
Submitted:
15 May 2025
Posted:
15 May 2025
You are already at the latest version
Abstract
Uterine leiomyoma (uterine fibroids, UF) are benign myometrium tumors, that affect up to 70% of the female population and may lead to severe clinical symptoms. Despite the high prevalence, pathogenesis of UF is not understood and involves cytokines, steroid hormones and growth factors. Additionally, an increased deposition and remodelling of the extracellular matrix is characteristic for UF. Vitamin D seems to play a new role in UF. Interestingly, hypovitaminosis D correlates with a higher prevalence of myomas and the severity of the myomas. Administration of vitamin D in insufficient women (serum level <30 ng/mL) proved to restore the vitamin D status and reduce the mild symptoms of myomas. In addition, inflammatory processes may play also a role. In the past years it has become clear, that cessation of inflammation is an active process driven by a class of lipid mediator molecules called specialized pro-resolving mediators (SPM). Inadequate resolution of inflammation is related to several chronic inflammatory diseases and several studies have proven the crucial role of SPMs in improving these diseases. In this review, we will give the overview on processes involved in UF growth and will give an overview on the modern view regarding the concept of inflammation and the role of SPMs in resolution of inflammation, especially in chronic inflammatory diseases.
Keywords:
Keywords Uterine myoma
; inflammation
; vitamin D
; specialized pro-resolving mediators (SPMs)
1. Introduction
Uterine leiomyoma (myoma or uterine fibroids [UF]) are the most common nonmalignant pelvic tumors during the reproductive span [1] and up to 70% of women are affected. In most cases myoma do not cause symptoms and grow very slowly [2]. However, about 25% of the affected patients suffer from clinically relevant symptoms [2,3]. The incidence of UF is especially high among African-American women [4,5], who also suffer from more severe symptoms than the Caucasian population [6]. Typical symptoms include strong uterine bleeding, dysmenorrheal, dyspareunia and constipation. In addition, uterine leiomyoma can also be a cause of infertility or abortion [2,7]. Myoma were described as monoclonal tumors of the smooth musculature. However, recent research on MED12(+) and MED12(-) fibroid variants presents them by single cell RNA sequencing as rather heterogeneous cell types [8]. The tumors, although benign, are characterized by abnormal vascularization. Initiation and progression is driven by a variety of factors including local hypoxia, gonadal steroids as well as locally expressed angiogenic growth factors including EGF (epidermal growth factor) and VEGF (vascular endothelial growth factor). Cytokines and chemokines that are also involved in the pathogenesis not only promote the proliferation of cells but also the deposition of ECM [9], which is the main component of UF and an important factor in its development [1,9]. There also seems to be a link between myoma formation and genetic aberrations, as cytogenetic tests showed abnormalities in 40% of the samples tested: 65% overexpressed the HMGA2 (high mobility group AT-hook 2) gene. Furthermore, 70% of leiomyoma samples carried mutations in the subunit of a transcription regulator [10,11]. Estrogens and progesterone (P4) play a role in the initiation and propagation of UF. Progesterone receptors A and B (PR-A and PR-B) expression is induced by estrogen signaling at the ER. This, in turn, increases the effectiveness of progesterone. As a result, the progesterone-induced increase in mitotic activity of UF cells increases [12,13]. Additional effects of growth factor regulation or microRNA expression and ECM accumulation seem to be mediated by P4 [1,13,14,15]. Progesterone is known to induce cell proliferation in UF tissue, while reducing apoptosis. Its activity is mediated by growth factors such as TGFβ (transforming growth factor β), although the action of these mediators would not occur in the absence of P4 [16]. In addition, the proliferation and growth of myoma may be induced and promoted by local and systemic inflammatory processes. Although the question is controversially debated [10,17,18], it is assumed that chronic inflammation promotes the pathogenesis of leimyoma in the following review. Tumor types of various etiology, including uterus fibroids, are associated with an inflammatory microenvironment, which is induced by pro-inflammatory cytokines produced by immune cells and undifferentiated cells [17]. Interestingly, pro-inflammatory cytokines (such as IL-6, IL-1 and tumor necrosis factor alpha) were shown to increase estrogen synthesis via modulation of aromatase, estradiol 17-β-dehydrogenase and estrone sulfatase activity. Inhibition of the synthesis of prostaglandin E2 was also demonstrated to exert an influence on this balance [19,20]. As mentioned above, ECM remodeling and accumulation is characteristic of inflammatory processes and is a key factor in the development of UF. It is associated with excessive collagen accumulation and a reduced number of micro vessels, which in turn promote degenerative atrophic changes of the myometrium with cellular senescence and involution. The collagenized areas exhibit a hyaline and hypocellular morphology [21]. ECM generation is driven by cytokine- and growth factor- signaling, on the other hand it acts as a source for the continuous availability of these molecules as it exerts a stabilizing effect. Immune cells, especially monocytes and macrophages, responding to these inflammatory signals stimulate fibrotic changes of the tumor tissue. The TGF-β is one of the major players in these processes [22,23], but also TNF-α and the granulocyte-macrophage colony- stimulating factor, as well as interleukins (IL) 1, IL6, IL11, IL13, IL15 and IL13 were found to be involved in the pathogenesis of UF. Drugs, that are commonly used in treatment of UF may interfere with these inflammatory processes and it may by worthwile to gain more understanding about these interactions [7].
2. Role of Vitamin D
Role of Vitamin D Lima et al. could show recently [24] a reduced Vitamin D Receptor (VDR) expression in leiomyoma tissue compared with myometrial tissue, which can be associated with the pathogenesis and development of human uterine leiomyomatosis. The cause of the reduction in the expression of these receptors is still unknown and it is also not clear whether it is an event that occurs in parallel to the onset or progression of uterine leiomyomas. Furthermore Halder et al. [25] analyzed the influence of vitamin D when studying the effect of 1,25-dihydroxyvitamin D3 on fibrosis-related protein expression in TGF-3 induced uterine leiomyoma cells in vitro. Myoma cells were treated with TGF-3 with or without vitamin D. They identified that TGF-3 induced the expression of fibronectin and collagen protein type 1 in myoma cells, which was suppressed by vitamin D and considered as an antifibrotic factor in the treatment of benign uterine myomas. Halder et al. in subsequent study [26] investigated the risk of benign uterine tumors in regard to VDR protein and determined the biological function of 1.25 (OH) 2D3 in the regulation of proteins related to the extracellular matrix, which is essential in the formation of leiomyomas. They identified reduced VDR levels in more than 60 % of the uterine tumors analyzed compared to the adjacent myometrium. In fact, the levels of VDR in the myoma uteri samples were significantly lower than the levels in the adjacent myometrial samples, similar to the results of the present study. Al-Hendy et al. [27], who investigated the role of 1.25 (OH) 2D3 in the expression of sex steroid receptors in leiomyoma cells, realized that the deregulation of steroid hormones and their receptors could be a starting point for myoma growth since 1.25 (OH) 2D3 VDR expression acts as an antiesterogenic agent in these cells. They also showed a significant decrease in estrogenic re- ceptor levels in leiomyoma cells treated with 1.25 (OH) 2D3 and analyzed for receptor expression and location. In contrast, 1.25 (OH) 2D3 induced the expression of its own VDR, suggesting that 1.25 (OH) 2D3 acts as an antagonist of hormone receptors with antiestrogenic and antiprogesteronic functions. Paffoni et al. [28] analyzed serum levels of vitamin D in women with myoma and recognized that the vitamin D concentration was significantly lower in women with myomas compared to women in the control group (11.1 and 18.0 ng/ml, respectively; p < 0.010 and OR = 2.2). Similar results were obtained by Baird et al. [29], who assessed vitamin D and the risk of uterine myomas and found that women with the sufficient vitamin D had an estimated 32 % reduction in the incidence of myomas com- pared with those with insufficient vitamin D. Sabry et al. [30] studied whether the low serum levels of vitamin D were associated with the increased risk and occurrence of uterine myomas and found that reduced serum levels of 25-(OH) vitamin D were significantly associated with the occurrence of myomas. A statistically significant inverse correlation was also observed between the serum levels of 25-(OH) vitamin D and the total leiomyoma volume within the case cohort. The above described studies indicate that the loss of vitamin D functions due to the reduction of vitamin D3 levels and/ or reduced expression of VDR may be associated with the growth and development of different types of neoplastic lesions. It reinforces the hypothesis that low VDR expression may be associated with the growth and development of myomas, presenting itself as an important biomarker in this pathology. Vitamin D is believed to regulate cell proliferation and differentiation, reduce angiogenesis, and stimulate apoptosis. Nowadays hypovitaminosis D is supposed to be a major risk factor in the development of fibroids. In many studies vitamin D appears to be a powerful factor against UFs, resulting in inhibition of tumor cell division and a significant reduction in its size, however, the exact role of this compound and its receptor in the pathophysiology of myomas is not fully elucidated. Based on available studies vitamin D and its analogs seem to be promising, effective and low-cost compounds in the management of myomas and their clinical symptoms and the anti-tumor activities of vitamin D play an important role in uterine myoma biology.
3. Inflammatory Processes and the Role of Lipid Acid-Derived Mediators
Acute inflammation occurs in response to a microbial infection or injury. The initiated processes are necessary to render pathogens innocuous, remove cell debris and restore affected tissue. Eicosanoid lipid mediator (LM) molecules that are synthesized from the ω-6 poly-unsaturated fatty acid (PUFA) arachidonic acid (AA) are characteristic pro-inflammatory signaling molecules. These prostanoids comprise the prostaglandins (PG), Leukotrienes (LT) and thromboxanes (TX) are synthesized via the enzymes cycloox-agenoses 1 and 2 (Cox-1/2) by cells of the innate immune system (granulocytes, macro- phages), which are quickly recruited to a site of injury or infection [31]. Further pro-inflammatory cytokines including TNFα, IL-1 and Il-6 are secreted by mast cells and classically activated M1 macrophages contribute to the inflammatory response and its characteristic symptoms (heat, swelling, redness, pain). Prostanoids and cytokines also attract neutrophils and monocytes, that migrate towards the site of event and enter the af-fected tissue, thereby further promoting the inflammatory response of the organism [33] Rapid and efficient initiation of inflammation is vital for the organism, but timely cessation of the processes is of equal importance, as excessive inflammation can lead to phenomena such as cytokine storm or sepsis, which is an life-threatening event [34]. Chronic inflammatory pathologies like cardiovascular disease (CVD), diabetes or autoimmune disorders are further examples for inadequate cessation of inflammation [35,36]. Resolu- tion of inflammation is an active process that is driven by so-called specialized pro-resolving lipid mediator molecules (SPM), which are able to initiate and drive resolu- tion in animal models [37,38]. Based on their chemical structure and biosynthetic path- ways, they are clustered in four families: the resolvins (Rvs), protectins (PD), lipoxins (LXs) and maresins (Mar) [37,38,39]. Rv, PD, LX derive from the ω-3 PUFAs EPA (Eicosa- pentanenoic acid) and DHA (docosahexaenoic acid) and their biosynthesis involves cer- tain lipoxygenases, but also the COX-enzymes. They are generated via the hydroxylated precursors 18HpETE, 17-HpDHA and 14-HpDHA. ω-6 PUFA AA is the precursor for LX [39,40]. In Figure 1 an overview on the biosynthetic pathways is given. Notably, Aspirin, which inhibits the COX-enzymes irreversibly, blocks the synthesis of prostanoids by mod-ifying the catalytic domains of COX. However, their capacity to catalyse the synthesis of the SPM precursors 18HpETE, 17-HpDHA and 14-HpDHA is not abolished. The emergin SPMs are referred to as aspirin-triggered SPMs (AT-SPMs) and are often used in experiments [38,39,42,43].
Eicosanoid lipid mediator LM) molecules (LT, TX, PG etc) from AA, and pro-inflammatory cyto-kines that induce the influx of PMN and moncytes and lead to the classical signs of acute in-flammation (calor, rubor, tumor, dolor). At the very beginning, eicosanoids initiate the LM class switch resulting in the production of SPMs from omega-3-PUFAs (See Fig. 1 and text). SPMs are critical for the resolution of the inflammation. LM: Lipid mediator; LT: Leukotriene, TX: Thromboxane, PG: Prostaglandins; PUFA: poly-unsaturated fatty acid, SPM Specialized pro-resolving mediators (modified from Serhan [40]).
4. SPMs are Essential for Resolution of Inflammation
SPMs are crucial for the resolution of inflammation. How the resolution of inflammation is achieved is of the utmost importance. There are several things that must be achieved: recruitment of neutrophiles must stop, dead cells must be eliminated, and macrophages need to switch towards the resolving M2 macrophages [44]. These and other key events in obtaining resolution are driven by SPMs, as reviewed elsewhere in detail [40,43,45,46]. In short, SPMs are necessary for stopping the recruitment of neutrophiles and triggering apoptosis cell phagocytosis. Further, they play a role in the downregulation of pro-inflammatory cytokines, such as TNF-α, IL-6, IL-8, IL-12 and PAF (platelet-activating factor)- and prostaglandin-production. In fact, SPMs are involved in clearing the infection site and in tissue re-generation, since they promote efferocytosis and wound healing. In the process, these resolutive events are initiated at the same time as the inflammatory process starts, which is reflected by the interlinkage of prostaglandin-synthesis with SPM biosynthesis. PGE2 and PGD2 are required to induce type1- lipoxygenases, the key enzymes in the SPM bio-synthetic pathways which produce LXs, Rvs, and PD2 [47]. Consequently, when PG-synthesis is inhibited the lipid mediator class switch from pro-inflammatory to pro-resolutive is interfered with, which may lead to delayed resolution [48,49]. To sum up, the inflammation is always started with its active resolution, since alfa signal omega in the whole cascade signaling pathways [47]. In this way a new perception of inflammation is shown in Figure 2.
Extensive research into SPMs led to detailed knowledge concerning the structure, biosynthetic pathways and function of SPMs and different options for the outcome of inflammatory responses are dealt with in Serhan et al [37,39,40]. Figure 3 shows the current view. Should the resolution of inflammation not take place adequately and remain “switched on”, a continuous production of pro-inflammatory signaling molecules and thus a state of chronic inflammation may be established. In addition to their activity in promoting the resolution of inflammation, the SPMs represent a link to the adaptive immune response. For example, lipoxin LX3 also activates natural killer cells (NKC) [50]. About the differentiation of CD4+-T-cells it was shown that this is also modulated by resolvins RvD1 and RvD2 and maresin Mar1 [51].
5. The Significance of SPMs in Chronic Inflammatory Diseases
Animal experiments have demonstrated the role of SPMs in prevention or improvement of chronic inflammatory diseases. In the case of periodontitis, pathogenesis is based on an inflammatory process resulting from an infection with P. gingivalis able to be mimicked in air pouche mouse models. In these experiments increased recruitment of neutrophils and upregulation of COX-2 enzymes was observed, which was decreased by supplementation with lipoxin LXA analogues. [52]. A further model in mice under the influence of 12/15 lipoxygenase demonstrated the role of the SPMs LXA, RvD and RvD in the pathogenesis of atherosclerosis, which is considered a chronic inflammatory disease. [53]. In mice under the influence of 12/15 lipoxygenase the increased production of RvD, PD and 17-HpDHA resulted in being protective, as the development of atherosclerosis was reduced in comparison to wild type mice. The anti-atherogenic activity of LXA, PD and RvD derived from different processes such as reduced expression of endothelial adhesion molecules and diminished secretion of cytokines. Notably, this mouse model also demonstrated the role of nutrition in the pathogenesis of atherosclerosis, since a standard high-fat western diet rendered the trans-genic mice as susceptible to atherosclerosis as the wild type animals [53,54]. In a rat model for arthritis, a further common chronic inflammatory disease, resolvin RvD, as well as its precursor metabolite (17-HDHA) led to a greater reduction in pain and tissue damage compared with medication with steroids [55]. Fibrosis, which is also a characteristic feature of uterine leiomyoma [1], may also be considered a consequence of an inappropriate resolution of inflammation. For pulmonary fibrosis, the experimental LX4-analogues (AT-LX4) that were administered exogenously, reduced fibrosis in animal models [56]. LXA4 and benzo-LXA4, an analogue, also reduced the extent of fibrotic changes in kidney in a rat model of early renal fibrosis [57] and for the resolvin RvE1, antifibrotic effects were demonstrated in a mouse model of obstructed kidney [58]. Although the role of SPMs in UF development has not been elucidated yet, there are some shared features with other chronic inflammatory diseases in which the role of in-adequate resolution has been demonstrated. Further research in this area may lead to new treatment options.
6. Conclusion
The pathogenesis of uterine fibroids has not been fully clarified yet. Cytokine-, growth factor-, and steroid hormone signaling together with inflammatory processes may be involved. Vitamin D acts on human reproduction not only by calcium homeostasis, but also due to its paramount importance as direct regulator of the aromatase gene expression. Accumulating evidence suggests that VDR-bound 1,25(OH)2D3 acts as a transcription factor to regulate the expression of the CYP19 gene. This gen encodes the fat and ovarian tissue aromatase. Hence this mechanism encodes the ostrogen regulation [58]. Vitamin D is also able to stimulate the production of progesterone, different estrogens and substances like the insulin like growth factor binding protein 1 (IGFBP-1) especially in cultured human ovarian cells. All together these different modes of vitamin D influence hormonal dependent benign tissues like e.g uterine fibroids having a modulation effect on growth and differentiation. Other estrogen dependent diseases like endometriosis seem to be also modulated by Vitamin D. Activated CD4 and CD8 cells, express widely vitamin D receptors and both the activated and metabolized enzymes, 1-a -hydroxylase and 24-hydroxylase. This induces a vitamin D local expression and has a negative effect on the potential regression of endometriosis implants. In addition, in a large prospective cohort study, a high plasma 25(OH)D level was associated with a lower risk of endometriosis [60]. Finally, vitamin D showed inhibitory capacities on the expression of ER and PR in a dose-dependent manner in leiomyoma cells. [61]. As has been described during the last years, active resolution of inflammation, which is mainly driven by specialized pro-resolving lipid mediator molecules (SPMs), is crucial for the adequate cessation of acute and chronic inflammation, as demonstrated for diseases like periodontitis, atherosclerosis or diabetes. Therefore, further research on the role of inflammation in general and SPMs might yield further important knowledge on uterine fibroids.
Author Contributions
Pedro-Antonio Regidor: Responsible for the concept of resolution of inflammation in PCOS. Manuela Mayr: Responsible for graphic design. Fernando Gonzalez Santos: Responsible for the development of SPMs use in humans and therfeore for basic scientific data. Beatriz Lazcoz Calvo: Responsible for the scientific background. Rocio Gutierrez: Responsible for literature research and writing. Jose Miguel Rizo: Responsible for PCOS chapter and data aquisition
Funding
Not applicable.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study.
Acknowledgments
Not applicable.
Conflicts of Interest
Pedro-Antonio Regidor and Beatriz Lazcoz are employees of Exeltis Healthcare. Manuela Mayr is emploeyee of Exeltis Germany. Fernando Gonzalez Santos declares no conflict of interest. Rocio Gutierrez and Jose Miguel Rizo are employees of Chemo OTC Spain.
References
- Islam, M.S. , et al., Extracellular matrix in uterine leiomyoma pathogenesis: a potential target for future therapeutics. Hum Reprod Update, 2018. 24(1): p. 59-85.
- Buttram, V.C., Jr. and R.C. Reiter, Uterine leiomyomata: etiology, symptomatology, and management. Fertil Steril, 1981. 36(4): p. 433-45.
- Cramer, S.F. and A. Patel, The frequency of uterine leiomyomas. Am J Clin Pathol, 1990. 94(4): p. 435-8.
- Marshall, L.M. , et al., Variation in the incidence of uterine leiomyoma among premenopausal women by age and race. Obstet Gynecol, 1997. 90(6): p. 967-73.
- Baird, D.D. , et al., High cumulative incidence of uterine leiomyoma in black and white women: ultrasound evidence. Am J Obstet Gynecol, 2003. 188(1): p. 100-7.
- Kjerulff, K.H. , et al., Uterine leiomyomas. Racial differences in severity, symptoms and age at diagnosis. J Reprod Med, 1996. 41(7): p. 483-90.
- Szydlowska, I. , et al., Markers of Inflammation and Vascular Parameters in Selective Progesterone Receptor Modulator (Ulipristal Acetate)-Treated Uterine Fibroids. J Clin Med, 2021. 10(16).
- Goad, J. , et al., Single Cell atlas of uterine myometrium and leiomyomas reveals diverse and novel cell types of non-monoclonal origin. bioRxiv, 2020: p. 2020.12.21.402313.
- Tal, R. and J.H. Segars, The role of angiogenic factors in fibroid pathogenesis: potential implications for future therapy. Hum Reprod Update, 2014. 20(2): p. 194-216.
- Ciavattini, A. , et al., Uterine fibroids: pathogenesis and interactions with endometrium and endomyometrial junction. Obstet Gynecol Int, 2013. 2013: p. 173184.
- Galindo, L.J. , et al., HMGA2 and MED12 alterations frequently co-occur in uterine leiomyomas. Gynecol Oncol, 2018. 150(3): p. 562-568.
- Grings, A.O. , et al., Protein expression of estrogen receptors alpha and beta and aromatase in myometrium and uterine leiomyoma. Gynecol Obstet Invest, 2012. 73(2): p. 113-7.
- Ishikawa, H. , et al., Progesterone is essential for maintenance and growth of uterine leiomyoma. Endocrinology, 2010. 151(6): p. 2433-42.
- Kawaguchi, K. , et al., Mitotic activity in uterine leiomyomas during the menstrual cycle. Am J Obstet Gynecol, 1989. 160(3): p. 637-41.
- Islam, M.S. , et al., Complex networks of multiple factors in the pathogenesis of uterine leiomyoma. Fertil Steril, 2013. 100(1): p. 178-93.
- Modugno, F. , et al., Inflammation and endometrial cancer: a hypothesis. Cancer Epidemiol Biomarkers Prev, 2005. 14(12): p. 2840-7.
- Orciani, M. , et al., Chronic Inflammation May Enhance Leiomyoma Development by the Involvement of Progenitor Cells. Stem Cells Int, 2018. 2018: p. 1716246.
- Protic, O. , et al., Possible involvement of inflammatory/reparative processes in the development of uterine fibroids. Cell Tissue Res, 2016. 364(2): p. 415-27.
- Macdiarmid, F. , et al., Stimulation of aromatase activity in breast fibroblasts by tumor necrosis factor alpha. Mol Cell Endocrinol, 1994. 106(1-2): p. 17-21.
- Purohit, A., S. P. Newman, and M.J. Reed, The role of cytokines in regulating estrogen synthesis: implications for the etiology of breast cancer. Breast Cancer Res, 2002. 4(2): p. 65-9.
- Leppert, P.C. et al., Summary of the Proceedings of the Basic Science of Uterine Fibroids Meeting: New Developments February 28, 2020. F S Sci, 2021. 2(1): p. 88-100.
- Ciebiera, M. , et al., The Evolving Role of Natural Compounds in the Medical Treatment of Uterine Fibroids. J Clin Med, 2020. 9(5).
- Zannotti, A. , et al., Macrophages and Immune Responses in Uterine Fibroids. Cells, 2021. 10(5).
- Lima, M.S.O. , et al., Evaluation of vitamin D receptor expression in uterine leiomyoma and nonneoplastic myometrial tissue: a cross-sectional controlled study. Reprod Biol Endocrinol, 2021. 19(1): p. 67.
- Halder, S.K., J. S. Goodwin, and A. Al-Hendy, 1,25-Dihydroxyvitamin D3 reduces TGF-beta3-induced fibrosis-related gene expression in human uterine leiomyoma cells. J Clin Endocrinol Metab, 2011. 96(4): p. E754-62.
- Halder, S.K., K. G. Osteen, and A. Al-Hendy, 1,25-dihydroxyvitamin d3 reduces extracellular matrix-associated protein expression in human uterine fibroid cells. Biol Reprod, 2013. 89(6): p. 150.
- Al-Hendy, A. , et al., 1,25-dihydroxyvitamin D3 regulates expression of sex steroid receptors in human uterine fibroid cells. J Clin Endocrinol Metab, 2015. 100(4): p. E572-82.
- Paffoni, A. , et al., Vitamin D status in women with uterine leiomyomas. J Clin Endocrinol Metab, 2013. 98(8): p. E1374-8.
- Baird, D.D. , et al., Vitamin d and the risk of uterine fibroids. Epidemiology, 2013. 24(3): p. 447-53.
- Sabry, M. , et al., Serum vitamin D3 level inversely correlates with uterine fibroid volume in different ethnic groups: a cross-sectional observational study. Int J Womens Health, 2013. 5: p. 93-100.
- Chiurchiu, V., A. Leuti, and M. Maccarrone, Bioactive Lipids and Chronic Inflammation: Managing the Fire Within. Front Immunol, 2018. 9: p. 38.
- Flower, R.J. , Prostaglandins, bioassay and inflammation. Br J Pharmacol, 2006. 147 Suppl 1: p. S182-92.
- Samuelsson, B. , Role of basic science in the development of new medicines: examples from the eicosanoid field. J Biol Chem, 2012. 287(13): p. 10070-10080.
- Hotchkiss, R.S. , et al., Sepsis and septic shock. Nat Rev Dis Primers, 2016. 2: p. 16045.
- Furman, D. , et al., Chronic inflammation in the etiology of disease across the life span. Nat Med, 2019. 25(12): p. 1822-1832.
- Nathan, C. and A. Ding, Nonresolving inflammation. Cell, 2010. 140(6): p. 871-82.
- Serhan, C.N. , Pro-resolving lipid mediators are leads for resolution physiology. Nature, 2014. 510(7503): p. 92-101.
- Serhan, C.N. , et al., Novel functional sets of lipid-derived mediators with antiinflammatory actions generated from omega-3 fatty acids via cyclooxygenase 2-nonsteroidal antiinflammatory drugs and transcellular processing. J Exp Med, 2000. 192(8): p. 1197-204.
- Serhan, C.N., N. Chiang, and J. Dalli, The resolution code of acute inflammation: Novel pro-resolving lipid mediators in resolution. Semin Immunol, 2015. 27(3): p. 200-15.
- Serhan, C.N. , Treating inflammation and infection in the 21st century: new hints from decoding resolution mediators and mechanisms. FASEB J, 2017. 31(4): p. 1273-1288.
- Regidor, P.A. , et al., Omega-3 long chain fatty acids and their metabolites in pregnancy outcomes for the modulation of maternal inflammatory- associated causes of preterm delivery, chorioamnionitis and preeclampsia. F1000Research 2024, 13:882.
- Serhan, C.N. , et al., Novel proresolving aspirin-triggered DHA pathway. Chem Biol, 2011. 18(8): p. 976-87.
- Serhan, C.N. , et al., Resolvins: a family of bioactive products of omega-3 fatty acid transformation circuits initiated by aspirin treatment that counter proinflammation signals. J Exp Med, 2002. 196(8): p. 1025-37.
- Schett, G. and M.F. Neurath, Resolution of chronic inflammatory disease: universal and tissue-specific concepts. Nat Commun, 2018. 9(1): p. 3261.
- Serhan, C.N. and N. Chiang, Resolution phase lipid mediators of inflammation: agonists of resolution. Curr Opin Pharmacol, 2013. 13(4): p. 632-40.
- Serhan, C.N. and B.D. Levy, Resolvins in inflammation: emergence of the pro-resolving superfamily of mediators. J Clin Invest, 2018. 128(7): p. 2657-2669.
- Serhan, C.N. and J. Savill, Resolution of inflammation: the beginning programs the end. Nat Immunol, 2005. 6(12): p. 1191-7.
- Bandeira-Melo, C. , et al., Cyclooxygenase-2-derived prostaglandin E2 and lipoxin A4 accelerate resolution of allergic edema in Angiostrongylus costaricensis-infected rats: relationship with concurrent eosinophilia. J Immunol, 2000. 164(2): p. 1029-36.
- Levy, B.D. , et al., Lipid mediator class switching during acute inflammation: signals in resolution. Nat Immunol, 2001. 2(7): p. 612-9.
- Barnig, C. , et al., Lipoxin A4 regulates natural killer cell and type 2 innate lymphoid cell activation in asthma. Sci Transl Med, 2013. 5(174): p. 174ra26.
- Chiurchiu, V. , et al., Proresolving lipid mediators resolvin D1, resolvin D2, and maresin 1 are critical in modulating T cell responses. Sci Transl Med, 2016. 8(353): p. 353ra111.
- Serhan, C.N. , et al., Reduced inflammation and tissue damage in transgenic rabbits overexpressing 15-lipoxygenase and endogenous anti-inflammatory lipid mediators. J Immunol, 2003. 171(12): p. 6856-65.
- Merched, A.J. , et al., Atherosclerosis: evidence for impairment of resolution of vascular inflammation governed by specific lipid mediators. FASEB J, 2008. 22(10): p. 3595-606.
- Merched, A.J., C. N. Serhan, and L. Chan, Nutrigenetic disruption of inflammation-resolution homeostasis and atherogenesis. J Nutrigenet Nutrigenomics, 2011. 4(1): p. 12-24.
- Lima-Garcia, J.F. , et al., The precursor of resolvin D series and aspirin-triggered resolvin D1 display anti-hyperalgesic properties in adjuvant-induced arthritis in rats. Br J Pharmacol, 2011. 164(2): p. 278-93.
- Martins, V. , et al., ATLa, an aspirin-triggered lipoxin A4 synthetic analog, prevents the inflammatory and fibrotic effects of bleomycin-induced pulmonary fibrosis. J Immunol, 2009. 182(9): p. 5374-81.
- Borgeson, E. , et al., Lipoxin A(4) and benzo-lipoxin A(4) attenuate experimental renal fibrosis. FASEB J, 2011. 25(9): p. 2967-79.
- Qu, X. , et al., Resolvins E1 and D1 inhibit interstitial fibrosis in the obstructed kidney via inhibition of local fibroblast proliferation. J Pathol, 2012. 228(4): p. 506-19.
- Lagana AS, Vitale SG, Ban Frangez H, Vrtacnik-Bokal E., D’Anna R. Vitamin D in human reproduction: the more, the better? An evidence-based critical appraisal. European Review for Medical and Pharmacological Sciences. 2017; 21: 4243-4251.
- Colonese F, Laganà AS, Colonese E, Sofo V, Salmeri FM, Granese R, Triolo O. The pleiotropic Effects of Vitamin D in Gynaecological and Obstetric Diseases: An Overview on a Hot Topic. BioMed Research International Volume 2015, Article ID 986281, 11 pages. [CrossRef]
- Vergara D, Catherino WH, Trojano G, Tinelli A. Vitamin D: Mechanism of Action and Biological Effects in Uterine Fibroids. Nutrients. 2021, 13, 597. [CrossRef]
Figure 1.
Biosynthesis of the SPMs Resolvins, Protectins and Maresins. EPA; Eicosapentaenoic Acid; 18-HpEDE, 17-HpDHA, 14-HPDHA: precursors of the SPMs during biosynthesis; Cox-1/2: Cyclooxygenases. LOX: lipoxygenase. ASS: aspirin. Aspirin triggers biosynthesis of 18-HpEDE, 17-HpDHA intermediates via modification of COX-enzymes. Maresins are produced by macrophages via a preliminary lipoxygenation step. Further lipoxygenases are required for SPM biosynthesis as depicted (modified from Serhan [40,41]).
Figure 1.
Biosynthesis of the SPMs Resolvins, Protectins and Maresins. EPA; Eicosapentaenoic Acid; 18-HpEDE, 17-HpDHA, 14-HPDHA: precursors of the SPMs during biosynthesis; Cox-1/2: Cyclooxygenases. LOX: lipoxygenase. ASS: aspirin. Aspirin triggers biosynthesis of 18-HpEDE, 17-HpDHA intermediates via modification of COX-enzymes. Maresins are produced by macrophages via a preliminary lipoxygenation step. Further lipoxygenases are required for SPM biosynthesis as depicted (modified from Serhan [40,41]).
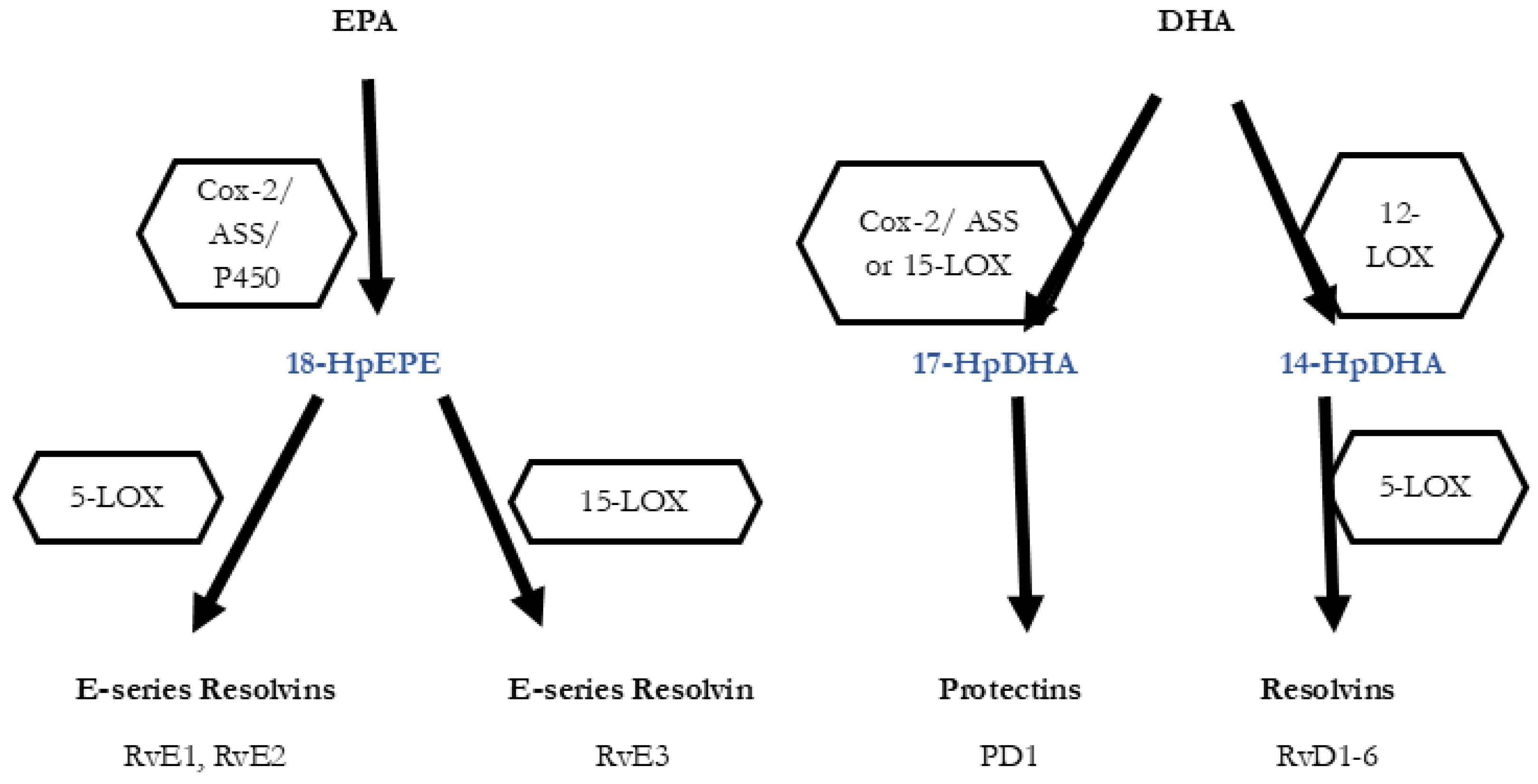
Figure 2.
Concomitant initiation and resolution of inflammation. Synthesis of pro-resolving lipid mediator molecules ist initiated in the beginning of inflammatory processes. LM: Lipid mediator; PUFA: poly-unsaturated fatty acid, PMN: polymorphonuclear neutrophils; SPM: specialized pro-resolving mediators (maresins, resolvins, protectins, lipoxins). Modified from Serhan and Levy [41,45].
Figure 2.
Concomitant initiation and resolution of inflammation. Synthesis of pro-resolving lipid mediator molecules ist initiated in the beginning of inflammatory processes. LM: Lipid mediator; PUFA: poly-unsaturated fatty acid, PMN: polymorphonuclear neutrophils; SPM: specialized pro-resolving mediators (maresins, resolvins, protectins, lipoxins). Modified from Serhan and Levy [41,45].
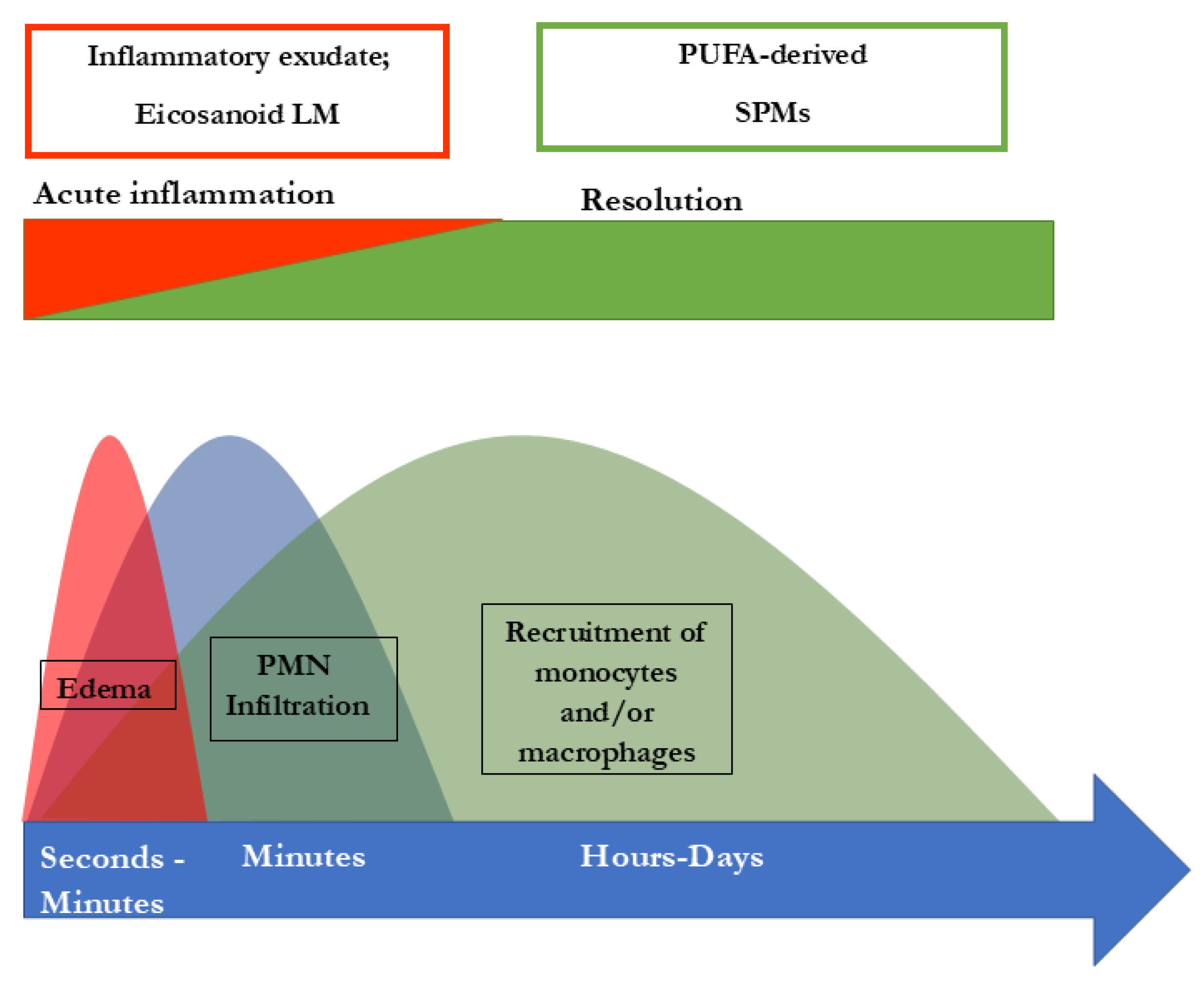
Figure 3.
Role of SPMs during resolution of inflammation and potential outcome of inflammatory processes. After an inflammation is triggered, cells of the innate immune system synthesize eicosanoid lipid mediator LM) molecules (LT, TX, PG etc) from AA, and pro-inflammatory cytokines that stimulate the influx of PMN and moncytes and result in the classical signs of acute inflammation (calor, rubor, tumor, dolor). Already in the beginning, eicosanoids trigger the LM class switch resulting in the synthesis of SPMs from omega-3-PUFAs (See Fig. 1 and text). SPMs are crucial for resolution of inflammation. LM: Lipid mediator; LT: Leukotriene, TX: Thromboxane, PG: Prostaglandins; PUFA: poly-unsaturated fatty acid, SPM Specialized pro-resolving mediators (modified from Serhan [40,41]).
Figure 3.
Role of SPMs during resolution of inflammation and potential outcome of inflammatory processes. After an inflammation is triggered, cells of the innate immune system synthesize eicosanoid lipid mediator LM) molecules (LT, TX, PG etc) from AA, and pro-inflammatory cytokines that stimulate the influx of PMN and moncytes and result in the classical signs of acute inflammation (calor, rubor, tumor, dolor). Already in the beginning, eicosanoids trigger the LM class switch resulting in the synthesis of SPMs from omega-3-PUFAs (See Fig. 1 and text). SPMs are crucial for resolution of inflammation. LM: Lipid mediator; LT: Leukotriene, TX: Thromboxane, PG: Prostaglandins; PUFA: poly-unsaturated fatty acid, SPM Specialized pro-resolving mediators (modified from Serhan [40,41]).

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



