
Submitted:
22 May 2025
Posted:
23 May 2025
You are already at the latest version
Abstract
Pancreatic islets play a central role in glucose homeostasis, and their dysfunction underlies the development of diabetes mellitus. While mesenchymal stromal cells (MSCs) have shown promise in supporting islet regeneration, limitations such as immunogenicity and invasiveness prompt the search for alternative sources. Salivary gland cells (SGCs), due to their accessibility and regenerative potential, are emerging as a candidate, though their role in pancreatic repair remains largely unexplored. This study evaluated the interactions of MSCs and SGCs with pancreatic islets using in vitro co-culture models, migration assays, apoptosis and proliferation analysis of HIT-T15 β-cells, and proteomic profiling. Both MSCs and SGCs demonstrated chemotactic responses to islet-derived factors. Co-culture with MSCs enhanced β-cell proliferation and reduced apoptosis, while SGCs increased proliferation without significant anti-apoptotic effects. Proteomic analysis and ligand-receptor mapping revealed that MSCs primarily influenced extracellular matrix remodeling and immunomodulation, whereas SGCs were enriched in adhesion and intercellular signaling pathways. Key pathways included collagen-integrin interactions, fibronectin signaling, and PECAM1-associated vascular support. These results indicate that SGCs, despite functional differences, possess regenerative properties comparable to MSCs and represent a promising alternative for cell-based support of pancreatic islets in diabetes therapy and transplantation contexts.
Keywords:
salivary gland cells
; mesenchymal stromal cells
; pancreatic islets
; pancreatic β-cells
; diabetes
; cell-based therapy
; ligand-receptor interactions
; apoptosis
; proliferation
; regenerative medicine
1. Introduction
Diabetes mellitus is a heterogeneous metabolic disease characterized by chronic hyperglycemia due to impaired insulin secretion and/or action. Type 1 diabetes (T1D) develops as a result of autoimmune destruction of the β-cells in the islets of Langerhans, leading to absolute insulin deficiency and the need for lifelong replacement therapy [1]. Type 2 diabetes (T2D) is associated with the development of peripheral insulin resistance and impaired compensatory insulin secretion by β-cells. Both types of the disease are accompanied by dysfunction of the endocrine pancreas and progressive remodeling of the tissue microenvironment [2] (pp.140-162). Modern therapeutic approaches aim not only to correct metabolic disturbances but also to restore the functional state of insulin-producing cells and their niche. In this context, cellular and molecular strategies in regenerative medicine, including the use of mesenchymal stromal cells and other types of regulatory cells to restore insulin secretory function and modulate the pancreatic islet microenvironment, are of particular interest.
The regeneration of pancreatic β-cells holds promise for restoring insulin-secreting cells in the islets of Langerhans through neogenesis from progenitor cells [3], replication of existing β-cells [4], or transdifferentiation from other cell types [5]. In adults, epimorphic regeneration is limited or absent, although β-cell regeneration may occur in response to metabolic demand [6]. Historically, islet transplantation has been a primary strategy; however, challenges such as immune rejection and donor scarcity persist [7]. In 2023, the FDA approved Lantidra for allogeneic transplantation of pancreatic islets from deceased donors for adults with T1D and severe hypoglycemia unresponsive to conventional therapies [8]. Advances in encapsulation and biocompatible substrates aim to improve transplantation outcomes, although further refinement is necessary for routine clinical application [9].
Cell-based approaches, including the differentiation of insulin-secreting cells from induced pluripotent or embryonic stem cells, offer additional therapeutic potential [10]. Moreover, reprogramming α-cells [11], pancreatic ductal cells [12], and other cell types [13] into β-cells via genetic modification presents a promising alternative. Gene-editing technologies may also correct β-cell defects, for example, in maturity-onset diabetes of the young (MODY) [14,15]. Despite the initial promise of islet transplantation, limited donor availability, poor engraftment, high islet attrition, and the need for prolonged immunosuppression hinder its widespread application.
Tissue engineering approaches, including organoids, biomimetic scaffolds, and “organs-on-a-chip,” have emerged to replicate pancreatic function and enhance insulin secretion, offering novel therapeutic solutions [16]. These advances highlight significant progress and the ongoing need for alternative cellular sources and regenerative strategies. Achieving success in this field requires interdisciplinary efforts not only to replace β-cells but also to regenerate the entire pancreatic microenvironment [17].
The islets of Langerhans comprise α-, β-, γ-, δ-, and endothelial cells, whose spatial organization and relative proportions govern endocrine function. Their architecture varies across species: in rodents, α-cells are predominantly located peripherally around a β-cell core, whereas in humans and primates, α- and β-cells are more interspersed [18].
Recent studies underscore the role of intercellular interactions in regulating insulin and glucagon secretion [19]. Cell adhesion molecules and the extracellular matrix are key determinants of islet functionality. Connexin 36 in β-cells and connexin 43 in endothelial cells facilitate synchronized insulin release, while ephrin signaling mediates α- and β-cell communication. Cadherins, particularly E-cadherin, promote intercellular adhesion and gap junction formation, while N-cadherin contributes to granule transport. The neural cell adhesion molecule (N-CAM) regulates F-actin dynamics and cadherin interactions, supporting vesicle trafficking and hormone secretion. The extracellular matrix, comprising laminin isoforms (211 and 511) and type IV collagen, provides structural support and integrative signaling cues. Collectively, these components enable adaptive endocrine activity and underscore the significance of islet architecture in functional regulation [20].
The pancreas and salivary glands share developmental, structural, and functional similarities. Both organs originate from the endoderm and undergo branched morphogenesis. Structurally, they contain acinar formations and ductal systems. While the pancreas possesses both exocrine and endocrine functions, the salivary gland, although solely exocrine, secretes a broad spectrum of bioactive peptides and growth factors, such as EGF, NGF, TGF-α, TGF-β, HGF, IGF-I, IGF-II, bFGF, cytokines and chemokines (IL-1, IL-6, IL-7, IL-18, TNF, BAFF, CXCL10, CXCL12, and CXCL13), and extracellular matrix components, contributing to tissue regeneration and immunomodulation [21,22].
Furthermore, salivary glands may influence pancreatic function via paracrine signaling. Growth factors secreted by salivary tissue, including EGF analogues, can exert protective effects on pancreatic tissue during injury, mitigating necrosis and dampening inflammatory responses [23]. Salivary glands consist of serous and mucous secretory cells, which produce enzyme-rich fluid and mucins, respectively. Notably, salivary gland cells have been shown to express genes for gastrointestinal hormones such as GIP, GLP-1, and PYY [24], and secrete these hormones contained in the plasma [25]. Additionally, insulin secretion by salivary cells has been demonstrated, offering potential for non-invasive diabetes monitoring [26].
Gene therapy approaches targeting salivary gland cells to express therapeutic proteins offer a novel strategy for systemic protein delivery. One promising method involves in situ genetic modification of salivary gland cells to secrete GLP-1 receptor agonists. For instance, AAV5-mediated delivery of Exendin-4 in Zucker (fa/fa) rats and high-fat diet mice achieved sustained expression, reducing weight gain and improving glycemic control [27]. Similarly, an AAV vector encoding a human proinsulin variant induced secretion of bioactive proinsulin, normalizing hyperglycemia in streptozotocin-induced diabetic mice [28].
Salivary glands are readily accessible for tissue biopsy, allowing ex vivo expansion and autologous or allogeneic therapeutic application [29]. However, diabetes may alter salivary gland secretory profiles, potentially compromising cell functionality and the feasibility of in vivo genetic modification or autotransplantation [30]. Nevertheless, ex vivo genetic engineering followed by transplantation remains a viable alternative.
Despite these advantages, research on salivary gland cell migration, homing capacity, and broader therapeutic applications remains limited. Further investigation into the molecular mechanisms and regenerative potential of these cells could expand their use in regenerative medicine.
Mesenchymal stromal cells have long been explored as therapeutic agents for various diseases. By 2024, over 590 clinical trials utilizing MSCs from different sources have been registered, including 29 focusing on diabetes treatment. The therapeutic potential of MSCs lies in their capacity to migrate to injured tissues and facilitate repair through paracrine signaling: secreting cytokines and growth factors [31] or differentiating into requisite cell types [32].
Numerous studies confirm the therapeutic efficacy of MSC-based therapies across a range of pathologies including diabetes [33,34,35]. MSCs exhibit low immunogenicity and secrete a diverse array of cytokines (including IL-6, IL-7, IL-8, and IL-28), chemokines (including CCL2, CCL20, CCR1, CCR2, and CXCR5), glycoproteins (FSTL1, TSG-6), and growth factors (VEGF, bFGF, KGF, IGF-1, IGF-2, and HGF) [36]. Even a single MSC administration can significantly improve glycemic control in both type 1 [37] and type 2 [38] diabetes, as well as in diabetic kidney disease [39] and nephropathy [40]. The MSC secretome enhances β-cell proliferation by modulating pathways such as FoxO1 [40], while also reducing pro-inflammatory cytokines like IL-1β and TNF-α [41].
MSCs possess a robust ability to home to inflammatory or damaged sites. Their migration involves interactions with endothelial adhesion molecules (e.g., P-selectin, CD44), and is primarily guided by the SDF-1/CXCR4 axis. Integrins and matrix metalloproteinases facilitate tissue penetration and anchorage [42]. MSCs have been shown to engraft in various tissues, including the pancreas, supporting their potential for islet repair and functional recovery [41]. Nonetheless, clinical translation of MSC therapies has faced challenges. One major limitation is the often insufficient activation of regenerative mechanisms in the target tissue. Strategies such as genetic modification or priming with bioactive molecules may enhance MSC efficacy [43].
Co-transplantation of MSCs with pancreatic islets improves islet viability and insulin secretion [44,45]. MSCs enhance transplant outcomes via immunomodulation and antifibrotic effects. Co-encapsulation strategies have demonstrated improved glycemic control in transplantation models [46,47], while microencapsulation creates a favorable microenvironment through N-cadherin-mediated interactions [48]. Indirect or mixed-contact cultures with MSCs show superior viability and insulin output compared with direct-contact models [49].
Despite their advantages, MSC-based therapies face several limitations. MSCs derived from different sources show heterogeneity in proliferative capacity, differentiation potential, and secretome composition [50,51]. This variability hinders standardization and therapeutic consistency [52]. Moreover, MSCs often exhibit poor long-term engraftment and survival after transplantation [35]. Extended in vitro expansion may reduce their functionality [53] and alter surface marker expression [54]. Although malignant transformation is rare, prolonged culture or genetic modification can increase oncogenic risk. Immune responses, particularly in allogeneic applications, may also reduce therapeutic efficacy [55].
In summary, MSCs remain a promising platform for regenerative therapies in diabetes due to their immunomodulatory properties and trophic effects. However, overcoming current challenges requires optimization through emerging biotechnologies.
Salivary gland cells (SGCs) and mesenchymal stromal cells (MSCs) differ in origin and biological function, which influences their therapeutic potential. SGCs show promise for genetic modification to express therapeutic proteins such as GLP-1 receptor agonists or insulin. AAV-based gene transfer has demonstrated improved glycemic control in preclinical models. Moreover, SGCs secrete bioactive factors akin to those in the pancreas, supporting tissue regeneration.
However, the absence of evidence regarding SGC migration and homing limits their application beyond the site of implantation. MSCs, by contrast, exhibit strong migratory and immunomodulatory properties via their rich secretome, supporting β-cell regeneration and immune regulation. Both cell types are readily isolated from human tissue. SGCs are accessible through minimally invasive biopsies for potential autologous use, though studies remain limited. MSCs are obtained from bone marrow, adipose tissue, or umbilical cord blood and can be expanded in vitro for therapeutic applications.
SGCs offer the advantage of physiological similarity to the pancreas and can be engineered to enhance therapeutic output. MSCs benefit from extensive research, clinical data, low immunogenicity, and regenerative capacity. However, inconsistent outcomes in MSC-based monotherapies highlight the need for further optimization and exploration of alternative or complementary cell types such as SGCs in regenerative medicine.
The aim of the study was to experimentally identify and analyze cell populations capable of modulating the state of the islets of Langerhans, including processes of regeneration, apoptosis, and the potential for targeted delivery of biological constructs.
The objectives of the study included the following tasks:
- Analysis of the migration potential of mesenchymal stromal cells (MSCs) and salivary gland cells (SGCs) towards pancreatic cells;
- Evaluation of proliferation and apoptosis levels in pancreatic cells under the influence of MSCs and SGCs;
- Identification of ligand-receptor interaction patterns involved in the formation of interactions between MSCs and pancreatic cells, and between SGCs and pancreatic cells;
- Identification of key signaling pathways involved in ligand-receptor interactions between MSCs, SGCs, and pancreatic cells.
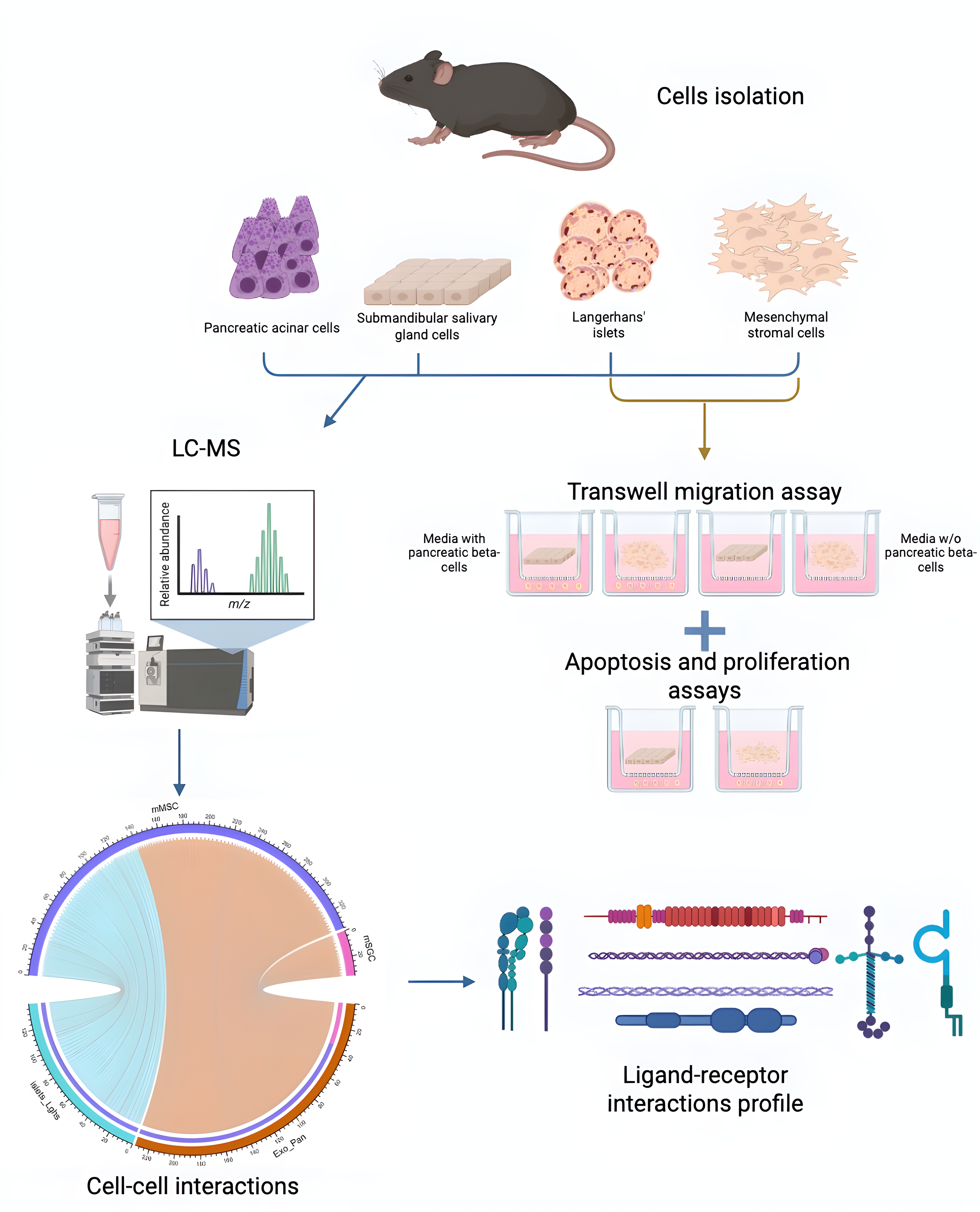
2. Materials and Methods
2.1. Animals and Experimental Procedures
Experiments were conducted on C57BL/6 mice with an average body weight of 22 grams. The animals were housed under standard conditions in the vivarium of the Institute of Developmental Biology of the Russian Academy of Sciences (IDB RAS), in cages containing 4–6 individuals, with ad libitum access to food and water. Animal care was supervised by the vivarium staff, and compliance with quarantine and sanitary regulations was monitored by the attending veterinarian. All experimental procedures were approved by the IDB RAS Institutional Animal Care and Use Committee (Protocol No. 74, dated 14.09.2023). Euthanasia was performed by decapitation.
2.2. Isolation and Cultivation of Mesenchymal Stromal Cells from Mouse Adipose Tissue
Following dissection of C57BL/6 mice, epididymal and intraperitoneal adipose tissues were collected and transferred into tubes containing 5 ml of 0.01% collagenase type II solution. The samples were incubated for 60 minutes at 37 °C. Subsequently, 10 ml of Dulbecco’s Modified Eagle Medium (DMEM) supplemented with 10% fetal bovine serum (FBS) was added. The suspension was centrifuged at 1500 rpm for 15 minutes at 4 °C. After removing the adipocyte fraction, the pellet was resuspended and centrifuged again at 1500 rpm for 5 minutes. The resulting pellet was resuspended in alpha-Minimum Essential Medium (α-MEM) supplemented with 10% FBS, 100 U/ml penicillin-streptomycin, and 4.5 mM Glutamax. The cells were seeded into T25 culture flasks. The primary culture of mouse mesenchymal stromal cells (mMSCs) was subsequently expanded in 75 cm² flasks using α-MEM supplemented with 10% FBS, 1% insulin-transferrin-selenium (ITS), 1% Glutamax, and 100 U/ml penicillin-streptomycin. Upon reaching approximately 70% confluence, the cells were passaged according to the standard protocol described below. The cultures were maintained in a CO₂ incubator at 37 °C with 5% CO₂ and constant humidity.
2.3. Isolation and Culture of Mouse SGCs
After dissection, mouse salivary glands were placed in Versene solution, minced (~2 mm fragments), and centrifuged twice (8000 rpm, 2 min). Collagenase type IV (2 mg/ml, 1.5–4 ml) was added and incubated (37°C, 5% CO2, 1 h). The cells were filtered (70 µm), centrifuged twice (8000 rpm, 2 min), and resuspended in DMEM/F12 medium. Mouse salivary gland cells (mSGCs) were cultured in T75 flasks with 12 ml DMEM/F12 + 10% FBS, 1% ITS, 1% Glutamax, 100 U/ml penicillin-streptomycin, and 50 ng/ml EGF. Culture plastic was pre-coated with collagen. At 70% confluence, the cells were passaged. The culture conditions were 37 °C, 5% CO2, and constant humidity.
2.4. Passaging of MSCs and SGCs
The cells were washed with Versene solution, then treated with 0.25% trypsin-EDTA (2 ml per 25 cm²). The detached cells were transferred to 15 ml tubes, diluted with DMEM + 10% FBS, centrifuged (200g, 5 min), resuspended in fresh medium, and seeded at 5000 cells/cm².
2.5. Boyden Chamber Assay
24-well plates with 6 µm pore polycarbonate membranes precoated with 300 µg/ml Matrigel were used. DiI-labeled MSCs or SGCs (30,000 cells/cm²) were seeded on the membrane. Langerhans islets in CnT-07 medium (with antibiotics, Glutamax, and 11 mM glucose) were added to the lower chamber as a chemoattractant. Control chambers received CnT-07 only. After 10 days, RFP+ cells in 7 fields of view were counted using a Juli Stage imager. Three biological replicates were analyzed.
2.6. Analysis of the Effects of MSCs and SGCs on Proliferation
24-well Transwell plates were taken. MSCs and SGCs were seeded into the upper well inserts of Transwell plate wells with a pore size of 0.4 μm at a density of 50 thousand cells per cm2. The inserts were precoated with Matrigel solution. 0.6 ml of culture medium was added to the lower well of the tablet and 0.2 ml of culture medium was added to the Transwell insert. In each variant, 10 thousand cells of immortalized Syrian hamster β-cell line HIT-T15 (CRL-1777) were seeded into the lower well chamber of the Transwell plate. Then immunocytochemical analysis was performed using antibodies to Ki67.
2.7. Immunocytochemical Analysis
Cell cultures were fixed in 10% buffered formalin for 15 minutes, washed three times with 0.01 M PBS, and then incubated for 30 minutes in a 0.1% Triton X-100 solution. Following permeabilization, samples were incubated for 30 minutes in a blocking buffer composed of 0.01 M PBS with 5% goat serum and 0.1% Tween 20. Samples were then stained with primary antibodies against Ki67 (ab16667, Rabbit, 1:200; Abcam, USA). Incubation with the primary antibody in blocking buffer was carried out overnight at +4 °C. The cultures were subsequently washed three times with 0.01 M PBS. For secondary antibody staining, Donkey anti-Rabbit IgG (H+L), Highly Cross-Adsorbed Secondary Antibody, Alexa Fluor 546 (A10040, Donkey, 1:1000; Invitrogen, USA) was applied. Staining was performed for 1 hour at room temperature in the dark, followed by three washes with 0.01 M PBS. Nuclei were stained with 100 ng/ml DAPI (Sigma Aldrich, USA) for 15 minutes, after which the cultures were washed three times with 0.01 M PBS (Sigma Aldrich, USA). Visualization was carried out using a Leica THUNDER imaging system, and data analysis was performed with Icy software.
2.8. Determination of DNA Fragmentation by Labeling DNA Fragments Using Terminal Deoxynucleotidyl Transferase and dUTP
The assay was performed according to the manufacturer's instructions according to Click-iT™ Plus TUNEL Assay Kit for In Situ Apoptosis Detection [56].
The cells were fixed for 15 min at room temperature in 4% paraformaldehyde and were then washed twice with PBS, each wash for 5 min. For permeabilization, the cells were incubated for 20 minutes in 0.25% Triton X-100 and washed. By way of a positive control, the tested cells were treated with DNase I for induction of DNA breaks for 30 minutes at room temperature and were then washed. The TdT reaction mixture was prepared according to the manufacturer's instructions: for 1 sample, 47 µl of buffer, 1 µl of EdUTP, and 2 µl of TdT enzyme were mixed. The mixture was then applied to the samples and incubated for 60 min at 37 °C. For the Click-iT reaction, a working solution with Alexa Fluor 594 secondary antibody was prepared by mixing 2625 µl of 1X Click-iT™ Plus TUNEL Reaction Buffer, 67 µl Copper Protectant, and 8.3 µl Alexa Fluor™ 594 picolyl azide according to the manufacturer's instructions. The solution was applied to the samples and the mixture was incubated for 30 minutes at 37 °C in the dark. For nuclei staining, the cells were incubated for 15 minutes with 4',6-diamidino-2-phenylindole. The cells were then washed 3 times with PBS. A Leica THUNDER imaging system was used for visualization. Data analysis was performed using the Icy software.
2.9. Proteomic Analysis
To investigate the key mechanisms underlying the interaction between cells demonstrating directed migration to the pancreas and endocrine pancreatic cells, a proteomic analysis of four cell types was performed:
1. Mouse salivary gland cells (mSGCs);
2. Mouse mesenchymal stem cells (mMSCs);
3. Mouse islets of Langerhans (islets);
4. Mouse exocrine pancreatic cells (Exo_Pan).
Sample preparation (three biological replicates per cell type) involved isolation of cells from C57BL/6 male mice. The cells were lysed, followed by trypsinolysis. Chromatography-mass spectrometry was performed on an OrbiTrap-type mass spectrometer using a chromatographic gradient of 120–180 minutes.
Primary data processing was performed using MaxQuant and the ms1searchpy package. General statistical analysis was conducted using Perseus and ms1searchpy. To assess interactions of MSCs and SGCs with pancreatic islets and exocrine cells, MSC and SGC proteins were treated as receptors, while proteins from islets and the exocrine pancreas were considered ligands.
Ligand-receptor analysis revealed stronger interactions between MSCs and both islets and exocrine pancreatic cells compared with SGCs, based on the number and affinity of ligand-receptor pairs. Functional enrichment analysis (GSEA) of these protein pairs was performed to annotate genes associated with cell migration processes using Gene Ontology (GO) terms.
2.10. Bioinformatics Analysis of Proteome Data
GSEA analysis was performed to identify specific mechanisms of cell-cell interactions. Differentially represented proteins were determined for each pair of comparison groups based on the ratio of mean protein abundance, as well as the combination of mean ratio and p-value.
Ligand-receptor interactions were constructed, statistically analyzed, and visualized using the scTalk and CellChat packages in R. Functional enrichment analysis was then conducted separately for ligands and receptors in each cell pair (FC ≥ 2, p-value ≤ 0.05) using the g:Profiler database (with gene IDs used instead of protein IDs).
3. Results
3.1. Analysis of SGC and MSC Migration Under the Influence of the Langerhans Islet Secretome
To assess the potential interaction between mMSCs/mSGCs and pancreatic islet cells, we hypothesized that the islet secretome may act as a chemoattractant for these cell types. To test this, a Boyden chamber assay was performed using mMSCs and mSGCs exposed to the pancreatic islet secretome, as described in the Materials and methods section.
The results demonstrated a statistically significant increase in the migration of both mMSCs and mSGCs in experimental groups where the islet secretome was present in the lower chamber, compared with the control wells without the secretome. In the absence of the islet secretome, migration levels did not differ significantly from the controls, confirming the chemoattractant effect of the secretome on both cell types. No significant difference in migration intensity was observed between mMSCs and mSGCs, suggesting similar chemotactic responses to islet-derived signals (Figure 2). These findings support the hypothesis that the pancreatic islet secretome promotes the migration of mMSCs and mSGCs, highlighting its potential role in guiding therapeutic cell populations for islet regeneration strategies.
Figure 1.
Results of mMSC and mSGC migration analysis during co-culture with Langerhans islets. Statistically significant differences in the migration level, compared with the negative control (wells without islets of Langerhans), are observed only in the variants where mMSCs and mSGCs were co-cultured with islets of Langerhans. No statistically significant differences in the migration level were found between mMSCs and mSGCs when co-cultured with islets of Langerhans. Statistical significance levels in the comparison groups: mMSCs + islets **p=0.001495; SGCs + pancreatic islets **p=0.003567.
Figure 1.
Results of mMSC and mSGC migration analysis during co-culture with Langerhans islets. Statistically significant differences in the migration level, compared with the negative control (wells without islets of Langerhans), are observed only in the variants where mMSCs and mSGCs were co-cultured with islets of Langerhans. No statistically significant differences in the migration level were found between mMSCs and mSGCs when co-cultured with islets of Langerhans. Statistical significance levels in the comparison groups: mMSCs + islets **p=0.001495; SGCs + pancreatic islets **p=0.003567.
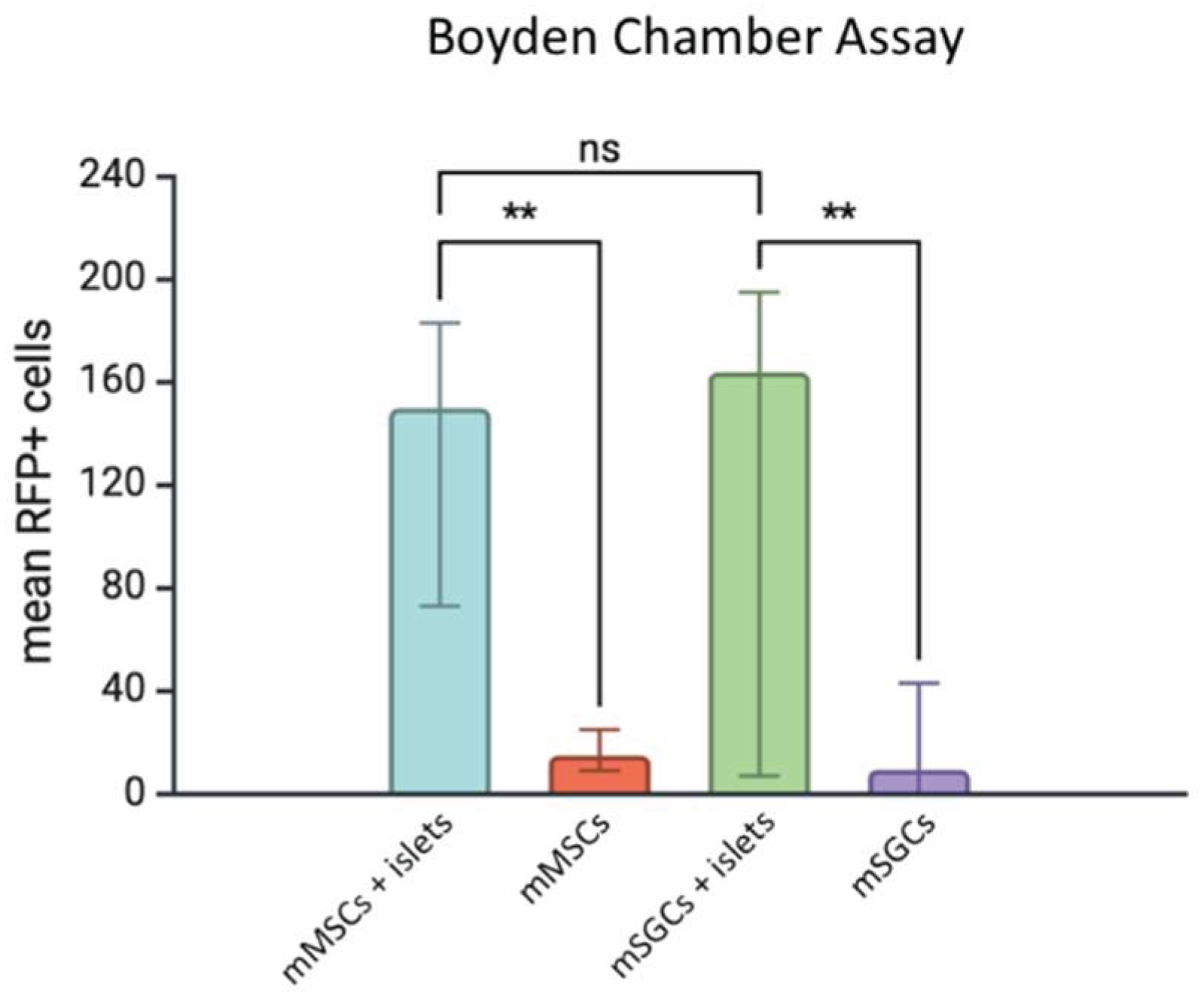
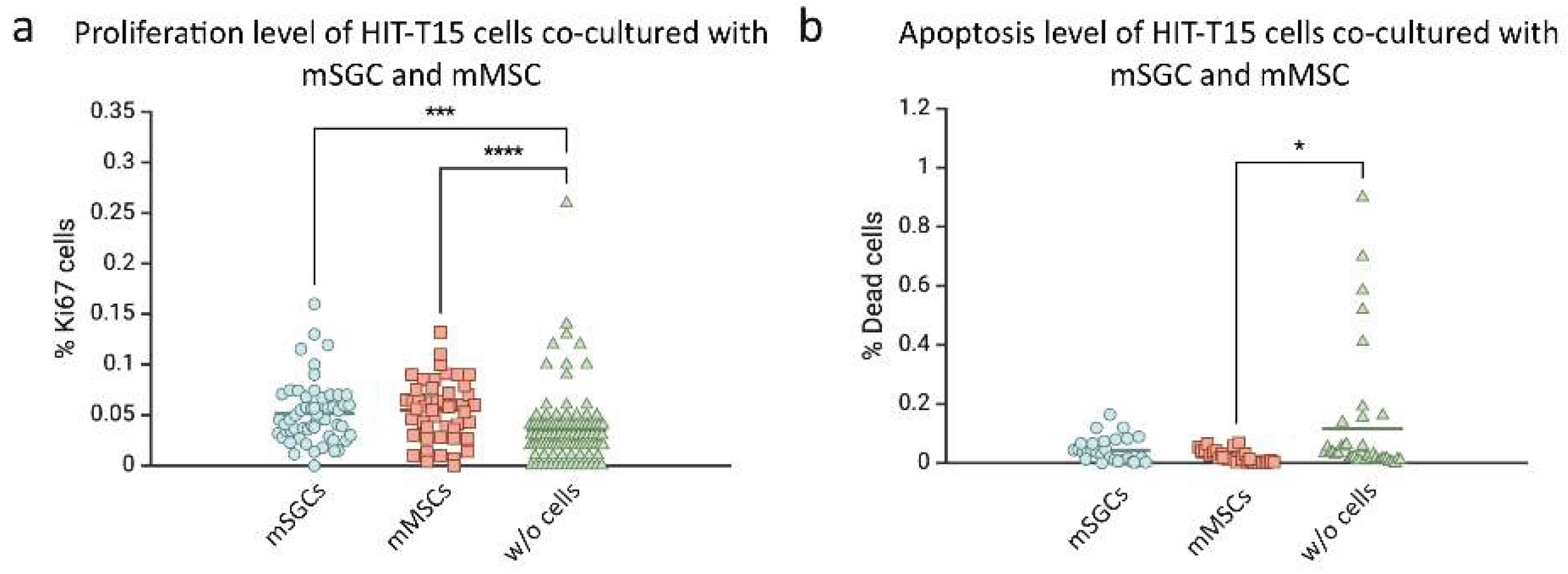
Figure 2.
Results of proliferation and apoptosis analysis in HIT-T15 (CRL-1777) cells during their co-culture with mSGCs and mMSCs. a) Analysis of HIT-T15 (CRL-1777) cell proliferation. In this case, there is a statistically significant increase in the level of HIT-T15 cell proliferation in the variants of co-culture with mMSCs and mSGCs, compared with the negative control (w/o cells: HIT-T15 cells cultured alone). b) Analysis of HIT-T15 cell apoptosis (CRL-1777). A statistically significant decrease in HIT-T15 cell apoptosis was observed only in the co-cultivation with mMSCs. Statistical significance levels in comparison groups: mMSCs+w/o cells ****p <0.0001; mSGCs+w/o cells ***p=0.0001030; mMSCs+w/o cells *p=0.02538.
Figure 2.
Results of proliferation and apoptosis analysis in HIT-T15 (CRL-1777) cells during their co-culture with mSGCs and mMSCs. a) Analysis of HIT-T15 (CRL-1777) cell proliferation. In this case, there is a statistically significant increase in the level of HIT-T15 cell proliferation in the variants of co-culture with mMSCs and mSGCs, compared with the negative control (w/o cells: HIT-T15 cells cultured alone). b) Analysis of HIT-T15 cell apoptosis (CRL-1777). A statistically significant decrease in HIT-T15 cell apoptosis was observed only in the co-cultivation with mMSCs. Statistical significance levels in comparison groups: mMSCs+w/o cells ****p <0.0001; mSGCs+w/o cells ***p=0.0001030; mMSCs+w/o cells *p=0.02538.
3.2. Influence of SGCs and mMSCs on the Proliferation and Apoptosis of HIT-T15 β-Cells
At the next stage, we analyzed the effect of mMSCs and mSGCs on the proliferation and apoptosis of HIT-T15 (CRL-1777) cells. HIT-T15 is a clonal cell line of SV40-immortalized pancreatic β-cells derived from the golden hamster (Mesocricetus auratus), widely used in experimental endocrinology as a stable and reproducible model with the ability to secrete insulin in a glucose-dependent manner. The use of a single cell line minimized biological variability and ensured data comparability when evaluating the effects of different cell types on the proliferation and apoptosis of insulin-producing cells.
To assess the impact of mMSCs and mSGCs, co-culture experiments were conducted in Transwell 24-well plates, as described in the Materials and methods section. The results demonstrated that co-culturing HIT-T15 cells with either mMSCs or mSGCs significantly increased β-cell proliferation compared with the control cultures without stromal cells. Proliferation levels were significantly higher in co-culture with mMSCs (p < 0.0001) and mSGCs (p = 0.000103) (Figure 3a). Apoptosis analysis revealed a cell type-dependent effect. Co-culture with mMSCs resulted in a significant reduction in the apoptosis of HIT-T15 cells (p = 0.02538), suggesting a cytoprotective effect of mMSCs. In contrast, co-culture with mSGCs did not alter apoptosis levels compared with the controls (Figure 3b). Proliferation and apoptosis were quantified using standardized methods, allowing direct comparison between the experimental groups based on total HIT-T15 cell counts per well.
Overall, the data indicate that mMSCs exert both proliferative and protective effects on β-cells, whereas mSGCs enhance proliferation but do not influence apoptosis. These findings suggest that the two cell types have differential mechanisms of action and potential therapeutic value for pancreatic tissue regeneration. The observed stimulation of proliferation in both co-culture conditions may reflect non-specific intercellular interactions, highlighting the need for further investigation of the underlying molecular mechanisms.
3.3. Proteomic Analysis of Intercellular Interactions Between SGCs, MSCs, and Pancreatic Cells
3.3.1. Volcano Plots and Functional Enrichment Analysis
To comprehensively investigate the intercellular interactions between mSGCs, mMSCs, and pancreatic cells (islets of Langerhans and exocrine pancreas), we identified ligands and receptors differentially upregulated in each cell type. A total of 6,301 proteins were identified across all samples. The number of proteins identified per cell type was: 5,018 for mSGCs, 5,630 for mMSCs, 4,142 for pancreatic islets (islets), and 5,037 for exocrine pancreatic cells (Exo_Pan).
Pairwise comparisons revealed the following numbers of differentially enriched proteins:
• mMSCs vs islets: 1,014 proteins enriched in mMSCs, 220 enriched in islets;
• mMSCs vs Exo_Pan: 758 enriched in mMSCs, 210 in Exo_Pan;
• mSGCs vs islets: 269 enriched in mSGCs, 327 in islets;
• mSGCs vs Exo_Pan: 54 enriched in mSGCs, 248 in Exo_Pan.
Based on these datasets, ligand-receptor pairs were identified using a confidence threshold of p < 0.05. The total number of ligand-receptor pairs was:
• 96 for mMSCs→Exo_Pan and 77 for Exo_Pan→ mMSCs;
• 76 for mMSCs→islets and 65 for islets→ mMSCs;
• 12 for mSGCs→Exo_Pan and 20 for Exo_Pan→ mSGCs;
• 9 for mSGCs→islets and 0 for islets→ mSGCs.
To improve the statistical accuracy and reduce background noise, proteins with a total spectral count (after normalization) below the number of samples were excluded. This filtering step enhanced the reliability of protein detection and improved data interpretability. The dataset was then re-normalized by recalculating each protein’s spectral count relative to the total number of spectra per sample, compensating for inter-sample variability and ensuring accurate comparisons of proteomic profiles.
Volcano plots were generated to visualize the distribution of proteins according to their statistical significance (–log10 p-value) and magnitude of change (log2 fold change). The majority of proteins clustered near the center of the plots, indicating no significant differences in expression. However, a substantial number of proteins were distributed beyond the significance thresholds (highlighted areas), reflecting differential expression (Figure 4). These data confirm specific proteomic differences between cell types and serve as a foundation for subsequent functional analysis of the identified proteins.
3.3.2. Identification of Key Proteins Involved in Intercellular Communication
At the next stage, to identify key proteins involved in intercellular signaling, interaction patterns were constructed for each cell type. Separate patterns were generated for outgoing interactions (ligands) and incoming interactions (receptors). Heat maps were used to visualize the distribution of outgoing and incoming signals across cell types (Figure 5).
The integrated heat map (Figure 5a) provides an overview of the contributions of each cell type to signal transmission and reception. Green and magenta areas indicate the most active signaling pathways involved in intercellular communication. A specific pattern of receptor protein expression in each cell subpopulation was also generated (Figure 5b), highlighting receptor complexes and signaling pathways predominantly engaged in specific cell groups, thereby reflecting the functional specificity of the interactions.
Additionally, a ligand-receptor interaction map was constructed, representing signal transmission between all four studied cell types (Figure 6). This analysis enabled the identification of cell populations that act primarily as signal senders or receivers, and revealed key receptor proteins involved in regulating intercellular communication. To quantify the overall strength of intercellular signaling between different cell types, a pie chart was generated (Figure 7), showing the number of ligand-receptor pairs identified in each group.
The analysis revealed that the highest number of ligand-receptor interactions occurred between mMSCs and Exo_Pan, indicating a high level of communication between these cell types. A substantial, though less prominent, number of interactions was observed between mMSCs and islets, suggesting a potential role of mMSCs in modulating endocrine pancreatic function. In contrast, the mSGC-islet group exhibited the minimal number of interactions, indicating relatively weak intercellular signaling in this pair.
In summary, these findings suggest that mMSCs demonstrate a high degree of interaction with both exocrine and endocrine pancreatic cells, supporting their potential role in maintaining tissue homeostasis and regulating intercellular signaling processes.
3.3.3. Functional Enrichment Analysis (GSEA) of mMSCs
GSEA analysis of interactions between mMSCs and pancreatic islets revealed a significant enrichment of biological processes related to cell migration (P < 0.01), suggesting a role for these interactions in modulating cell motility (Figure 8a). In the mMSC + Exo_Pan group, a broader range of enriched Gene Ontology Biological Processes (GO:BP) was observed, including terms related to cell adhesion, heterotypic cell-cell adhesion, cell migration, and cell spreading (Figure 8b).
To visualize ligand-receptor interactions contributing to these processes, a bubble plot was constructed (Figure 8c). The analysis demonstrated that mMSCs exhibit extensive interaction networks with both Langerhans islets and exocrine pancreatic cells. Key interactions were mediated by ligands such as collagen type I and VI subunits (Col1, Col6), THY1, and laminins (Lamb1, Lama2, and Lamc1), which bind to various integrin receptors expressed on pancreatic cells.
Notably, specific ligand-receptor interactions differed between cell groups. In the mMSC + islets group, unique interactions were observed between laminin subunits (Lamb1, Lama2, and Lamc1) and islet integrins. In the mMSC + Exo_Pan group, distinctive interactions included:
• Col6a4/Col6a5 binding to Itga3+Itgb1
• ESAM-ESAM homotypic interaction
• Pecam1-Pecam1 (potentially involving Cd38 co-expression)
• Vcam1-integrin interactions
• VWF-integrin binding
These results indicate the involvement of extracellular matrix components and adhesion molecules in mMSC-mediated communication with pancreatic cells.
3.4. Functional Enrichment Analysis of mSGCs
In the mSGC + Exo_Pan group, GSEA revealed enrichment of GO:BP terms associated with cell adhesion and migration (Figure 9a). This group displayed the most diverse set of ligand-receptor interactions, including ligands such as complement component 3 (C3), CD22, ICAM1, Pecam1, and Ptprc, interacting with receptors on exocrine pancreatic cells, including:
• Integrin complexes Itgam+Itgb2
• Ptprc
• Cd38
• Cd177
In contrast, the mSGC + islets group exhibited a limited number of interactions, represented by only two high-confidence ligand-receptor pairs:
• MIF - (Cd74+Cd44)
• THY1 - (Itgam+Itgb2)
Despite the low number of interactions, these pairs showed the highest predicted interaction probabilities among all analyzed groups (Figure 9b), suggesting a selective but potent communication axis between mSGCs and islet cells.
3.4.1. Similarities and Differences in mMSC and mSGC Protein Patterns
mMSCs exhibited a broad range of interactions with islets and Exo_Pan (96 pairs for mMSC+Exo_Pan, 77 pairs for Exo_Pan+mMSC; 76 pairs for mMSC+islets, and 65 pairs for islets+mMSC), including several signaling molecules involved in adhesion and migration. This highlights the substantial regulatory potential of mMSCs in intercellular communication. In contrast, mSGCs displayed fewer interactions (12 pairs for mSGC+Exo_Pan, 20 pairs for Exo_Pan+mSGC, and 9 pairs for mSGC+islets), with no interactions observed for islets+mSGC. This suggests a more limited role of mSGCs in the direct regulation of pancreatic endocrine cells. The primary differences between mMSC and mSGC protein patterns lie in their levels of cellular plasticity and intercellular communication. mMSCs are involved in more complex signaling networks, influencing cell adhesion, migration, and tissue regeneration, whereas mSGCs appear to engage in more specialized interactions.
3.4.2. Cell-Cell Interaction Patterns Across Different Roles
To better understand the functional differences between cell types, we examined their roles in specific signaling pathways. The roles of each cell type (Sender, Receiver, Mediator, Influencer) in pathways involving Collagen, FN1, ICAM, JAM, Laminin, MIF, PECAM1, and THY1 were compiled and presented in Figure 10 and Figure 11. Based on the ligand-receptor protein pairs analyzed, the distribution of roles is as follows: sender cells acting as the signal source; receiver cells that receive the signal; mediator cells that facilitate signal propagation; influencer cells that regulate or modulate signal transmission.
3.4.3. The Role of mMSCs in Intercellular Interactions
In the COLLAGEN signaling pathway, mMSCs demonstrated a strong ability to receive signals from islets and Exo_Pan, suggesting their key role in perceiving signals that regulate collagen-associated processes, such as tissue remodeling and fibrosis. Notably, MSCs can differentiate into myofibroblasts, contributing to fibrosis development. However, MSC infusion studies indicate their potential to alleviate fibrotic diseases by modulating inflammation, promoting tissue regeneration, and remodeling the extracellular matrix (ECM) [58].
In the FN1 (fibronectin) signaling pathway, mMSCs primarily acted as Senders of fibronectin signals, while Exo_Pan served as the Receiver. A similar, though less pronounced, pattern was observed for pancreatic islets, indicating the potential of mMSCs to modulate the extracellular matrix and their recruitment to pancreatic tissues for microenvironment regulation. In the ICAM signaling pathway, mMSCs influenced Exo_Pan, suggesting their involvement in regulating inflammatory processes and cell motility.
In the JAM signaling pathway, mMSCs interacted with Exo_Pan, implying their possible role in forming tight junctions that maintain barrier functions and facilitate immune interactions. In the PECAM1 (platelet endothelial cell adhesion molecule) signaling pathway, mMSCs exhibited potential interactions with Exo_Pan, possibly contributing to the stabilization of the endothelial network and supporting vascular structures.
4. Discussion
The clinical application of MSCs is limited by several factors, most notably the risk of neoplastic transformation during prolonged in vitro expansion [35] and potential immunogenicity, particularly in allogeneic use [55]. These limitations highlight the need for alternative cellular sources with high regenerative potential and improved safety profiles. In this context, salivary gland cells (SGCs) represent a promising candidate for cell-based therapies.
SGCs are capable of secreting a broad spectrum of bioactive molecules, including growth factors (e.g., EGF, NGF, TGF-α, TGF-β, HGF, IGF-I, IGF-II, and bFGF) [21,22], cytokines, and exosomes, containing different types of lipids, enzymes, proteins, and nucleic acids [59,60]. Studies have shown that extracellular vesicles derived from SGCs contain microRNAs and proteins that enhance the survival and functional activity of insulin-producing cells [61]. Furthermore, SGCs secrete key extracellular matrix components such as laminin, collagen, and fibronectin, thereby contributing to a supportive microenvironment for tissue repair [62] (pp. 48–77).
An additional advantage of SGCs is their accessibility and the minimally invasive nature of their collection. Unlike MSCs, which typically require bone marrow or adipose tissue biopsies, SGCs can be obtained via less invasive procedures, such as fine-needle aspiration, resection biopsy, or labial minor salivary gland biopsy [63], making them more practical for clinical applications.
In summary, SGCs offer a promising alternative to MSCs due to their immunomodulatory properties, secretion of bioactive molecules, and regenerative potential. Their use may provide an effective strategy for treating diseases associated with insulin-producing cell dysfunction and broader applications in regenerative medicine.
4.1. Migration Properties of SGCs and MSCs Under the Influence of the Langerhans Islet Secretome
The results of this study demonstrated that the presence of the Langerhans islet secretome in the lower chamber significantly enhanced the migration of both SGCs (p = 0.003567) and MSCs (p = 0.001495), compared with the control groups with empty wells. In contrast, conditions lacking pancreatic islets did not induce statistically significant migration, confirming the chemoattractant role of the islet secretome.
No significant difference between SGCs and MSCs was observed in the migration levels, suggesting that both cell types exhibit comparable chemotactic responsiveness to pancreatic islet-derived signals. Evaluating cell migration potential is crucial for assessing therapeutic applicability, as targeted cell homing to injury sites or their use as delivery vectors can enhance the efficacy of regenerative therapies. MSCs are well known for their robust chemotactic response to signals from injured tissues [41], and our results confirm their directed migration toward Langerhans islets in vitro, consistent with previous findings.
Notably, SGCs exhibited a similar level of migration, highlighting their potential utility in the treatment of diabetes and other disorders associated with islet dysfunction. This suggests that SGCs possess receptor systems capable of responding to inflammatory and chemotactic cues, making them promising candidates for therapeutic transplantation. Further investigation into the molecular mechanisms underlying SGC migration is warranted to better understand their interaction with injured tissues and to identify key regulatory signals. If SGCs share homing mechanisms with MSCs, their clinical application may be simplified, particularly given their easier accessibility and lower risk of tumorigenic transformation.
Overall, the findings support the high potential of SGCs as an alternative cell source for diabetes therapy and regenerative medicine, warranting further research into their molecular and functional properties.
4.1.1. Effect of SGCs and MSCs on the Proliferation and Apoptosis of HIT-T15 β-Cells
The results showed that both SGCs (p = 0.0001030) and MSCs (p < 0.0001) significantly stimulated the proliferation of the pancreatic β-cell line HIT-T15 (CRL-1777). Additionally, MSCs exerted a protective effect by reducing apoptosis in HIT-T15 cells (p = 0.02538), while SGCs did not induce statistically significant changes in cell death.
Despite the active use of MSCs in regenerative medicine, their clinical application is constrained by several limitations, as discussed above. This underscores the need to identify alternative cell sources. Compared with MSCs, salivary gland cells are more accessible and can be obtained via less invasive procedures. SGCs secrete a broad array of bioactive factors, including growth factors, cytokines, and exosomes, which may contribute to β-cell proliferation.
Moreover, SGC-derived factors may also promote angiogenesis, supporting improved vascularization of transplanted Langerhans islets. In addition, cell-cell interactions mediated by integrins and N-cadherin are critical for maintaining β-cell function. In this study, ligand-receptor protein pairs involved in interactions between integrins and Langerhans islets or pancreatic acinar cells were identified, further highlighting the potential of SGCs for supporting β-cell survival and function.
These findings reinforce the therapeutic relevance of SGCs as an alternative or complementary cellular component to MSCs in β-cell support and regenerative medicine.
4.2. Proteomic Analysis
Proteomic analysis revealed distinct patterns of intercellular interactions between MSCs, SGCs, and both pancreatic exocrine (Exo_Pan) and endocrine (pancreatic islets) cells. Ligand-receptor pair and GSEA analyses showed that MSCs exhibited a higher degree of potential interactions with both pancreatic exocrine and endocrine cells, whereas SGCs showed weaker interactions with islet cells. Despite the reduced number of interactions with islets, GSEA analysis indicated that SGC interactions with exocrine cells activated key processes such as cell adhesion, migration, and intercellular communication.
The interaction of MSCs with Exo_Pan was more pronounced than that with islet cells, suggesting that MSCs may exert a less specific effect on pancreatic tissues. This could explain their propensity to induce fibrosis rather than promote targeted regeneration of islets. Interaction analysis identified several key MSC-secreted ligands, including collagens (Col1, Col6), laminins (Lamb1, Lama2, and Lamc1), and cell adhesion molecules (THY1, Pecam1, and Vcam1). However, these ligands primarily bind to exocrine cell integrins rather than the β-cells of the islets of Langerhans, raising questions about the specificity of their therapeutic effects.
Although the number of interactions between SGCs and islets of Langerhans was lower than that observed with MSCs, ligand-receptor analysis indicated that SGC signaling may be more selective. The most significant interactions in the SGC+pancreatic islet group involved the Mif+(Cd74+Cd44) and THY1+(Itgam+Itgb2) pairs, which may play critical roles in modulating β-cell metabolic activity and stress resistance. SGC interactions with Exo_Pan were more diverse and included activation of cell adhesion mechanisms, which may contribute to the formation of a favorable microenvironment for Langerhans islets.
These findings suggest that salivary gland cells hold promise as a viable alternative to MSCs in cell therapy for diabetes. They offer several advantages, including easier derivation, greater safety, and more targeted intercellular interactions with islets of Langerhans.
4.2.1. Protein Patterns Involved in the Interaction Between MSCs and Islets of Langerhans
Integrins are crucial cell adhesion receptors that regulate the proliferation, migration, and functional activity of β-cells in the islets of Langerhans. The interaction between integrins and the extracellular matrix ensures the structural integrity of the islets and modulates insulin secretion.
In our study, the ligand-receptor interaction analysis identified key MSC proteins involved in interactions with pancreatic islets:
• Integrin α6β1: Binds to laminin, a major component of the basal membrane in islets of Langerhans. Elevated levels of integrin α6β1 are associated with enhanced glucose-stimulated insulin secretion in rat islets of Langerhans [20].
• Integrin α3β1: Binds both laminin and collagen and is involved in cell adhesion, migration, and proliferation. Loss of α3β1 impairs β-cell proliferation in rodents during both pre- and postnatal periods [20].
• Integrin αvβ3: Interacts with vitronectin and osteopontin to regulate MSC adhesion and migration.
It is important to note that integrins β1, β3, and β5 exhibit functional compensation at the transcriptional, translational, and post-translational levels. Blockade of β1 results in the activation of β3 and/or β5 functions, supporting cell differentiation [64], indicating the ability of cells to adapt to microenvironmental changes while maintaining adhesion and signaling functions.
The THY1 protein on MSCs plays a role in adhesion, differentiation, and immune response modulation, which is critical for protecting islets of Langerhans from immune rejection during transplantation.
Thus, the identified interaction pattern highlights the mechanisms by which MSCs influence the maintenance of the microenvironment and functional activity of Langerhans islets.
4.2.2. Key Proteins Involved in the Interactions Between MSCs and Exocrine Cells of the Pancreas
Analysis of MSC interactions with pancreatic exocrine cells identified several key proteins:
• PECAM-1 (CD31) and CD38: These proteins regulate intercellular contacts and influence islet survival and insulin secretion. They also play a role in the transformation of vascular cells into myofibroblast-like cells, aiding tissue healing [46].
• N-cadherin and ICAM-1: These proteins facilitate MSC adhesion to islets, improving the viability and functionality of the latter.
• Adiponectin (ADIPOQ): Through its receptor AdipoR1, adiponectin protects β-cells from apoptosis under metabolic stress (e.g., increased fatty acids or inflammation), enhances insulin secretion, and reduces lipotoxicity. This protein is crucial for maintaining β-cell function and supports β-cell survival and function in diabetes [65,66].
• Von Willebrand Factor (VWF): A key glycoprotein involved in hemostasis, angiogenesis, and cell adhesion [67]. It works in conjunction with integrins ITGAV (alpha-V) and ITGB3 (beta-3) to support new blood vessel formation and endothelial cell stabilization.
• VCAM-1: Interacting with integrins ITGA4 and ITGB1, VCAM-1 promotes MSC migration to damaged tissues, enhances their regenerative properties, and supports islet survival. It also has an immunomodulatory effect, reducing inflammation around islets and potentially improving the microenvironment for insulin-producing β-cells.
4.2.3. Interactions Between SGCs and Islets of Langerhans
This study identified key protein pairs involved in the interactions between salivary gland cells (SGCs) and islets of Langerhans:
• THY-1 (CD90): The interaction of THY-1 with integrins such as ITGAM (CD11b) and ITGB2 is critical for adhesion, migration, and immune modulation, particularly around islets of Langerhans. THY-1 regulates adhesion and migration through integrin connections, influencing inflammation in islet transplantation, such as in type 1 diabetes. These interactions involve mechanotransduction, modulate immune responses, and maintain cell survival through actin cytoskeleton remodeling. In transplantation, THY-1 on MSCs reduces inflammation and enhances angiogenesis, promoting islet viability [68].
• CD44: Binds to hyaluronan, increasing adhesion, survival, and functionality of islets, reducing inflammation and promoting regeneration [69].
• CD74: Regulates immune responses by modulating cell proliferation and migration in complex with CD44, maintaining the islet microenvironment and supporting islet viability.
4.2.4. SGC Interaction with the Exocrine Pancreas
The interaction of SGCs with exocrine pancreatic cells reveals a potential interaction between C3 complement components and ITGAM (CD11b) and ITGB2 (CD18) integrins, mediated through the MAC-1 complex. This interaction plays a key role in leukocyte adhesion and migration, crucial for immune regulation in the pancreas. These processes may help reduce inflammation and improve the survival of Langerhans islet grafts, with SGCs interacting with ITGB2 to regulate adhesion and the innate immune response.
• CD22: This protein regulates B cells via CD45 (PTPRC) and is involved in modulating immune cell signaling. It affects tyrosine phosphorylation, which reduces B cell signaling activity, potentially regulating inflammation in the Langerhans islet microenvironment [70].
• ICAM1: On SGCs, ICAM1 binds to ITGAL (CD11a) and ITGB2 to form the LFA-1 complex, regulating the adhesion and migration of immune cells, particularly macrophages and neutrophils. This interaction plays a key role in mediating immune interactions between SGCs and the local immune environment of the islets. Through ICAM1, SGCs can modulate the local immune response, potentially reducing inflammation and enhancing islet cell survival [71].
These findings suggest that SGCs possess regenerative properties comparable with MSCs and may be effective in improving the transplantation of Langerhans islets and supporting their functional activity.
4.2.5. Signaling Pathways
The study of signaling pathways active in SGCs and MSCs has revealed key differences and similarities in their regulatory functions, which influence their potential for pancreatic tissue regeneration.
Analysis of intercellular interaction patterns has demonstrated that MSCs are actively involved in signaling pathways associated with the extracellular matrix. In the COLLAGEN pathway, MSCs exhibit a strong ability to receive signals from pancreatic islets and exocrine pancreatic cells (Exo_Pan), highlighting the role of MSCs in tissue remodeling and fibrosis. In the fibronectin (FN1) signaling pathway, MSCs primarily act as a signal source, influencing the microenvironment of β-cells. The active participation of MSCs in ICAM and JAM signaling cascades underscores their role in regulating inflammatory processes and maintaining the barrier functions of pancreatic cells. Interactions in the PECAM1 pathway suggest that MSCs contribute to the maintenance of vascular structures, which is crucial for angiogenesis and the survival of islets of Langerhans.
SGCs exhibit a similar potential for regulating the tissue microenvironment but with greater specificity towards pancreatic islets. In the Laminin signaling pathway, β-cells serve as the primary Senders, maintaining interactions within islets, which aligns with their function in insulin secretion and cellular homeostasis regulation. SGCs show a high activity in the MIF signaling pathway, which indicates their involvement in regulating inflammation and tissue stability. Their significant role in THY1 signaling further emphasizes their contribution to maintaining structural organization and interactions with pancreatic islets.
A comparison of both cell types reveals that MSCs are more actively involved in matrix-dependent processes, including tissue remodeling, interaction with vascular structures, and immune modulation. In contrast, SGCs display a high activity in inflammation-related pathways (MIF, THY1) and contribute to tissue adhesion and structural support (Laminin), which positions them as strong candidates for pancreatic regeneration.
These findings suggest that SGCs possess regenerative properties comparable with MSCs and can be considered a promising alternative for cell therapy focused on β-cell regeneration and enhancing the islet microenvironment.
5. Conclusions
The proteomic analysis and evaluation of protein interaction patterns in this study demonstrate that SGCs are capable of interacting with pancreatic cells and activating key signaling pathways, including ICAM, JAM, FN1, and THY1. These findings suggest that SGCs play a significant role in maintaining tissue homeostasis, supporting cell adhesion and modulating immune responses.
When comparing the signaling pathways of MSCs and SGCs, it is evident that while SGCs can secrete factors that influence the microenvironment of islets of Langerhans, their immunomodulatory activity is less pronounced than that of MSCs. Nonetheless, SGCs secrete factors similar to those secreted by MSCs, positioning them as a potential alternative for cell therapy.
The biological similarity between the salivary gland and the pancreas further supports the potential of SGCs as a promising source for cell therapy, particularly in the context of β-cell transplantation and regeneration. Given their regenerative properties, SGCs could serve as a viable alternative to MSCs in diabetes therapy, with particular promise in the development of novel approaches for pancreatic β-cell regeneration.
In conclusion, the results of this study highlight the need for further investigation into SGCs as a promising tool for cell therapy. Additionally, exploring methods for gene modification to enhance the therapeutic potential of SGCs could significantly improve their applicability in regenerative medicine.
Author Contributions
Writing - original draft, investigation, validation and methodology E.R., M.C.; writing - review & editing, software, data curation, visualization and formal analysis E.C., I.S.; supervision, resources, funding acquisition, project administration and writing - review & editing V.G. and A.V. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Government programs of basic research in Koltzov Institute of Developmental Biology of the Russian Academy of Sciences in 2025 № 0088-2025-0002 and № 0088- 2025-0005.
Institutional Review Board Statement
The animal study protocol was approved by the Institutional Ethics Committee of the Institute of Developmental Biology of the Russian Academy of Sciences (Protocol No. 74, dated 14.09.2023).
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in this study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author.
Acknowledgments
We thank Dr. Natasha Grigorian for her help with proofreading the manuscript. Also we would like to acknowledge the Core Centrum «Genomic Technologies Group», the Cell Culture Collections for Biotechnological and Biomedical Research of the Koltzov IDB RAS and Core Centrum «Genomics, proteomics, metabolomics» of the Lopukhin FRCC PCM for providing the necessary facilities and equipment.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| MDPI | Multidisciplinary Digital Publishing Institute |
| DOAJ | Directory of open access journals |
| TLA | Three letter acronym |
| LD | Linear dichroism |
| MSCs | Mesenchymal stromal cells |
| SGCs | Salivary gland cells |
| iPSCs | Induced pluripotent stem cells |
| MODY | Maturity-onset diabetes of the young |
| GLP-1 | Glucagon-like peptide-1 |
| GSEA | Gene Set Enrichment Analysis |
| GO | Gene Ontology |
| FC | Flow cytometry |
| IHC | Immunohistochemistry |
| Exo_Pan | Exocrine pancreas |
| FN1 | Fibronectin-1 |
References
- Katsarou, A.; Gudbjörnsdottir, S.; Rawshani, A.; Dabelea, D.; Bonifacio, E.; Anderson, B.J.; Jacobsen, L.M.; Schatz, D.A.; Lernmark, Å. Type 1 Diabetes Mellitus. Nat Rev Dis Primers 2017, 3, 17016. [Google Scholar] [CrossRef]
- Edwards, N. Disease Pathways: An Atlas of Human Disease Signaling Pathways by Anastasia Nesterova, Anton Yuryev, Eugene Klimov, Maria Zharkova, Maria Shkrob, Natalia Ivanikova, Sergey Sozin and Vladimir Sobolev. The Biochemist 2020, 42, 140–162. [Google Scholar] [CrossRef]
- Niu, F.; Liu, W.; Ren, Y.; Tian, Y.; Shi, W.; Li, M.; Li, Y.; Xiong, Y.; Qian, L. β-Cell Neogenesis: A Rising Star to Rescue Diabetes Mellitus. Journal of Advanced Research 2024, 62, 71–89. [Google Scholar] [CrossRef]
- Spears, E.; Serafimidis, I.; Powers, A.C.; Gavalas, A. Debates in Pancreatic Beta Cell Biology: Proliferation Versus Progenitor Differentiation and Transdifferentiation in Restoring β Cell Mass. Front. Endocrinol. 2021, 12, 722250. [Google Scholar] [CrossRef]
- Guney, M.A.; Lorberbaum, D.S.; Sussel, L. Pancreatic β Cell Regeneration: To β or Not to β. Current Opinion in Physiology 2020, 14, 13–20. [Google Scholar] [CrossRef]
- Soldovieri, L.; Di Giuseppe, G.; Ciccarelli, G.; Quero, G.; Cinti, F.; Brunetti, M.; Nista, E.C.; Gasbarrini, A.; Alfieri, S.; Pontecorvi, A.; et al. An Update on Pancreatic Regeneration Mechanisms: Searching for Paths to a Cure for Type 2 Diabetes. Molecular Metabolism 2023, 74, 101754. [Google Scholar] [CrossRef]
- Wang, Q.; Huang, Y.; Liu, L.; Zhao, X.; Sun, Y.; Mao, X.; Li, S. Pancreatic Islet Transplantation: Current Advances and Challenges. Front. Immunol. 2024, 15, 1391504. [Google Scholar] [CrossRef]
- Parums, D.V. Editorial: First Regulatory Approval for Allogeneic Pancreatic Islet Beta Cell Infusion for Adult Patients with Type 1 Diabetes Mellitus. Med Sci Monit 2023, 29. [Google Scholar] [CrossRef]
- Liu, S.S.; Shim, S.; Kudo, Y.; Stabler, C.L.; O’Cearbhaill, E.D.; Karp, J.M.; Yang, K. Encapsulated Islet Transplantation. Nat Rev Bioeng 2024, 3, 83–102. [Google Scholar] [CrossRef]
- Wang, X.; Gao, M.; Wang, Y.; Zhang, Y. The Progress of Pluripotent Stem Cell-Derived Pancreatic β-Cells Regeneration for Diabetic Therapy. Front. Endocrinol. 2022, 13, 927324. [Google Scholar] [CrossRef]
- Guo, P.; Zhang, T.; Lu, A.; Shiota, C.; Huard, M.; Whitney, K.E.; Huard, J. Specific Reprogramming of Alpha Cells to Insulin-Producing Cells by Short Glucagon Promoter-Driven Pdx1 and MafA. Molecular Therapy - Methods & Clinical Development 2023, 28, 355–365. [Google Scholar] [CrossRef]
- Lee, J.; Sugiyama, T.; Liu, Y.; Wang, J.; Gu, X.; Lei, J.; Markmann, J.F.; Miyazaki, S.; Miyazaki, J.; Szot, G.L.; et al. Expansion and Conversion of Human Pancreatic Ductal Cells into Insulin-Secreting Endocrine Cells. eLife 2013, 2, e00940. [Google Scholar] [CrossRef]
- Fontcuberta-PiSunyer, M.; García-Alamán, A.; Prades, È.; Téllez, N.; Alves-Figueiredo, H.; Ramos-Rodríguez, M.; Enrich, C.; Fernandez-Ruiz, R.; Cervantes, S.; Clua, L.; et al. Direct Reprogramming of Human Fibroblasts into Insulin-Producing Cells Using Transcription Factors. Commun Biol 2023, 6, 256. [Google Scholar] [CrossRef]
- Ng, N.H.J.; Ghosh, S.; Bok, C.M.; Ching, C.; Low, B.S.J.; Chen, J.T.; Lim, E.; Miserendino, M.C.; Tan, Y.S.; Hoon, S.; et al. HNF4A and HNF1A Exhibit Tissue Specific Target Gene Regulation in Pancreatic Beta Cells and Hepatocytes. Nat Commun 2024, 15, 4288. [Google Scholar] [CrossRef]
- Hu, M.; Cherkaoui, I.; Misra, S.; Rutter, G.A. Functional Genomics in Pancreatic β Cells: Recent Advances in Gene Deletion and Genome Editing Technologies for Diabetes Research. Front. Endocrinol. 2020, 11, 576632. [Google Scholar] [CrossRef]
- Berishvili, E.; Peloso, A.; Tomei, A.A.; Pepper, A.R. The Future of Beta Cells Replacement in the Era of Regenerative Medicine and Organ Bioengineering. Transpl Int 2024, 37, 12885. [Google Scholar] [CrossRef]
- Bourgeois, S.; Coenen, S.; Degroote, L.; Willems, L.; Van Mulders, A.; Pierreux, J.; Heremans, Y.; De Leu, N.; Staels, W. Harnessing Beta Cell Regeneration Biology for Diabetes Therapy. Trends in Endocrinology & Metabolism 2024, 35, 951–966. [Google Scholar] [CrossRef]
- Arrojo E Drigo, R.; Ali, Y.; Diez, J.; Srinivasan, D.K.; Berggren, P.-O.; Boehm, B.O. New Insights into the Architecture of the Islet of Langerhans: A Focused Cross-Species Assessment. Diabetologia 2015, 58, 2218–2228. [Google Scholar] [CrossRef]
- Cottet Dumoulin, D.; Fonseca, L.M.; Bignard, J.; Hanna, R.; Parnaud, G.; Lebreton, F.; Bellofatto, K.; Berishvili, E.; Berney, T.; Bosco, D. Identification of Newly Synthetized Proteins by Mass Spectrometry to Understand Palmitate-Induced Early Cellular Changes in Pancreatic Islets. American Journal of Physiology-Endocrinology and Metabolism, 0019. [Google Scholar] [CrossRef]
- Wieland, F.C.; Van Blitterswijk, C.A.; Van Apeldoorn, A.; LaPointe, V.L.S. The Functional Importance of the Cellular and Extracellular Composition of the Islets of Langerhans. Journal of Immunology and Regenerative Medicine 2021, 13, 100048. [Google Scholar] [CrossRef]
- Chibly, A.M.; Aure, M.H.; Patel, V.N.; Hoffman, M.P. Salivary Gland Function, Development, and Regeneration. Physiological Reviews 2022, 102, 1495–1552. [Google Scholar] [CrossRef]
- Shang, Y.F.; Shen, Y.Y.; Zhang, M.C.; Lv, M.C.; Wang, T.Y.; Chen, X.Q.; Lin, J. Progress in Salivary Glands: Endocrine Glands with Immune Functions. Front. Endocrinol. 2023, 14, 1061235. [Google Scholar] [CrossRef]
- Tiffon, C. Defining Parallels between the Salivary Glands and Pancreas to Better Understand Pancreatic Carcinogenesis. Biomedicines 2020, 8, 178. [Google Scholar] [CrossRef]
- Sangsuriyothai, P.; Watari, I.; Serirukchutarungsee, S.; Satrawaha, S.; Podyma-Inoue, K.A.; Ono, T. Expression of Glucagon-like Peptide-1 and Glucose-Dependent Insulinotropic Polypeptide in the Rat Submandibular Gland Is Influenced by Pre- and Post-Natal High-Fat Diet Exposure. Front. Physiol. 2024, 15, 1357730. [Google Scholar] [CrossRef]
- Li, B.-B.; Chen, Z.-B.; Li, B.-C.; Lin, Q.; Li, X.-X.; Li, S.-L.; Ding, C.; Wu, L.-L.; Yu, G.-Y. Expression of Ghrelin in Human Salivary Glands and Its Levels in Saliva and Serum in Chinese Obese Children and Adolescents. Archives of Oral Biology 2011, 56, 389–394. [Google Scholar] [CrossRef]
- Sahakian, B.J.; Lean, M.E.J.; Robbins, T.W.; James, W.P.T. Salivation and Insulin Secretion in Response to Food in Non-Obese Men and Women. Appetite 1981, 2, 209–216. [Google Scholar] [CrossRef]
- Di Pasquale, G.; Dicembrini, I.; Raimondi, L.; Pagano, C.; Egan, J.M.; Cozzi, A.; Cinci, L.; Loreto, A.; Manni, M.E.; Berretti, S.; et al. Sustained Exendin-4 Secretion through Gene Therapy Targeting Salivary Glands in Two Different Rodent Models of Obesity/Type 2 Diabetes. PLoS ONE 2012, 7, e40074. [Google Scholar] [CrossRef]
- Rowzee, A.M.; Perez-Riveros, P.J.; Zheng, C.; Krygowski, S.; Baum, B.J.; Cawley, N.X. Expression and Secretion of Human Proinsulin-B10 from Mouse Salivary Glands: Implications for the Treatment of Type I Diabetes Mellitus. PLoS ONE 2013, 8, e59222. [Google Scholar] [CrossRef]
- Beucler, M.J.; Miller, W.E. Isolation of Salivary Epithelial Cells from Human Salivary Glands for In Vitro Growth as Salispheres or Monolayers. JoVE 2019, 59868. [Google Scholar] [CrossRef]
- Fouani, M.; Basset, C.A.; Jurjus, A.R.; Leone, L.G.; Tomasello, G.; Leone, A. Salivary Gland Proteins Alterations in the Diabetic Milieu. J Mol Histol 2021, 52, 893–904. [Google Scholar] [CrossRef]
- Sun, F.; Sun, Y.; Wu, F.; Xu, W.; Qian, H. Mesenchymal Stem Cell-Derived Extracellular Vesicles: A Potential Therapy for Diabetes Mellitus and Diabetic Complications. Pharmaceutics 2022, 14, 2208. [Google Scholar] [CrossRef]
- Badr, O.I.; Kamal, M.M.; El-Maraghy, S.A.; Ghaiad, H.R. The Effect of Diabetes Mellitus on Differentiation of Mesenchymal Stem Cells into Insulin-Producing Cells. Biol Res 2024, 57, 20. [Google Scholar] [CrossRef]
- Habiba, U.E.; Khan, N.; Greene, D.L.; Ahmad, K.; Shamim, S.; Umer, A. Meta-Analysis Shows That Mesenchymal Stem Cell Therapy Can Be a Possible Treatment for Diabetes. Front. Endocrinol. 2024, 15, 1380443. [Google Scholar] [CrossRef]
- Ahmed, Z.T.; Zain Al-Abeden, M.S.; Al Abdin, M.G.; Muqresh, M.A.; Al Jowf, G.I.; Eijssen, L.M.T.; Haider, K.H. Dose-Response Relationship of MSCs as Living Bio-Drugs in HFrEF Patients: A Systematic Review and Meta-Analysis of RCTs. Stem Cell Res Ther 2024, 15, 165. [Google Scholar] [CrossRef]
- Shan, Y.; Zhang, M.; Tao, E.; Wang, J.; Wei, N.; Lu, Y.; Liu, Q.; Hao, K.; Zhou, F.; Wang, G. Pharmacokinetic Characteristics of Mesenchymal Stem Cells in Translational Challenges. Sig Transduct Target Ther 2024, 9, 242. [Google Scholar] [CrossRef]
- Han, Y.; Yang, J.; Fang, J.; Zhou, Y.; Candi, E.; Wang, J.; Hua, D.; Shao, C.; Shi, Y. The Secretion Profile of Mesenchymal Stem Cells and Potential Applications in Treating Human Diseases. Sig Transduct Target Ther 2022, 7, 92. [Google Scholar] [CrossRef]
- Wang, M.; Song, L.; Strange, C.; Dong, X.; Wang, H. Therapeutic Effects of Adipose Stem Cells from Diabetic Mice for the Treatment of Type 2 Diabetes. Molecular Therapy 2018, 26, 1921–1930. [Google Scholar] [CrossRef]
- Kawada-Horitani, E.; Kita, S.; Okita, T.; Nakamura, Y.; Nishida, H.; Honma, Y.; Fukuda, S.; Tsugawa-Shimizu, Y.; Kozawa, J.; Sakaue, T.; et al. Human Adipose-Derived Mesenchymal Stem Cells Prevent Type 1 Diabetes Induced by Immune Checkpoint Blockade. Diabetologia 2022, 65, 1185–1197. [Google Scholar] [CrossRef]
- Xu, N.; Liu, J.; Li, X. Therapeutic Role of Mesenchymal Stem Cells (MSCs) in Diabetic Kidney Disease (DKD). Endocr J 2022, 69, 1159–1172. [Google Scholar] [CrossRef]
- Khatri, R.; Mazurek, S.; Petry, S.F.; Linn, T. Mesenchymal Stem Cells Promote Pancreatic β-Cell Regeneration through Downregulation of FoxO1 Pathway. Stem Cell Res Ther 2020, 11, 497. [Google Scholar] [CrossRef]
- Zhao, H.; He, Z.; Huang, D.; Gao, J.; Gong, Y.; Wu, H.; Xu, A.; Meng, X.; Li, Z. Infusion of Bone Marrow Mesenchymal Stem Cells Attenuates Experimental Severe Acute Pancreatitis in Rats. Stem Cells International 2016, 2016, 7174319. [Google Scholar] [CrossRef]
- Mei, R.; Wan, Z.; Yang, C.; Shen, X.; Wang, R.; Zhang, H.; Yang, R.; Li, J.; Song, Y.; Su, H. Advances and Clinical Challenges of Mesenchymal Stem Cell Therapy. Front. Immunol. 2024, 15, 1421854. [Google Scholar] [CrossRef]
- Levy, O.; Kuai, R.; Siren, E.M.J.; Bhere, D.; Milton, Y.; Nissar, N.; De Biasio, M.; Heinelt, M.; Reeve, B.; Abdi, R.; et al. Shattering Barriers toward Clinically Meaningful MSC Therapies. Sci. Adv. 2020, 6, eaba6884. [Google Scholar] [CrossRef]
- Mou, L.; Wang, T.B.; Wang, X.; Pu, Z. Advancing Diabetes Treatment: The Role of Mesenchymal Stem Cells in Islet Transplantation. Front. Immunol. 2024, 15, 1389134. [Google Scholar] [CrossRef]
- Gamble, A.; Pawlick, R.; Pepper, A.R.; Bruni, A.; Adesida, A.; Senior, P.A.; Korbutt, G.S.; Shapiro, A.M.J. Improved Islet Recovery and Efficacy through Co-Culture and Co-Transplantation of Islets with Human Adipose-Derived Mesenchymal Stem Cells. PLoS ONE 2018, 13, e0206449. [Google Scholar] [CrossRef]
- Koehler, N.; Buhler, L.; Egger, B.; Gonelle-Gispert, C. Multipotent Mesenchymal Stromal Cells Interact and Support Islet of Langerhans Viability and Function. Front. Endocrinol. 2022, 13, 822191. [Google Scholar] [CrossRef]
- Rawal, S.; Williams, S.J.; Ramachandran, K.; Stehno-Bittel, L. Integration of Mesenchymal Stem Cells into Islet Cell Spheroids Improves Long-Term Viability, but Not Islet Function. Islets 2017, 9, 87–98. [Google Scholar] [CrossRef]
- Montanari, E.; Meier, R.P.H.; Mahou, R.; Seebach, J.D.; Wandrey, C.; Gerber-Lemaire, S.; Buhler, L.H.; Gonelle-Gispert, C. Multipotent Mesenchymal Stromal Cells Enhance Insulin Secretion from Human Islets via N-Cadherin Interaction and Prolong Function of Transplanted Encapsulated Islets in Mice. Stem Cell Res Ther 2017, 8, 199. [Google Scholar] [CrossRef]
- De Souza, B.M.; Bouças, A.P.; Oliveira, F.D.S.D.; Reis, K.P.; Ziegelmann, P.; Bauer, A.C.; Crispim, D. Effect of Co-Culture of Mesenchymal Stem/Stromal Cells with Pancreatic Islets on Viability and Function Outcomes: A Systematic Review and Meta-Analysis. Islets 2017, 9, 30–42. [Google Scholar] [CrossRef]
- Giannasi, C.; Niada, S.; Della Morte, E.; Casati, S.; Orioli, M.; Gualerzi, A.; Brini, A.T. Towards Secretome Standardization: Identifying Key Ingredients of MSC-Derived Therapeutic Cocktail. Stem Cells International 2021, 2021, 1–13. [Google Scholar] [CrossRef]
- Chen, S.; Liang, B.; Xu, J. Unveiling Heterogeneity in MSCs: Exploring Marker-Based Strategies for Defining MSC Subpopulations. J Transl Med 2024, 22, 459. [Google Scholar] [CrossRef]
- Chouaib, B.; Haack-Sørensen, M.; Chaubron, F.; Cuisinier, F.; Collart-Dutilleul, P.-Y. Towards the Standardization of Mesenchymal Stem Cell Secretome-Derived Product Manufacturing for Tissue Regeneration. IJMS 2023, 24, 12594. [Google Scholar] [CrossRef]
- Zhang, L.; Mack, R.; Breslin, P.; Zhang, J. Molecular and Cellular Mechanisms of Aging in Hematopoietic Stem Cells and Their Niches. J Hematol Oncol 2020, 13, 157. [Google Scholar] [CrossRef]
- Preda, M.B.; Neculachi, C.A.; Fenyo, I.M.; Vacaru, A.-M.; Publik, M.A.; Simionescu, M.; Burlacu, A. Short Lifespan of Syngeneic Transplanted MSC Is a Consequence of in Vivo Apoptosis and Immune Cell Recruitment in Mice. Cell Death Dis 2021, 12, 566. [Google Scholar] [CrossRef]
- Wang, P.; Zeng, F.; He, L.; Wang, J.; Zhang, T.; Zhang, D. Alteration of the Immune Status of Umbilical Cord Mesenchymal Stem Cells Stimulated by TLR1/2 Agonist, Pam3Csk. Molecular Medicine Reports 2016, 14, 2206–2212. [Google Scholar] [CrossRef]
- Click-ITTM Plus TUNEL Assay Kit for In Situ Apoptosis Detection Available Online:. Available online: Https://Assets.Fishersci.Com/TFS-Assets/BID/Manuals/MAN0010877_Click_iT_Plus_TUNEL_Assay_UG.Pdf (accessed on day month year).
- Deskins, D.L.; Bastakoty, D.; Saraswati, S.; Shinar, A.; Holt, G.E.; Young, P.P. Human Mesenchymal Stromal Cells: Identifying Assays to Predict Potency for Therapeutic Selection. Stem Cells Translational Medicine 2013, 2, 151–158. [Google Scholar] [CrossRef]
- Qin, L.; Liu, N.; Bao, C.; Yang, D.; Ma, G.; Yi, W.; Xiao, G.; Cao, H. Mesenchymal Stem Cells in Fibrotic Diseases—the Two Sides of the Same Coin. Acta Pharmacol Sin 2023, 44, 268–287. [Google Scholar] [CrossRef]
- Cheshmi, B.; Cheshomi, H. Salivary Exosomes: Properties, Medical Applications, and Isolation Methods. Mol Biol Rep 2020, 47, 6295–6307. [Google Scholar] [CrossRef]
- Liu, A.; Hefley, B.; Escandon, P.; Nicholas, S.E.; Karamichos, D. Salivary Exosomes in Health and Disease: Future Prospects in the Eye. IJMS 2023, 24, 6363. [Google Scholar] [CrossRef]
- Reseco, L.; Molina-Crespo, A.; Atienza, M.; Gonzalez, E.; Falcon-Perez, J.M.; Cantero, J.L. Characterization of Extracellular Vesicles from Human Saliva: Effects of Age and Isolation Techniques. Cells 2024, 13, 95. [Google Scholar] [CrossRef]
- Sequeira, S.J.; Larsen, M.; DeVine, T. Extracellular Matrix and Growth Factors in Salivary Gland Development. In Frontiers of Oral Biology; Tucker, A.S., Miletich, I., Eds.; S. Karger AG, 2010; Vol. 14, pp. 48–77 ISBN 978-3-8055-9406-6.
- Pellegrini, M.; Pulicari, F.; Zampetti, P.; Scribante, A.; Spadari, F. Current Salivary Glands Biopsy Techniques: A Comprehensive Review. Healthcare 2022, 10, 1537. [Google Scholar] [CrossRef]
- Diaferia, G.R.; Jimenez-Caliani, A.J.; Ranjitkar, P.; Yang, W.; Hardiman, G.; Rhodes, C.J.; Crisa, L.; Cirulli, V. Β1 Integrin Is a Crucial Regulator of Pancreatic β-Cell Expansion. Development 2013, 140, 3360–3372. [Google Scholar] [CrossRef]
- Li, W. Targeting Ceramides and Adiponectin Receptors in the Islet of Langerhans for Treating Diabetes. Molecules 2022, 27, 6117. [Google Scholar] [CrossRef]
- Gilani, A.; Stoll, L.; Homan, E.A.; Lo, J.C. Adipose Signals Regulating Distal Organ Health and Disease. Diabetes 2024, 73, 169–177. [Google Scholar] [CrossRef]
- Cortes, G.A.; Moore, M.J.; El-Nakeep, S. Physiology, Von Willebrand Factor. In StatPearls; StatPearls Publishing: Treasure Island (FL), 2025. [Google Scholar]
- Valdivia, A.; Avalos, A.M.; Leyton, L. Thy-1 (CD90)-Regulated Cell Adhesion and Migration of Mesenchymal Cells: Insights into Adhesomes, Mechanical Forces, and Signaling Pathways. Front. Cell Dev. Biol. 2023, 11, 1221306. [Google Scholar] [CrossRef]
- Zhu, H.; Mitsuhashi, N.; Klein, A.; Barsky, L.W.; Weinberg, K.; Barr, M.L.; Demetriou, A.; Wu, G.D. The Role of the Hyaluronan Receptor CD44 in Mesenchymal Stem Cell Migration in the Extracellular Matrix. Stem Cells 2006, 24, 928–935. [Google Scholar] [CrossRef]
- Alborzian Deh Sheikh, A.; Akatsu, C.; Abdu-Allah, H.H.M.; Suganuma, Y.; Imamura, A.; Ando, H.; Takematsu, H.; Ishida, H.; Tsubata, T. The Protein Tyrosine Phosphatase SHP-1 (PTPN6) but Not CD45 (PTPRC) Is Essential for the Ligand-Mediated Regulation of CD22 in BCR-Ligated B Cells. The Journal of Immunology 2021, 206, 2544–2551. [Google Scholar] [CrossRef]
- Sionov, R.V.; Ahdut-HaCohen, R. A Supportive Role of Mesenchymal Stem Cells on Insulin-Producing Langerhans Islets with a Specific Emphasis on The Secretome. Biomedicines 2023, 11, 2558. [Google Scholar] [CrossRef]
Figure 3.
Volcano plots of protein-coding gene expression. a) Volcano plot: mMSCs vs islets; b) Volcano plot: mMSCs vs Exo_Pan; c) Volcano plot: mSGCs vs islets; d) Volcano plot in mSGC vs Exo_Pan.
Figure 3.
Volcano plots of protein-coding gene expression. a) Volcano plot: mMSCs vs islets; b) Volcano plot: mMSCs vs Exo_Pan; c) Volcano plot: mSGCs vs islets; d) Volcano plot in mSGC vs Exo_Pan.
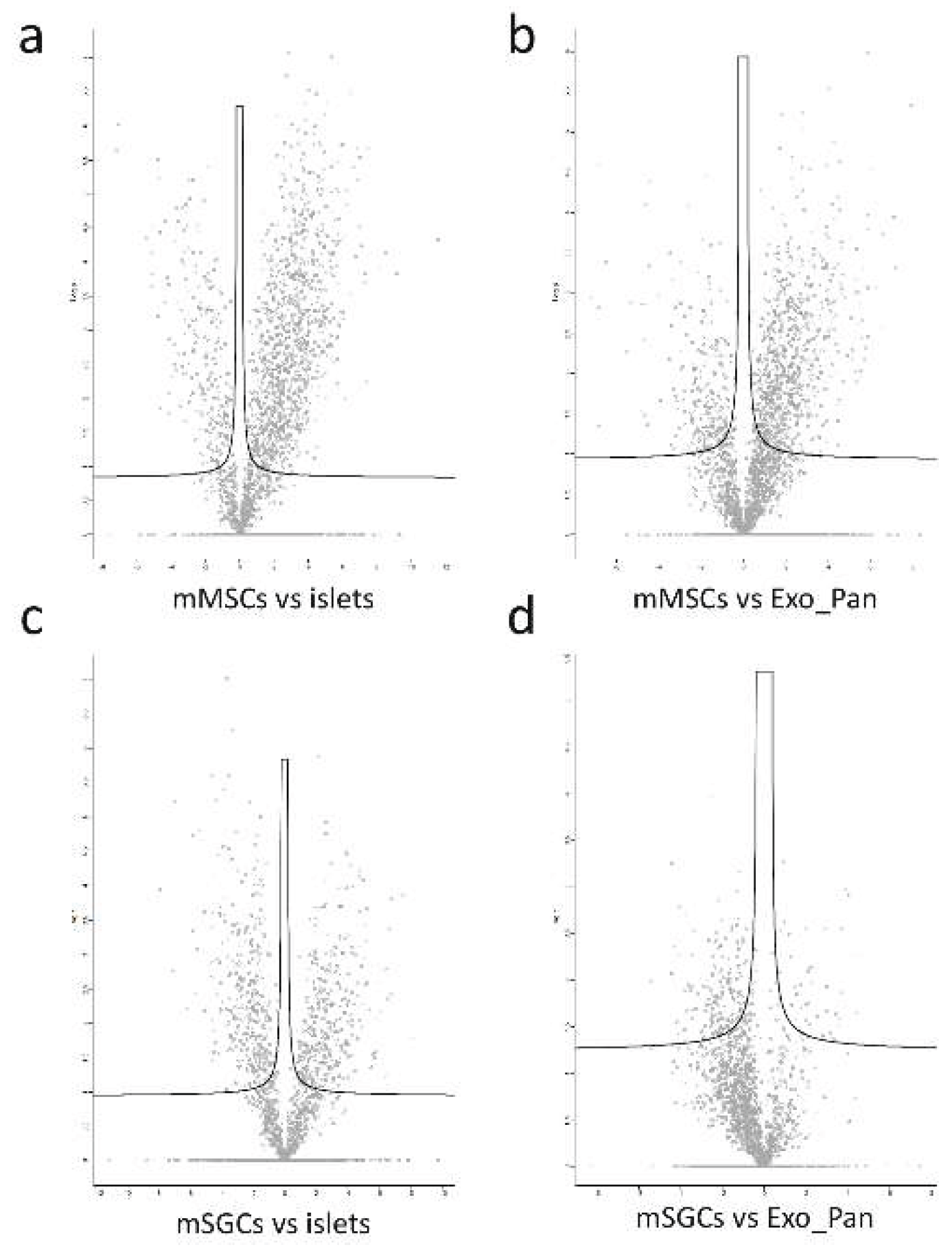
Figure 4.
Heat maps demonstrating the contribution of all received signals to the investigated cell groups. a) Outgoing signaling patterns; b) Incoming signaling patterns of cell types.
Figure 4.
Heat maps demonstrating the contribution of all received signals to the investigated cell groups. a) Outgoing signaling patterns; b) Incoming signaling patterns of cell types.
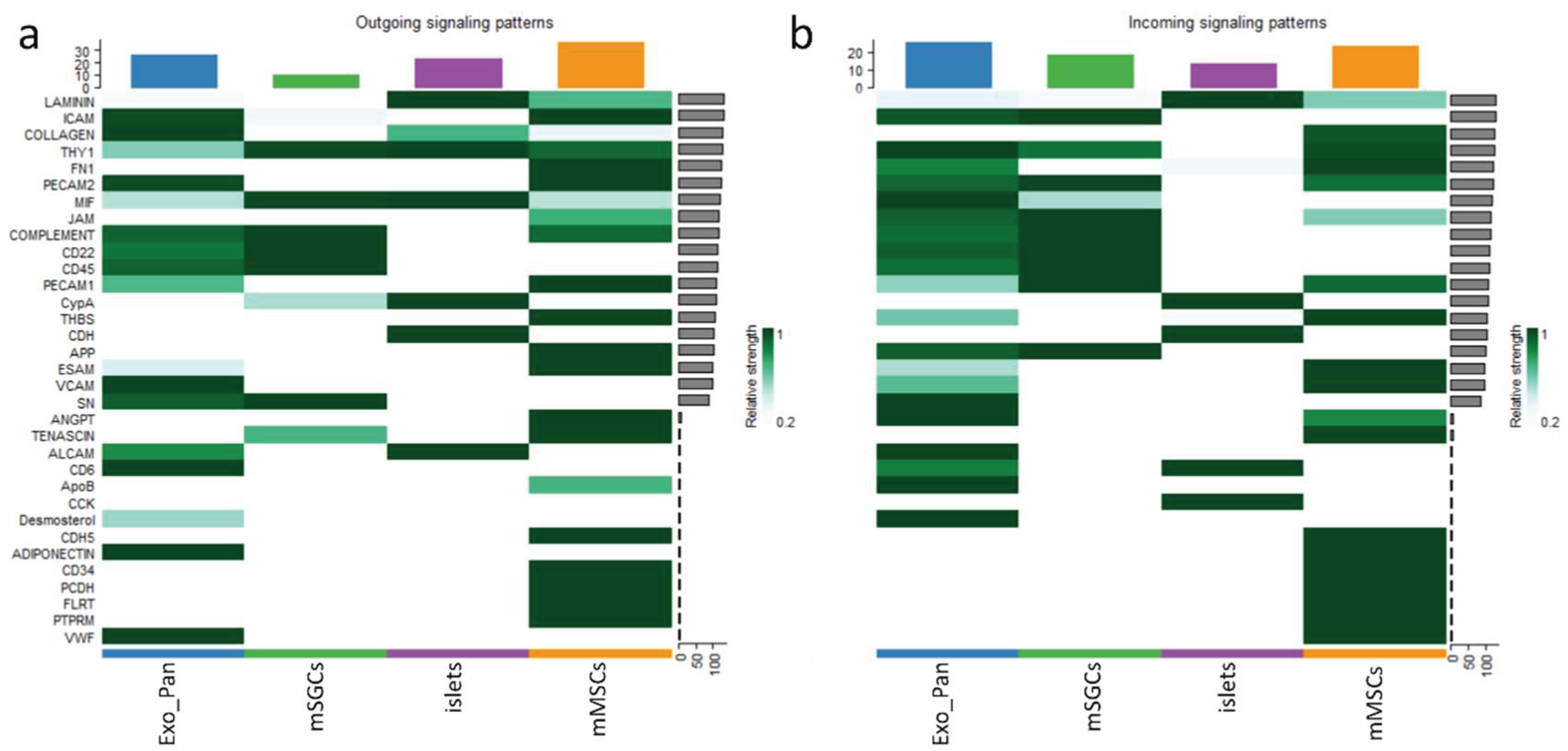
Figure 5.
Heatmaps demonstrating the four global cell-to-cell communication patterns calculated from major signals for the subpopulations studied: (a) Outgoing_communication_patterns; (b) Incoming_communication_patterns.
Figure 5.
Heatmaps demonstrating the four global cell-to-cell communication patterns calculated from major signals for the subpopulations studied: (a) Outgoing_communication_patterns; (b) Incoming_communication_patterns.
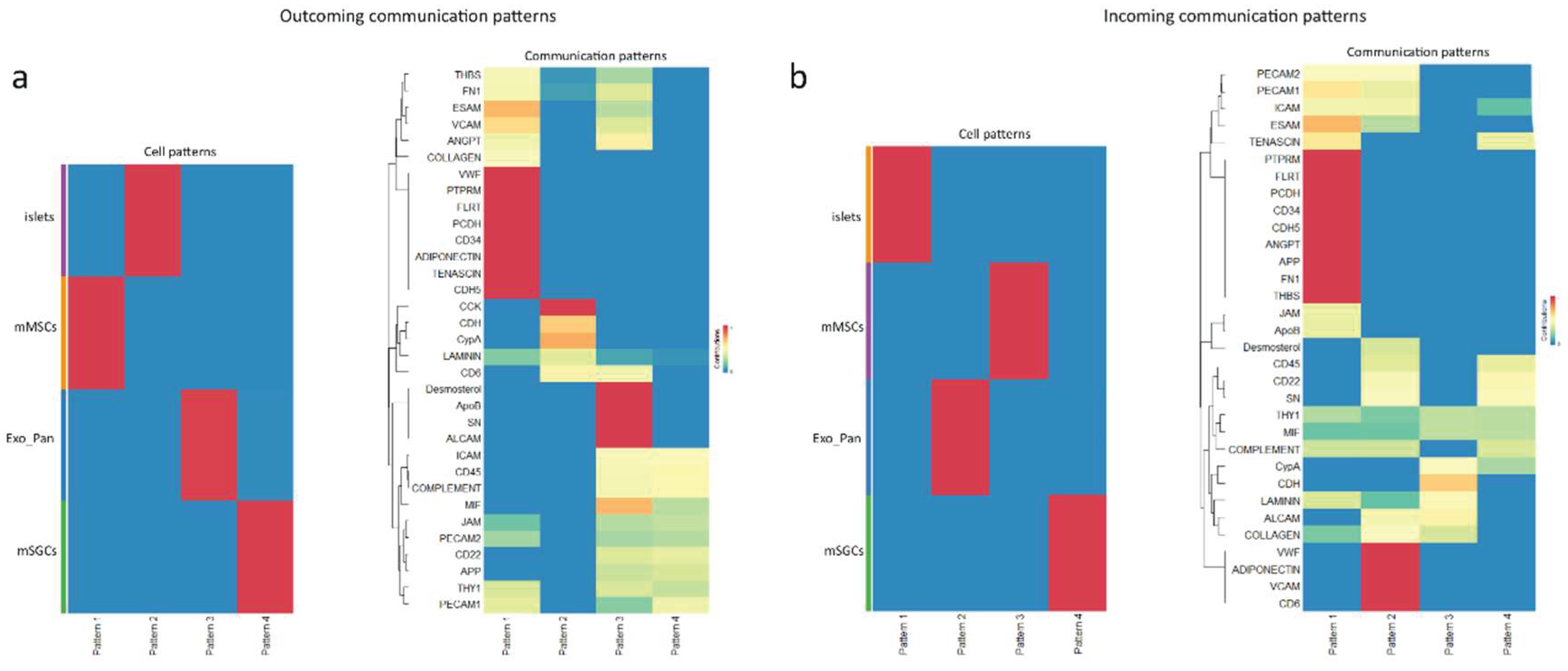
Figure 6.
Chord diagram illustrating the main communication pathways used by the different cell types to interact. The connections start from the ligand islets and Exo_Pan and end with the mMSC receptor and mSGCs, which are grouped for each cell type (colored outer arcs).
Figure 6.
Chord diagram illustrating the main communication pathways used by the different cell types to interact. The connections start from the ligand islets and Exo_Pan and end with the mMSC receptor and mSGCs, which are grouped for each cell type (colored outer arcs).
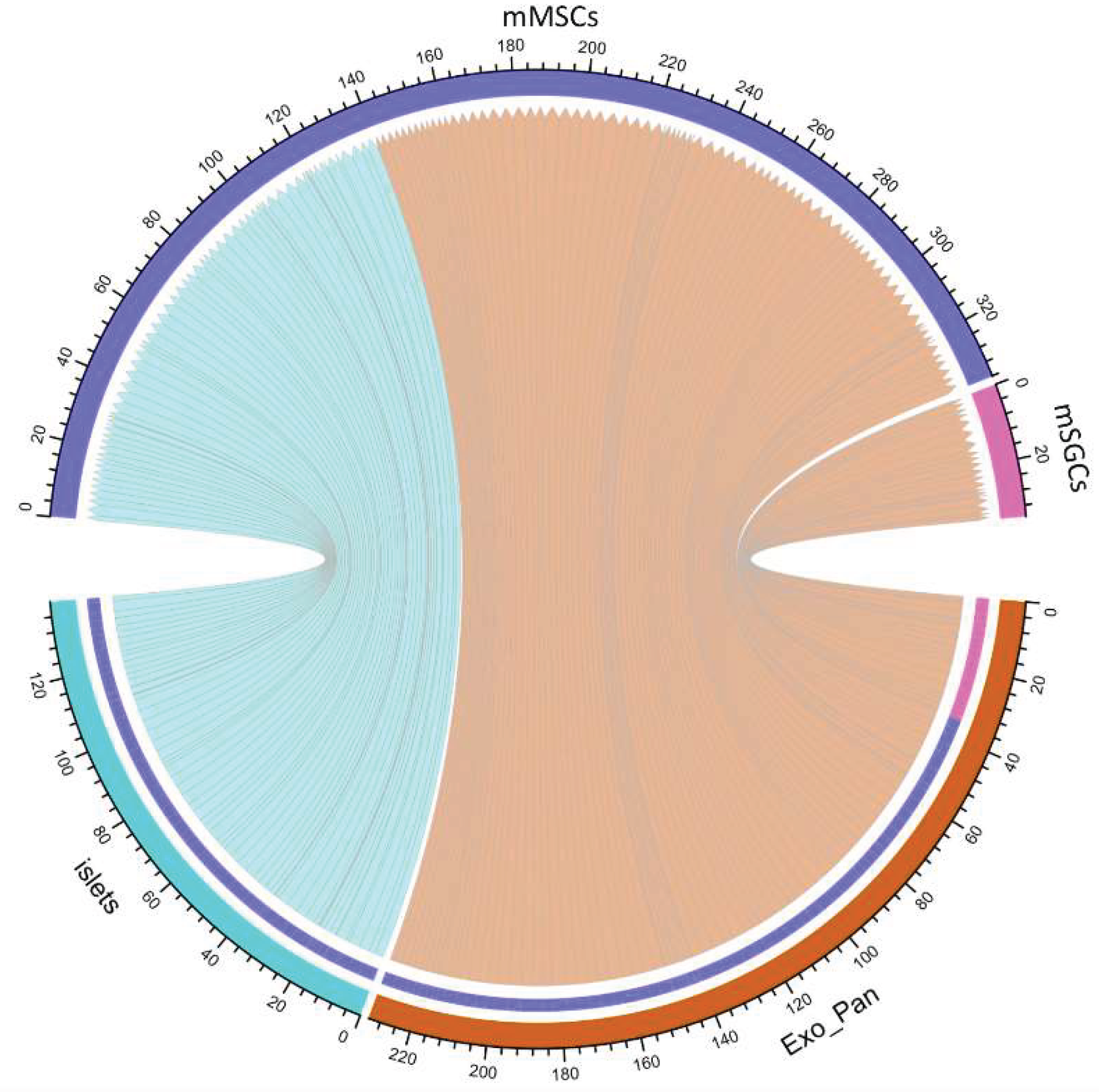
Figure 7.
Results of protein function analysis of proteins found in mMSCs and analysis of ligand-receptor pair interactions between mMSC receptor proteins and pancreatic cell ligand proteins. a) GSEA results of mMSC receptor proteins in the mMSC + islets comparison group. Analyses were performed using the Gene Ontology: Biological Process (GO:BP) database. -log (p-value) is the negative logarithm of the significance level; (b) GSEA results of mMSC receptor proteins in the mMSC + Exo_Pan comparison group. Analyses were performed using the Gene Ontology: Biological Process (GO:BP) database. -log (p-value) is the negative logarithm of the significance level; (c) Bubble diagram showing the relationship between ligands and receptors detected in the analysis of mMSC + islets and mMSC + Exo_Pan pairs. (The color of the dots indicates the probability of interaction (Commun. Prob.), where lighter tones indicate low probability and more saturated tones indicate high probability. The size of the dots corresponds to the significance level (p-value) of the interaction: Small dots (pale) indicate p > 0.05 (not significant). Medium-sized dots (blank) indicate 0.01 < p < 0.05 (significant). Large dots (black) indicate p < 0.01 (high significance)).
Figure 7.
Results of protein function analysis of proteins found in mMSCs and analysis of ligand-receptor pair interactions between mMSC receptor proteins and pancreatic cell ligand proteins. a) GSEA results of mMSC receptor proteins in the mMSC + islets comparison group. Analyses were performed using the Gene Ontology: Biological Process (GO:BP) database. -log (p-value) is the negative logarithm of the significance level; (b) GSEA results of mMSC receptor proteins in the mMSC + Exo_Pan comparison group. Analyses were performed using the Gene Ontology: Biological Process (GO:BP) database. -log (p-value) is the negative logarithm of the significance level; (c) Bubble diagram showing the relationship between ligands and receptors detected in the analysis of mMSC + islets and mMSC + Exo_Pan pairs. (The color of the dots indicates the probability of interaction (Commun. Prob.), where lighter tones indicate low probability and more saturated tones indicate high probability. The size of the dots corresponds to the significance level (p-value) of the interaction: Small dots (pale) indicate p > 0.05 (not significant). Medium-sized dots (blank) indicate 0.01 < p < 0.05 (significant). Large dots (black) indicate p < 0.01 (high significance)).
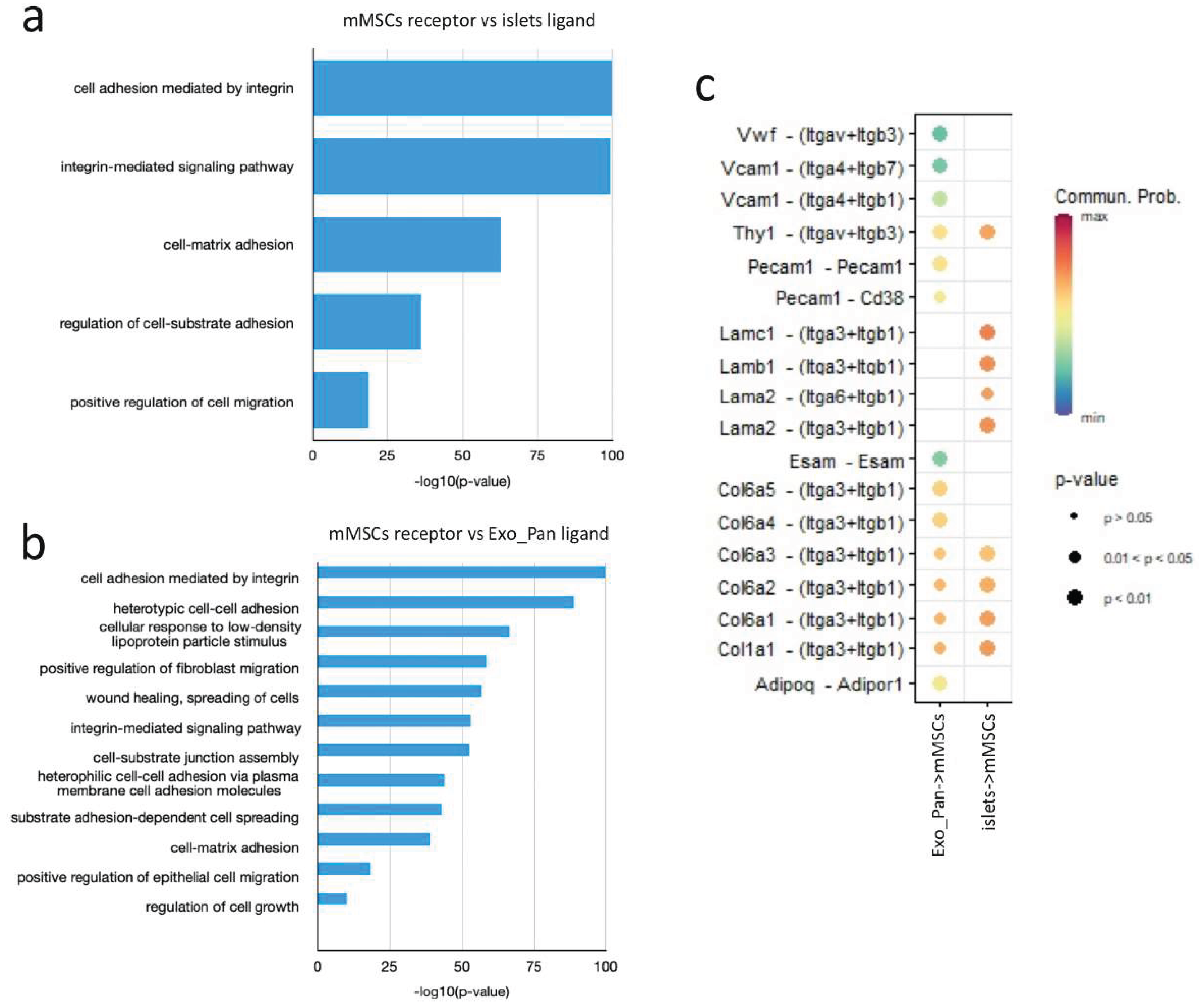
Figure 8.
The results of the analysis of mSGC receptor protein functions and the analysis of ligand-receptor pair interactions between mSGC receptor proteins and pancreatic cell ligand proteins. a) List of functions of mSGC receptor proteins in the mSGC + Exo_Pan comparison group. Analyses were performed using the Gene Ontology: Biological Process (GO:BP) database. -log (p-value) is the negative logarithm of the significance level; (b) Bubble diagram showing the relationship between ligands and receptors detected in the analysis of mSGC + islets and mSGC + Exo_Pan pairs.
Figure 8.
The results of the analysis of mSGC receptor protein functions and the analysis of ligand-receptor pair interactions between mSGC receptor proteins and pancreatic cell ligand proteins. a) List of functions of mSGC receptor proteins in the mSGC + Exo_Pan comparison group. Analyses were performed using the Gene Ontology: Biological Process (GO:BP) database. -log (p-value) is the negative logarithm of the significance level; (b) Bubble diagram showing the relationship between ligands and receptors detected in the analysis of mSGC + islets and mSGC + Exo_Pan pairs.
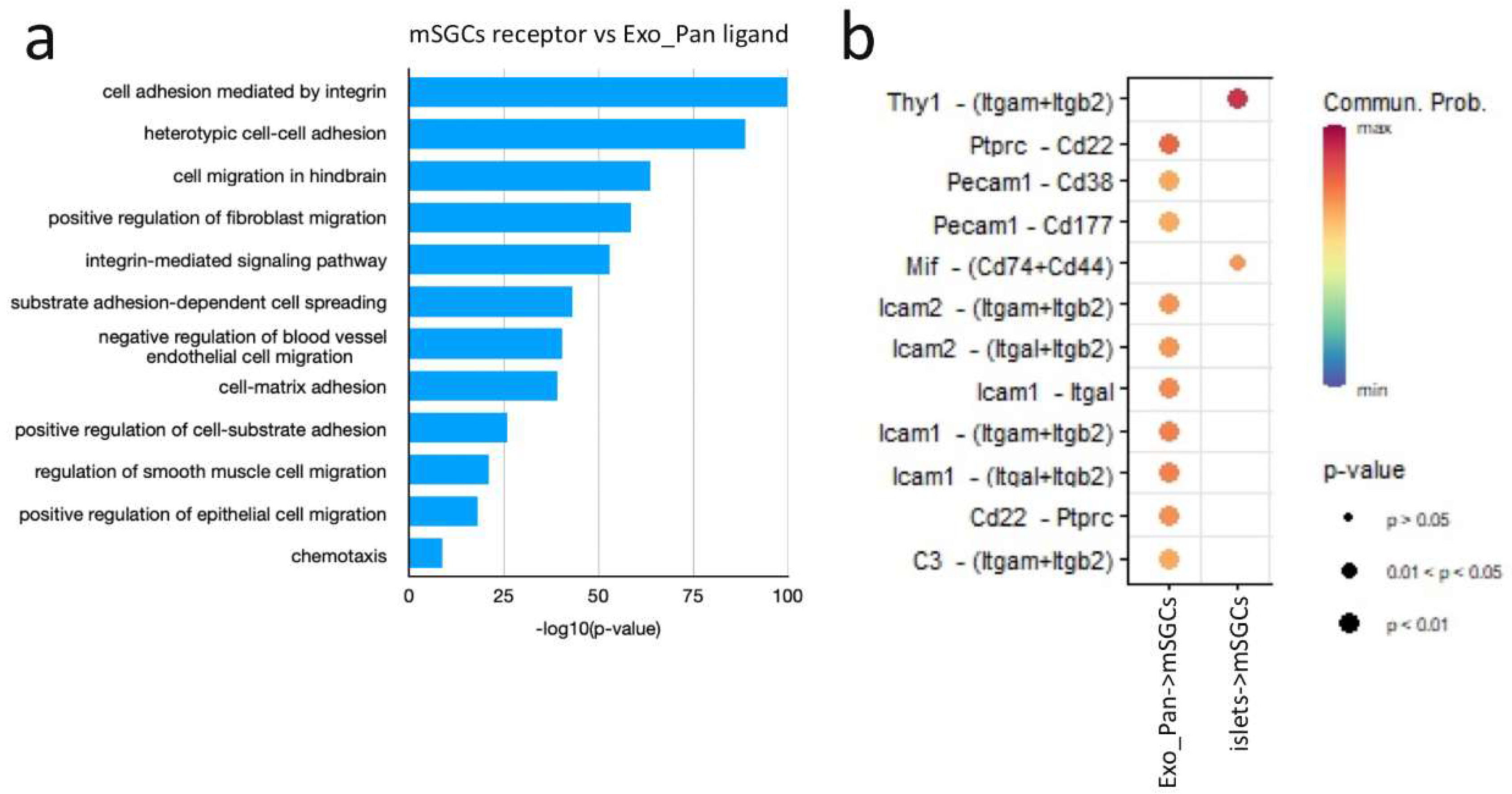
Figure 9.
Heat map showing the activity of the researched cell types (Exo_Pan, mSGC, islets, mMSC) in the signaling pathways: Collagen, FN1, ICAM, JAM, Laminin, Mif, PECAM1, and THY1. Each bar indicates how much a particular cell type is involved in the network. The dark red color indicates the greater involvement of a given cell type in a certain signaling pathway.
Figure 9.
Heat map showing the activity of the researched cell types (Exo_Pan, mSGC, islets, mMSC) in the signaling pathways: Collagen, FN1, ICAM, JAM, Laminin, Mif, PECAM1, and THY1. Each bar indicates how much a particular cell type is involved in the network. The dark red color indicates the greater involvement of a given cell type in a certain signaling pathway.
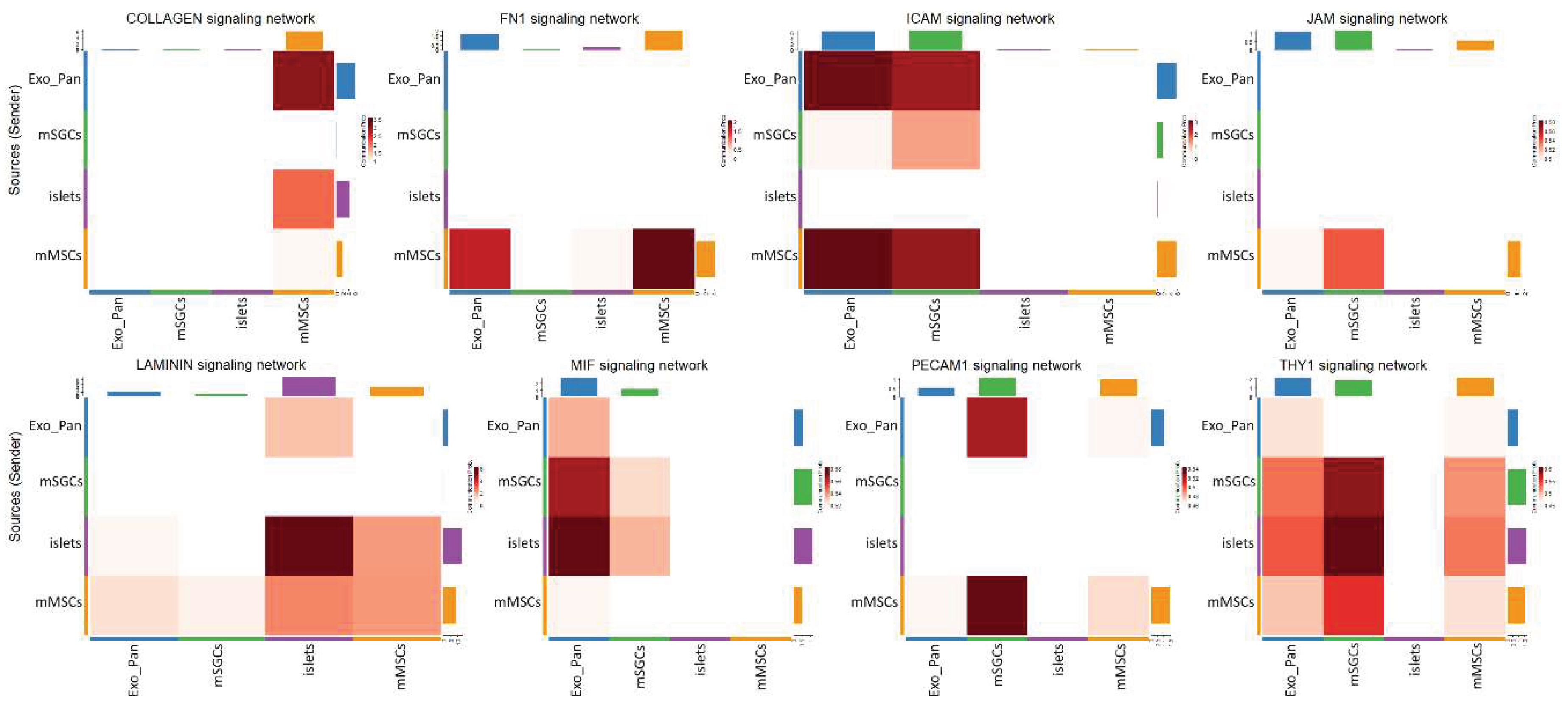
Figure 10.
Heat map showing the prevailing roles of intercellular communication (Sender, Receiver, Mediator, and Influencer) of the analyzed cell types (Exo_Pan, mSGCs, islets, and mMSCs) in the signaling pathways: Collagen, FN1, ICAM, JAM, Laminin, Mif, PECAM1, and THY1. Each bar indicates how much a cell type is involved in a particular role. The dark green color indicates the higher importance of that cell type in a particular role in the signaling pathway examined.

Table 1.
The table shows the dominant roles and cell-cell interactions for mMSCs and mSGCs in Collagen, FN1, ICAM, ICAM, JAM, Laminin, Mif, PECAM1, and THY1 signaling pathways. mMSC+Exo_Pan/islets and mSGC+Exo_Pan/islets interactions are considered.
Table 1.
The table shows the dominant roles and cell-cell interactions for mMSCs and mSGCs in Collagen, FN1, ICAM, ICAM, JAM, Laminin, Mif, PECAM1, and THY1 signaling pathways. mMSC+Exo_Pan/islets and mSGC+Exo_Pan/islets interactions are considered.
| Signaling pathway | Dominant role of mMSCs (Importance) |
Dominant role of mSGCs (Importance) |
Dominant interaction of mMSCs (Communication. Prob.) |
Dominant interaction of mSGCs (Communication. Prob.) |
|---|---|---|---|---|
| COLLAGEN | Mediator (100%) | Influencer (30%) | mMSC+Exo_Pan (3.25) | - |
| FN1 | Sender (90%) | Influencer (30%) | mMSC+Exo_Pan (1.5) | - |
| ICAM | Influencer (100%) | Receiver (90%) | mMSC+Exo_Pan (3) | mSGC+Exo_Pan (2.5) |
| JAM | Influencer (80%) | Receiver (70%) | mMSC+Exo_Pan (0.5) | - |
| Laminin | Influencer (70%) | Influencer (40%) | mMSC+islets (3) | - |
| MIF | Influencer (50%) | Mediator (80%) | mMSC+Exo_Pan (0.52) | mSGC+islets (0.54) |
| PECAM1 | Sender (80%) | Receiver (90%) | mMSC+Exo_Pan (0.46) | mSGC+islets (0.52) |
| THY1 | Mediator (90%) | Influencer (70%) | mMSC+islets (0.55) | mSGC+islets (0.6) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



