexpression of AQPs in constipation is often driven by inflammatory cytokines (e.g., TNF-α, IL-6) that upregulate AQP transcription via NF-κB/MAPK pathways. LRa05-derived SCFAs could suppress these pathways, reducing AQP4/AQP8 expression [43,44]. SCFAs could interact with GPR43 receptors on enterocytes, activating intracellular calcium signaling to modulate AQP trafficking or degradation, thereby reducing membrane AQP density [45]. From a gastrointestinal peptide synergy perspective, Lactobacillus rhamnosus LRa05 lowers inhibitory peptides (VIP, SS; Table. 1), which were known regulators of AQP activity. Reduced VIP/SS levels could decrease cAMP signaling in enterocytes, thereby indirectly normalizing AQP-mediated water reabsorption. Concurrently, elevated 5-hydroxytryptamine (5-HT; Table 1) and restored MTL enhance fluid secretion by stimulating chloride channels (e.g., cystic fibrosis transmembrane conductance regulator, CFTR), counteracting AQP-driven water retention. The coordinated actions of AQP modulation and neuropeptide regulation synergistically maintain optimal fecal viscoelasticity (rheological plasticity) and prevent stool dehydration, thereby preserving intestinal transit efficiency.
This dual modulation of inhibitory-excitatory peptide networks likely rebalanced intestinal water flux, alleviating constipation-associated dehydration. At the motility level, SCF/C-Kit activation rebuilds ICC networks, rescuing slow-wave activity for propulsive contractions [46]. Concurrently, at the osmoregulatory level, AQP suppression prevents excessive water extraction, maintaining stool plasticity [47]. This mechanistic synergy could explain the accelerated fecal transit, improved stool hydration, and normalized defecation frequency observed in LRa05-treated groups(Figure 1a).
3.6. Functional Role of LRa05 in Modulating the Gut Microbiota
To evaluate the microbiota-modulatory effects of Lactobacillus rhamnosus LRa05 in constipated mice, 16S rRNA gene sequencing targeting the V3-V4 hypervariable regions was conducted on colonic contents collected at the study endpoint. Alpha diversity metrics (Chao1 and observed otus for richness, Shannon and Simpson for diversity) were analyzed via species richness and community diversity indices (Figure 3a). Beta diversity was assessed using PCoA and NMDS based on Bray-Curtis dissimilarity (Figure 3b).
In the MC group, significant (p < 0.05) reductions were observed in both microbial richness (Chao1 and observed OTUs) and diversity (Shannon and Simpson indices) compared to the NC group. The LRa05 intervention reversed these perturbations in a dose-dependent manner, with the LRa05.H group demonstrating significantly enhanced Chao1 richness and observed OTUs (p < 0.001), as well as Shannon and Simpson indices (p < 0.01), surpassing both the low-dose LRa05.L and PC groups (Figure 3a).
PCoA and NMDS revealed distinct clustering patterns. Compared to the NC group, the MC group exhibited a marked separation along the second principal component (PC2, 19.1% variance), confirming constipation-induced gut microbiota dysbiosis (Figure 3b). Notably, both LRa05.H and LRa05.L groups overlapped with the NC group, indicating partial restoration of microbiota composition. Conversely, the PC group showed a clear separation along the first principal component (PC1, 44.0% variance) relative to other groups, highlighting the distinct role of moxapride (used in the PC group) in modulating gut microbiota composition.
Consistent with established gut microbiota profiles, Bacteroidetes and Firmicutes dominated (>90% combined abundance) at the phylum level (Figure 4a) [48]. Loperamide-induced constipation disrupted gut microbiota composition, with Bacteroidetes abundance decreasing by 21.2% and Firmicutes increasing by 26.1% relative to the NC control (p < 0.01). This microbial dysbiosis resulted in a significantly reduced Firmicutes/Bacteroidetes (F/B) ratio in the MC group (1.01±0.12) compared to NC group (1.62 ± 0.18; p < 0.05). LRa05 intervention restored the F/B ratio in a dose-dependent manner (LRa05.L: 1.38 ± 0.15; LRa05.H: 1.52 ± 0.14), with the high-dose group achieving NC-equivalent levels (p > 0.05). This finding aligns with previous studies demonstrating that lactic acid bacteria stabilize F/B ratios to mitigate inflammation and enhance epithelial barrier function [49].
Constipated microbiota is characterized by decreased beneficial bacteria and increased potential pathogens [50]. At the genus level (Figure 4b, 4c), loperamide induced significant compositional shifts. LRa05 intervention notably restored beneficial taxa including Parabacteroides, Lachnospiraceae_NK4A136_group, Oscillibacter, Alloprevotella, and Rikenellaceae_RC9_gut_group, aligning with prior studies. For instance, functional fruit drinks relieved constipation by upregulating Alloprevotella and enhancing barrier function [51], while Lactobacillus salivus Li01 restored Lachnospiraceae_NK4A136_group and Rikenellaceae_RC9_gut_group abundances to reduce inflammation [52]. Emerging evidence indicates Parabacteroides and Oscillibacter metabolize dietary tryptophan into indole-3-propionic acid (IPA), a high-affinity aryl hydrocarbon receptor (AhR) ligand that suppresses neuroinflammation and stabilizes enteric nervous system (ENS) function through gut-brain axis modulation. Mechanistically, Alloprevotella negatively correlates with pro-inflammatory cytokines and produces SCFAs like butyrate to mitigate intestinal inflammation [53]. Lachnospiraceae_NK4A136_group, an anaerobic spore-forming probiotic, ferments plant polysaccharides into SCFAs to protect intestinal mucosa and inversely correlates with inflammation markers [54]. Rikenellaceae_RC9_gut_group mediates anti-inflammatory effects through immune modulation, strengthening intestinal barrier integrity. In summary, Lactobacillus rhamnosus LRa05 exerted functional effects by restoring beneficial bacterial genera, promoting SCFA production, modulating inflammatory pathways, and enhancing intestinal barrier function. These concerted actions contribute to microbiota rehabilitation and defecation function recovery in constipation.
The heatmap and LEfSe analysis (LDA > 3.0, Figure 5a and 5b) identified distinct microbial signatures across groups. Compared with the other group, the MC group exhibited significant enrichment of Desulfovibrio, Candidatus_Saccharimonas, Enterorhabdus, and Rikenella (LEfSe analysis, LDA score > 3.0, p < 0.05). These microbial taxa (Desulfovibrio, Candidatus_Saccharimonas, Enterorhabdus, and Rikenella) have been extensively documented to be associated with intestinal mucosal barrier dysfunction (e.g., reduced tight junction protein expression and increased permeability) and exacerbation of pro-inflammatory cytokine production (particularly IL-6, TNF-α, and IL-1β) in both preclinical models and human cohort studies [55]. Notably, Enterorhabdus has been specifically implicated in triggering NF-κB-mediated inflammatory cascades through its metabolite interactions with intestinal epithelial cells, as demonstrated by in vitro transwell co-culture systems [56]. It is worth noting that these inflammatory cytokines or signaling pathways may exacerbate constipation symptoms. For instance, IL-6 slows intestinal peristalsis by inhibiting cholinergic neuron activity in the ENS. TNF-α induces segmental hyperkinesia and transmission delay via RhoA/ROCK-mediated enhancement of smooth muscle contraction persistence. While IL-1β exacerbates fecal dryness through downregulating intestinal epithelial aquaporin and reducing luminal water secretion. This finding was further validated in correlation analysis (Figure 5c), where Enterorhabdus and Rikenella demonstrated significant positive correlations with AQP4 and AQP8, respectively.
The LRa05.H group restored ecological equilibrium via enrichment of bacteria with SCFA synthesis, or immune regulatory functions (such as Bacteroides, Parabacteroides, Paraprevotella, Colidextribacter, Rikenellaceae_RC9_gut_group) [57]. For instance, Bacteroides and Parabacteroides species predominantly mediate SCFA production and mucin degradation, creating a prebiotic niche for secondary fermenters. These taxa provide metabolic substrates (e.g., acetate, propionate) and mucin-derived oligosaccharides that sustain microbial communities critical for intestinal homeostasis [58]. The LRa05.L group highlighted enrichment of beneficial bacteria such as Eubacterium_xylanophilum_group (linked to carbohydrate metabolism and gut barrier integrity), Oscillibacter (implicated in neurotransmitter production and mental health), Alistipes (involved in immune regulation and inflammation reduction), Odoribacter (correlated with improved lipid metabolism), and Lachnospiraceae_NK4A136_group (promoting SCFA production critical for colonic health) [59]. Notably, preferential proliferation of Oscillibacter and Alistipes—which are specialized in synthesizing neurotransmitter precursors (e.g., GABA, glutamate) and promoting immune tolerance through regulatory T cell activation—exerts synergistic neuroprotective and immunomodulatory effects [60].
The LRa05.L (low-dose) and LRa05.H (high-dose) groups exhibited distinct induction patterns of beneficial taxa, demonstrating dose-dependent probiotic modulation of gut microbial ecology. Additionally, the PC group exhibited a unique gut microbiota modulation profile distinct from both LRa05.L and LRa05.H probiotic interventions. Specifically, Akkermansia, Alloprevotella, Prevotellaceae_UCG-001, and Muribaculum were significantly enriched in the PC group compared to probiotic-treated cohorts, with Alloprevotella demonstrating the most pronounced enrichment. The PC-induced microbiota profile reflected the ecological mechanisms favoring mucin- and plant glycan-adapted taxa rather than functional resilience, distinct from the probiotic cross-feeding networks observed in LRa05 groups [61].
The ecological divergence was further supported by the correlation analysis (Figure 5c). The MC group exhibited significant microbial-metabolic correlations distinct from the probiotic-treated LRa05.H group. The Desulfovibrio and Candidatus_Saccharimonas with higher abundances in the MC group, were significantly negatively correlated with fecal water content, propulsion rate, levels of gut motility activators (GAS, MTL, SP, 5-HT), and expression of SCF/C-kit, while showing significant positive correlations with time to first black feces and levels of motility inhibitors (SS, VIP, ET-1). Notably, Rikenella-another constipation-associated genus enriched in the MC group demonstrated a notable negative correlation with SCFA production (butyrate: r = -0.65; acetate: r = -0.61; p < 0.05). This aligned with its known role in degrading mucin glycans and releasing pro-inflammatory mediators (e.g., IL-6, TNF-α), which suppress colonic fermentation and epithelial energy metabolism. The dominance of Desulfovibrio in the MC group likely impairs gut motility through hydrogen sulfide-mediated smooth muscle relaxation, while its mucolytic activity exacerbates intestinal barrier dysfunction, triggering upregulation of SS/VIP [62]. Concurrently, Rikenella thrives in dysbiotic, inflamed environments by preferentially metabolizing host glycans over dietary fibers. This metabolic shift diverts microbial activity toward lactate and ammonia production, further acidifying the intestinal lumen and inhibiting SCFA biosynthesis. Collectively, these distinct microbial-metabolic interactions in the MC group highlight how constipation-associated dysbiosis disrupts host-microbe co-metabolism, perpetuating motility arrest and inflammatory cascades [63].
In contrast, probiotic-enriched genera Paraprevotella and Parabacteroides in the LRa05.H group exhibited inverse correlations with these motility and metabolic parameters, suggesting beneficial modulation of gut function. Specifically, Parabacteroides likely enhances motility via succinate-TGR5 receptor signaling, which potentiates 5-HT biosynthesis in enterochromaffin cells [64]. Additionally, Paraprevotella-derived anti-inflammatory metabolite 3-indolepropionic acid may suppress ET-1 production through nuclear factor kappa-light-chain-enhancer of NF-κB inhibition [65]. Furthermore, genera Eubacterium_xylanophilum_group, Colidextribacter, and Rikenellaceae_RC9_gut_group demonstrated significant positive correlations with SCFA production, underscoring their roles in metabolic regulation. Notably, taxa enriched in the positive control PC group (Akkermansia, Alloprevotella, Prevotellaceae) exhibited negligible significant correlations with these functional parameters. The above findings suggested that dietary probiotic supplementation alleviates constipation symptoms through a multi-tiered microbiota-host signaling axis, for instance, SCFA biosynthesis, gastrointestinal regulatory reptide modulation, SCF/C-Kit pathway reactivation, AQP homeostasis restoration.
4. Conclusions
This mechanistic framework offered an academic groundwork for the precise intervention in constipation cases (such as distinguishing between dehydration-induced constipation and hypomotility-induced constipation), and at the same time highlighted the unique advantages of multi-target intervention by probiotics. Unlike previous single-target studies (e.g., microbiota or SCFAs), this study established Lactobacillus rhamnosus LRa05 as a mechanistically defined, multitarget probiotic for constipation management, with functional efficacy spanning motility enhancement, mucosal repair, and microbiome-metabolite crosstalk regulation etc. The LRa05 intervention demonstrated significant efficacy in enhancing gastrointestinal transit, as indicated by the decreased time to initial black feces passage and increased fecal moisture content. This treatment strategy also restored enteroendocrine homeostasis by modulating key gastrointestinal hormones. It promoted the upregulation of motilin while concurrently reducing the levels of vasoactive bowel peptide. At the molecular level, LRa05 rectified dysregulated signaling pathways, upregulating markers of Cajal interstitial cells axis while suppressing the overexpression of pathological aquaporins, specifically AQP4 and AQP8. Additionally, the intervention reshaped the gut microbiota composition by increasing the production of SCFAs and enriching commensal microbial taxa associated with improved laxation. These findings highlight the probiotic's dual role in mediating microbiota-gut barrier crosstalk through SCFA-dependent epithelial modulation, and regulating neuroendocrine axes, including the GBA and HPA axis, via integrated ICC-hormonal-microbial signaling. Mechanistically, LRa05 demonstrates translational potential for constipation management by simultaneously addressing motility dysfunction, mucosal injury, and luminal dehydration. Notably, this probiotic strain selectively enriched anti-inflammatory and SCFA-producing genera (Parabacteroides, Lachnospiraceae), with microbial profiles exhibiting marked divergence from the antibiotic-treated control group. Such ecological specificity highlights the functional advantages of probiotic interventions over broad-spectrum antimicrobials. Therefore, Lactobacillus rhamnosus LRa05 has translational potential for gut health as a probiotic intervention or supplement. Regarding translation, while rodent models validated its pre-clinical efficacy, critical steps remain. For example, human trials for dose-response and safety in diverse groups (such as elderly, IBS-C subtypes), formulation optimization for colonic delivery and microbiome engraftment, and omics-based mechanistic deconvolution are needed to find host-microbe interaction nodes.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Jingxin Zhang and Qian Li contributed to the work equally and should be regarded as co-first authors, corresponding for conceptualization, data curation, formal analysis, investigation, writing the initial draft, etc. Shanshan Liu, Ning Wang, Yu Song, corresponded for the investigation, data curation and formal analysis. Tao Wu and Min zhang, corresponded for the funding acquisition, project administration and supervision.
Funding
This research was funded by the Key Research and Development Program of China (2022YFF1100201), 2024 Tianjin "the Belt and Road" Innovation Platform Project (24PTLYHZ00020), Natural Science Foundation of Tianjin Municipality (24JCQNJC00740).
Data Availability Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Acknowledgments
We thank Wecare Probiotics Co., Ltd. for their instrumental contribution of materials utilized in this experimental investigation.
Conflicts of Interest
The authors have declared no conflict of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| ICC | Interstitial cells of Cajal |
| AQPs | Aquaporins |
| MTL | Motilin |
| GAS | Gastrin |
| SP | Substance P |
| SS | Somatostatin |
| ET-1 | Endothelin-1 |
| VIP | Vasoactive intestinal peptide |
| 5-HT | 5-hydroxytryptamine |
| SCF | Stem cell factor |
References
- Bharucha, A.E.; Lacy, B.E. Mechanisms, evaluation, and management of chronic constipation. Gastroenterology 2020, 158, 1232–1249.e3. [Google Scholar] [CrossRef] [PubMed]
- Yuan, M.; Zhang, Z.; Liu, T.; Feng, H.; Liu, Y.; Chen, K. The role of nondigestible oligosaccharides in alleviating human chronic diseases by regulating the gut microbiota: A review. Foods 2024, 13, 2157. [Google Scholar] [CrossRef] [PubMed]
- Gershon, M.D.; Margolis, K.G. The gut, its microbiome, and the brain: Connections and communications. J Clin Invest 2021, 131, e143768. [Google Scholar] [CrossRef]
- Zhao, X.; Qiu, Y.; Liang, L.; Fu, X. Interkingdom signaling between gastrointestinal hormones and the gut microbiome. Gut Microbes 2025, 17, 2456592. [Google Scholar] [CrossRef] [PubMed]
- Bubeck, M.; Becker, C.; Patankar, J.V. Guardians of the gut: Influence of the enteric nervous system on the intestinal epithelial barrier. Front Med 2023, 10, 1228938. [Google Scholar] [CrossRef]
- Foong, D.; Zhou, J.; Zarrouk, A.; Ho, V.; O’Connor, M.D. Understanding the Biology of Human Interstitial Cells of Cajal in Gastrointestinal Motility. Int J Mol Sci 2020, 21, 4540. [Google Scholar] [CrossRef]
- Gao, X.; Guo, K.; Liu, S.; Yang, W.; Sheng, J.; Tian, Y.; Peng, L.; Zhao, Y. A Potential use of vidarabine: Alleviation of functional constipation through modulation of the adenosine A2A receptor-MLC signaling pathway and the gut microbiota. Int J Mol Sci 2024, 25, 12810. [Google Scholar] [CrossRef]
- Zhu, S.; Yu, Q.; Xue, Y.; Li, J., Huang; Y., Liu, W.; Wang, G.; Wang, L.; Zhai, Q.; Zhao, J.; Zhang, H.; Chen, W. Bifidobacterium bifidum CCFM1163 alleviates cathartic colon by activating the BDNF-TrkB-PLC/IP3 pathway to reconstruct the intestinal nerve and barrier. Food Funct 2025, 16, 2057–2072. [Google Scholar]
- Brigstocke, S.; Yu, V.; Nee, J. Review of the Safety Profiles of Laxatives in Pregnant Women. J Clin Gastroenterol 2022, 56, 197–203. [Google Scholar] [CrossRef]
- Huang, Y.P.; Shi, J.Y.; Luo, X.T.; Luo, S.C.; Cheung, P.C.K.; Corke, H.; Yang, Q.Q.; Zhang, B.B. How do probiotics alleviate constipation? A narrative review of mechanisms. Crit Rev in Biotechnol 2025, 45, 80–96. [Google Scholar] [CrossRef]
- Zhu, M.; Yang, L.; Kong, S.; Bai, Y.; Zhao, B. Lacticaseibacillus Rhamnosus LRa05 alleviates cyclophosphamide-induced immunosuppression and intestinal microbiota disorder in mice. J Food Sci 2024, 89, 10003–10017. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Shao, Y.; Zhang, Y.; Zhao, Y.; Han, M.; Gai, Z. In Vitro and in Vivo genome-based safety evaluation of Lacticaseibacillus Rhamnosus LRa05. Food Chem Toxicol 2024, 186, 114600. [Google Scholar] [CrossRef]
- Vallejos, O.P.; Bueno, S.M.; Kalergis, A.M. Probiotics in inflammatory bowel disease: microbial modulation and therapeutic prospects. Trends in Mol Med 2025. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.; Chen, Y.; Wang, J.; Gao, Y.; Gai, Z.; Zhao, Y.; Xu, F. Lacticaseibacillus Rhamnosus LRa05 alleviated liver injury in mice with alcoholic fatty liver disease by improving intestinal permeability and balancing gut microbiota. Benef Microbes 2024, 15, 481–493. [Google Scholar] [CrossRef]
- Wu, T.; Zhang, Y.; Li, W.; Zhao, Y.; Long, H.; Muhindo, E.M.; Liu, R.; Sui, W.; Li, Q.; Zhang, M. Lactobacillus rhamnosus LRa05 ameliorate hyperglycemia through a regulating glucagon-mediated signaling pathway and gut microbiota in type 2 diabetic Mice. J Agric Food Chem 2021, 69, 8797–8806. [Google Scholar] [CrossRef] [PubMed]
- Bahari, H.; Akhgarjand, C.; Mirmohammadali, S.N.; Malekahmadi, M. Probiotics and eating disorders: A systematic review of humans and animal model studies. J Eat Disord 2024, 12, 193. [Google Scholar] [CrossRef]
- Ma, W.; Zhao, Y.; Liu, Y.; Wang, Y.; Yu, S.; Huang, L. Bifidobacterium Animalis Subsp. Lactis TG11 ameliorates loperamide-induced constipation in mice by modulating gut microbiota. Front Microbiol 2025, 16, 1525887. [Google Scholar]
- Dong, L.; Xu, Z.; Huang, G.; Zhang, R.; Deng, M.; Huang, F.; Su, D. Lychee pulp-derived dietary fiber-bound phenolic complex upregulates the SCFAs-GPRs-ENS pathway and aquaporins in loperamide-induced constipated mice by reshaping gut microbiome. J Agric Food Chem 2023, 1, 15087–15096. [Google Scholar] [CrossRef]
- Nobels, A.; van Marcke, C.; Jordan, B.F.; Hul, M.V.; Cani, P.D. The gut microbiome and cancer: from tumorigenesis to therapy. Nat Metab 2025. [Google Scholar] [CrossRef]
- Shi, Y.; Chen, F.; Wang, Z.; Cao, J.; Li, C. Effect and mechanism of functional compound fruit drink on gut microbiota in constipation mice. Food Chem 2023, 401, 134210. [Google Scholar] [CrossRef]
- Ma, W.; Zhao, Y.; Liu, Y.; Wang, Y.; Yu, S.; Huang, L. Bifidobacterium animalis subsp. lactis TG11 ameliorates loperamide-induced constipation in mice by modulating gut microbiota. Front Microbiol 2025, 16, 1525887. [Google Scholar]
- Wang, G.; Yang, S.; Sun, S.; Si, Q.; Wang, L.; Zhang, Q.; Wu, G.; Zhao, J.; Zhang, H.; Chen, W. Lactobacillus Rhamnosus strains relieve Loperamide-induced constipation via different pathways independent of short-chain fatty acids. Front Cell Infect Microbiol 2020, 10, 423. [Google Scholar] [CrossRef] [PubMed]
- Vivot, K.; Meszaros, G.; Pangou, E.; Zhang, Z.R.; Qu, M.D.; Erbs, E.; et al. CaMK1D signalling in AgRP neurons promotes ghrelin-mediated food intake. Nat Metab 2023, 5, 1045–1058. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Zhao, W.; Luo, J.; Shi, S.; Niu, X.; He, J.; et al. Synergistic Defecation effects of Bifidobacterium Animalis Subsp. Lactis BL-99 and fructooligosaccharide by modulating gut microbiota. Front Immunol 2025, 15, 1520296. [Google Scholar]
- Haq, S.; Wang, H.Q; Grondin, J.; Banskota, S.; Marshall, J.K.; Khan, I.I.; et al. , Disruption of autophagy by increased 5-HT alters gut microbiota and enhances susceptibility to experimental colitis and Crohn’s disease. Sci Adv 2021, 7, eabi6442. [Google Scholar] [CrossRef]
- Xia, T.; Huang, F.; Yun, F.; Liu, Y.; Wang, T.; Wang, S.; Jin, S.; Ma, X.; Wang, W.; He, J.; et al. Lacticaseibacillus Rhamnosus LRJ-1 Alleviates Constipation through Promoting Gut Bacteroides-Derived γ-Aminobutyric Acid Production. Curr Res Food Sci 2024, 9, 100924. [Google Scholar] [CrossRef] [PubMed]
- Hickey, J.W.; Becker, W.R.; Nevins, S.A.; Horning, A.; Perez, A.E.; Zhu, C.C.; et al. Organization of the human intestine at single-cell resolution. Nature 2023, 619, 572–584. [Google Scholar] [CrossRef]
- Piechka, A.; Sparanese, S.; Witherspoon, L.; Hach, F.; Flannigan, R. Molecular mechanisms of cellular dysfunction in testes from men with non-obstructive azoospermia. Nat Rev Urol 2024, 21, 67–90. [Google Scholar] [CrossRef]
- Hu, M.; Fang, C.; Liu, Y.; Gao, M.; Zhang, D.; Shi, G.; Yin, Z.; Zheng, R.; Zhang, J. Comparative study of the laxative effects of konjac oligosaccharides and konjac glucomannan on loperamide-induced constipation in rats. Food Funct 2021, 2021. 12, 7709–7717. [Google Scholar] [CrossRef]
- Zhao, Q.; Xing, F.; Tao, Y.; Liu, H.; Huang, K.; Peng, Y.; Feng, N.; Liu, C. Xiaozhang Tie improves intestinal motility in rats with cirrhotic ascites by regulating the stem cell factor/c-kit pathway in interstitial cells of Cajal. Front Pharmacol 2020, 11, 1. [Google Scholar] [CrossRef]
- Tang, Q.; Huang, H.; Xu, H.; Xia, H.; Zhang, C.; Di, Y.; Bi, F. Endogenous coriobacteriaceae enriched by a high-fat diet promotes colorectal tumorigenesis through the CPT1A-ERK axis. NPJ Biofilms Microbi 2024, 10, 1–15. [Google Scholar] [CrossRef]
- Sun, M.; Wu, T.; Zhang, G.; Liu, R.; Sui, W.; Zhang, M.; Geng, J.; Yin, J.; Zhang, M. Lactobacillus rhamnosus LRa05 improves lipid accumulation in mice fed with a high fat diet via regulating the intestinal microbiota, reducing glucose content and promoting liver carbohydrate metabolism. Food Funct 2020, 11, 9514–9525. [Google Scholar] [CrossRef] [PubMed]
- Bárcenas-Preciado, V.; Mata-Haro, V. Probiotics in miRNA-mediated regulation of intestinal immune homeostasis in pigs: A physiological narrative. Microorganisms 2024, 12, 1606. [Google Scholar] [CrossRef] [PubMed]
- Lv, H.; Niu, J.; Pan, W.; Wang, Y.; Wang, L.; Wang, M.; Shi, Y.; Zhang, G.; Al Hamyari, B.; Wang, S.; et al. Stool-softening effect and action mechanism of free anthraquinones extracted from Rheum Palmatum L. on water deficit-induced constipation in rats. J Ethnopharmacol 2024, 319, 117336. [Google Scholar] [CrossRef] [PubMed]
- Hays, K.E.; Pfaffinger, J.M.; Ryznar, R. The interplay between gut microbiota, short-chain fatty acids, and implications for host health and disease. Gut Microbes 2024, 16, 2393270. [Google Scholar] [CrossRef]
- Pimpão, C.; da Silva, I.V.; Soveral, G. The expanding role of aquaporin-1, aquaporin-3 and aquaporin-5 as transceptors: involvement in cancer development and potential druggability. Int J Mol Sci 2025, 26, 1330. [Google Scholar] [CrossRef]
- Sanders, K.M.; Santana, L.F.; Baker, S.A. Interstitial cells of Cajal - pacemakers of the gastrointestinal tract. J Physiol 2023, 2023. 10, 1113. [Google Scholar] [CrossRef]
- Huang, Z.P.; Qiu, H.; Wang, K.; Chao, W.B.; Zhu, H.B.; Chen, H.; Liu, Y.; Yu, B.P. The impact of acute stress disorder on gallbladder interstitial cells of Cajal. J Cell Physiol 2020, 235, 8424–8431. [Google Scholar] [CrossRef]
- Yuhan, L.; Khaleghi Ghadiri, M.; Gorji, A. Impact of NQO1 dysregulation in CNS disorders. J Transl Med 2024, 22, 4. [Google Scholar] [CrossRef]
- Huang, S.; Xu, P.; Shen, D. D.; Simon, I. A.; Mao, C.; Tan, Y.; et al. GPCRs steer Gi and Gs selectivity via TM5-TM6 switches as revealed by structures of serotonin receptors. Molecular cell 2022, 82, 2681–2695.e6. [Google Scholar]
- Yoshimoto, T.; Kittaka, M.; Doan, A.A.P.; Urata, R.; Prideaux, M.; Rojas, R.E. Osteocytes directly regulate osteolysis via MYD88 signaling in bacterial bone infection. Nat Commun 2022, 13, 6648. [Google Scholar] [CrossRef]
- Xia, T.; Huang, F.; Yun, F.; Liu, Y.; Wang, T.; Wang, S.; Jin, S.; Ma, X.; Wang, W.; He, J.; Teng, K.; Zhong, J. Lacticaseibacillus rhamnosus LRJ-1 alleviates constipation through promoting gut Bacteroides-derived γ-aminobutyric acid production. Curr. Res. Food Sci 2024, 9, 100924. [Google Scholar] [CrossRef]
- Lv, H.; Niu, J.; Pan, W.; Wang, Y.; Wang, L.; Wang, M.; Shi, Y.; Zhang, G.; Al, H.B.; Wang, S.; Li, X.; Shi, Y. Stool-softening effect and action mechanism of free anthraquinones extracted from Rheum palmatum L on water deficit-induced constipation in rats. J. Ethnopharmacol 2024, 319, 117336. [Google Scholar]
- Liu, Q.; Yu, Z.; Tian, F.; Zhao, J.; Zhang, H.; Zhai, Q.; Chen, W. Surface components and metabolites of probiotics for regulation of intestinal epithelial barrier. Microb. Cell Fact 2020, 19, 23. [Google Scholar] [CrossRef]
- Shi, R.; Yu, F.; Hu, X.; Liu, Y.; Jin, Y.; Ren, H.; Lu, S.; Guo, J.; Chang, J.; Li, Y.; et al. Protective effect of Lactiplantibacillus Plantarum Subsp. Plantarum SC-5 on dextran sulfate sodium—induced colitis in mice. Foods 2023, 12, 897. [Google Scholar] [PubMed]
- Miao, C.; Wang, L.; Wang, H.; Shen, Y.; Man, C.; Zhang, W.; Zhang, Y.; Zhao, Q.; Jiang, Y. Lacticaseibacillus plantarum postbiotics prepared by the combined technique of pasteurization and ultrasound: effective measures to alleviate obesity based on the SCFAs-GPR41/GPR43 signaling pathway. Food Funct 2024, 15, 11005–11019. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Chen, F.; Wang, Z.; Cao, J.; Li, C. Effect and mechanism of functional compound fruit drink on gut microbiota in constipation mice. Food Chem. 2023, 401, 134210. [Google Scholar] [CrossRef]
- Olesen, E.T.B.; Fenton, R.A. Aquaporin 2 regulation: implications for water balance and polycystic kidney diseases. Nat Rev Nephrol 2021, 17, 765–781. [Google Scholar] [CrossRef]
- Sequeira, R.P.; McDonald, J.A.K.; Marchesi, J.R. Clarke; Thomas, B. Commensal Bacteroidetes protect against Klebsiella pneumoniae colonization and transmission through IL-36 signalling. Nat Microbiol 2020, 5, 304–313. [Google Scholar] [CrossRef]
- Yue, Z.; Zhao, F.; Guo, Y.; Zhang, Y.; Chen, Y.; He, L.; Li, L. Lactobacillus reuteri JCM 1112 ameliorates chronic acrylamide-induced glucose metabolism disorder via the bile acid-TGR5-GLP-1 axis and modulates intestinal oxidative stress in mice. Food Funct 2024, 15, 6450–6458. [Google Scholar] [CrossRef]
- Zhang, T.; Lu, H.; Cheng, T.; Wang, L.; Wang, G.; Zhang, H.; Chen, W. Bifidobacterium Longum S3 alleviates loperamide-induced constipation by modulating intestinal acetic acid and stearic acid levels in mice. Food Funct 2024, 15, 6118–6133. [Google Scholar] [CrossRef]
- Han, B.; Shi, L.; Bao, M.Y.; Yu, F.L.; Zhang, Y.; Lu, X.Y.; Wang, Y.; Li, D.X.; Lin, J.C.; Jia, W.; et al. Dietary ellagic acid therapy for CNS autoimmunity: Targeting on Alloprevotella Rava and propionate metabolism. Microbiome 2024, 12, 114. [Google Scholar] [CrossRef]
- Yang, W.; Cong, Y. Gut microbiota-derived metabolites in the regulation of host immune responses and immune-related inflammatory diseases. Cell Mol Immunol 2021, 18, 866–877. [Google Scholar] [CrossRef] [PubMed]
- Katsirma, Z.; Dimidi, E.; Rodriguez-mateos, A.; Whelan, K. Fruits and their impact on the gut microbiota, gut motility and constipation. Food Funct 2021, 12, 8850–8866. [Google Scholar]
- Huang, S.; Chen, J.; Cui, Z.; Ma, K.; Wu, D.; Luo, J.; Li, F.; Xiong, W.; Rao, S.; Xiang, Q.; et al. Lachnospiraceae-derived butyrate mediates protection of high fermentable fiber against placental inflammation in gestational diabetes mellitus. Scince Adv 2023, 9, eadi7337. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Zhang, L.; Wang, X.; Yi, Y.; Shan, Y.; Liu, B.; Zhou, Y.; Lü, X. Roles of Intestinal Parabacteroides in human health and diseases. FEMS Microbiol Lett 2022, 369, fnac072. [Google Scholar] [CrossRef]
- Mukherjee, A.; Lordan, C.; Ross, R.P.; Cotter, P.D. Gut microbes from the phylogenetically diverse genus Eubacterium and their various contributions to gut Health. Gut Microbes 2020, 12, 1802866. [Google Scholar] [CrossRef]
- Paone, P.; Cani, P.D. Mucus barrier, mucins and gut microbiota: The expected slimy partners? Gut 2020, 69, 2232–2243. [Google Scholar] [CrossRef] [PubMed]
- Dong, L.; Xu, Z.; Huang, G.; Zhang, R.; Deng, M.; Huang, F.; Su, D. Lychee pulp-derived dietary fiber-bound phenolic complex upregulates the SCFAs-GPRs-ENS pathway and aquaporins in loperamide-induced constipated mice by reshaping gut microbiome. J. Agric. Food Chem 2023, 71, 15087–15096. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, Y.; Li, W.; Zhang, B.; Yin, J.; Liuqi, S.; Wang, J.; Peng, B.; Wang, S. Fucoidan ameliorated dextran sulfate sodium-induced ulcerative colitis by modulating gut microbiota and bile acid metabolism. J Agric Food Chem 2022, 70, 14864–14876. [Google Scholar] [CrossRef]
- Yang, J.; Li, Y.; Wen, Z.; Liu, W.; Meng, L.; Huang, H. Oscillospira - a candidate for the next-generation probiotics. Gut microbes 2021, 13, 1987783. [Google Scholar] [CrossRef]
- Qu, S.Y.; Yu, Z.J.; Zhou, Y.X.; Wang, S.Y.; Jia, M.Q.; Chen, T.; Zhang, X.J. Gut microbiota modulates neurotransmitter and gut-brain signaling. Microbiol Res 2024, 287, 127858. [Google Scholar] [CrossRef]
- Qi, Q.; Zhang, H.; Jin, Z; Wang, C.C.; Xia, M.Y.; Chen, B.; et al. Hydrogen sulfide produced by the gut microbiota impairs host metabolism via reducing GLP-1 levels in male mice. Nat Metab 2024, 6, 1601–1615. [Google Scholar] [CrossRef]
- Okamoto, Y.; Miyabe, Y.; Seki, M.; Ushio, Y.; Sato, K.; Kasama, E.; Akiyama, K.; Karasawa, K.; Uchida, K.; Kikuchi, K.; et al. First case of a renal cyst infection caused by Desulfovibrio: A case report and literature review. BMC Nephrol 2022, 23, 194. [Google Scholar] [CrossRef] [PubMed]
- Thingholm, L.B.; Rühlemann, M.C.; Koch, M.; Fuqua, B.; Laucke, G.; Boehm, R.; Bang, C.; Franzosa, E.A.; Hübenthal, M.; Rahnavard, A.; et al. Obese individuals with and without type 2 diabetes show different gut microbial functional capacity and composition. Cell Host & Microbe 2019, 26, 252–264.e10. [Google Scholar]
Figure 1.
Effects of Lactobacillus rhamnosus LRa05 on constipation-related parameters and intestinal tissue damage. (a) Fecal water content (%), times to first black feces (min) and intestinal propulsion rate (%); (b) HE-stained images of mice colon tissue; (c) Thickness of the colon wall, crypt depth and villus height (μm). Data are presented as mean ± standard error of the mean (SEM) with a sample size of n=10. For statistical significance, the symbol # denotes p < 0.05, ## denotes p < 0.01, ### denotes p < 0.001 (vs. NC group); * representing p < 0.05, ** representing p < 0.01, and *** representing p < 0.001 (vs. MC group).
Figure 1.
Effects of Lactobacillus rhamnosus LRa05 on constipation-related parameters and intestinal tissue damage. (a) Fecal water content (%), times to first black feces (min) and intestinal propulsion rate (%); (b) HE-stained images of mice colon tissue; (c) Thickness of the colon wall, crypt depth and villus height (μm). Data are presented as mean ± standard error of the mean (SEM) with a sample size of n=10. For statistical significance, the symbol # denotes p < 0.05, ## denotes p < 0.01, ### denotes p < 0.001 (vs. NC group); * representing p < 0.05, ** representing p < 0.01, and *** representing p < 0.001 (vs. MC group).
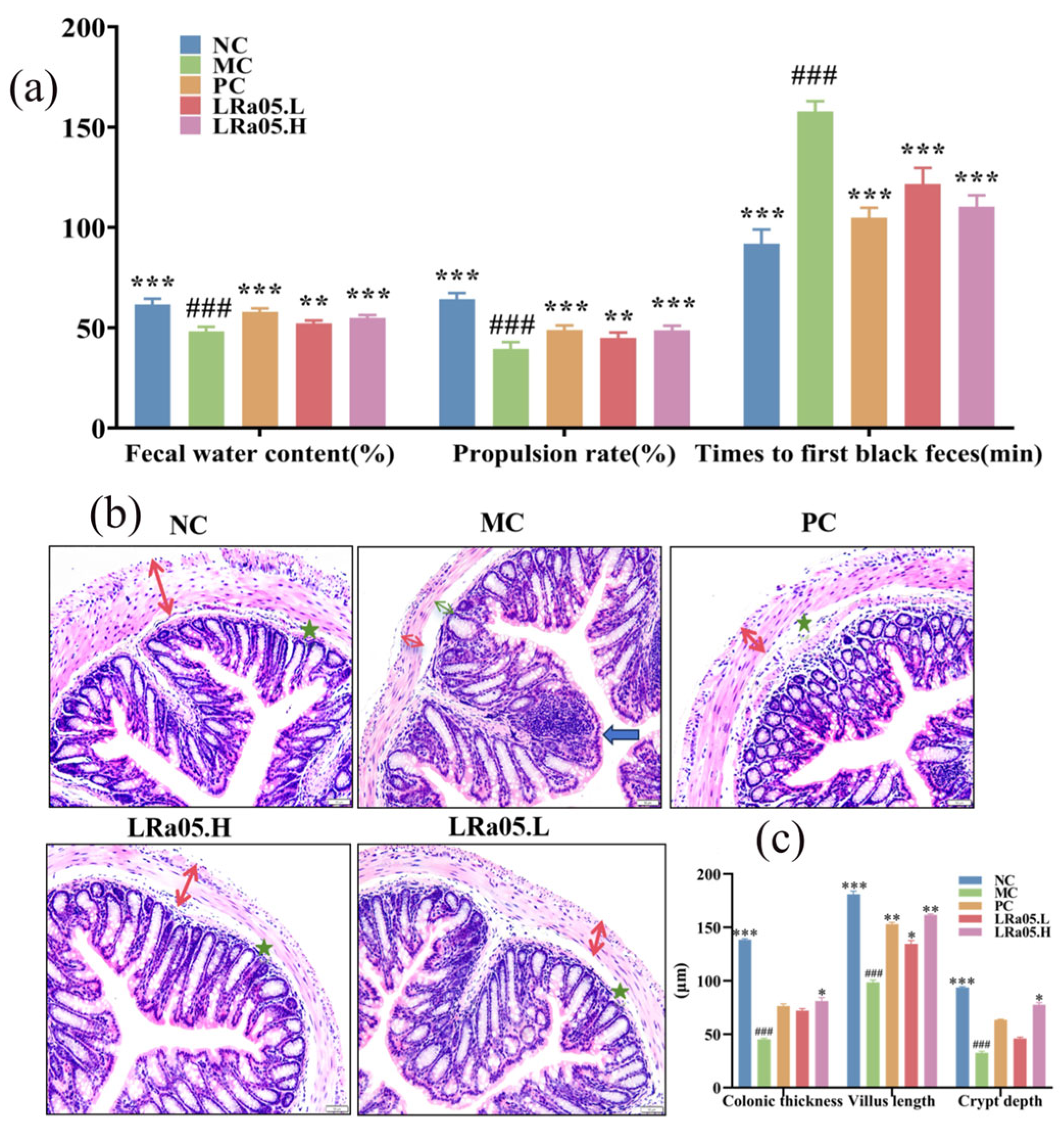
Figure 2.
Transcriptional regulation of SCF (a), C-Kit (b), AQP4 (c), and AQP8 (d) in the colons of mice. Values are expressed as mean ± SEM (n=10). Data are presented as mean ± standard error of the mean (SEM) with a sample size of n=10. For statistical significance, the symbol # denotes p < 0.05, ## denotes p < 0.01, ### denotes p < 0.001 (vs. NC group); * representing p < 0.05, ** representing p < 0.01, and *** representing p < 0.001 (vs. MC group).
Figure 2.
Transcriptional regulation of SCF (a), C-Kit (b), AQP4 (c), and AQP8 (d) in the colons of mice. Values are expressed as mean ± SEM (n=10). Data are presented as mean ± standard error of the mean (SEM) with a sample size of n=10. For statistical significance, the symbol # denotes p < 0.05, ## denotes p < 0.01, ### denotes p < 0.001 (vs. NC group); * representing p < 0.05, ** representing p < 0.01, and *** representing p < 0.001 (vs. MC group).
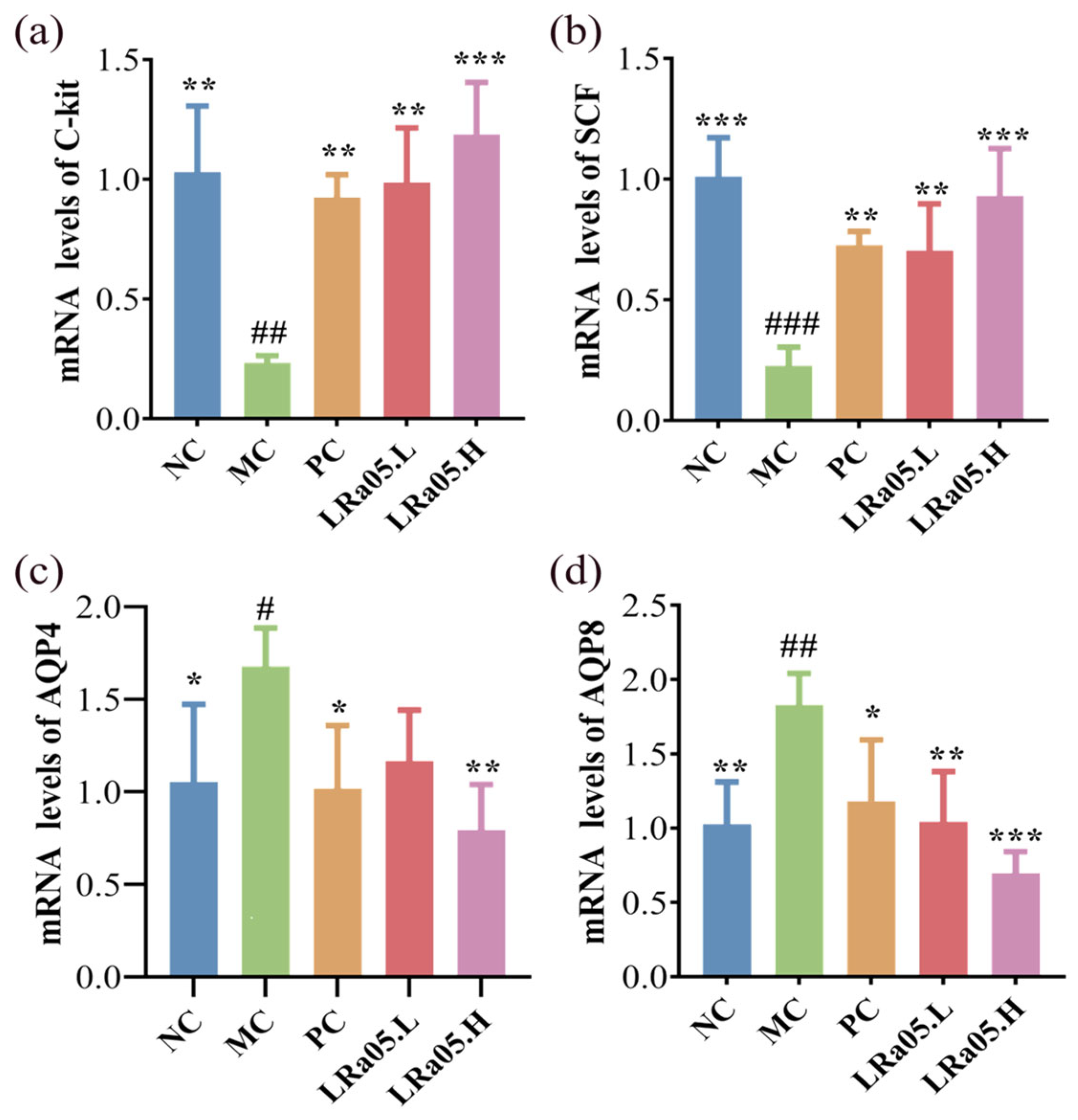
Figure 3.
LRa05-mediated modulation of species diversity and differentiation in the gut microbiota. (a) Alpha diversity; (b) Beta diversity. Values are expressed as mean ± SEM (n=10). Data are presented as mean ± standard error of the mean (SEM) with a sample size of n=10. For statistical significance, the symbol # denotes p < 0.05, ## denotes p < 0.01, ### denotes p < 0.001 (vs. NC group); * representing p < 0.05, ** representing p < 0.01, and *** representing p < 0.001 (vs. MC group).
Figure 3.
LRa05-mediated modulation of species diversity and differentiation in the gut microbiota. (a) Alpha diversity; (b) Beta diversity. Values are expressed as mean ± SEM (n=10). Data are presented as mean ± standard error of the mean (SEM) with a sample size of n=10. For statistical significance, the symbol # denotes p < 0.05, ## denotes p < 0.01, ### denotes p < 0.001 (vs. NC group); * representing p < 0.05, ** representing p < 0.01, and *** representing p < 0.001 (vs. MC group).
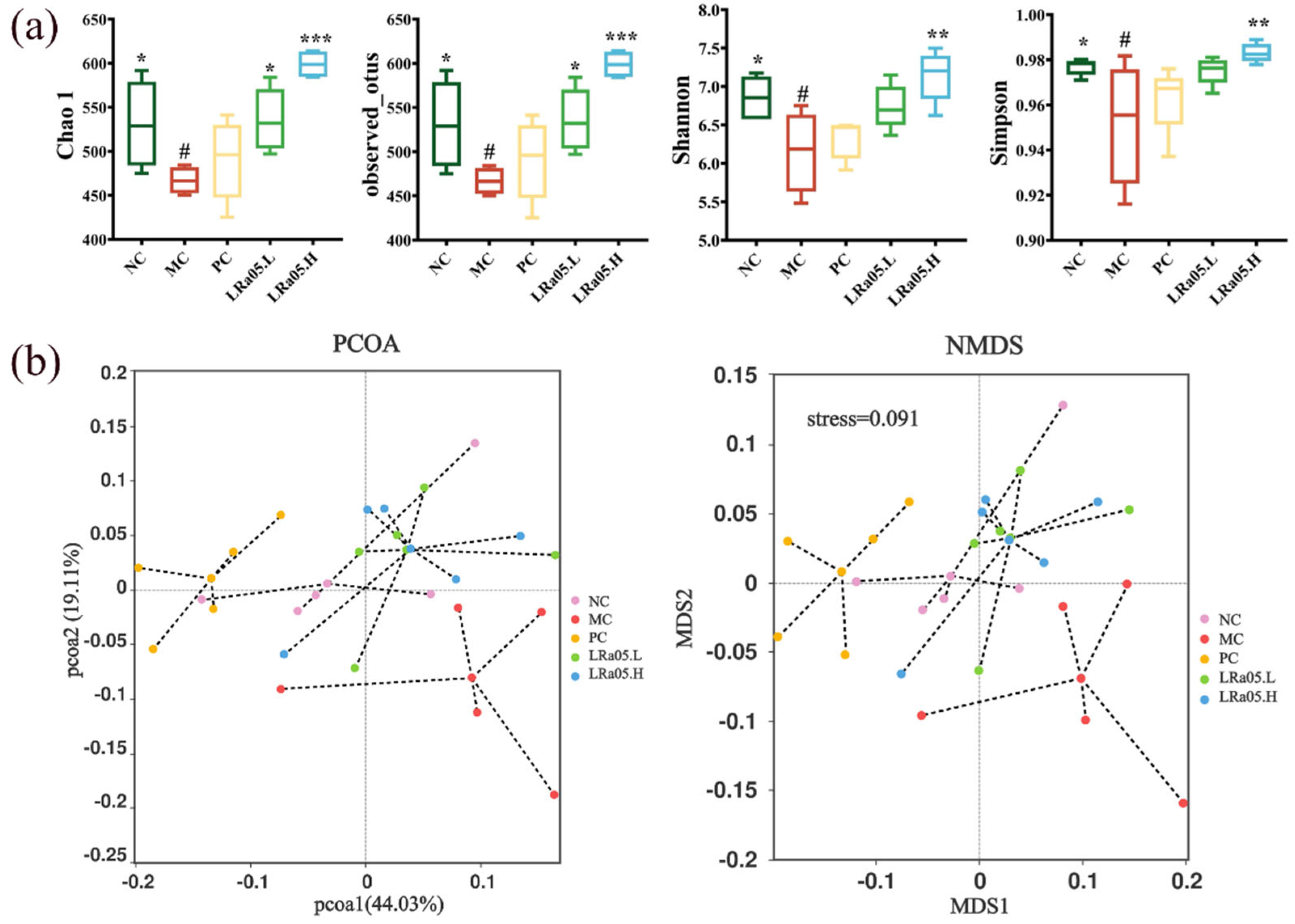
Figure 4.
Modulation of gut microbiota composition by LRa05 intervention. (a) Phylum-level distribution of intestinal microbiota; (b) Genus-level compositional profiles; (c) Relative abundances of selected bacterial genera demonstrating significant alterations. Data are presented as mean ± standard error of the mean (SEM) with a sample size of n=10. For statistical significance, the symbol # denotes p < 0.05, ## denotes p < 0.01, ### denotes p < 0.001 (vs. NC group); * representing p < 0.05, ** representing p < 0.01, and *** representing p < 0.001 (vs. MC group).
Figure 4.
Modulation of gut microbiota composition by LRa05 intervention. (a) Phylum-level distribution of intestinal microbiota; (b) Genus-level compositional profiles; (c) Relative abundances of selected bacterial genera demonstrating significant alterations. Data are presented as mean ± standard error of the mean (SEM) with a sample size of n=10. For statistical significance, the symbol # denotes p < 0.05, ## denotes p < 0.01, ### denotes p < 0.001 (vs. NC group); * representing p < 0.05, ** representing p < 0.01, and *** representing p < 0.001 (vs. MC group).
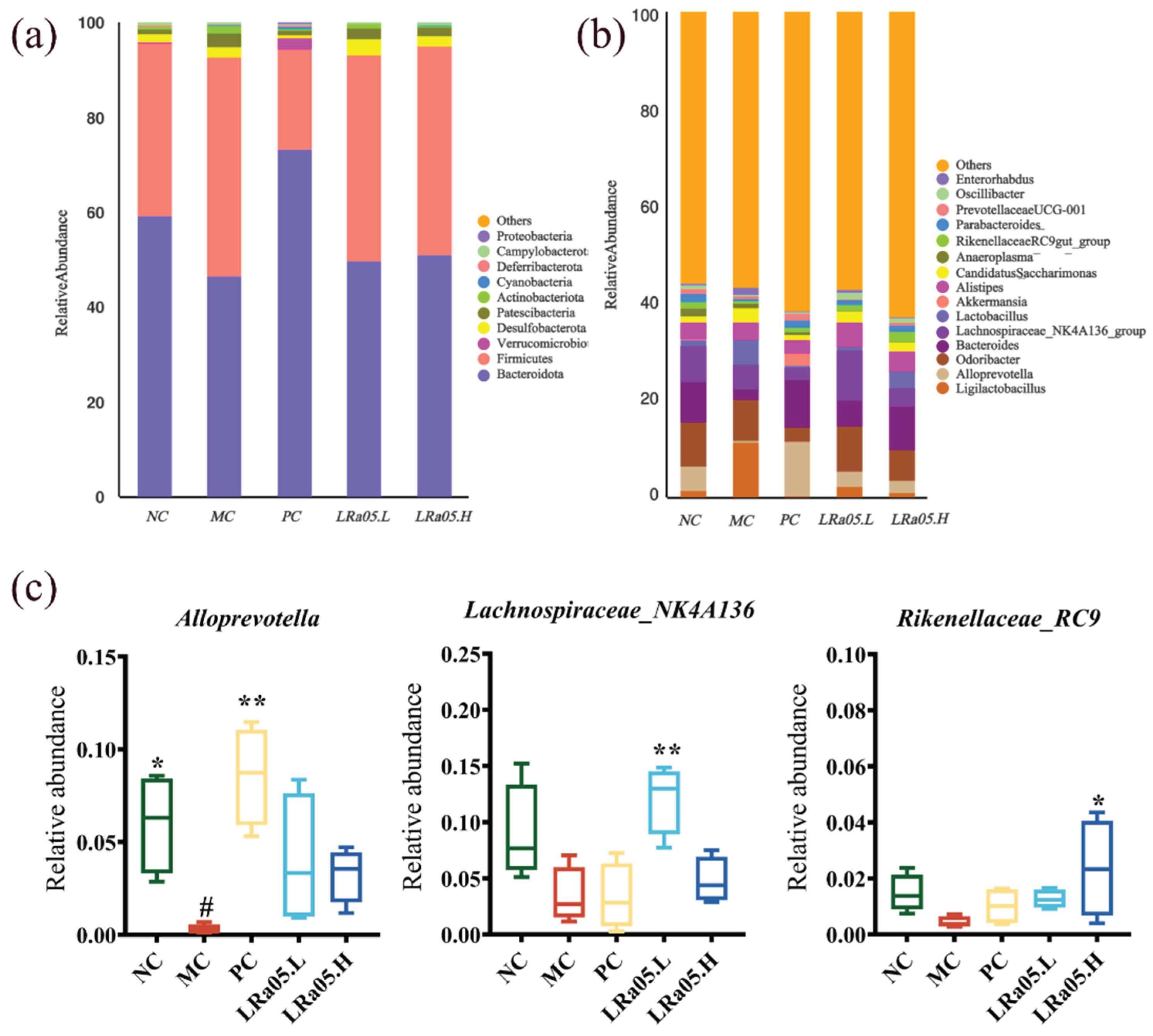
Figure 5.
Microbial biomarkers and functional correlations. (a) Hierarchical clustering was visualized using a heatmap; (b) Linear discriminant analysis (LDA) effect size (LEfSe) (LDA score ≥ 3.0); (c) Spearman correlation heatmap depicting genus-level associations with constipation-related phenotypes. Color intensity reflects correlation strength (blue = negative, red = positive), with significant correlations denoted by * p < 0.05 and ** p < 0.01.
Figure 5.
Microbial biomarkers and functional correlations. (a) Hierarchical clustering was visualized using a heatmap; (b) Linear discriminant analysis (LDA) effect size (LEfSe) (LDA score ≥ 3.0); (c) Spearman correlation heatmap depicting genus-level associations with constipation-related phenotypes. Color intensity reflects correlation strength (blue = negative, red = positive), with significant correlations denoted by * p < 0.05 and ** p < 0.01.
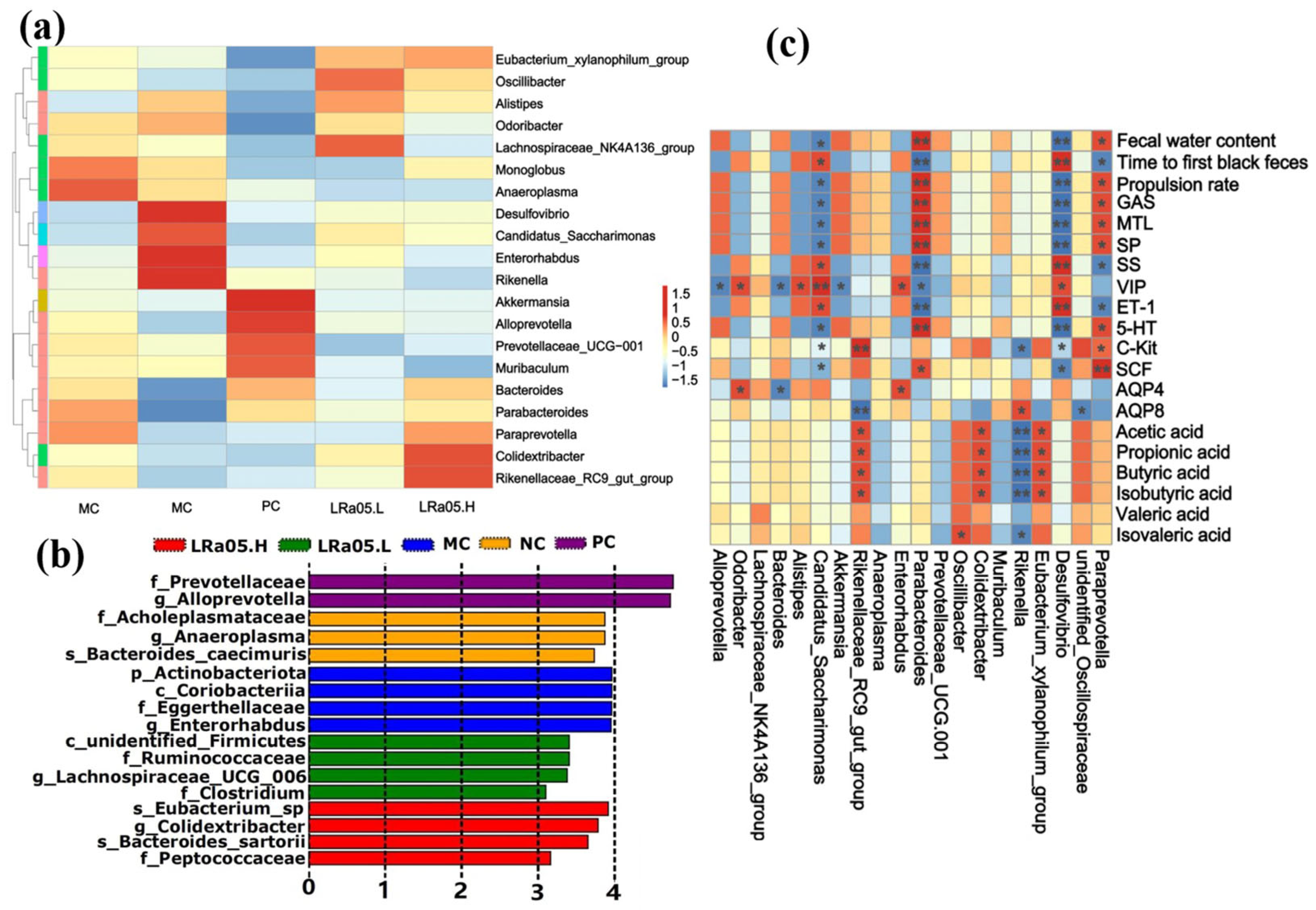

Table 1.
The serum concentrations of gastrointestinal regulatory peptides and the contents of SCFAs in various groups of mice
Table 1.
The serum concentrations of gastrointestinal regulatory peptides and the contents of SCFAs in various groups of mice
| Index | Group | ||||||
|---|---|---|---|---|---|---|---|
| NC | MC | PC | LRa05.L | LRa05.H | |||
| serum concentrations of gastrointestinal regulatory peptides | |||||||
| MTL | 395.05±8.57a | 264.75±13.50b | 376.81±10.92a | 275.43±16.47b | 300.24±14.22b | ||
| GAS | 66.36±2.24a | 36.79±2.35c | 64.43±2.11a | 43.72±2.14bc | 48.24±1.55b | ||
| SP | 304.12±7.99a | 176.36±8.54c | 280.57±12.45a | 183.89±10.75bc | 223.52±5.64b | ||
| SS | 113.06±3.34c | 174.10±5.68a | 103.09±3.95c | 160.71±6.69ab | 142.66±7.07b | ||
| ET-1 | 60.91±1.92d | 135.83±5.47a | 68.52±1.10d | 110.82±3.78b | 86.33±2.51c | ||
| VIP | 120.01±5.93c | 216.68±10.02a | 108.45±8.53c | 182.68±12.12b | 133.70±7.87c | ||
| 5-HT | 5.68±0.20d | 2.24±0.06a | 5.06±0.16d | 2.90±0.08b | 3.66±0.11c | ||
| SCFAs contents | |||||||
| Acetic acid | 27.11±1.99bc | 9.46±0.91d | 21.64±1.33c | 26.68±.51b | 38.15±1.13a | ||
| Propionic acid | 9.25±0.65ab | 5.43±0.23d | 7.77±0.65c | 9.59±0.72ab | 10.75±0.49a | ||
| Butyric acid | 8.55±0.50bc | 4.17±0.27d | 7.03±0.50c | 9.61±1.22b | 12.34±0.58a | ||
| Isobutyric acid | 4.20±0.10a | 3.78±0.04b | 4.01±0.07ab | 4.21±0.08a | 4.23±0.10a | ||
| Valeric acid | 1.83±0.10a | 1.32±0.05b | 1.60±0.09ab | 1.98±0.14a | 1.95±0.17a | ||
| Isovaleric acid | 2.53±0.11a | 2.10±0.05b | 2.32±0.08ab | 2.58±0.12a | 2.52±0.12a | ||
* Data represent the mean ± SEM (n = 10). Different letters within the same row denote significant differences among distinct groups, with a significance level of p < 0.05.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



