
Submitted:
14 May 2025
Posted:
16 May 2025
You are already at the latest version
Abstract
Previous studies performed in the European eel Anguilla anguilla showed changes in the morphology and physiology of several tissues after exposure to environmental cocaine concentrations. To better understand the model through which cocaine produced its effects on these tissues, we investigated whether there were alterations in the expression of cannabinoid CB1 receptor (CB1R). Indeed, the endocannabinoid system, and CB1R, regulate neurotransmission, neurodevelopment, embryonic development, reproduction, and the activity of the gastrointestinal system. CB1R has been detected in nervous and peripheral tissues in mammals, and orthologues of the mammalian CB1R are found throughout vertebrates including chicken, turtle, frog, and fish. Therefore, samples of gut, kidney, ovary, muscle, liver, skin, and gills from cocaine-exposed and non-exposed eels were processed for routine histology. Immunohistochemical analysis was carried out to evaluate the immunolocalization of the CB1R. Our results showed for the first time 1) the presence of CB1R in the peripheral tissues of the eel 2) statistically significant differences in the localization of CB1R in gut, kidney, ovary, muscle, and liver of the eels exposed to cocaine, compared to controls. These results demonstrate the involvement of CB1R in cocaine effects and suggest its potential role as a biomarker of tissue alteration.
Keywords:
cannabinoid receptor
; cocaine
; European eel
; gills
; gut
; kidney
; liver
; muscle
; ovary
; skin
1. Introduction
The results of numerous studies have shown that the presence of illicit drugs and their metabolites in both fresh and coastal water is a widespread problem [1,2,3,4,5,6], which has its origin in the growing consumption of illicit substances by the human population [7] and, in many cases, in the lack of effective removal of these substances by wastewater treatment plants [8,9,10]. As a result, the aquatic organisms, if living in contaminated waters, are exposed to continuous contact with these drugs and their metabolites [11,12,13,14,15]. The scientific literature reports many effects, bioaccumulation, genetic, metabolic, systemic, nervous, and endocrine, observed in the exposed organisms, due to the remarkable pharmacological properties exhibited both by illicit drugs and their metabolites [5,12,13,14,16]. Previous studies on the European eel (Anguilla anguilla) have shown that chronic exposure to low environmental concentrations of cocaine, one of the most common illicit drugs found in aquatic environments, led to its accumulation in the brain and peripheral tissues of exposed specimens, as well as alterations of organs such as liver, kidney, intestine, gills, muscle, and skin, as well as affecting the endocrine system [14]. To better understand the mechanism of action of cocaine, we decided to test whether it can induce changes in the expression of the CB1 cannabinoid receptor (CB1R) in altered tissues compared to controls. CB1R is part of the endocannabinoid system (eCB), a widely preserved signalling system in vertebrates and many invertebrates, whose main function is to maintain homeostasis [17,18,19,20]. eCB consists of the endocannabinoids themselves, the enzymes involved in their synthesis and degradation, and the cannabinoid receptors. The best known and studied endocannabinoids are two derivatives of arachidonic acid; N-arachidonoyl-ethanolamine (AEA; anandamide) and 2-arachidonoylglycerol (2-AG). They are produced respectively by N-acyl-phosphatidylethanolamine-specific phospholipase D (NAPE-PLD), which synthesizes AEA from N-acyl-phosphatidylethanolamine (NAPE), and diacylglycerol lipase (DAGL) which synthesises 2-AG from diacylglycerol (DAG), although these are not the only ways of their synthesis. The degradation of endocannabinoids occurs by oxidation and/or hydrolysis; the main but not the only enzymes involved are fatty acid amide hydrolase (FAAH) which degrades AEA into free arachidonic acid and ethanolamine, and monoacylglycerol lipase (MAGL) which hydrolyses 2-AG into arachidonic acid and glycerol [21,22]. Finally, the main cannabinoid receptors, often present in different isoforms and subtypes in different organisms, are CB1R and the CB2 cannabinoid receptor (CB2R), belonging to the large family of seven transmembrane-spanning G-protein-coupled receptors (GPCRs) [23,24,25]. However, both AEA and 2-AG may also interact with different receptors such as the transient receptor potential cation channel subfamily V member 1 (TRPV1), peroxisome proliferator-activated receptor alpha receptors and the orphan G-protein coupled receptors, GPR55 and GPR119 [26,27].
The choice to investigate whether cocaine effects may at least partially involve changes in CB1R expression was justified by 1) the widespread presence of CB1R throughout the animal kingdom, in peripheral tissues, as well as in the nervous system; 2) CB1R involvement in the response to stress and addiction as well as in the main physiologic activities of the organism. CB1R is a heptameric membrane receptor, with the extracellular N-terminal portion and the cytoplasmic C-terminal tail, coupled to G proteins. The CB1-activated signal transduction pathway includes both the inhibition of adenylate cyclase through the action of protein Gi and the consequent activation/inhibition of Ca2+ and K+ channels and the activation of mitogen-activated kinases. The G proteins associated with these receptors are of type Gi/o; the subunit Gα negatively regulates adenylate cyclase (AC), inducing inhibition of cAMP production, resulting in decreased phosphorylation of protein kinase A (PKA), which is involved in cellular communication pathways such as the regulation of ion channels and FAK kinases. CB1R may also be associated with the Gs protein, thus increasing cAMP levels, although the physiological significance of this duality has yet to be investigated. CB1R bound to the Gq/11 protein activates phospholipase C (PLC), and constitutively activates nitric oxide synthase cNOS, also mediates the release of Ca2+ from intracellular deposits resulting in increased concentration of intracellular Ca2+. Finally, CB1R positively regulates the pathway of MAP (mitosis activating protein) kinases [23,28,29]. CB1R is involved in many activities such as neurotransmission, neuro- and embryonic development, reproduction, food intake, thermoregulation, energy balance, metabolism, stress reduction and motivation, among others [18,30]. Moreover, several studies suggest its involvement in drug addiction in general, and particularly in the effects of cocaine [31,32,33,34,35]. The CB1R is not only widely present in the central nervous system but has also been found in adipocytes, endothelial cells, hepatocytes, in skeletal and smooth muscle, skin, bone, in the cardiovascular system, the gastrointestinal tract, the enteric nervous system, the reproductive system and different types of cancer [20. 22]. In mammals, CB1R has been detected in nervous and peripheral tissues, and orthologues of the mammalian CB1R are found throughout vertebrates including chicken, turtle, frog, and fish [36]. In teleost, a likely gene duplication event in a common ancestor followed by retention and/or loss of duplicate genes has in many cases resulted in copies of CBIR and CB2R. In zebrafish, there is only one CB1 gene and two CB2 genes, while in Fugu rubripes two CB1-type genes (CB1A and CB1B) and a single CB2 gene have been identified. In Solea solea, two forms of CB1R have also been found, CB1A and CB1B, whose deduced amino acid sequences are 80% identical, whereas only one CB1 receptor type was identified in Pelvicachromis pulcher, Carassius auratus and Sparus aurata [17]. In the European eel, the existence of a cannabinoid receptor of 469 aa has been inferred from homology [37] (UniProt: A0A9D3MIM1_ANGAN). Moreover, a cannabinoid receptor 2 gene (cnr2), located on chromosome 1, has been identified (Bethesda, National Library of Medicine: https://www.ncbi.nlm.nih.gov/gene/118235455/#general-protein-info).To explore the mechanism underlying histological changes in the tissues of cocaine-exposed eels, the presence and distribution of CB1R was investigated by an immunohistochemical analysis and a semiquantitative analysis of the immunoreactivity.
2. Materials and Methods
2.1. Animals
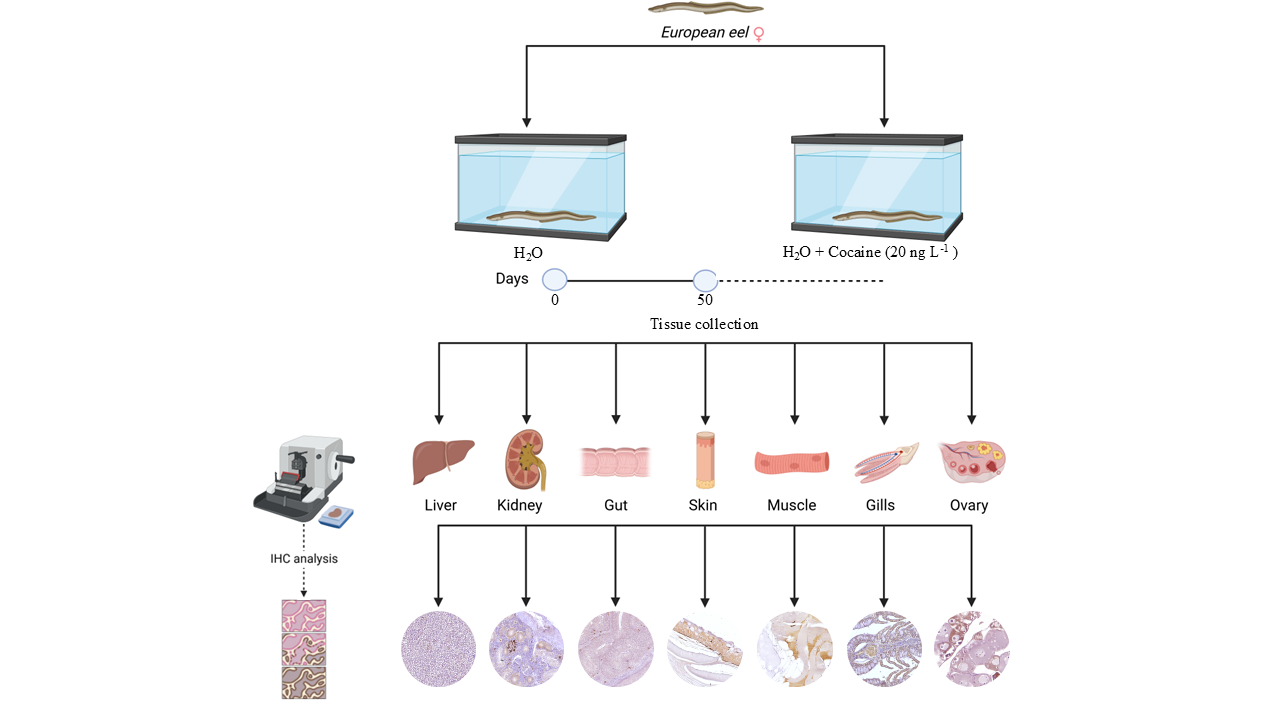
Twenty-seven female specimens of the European eel (A. anguilla) in silver stage (52.0 ± 3.93 cm and 290.70 ± 36.40 g, mean ± s.d.) were purchased from a fish dealer and acclimatized in our laboratory for one month in 300 L glass aquaria under natural photoperiod. The water in the aquaria was dechlorinated and well-aerated, with the following characteristics: dissolved oxygen 8.0 ± 0.7 mg L-1, ammonia <0.1 mg L-1, salinity 1°/oo, pH 7.4 ± 0.8, and temperature 15°C ± 1°C. The water in all the aquaria was replaced with fresh tap water every 24 h; furthermore, since silver stage eels do not eat, they were not fed. This study was carried out in accordance with EU Directive 2010/63/EU for animal experimentation and institutional guidelines for care and use of laboratory animals and was authorized by General Direction of Animal Health and Veterinary Drugs of the Italian Ministry of Health (Authorization n.221/2015-PR and Authorization n. 22/2015-PR).
2.2. Experimental Design
A stock solution of 0.006 mg mL-1 free base cocaine (≥97% purity, Sigma-Aldrich Inc., St. Louis, MO, USA) in ethanol was prepared. The eels were randomly divided into three groups (eels exposed to cocaine, eels exposed to ethanol at the same concentration of the cocaine-exposed specimens, and eels exposed to tap water only) of 9 animals each. The experiment was carried out in triplicate, therefore, the specimens in each experimental group were divided into three aquaria containing 3 specimens each one. Three control groups of three eels each were dipped in tap water only, while three groups of three eels were exposed to a nominal dose of 20 ng L-1 cocaine (1 mL of stock solution, added to the aquaria after water changes every 24 h) and three control groups of three eels each were exposed to ethanol. Water removed from the aquaria containing experimental groups was stored in special containers for three days before being discharged as wastewater since nearly 90% of cocaine is degraded in 24 h at room temperature in water [38]. All groups were kept in 300 L glass aquaria under the conditions described above, and water was changed every 24 h. After the exposure, eels were anesthetized using MS-222 (ethyl 3-aminobenzoate, methanesulfonic acid salt 98%, Aldrich Chemical Corporation Inc., Milwaukee, WI, USA) at a concentration of 100 mg L-1, buffered with sodium bicarbonate at neutral pH. The specimens were weighed, measured, and killed by decapitation (Authorization n.221/2015-PR and Authorization n. 22/2015-PR). Gut, skeletal muscle, kidney, liver, ovaries, dorsal skin, and gills were sampled from each animal and processed for light microscopy. After decapitation, the tissues were immediately fixed in Bouin’s solution for 24 hours, dehydrated in graded alcohols, cleared in Histolemon and embedded in Paraplast. Tissue specimens were sectioned with a microtome (Rotary 3003, pfm, BioOptica, Milan, Italy) into 4 μm thick sections and placed on poly-L-lysine slides (Menzel-Glaser, Braunschweig, Germany).
2.3. Western Blot
Since no eel specific CB1R has been characterized, we used Western blot to validate our antibody to CB1R. The Anguilla anguilla kidney was potter homogenised at 4 °C with 5 mL of RIPA buffer containing 50 mM Tris-HCl (pH 7.4), 150 mM NaCl, 1 mM EDTA, 10% NP-40 and protease inhibitor cocktail (3 mM aprotinin, 1 mM leupeptin, 1 mM e-64, 130 μM bestain, 1 mM EDTA and 2 mm AEBSF) [39]. The homogenate was centrifuged at 7000 g for 10 min at 4 °C. Proteins were quantified using the PIERCE method. This type of analysis was carried out with the aim of validating the anti-CB1R (SAB4500345, Sigma-Aldrich). For this purpose, 40μg of protein from each sample was loaded, after boiling for 5 minutes in SDS buffer [50 mM Tris-HCl (pH 6.8), 2 g 100 mL-1 SDS, 10% (v/v) glycerol, 0.1 g 100 mL-1 Bromophenol blue], run on 12.5% SDS/polyacrylamide gels and transferred onto a nitrocellulose membrane using a Mini trans-blot apparatus (Bio-Rad Laboratories). The membrane was blocked at room temperature, then blocked with 7% non-fat milk dissolved in TBS 1x- 0,025% Tween20 for 45 minutes and then with 7% BSA in TBS 1x-Tween20 for a further 45 minutes. The membranes were incubated overnight at 4 °C with anti-rabbit CB1R (Sigma-Aldrich), diluted 1: 500 in TBS-T 2,5% BSA. Then, the membranes were washed three times with TBS-T and incubated for 1 h with anti-rabbit IgG secondary antibody (Santa Cruz, Cat. Sc-2005) diluted 1: 2000 in TBS-T and 2.5% BSA. The results were acquired with Chemidoc Biorad.
2.4. Immunohistochemical Analysis
Slides were first deparaffined in xylene and then rehydrated using a graded series of alcoholic solutions. For antigen unmasking, slides were heat-treated in the microwave for 20 minutes in 10 mM citrate buffer (pH 6.0). Further, slides were incubated with 2.5% hydrogen peroxide (H₂O₂) for 30 minutes to inhibit endogenous peroxidase activity and, non-specific blockade was performed by incubation for 1 hour at room temperature with normal goat serum (Pierce, Rockford, IL, USA). Sections were then incubated at 4 °C overnight with following primary antibody diluted in normal goat serum: rabbit anti-CB1R (1:250, Abcam). The following day, slides were washed two times (5 minutes each) in PBS, and a 1-hour incubation at room temperature with HRP-conjugated goat anti-rabbit secondary antibody diluted 1:200 in normal goat serum was performed. Immunoreactivity signal was detected using diaminobenzidine (DAB) as chromogen, followed by counterstaining with Carazzi's haematoxylin [40]. For negative controls, the primary antibody was omitted during incubation. The images were acquired using an Axiocam MRc5 camera (Carl Zeiss) and Axiovision 4.7 software (Carl Zeiss) [41].
2.5. Semiquantitative Analysis of the Immunoreactivity
The immunoreactivity of the antibody was evaluated, and its cellular localisation was reported. A semiquantitative assessment of immunohistochemical stained sections was performed on 5 randomly selected slides for each tissue. For each slide, twenty randomly chosen not overlapping fields at high power magnification (40x) were photographed under a Zeiss Axioskop microscope coupled to a video camera Zeiss Axiocam 305 color and Zen microscopy software-blue edition. Each photo was elaborated with Fiji (ImageJ, National Institutes of Health): a color deconvolution filter (H DAB) was applicated to each photo to discriminate browns and blues; the obtained 8-bit browns level was binarized using a threshold manually adjusted (between 0–100 and 0–120), as needed, for each image to reflect chromogen distribution in the regions of interest [42,43]. The overall score of immunopositive cells was calculated by averaging scores of all examined cases of the same treatment group (three tissue per treatment) while the relative percentages of immunopositive cells were expressed as mean ± standard deviation (SD). The immunolabeling grade was scored as follows: 0 (Absent), 1 (Faint <10%), 2 (Moderate 10–30%), and 3 (Strong >30%).
2.6. Statistical Analysis
Statistical analysis was performed using GraphPad (version 10.4.1; GraphPad Software Inc., San Diego, CA, USA). Differences among each histological semiquantitative score were evaluated using two nonparametric tests: Mann–Whitney test and Kruskal–Wallis test followed by a post-hoc multiple comparison using Dunn's test. The nonparametric tests were used to assess differences in the number of CB1R-positive cells evaluated between control and exposed groups; p-values < 0.05 were considered statistically significant.
3. Results
Since no differences were found between the specimens exposed to water and to vehicle (ethanol), the results related to the vehicle-exposed specimens have not been shown.
3.1. Western Blot
As regards Western blot, the CB1R antibody detected a single molecular band of approximately 52 kDa (Figure 1), corresponding to the expected molecular weight of the CB1 protein, confirming the specificity of antibody binding. The absence of signals under negative control conditions, obtained by omitting the primary antibody, excludes the possibility of non-specific reactions or interference by the secondary antibody (Figure 1).
3.2. Gut and Liver Immunoistochemistry
Both in the gut and liver, CB1R distribution was similar. In control specimens, gut epithelium and smooth muscle tissue showed an absent to faint immunopositivity, whereas in cocaine-exposed specimens, both tissues showed a moderate to strong immunopositivity (Figure 2). In the same way, in the liver of control specimens, the immunopositivity appeared poorly distributed, whereas in cocaine-exposed specimens a greater immunopositivity was found (Figure 2). These results were confirmed by statistical analysis that showed statistically significant differences in the localization of CB1R among the control and the exposed specimens (p < 0.05) (Figure 2).
3.3. Kidney Immunoistochemistry
In control specimens, the hematopoietic tissue showed a moderate to strong CB1R immunopositivity, whereas both the proximal and the distal tubules appeared generally immunonegative. In the exposed specimens, the immunopositivity of the hematopoietic tissue appeared absent/faint, whereas a mild to moderate immunopositivity to CB1R was found in proximal tubules. The distal tubules appeared generally immunonegative (Figure 3). Statistical analysis showed statistically significant differences in the localization of CB-1R among the control and the exposed specimens (p < 0.05) (Figure 3).
3.4. Ovary Immunoistochemistry
Histological analysis revealed an asynchronous ovarian development in the exposed animals compared to controls. Specifically, the ovaries of control animals were mainly composed of fully vitellogenic oocytes, that is large oocytes that had completed vitellogenesis. In contrast, the ovaries of cocaine-exposed animals exhibited an altered oocyte developmental progression, characterized by the presence of oocytes at early growth stages, including previtellogenic oocytes, which lacked yolk vesicles, and early vitellogenic oocytes, which showed partial yolk vesicle accumulation in the cytoplasm and a centrally located nucleus. Immunohistochemical analysis revealed a differential distribution of the CB1R. As a result of this asynchronous development determined by cocaine, in control specimens, CB1R was mainly expressed in previtellogenic oocytes and in mid vitellogenic oocytes (Figure 4) whereas, in cocaine exposed specimens, only previtellogenic oocytes showed a strong positivity to CB1R (Figure 4). Statistical analysis showed statistically significant differences in the localization of CB1R among the control and the exposed specimens (p < 0.05) (Figure 4).
3.5. Skeletal Muscle Immunoistochemistry
Immunohistochemical analysis showed a different expression of CB1R depending on the muscle fiber type. In the control specimens the white muscle fibers appeared immunopositive while the red ones did not show positivity to CB1R (Figure 5). In cocaine exposed specimens, white fibers showed a reduced immunopositivity, whereas the red fibers showed immunopositivity to CB1R (Figure 5). These results were confirmed by statistical analysis that showed statistically significant differences in the localization of CB1R among the control and the exposed specimens (p < 0.05) (Figure 5).
3.6. Skin and Gills Immunoistochemistry
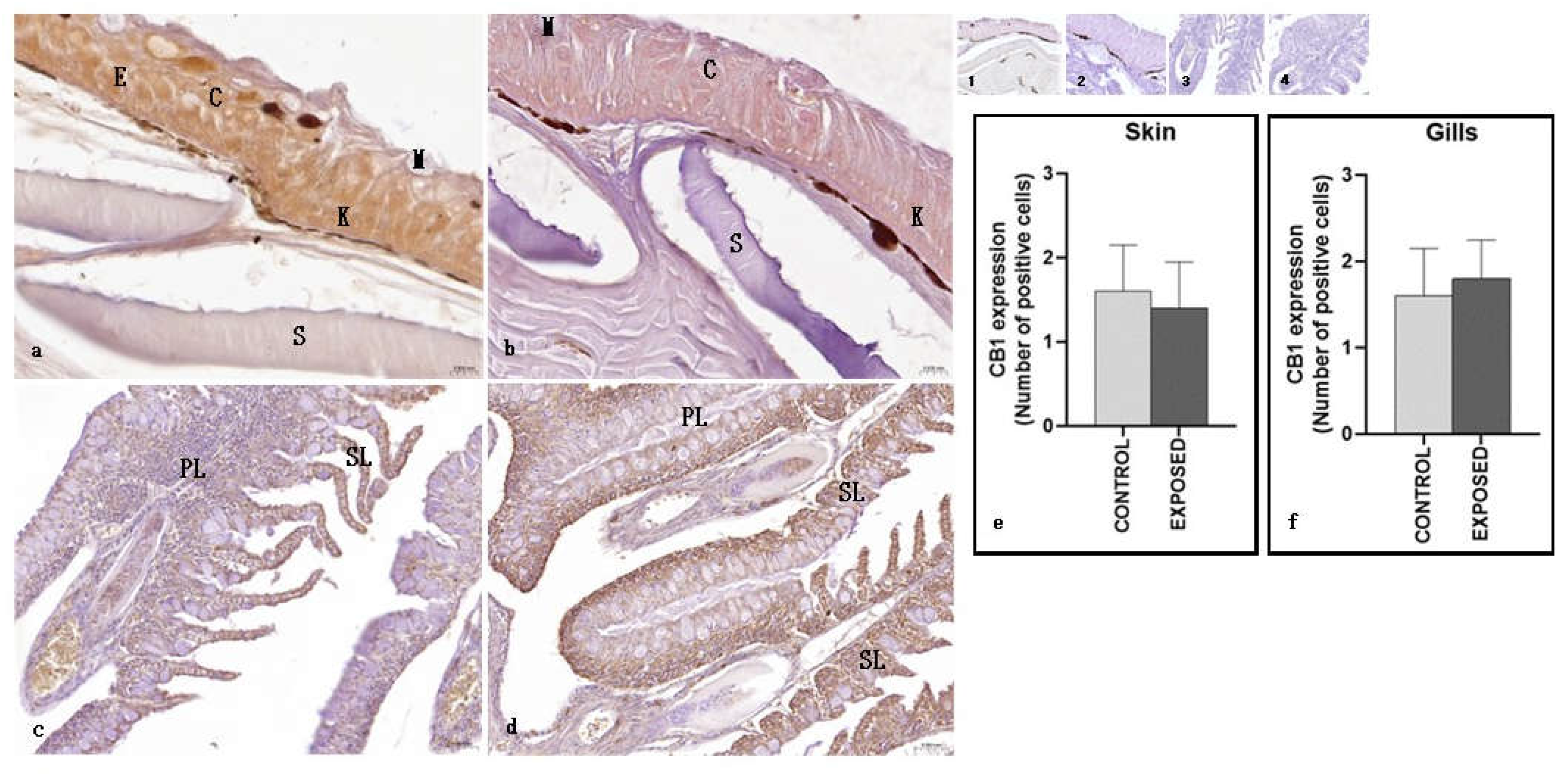
As regards the skin, both in the control and the exposed specimens, CB1R positivity was observed in the epithelial layer, both in keratinocytes and club cells, but not in mucosal cells. No changes due to cocaine exposure were observed (Figure 6). Statistical analysis showed no statistically significant differences in the localisation of CB1R between the control and the exposed specimens (Figure 6). As regards the gills, both in the control and the exposed specimens, a moderate immunopositivity to CB1R was observed both in primary and secondary lamellae. No changes due to cocaine exposure were observed (Figure 6). Statistical analysis showed no statistically significant differences in the localisation of CB1R between the control and the exposed specimens (p < 0.05) (Figure 6).
4. Discussion
In our study, we used immunohistochemistry to evidence in the peripheral tissues of the European eel the presence of CB1R, and its possible variations due to exposure to cocaine. As no eel-specific CB1R antibody was available, we used a rabbit antibody raised against the human CB1R. Indeed, the CB1R is well conserved throughout evolution, as 97% of mouse CB1R, 84% of amphibian CB1R and 72% of fish CB1R are identical to the human CB1 [44]. Moreover, we performed a western blot analysis showing a molecular band of approximately 52 kDa, corresponding to the expected molecular weight of the CB1R protein. Therefore, we were confident that our antibody could recognize the eel protein.
Several conclusions can be drawn from the results of this study. First, the results of the immunohistochemical analysis showed for the first time the presence of the CB1R in the examined tissues of the European eel, although at different levels, as evidenced by the semiquantitative analysis. Secondly, the results showed that in many tissues cocaine exposure was able to vary the expression of CB1R, suggesting a possible involvement of the endocannabinoid system in the mechanism of action of cocaine. Such involvement has been previously shown in the nervous tissue [31,32,33,34,35] but not in peripheral tissues.
The presence of CB1R immunopositivity was variable and varied in the different tissues examined. In the control specimens, immunopositivity appeared low/weak in the intestinal epithelium; moderate/strong in the renal hematopoietic tissue; evident in the degenerating and previtellogenic oocytes in the ovary, and in the white fibers of striated muscle tissue; weak in the hepatic parenchyma; moderate in the epidermis and gill epithelium. These results are consistent with data in the literature demonstrating the wide presence of CB1R in peripheral tissues in the animal kingdom [17,20,21,22,36,45,46,47] and in fish [17,18,30].
As regards the gut, in the intestinal epithelium and in the smooth muscle tissue of the control specimens the presence of CB1R appeared to be very low, while the exposure to cocaine resulted in a significant increase in both tissues. Literature data indicate the presence of CB1R in the gastrointestinal system, where the receptor is abundant in the enteric nervous system, in non-neuronal cells as intestinal mucosa and enterocytes, and in smooth muscle tissue [20,22,26,48]. Our results, which show a low presence of CB1R in the gut of the control specimens, contrast with those generally reported in the literature. There is no data on the presence of the CB1R in peripheral tissues of eel, and its expression in relation to different eel metamorphic stages, but one possible explanation could be that the specimens were in the silver stage, characterized by physiological bowel regression in preparation for reproductive migration, during which the eels do not feed [49]. CB1R is involved in the regulation of intestinal epithelium permeability, gastrointestinal motility, energy balance and the increase of food intake, among others [22,40,50,51], therefore it is possible that in the silver stage CB1R has been physiologically downregulated. Previous studies showed that the chronic exposure to cocaine increased the thickness of the epithelium and reactivated the structure of the intestine and of the intestinal musculature [52]. It could be assumed that the mechanism of action of cocaine involved the activation of the CB1R receptor, which is known to increase gastrointestinal epithelial integrity and tight-junctions proteins [53].
As for the liver, which is one of the main organs involved in the metabolization of cocaine [54], CB1R immunopositivity, which was poor in the control specimens, increased significantly after exposure to cocaine. Our results agree with those in the literature, which report a low presence of the CB1R in the whole liver and hepatocytes, and its increase in various liver diseases, where it contributes to the underlying pathologies [55,56]. Such CB1R increase in human and mouse hepatocytes was observed under conditions of high fat or alcohol diets [57] and it was found to contribute to hemodynamic abnormalities, to promote hepatic lipogenesis [58] and fibrosis in liver cirrhosis [55]. Previous studies [59] found in the liver of eels chronically exposed to cocaine morphologic alterations such as necrotic areas, karyolitic and pyknotic nuclei; increased cytochrome oxidase (COX) and caspase-3 activities, markers of oxidative metabolism and apoptosis activation, respectively. Moreover, increase in serum levels of alanine aminotransferase (ALT), marker of liver injury, was observed. The increase in CB1R in liver of our cocaine-exposed eels agrees with the increase of CB1R observed in hepatic diseases. As far as further studies are needed, it could be assumed that the hepatotoxicity induced by cocaine involved the activation of the CB1R.
The teleost kidney is a mixed organ performing excretory and endocrine functions. It comprises hematopoietic, reticuloendothelial, endocrine, and excretory elements, the nephrons, consisting of a glomerulus within Bowman’s capsule, proximal, distal, and collecting convolute tubules [60,61]. A moderate to strong CB1R immunopositivity was found in the hematopoietic tissue of the control specimens, whereas both the proximal and the distal tubules appeared generally immunonegative. After cocaine exposure, the immunopositivity of the hematopoietic tissue appeared absent/faint, whereas a mild to moderate immunopositivity to CB1R was observed only in the proximal tubules, causing a general decrease in kidney CB1R expression. Our results, showing CB1R immunopositivity in the hematopoietic tissue of control specimens, agree with those of studies that demonstrate the presence of CB1R in murine and human hematopoietic tissue, suggesting a role of these receptors in the regulation of immunity and blood cells development [52,62]. However, the strong attenuation of immunopositivity in the hematopoietic tissue of the exposed specimens needs further studies to be better evaluated. As for the presence of CB1R in the fish kidney, there is little literature; as far as we know, CB1R has been identified in goldfish [63] and zebrafish [46] kidney. Most studies were conducted on mammals, where CB1R was considered the predominant eCB kidney receptor. Its expression, low in healthy kidneys, has been identified in all components of the nephron and in the collecting duct; the activation of CB1R contributes to multiple renal diseases through the mitogen-activated protein kinase (MAPK) pathway [64,65,66]. Previous studies found, in the kidney of eels exposed to cocaine, nephrotoxic effects as morphologic alterations and the increase of COX and caspase-3 activity, like that observed in the liver [58]. Our results, showing immunonegativity in renal tubules of control eels, and immunopositivity in proximal tubules of cocaine exposed eels, suggest an activation of CB1R induced by cocaine in proximal tubules. Interestingly, in the kidney, cocaine is metabolized to ecgonine methyl ester and norcocaine (NCOC), the last known to induce apoptosis in primary cultured human proximal tubular epithelial cells [67]. Therefore, although further studies are needed, it could be assumed that at least partially the nephrotoxicity induced by cocaine/norcocaine involved the activation of the CB1R, known to induce apoptosis in colon cancer cells [68].
Regarding the ovary, in the controls CB1R was mainly expressed in previtellogenic and in mild vitellogenic oocytes. On the contrary, in cocaine-treated eels, CB1R immunopositivity was observed only in early developmental stages, with especially localization in previtellogenic oocytes.
This distribution suggests that the early presence of the receptor may make immature follicles more vulnerable to the action of cocaine, thus contributing to the slowing of follicular progression.
These observations are consistent with previous studies [69], in which cocaine-treated eels showed a reduced rate of ovarian maturation compared to controls. In these animals, in fact, there was a prevalence of previtellogenic oocytes, large areas of connective tissue and larger and more numerous yolk vesicles.
The increased expression of CB1R in control samples could therefore reflect the normal course of folliculogenesis, during which the receptor appears even in the most advanced stages, as indicated by the positivity observed in mid-stage vitellogenic oocytes. In cocaine-treated eels, on the other hand, follicular development slowed down by the action of the drug could determine the non-appearance of mid vitellogenic oocytes; this phenomenon could therefore explain why CB1R levels in exposed people are lower than in controls. This is in line with the literature where it is known that high levels of endocannabinoids and exogenous cannabinoids (abnormal activation of the endocannabinoid system) suppress the release of gonadotropin-releasing hormone (GnRH), luteinizing hormone (LH), follicle-stimulating hormone (FSH), oestrogen and progesterone. Furthermore, since mitochondrial function is associated with oocyte quality, the presence of CB1 on the outer mitochondrial membrane can disturb ovarian function and therefore oogenesis [70,71].
In control animals, the appearance of the receptor in mid vitellogenic oocytes is in line with numerous evidence in the literature confirming the key role of the endocannabinoid system in the regulation of folliculogenesis, oocyte maturation and endocrine function of the ovary [72]. In mammals, CB1R has been identified in the ovaries, and an entire endocannabinoid system has been localized within human ovarian follicles, suggesting a regulatory function of these signals in follicular development [70]. In addition, CB1R has also been detected in the ovary of zebrafish [46], confirming the evolutionary conservation of this system in vertebrates.
As for the skeletal muscle, in the control specimens the white muscle fibers appeared immunopositive and the red ones did not, whereas in cocaine exposed specimens, white fibers reduced their immunopositivity, and the red fibers became immunopositive to CB1R. In humans and rodents, CB1R is localized on cell membrane and in mitochondria, and is involved in oxidative metabolism [73,74,75]. The observation of a prevalent association of the CB1 receptor with slow versus fast fibers suggests its role in influencing the type of muscle fiber [76]. The activation of CB1 is associated with metabolic dysfunction, chronic inflammation, and muscle loss, while its antagonism can improve insulin sensitivity, increase glucose absorption, reduce fat mass, and promote muscle growth through molecular pathways such as PI3K/Akt and mTOR [77]. Moreover, CB1R also regulates muscle apoptosis, indeed agonism of CB1R induces various markers of apoptosis as caspase-3 in human embryonic rhabdomyosarcoma cells and in human primary and in rat cardiomyocytes [78]. Our results, showing the presence of CB1R in the skeletal muscle of A. anguilla are consistent with literature data showing the presence of CB1R in skeletal muscle [73,74,75], but are in contrast with the prevalent association of CB1R with slow fibers; indeed, CB1R was associated in the control eels to fast fibres. The reason of this difference could be related to species differences and needs further studies to be evaluated. Previous studies showed in the skeletal muscle of cocaine exposed eels, signs of injury as muscle breakdown and swelling, reminiscent of rhabdomyolysis, and more evident in slow fibres; increase in caspase-3 activity, and of serum levels of creatine kinase, lactate dehydrogenase and aspartate aminotransferase, markers of the functional state of the muscle [79]. The decrease in the immunopositivity of fast fibres, and the appearance of CB1R immunoreactivity in slow fibers after exposure to cocaine could be explained with a higher sensitivity of these fibers to cocaine, and with the greatest morphological damages, observed in the slow fibres compared to fast ones. Probably, the cocaine-induced injury involved the activation of CB1R in the slow fibers, where it was not present in normal conditions.
As for the skin and the gills, no changes due to cocaine exposure were observed in the CB1R immunopositivity, that was found in the skin epithelial layer (keratinocytes and club cells) and in the gills primary and secondary lamellae. In humans and rodent skin, a functional eCB has been identified, involved in the modulation of cell growth, proliferation, and death. CB1R and CB2R immunoreactivities have been observed in human and murine skin cells including epidermal keratinocytes, in which the activation of CB1R and CB2R by locally produced endocannabinoids induces suppression of cellular proliferation, differentiation and the release of inflammatory mediators as well as the induction of apoptosis [80,81]. As for the gills, as far as we know, CB1R was found in the gills of goldfish [63] whereas CB2a and CB2b receptors have been identified in the gills of zebrafish [82]. Our results, showing CB1R immunopositivity in the eel keratinocytes and in the gills, agree with those in the literature and suggest that probably even in the eel the eCB could exert a modulating function in these tissues, that however needs further studies to be evaluated. However, despite the morphological changes induced by cocaine in the epidermis (increased thickness of the epithelium and thickening and folding of the underlying basal lamina) [52] and in the gills (partial and total fusion of the secondary lamellae, proliferation of the interlamellar epithelium, epithelial lifting and aneurism in the secondary lamellae) [83], there were no changes in CB1R immunopositivity in both tissues. Although further studies are needed, the action of cocaine could also involve in these tissues receptors others than CB1R.
5. Conclusions
In conclusion, the results of our study showed for the first time the presence of CB1R in some peripheral tissues of the eel, and changes in this receptor in many of these tissues after cocaine exposure, suggesting an involvement of CB1R in the response of the organism to cocaine. This involvement had already been observed in the nervous system, but not in peripheral tissues. In the different eel tissues, the alterations of the CB1 receptor were different, going from increase to decrease or lack of alterations of the expression of the receptor; this difference could be explained by the fact that different signalling pathways were activated in different tissues, and/or even different receptors. The European eel is an endangered species; considering the important physiological activities carried out by the CB1R receptor in this as in other species, the study of variations of this receptor, and of the endocannabinoid system more generally, can be of great use for the better knowledge and conservation of this species.
Author Contributions
Conceptualization: A.C., L.RO., R.S. and M.D.F. ; methodology: L.R.; T.C. and A.M.; software, L.R.; validation, L.R.; formal analysis, L.R.; investigation, L.R.; T.C.; S.B; R.G.; data curation, L.R.; writing—original draft preparation: L.RO; A.C..; writing—review and editing, A.C.; L.R.; L.RO., M.D.F.; visualization, L.R.; L.RO; supervision, A.C., L.RO and M.D.F.
All authors have read and agreed to the published version of the manuscript.”
Funding
This research received no external funding.
Institutional Review Board Statement
This study was carried out in accordance with EU Directive 2010/63/EU for animal experimentation and institutional guidelines for care and use of laboratory animals and was authorized by General Direction of Animal Health and Veterinary Drugs of the Italian Ministry of Health (Authorization n.221/2015-PR and Authorization n. 22/2015-PR).
Data Availability Statement
The data supporting the conclusions of this article will be made available by the authors on request.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Pal, R., Megharaj, M., Kirkbride, K.P., Naidu, R. Illicit drugs and the environment-A review. Sci. Total Environ. 2013, 463-464, 1079-1092. [CrossRef]
- Rosi-Marshall, E.J., Snow, D., Bartelt-Hunt, S.L., Paspalof, A., Tank, J.L. A review of ecological effects and environmental fate of illicit drugs in aquatic ecosystems. J. Hazard. Mater. 2015, 282, 18-25. [CrossRef]
- Aligizakis, N.A., Gago-Ferrero, P., Borova, V.L., Pavlidou, A., Hatzianestis, I., Thomaidis N.S.. Occurrence and spatial distribution of 158 pharmaceuticals, drugs of abuse and related metabolites in offshore seawater. Sci. Total Environ. 2016, 541, 1097-1105. [CrossRef]
- Seabra-Pereira, C.D., Maranho, L.A., Cortez, F.S., Pusceddu, F.H., Santos, A.R., Ribeiro, D.A., Cesar, A., Guimarães L.L. Occurrence of pharmaceuticals and cocaine in a Brazilian coastal zone. Sci. Total Environ. 2016, 548-549, 148-154. [CrossRef]
- Fontes, M.K., Maranho, L., Pereira, C.D.S. Review on the occurrence and biological effects of illicit drugs in aquatic ecosystems. Environ. Sci. Pollut. Res. 2020, 27, 30998–31034. [CrossRef]
- Muñiz-Bustamante, L., Caballero-Casero, N., Rubio, S. Drugs of abuse in tap water from eight European countries: Determination by use of supramolecular solvents and tentative evaluation of risks to human health. Environment International. 2022, 164, 107281. [CrossRef]
- UNODC, World Drug Report 2024 (United Nations Publication 2024). https://www.unodc.org/documents/data-and-analysis/WDR_2024/WDR24_Key_findings_and_conclusions.pdf.
- Kasprzyk-Hordern, B., Dinsdale, R.M., Guwy, A. J. The removal of pharmaceuticals, personal care products, endocrine disruptors and illicit drugs during wastewater treatment and its impact on the quality of receiving waters. Water Res. 2009, 43, 363–380. [CrossRef]
- Deng, Y., Guo, C., Zhang, H., Yin, X., Chen, L., Wu, D., Xu, J. Occurrence and removal of illicit drugs in different wastewater treatment plants with different treatment techniques. Environ. Sci. Eur. 2020, 32, 28. [CrossRef]
- Davey, C.J.E., Kraak, M.H.S., Pratorius, A., ter Laak, T.L., van Wezel, A. P. Occurrence, hazard, and risk of psychopharmaceuticals and illicit drugs in European surface waters. Water Res. 2022, 222, 118878. [CrossRef]
- Binelli, A., Pedriali, A., Riva, C., Parolini, M. Illicit drugs as new environmental pollutants: Cyto-genotoxic effects of cocaine on the biological model Dreissena polymorpha. Chemosphere. 2012, 86, 906-911. [CrossRef]
- Chen, L., Guo, C., Sun, Z., Xu, J. Occurrence, bioaccumulation and toxicological effect of drugs of abuse in aquatic ecosystem: A review. Environ. Res. 2021, 200, 111362. [CrossRef]
- Fontes, M.K., Dourado, P.L.R. de Campos, B.G. Maranho, L.A. de Almeida, E.A. de Souza Abessa, D.M. Pereira, C.D.S. Environmentally realistic concentrations of cocaine in seawater disturbed neuroendocrine parameters and energy status in the marine mussel Perna perna. Comp. Biochem. Physiol. 2022a, 251, 109198. [CrossRef]
- Rosati, L., Caputo, I., Lionetti, L., Fontes, M.K., Seabra Pereira, C.D., Capaldo, A. Side effects of human drug use: an overview of the consequences of eels’ exposure to cocaine. Fishes. 2023, 8, 166. [CrossRef]
- Sokołowski, A., Mordec, M., Caban, M., Øverjordet, I.B., Wielogorska, E., Włodarska-Kowalczuk, M., Balazy, P., Chełchowski, M., Lepoint. G. Bioaccumulation of pharmaceuticals and stimulants in macrobenthic food web in the European Arctic as determined using stable isotope approach. Sci. Total Environ. 2024, 909, 168557. [CrossRef]
- De Farias Araujo, G., de Oliveira, L.V.A., Hoff, R.B., Wosnick, N., Vianna, M., Verruck, S., Hauser-Davis, R.A., Saggioro, E.M. “Cocaine Shark”: First report on cocaine and benzoylecgonine detection in sharks. Sci. Total Environ. 2024, 948, 174798. [CrossRef]
- Elphick, M.R. The evolution and comparative neurobiology of endocannabinoid signalling. Phil. Trans. R. Soc. B. 2012, 367, 3201–3215. [CrossRef]
- Oltrabella F., Melgoza A., Nguyen B., Guo S. Role of the endocannabinoid system in vertebrates: Emphasis on the zebrafish model. Dev Growth Differ. 2017 May;59(4):194-210. [CrossRef]
- Silver, R.J. The endocannabinoid system of animals. Animals. 2019, 9, 686-701. [CrossRef]
- Bailone R.L., Fukushima H.C.S., Kluwe de Agular, Borra R.C. The endocannabinoid system in zebrafish and its potential to study the effects of Cannabis in humans. Laboratory Animal Research. 2022, 38, 5. [CrossRef]
- Pacher, P., Batkai, S., Kunos, G. The endocannabinoid system as an emerging target of pharmacotherapy. Pharmacol. Rev. 2006, 58, 389–462. [CrossRef]
- Zou, S., Kumar, U. Cannabinoid Receptors and the Endocannabinoid System: Signaling and Function in the Central Nervous System. Int. J. Mol. Sci., 2018, 19, 833. [CrossRef]
- Matsuda LA, Lolait SJ, Brownstein MJ, Young AC, Bonner TI. Structure of a cannabinoid receptor and functional expression of the cloned cDNA. Nature. 1990; 346:561–4. [CrossRef]
- Munro S, Thomas KL, Abu-Shaar M. Molecular characterization of a peripheral receptor for cannabinoids. Nature. 1993; 365:61–5. [CrossRef]
- Shahbazi, F., Grandi, V., Banerjee, A., Trant, J. Cannabinoids and Cannabinoid Receptors: The Story so Far. IScience. 2020, 23, 101301. [CrossRef]
- Izzo, A.A., Sharkey, K.A. Cannabinoids and the gut: New developments and emerging concepts. Pharmacol. Ther. 2010, 126, 21–38. [CrossRef]
- Di Marzo, V., de Petrocellis, L. Why do cannabinoid receptors have more than one endogenous ligand? Philos. Trans. R. Soc. B 2012, 367, 3216–3228. [CrossRef]
- Mukhopadhyay S., Shim J. Y., Assi A. A., Norford D., Howlett A. C. CB1 cannabinoid receptor-g protein association: a possible mechanism for differential signaling. Chemistry and Physics of Lipids. 2002, 121: 61-65. [CrossRef]
- Cacciola G, Chianese R, Chioccarelli T, Ciaramella V. Fasano S, Pierantoni R, Meccariello R, Cobellis G.Cannabinoids and Reproduction: A Lasting and Intriguing History. Pharmaceuticals. 2010 3: 3275-332. [CrossRef]
- Migliarini, B., Carnevali, O. Anandamide modulates growth and lipid metabolism in the zebrafish Danio rerio. Molecular and Cellular Endocrinology. 2008, 286S, S12–S16. [CrossRef]
- Fattore, L., Martellotta, C., Cossu, G., Mascia, M.S., Fratta, W. CB1 cannabinoid receptor agonist WIN 55, 212-2 decreases intravenous cocaine self-administration in rats. Behavioural Brain Research. 1999, 104, 141–146. [CrossRef]
- Gonzàlez, S., Fernàndez-Ruiz, J., Sparpaglione, V., Parolaro, D., Ramos, J. Chronic exposure to morphine, cocaine or ethanol in rats produced different effects in brain cannabinoid CB1 receptor binding and mRNA levels. Drug and Alcohol Dependence. 2002, 66, 77–84. [CrossRef]
- Centonze, D., Battista, N., Rossi, S., Mercuri, N.B., Finazzi-Agro, A., Bernardi, G., Calabresi, P., Maccarrone, M. A critical interaction between Dopamine D2 receptors and endocannabinoids mediates the effects of cocaine on striatal GABAergic transmission. Neuropsychopharmacology. 2004, 29, 1488–1497. [CrossRef]
- Soria G., Mendizabal, V., Tourin, C., Robledo, P., Ledent, C., Parmentier, M., Maldonado, R., Valverde, O. Lack of CB1 cannabinoid receptor impairs cocaine self-administration. Neuropsychopharmacology. 2005, 30, 1670–1680. [CrossRef]
- Tanda, G. Modulation of the endo-cannabinoid system: therapeutic potential against cocaine dependence. Pharmacol Res. 2007, 56(5): 406–417. [CrossRef]
- Harvey-Girard, E., Giassi, A.C.C, Ellis, W., Maler, L. Expression of the cannabinoid CB1 receptor in the Gymnotiform fish brain and its implications for the organization of the teleost pallium. The Journal of Comparative Neurology-Research in Systems Neuroscience. 2013, 521:949–975. [CrossRef]
- Henkel C., Jong-Raadsen S.A., Dufour S., Weltzien F.-A., Palstra A.P., Pelster B., Spaink H.P., Van Den Thillart G.E., Jansen H., Zahm M., Klopp C., Cedric C., Louis A., Berthelot C., Parey E., Roest Crollius H., Montfort J., Robinson-Rechavi M., Bucao C., Bouchez O., Gislard M., Lluch J., Milhes M., Lampietro C., Lopez Roques C., Donnadieu C., Braasch I., Desvignes T., Postlethwait J., Bobe J., Guiguen Y., Dirks R. A chromosome-scale assembly of European eel, Anguilla anguilla. UniProt: A0A9D3MIM1_ANGAN; submitted to MBL/GenBank/DDBJ databases JAN-2021. https://www.uniprot.org/citations/CI-4V42E84B85DHN.
- Gheorghe, A., van Nuijs, A., Pecceu, B., Bervoets, L., Jorens, P.G., Blust, R., Neels, R., Covaci, A. Analysis of cocaine and its principal metabolites in waste and surface water using solid-phase extraction and liquid chromatography–ion trap tandem mass spectrometry. Anal Bioanal Chem.2008, 391:1309–1319. [CrossRef]
- Chianese, T., Trinchese, G., Leandri, R., De Falco, M., Mollica, M.P., Scudiero, R., Rosati, L. Glyphosate Exposure Induces Cytotoxicity, Mitochondrial Dysfunction and Activation of ERα and ERβ Estrogen Receptors in Human Prostate PNT1A Cells. Int. J. Mol. Sci. 2024, 25, 7039. [CrossRef]
- d’Aquino, I., Piegari, G., Miletti, G., Sannino, E., Costanza, D., Meomartino, L., Fico, R., Riccio, L., Vaccaro, E., De Biase, D., Paciello, O. Morphometrical and Immunohistochemical Evaluation of Kidney as an Indirect Parameter to Estimate Age in Puppies in Veterinary Forensic Pathology. Animals. 2023, 13(16), 2665. [CrossRef]
- Spampinato, M., Siciliano, A., Travaglione, A., Chianese, T., Mileo, A., Libralato, G., Guida, M., Trifuoggi, M., De Gregorio, V., Rosati L. Unravelling the ecotoxicological impacts of gadolinium (Gd) on Mytilus galloprovincialis embryos and sperm in seawater: A preliminary study. Heliyon. 2024, 17;10(10):e31087. [CrossRef]
- Johnson, S.J., Walker, F. R. Strategies to improve quantitative assessment of immunohistochemical and immunofluorescent labelling. Scientific reports. 2015, 5(1), 10607. [CrossRef]
- Law, A.M., Yin, J.X., Castillo, L., Young, A.I., Piggin, C., Rogers, S., Caldon, C.E., Burgess, A., Millar, E.K.A., O'Toole, S.A., Gallego-Ortega, D., Ormandy, C.J., Oakes, S.R. Andy’s Algorithms: new automated digital image analysis pipelines for FIJI. Scientific reports. 2017, 7(1), 15717. [CrossRef]
- Lutz B. Molecular biology of cannabinoid receptors. Prostaglandins Leukot Essent Fat Acids. 2002, 66(2–3):123–42. [CrossRef]
- Yamaguchi F., Macrae A.D., Brenner S. Molecular cloning of two cannabinoid type 1-like receptor genes from the puffer fish Fugu rubripes. Genomics. 1996, 35:603-605. [CrossRef]
- Elphick M. R. Evolution of cannabinoid receptors in vertebrates: identification of a CB2 gene in the puffer fish Fugu rubripes. Biol Bull. 2002, 202:104-107. [CrossRef]
- Lachowicz, J., Szopa, A., Ignatiuk, K., Swiader, K., Serefko, A. Zebrafish as an animal model in cannabinoid research. Int. J. Mol.Sci. 2023, 24, 10455. [CrossRef]
- Casu, M.A., Porcella, A., Ruiu, S., Saba, P., Marchese, G., Carai, M.A.M., Reali, R., Gessa, G.L., Pani, L. Differential distribution of functional cannabinoid CB1 receptors in the mouse gastroenteric tract. Eur. J. Pharmacol. 2003. 459, 97-105. [CrossRef]
- Tesch, S.W. The Eel. Blackwell Science, Ltd, Oxford. 2003. [CrossRef]
- Di Marzo, V., Matias, I. Endocannabinoid control of food intake and energy balance. Nature Neurosci. 2005, 8(5), 585-589. [CrossRef]
- Di Patrizio, N. Endocannabinoids in the Gut. Cannabis and Cann. Res, 2016, 1.1, 67-77. [CrossRef]
- Gay, F., Ferrandino, I., Monaco, A., Cerulo, M., Capasso, G., Capaldo, A. Histological and hormonal changes in the European eel (Anguilla anguilla) after exposure to environmental cocaine concentration. J. Fish Dis. 2016, 39, 295-308. [CrossRef]
- Becker, W., Alrafas, H.R., Busbee, P.B., Walla, M.D., Wilson, K., Miranda, K., Cai, G., Putluri, V., Putluri, N., Nagarkatti, M., Nagarkatti, P. Cannabinoid Receptor Activation on Haematopoietic Cells and Enterocytes Protects against Colitis. J. Chrohn’s Colitis. 2021, 15(6), 1032-1048. [CrossRef]
- Arinç, E., Bozcaarmutlu, A. Catalyzation of Cocaine N-Demethylation by Cytochromes P4502B, P4503A, and P4502D in Fish Liver. J. Biochem. Mol. Toxicol. 2003, 17(3), 169-176. [CrossRef]
- Tam, J., Liu, J., Mukhopadhyay S., Cinar, R., Godlewski, G., Kunos, G. Endocannabinoids in Liver Disease. Hepatology. 2011, 53(1), 346-355. [CrossRef]
- Bazwinsky-Wutschke, I., Zipprich, A., Dehghani, F. Endocannabinoid System in Hepatic Glucose Metabolism, Fatty Liver Disease, and Cirrhosis. Int.J.Mol.Sci. 2019, 20(10), 2516. [CrossRef]
- Howlett, A.C., Abood, M.E. CB1 & CB2 Receptor Pharmacology. Adv. Pharmacol. 2017, 80, 169-206. [CrossRef]
- Pagotto, U., Pasquali, R. Endocannabinoids and energy metabolism. J. Endocrinol. Invest. 2006, 29 (3), 58-68. PMID: 16751710.
- Capaldo A., Gay F., Caputo I., Lionetti L., Paolella G., Di Gregorio I., Martucciello S., Di Lorenzo M., Rosati L., Laforgia V. Effects of environmental cocaine concentrations on COX and caspase-3 activity, GRP-78, ALT, CRP and blood glucose levels in the liver and kidney of the European eel (Anguilla anguilla). Ecotoxicology and Environmental Safety. 2021, 208, 111475. [CrossRef]
- Mumford, S., Heidel, J., Smith, C., Morrison, J., Mac Connell, B., Blazer, V. Fish Histology and Histopathology. U.S. Fish & Wildlife Service-National Conservation Training Center. 2007. https://nctc.fws.gov/resources/course-resources/fish-histology/.
- Watanabe, T., Takei, Y. Molecular physiology and functional morphology of SO4 2– excretion by the kidney of seawater-adapted eels. J. Exper.Biol. 2011, 214, 1783-1790. [CrossRef]
- Jiang, S., Fu, Y., Avraham, H.K. Regulation of hematopoietic stem cell trafficking and mobilization by the endocannabinoid system. Transfusion 2011, 51 supplement, 65S-71S. [CrossRef]
- Cottone, E., Pomatto, V., Cerri, F., Campantico, E., Mackie, K., Delpero, M., Guastalla, A., Dati, C., Bovolin, P., Franzoni, M.F. Cannabinoid receptors are widely expressed in goldfish: molecular cloning of a CB2-like receptor and evaluation of CB1 and CB2 mRNA expression profiles in different organs. Fish Physiol Biochem. 2013, 39:1287–1296. [CrossRef]
- Tam, J. The emerging role of the endocannabinoid system in the pathogenesis and treatment of kidney diseases. J. Basic Clin. Physiol. Pharmacol. 2015, 27(3), 267-276. [CrossRef]
- Dao, M., Francois, H. Cannabinoid Receptor 1 Inhibition in Chronic Kidney Disease: A New Therapeutic Toolbox. Front. Endocrinol. 2021, 12:720734. [CrossRef]
- Arceri, L., Nguyen, T.K., Gibson, S., Baker, S., Wingert, R.A. Cannabinoid Signaling in Kidney Disease. Cells. 2023, 12, 1419. [CrossRef]
- Valente, M.J., Henrique, R., Vilas-Boas, V., Silva, R., de lourdes Bastos, M., Carvalho, F., Guedes de Pinho, P., Carvalho, M. Cocaine-induced kidney toxicity: an in vitro study using primary cultured human proximal tubular epithelial cells. Arch. Toxicol. 2012, 86 (2), 249–261. [CrossRef]
- Cianchi, F., Papucci, L., Schiavone, N., Lulli, M., Magnelli, L., Vinci, M.C., Messerini, L., Manera, C., Ronconi, E., Romagnani, P., Donnini, M., Perigli, G., Trallori, G., Tanganelli, E., Capaccioli, S., Masini, E. Cannabinoid receptor activation induces apoptosis through tumor necrosis factor α-mediated ceramide de novo synthesis in colon cancer cells. Clin. Cancer Res. 2008, 14(23), 7691-7700. [CrossRef]
- Fontes, M. K., Rosati, L., Di Lorenzo, R., Seabra Pereira, C.D., Maranho, L. A., Laforgia, V., Capaldo, A.Aquatic Pollution and Risks to Biodiversity: The Example of Cocaine Effects on the Ovaries of Anguilla anguilla. Animals 2022b, 12(14), 1766. [CrossRef]
- Walker, O.S., Holloway, A.C., Raha, S. The role of the endocannabinoid system in female reproductive tissues. J. Ovar. Res., 2019, 12, 3. [CrossRef]
- Rosati, L., Chianese, T., Mileo, A., De Falco, M., Capaldo, A. Cocaine Effects on Reproductive Behavior and Fertility: An Overview. Vet. Sci. 2023, 10, 484. [CrossRef]
- Di Blasio, A.M., Vignali, M., Gentilini, D. The endocannabinoid pathway and the female reproductive organs. J Mol Endocrinol. 2013, 11;50(1), R1-9. [CrossRef]
- Mendizabal-Zublaga, J.M., Melser, Su., Bènard, G., Ramos, A., Reguero, L., Arrabal, S., Elezgaral, I., Gerrikagoitia, I., Suarez, J., De Fonseca, F.R., Puente, N., Marsicano, G., Grandes, O. Cannabinoid CB1 Receptors Are Localized in Striated Muscle Mitochondria and Regulate Mitochondrial Respiration. Front. Physiol. 2016, 7, 476. [CrossRef]
- Cavuoto, P., McAinch, A.J., Hatzinikolas, G., Cameron-Smith, G.A., Wittert, G.A. Effects of cannabinoid receptors on skeletal muscle oxidative pathways. Mol. Cell Endocrinol. 2007a, 267, 63-69. [CrossRef]
- Cavuoto, P., McAinch, A.J., Hatzinikolas, G., Janovskà, A., Game, P., Wittert, G.A. The expression of receptors for endocannabinoids in human and rodent skeletal muscle. Biochem. Biophys. Res Comm. 2007b, 364, 105-110. [CrossRef]
- Dalle, S., Schouten, M., Meeus, G., Slagmolen, L., Koppo, K. Molecular networks underlying cannabinoid signaling in skeletal muscle plasticity. J. Cell. Physiol.2022, 237, 3517–3540. [CrossRef]
- de Abreu, N.K., Fabro Feltrin, I., Russiano Pereira, D.B., Penasso bezerra, P., Aguiar Jr, A.S. Impact of CB1 receptor antagonism on skeletal muscle hypertrophy and metabolic health: a systematic review of preclinical studies. Hormones. 2025, 1-12. [CrossRef]
- Tomiyama, K.I, Funada M, Synthetic cannabinoid CP-55,940 induces apoptosis in a human skeletal muscle model via regulation of CB1 receptors and l-type Ca2+channels, Arch. Toxicol. 2021, 95, 617-630. [CrossRef]
- Capaldo, A., Gay, F., Lepretti, M., Paolella, G., Martucciello, S., Lionetti, L., Caputo, I., Laforgia, V. Effects of environmental cocaine concentrations on the skeletal muscle of the European eel (Anguilla anguilla). Sci. Total Environ. 2018, 640-641, 862-873. [CrossRef]
- Birò, T., Tòth, B., Haskò, G., Paus, R., Pacher, P. The endocannabinoid system of the skin in health and disease: novel perspectives and therapeutic opportunities. Trends Pharmacol Sci. 2009, 30(8), 411-420. [CrossRef]
- Kupczyk, P., Reich, A., Szepietowski, J.C. Cannabinoid system in the skin – a possible target for future therapies in dermatology. Exp. Dermatol. 2009, 18, 669-679. [CrossRef]
- Rodriguez-Martin, I., Herrero-Turrion, M.J., Marron Fdez de Velasco, E., Gonzalez-Sarmiento, R., Rodriguez, R.E. Characterization of two duplicate zebrafish Cb2-like cannabinoid receptors. Gene. 2007, 389, 36-44. [CrossRef]
- Capaldo, A., Gay, F., Laforgia, V. Changes in the gills of the European eel (Anguilla anguilla) after chronic exposure to environmental cocaine concentration. Ecotoxicol. Environ. Saf. 2019, 169, 112-119. [CrossRef]
Figure 1.
Immunoblot on Anguilla anguilla kidney proteins. The antibody reacts with a band of about 52 kDa corresponding to the molecular weight of CB1R. M: known molecular weight protein markers. C: negative control shows no band.
Figure 1.
Immunoblot on Anguilla anguilla kidney proteins. The antibody reacts with a band of about 52 kDa corresponding to the molecular weight of CB1R. M: known molecular weight protein markers. C: negative control shows no band.
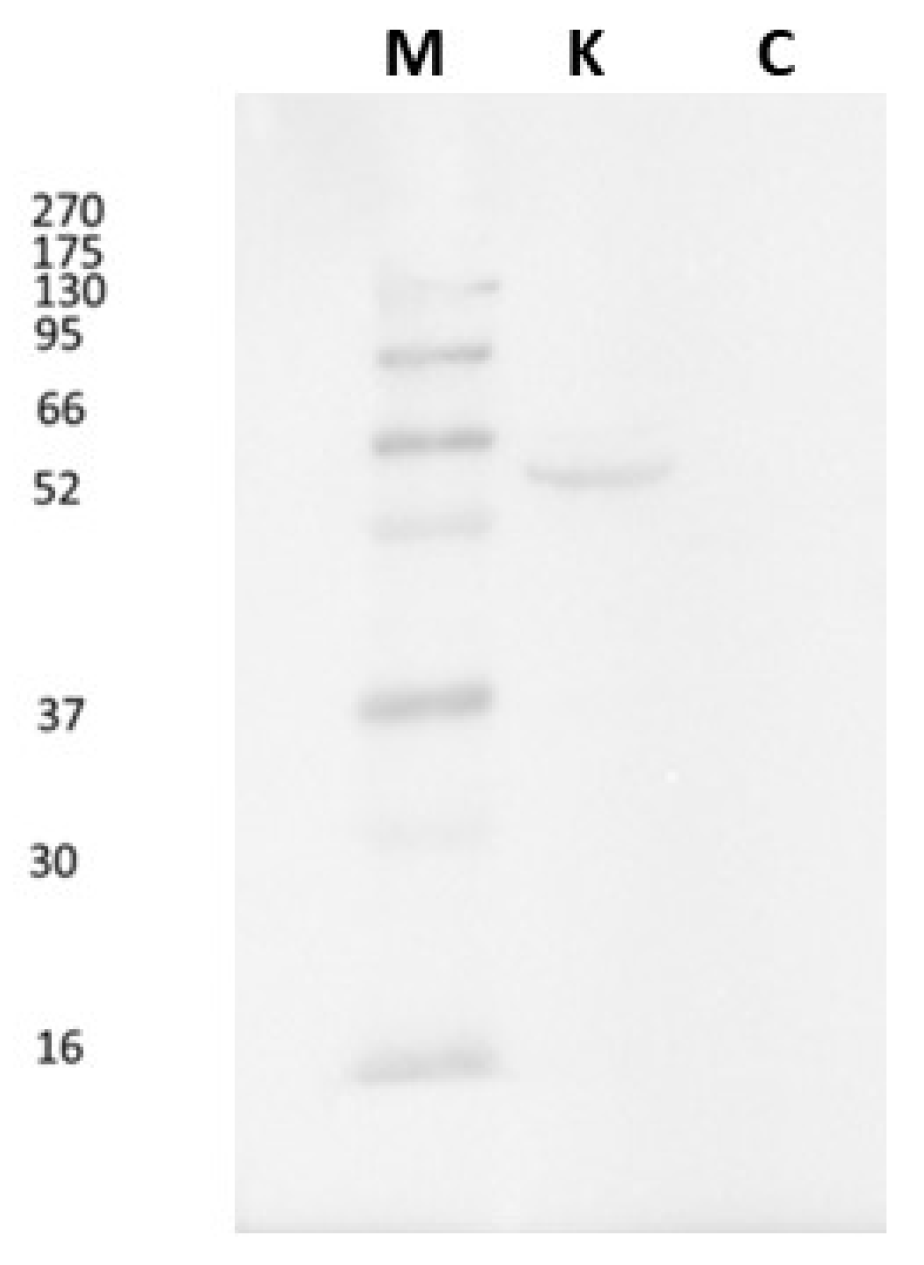
Figure 2.
Light micrographs of A. anguilla gut and liver. Immunoreactivity to CB1R. a, c, e: control specimens; b, d, f: exposed specimens. Insets: 1, 2, 3, 4: negative controls of a, b, e, f respectively. In the gut of control specimens (a, c), the immunoreactivity was almost absent, both in epithelium (E) and in smooth muscle (M) tissue. In the exposed specimens (b, d), the smooth muscle (M) tissue and the epithelium (E) appeared immunopositive. C; connective tissue. The hepatic parenchyma of cocaine-exposed specimens (f) showed an increased immunopositivity to CB1R compared with control specimens (e). g, h: semi-quantitative analysis of gut and liver immunoreactivity, respectively. The Mann Whitney test showed a statistically significant difference among the control and the exposed specimens. ** p < 0.01 (0.079); * p < 0.05 (0.0397).
Figure 2.
Light micrographs of A. anguilla gut and liver. Immunoreactivity to CB1R. a, c, e: control specimens; b, d, f: exposed specimens. Insets: 1, 2, 3, 4: negative controls of a, b, e, f respectively. In the gut of control specimens (a, c), the immunoreactivity was almost absent, both in epithelium (E) and in smooth muscle (M) tissue. In the exposed specimens (b, d), the smooth muscle (M) tissue and the epithelium (E) appeared immunopositive. C; connective tissue. The hepatic parenchyma of cocaine-exposed specimens (f) showed an increased immunopositivity to CB1R compared with control specimens (e). g, h: semi-quantitative analysis of gut and liver immunoreactivity, respectively. The Mann Whitney test showed a statistically significant difference among the control and the exposed specimens. ** p < 0.01 (0.079); * p < 0.05 (0.0397).
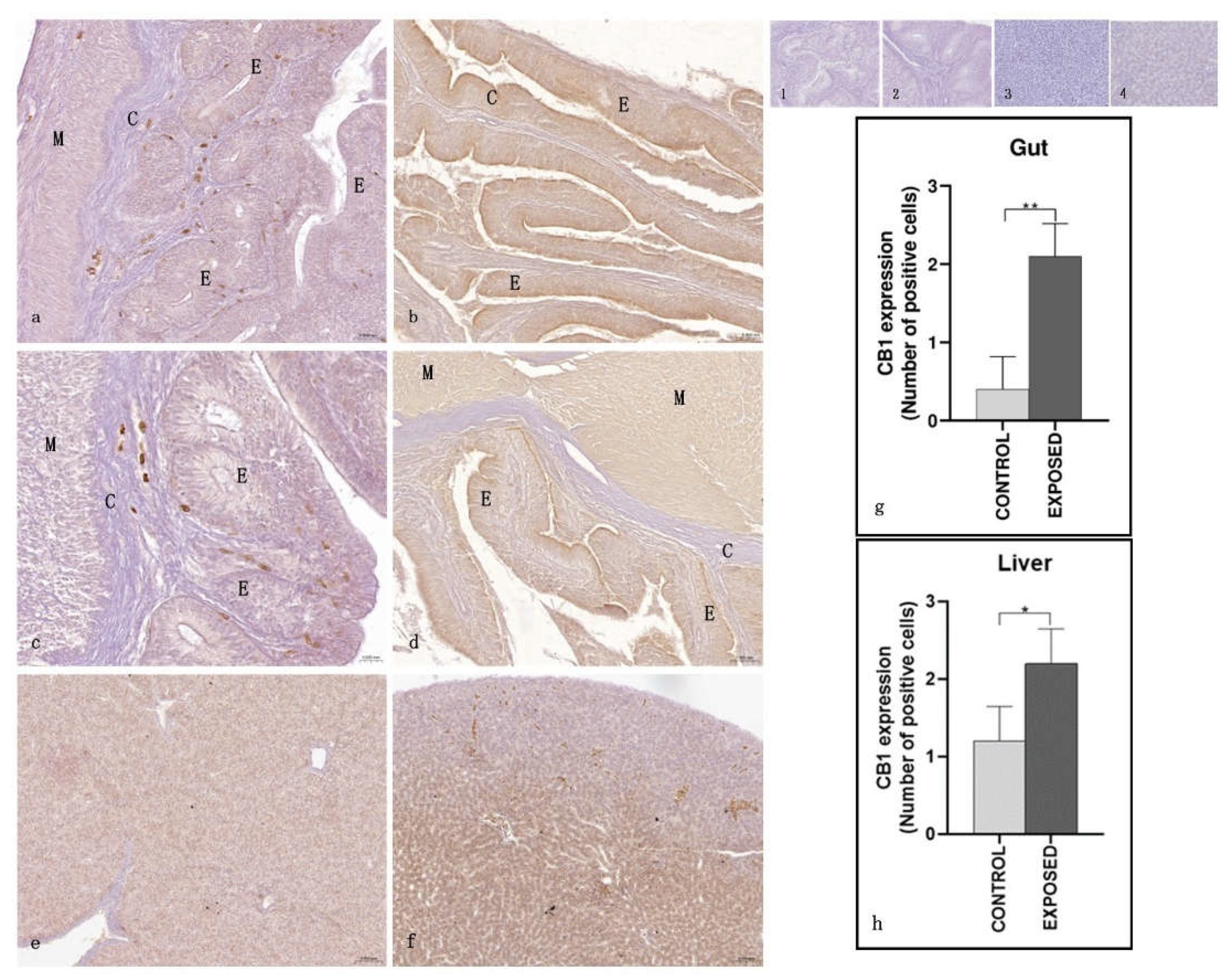
Figure 3.
Light micrographs of A. anguilla kidney. Immunoreactivity to CB1R. a, c: control specimens; b, d: exposed specimens. Insets: 1, 2: negative control of a, b, respectively. In the control specimens (a, c), the hematopoietic (H) tissue appeared immunopositive to CB1R, while the proximal (arrow) and the distal (asterisk) tubules) were not. In the exposed specimens (b, d), the immunopositivity of the hematopoietic (H) tissue appeared absent/faint, while the proximal (arrow) tubules were generally immunopositive, and the distal (asterisk) tubules were generally not. M: melanomacrophage centers. e: semi-quantitative analysis of immunoreactivity. The Mann Whitney test showed a statistically significant difference among the control and the exposed specimens. * p < 0.05 (0.0159).
Figure 3.
Light micrographs of A. anguilla kidney. Immunoreactivity to CB1R. a, c: control specimens; b, d: exposed specimens. Insets: 1, 2: negative control of a, b, respectively. In the control specimens (a, c), the hematopoietic (H) tissue appeared immunopositive to CB1R, while the proximal (arrow) and the distal (asterisk) tubules) were not. In the exposed specimens (b, d), the immunopositivity of the hematopoietic (H) tissue appeared absent/faint, while the proximal (arrow) tubules were generally immunopositive, and the distal (asterisk) tubules were generally not. M: melanomacrophage centers. e: semi-quantitative analysis of immunoreactivity. The Mann Whitney test showed a statistically significant difference among the control and the exposed specimens. * p < 0.05 (0.0159).
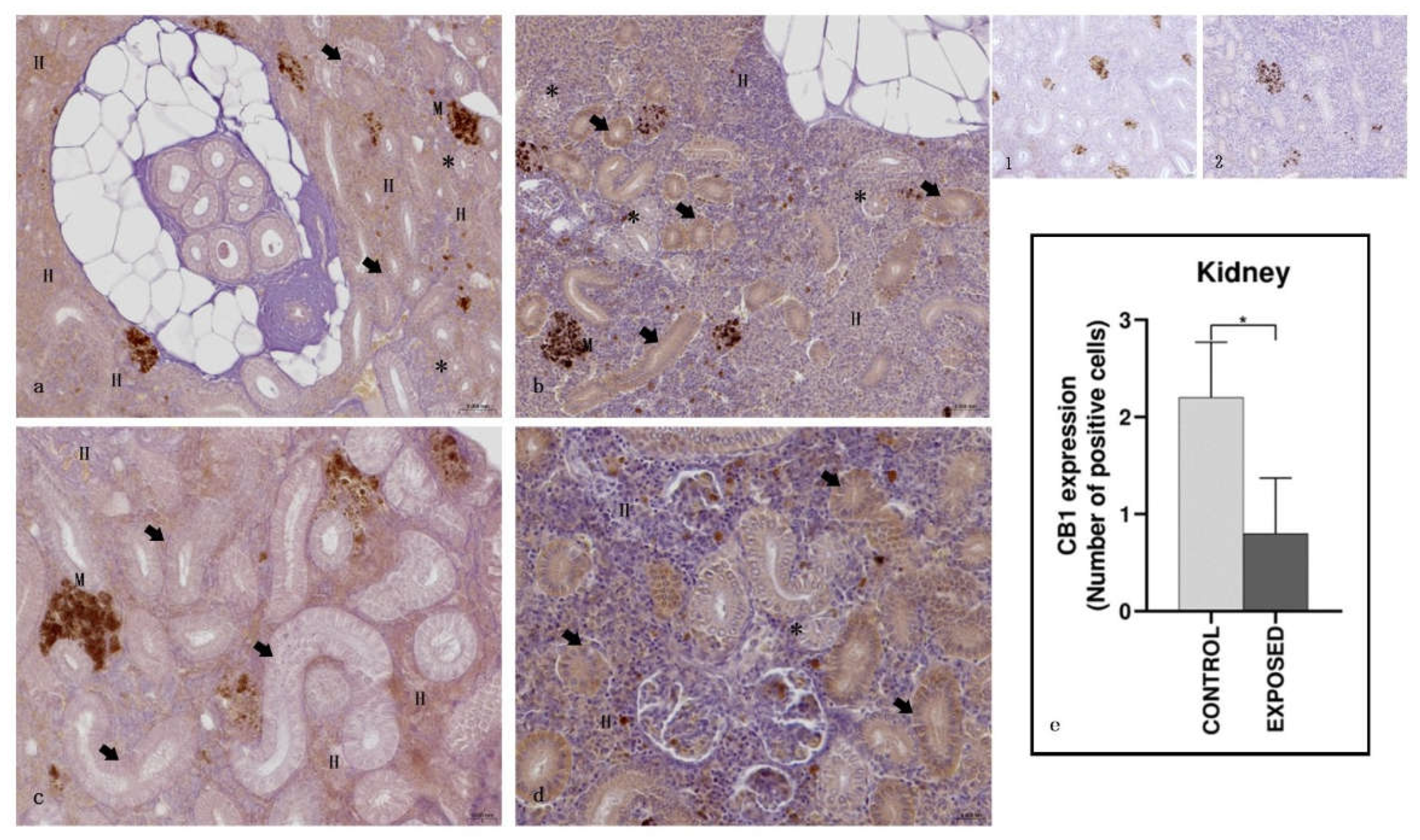
Figure 4.
Light micrographs of A. anguilla ovary. Immunoreactivity to CB1R. a, c: control specimens; b, d: exposed specimens. Insets: 1, 2: negative control of a, b, respectively. In control specimens (a, c), CB1R signaling was mainly observed in previtellogenic (pvOo) oocytes and mid vitellogenic oocytes (mvOo), with no immunopositivity detected in early vitellogenesis (evOo) or full vitellogenesis (fvOo) oocytes. In contrast, the exposed specimens (b, d) showed immunoreactivity to CB1R only in previtellogenic oocytes (pvOo), while early vitellogenic oocytes (evOo) remained negative. e: semi-quantitative analysis of immunoreactivity. The Mann Whitney test showed a statistically significant difference among the control and the exposed specimens. * p < 0.05 (0,0286).
Figure 4.
Light micrographs of A. anguilla ovary. Immunoreactivity to CB1R. a, c: control specimens; b, d: exposed specimens. Insets: 1, 2: negative control of a, b, respectively. In control specimens (a, c), CB1R signaling was mainly observed in previtellogenic (pvOo) oocytes and mid vitellogenic oocytes (mvOo), with no immunopositivity detected in early vitellogenesis (evOo) or full vitellogenesis (fvOo) oocytes. In contrast, the exposed specimens (b, d) showed immunoreactivity to CB1R only in previtellogenic oocytes (pvOo), while early vitellogenic oocytes (evOo) remained negative. e: semi-quantitative analysis of immunoreactivity. The Mann Whitney test showed a statistically significant difference among the control and the exposed specimens. * p < 0.05 (0,0286).
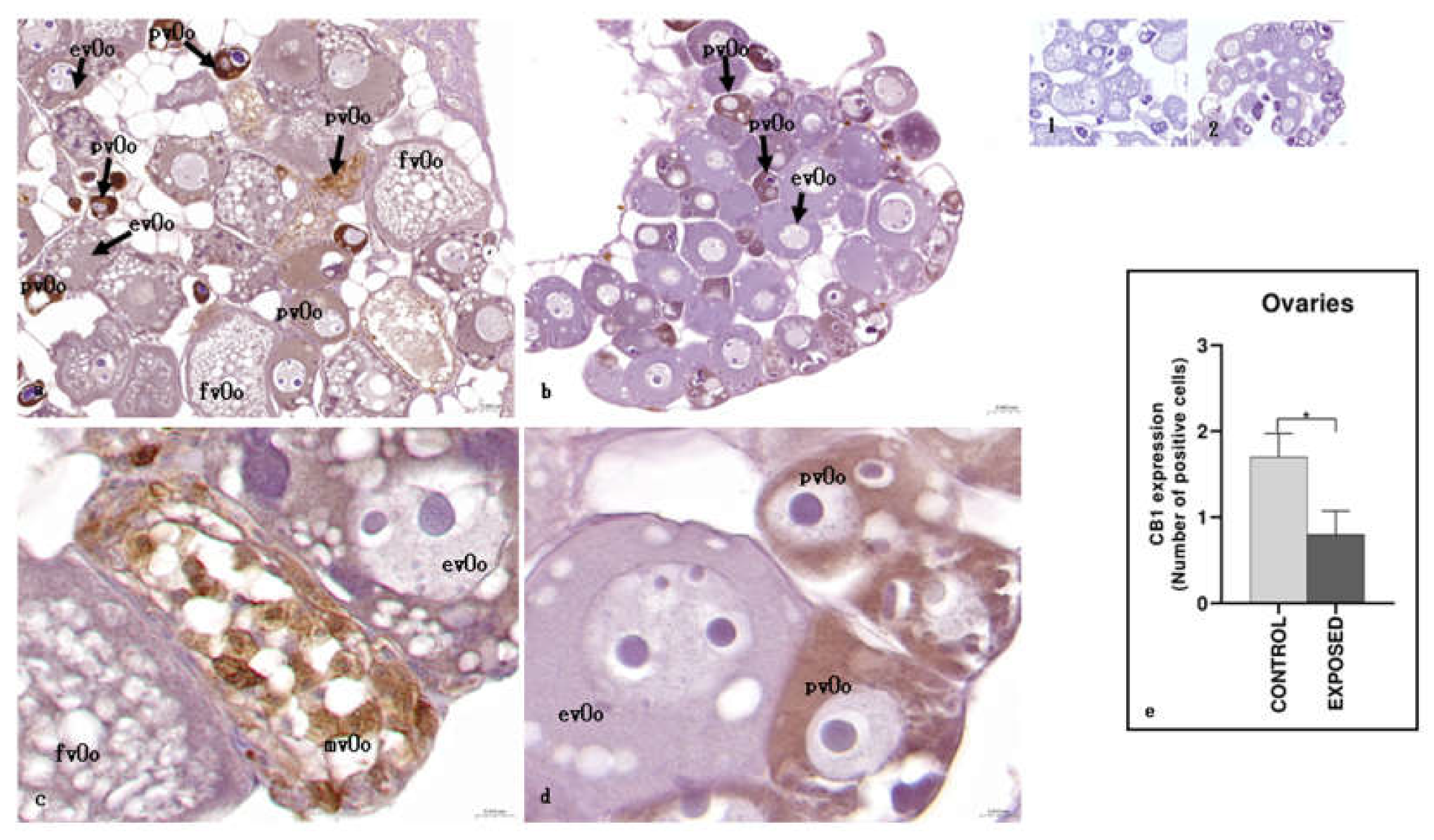
Figure 5.
Light micrographs of A. anguilla skeletal muscle. Immunoreactivity to CB1R. a, c: control specimens; b, d: exposed specimens; a,b: white fibers; c,d: red fibres. Insets: 1, 2, 3, 4: negative control of a, b, c, d, respectively. The white fibres (WF) of control specimens (a) appear positive to CB1R, also after cocaine exposure (b), when they show an equal or slightly reduced positivity. The red fibres (RF) appear negative in the control specimens (c) and then become positive for CB1R after exposure to cocaine (d). The adipose tissue (AT) appears always negative. e: semi-quantitative analysis of immunoreactivity. The Mann Whitney test showed a statistically significant difference among the control and the exposed specimens (p < 0.05). White fibers of control specimens vs. white fibers of exposed specimens: * p = 0.0476; red fibers of control specimens vs. white fibers of exposed specimens: * p = 0.0238.
Figure 5.
Light micrographs of A. anguilla skeletal muscle. Immunoreactivity to CB1R. a, c: control specimens; b, d: exposed specimens; a,b: white fibers; c,d: red fibres. Insets: 1, 2, 3, 4: negative control of a, b, c, d, respectively. The white fibres (WF) of control specimens (a) appear positive to CB1R, also after cocaine exposure (b), when they show an equal or slightly reduced positivity. The red fibres (RF) appear negative in the control specimens (c) and then become positive for CB1R after exposure to cocaine (d). The adipose tissue (AT) appears always negative. e: semi-quantitative analysis of immunoreactivity. The Mann Whitney test showed a statistically significant difference among the control and the exposed specimens (p < 0.05). White fibers of control specimens vs. white fibers of exposed specimens: * p = 0.0476; red fibers of control specimens vs. white fibers of exposed specimens: * p = 0.0238.
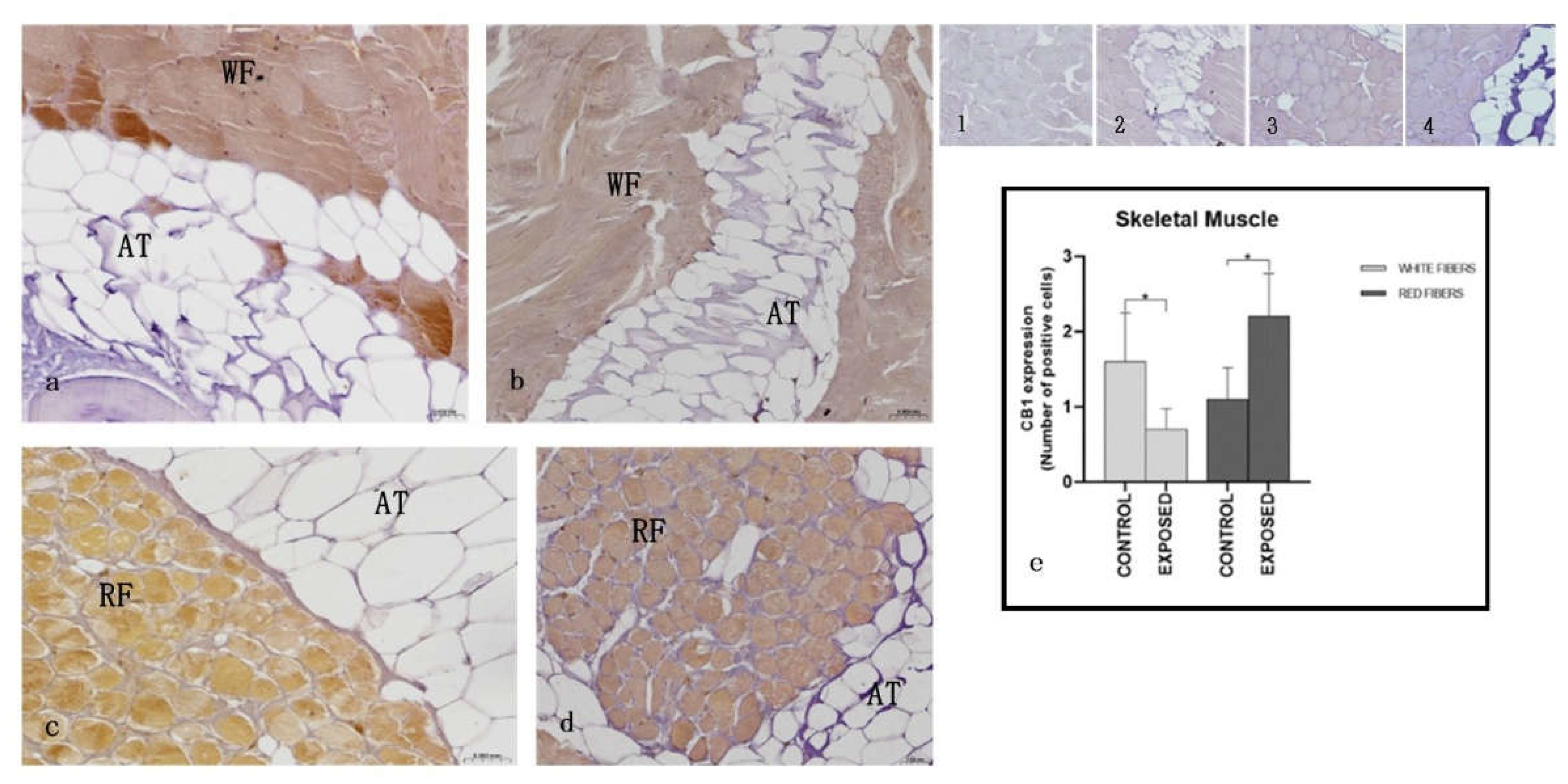
Figure 6.
Light micrographs of A. anguilla skin and gills. Immunoreactivity to CB1R. a, c: control specimens; b, d: exposed specimens. Insets: 1, 2, 3, 4: negative control of a, b, c, d respectively. In the skin of control (a) and exposed (b) specimens, the epithelium (E) appeared immunopositive, with keratinocytes (K) and club (C) cells well stained. M: mucous cells; S: scales. The gills of both control (c) and exposed (d) specimens showed CB1R immunopositivity in the primary (PL) and secondary (S) lamellae. e, f: semi-quantitative analysis of skin and gills immunoreactivity, respectively. The Mann Whitney test didn’t show a statistically significant difference among the control and the exposed specimens (p < 0.05).
Figure 6.
Light micrographs of A. anguilla skin and gills. Immunoreactivity to CB1R. a, c: control specimens; b, d: exposed specimens. Insets: 1, 2, 3, 4: negative control of a, b, c, d respectively. In the skin of control (a) and exposed (b) specimens, the epithelium (E) appeared immunopositive, with keratinocytes (K) and club (C) cells well stained. M: mucous cells; S: scales. The gills of both control (c) and exposed (d) specimens showed CB1R immunopositivity in the primary (PL) and secondary (S) lamellae. e, f: semi-quantitative analysis of skin and gills immunoreactivity, respectively. The Mann Whitney test didn’t show a statistically significant difference among the control and the exposed specimens (p < 0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



