
Submitted:
21 May 2025
Posted:
21 May 2025
You are already at the latest version
Abstract
Annexins (ANNs), which are calcium (Ca2+)-dependent and phospholipid-binding protein families, are implicated in the regulation of plant growth and development as well as protection from various stresses. However, little is known about ANNs in Corydalis Saxicola, an endangered benzylisoquinoline alkaloid (BIA)-riched herbaceous plant widely used in traditional Chinese medicine and endemic to the calciphilic karst region in China. Here, nine CsANN genes were identified from C. saxicola, and they were divided into three subfamilies based on the phylogenetic analysis. The CsANNs clustered into the same clade, sharing similar gene structures and conserved motifs. The nine CsANN genes were located on five chromosomes, and their expansions were mainly attributed to tandem duplication and segmental duplication. The CsANN transcripts showed variable organ-specific and Ca2+-responsive expression patterns. Further transient overexpression assays showed that CsANN1 could positively regulate the accumulation of BIA compounds in C. saxicola leaves, probably through directly interacting with key BIA-biosynthesis-pathway enzymes or by interacting with BIA-biosynthesis-regulating factors, such as MYBs. This study sheds light on profiles and functions of the CsANN gene family and paves the way for unraveling the molecular mechanism of BIA accumulation regulated by Ca2+ through CsANNs.
Keywords:
annexin
; Corydalis saxicola
; calcium
; benzylisoquinoline alkaloid
1. Introduction
Corydalis saxicola Bunting, which belongs to the genus Corydalis within the Papaveraceae family, is an endangered herbaceous plant widely used in traditional Chinese medicine due to its various pharmacological activities, including anti-cancer, anti-inflammatory, antibacterial, antioxidant, and analgesic properties [1,2,3]. These pharmacological activities of Yanhuanglian (the Chinese name of C. saxicola) are primarily attributed to the presence of benzylisoquinoline alkaloid (BIA) compounds, with dehydrocavidine (DHCA) identified as one of the key active ingredients [4,5]. As a species endemic to the karst region in China, C. saxicola lives exclusively in or around rock crevices and demonstrates good adaptability to the arid, infertile, and calcium-rich soils, which are the typical karst environments [6]. However, both its narrow, stringent karst-adapted habitat and the imbalance of large market demand make it endangered [6,7,8]. Therefore, understanding the adaptation mechanism of C. saxicola in calcium-rich karst habitats, as well as the regulatory molecular mechanism of the key active BIAs, such as DHCA, in C. saxicola, are considered as two of the most efficient and promising approaches for the conservation and utilization of this endangered medicinal plant.
Annexins (ANNs) are evolutionarily conserved, water-soluble proteins [9]. The plant ANNs, which can bind to or insert into cell membranes and regulate the homeostasis of free Ca2+ in the cytoplasm, are the members of voltage-gated Ca2+ channels [10]. In addition, it has peroxidase as well as ATPase/GTPase activities, which give ANNs functional specificity [11,12]. The core structures of plant ANNs are characterized by a N-terminal region and a C-terminal region [12]. The short N-terminal region comprises approximately a dozen amino acids and is commonly as the site for secondary modifications, including phosphorylation, nitrosylation, S-glutathionylation, and N-myristoylation, which are regulated by various signaling pathways [9,13,14]. Biochemical experiments have demonstrated that the N-terminal domain modulates different ANN properties through its allosteric mechanism [9,15]. The C-terminal region of plant ANNs contains four conservative ANN repeats, each repeat containing about 70 amino acid residues and then folding into five α helices [16]. Specifically, both of the repeats I and IV include a motif of GXGT-(38 variable amino acid residues)-D/E, which was considered as a Ca2+ binding site [17]. However, compared to that in the repeat I, this motif is less conserved in the repeat IV [18]. Recently, the ANN families have been identified in many plant species, including Carica papaya, Glycine max, Oryza sativa, Raphanus sativus, Vitis vinifera, Zea mays, and so on [19,20,21,22,23]. These ANNs are ubiquitously distributed across various plant tissues, including embryos, seedlings, roots and tubers, stems, hypocotyls and cotyledons, leaves, inflorescences, fruits, vascular systems, and phloem sap [24,25,26,27,28], demonstrating their wide involvements in regulating of biochemical and cellular processes, plant growth and development, and response to biotic and abiotic stresses [9,10,11,14].
Plants in karst areas generally have the tolerance to drought and calciphilic characteristics [29,30]. Our previous studies found that the cultivated C. saxicola plants had the ability to endure the treatment of high concentrations of exogenous CaCl2 solutions for nearly a month at least, and the contents of DHCA in the roots of C. saxicola were significantly enhanced after the treatments. Nevertheless, how extracellular Ca2+ signaling was recognized and converted into intracellular ones to regulate the biosynthesis of DHCA in C. saxicola is largely unknown. As plant ANNs are members of voltage-gated Ca2+ channels and are involved in Ca2+ uptake and transport, we speculated that ANNs in C. saxicola (CsANNs) might be one of the key executors during this process.
In this study, a global survey of the nine ANN genes in the C. saxicola genome was conducted. The physicochemical properties, conserved motifs, gene structures, and cis-elements in the promoter regions of CsANNs were identified. In addition, the homologous ANN genes from different species were determined based on a collinearity analysis. Furthermore, the expression profiles of all nine CsANNs in different tissues and under CaCl2 treatments were analyzed. The transient overexpression assays demonstrated that CsANN1 could induce the accumulation of DHCA in the leaves of C. saxicola. In summary, the present study might provide substantial useful information for the understanding of CsANNs as the key effectors during Ca2+ uptake and transport, and the role played in the biosynthesis of DHCA in C. saxicola.
2. Results
2.1. Identification and Physicochemical Analysis of CsANNs
A total of nine candidate ANNs were identified in the C. saxicola genome. All these CsANNs contained conserved ANN domains, indicating that these peptides were putative ANNs. Subsequently, all nine CsANN genes were successfully cloned by PCR and verified by Sanger sequencing. Consequently, these CsANNs were named as CsANN1-9 according to their chromosomal localization (Table S1).
Physicochemical property analysis results showed that the CsANN proteins contained 316 to 341 amino acids, with molecular weights (MWs) ranging from 35.48 kDa to 38.67 kDa (Table S1). The hydrophilicities of the nine CsANN proteins were all below 0, indicating their hydrophilic nature. The isoelectric points (pIs) of CsANN1, 5, 6, 7, and 9 were all below 7, indicating that they were acidic hydrophilic proteins; the others were basic hydrophilic proteins. The protein instability indices of CsANN1, 3, 4, 5, and 7 were below 40, and these are stable proteins; the others were unstable proteins. The results of subcellular localization of CsANNs predicted by different software were variable, but most of them localized in the cytosol (Figure 1A).
2.2. Sequences Alignment and Phylogenetic Analysis of ANNs
The amino acid sequence alignment results indicated that CsANN proteins contained four conserved ANN repeats, each of which consisted of approximately 70 amino acids and was similar to that of AtANN1 (Figure 1B). Particularly, the specific Ca2+-binding sites (G/KXGT-38-D/E), which could functionally bind to phospholipid membranes, were characterized in the repeat I of CsANN1, 5, 7, and 9 (Figure 1B). In addition, a conserved peroxidase residue (ILAHR) was identified in CsANN1, 5, and 9, indicating that they were likely to have peroxidase activity. Furthermore, the IRI site, which could bind to F-actin, existed in the repeat III of CsANN1 and 9, and the DXXG site, which could bind to GTP, was found in the repeat IV of CsANN1, 5, and 9 (Figure 1B).
To elucidate the evolutionary patterns and potential functions of CsANNs, we used the neighbor-joining (NJ) method to construct a phylogenetic tree based on 111 ANN proteins from nine species. As shown in Figure 2, all the ANNs could be divided into four subfamilies according to the phylogenetic tree analysis. CsANNs were distributed among all subfamilies except subfamily III, and their distribution pattern was similar with other ANNs except two ones of C. yanhusuo (CyANN16 and CyANN24) (Figure 2). Among all nine CsANNs, CsANN1, 7, and 9 were classified within subfamily I, CsANN2, 3, 4, 6, and 8 were assigned to subfamily IV, and only CsANN5 was categorized under subfamily II (Figure 2). Typically, CsANNs were initially grouped with CtANNs and CyANNs, followed by PtANNs and AtANNs within each clade, suggesting a closer evolutionary relationship among CtANNs, CyANNs, and CsANNs (Figure 2). Conversely, the evolutionary relationships among ANN proteins from C. saxicola, Hordeum vulgare, and O. sativa were found to be the farthest (Figure 2). These findings indicated that CsANNs were more closely related to those of dicotyledons than those of monocotyledons, which was consistent with previous studies [18,19,21,22].
2.3. Conserved Motifs and Gene Structure of CsANNs
The gene structure characteristics of CsANNs, and the conserved motifs and domain compositions of CsANN proteins were analyzed and shown according to their phylogenetic relationships (Figure 3A). Four or five motifs were characterized in CsANNs, and motif 1~4 were commonly identified in all CsANNs (Figure 3B). CsANN2 and CsANN8, two members of Subfamily Ⅳ, lack motif 5, potentially causing functional loss (Figure 3B). Additionally, all CsANNs have four ANN domains except CsANN2 and CsANN8, which lack one and two ANN domains, respectively (Figure 3C). The exon-intron structural analysis showed that the number of exons within the CsANN genes varied between 4 and 7 (Figure 3D). Notably, CsANN3 contained 7 exons, and CsANN8 contained 4 exons (Figure 3D). It should be mentioned that among all CsANNs, CsANN1 possesses the longest introns (Figure 3D). Furthermore, CsANNs within the same evolutionary branches, specifically subfamily I (CsANN1 and CsANN9) and subfamily IV (CsANN4 and CsANN6), exhibited similar intron/exon patterns (Figure 3D).
2.4. Chromosomal Location and Collinearity Analysis of CsANNs
A chromosomal location map was constructed to investigate the genetic divergence and duplication within the CsANNs. Nine CsANN genes were identified and localized on five chromosomes. The distribution of CsANNs on each chromosome appeared relatively independent and irregular. Among these chromosomes, chromosome 8 contained the highest number of CsANNs, with three identified, followed by chromosomes 6 and 7, each with two CsANNs. Chromosomes 1 and 2 each contained a single CsANN gene (Figure 4A).
To further elucidate the expansion mechanism of the CsANN gene family, we performed collinearity and duplication analyses using TBtools [31]. Intraspecific collinearity analysis revealed the presence of one pair of collinear genes in C. saxicola (CsANN1-CsANN9), both were attributed to segmental duplication, and CsANN3, 4, 5, and 6 came from tandem repetition (Figure 4B). These indicate that segmental duplication events and tandem repetition events were pivotal for the expansion of CsANNs during evolution.
To explore the underlying evolutionary mechanisms of CsANNs, we selected five representative angiosperm species, including A. thaliana, C. tomentella, H. vulgare, O. sativa, and P. tremula, to construct collinearity analysis maps with C. saxicola. The analysis revealed that the CsANNs exhibited the highest syntenic relationships with ANNs in P. tremula (10), followed by C. tomentella (8), A. thaliana (5), H. vulgare (4), and O. sativa (3) (Figure 5). Those results indicated that the ANN genes from C. saxicola, P. tremula and C. tomentella had a close relationship. In addition, a greater number of ANN homologous genes were identified in dicotyledons than those in monocotyledons (Figure 5).
2.5. Analysis of Cis-Acting Elements in CsANN Promoters
To further investigate the response of CsANNs to stresses, we used PlantCARE to extract cis-acting elements involved in plant hormone regulation and abiotic stress response from 2.0 kb 5′ upstream regions of CsANNs [32]. As shown in Figure 6, five main categories of cis-acting elements were predicted, including oxidative stress- (ARE-motif), cold stress- (LTR-motif), drought stress- (MBS-motif), light- (Box 4, GATA-motif, G-Box, G-Box, GT1-motif, TCT-motif), and plant hormone- (abscisic acid response element, ABRE; methyl jasmonate response element, CGTCA-motif, TGACG-motif; gibberellin response element, GARE-motif, P-box; auxin response element, TGA-element) responsive elements. All CsANNs, except CsANN4, contain light-responsive elements. Cis-acting elements associated with hormonal responses were also found in all CsANN promoters, except CsANN1. CsANN1, 7, and 4 contain elements related to drought response. CsANN4, 7, and 9 contain elements related to low temperature response. Only CsANN7 promoters have been identified as a role involved in the antioxidant response. In addition, the GTGGC-motif was only present in CsANN9. The TGACG-motif, which was suggested to be involved in the methyl jasmonate response, appeared only in CsANN3. The P-box, one of the gibberellin response elements, was present in the promoters of CsANN4 and CsANN9. The GARE-motif was present in the promoters of CsANN8 and CsANN9. It should be mentioned that all CsANNs had MYB transcription factor binding sites (Table S2). All the observations indicated that CsANN genes might be involved in environmental stress response and hormone regulation, and play essential roles in physiological and developmental processes of C. saxicola.
2.6. Tissue-Specific Expression Profile of CsANNs
The quantitative real time PCR (qRT-PCR) analysis results indicated that the expression patterns of the CsANNs were variable in different tissues of C. saxicola. As shown in Figure 7, the CsANN2, CsANN3, CsANN4, and CsANN8 were greatly expressed in flowers, suggesting their potentially significant biological role in flowering phase transition or flowering organ development. CsANN5 and CsANN6 were particularly highly expressed in the stems, indicating that they might be involved in the development of the stems of C. saxicola. We also observed that some CsANN genes (CsANN2, CsANN7, and CsANN8) displayed high expression in fruit pods of C. saxicola. Interestingly, CsANN1, CsANN3, CsANN4, and CsANN9 showed relatively high accumulation in the roots of C. saxicola.
2.7. The Effects of Exogenous CaCl2 Treatments on C. saxicola Seedlings
In order to investigate whether Ca2+ influences the growth and development of C. saxicola, different concentrations of exogenous CaCl2 were used to treat one-month-old C. saxicola seedlings. As shown in Figure 8A-C, the calcium content, proline content, and soluble sugar content of C. saxicola leaves were significantly increased with the elevation of exogenous CaCl2 concentrations (P < 0.05). In addition, significant differences in the DHCA content in the stems, leaves, and roots of C. saxicola after treatments were also observed (P < 0.05), and the content of DHCA in roots was much higher than that in stems and leaves (Figure 8D-F). Interestingly, the high concentration of exogenous CaCl2 significantly induced the accumulation of DHCA in the roots of C. saxicola (Figure 8F).
We then analyzed the FPKM values of CsANNs based on the transcriptomic data. It was found that, except for CsANN2 and CsANN8, the FPKM values of other CsANNs showed significant changes with the increase of CaCl2 concentration (P < 0.05) (Figure 8G-O). Especially, the FPKM values of CsANN1 and CsANN9 are positively correlated with calcium concentration (Figure 8G, O).
2.8. CsANNs Correlated with DHCA Biosynthesis
According to the predicted biosynthetic pathway of DHCA, C-methyltransferases (CMTs), O-methyltransferases (OMTs), and berberine bridge enzyme-likes (BBELs) were involved in the transition of cheilanthifoline into DHCA [33]. In order to elucidate the relationship between CsANNs and the above key enzymes involved in the DHCA biosynthesis, we conducted a correlation analysis among CsCMTs, CsOMTs, CsBBELs, and CsANNs. An interaction network that comprised 34 nodes and 66 relational pairs with high correlations (R ≥ 0.8 and P ≤ 0.05 or R ≤ -0.8 and P ≤ 0.05) was identified. Among them, four major sub-networks (CsANN1, CsANN5, CsANN6, CsANN9), which collectively included 48 relational pairs and 25 nodes, were summarized (Figure S2). For CsANN1, the connectivity number is 9, and it shows significant correlation with the expression of 3 CsOMTs and 6 CsBBELs. Similarly, the connectivity number for CsANN9 is 11, which is significantly correlated with the expression of 5 CsOMTs and 6 CsBBELs (Figure S2).
As is mentioned above, the transcripts of both CsANN1 and CsANN9 were not only relatively high in the roots of C. saxicola, but also positively correlated with the content of DHCA in the roots of C. saxicola treated by exogenous CaCl2 solutions. Furthermore, the expression of CsANN1 and CsANN9 had high correlations with the expression of DHCA-biosynthetic-related genes (Figure S2). These indicated that CsANN1 and CsANN9 might play positive roles in the biosynthesis of DHCA.
To confirm this speculation, transient overexpression assays of CsANN1 and CsANN9 were conducted in C. saxicola leaves, respectively (Figure 9A). The transcript levels of CsANN1 and CsANN9 in C. saxicola leaves infiltrated with CsANN1-pCAMBIA1301 and CsANN9-pCAMBIA1301 increased significantly compared to the empty vector, respectively (Figure 9B, Figure S3A). Furthermore, the contents of Cheilanthifoline and DHCA in C. saxicola leaves infiltrated with CsANN1-pCAMBIA1301 were also significantly increased compared to the empty vector (Figure 9C, D). However, compared to the empty vector, the contents of Cheilanthifoline and DHCA in C. saxicola leaves infiltrated with CsANN9-pCAMBIA1301 did not show significant difference (Figure S3B, C). These results suggested that CsANN1 but not CsANN9 might play a positive role in the biosynthesis of DHCA.
2.9. Yeast Two Hybrid Assay and Protein Interaction Networks of the CsANNs
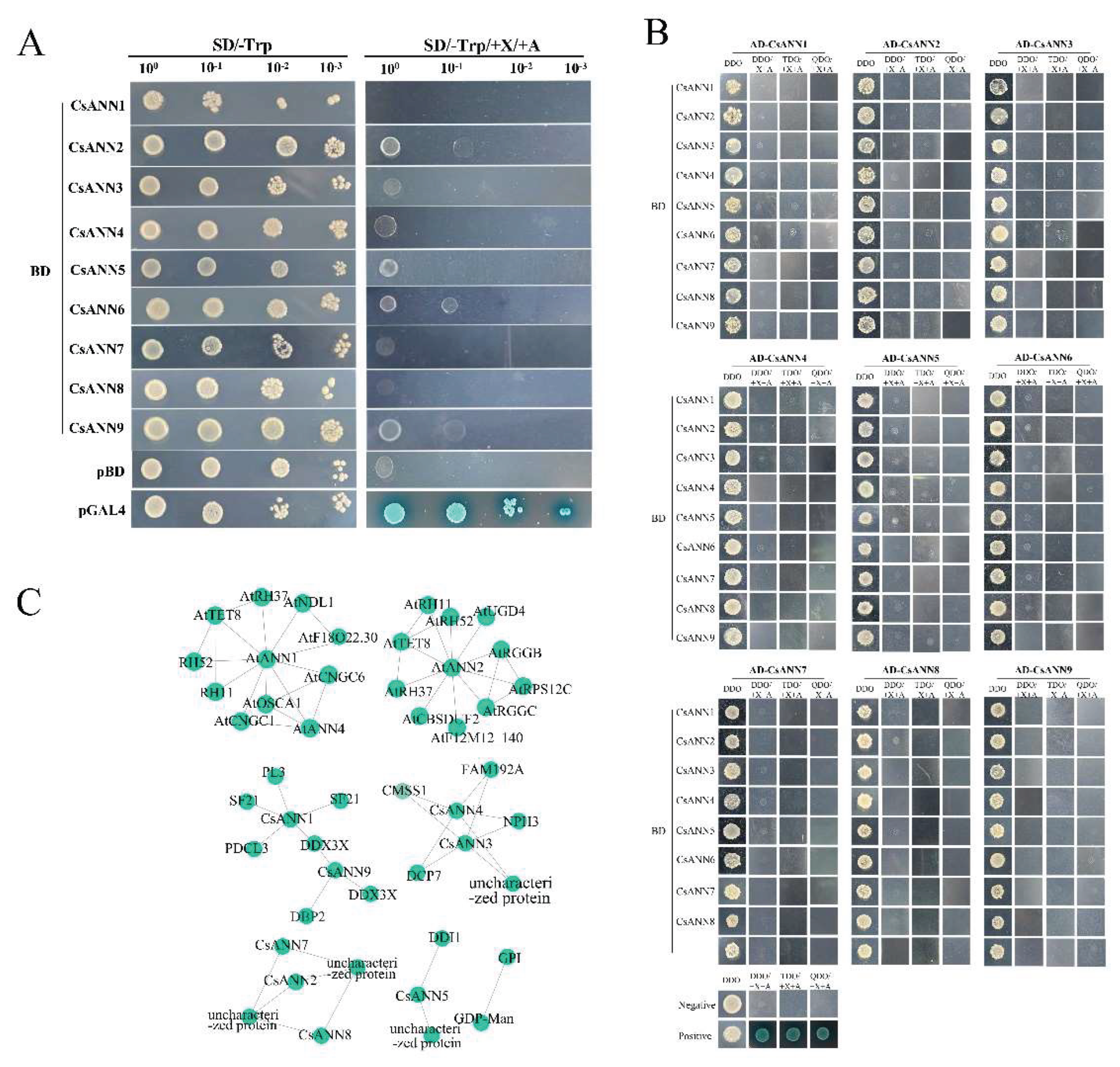
Numerous proteins could form dimers, polymers or more complicated complexes to execute biological processes in vivo [34,35]. In order to explore whether CsANNs could form homodimers or heterodimers, yeast two-hybrid assays were conducted. As shown in Figure 10A, similar to the negative control pBD, the yeast cells carrying the tested vectors could not grow on the selective solid medium lacking Tyrosine (Trp) but containing 5-Bromo-4-chloro-3-indoxyl-α-D-galactopyranoside (X-α-gal) and Aureobasidin A (AbA), indicating that all CsANNs had no auto-activation activities in Y2HGold cells (Figure 10A). Further assays indicated that either the yeast cells containing the pair of cotransformed indicated plasmids could grow on the selective medium containing X-α-gal and AbA, indicating that none of these CsANNs could form homodimers or heterodimers in yeast cells (Figure 10B).
We then utilized the STRING database and TBtools to further reveal the interactions between CsANNs and other proteins. The results showed that the interaction patterns of CsANN1 with proteins closely resemble those of AtANN1 (Figure 10C). Proteins interacting with AtANN1 include the DEAD-box ATP-dependent RNA helicase (AtRH11, 37, 52) belongs to the DDX3/DED1 subfamily of DEAD box helicase family; probable cyclic nucleotide-gated ion channel (AtCNGC1, 6) belongs to the cyclic nucleotide-gated cation channel; the tetraspanin-8 (AtTET8) belongs to the tetraspanin family; the hyperosmolarity-gated non-selective cation channel (AtOSCA1) belongs to the CSC1 family, among others. Similarly, the interaction patterns of CsANN9 with proteins are similarities to those of AtANN2. AtANN2 is predicted to interact with AtTET8; RGG repeats nuclear RNA binding protein C (AtRGGC); RGG repeats nuclear RNA binding protein B (AtRGGB) belongs to the RGGA protein family; UDP-glucose 6-dehydrogenase 4 (AtUGD4) belongs to the UDP-glucose/GDP-mannose dehydrogenase family, and other proteins. Furthermore, the interaction network of CsANNs suggested potential interactions with proteins such as Glucose-6-phosphate isomerase (GPI); GDP-mannose transporter GONST1 (GDP-Man); ATP dependent RNA helicase DDX3X (DDX3X), among others (Figure 10C). These results indicated that CsANN proteins might be involved in various physiological functions, including transcriptional regulation, RNA stability, signal transduction, mRNA transport, Golgi secretion, maintenance of cellular osmotic balance, synthesis of cell wall precursors, as well as regulation of calcium channels.
3. Discussion
3.1. The Structure of CsANNs Were Conserved
The ANNs had been identified and functionally analyzed in a variety of plant species, including model plants and crops, but the informations about ANN gene families in medicinal plants were scarce [19,21,22,23,36,37]. In this study, nine CsANN genes were identified based on the genome and transcriptome data of C. saxicola, an endangered herbaceous plant that exclusively lived in Chinese karst landforms and was widely used in traditional Chinese medicine. All the CsANN proteins have four typical ANN domains, except CsANN2 and CsANN8, which lacked one and two ANN domains, respectively (Figure 3C). It is well known that a highly conserved sequence (G/KXGT-38-D/E) for binding sites of type II Ca2+ is present in each ANN domain in vertebrates [10]. However, the type II Ca2+ binding sites were absent in the repeats II and III in plant ANNs [9]. Our results also showed that repeats II and III of CsANNs did not contain the type II Ca2+ binding sites, which was in accordance with other plant ANNs [9]. In addition, repeat I was absent in CsANN2 and CsANN8, and repeat IV was absent in CsANN8 (Figure 1B). Furthermore, although four repeats were all present in the other seven CsANNs, the type II Ca2+ binding sites were less conserved, with two or three substitutions in repeat IV of CsANNs. Actually, the type II Ca2+ binding sites were only highly conserved in repeat I of CsANN1, 5, 7, and 9 (Figure 1B). Those absences of type II Ca2+ binding sites or the entire repeats might result in different protein conformations and largely weaken the ability of the CsANN proteins to bind to phospholipids of cell membranes in a Ca2+-dependent manner, further influencing their capacities involved in plant growth and development regulation [10,11].
Another motif present in repeat I was the ILAHR sequence (Figure 1B). It was demonstrated that this sequence, especially the conserved His40 residue, played an essential role in peroxidase activity [37,38]. For example, Arabidopsis AnnAt1 has shown the His-residue-dependent peroxidase activity in vitro [16]. An AnnBr1-contained complex protein purified from B. rapa floral buds had peroxidase activity [39]. A maize ANN preparation also demonstrated peroxidase activity that appeared independent of heme association [37]. It is believed that the peroxidase activity of ANNs could protect the membrane from peroxidation and then subsequently improve the abiotic tolerance in plants, such as drought tolerance, etc [40,41]. As plants such as C. saxicola in karst regions experienced the harsh environmental conditions, the CsANNs (CsANN1, 5, 9), which present the ILAHR sequences, might contribute calcareous and drought tolerance abilities to C. Saxicola plants partly by the peroxidase activities.
The IRI sequence in repeat III of ANNs was highly conserved in CsANN1 and 9, and with one or two substitutions in other CsANNs except CsANN2 (Figure 1B). The IRI present in ANNs were demonstrated to bind filamentous actin (F-actin) but not globular actin in a Ca2+-dependent manner, indicating these ANNs might be involved in exocytosis regulation [42]. Moreover, several ANNs of maize, cotton, and tomato were proved to hydrolyze ATP and GTP, and the nucleotide hydrolyzation abilities were attributed to the Walker A motif (GXXXXGKT/S) and the GTP binding domain (GXXG) which were retained in repeat IV of ANNs [11]. CsANN1, 5, and 9 contain a modified Walker A motif with one or two substitutions and the conserved GXXG sequence in the repeat IV, indicating their potential abilities to bind and hydrolysis GTP/ATP [45]. Interestingly, similar to A. thaliana, cotton, maize, and poplar, these nucleotide binding domains were partly overlapped with the Ca2+ binding site in repeat IV, indicating the two different binding properties were contradictory to each other and thus controlled the spatiotemporal functions of ANNs [13,18,19,46].
The phylogenetic analysis showed that CsANNs were distributed in three subfamilies, and clustered with ANNs from the dicotyledonous plants, especially the ANNs in the same genus (C. tomentella and C. yanhusuo) (Figure 2). Similar to other plant ANNs, the closely related CsANNs tend to exhibit similar structural features [22,39,47,48]. As shown in Figure 3B, motifs 1–4 are common to all CsANN members, indicating that it’s crucial for CsANNs. Nevertheless, the members of subfamily II lacked motif 5, and those of subfamily III had significant differences in exon-intron structure, which may be related to functional differentiation. It is worth noting that CsANN1 had more exons and longer introns, implying that the gene plays a more crucial role in function, expression regulation, and evolution [49,50,51,52].
3.2. CsANNs were Involved in the Biosynthesis of DHCA
It was interesting that the expression profiles of CsANN1 and CsANN9 in variable tissues and Ca2+-treated roots of C. saxicola were similar to the accumulation profiles of DHCA in these treated or untreated organs, the fact of which might be an indicative of the involvement of CsANN1/9 in DHCA biosynthesis (Figure 7, 8). In fact, transient overexpression of CsANNs in C. saxicola leaves demonstrated that higher expression of CsANN1 could really result in the higher accumulation of BIAs, including palmatine, cheilanthifoline, dehydrocheilanthifoline, and cavidine, compared to that of EV (Figure 9C, D). Especially, compared to EV, the contents of DHCA and cheilanthifoline, a crucial upstream precursor compound of DHCA, were significantly elevated in CsANN1-overexpressed C. saxicola leaves, indicating that CsANN1 might positively regulate the expression of the genes or proteins involved in the biosynthesis pathway of cheilanthifoline and DHCA (Figure 9D). Further analysis of candidates participated in the biosynthesis pathway of DHCA showed that the expression level of CsBBEL9 transcripts was significantly upregulated in CsANN1-overexpressed C. saxicola leaves compared to that of EV (Figure 9B). As a result, we speculated that CsANN1 might bind directly to the DNA sequence of CsBBEL9 in the nucleus, similar to trans-acting factors, to regulate the expression of CsBBEL9, and thus induce the accumulation of DHCA and its precursors. Subcellular localization analyses of CsANN1 and CsANN9 suggested a more likely presence in the cytoplasm. However, studies have shown that some ANNs could be introduced into the nucleus through unknown mechanisms, especially under stress conditions [9,12,47,48]. One possible assistance might be supported by the conformational changes of ANN proteins induced by N-terminal tyrosine phosphorylation, allowing the proteins to enter the nucleus [47,48,49,50,51,52]. However, more biochemical and structural evidence should be provided to confirm the subcellular localizations of ANNs, including CsANNs, during physiological processes.
Another factor influencing the accumulation of DHCA by CsANN1 might occur during the translational level. A number of cis-acting elements, including MYB binding sites, were identified in the promoters of CsBBELs, CsCMTs, CsOMTs and CsANNs, and the Y2H assays demonstrated that CsANNs could interact with several CsMYBs in Y2HGold yeast cells (data not shown). Therefore, CsANNs-CsMYBs modules might play crucial roles in the biosynthesis pathway of DHCA. Further studies should be conducted to unravel the precise mechanisms of the biosynthesis of DHCA at transcriptional and translational levels, which could help to elucidate the adaptations of medicinal plants in karst regions.
3.3. CsANNs Might Play an Important Role in the Adaptability of C. saxicola to Karst Environment
It was reported that the calcium content in karst soil could reach up to 37.68 g/kg at most, and the average calcium availability of these soils were as high as 50.9% [29,53]. Preventing excessive absorption of Ca2+ is crucial for the normal growth of karst plants. For this purpose, variable unique physiological mechanisms and responsive pathways have been evolved to maintain the balance of Ca2+ within plant cells: (1) fixing excess Ca2+ by forming calcium oxalate crystals [54,55]; (2) releasing excess Ca2+ through stomata on mature leaves [56,57]; (3) regulating the concentration of Ca2+ by storing it in the intercellular spaces and organelles, such as vacuoles, endoplasmic reticulum, mitochondria, and chloroplasts [58,59]. To date, several key factors were demonstrated to be involved in these processes. For example, the Ca2+/cation antiporter superfamily, which are classified into four families, H+/cation exchangers (CAXs); Na+/Ca2+ exchanger-like proteins; cation/Ca2+ exchangers; and Mg2+/H+ exchangers, is critical in regulating and accumulating calcium [58,60].Physilogical and genetic evidences have demonstrated that CAXs could mainly remove excess Ca2+ from the cytoplasm into the vacuole to maintain calcium homeostasis and alleviating calcium-induced stresses, thus granting plants the adaptation to the high-calcium karst environment [59,60]. In addition, Ca2+-ATPase (ACA) could also prompt Ca2+ efflux from cytosol to endoplasmic reticulum, or facilate Ca2+ uptake from cytosol to vacuole, in an ATP-dependent manner [56]. In our study, compared with those in CK, the expression levels of several CsCAXs, CsACAs and other Ca2+-balancing genes were significantly upregulated in C. saxicola roots treated with high concentrations of CaCl2 solution (Figure S4), indicating the involvement of these factors in the adaptation of C. saxicola in karst landforms. In addition, the expression levels of some CsCAXs were much higher than those of CsACAs and other Ca2+-balancing genes (Figure S4). This meant that the shaping of the efflux element of Ca2+ signatures in C. saxicola was mainly contributed to the expression of CsCAXs other than CsACAs, which was consistent with the fact that CAXs has lower energy consumption than ACAs and mainly act on the vacuole membrane and are more conducive to the secretion and long-term storage of Ca2+ [56,61]. Interestingly, significant correlations between CsANNs and Ca2+-balancing genes have been identified (Figure S5), indicating the complicated adaptation styles of C. saxicola in calcium-rich karst ecosystems.
Except for the involvement in the adaptation of high-calcium environment, ANNs could also response to other stresses to help regulate plants’ growth and development, at least partly attributing to the diversity of their cis-acting elements in promoters [9,18,22,25,37,48]. Multiple cis-acting elements were also identified in the promoters of CsANN genes (Figure 6), indicating the role of CsANNs in the adaptation of C. saxicola to the complex ecological environment in karst areas. For example, the ABRE elements binding sites, which could respond to ABA signals and balance cellular osmotic pressure, present in the promoters of CsANN1, 3, 4, and 9 [62,63]. All CsANNs possess the cis-elements of MYB transcription factors, which indicated their involvements in drought resistance by regulating ion homeostasis and reducing water loss [64,65]. In addition, ARE elements, which could activate antioxidant enzymes (such as superoxide dismutase and catalase) by binding to the nuclear factor-erythroid 2-related factor 2 or basic (region-leucine) zipper transcription factors, and thus reduce the damage caused by reactive oxygen species produced by adverse environmental factors to the membrane lipids of plants, existed in all CsANNs except CsANN7 [62,66,67,68]. Furthermore, the CsANNs’ promoters also contain a variety of hormone-responsive elements, such as CGTCA-motif (cis-acting regulatory element involved in the MeJA reaction), TGACG-motif (methyl jasmonate response), P-box (gibberellin response), and TGA-element (auxin response) (Table S2), all of which are conducive to alleviating stresses to the growth, development and differentiation of plants caused by arid, infertile and heavy metal pollutions [69,70,71,72]. Based on these, we speculated that CsANNs may alter the concentration of free Ca2+ in the cytoplasm by interacting with multiple signaling pathway genes. As Ca2+ is a second messenger in plants, the characteristic changes of [Ca2+]cyt in different environments are recognized and decoded as regulatory responses in DNA, RNA, protein, or metabolic levels to help C. saxicola adapt in harsh karst environments.
4. Materials and Methods
4.1. Plant Materials and Treatments
C. saxicola seeds were germinated and grown in pots containing a matrix with a 3:1 mixture ratio of peat soil and vermiculite in the greenhouse (23 ℃, 16 h light, a photo flux density of 120 µmol m-2 s-1). For the exogenous CaCl2 treatment experiments, the one-month-old C. saxicola plants were treated with the equal volume of CaCl2 at 4 mmol/L, 30 mmol/L, 100 mmol/L, 200 mmol/L, 300 mmol/L, and 400 mmol/L, respectively, for 25 d. The same batch of plants treated with the equal volume of water were set as the control (CK). The organs were then cleaned, physically isolated, and immediately frozen or dried for further research.
4.2. Genome-Wide Identification of the CsANN Genes
Firstly, the Hidden Markov Model (HMM) with the canonical ANN domain (Pfam00191) was used to search against the C. saxicola genome dataset (P < 0.001), which was assembled by ourselves, to identify CsANN genes [73]. Secondly, all eight AtANNs downloaded from The Arabidopsis Information Resource (TAIR) database were used as queries to retrieve CsANNs by BLAST in TBtools v2.096 [31]. Lastly, putative CsANNs were further confirmed using the InterPro and SMART databases [74]. The above non-redundant sequences were considered as CsANN candidates. The protein physicochemical properties of CsANNs were analyzed using the online website ExPASy [75]. Subcellular localizations of CsANNs were predicted using the online websites Wolf-psort, Cell-PLoc 2.0, CELLO, PSORT2, and Euk-mPLoc 2.0 [76,77,78].
4.3. Phylogenetic Relationship, Conserved Motif, and Gene Structure Analysis
The multiply alignment of the CsANNs was performed by the TBtools v2.096 [31]. The results were then visualized using the Jalview 9.0.5 [79]. The ANN protein sequences of A. thaliana, B. oleracea, B. rapa, C. saxicola, C. tomentella, C. yanhusuo, H. vulgare, O. sativa, and P. tremula were aligned using MEGA 11.0 [18]. The phylogenetic tree of the ANNs was constructed using the neighbor-joining method, with the bootstrap value of 1000 and other parameters set to default parameters, and then was visualized by FigTree v1.4.4. Conserved motifs of CsANNs were identified using the MEME Suite 5.5.7 [80]. The intron and exon regions of CsANNs were analyzed using the Visualize Gene Structure program in TBtools v2.096 [31].
4.4. Chromosomal Location, Collinearity, and Gene Duplication Analysis
The chromosome location information of CsANN genes was extracted from the C. saxicola plant genome gff file using the TBtools software, and visualizations were generated [31]. TBtools was further employed to conduct and visualize the collinearity analysis of the CsANNs. Additionally, the collinearity analysis of A. thaliana, C. tomentella, H. vulgare, O. sativa, and P. tremula with C. saxicola was performed using TBtools [31].
4.5. Cis-Acting Elements, and Protein-Protein Interaction Analysis
The 2 kb sequences upstream of the 5’ UTR of CsANN genes were extracted using the TBtools software and then used for the prediction of cis-acting elements based on the PlantCARE database [32]. Protein-protein interactions were predicted using the STRING database [81]. Cytoscape V3.9 was used for visualization [82].
4.6. Measurement of the BIAs Contents
An Agilent 1260 Infinity II (Agilent Co. Ltd, NY, USA) was employed to determine the relative content of the BIAs (cavidine, (s)-cheilanthifoline, dehydrocheilanthifoline, DHCA, and palmatine). Initially, 200.0 mg of dried C. Saxicola samples were dissolved in 10 mL of methanol and then ultrasonicated for 60 minutes. Next, the samples were centrifuged at 13,000 g for 15 min, and the supernatant was removed to a new 15-mL tube and dried using a nitrogen pressure reduction method at room temperature. Thereafter, the sample was dissolved by 1 ml of the mobile phase (acetonitrile: 0.01% K2HPO4 aqueous solution = 21:79 (v/v)) and filtered through a 0.22-μm filter prior to analysis. The Agilent XDBC 18 chromatographic column (150 mm × 4.6 mm, 5 µm) was used to elute the BIAs in samples under 347 nm of detection wavelength at 30 °C. The volumetric flow rate of the mobile phase (acetonitrile: 0.01% of K2HPO4 = 21:79 (v/v)) is 1.0 mL/min.
4.7. CsANN Cloning
The RNA was extracted from leaves, stems, and roots of C. saxicola before and after various treatments by FastPure Universal Plant Total RNA Isolation kit (Vazyme, Nanjing, China) based on the instruction. The first-strand cDNA was synthesized according to the instruction of HiScript Ⅲ 1st Strand cDNA Synthesis Kit (Vazyme, Nanjing, China) with the extracted total RNA as templates. To clone the full-length CDSs of CsANNs, the gene-specific primers (Table S3) were designed and the PCR reaction procedures were as follows: pre-denaturation at 95 ℃ for 3 min; followed by 33 cycles of denaturation at 95 ℃ for 15 s, annealing at 56 ℃ (CsANN2, 6) or 54 ℃ (other CsANNs) for 15 s, and extension at 72 ℃ for 50 s; and a final extension at 72 ℃ for 5 min. Subsequently, PCR products were ligated into the TA/Blunt-Zero Cloning Kit vector (Vazyme, Nanjing, China) and then transformed into DH5α competent cells for incubation on LB solid medium plus 100 µg mL-1 kanamycin overnight. Colony PCR with the same procedure above were conducted to verify the putative positive transformants. The final CDSs of CsANNs were ascertained by Sanger sequencing.
4.8. qRT-PCR Analysis
The qRT-PCR was performed using ChamQ Universal SYBR qPCR Master Mix (Vazyme, Nanjing, China) with three technical replicates. The relative expression levels of specific genes were calculated using the 2 delta delta Ct method (2−△△Dt method), with glyceraldehyde-3-phosphate dehydrogenase 8 (CsGAPDH8) used as the internal control [82,83,84,85]. The qRT-PCR primers were listed in Table S3.
4.9. Yeast Two-Hybrid Assay
The full-length CDS of CsANNs were amplified and cloned into the destination vectors pGADT7 (pAD) and pGBKT7 (pBD), respectively. The primers used were listed in Table S3. Transactivation analysis assays were carried out in the yeast strain Y2HGold using the Yeastmaker Yeast Transformation System 2 (Clontech, Tokyo, Japan). For Y2H assays, CsANN-pAD and CsANN-pBD were transformed into Y187 and Y2HGold yeast cells, respectively. Yeast mating was conducted and the putative cotransformants were screened on SD/-Trp/-Leu (DDO) selective solid medium and then verified by PCR. SD/-Ade/-Trp/-Leu (TDO)+X+A and SD/-His/-Ade/-Trp/-Leu (QDO)+X+A were further adopted to verify the protein-protein interactions.
4.10. Transient Overexpression of CsANN1 and CsANN9 in C. saxicola Leaves
The full-length CDS of CsANN1 and CsANN9 were amplified and cloned into the pCAMBIA1301 vector, respectively. The primers used were listed in Table S3. The resulting plasmids were introduced into Agrobacterium tumefaciens GV3101. After propagation, the cells were resuspended in infiltration buffer (composed of 10 mmol/L MgCl2, 10 mmol/L 2-morpholinoethanesulfonic acid, and 150 mmol/L Acetosyringone; pH 5.6) to achieve the OD600 of 0.8. Suspensions containing either the overexpressing constructs of the target genes CsANN1 and CsANN9 or the empty pCAMBIA1301 plasmid vector (serving as a control) were injected into opposite sides of the same leaf. Two days later, the injected leaves of C. saxicola were collected for metabolite content and gene expression analysis using HPLC and qRT-PCR, respectively [86].
5. Conclusions
In this study, we first provided a systematic analysis of the nine CsANN gene families in C. saxicola. Phylogenetic analysis of ANNs within nine species indicated that all nine CsANNs can be classified into three different subfamilies, with closely related members having similar structural characteristics. In addition, gene structures, conserved motifs, chromosomal distributions, collinearities, protein interactions, and cis-acting elements of CsANN genes or CsANN proteins were also analyzed. Expression pattern analysis unraveled two candidates, CsANN1 and CsANN9, as the putative positive regulators involved in the biosynthesis of DHCA in C. saxicola. Further experiments indicated that transient overexpression of CsANN1, but not CsANN9, could increase the content of DHCA in C. saxicola leaves. This study provides a theoretical basis for understanding the biological functions of the CsANNs in secondary metabolites and adaptation to adverse stresses of C. saxicola.
Supplementary Materials
The following supporting information can be downloaded at Preprints.org, Figure S1: The biosynthetic pathway of BIAs. (Each color represents a branch of different secondary metabolites synthesized by scoulerine, green: the biosynthetic pathway of palmatine synthesized by scoulerine; blue: the predicted biosynthetic pathway of DHCA synthesized from scoulerine; yellow: the biosynthetic pathway of sanguinarine synthesized from scoulerine; purple: the biosynthetic pathway of codeine and morphine synthesized from scoulerine. TYDC: L-tyrosine decarboxylase; TAT: tyrosine aminotransferase; TYR: tyrosine 3-monooxygenase; HPPDC: 4-hydroxyphenylpyruvate dioxygenase; NCS: norcoclaurine synthase; 6OMT: norcoclaurine 6 O-methyltransferase; CNMT: coclaurine N-methyltransferase; NMCH: N-methylcoclaurine 3’-hydroxylase; 4’OMT: 3’-hydroxy-N-methylcoclaurine 4’-O-methyltransferase; BBE: berberine bridge enzyme; 9OMT: scoulerine 9-O-methyltransferase; BBEL: berberine bridge enzyme-like; OMT: O-methyltransferase; CMT: C-methyltransferase; CFS: cheilanthifoline synthase; SPS: stylopine synthase; TNMT: tetrahydroprotoberberine N-methyltransferase; MSH: N-methylstylopine 14-hydroxylase; P6H: protopine 6-hydroxylase; DBOX: dihydrobenzophenanthridine oxidase; REPI: reticuline epimerase; SalSyn, salutaridine synthase); Figure S2: Correlation analysis of CsOMTs, CsCMTs, and CsBBELs with CsANNs; Figure S3: The transient overexpression and functional analysis of and CsANN9 (Expression profiles of CsANN9 and other genes in C. saxicola leaves infiltrated with CsANN9-pCAMBIA1301 or EV (A), Changes in BIA contents in C. saxicola leaves infiltrated with CsANN9-pCAMBIA1301 or EV (B), Statistic analysis of BIA contents in C. saxicola leaves infiltrated with CsANN9-pCAMBIA1301 or EV(C)); Figure S4: Expression of the genes involved in Ca2+ sensor and signal-relay pathways in C. saxicola under different concentrations of CaCl2 solution (CIPK, CBL-interacting protein; CAM, calmodulin; CML, calmodulin-like gene; GLR, glutamate-like receptors; CDPK, calcium-dependent protein kinase; CBL, calcineurin B-like protein; CAX, vacuolar Ca2+ exchanger; ACA, Ca2+-ATPase; CNGC, cyclic nucleotide-gated channel); Figure S5: Correlation analysis of the genes involved in Ca2+ sensor and signal-relay pathways with CsANNs; Table S1: Identification and physicochemical analysis of CsANNs; Table S2: The cis-acting elements identified in CsANN promoters; Table S3: Primers used in this study; Table S4. The accession numbers of genes involved in the study.
Author Contributions
Conceptualization, L.H. and X.O.; methodology, L.H. and Z.W.; data curation, L.K. and Y.L.; writing—original draft preparation, H.L. and M.L.; writing—review and editing, H.L., J.W. and M.L.; project administration, M.Q. and Z.W.; funding acquisition, C.L. and Z.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Guangxi Science and Technology Base and Talent Project (Guike AD22080016); the Natural Science Foundation of Guangxi (2025GXNSFBA069023; 2025GXNSFAA069814; 2023GXNSFAA026509); the National Natural Science Foundation of China (32460105 and 32260107); Guangxi Key R&D Program (Guike AB23026092); the Central Guidance on Local Science and Technology Development Fund of Guangxi (GK ZY24212031); Guangxi Key Laboratory of Medicinal Resources Protection and Genetic Improvement (KL2022ZZ04, KL2023ZZ09 and KL2023KF04).
Data Availability Statement
All relevant data are within the manuscript and its Additional files.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Yan, G.; Lin, Z.; Bo, C.; Jia, Y.; Jiang, B.; Qi, Y.; Jun, L. The Traditional Uses, Phytochemistry, Pharmacokinetics, Pharmacology, Toxicity, and Applications of Corydalis Saxicola Bunting: A Review. Front. Pharmacol. 2022, 13, 822792. [Google Scholar] [CrossRef]
- Xu, L.; Chao, T.; Hua, Z.; Jin, W.; Fang, W.; Yi, Y.M.; Xi, L.; Hong, Z.; Chun, Y.; Bang, C.; et al. Investigation of the Hepato-protective Effect of Corydalis Saxicola Bunting on Carbon Tetrachloride-Induced Liver Fibrosis in Rats by 1H-NMR-Based Metabonomics and Network Pharmacology Approaches. J. Pharm. Biomed. Anal. 2018, 159, 252–261. [Google Scholar] [CrossRef]
- Feng, Q.; Yao, C.; Fan, W.; Shao, T.; Yi, F. Corydalis Saxicola Bunting: A Review of Its Traditional Uses, Phytochemistry, Pharmacology, and Clinical Applications. Int. J. Mol. Sci. 2023, 24, 1626. [Google Scholar] [CrossRef]
- Fan, Z.; Yang, X.; Zhen, L.; Xiao, W.; Dan, H.; Sui, Z.; Min, L.; De, Y.; Dong, W.; Yi, W. Anti-Hepatitis B Virus Effects of Dehydrocheilanthifoline from Corydalis Saxicola. Am. J. Chin. Med. 2013, 41, 119–130. [Google Scholar] [CrossRef]
- Wang, T.; Sun, N.L.; Zhang, W.D.; Li, H.L.; Lu, G.C.; Yuan, B.J.; Jiang, H.; She, J.H.; Zhang, C. Protective Effects of Dehy-drocavidine on Carbon Tetrachloride-Induced Acute Hepatotoxicity in Rats. J. Ethnopharmacol. 2008, 117, 300–308. [Google Scholar] [CrossRef]
- Li, C.; Liu, H.; Qin, M.; Tan, Y.J.; Ou, X.L.; Chen, X.Y.; Wei, Y.; Zhang, Z.J.; Lei, M. RNA Editing Events and Expression Profiles of Mitochondrial Protein-Coding Genes in the Endemic and Endangered Medicinal Plant, Corydalis Saxicola. Front. Plant Sci. 2024, 15, 1332460. [Google Scholar] [CrossRef]
- Liu, C.X.; Duan, Y.; Xie, H.T.; Tian, J. Challenges in Research and Development of Traditional Chinese Medicines. Institute of Pharmaceutical Research. Nanjing, China, 2007: 7102–8.
- Hui, L.; Wei, Z.; Chuan, Z.; Ting, H.; Run, L.; Jiang, H.; Hai, C. Comparative Analysis of the Chemical Profile of Wild and Cultivated Populations of Corydalis Saxicola by High-Performance Liquid Chromatography. Phytochem. Anal. 2007, 18, 393–400. [Google Scholar] [CrossRef]
- Xiao, W.; Yan, W.; Yu, B.; Yan, R.; Xiao, X.; Fu, Z.; Hai, D. A Critical Review on Plant Annexin: Structure, Function, and Mechanism. Plant Physiol. Biochem. 2022, 190, 81–89. [Google Scholar] [CrossRef]
- Volker, G.; Carl, C.; Stephen, M. Annexins: Linking Ca; Signalling to Membrane Dynamics. Nat. Rev. Mol. Cell Biol. 2005, 6, 449–461. [Google Scholar] [CrossRef]
- C, M.J.; Anuphon, L.; Neil, M.; Alex, W.; Colin, B.; H, B.N.; M, D.J. Annexins: Multifunctional Components of Growth and Adaptation. J. Exp. Bot. 2008, 59, 533–544. [Google Scholar] [CrossRef]
- Joanna, P.; Aneta, K.; Ruud, D.; Marcin, G.; Michael, K.; Jean, L.; Slawomir, P.; René, B. A Putative Consensus Sequence for the Nucleotide-Binding Site of Annexin A6. Biochemistry. 2003, 42, 9137–9146. [Google Scholar] [CrossRef]
- Michaela, T.; Hendrik, R.; Miroslav, O.; Nicola, M.; Miroslava, H.; Petr, D.; Jozef, Š.; Olga, Š. Advanced Microscopy Reveals Complex Developmental and Subcellular Localization Patterns of ANNEXIN 1 in Arabidopsis. Front. Plant Sci. 2020, 11, 1153. [Google Scholar] [CrossRef]
- Volker, G.; E, M.S. Annexins: From Structure to Function. Physiol. Rev. 2002, 82, 331–371. [Google Scholar] [CrossRef]
- A, H.; J, P.; A, D.; R, S.; R, H. Annexin 24 from Capsicum Annuum X-Ray Structure and Biochemical Characterization. J. Biol. Chem. 2000, 275, 8072–8082. [Google Scholar] [CrossRef]
- Dorota, P.; Greg, C.; Grazyna, G.; Janusz, D.; Krzysztof, F.; Araceli, C.; Bartlomiej, F.; Stanley, R.; Jacek, H. The Role of Annexin 1 in Drought Stress in Arabidopsis. Plant Physiol. 2009, 150, 1394–1410. [Google Scholar] [CrossRef]
- B, C.G.; A, S.; J, E.D.; J, R.S. Differential Expression of Members of the Annexin Multigene Family in Arabidopsis. Plant Physiol. 2001, 126, 1072–1084. [Google Scholar] [CrossRef]
- Hui, W.; Ali, M.; Guo, L.; Yi, L.; Shi, L.; Chun, Y.; Yan, C.; Fei, Z.; Jian, Z. Genome-Wide Characterization and Abiotic Stresses Expression Analysis of Annexin Family Genes in Poplar. Int. J. Mol. Sci. 2022, 23, 515. [Google Scholar] [CrossRef]
- Mei, Z.; Xiong, Y.; Qian, Z.; Ming, Z.; En, Z.; Yi, T.; Xue, Z.; Ji, S.; Yan, W. Induction of Annexin by Heavy Metals and Jasmonic Acid in Zea Mays. Funct. Integr. Genomics 2013, 13, 241–251. [Google Scholar] [CrossRef]
- Yadav, D.; Ahmed, I.; Kirti, P.B. Genome-Wide Identification and Expression Profiling of Annexins in Brassica Rapa and Their Phylogenetic Sequence Comparison with B. Juncea and A. Thaliana Annexins. Plant Gene 2015, 4, 109–124. [Google Scholar] [CrossRef]
- Feng, S.; Jia, Y.; Liang, X.; Xiao, S.; Ji, W.; Yan, W.; Yi, M.; Yue, Z.; Li, L. Characterization of Annexin Gene Family and Func-tional Analysis of RsANN1a Involved in Heat Tolerance in Radish (Raphanus Sativus L.). Physiol. Mol. Biol. Plants. [CrossRef]
- Kumar, J.S.; B, C.G.; T, A.B.; J, R.S.; B, K.P. Identification and Characterization of Annexin Gene Family in Rice. Plant Cell Rep. 2012, 31, 813–825. [Google Scholar] [CrossRef]
- Feng, Y.M.; Wei, X.K.; Liao, W.X.; Huang, L.H.; Zhang, H.; Liang, S.C.; Peng, H. Molecular Analysis of the Annexin Gene Family in Soybean. Biol. plant. 2013, 57, 655–662. [Google Scholar] [CrossRef]
- Hee, Y.; Pat, H.; Venkatesan, S. Analysis of the Female Gametophyte Transcriptome of Arabidopsis by Comparative Expression Profiling. Plant Physiol. 2005, 139, 1853–1869. [Google Scholar] [CrossRef]
- Cantero, A.; Barthakur, S.; Bushart, T.; Chou, S.; Morgan, R.; Fernandez, M.; Clark, G.; Roux, S. Expression Profiling of the Arabidopsis Annexin Gene Family during Germination, de-etiolation and Abiotic Stress. Plant Physiol. Biochem. (Issy-les-Moulineaux, Fr.) 2006, 44, 13–24. [CrossRef]
- Guy, B.; Heidi, N.; Jeppe, E.; L, N.K.; Malene, J.; G, W.K. Patatins, Kunitz Protease Inhibitors and Other Major Proteins in Tuber of Potato Cv. Kuras. FEBS J. 2006, 273, 3569–3584. [Google Scholar] [CrossRef]
- D, S.; N, R.; J, D.A.; J, J. Sulfolipid Is a Potential Candidate for Annexin Binding to the Outer Surface of Chloroplast. Biochem. Biophys. Res. Commun. 2000, 272, 519–524. [Google Scholar] [CrossRef]
- Patrick, G.; Kristin, K.; Anna, K.; Anja, B.; Julia, K. Towards the Proteome of Brassica Napus Phloem Sap. Proteomics 2006, 6, 896–909. [Google Scholar] [CrossRef]
- Tang, J.; Tang, X.; Qin, Y.; He, Q.; Yi, Y.; Ji, Z. Karst Rocky Desertification Progress: Soil Calcium as a Possible Driving Force. Sci. Total. Environ. 2019, 649, 1250–1259. [Google Scholar] [CrossRef]
- Zhi, L.; Li, Z.; Zhong, Y.; Zhi, R.; Feng, H.; Xin, T.; Zhi, X.; Fang, D.; Zuo, Y.; Guang, L.; et al. Genomic Mechanisms of Physiological and Morphological Adaptations of Limestone Langurs to Karst Habitats. Mol. Biol. Evol. 2020, 37, 952–968. [Google Scholar] [CrossRef]
- Cheng, C.; Hao, C.; Yi, Z.; Hannah, T.; Margaret, F.; Ye, H.; Rui, X. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Magali, L.; Patrice, D.; Gert, T.; Kathleen, M.; Yves, M.; Peer, V.; Pierre, R.; Stephane, R. PlantCARE, a Database of Plant Cis-Acting Regulatory Elements and a Portal to Tools for in Silico Analysis of Promoter Sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Xu, Z.; Li, Z.; Ren, F.; Gao, R.; Wang, Z.; Zhang, J.; Zhao, T.; Ma, X.; Pu, X.; Xin, T.; et al. The Genome of Corydalis Reveals the Evolution of Benzylisoquinoline Alkaloid Biosynthesis in Ranunculales. Plant J. 2022, 111, 217–230. [Google Scholar] [CrossRef]
- Ferré, S.; Casadó, V.; Devi, L.A.; Filizola, M.; Jockers, R.; Lohse, M.J.; Milligan, G.; Pin, P.; Guitart, X. G Protein–Coupled Re-ceptor Oligomerization Revisited: Functional and Pharmacological Perspectives. Pharmacol. Rev. 2014, 66, 413–434. [Google Scholar] [CrossRef]
- Delaux, M.; Séjalon, N.; Bécard, G.; Ané, M. Evolution of the Plant-Microbe Symbiotic “Toolkit”. Trends Plant Sci 2013, 18, 298–304. [Google Scholar] [CrossRef]
- Jami, K.; Clark, B.; Ayele, T.; Ashe, P.; Kirti, B. Genome-Wide Comparative Analysis of Annexin Superfamily in Plants. PLoS One 2012, 7, e47801. [Google Scholar] [CrossRef] [PubMed]
- C, M.J.; M, C.K.; Anuphon, L.; M, D.J. Heme-Independent Soluble and Membrane-Associated Peroxidase Activity of a Zea Mays Annexin Preparation. Plant signaling behave. 2009, 4, 428–430. [Google Scholar] [CrossRef]
- M, G.K.; Dorota, K.; Jacek, H.; Rene, B.; Slawomir, P. Peroxidase Activity of Annexin 1 from Arabidopsis Thaliana. Biochem. Biophys. Res. Commun. 2005, 336, 868–875. [Google Scholar]
- Xin, H.; Li, L.; Sai, X.; Min, Y.; Pan, X.; Wei, L.; Yu, K.; Lu, H.; Mei, W.; Lun, Q.; et al. Comprehensive Analyses of the Annexin (ANN) Gene Family in Brassica Rapa, Brassica Oleracea and Brassica Napus Reveals Their Roles in Stress Response. Sci. Rep. 2020, 10, 4295. [Google Scholar]
- Raina, I.; Javeria, E.; Sheng, G.; Teng, L.; Muhammad, I.; Zhi, Y.; Taotao, W. Overexpression of Annexin Gene AnnSp2, Enhances Drought and Salt Tolerance through Modulation of ABA Synthesis and Scavenging ROS in Tomato. Sci. Rep. 2017, 7, 12087. [Google Scholar]
- Feng, Z.; Shufen, L.; Shu, Y.; Like, W.; Wang, G. Retraction Note to: Overexpression of a Cotton Annexin Gene, GhAnn1, Enhances Drought and Salt Stress Tolerance in Transgenic Cotton. Plant Mol. Biol. 2018, 98, 185. [Google Scholar] [CrossRef]
- Hoshino, D.; Hayashi, A.; Temi, Y.; Kan, N.; Tsuchiya, T. Biochemical and Immunohistochemical Characterization of Mimosa Annexin. Planta 2004, 219, 867–875. [Google Scholar] [CrossRef]
- Forero, C.; Wasserman, M. Isolation and Identification of Actin-Binding Proteins in Plasmodium Falciparum by Affinity Chromatography. Mem. Inst. Oswaldo Cruz 2000, 95, 329–337. [Google Scholar] [CrossRef]
- Calvert, C.M.; Gant, S.J.; Bowles, D.J. Tomato Annexins P34 and P35 Bind to F-Actin and Display Nucleotide Phosphodiesterase Activity inhibited by Phospholipid Binding. Plant cell 1996, 8, 333–342. [Google Scholar] [CrossRef] [PubMed]
- Nien, H.; Mohd, Y.A.; An, W.; Asiah, O.; K, R.A.; Andreas, H. The Crystal Structure of Calcium-Bound Annexin Gh1 from Gossypium Hirsutum and Its Implications for Membrane Binding Mechanisms of Plant Annexins. J. Biol. Chem. 2008, 283, 18314–18322. [Google Scholar] [CrossRef]
- Feng, Z.; Xuan, J.; Like, W.; Shu, L.; Shuang, W.; Chao, C.; Tian, Z.; Wang, G. A Cotton Annexin Affects Fiber Elongation and Secondary Cell Wall Biosynthesis Associated with Ca2+ Influx, ROS Homeostasis, and Actin Filament Reorganization. Plant Physiol. 2016, 171, 1750–1770. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, M.; Zhao, X.; Shen, M.; Feng, R.; Wei, Q. The Evolution, Variation and Expression Patterns of the Annexin Gene Family in the Maize Pan-Genome. Sci. Rep. 2025, 15, 5711. [Google Scholar] [CrossRef]
- Chen, L.; Chen, K.; Xi, X.; Du, X.; Zou, X.; Ma, Y.; Song, Y.; Luo, C.; Wei, S. The Evolution, Expression Patterns, and Domes-tication Selection Analysis of the Annexin Gene Family in the Barley Pan-Genome. Int. J. Mol. Sci. 2024, 25. [Google Scholar] [CrossRef]
- Gil, A. The Alternative Genome. Sci. Am. 2005, 292, 40–47. [Google Scholar]
- Barmak, M.; Christopher, L. A Genomic View of Alternative Splicing. Nat. Genet. 2002, 30, 13–19. [Google Scholar] [CrossRef]
- Jeffares, D.C.; Penkett, J.; Bähler, J. Rapidly Regulated Genes Are Intron Poor. Trends Genet. 2008, 24, 375–378. [Google Scholar] [CrossRef]
- William, R.S.; Walter, G. The Evolution of Spliceosomal Introns: Patterns, Puzzles and Progress. Nat. Rev. Genet. 2006, 7, 211–221. [Google Scholar] [CrossRef]
- Maranhão, C.; Pereira, G.; Collier, L.S.; Anjos, L.H.C.; Azevedo, A.C.; Souza, R. Pedogenesis in a Karst Environment in the Cerrado Biome, Northern Brazil. Geoderma 2020, 365, 114169. [Google Scholar] [CrossRef]
- Volk, G.M.; Lynch, V.J.; Kostman, T.A.; Goss, L.J.; Franceschi, V.R. The Role of Druse and Raphide Calcium Oxalate Crystals in Tissue Calcium Regulation in Pistia stratiotes Leaves. Plant Biol. (Berlin, Ger.) 2002, 4, 34–45. [Google Scholar] [CrossRef]
- Ilarslan, H. Calcium Oxalate Crystals in Developing Seeds of Soybean. Ann. Bot. (Oxford, U. K.) 2001, 88, 243–57. [Google Scholar] [CrossRef]
- Wei, X.; Deng, X.; Xiang, W. Calcium content and high calcium adaptation of plants in karst areas of southwestern Hunan, China. Biogeosciences 2018, 15, 2991–3002. [Google Scholar] [CrossRef]
- Musetti, R.; Favali, M. Cytochemical localization of calcium and X-ray microanalysis of Catharanthus roseus L. infected with phytoplasmas. Micron 2003, 34, 387–393 [CrossRef] [PubMed]. [Google Scholar] [CrossRef]
- Hanger, B.C. The movement of calcium in plants. Commun. Soil Sci. Plant Anal. 1979, 10, 171–193. [Google Scholar] [CrossRef]
- Poovaiah, B.W.; Reddy, A.S.N.; Leopold, A.C. Calcium messenger system in plants. Crit. Rev. Plant Sci. 1987, 6, 47–103. [Google Scholar] [CrossRef]
- Zhou, Y.; Fan, W.; Zhang, H. Marsdenia tenacissima genome reveals calcium adaptation and tenacissoside biosynthesis. Plant J. 2022, 113, 1146–1159. [Google Scholar] [CrossRef]
- Huang, H.; Yang, Y.; Li, J. Effects of Rocky Desertification Stress on Oat (Avena sativa L.) Seed Germination and Seedling Growth in the Karst Areas of Southwest China. Plants 2024, 13, 3260–3260. [Google Scholar] [CrossRef]
- Ya, L.; Xuan, X.; Lu, G.; Jia, Z.; Chao, G.; Zhi, W.; Hong, L.; Xing, L.; Xiao, Y.; Na, Z.; et al. CsbZIP50 Binds to the G-Box/ABRE Motif in CsRD29A Promoter to Enhance Drought Tolerance in Cucumber. Environ. Exp. Bot. 2022, 199. [Google Scholar]
- Zheng, J.; Wen, D.; Tang, C.; Lai, S.; Yan, Y.; Du, C.; Zhang, Z. The Transcriptional Regulation of Arabidopsis ECT8 by ABA-Responsive Element Binding Transcription Factors in Response to ABA and Abiotic Stresses. Physiol. Mol. Biol. Plants 2025, 31, 1–13. [Google Scholar] [CrossRef]
- Wang, X.; Wei, H.; Wang, K.; Tang, X.; Li, S.; Zhang, N.; Si, H. MYB Transcription Factors: Acting as Molecular Switches to Regulate Different Signaling Pathways to Modulate Plant Responses to Drought Stress. Ind. Crops Prod. 2025, 226, 120676. [Google Scholar] [CrossRef]
- Liu, Z.; Li, J.; Li, S.; Song, Q.; Miao, M.; Fan, T.; Tang, X. The 1R-MYB Transcription Factor SlMYB1L Modulates Drought Tol-erance via an ABA-Dependent Pathway in Tomato. Plant Physiol. Biochem. 2025, 222, 109721. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, Y.; Lu, H. The bZIP transcription factor ATF1 regulates blue light and oxidative stress responses in Trichoderma guizhouense. mLife 2023, 2, 365–77. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wu, F.; Liu, L. The bZIP transcription factor PfZipA regulates secondary metabolism and oxidative stress response in the plant endophytic fungus Pestalotiopsis fici. Fungal Genet. Biol. 2015, 81, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zha, W.; Liang, L.; Faso, O.E.; Wu, L.; Wang, S. The bZIP Transcription Factor AflRsmA Regulates Aflatoxin B1 Biosynthesis, Oxidative Stress Response and Sclerotium Formation in Aspergillus flavus. Toxins 2020, 12, 271. [Google Scholar] [CrossRef]
- M, G.; N, F.; E, G.; E, M.; G, P.M. The ACA4 Gene of Arabidopsis Encodes a Vacuolar Membrane Calcium Pump That Improves Salt Tolerance in Yeast. Plant Physiol. 2000, 124, 1814–1827. [Google Scholar] [CrossRef]
- Liu, L.; Yahaya, S.; Li, J.; Wu, F. Enigmatic Role of Auxin Response Factors in Plant Growth and Stress Tolerance. Front. Plant Sci. 2024, 15, 1398818. [Google Scholar] [CrossRef]
- Yuan, Y.; Stéphanie, S.; Aurore, C. A downstream mediator in the growth repression limb of the jasmonate pathway. Plant Cell 2007, 19, 2470–2483. [Google Scholar] [CrossRef]
- Kazuko, Y.S.; Kazuo, S. Organization of Cis-Acting Regulatory Elements in Osmotic- and Cold-Stress-Responsive Promoters. Trends Plant. Sci. 2005, 10, 88–94. [Google Scholar] [CrossRef]
- M, S.; C, S.; J, C.E. Gene Hunting with Hidden Markov Model Knockoffs. Biometrika 2019, 106, 1–18. [Google Scholar] [CrossRef]
- Lorna, R.; Neil, R.; Gustavo, S.; Alexandre, A.; David, H.; Gregory, D.; Granger, S.; Robert, F. Genome Properties in 2019: A New Companion Database to InterPro for the Inference of Complete Functional Attributes. Nucleic Acids Res. 2019, 47, D564–D572. [Google Scholar] [CrossRef]
- Séverine, D.; Chiara, G.; Frédérique, L.; Heinz, S.; Vassilios, I.; Christine, D. Expasy, the Swiss Bioinformatics Resource Portal, as Designed by Its Users. Nucleic Acids Res. 2021, 49, W216–W227. [Google Scholar]
- Ivica, L.; Leo, G.; J, D.N.; Tobias, D.; Joerg, S.; Richard, M.; Francesca, C.; R, C.R.; P, P.C.; Peer, B. Recent Improvements to the SMART Domain-Based Sequence Annotation Resource. Nucleic Acids Res. 2002, 30, 242–244. [Google Scholar] [CrossRef]
- Kuo, C.; Hong, S. Cell-PLoc: A Package of Web Servers for Predicting Subcellular Localization of Proteins in Various Organisms. Nat. Protoc. 2008, 3, 153–162. [Google Scholar] [CrossRef]
- Kuo, C.; Hong, S. Euk-mPLoc: A Fusion Classifier for Large-Scale Eukaryotic Protein Subcellular Location Prediction by Incorporating Multiple Sites. J. Proteome Res. 2007, 6, 1728–1734. [Google Scholar] [CrossRef]
- Andrew, W.; Jim, P.; David, M.; Geoffrey, B. Jalview: Visualization and Analysis of Molecular Sequences, Alignments, and Structures. BMC Bioinform. 2005, 6, P28. [Google Scholar] [CrossRef]
- Bailey, T.L.; Johnson, J.; Grant, C.E.; Noble, W.S. The MEME Suite. Nucleic Acids Res. 2015, 43, W39–W49. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Nastou, K.; Koutrouli, M.; Kirsch, R.; Mehryary, F.; Hachilif, R.; Hu, D.; Peluso, M.E.; Huang, Q.; Fang, T.; et al. The STRING Database in 2025: Protein Networks with Directionality of Regulation. Nucleic Acids Res. 2024. [CrossRef]
- David, O.; John, M.; Jorge, B.; Alexander, P.; Barry, D. Cytoscape Automation: Empowering Workflow-Based Network Analysis. Genome Biol. 2019, 20, 185. [Google Scholar] [CrossRef]
- Zhu, Y.; Niu, S.; Lin, J.; Yang, H.; Zhou, X.; Wang, S.; Liu, X.; Yang, Q.; Zhang, C.; Zhuang, Y.; et al. Genome-Wide Identification and Expression Analysis of TCP Transcription Factors Responding to Multiple Stresses in Arachis Hypogaea L. Int. J. Mol. Sci. 2025, 26, 1069. [Google Scholar] [CrossRef]
- Shao, Y.; Mu, D.; Zhou, Y.; Liu, X.; Huang, X.; Wilson, I.W.; Qi, Y.; Lu, Y.; Zhu, L.; Zhang, Y.; et al. Genome-Wide Mining of CULLIN E3 Ubiquitin Ligase Genes from Uncaria Rhynchophylla. Plants 2024, 13, 532. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Lv, M.; Zeng, M.; Chen, X.; Wang, J.; Liang, X.F. Genome-Wide Identification, Characterization of the ORA (Olfactory Receptor Class A) Gene Family, and Potential Roles in Bile Acid and Pheromone Recognition in Mandarin Fish (Siniperca Chuatsi). Cells 2025, 14, 189. [Google Scholar] [CrossRef] [PubMed]
- Jia, W.; Ya, W.; Xu, L.; Hui, P.; Dian, J.; Jia, W.; Can, J.; Kumar, S.S.; Jian, S.; Xin, L.; et al. An Integrated Multi-Omics Approach Reveals Polymethoxylated Flavonoid Biosynthesis in Citrus Reticulata cv. Chachiensis. Nat. Commun. 2024, 15, 3991. [Google Scholar] [CrossRef]
Figure 1.
The heatmap of the subcellular localizations of CsANNs (A) and multiple sequence alignments of deduced amino acid sequences of ANN proteins from C. saxicola and Arabidopsis thaliana (B).
Figure 1.
The heatmap of the subcellular localizations of CsANNs (A) and multiple sequence alignments of deduced amino acid sequences of ANN proteins from C. saxicola and Arabidopsis thaliana (B).

Figure 2.
The phylogenetic tree of the ANN proteins. At: Arabidopsis thaliana; Bo: Brassica oleracea; Br: Brassica rapa; Ct: Corydalis tomentella; Cy: Corydalis yanhusuo; Hv: Hordeum vulgare; Os: Oryza sativa; Pt: Populus tremula x Populus.
Figure 2.
The phylogenetic tree of the ANN proteins. At: Arabidopsis thaliana; Bo: Brassica oleracea; Br: Brassica rapa; Ct: Corydalis tomentella; Cy: Corydalis yanhusuo; Hv: Hordeum vulgare; Os: Oryza sativa; Pt: Populus tremula x Populus.
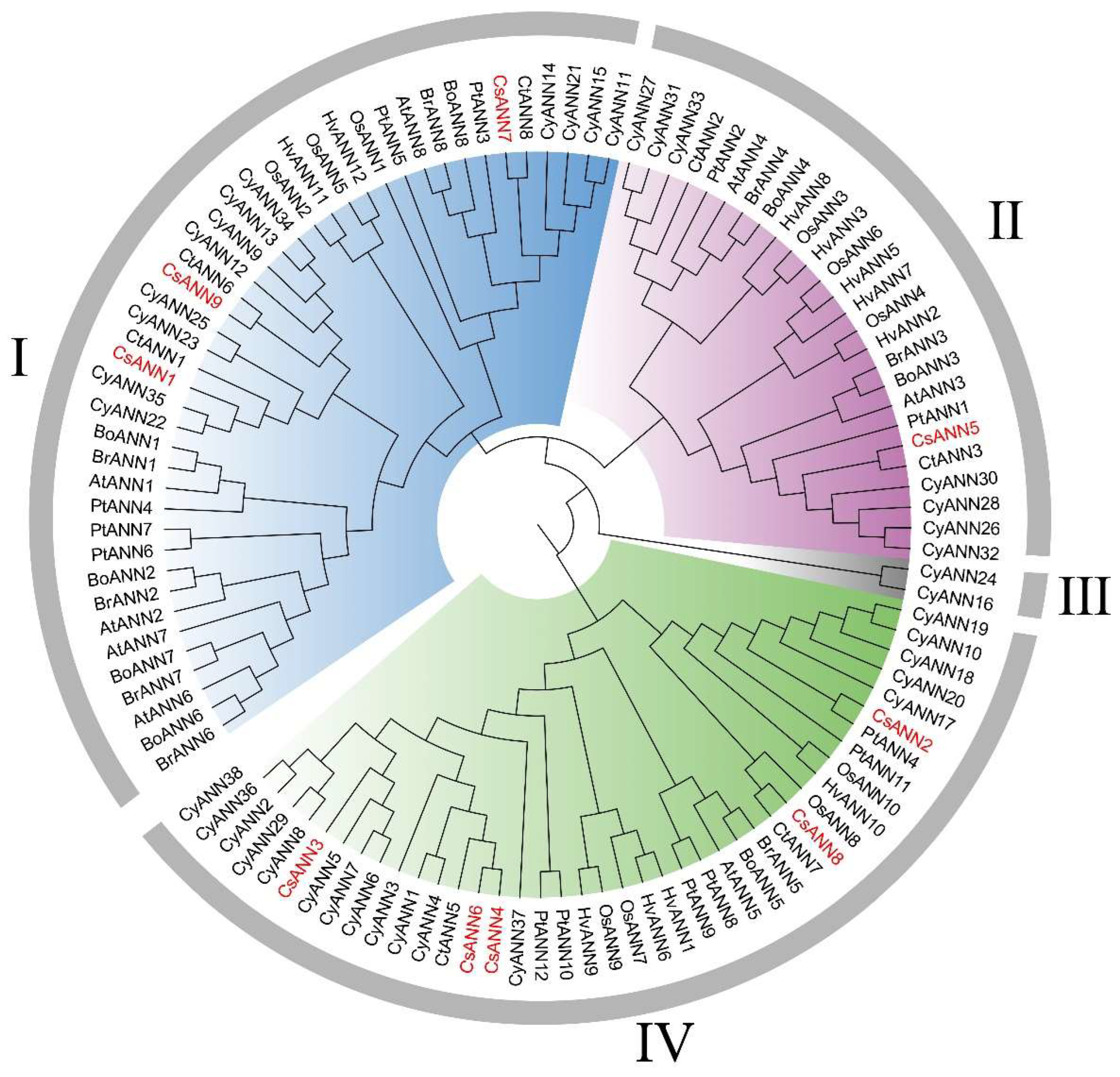
Figure 3.
The phylogenetic relationship, conserved motifs, ANN domains of CsANN proteins, and the intron and exon compositions of CsANN genes.
Figure 3.
The phylogenetic relationship, conserved motifs, ANN domains of CsANN proteins, and the intron and exon compositions of CsANN genes.
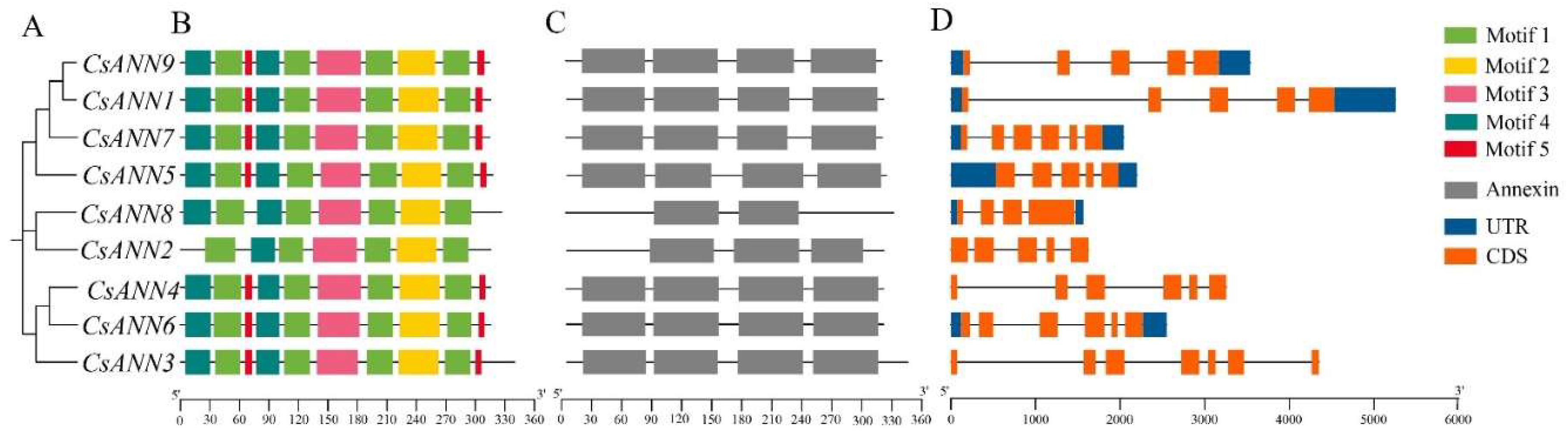
Figure 4.
Location distribution of CsANN genes on C. saxicola chromosomes (A) and synteny analysis of CsANN genes (B).
Figure 4.
Location distribution of CsANN genes on C. saxicola chromosomes (A) and synteny analysis of CsANN genes (B).
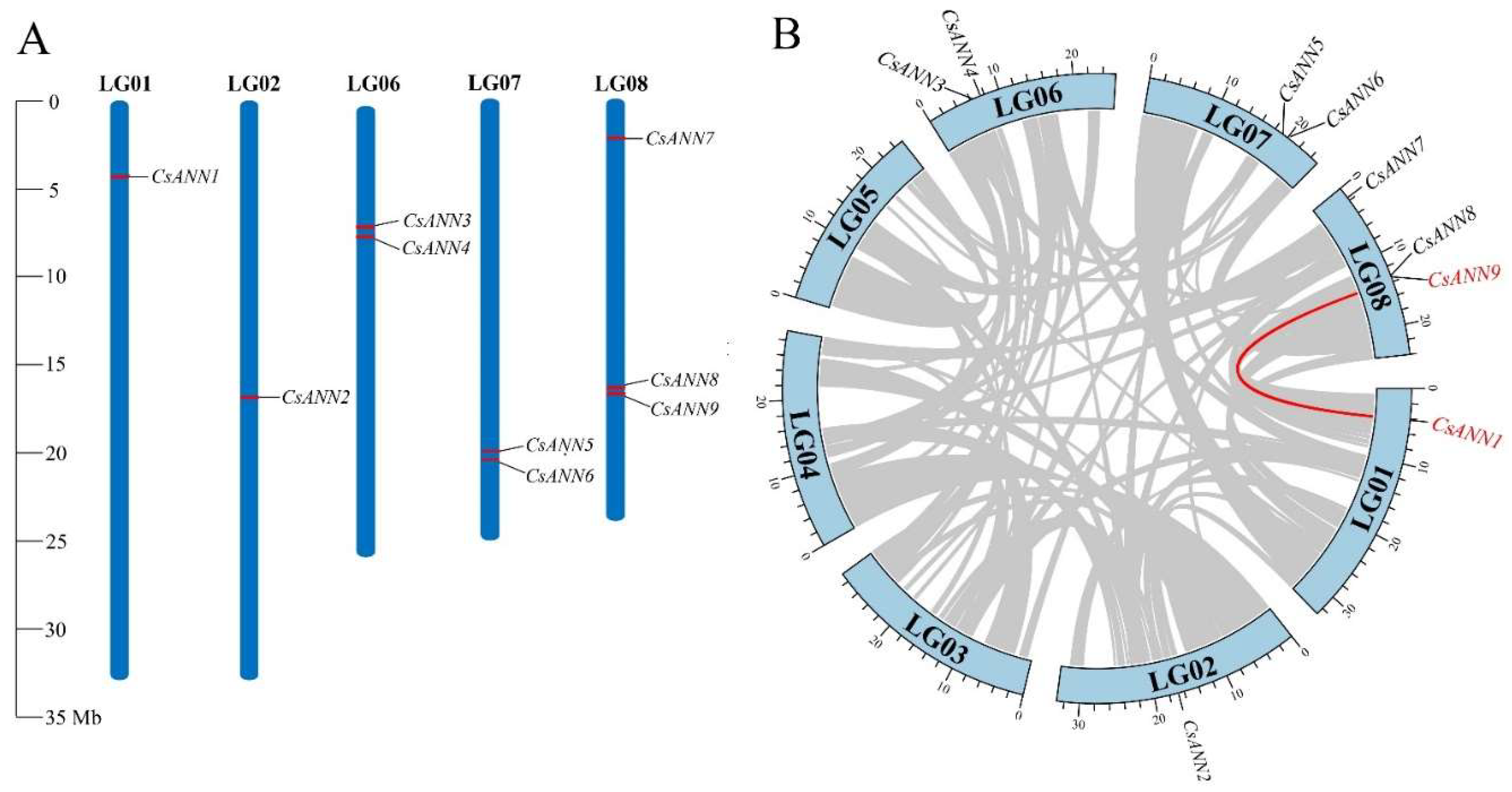
Figure 5.
Extra-genomic collinearity related to CsANN genes in A. thaliana, C. saxicola, C. tomentella, H. vulgare, O. sativa, and P. tremula.
Figure 5.
Extra-genomic collinearity related to CsANN genes in A. thaliana, C. saxicola, C. tomentella, H. vulgare, O. sativa, and P. tremula.
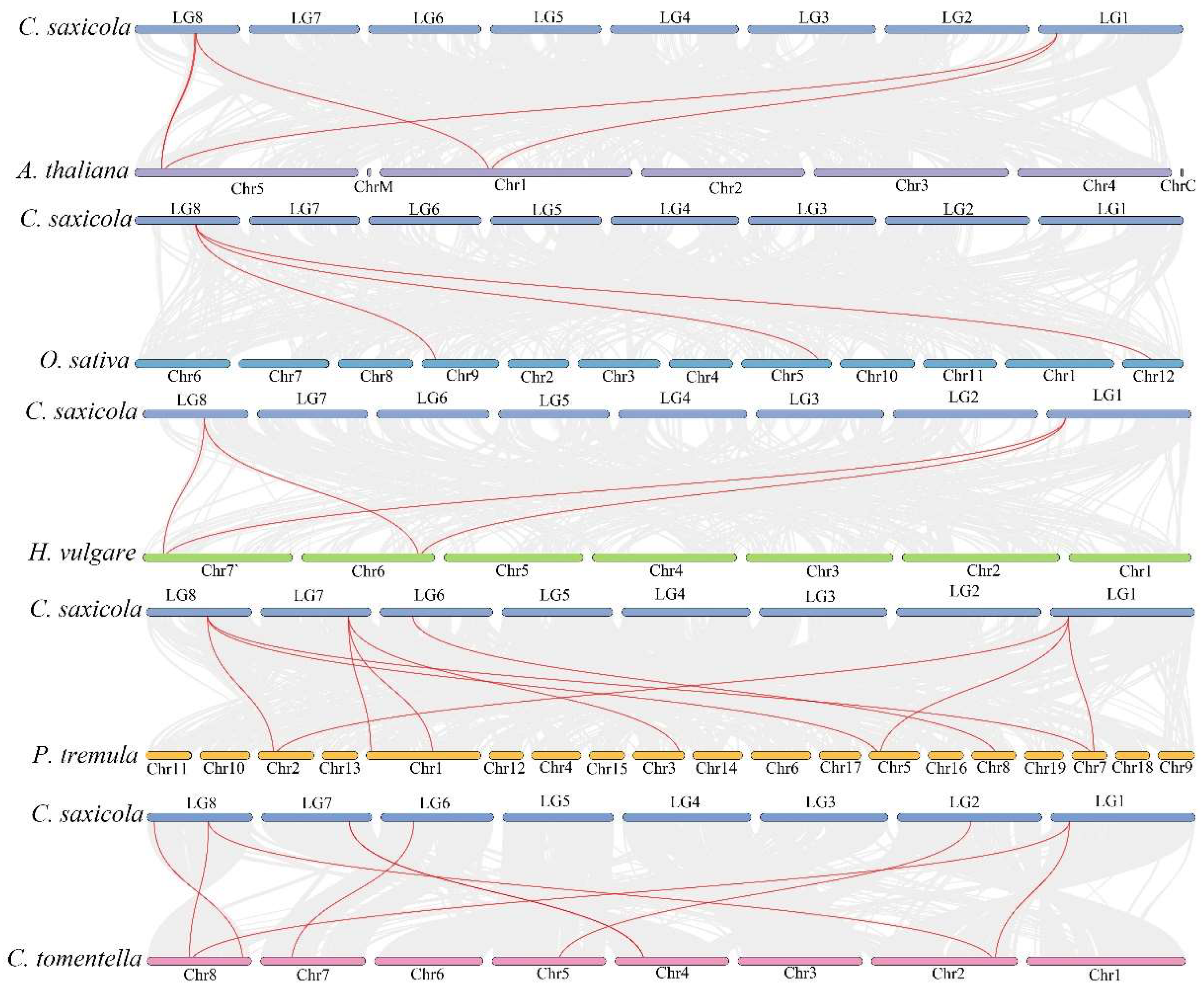
Figure 6.
The distribution of cis-acting elements in promoters of CsANNs.
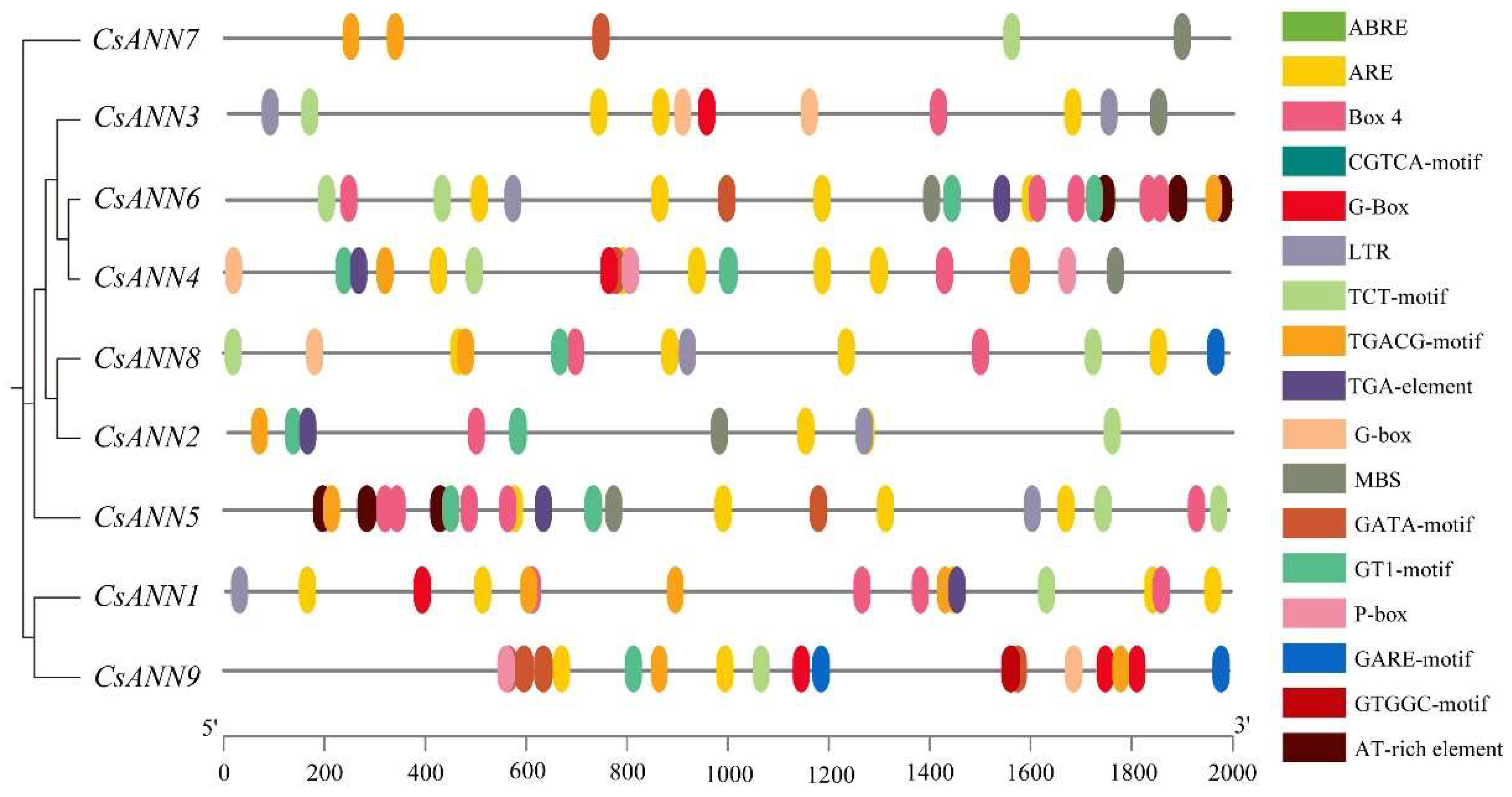
Figure 7.
Relative expression (RE) levels of CsANNs in different tissues of C. saxicola. R: roots, S: ems, LS: lateral stems, ML: mature leaves, YL: young leaves, FP: fruit pods, F: flowers.
Figure 7.
Relative expression (RE) levels of CsANNs in different tissues of C. saxicola. R: roots, S: ems, LS: lateral stems, ML: mature leaves, YL: young leaves, FP: fruit pods, F: flowers.
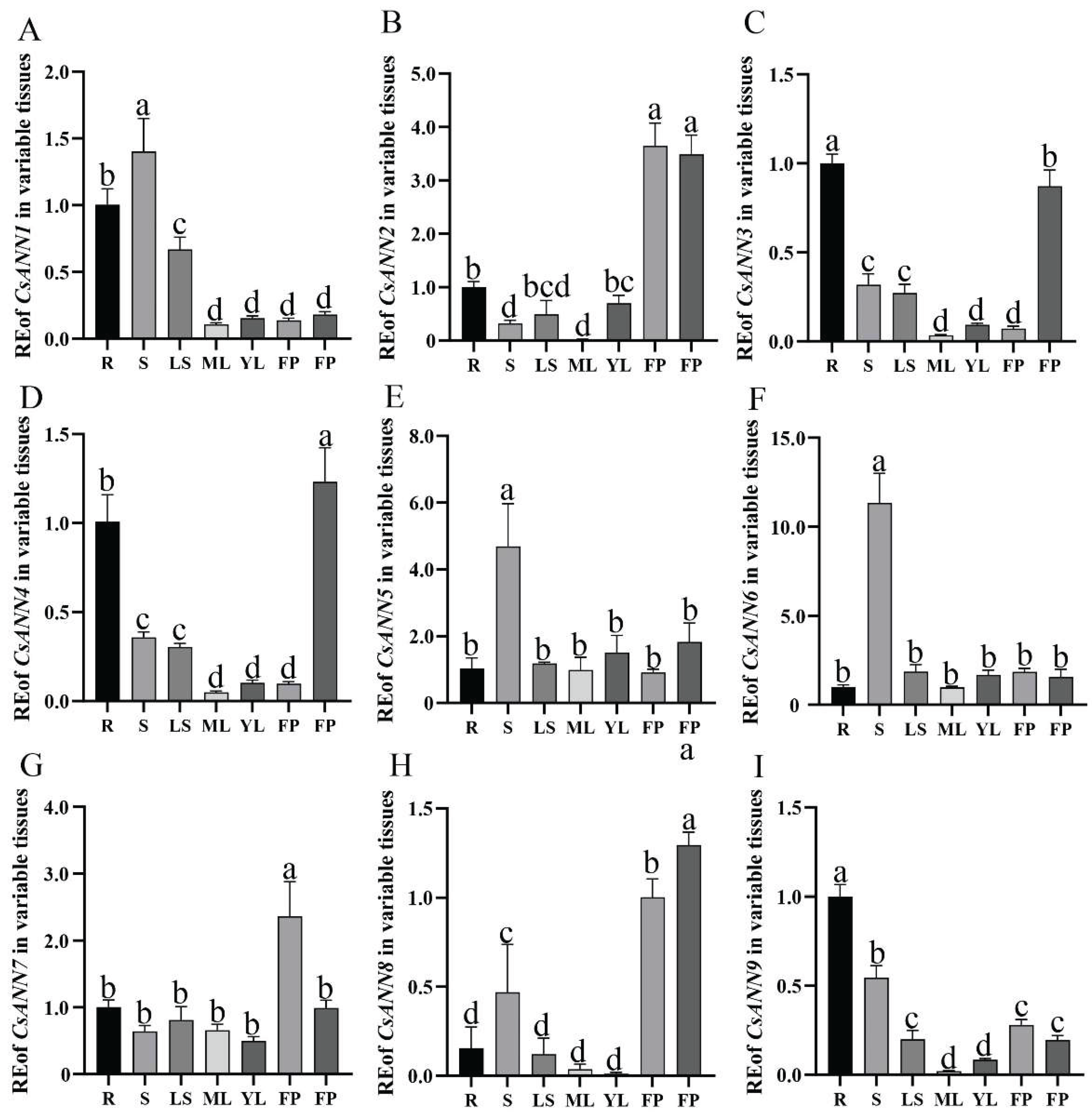
Figure 8.
The effects of exogenous CaCl2 treatments on C. saxicola seedlings. A-C: The calcium content, proline content, and soluble sugar content in mature leaves of C. saxicola under the treatments of different CaCl2 concentrations. D-F: The DHCA content in stems, mature leaves, and roots of C. saxicola under the treatments of different CaCl2 concentrations. G-O: Relative expression levels of CsANNs treated with different concentrations of CaCl2.
Figure 8.
The effects of exogenous CaCl2 treatments on C. saxicola seedlings. A-C: The calcium content, proline content, and soluble sugar content in mature leaves of C. saxicola under the treatments of different CaCl2 concentrations. D-F: The DHCA content in stems, mature leaves, and roots of C. saxicola under the treatments of different CaCl2 concentrations. G-O: Relative expression levels of CsANNs treated with different concentrations of CaCl2.
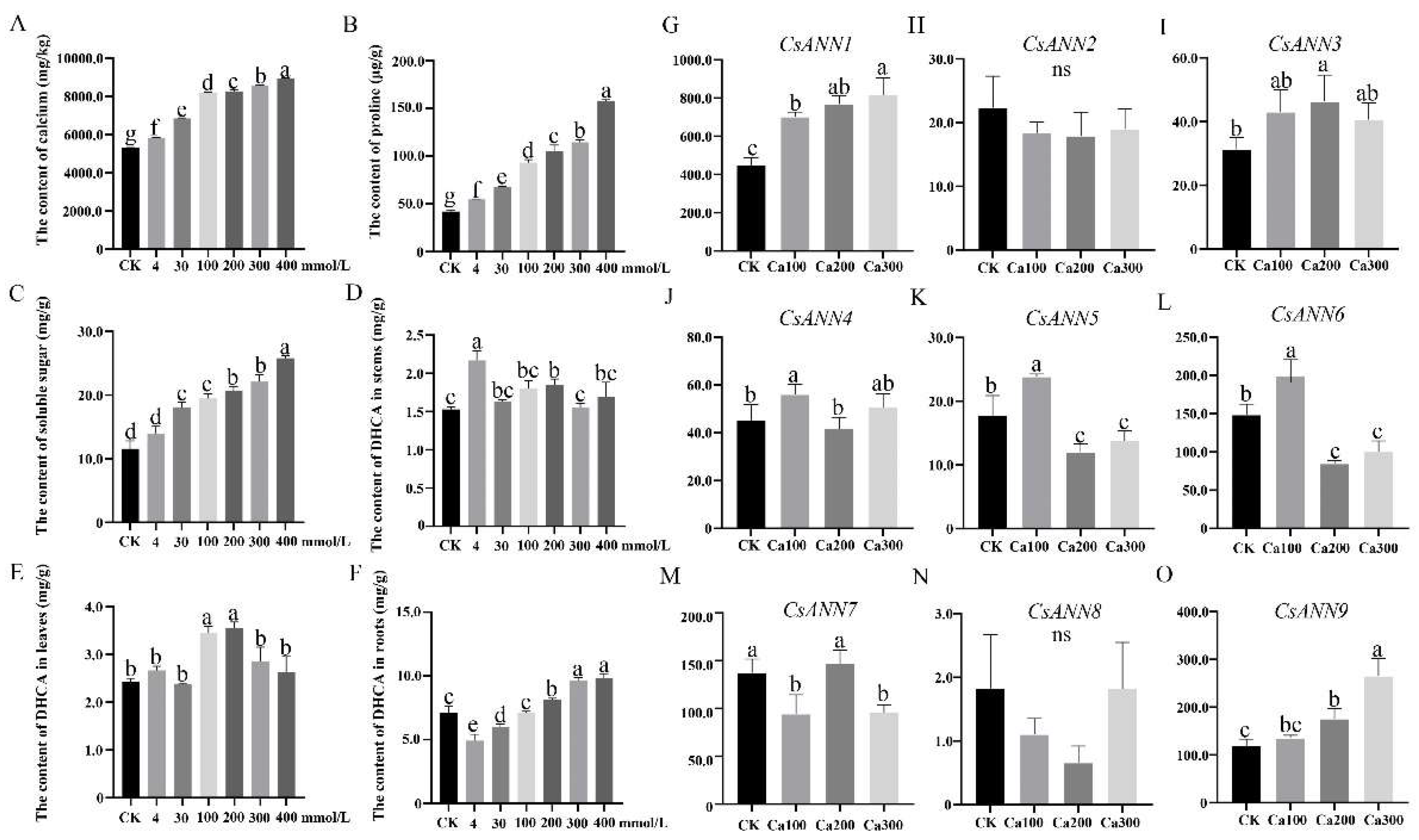
Figure 9.
The transient overexpression and functional analysis of CsANN1. (A) The schematic diagram of infiltration of C. saxicola leaves with CsANN-pCAMBIA1301 or EV. (B) Expression profiles of CsANN1 and other genes in C. saxicola leaves infiltrated with CsANN1-pCAMBIA1301 or EV. (C) Changes in BIA contents in C. saxicola leaves infiltrated with CsANN1-pCAMBIA1301 or EV. (D) Statistic analysis of BIA contents in C. saxicola leaves infiltrated with CsANN1-pCAMBIA1301 or EV.
Figure 9.
The transient overexpression and functional analysis of CsANN1. (A) The schematic diagram of infiltration of C. saxicola leaves with CsANN-pCAMBIA1301 or EV. (B) Expression profiles of CsANN1 and other genes in C. saxicola leaves infiltrated with CsANN1-pCAMBIA1301 or EV. (C) Changes in BIA contents in C. saxicola leaves infiltrated with CsANN1-pCAMBIA1301 or EV. (D) Statistic analysis of BIA contents in C. saxicola leaves infiltrated with CsANN1-pCAMBIA1301 or EV.
Figure 10.
Yeast two hybrid assay and protein interaction networks of the CsANNs. A: Transactivation assay. SD/-Trp: SD medium without Trp, SD/-Trp/+X/+A: SD medium without Trp supplemented with X-α-gal at a concentration of 40 ng/mL, AbA at a concentration of 100 ng/mL. B: Yeast two hybrid assay. DDO: SD medium without Trp and Leu; TDO: SD medium without Trp, Leu, and His; QDO: SD medium without Trp, Leu, His and Ade. C: The protein interaction networks of the CsANNs.
Figure 10.
Yeast two hybrid assay and protein interaction networks of the CsANNs. A: Transactivation assay. SD/-Trp: SD medium without Trp, SD/-Trp/+X/+A: SD medium without Trp supplemented with X-α-gal at a concentration of 40 ng/mL, AbA at a concentration of 100 ng/mL. B: Yeast two hybrid assay. DDO: SD medium without Trp and Leu; TDO: SD medium without Trp, Leu, and His; QDO: SD medium without Trp, Leu, His and Ade. C: The protein interaction networks of the CsANNs.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



