
Submitted:
17 June 2025
Posted:
19 June 2025
You are already at the latest version
Abstract
The amygdala is composed of several nuclei, including the lateral nucleus which is the main receiving area for the input from cortical and subcortical brain regions. It mediates fear, anxiety, stress, and pain across species. Evidence suggests that the endocannabinoid system (ECS) may be a promising target for modulating these processes. Cannabinoid and cannabinoid-related receptors have been identified in the amygdala of rodents, carnivores, and humans, but not in horses. This study aimed at investigating the gene expression of cannabinoid receptors 1 (CB1R) and 2 (CB2R), transient receptor potential vanilloid 1 (TRPV1), and peroxisome proliferator-activated receptor gamma (PPARɣ) within the lateral nucleus of six equine amygdalae collected post mortem from an abattoir using quantitative Real-Time PCR (RT-PCR), cellular distribution, and immunofluorescence. mRNA expression of CB1R and CB2R, but not TRPV1 or PPARɣ, was detected. The percentage of immunoreactivity (IR) was calculated using ImageJ software. Cannabinoid receptor 1 immunoreactivity was absent in the somata but was strongly detected in the surrounding neuropil and varicosities and CB2R-IR was observed in the varicosities; TRPV1-IR showed moderate expression in the cytoplasm of somata and processes, while PPARγ-IR was weak-to-moderate in the neuronal nuclei. These findings demonstrate ECS components in the equine amygdala and may support future studies on Cannabis spp. molecules acting on these receptors
Keywords:
cannabinoid receptor 1
; cannabinoid receptor 2
; equine
; peroxisome proliferator-activated receptor gamma
; transient receptor potential vanilloid type 1
1. Introduction
The amygdala, or amygdaloid complex, is an anatomically and functionally heterogeneous structure [1] located in the medial part of the temporal lobe [2]. Current neurochemical, functional and developmental data suggest that the amygdala is composed of pallial and subpallial structures. The pallial amygdala is composed of deep and superficial nuclei. The deep pallial nuclei include the basolateral amygdala, the anterior amygdaloid area and the amygdalohippocampal area. The basolateral amygdala (BLA) includes the lateral, basal and accessory basal nuclei [2,3]. The lateral nucleus is the main receiving structure of the amygdala. In particular, it collects most of the input coming from other brain regions and provides stimuli to other amygdalar nuclei and higher brain structures [4,5,6]. The lateral nucleus is critical for the formation of emotional memories and the generation of appropriate behavioral responses to salient sensory stimuli and emotionally arousing events in the external world, including nociceptive information [7,8,9,10,11,12,13]. The growing interest in amygdala function and dysfunction, particularly in relation to neuropsychiatric disorders, has stimulated numerous investigations aimed at elucidating how amygdala processing of emotional stimuli is organized, both in terms of connectivity, and at the cellular and molecular level. In horses, the amygdala is relatively well developed [14] and may be of great importance in this prey species, which is particularly alert to signs of threat and responds with rapid avoidance behavior [15]. However, little is known about the neurochemical properties of the equine amygdala [14,15].
The endocannabinoid system (ECS) consists of cannabinoid receptors (CB1R, CB2R) and cannabinoid-related receptors, their ligands (anandamide, AEA; 2-arachidonoylglycerol, 2-AG; and other ligands), and the enzymes necessary for their synthesis and degradation. Cannabinoid-related receptors include transient receptor potential (TRP) channels, G protein-coupled receptors (GPRs), nuclear peroxisome proliferator-activated receptors (PPARs) and serotonin receptors [16,17,18,19,20]. In the past decade, the ECS has emerged as a key neuromodulatory system which regulates synaptic efficacy within and between the amygdaloid nuclei. The endocannabinoid system, interacting with other neuromodulators such as opioids, influences all vital bodily functions and sets the tone for all conscious and unconscious thoughts and feelings. The ECS is probably the most powerful and ancient modulatory system capable of influencing every conscious thought and feeling [16,17,18,19,20]. The molecular components of the ECS are abundantly expressed in the mammalian amygdaloid complex, consistent with the robust role of the ECS in modulating emotional behavior, learning and stress response physiology [21,22].
The morphofunctional knowledge of the equine amygdala could improve the understanding of the behavioral domain of this species, such as cognition, learning, stereotyping, separation anxiety, the impact of equine temperament on welfare and equine psychopharmacology. To date, no studies have been published regarding the expression of cannabinoid and cannabinoid-related receptors in the equine amygdaloid complex. Therefore, the present study was designed to evaluate the mRNA of Cnr1, Cnrr2, PPARɣ and TRPV1 gene expression on fixed amygdala samples, and to immunohistochemically localize these receptors in the equine amygdala, particularly in the extensively studied lateral nucleus of the amygdala.
2. Results
2.1. Quantitative Real-Time PCR (RT-PCR) for Cnr1, Cnr2, TRPV1, and PPARgamma
The reference utilized for data normalization was Glyceraldehyde-3-phosphate dehydrogenase (GAPDH), as hypoxanthine phosphoribosyltransferase 1 (HPRT) was undedectable and Actin was characterized by a huge variability in the samples. Quantitative Rea Time PCR showed that the Cnr1, and Cnr2 transcripts were detected in all the 4% paraformaldehyde fixed amigdala samples (n=6) (Figure 1). On the contrary, TRPV1 and PPARγ were undetectable.
2.2. Immunofluorescence
All the receptors expressed immunoreactivity in the lateral nucleus of the horse amygdaloid complex with, however, different degrees of intensity.
2.2.1. CB1R
Cannabinoid receptor 1 immunoreactivity (CB1R-IR) was brightly expressed by the neuropil which extended within the lateral nucleus. The CB1-IR neuropil was characterized by the classic basket shape surrounding the bodies of other neurons (Figure 2). The calculated percentage of immunoreactivity was 4.96±3.16%.
2.2.2. CB2R
Cannabinoid receptor 2 immunoreactivity (CB2R-IR) was moderately expressed by the neuropil within the lateral nucleus. The neuropil expressing CB2-IR was characterized by a diffuse punctate appearance (Figure 3). The calculated percentage of CB2R-IR was the lowest (as compared to the other receptors) 3.22±1.84%.
2.2.3. TRPV1
Transient receptor potential vanilloid 1 immunoreactivity (TRPV1-IR) was moderately expressed by the neuronal bodies and processes within the lateral nucleus; TRPV1-IR was distributed in the neuronal cell bodies and processes, which formed a reticular pattern (Figure 4). The calculated percentage of TRPV1-IR was the highest (as compared to the other receptors) 10.72±5.92%.
2.2.4. PPARɣ
Peroxisome proliferator-activated receptor gamma immunoreactivity (PPARɣ-IR) was highly expressed in the nuclei of neuronal cells and glial cells (Figure 5). Virtually all the NeuroTrace® labeled neuronal cell bodies showed PPARɣ-IR; the calculated percentage of PPARɣ-IR was 9.03±3.83%
Figure 6 shows the percentages of CB1R, CB2R, TPRV1 and PPARɣ immunostaining within the lateral nucleus of the horse amygdaloid complex.
3. Discussion
The mammalian amygdala is composed of more than a dozen nuclei which have distinctive connections [4,23]. The BLA consists of three main nuclei, the lateral nucleus, the basal nucleus and the accessory basal nucleus, each of which has several subdivisions [4]. The lateral nucleus is a cortical-like structure with two main types of neurons: spiny glutamatergic pyramidal projection neurons, and spine-sparse or spine-free GABAergic nonpyramidal neurons [24,25,26]. Immunohistochemical studies suggest that the BLA contains three largely non-overlapping interneuron populations which can be distinguished based on their expression of parvalbumin (PV), cholecystokinin (CCK) and somatostatin (SOM). Somatostatin-immunoreactive (IR) interneurons target distal dendrites [27], whereas PV- and CCK-IR perisomatically innervate local BLA excitatory pyramidal neurons [28,29]. In the amygdala, large CCK immunoreactive neurons are GABAergic interneurons, and they densely innervate pyramidal cells [30].
In general, cannabinoid receptors are located densely in the animal and human brain areas involved in the regulation of emotional states, and they induce neurochemical responses which may have anxiolytic or anxiogenic effects [31] as in other central and peripheral nervous system areas of the horse, such as trigeminal ganglion, dorsal root ganglia, and also in synovial membrane [32,33,34,35]. Typically, increased transmission at the CB1R reduces anxiety, whereas inhibition at the CB1R induces an anxiogenic effect [36,37]. Therefore, compounds which directly or indirectly regulate cannabinoid receptor activity have the potential of reducing anxiety and may be the focus of future pharmaceutical development of anxiolytic compounds in equine medicine. Since cannabidiol (CBD) does not act directly on CB1R [38], but rather as a hydrolyzer of fatty acid amide hydrolase (FAAH)which is the membrane enzyme hydrolyzing AEA [39], it could potentially reduce anxiety [31].
Fear extinction is a form of inhibitory learning in which newly formed associations compete with fearful ones to inhibit fear formation and fear-related neural circuits [40]. In rodents, the connections between the prefrontal cortex and the basolateral amygdala stimulate the endocannabinoid system during fear extinction [41]. Specifically, glutamate released by neurons in the prefrontal cortex causes the production of cannabinoids in the BLA which act presynaptically on CB1 receptors, leading to the extinction of the cued fear by suppressing additional glutamate release [42]. Endocannabinoid production during periods of stress may influence emotion by regulating transmission from the amygdala [43]; typically, anxiety increases endocannabinoid tone and endocannabinoids, in turn, decrease anxiety, as shown by Marsicano et al. [44]. In addition, endocannabinoids appear to be crucial for the extinction of aversive memories and this process is likely to be mediated by the amygdala [45]. In addition to its role in fear extinction, the ECS has also been implicated in fear-conditioned analgesia, a phenomenon whereby pain perception is suppressed in the presence of a fear-related context. In rodents, Rea et al. [46] demonstrated that CB1R activity in the BLA is necessary for this form of analgesia, as intra-BLA administration of a CB1R antagonist abolished the antinociceptive effect induced by contextual fear. Rubino et al. [47] showed that treatment with the phytocannabinoid ∆9-THC, an agonist of CB1R, significantly decreased c-Fos levels in the amygdala, and that this effect was reversed by the CB1 inverse agonist AM251. Taken together, these findings underscore the central role of CB1R in modulating emotional and nociceptive processes through the amygdaloid circuitry. The identification of CB1R protein expression within the lateral nucleus of the equine amygdala in the present study supports the hypothesis that similar regulatory mechanisms of fear, anxiety, and pain may be conserved in horses, providing a neuroanatomical foundation for the potential therapeutic use of cannabinoid-based compounds in equine behavioral medicine.
Although traditionally considered a peripheral receptor mainly involved in immune function, CB2R expression has been increasingly reported in the central nervous system (CNS), including neurons and glial cells, challenging the early notion of its absence from the brain [48,49]. In the present study, CB2R-IR was observed in varicosities within the lateral nucleus of the amygdala, and Cnr2 gene expression was detected by RT-PCR, supporting the presence of this receptor at both protein and transcript levels. These findings are in line with previous studies showing CB2R localization in the neuropil and dendritic compartments of several brain regions, including the amygdala, hippocampus, and substantia nigra of rodents [50,51]. Ultrastructural investigations have demonstrated that CB2R can localize postsynaptically, often associated with dendritic membranes and a rough endoplasmic reticulum, implying endogenous synthesis and functional integration within synaptic networks [50]. Although CB2R-IR expression is lower than that of CB1R, CB2R in the CNS appears to play a modulatory role, particularly under pathological conditions, such as neuroinflammation, stress, and neuropsychiatric disorders [51,52]. Functionally, CB2R activation in the brain has been associated with the regulation of microglial activity, immune response modulation, and neuroprotection [49]. Moreover, behavioral studies in animal models have suggested that CB2R agonists, such as β-caryophyllene, exert anxiolytic and antidepressant effects, reinforcing the potential involvement of CB2R in the modulation of emotional states [53]. Several lines of evidence support a functional role for CB2R within the amygdala. Its expression has been documented under both physiological and stress-related conditions [54,55], and CB2R activation in the central nucleus has been shown to mediate cannabinoid-induced antinociceptive effects [56]. In the medial amygdala, early-life stimulation of CB2R influences neuronal morphology and contributes to the development of social behaviors [57]. Although most of the research has focused on CB1R within the BLA, recent findings have suggested that CB2R activation in this region may also modulate stress-induced alterations in synaptic plasticity and affective responses. Specifically, intra-BLA administration of the CB1R/CB2R agonist WIN55,212-2 reversed stress-impaired hippocampal–accumbens long-term potentiation, suggesting that CB2R could contribute to the amygdaloid regulation of stress-related neural circuits [58]. While direct evidence of CB2R expression in the BLA, and especially in the lateral nucleus, remains limited, the present findings aligned with the broader observations of CB2R functional relevance across the amygdaloid complex [48], supporting the hypothesis that CB2R may also play a modulatory role in emotional and sensory integration within these subregions.
The transient receptor potential vanilloid type 1 receptor is a non-selective cation channel known for its role in nociception and thermoregulation. However, increasing evidence has revealed its expression in the brain areas involved in emotional regulation, including the amygdala [59]. In particular, TRPV1 has been identified in the lateral nucleus of the amygdala, where it modulates synaptic plasticity and long-term potentiation, thereby influencing fear memory formation in mice [59]. Although the precise physiological role of TRPV1 in the amygdala remains unclear, functional studies in TRPV1 knockout mice demonstrated reduced anxiety-like behavior, and diminished fear conditioning and stress-sensitization, additionally implicating this receptor in emotional processing circuits [60,61]. Transient receptor potential vanilloid type 1 signaling intersects with the ECS sinces it can be activated by anandamide, a key endocannabinoid molecule [62]. This convergence has prompted the development of dual FAAH/TRPV1 inhibitors which have shown dose-dependent antidepressant-like effects in rodent models of depression [63], underscoring the therapeutic relevance of this pathway. Beyond its role in emotional modulation, TRPV1 is also critically involved in pain perception and neurogenic inflammation. The activation of TRPV1 on the peripheral terminals of the primary sensory neurons leads to a rapid influx of calcium and sodium, triggering the release of neuropeptides, such as substance P, neurokinin A (NKA), and calcitonin gene-related peptide (CGRP) [64]. In addition to peripheral effects, TRPV1 activation in primay sensory neurons also modulates central nociceptive processing, supporting its role in the higher-order processing of nociceptive signals [64]. At the molecular level, TRPV1 protein has been shown to localize to the plasma membrane, cytoplasmic vesicles, and membranes of the endoplasmic reticulum in neurons, compartments which support its involvement in both calcium influx and intracellular calcium release [65] with some studies also describing its presynaptic localization in amygdaloid circuits [66]. These data are in line with the immunofluorescence results in the present study which revealed TRPV1 expression predominantly in the neuronal somata and neuropil within the lateral nucleus of the equine amygdala, with notable cytoplasmatic distribution patterns. The functional outcome of TRPV1 activation on synaptic transmission appears to be brain-region specific; as Edwards et al. [67] reported, TRPV1 can exert both excitatory and inhibitory effects on glutamatergic transmission depending on the neural context. Altogether, these findings support a multifaceted role for TRPV1 as a molecular integrator of nociceptive and emotional signals, and highlight its conserved expression and functional relevance within the amygdaloid circuitry across mammalian species, including the horse, as demonstrated by the present study.
Peroxisome proliferator-activated receptor gamma is a nuclear receptor traditionally associated with metabolic regulation and anti-inflammatory signaling; however, it has also been implicated in CNS functions, including emotion, stress, and pain modulation. It has been shown to be expressed in rat, mouse, monkey and human neurons, astrocytes, and rat microglial and oligodendrocyte cell cultures where it contributes to mitochondrial protection, redox balance, and modulation of neuroinflammatory responses [68,69,70,71,72,73,74]. In the present study, PPARγ-IR was detected in the lateral nucleus of the equine amygdala, localized to the nuclear compartment of both neurons and glial fibrillary acidic protein (GFAP)-positive glial cells. This distribution is consistent with its role as a ligand-activated transcription factor. Upon activation by selective agonists, such as rosiglitazone and pioglitazone, PPARγ forms a heterodimer with the retinoid X receptor and binds to peroxisome proliferator response elements. It has been shown to reduce anxiety and depression-like behavior, attenuate fear expression, and alleviate mechanical allodynia and thermal hyperalgesia by means of mechanisms involving both central and peripheral pathways [75,76]. Taken together, these findings reinforce the view that PPARγ plays a modulatory role in emotional and nociceptive processing. The Authors’ identification of PPARγ in both neurons and GFAP-positive glial cells of the lateral amygdaloid nucleus in the horse adds novel comparative neuroanatomical evidence to this growing body of literature, and supports a conserved role for PPARγ in the regulation of limbic functions across mammalian species.
Despite the well-known effects of cannabinoids on emotional state and memory, previous studies have not investigated in detail how cannabinoids may affect neuronal networks in the equine brain, including the amygdala. In the present experiment, the Authors focused on the lateral nucleus which is the best described region of the mammalian amygdala. The present investigation demonstrated the presence of the CB1, CB2, TRPV1 and PPARγ receptors in the equine amygdaloid complex for the first time. Furthermore, the present investigation showed that there was a very high density of CB1R axon terminals forming pericellular baskets presumably contacting pyramidal somata, suggesting that cannabinoids may reduce tonic GABAergic inhibitory control over pyramidal cells in the lateral nucleus. The distribution of other cannabinoid receptors in neuropilar and somatal structures supports strong cannabinoids modulations on amygdalar activities. Consistent with the present findings the BLA of primates [22] and rodents [21] contains many CCK-IR axon terminals containing the CB1R. These axons form a pericellular basket which contacts the somata of putative pyramidal cells and may be critical for fear expression and extinction [22,77]. Regarding the transcripts of the genes of interest, gene expression for Cnr1 and Cnr2, consistent with the protein (CB1 and CB2) data was detected. In contrast, TRPV1 and PPARγ transcripts were not detected in the fixed amygdala samples. However, since the Authors were able to detect these transcripts in non-fixed samples (preliminary data, not shown), and in line with the protein expression data, they suggested that the absence of TRPV1 and PPARγ transcripts in the fixed samples described in this manuscript was due to the known negative effect of formaldehyde fixation on RNA integrity as has been well documented in the literature [78,79].
4. Materials and Methods
The brains of six healthy horses, between 3 and 25 years of age, 4 female and 2 male, 4 slaughtered for human consumption, and 2 died from causes unrelated to the central nervous system, were removed from the skull post mortem. The breeds included 1 Avelignese, 1 Italian thoroughbred and 2 half-breeds, 2 warmbloods. As the abattoir procedure involves the longitudinal mechanical opening of the skull along its dorsal part, twelve halves of the brain were quickly removed from the skull. This procedure usually does not damage the most ventral part of the brain, the piriform lobe in which the amygdala is located. After severing the connections to some of the cranial nerves, especially the optic nerve which anchors the diencephalon to the sphenoid bone, the brains were removed from those skulls in which the entire neural anatomy of the brain was better preserved. The brains used in the current study came from a number of the same horses used in a previous study [32]. Horses showing no neurological symptoms or behavioral problems were considered to be healthy on the basis of a summary clinical examination. In accordance with Directive 2010/63/EU of the European Parliament and of the Council of 22 September 2010 regarding the protection of animals used for scientific purposes, Italian legislation (D. Lgs. n. 26/2014) does not require approval by the competent authorities or ethics committees as this study did not influence any therapeutic decisions.
4.1. RNA Isolation and Quantitative Real Time PCR (RT- PCR) for Cnr1, Cnr2, PPARγ and TRPV1
For gene expression analysis, total mRNA extraction was carried out using a RNeasy FFPE Kit (Qiagen Hilden, Germany) with a few modifications. Fifty mg of formalin fixed amigdala samples (n=6) were washed in phosphate-buffered saline (PBS) and cut finely with disposable scalpel. After adding 240ul of Buffer PKD, the samples were briefly centrifuged and Proteinase k (10ul) was added. The samples were incubated at 56°C for 4 hours; every 30 min, the samples were mixed/homogenized using a micro-pestle (Eppendorf, Hamburg, Germany). The samples were then incubated at 80°C for 15 min; the manufacturer’s protocol was followed up to elution (20ul). After spectrophotometric quantification, the total RNA (500 ng) was reverse transcribed to cDNA using 5X iScript RT Supermix (Bio-Rad Laboratories Inc., Hercules, CA, USA) at a final volume of 20 μL. To evaluate the gene expression profiles, RT-qPCR was carried out in a CFX96 thermal cycler (Bio-Rad Laboratories Inc.) using SYBR green detection to target the genes. Specific primers for horses (Zamith Cunha et al. 2023d) were used to evaluate the gene expression for the interest gene (IG) Cannabinoid receptors 1 and 2 (Cn1r and Cn2r), peroxisome proliferator-activated receptor gamma (PPARγ) and potential vanilloid type 1 (TRPV1) Primers for peroxisome proliferator-activated receptor gamma (PPARγ) were designed on a horse sequence (Accession Number KF788296, ) by using Beacon Designer 2.07 (Premier Biosoft International, Palo Alto, CA, USA) (For: 5’-CTAAAGAGCCTGAGAAAG-3’; Rev:5’- CCACTGAGAATAATGACA-3’): Regarding the reference genes (RG), GAPDH, HPRT and β-Act (beta Actin) were based on horse sequences as previously reported (Zannoni et al. 2014). All the amplification reactions were carried out in 20 µl and analyzed in duplicate; the reaction contained 10µl of iTaq Universal SYBR Green Supermix (Bio-Rad Laboratories Inc.), 0.8 µl of the forward and reverse primers (5mM each) of each target gene, 2 µl cDNA, and 7.2µl of water. The real-time procedure included an initial denaturation period of 3 min at 95°C, 40 cycles at 95°C for 15s, and 60°C for 30s, followed by a melting step with ramping from 55°C to 95°C at a rate of 0.5°C/10s. The specificity of the amplified PCR products was confirmed by agarose gel electrophoresis and melting curve analysis. The relative expression of the IGs were normalized based on the RGs. The relative mRNA expression of the genes tested was evaluated using the ΔCt method with ΔCt = (Ct RG– Ct IG) which directly correlated with the expression level (ΔCt values very negative, lower expression; ΔCt values less negative higher expression).
4.2. Immunofluorescence
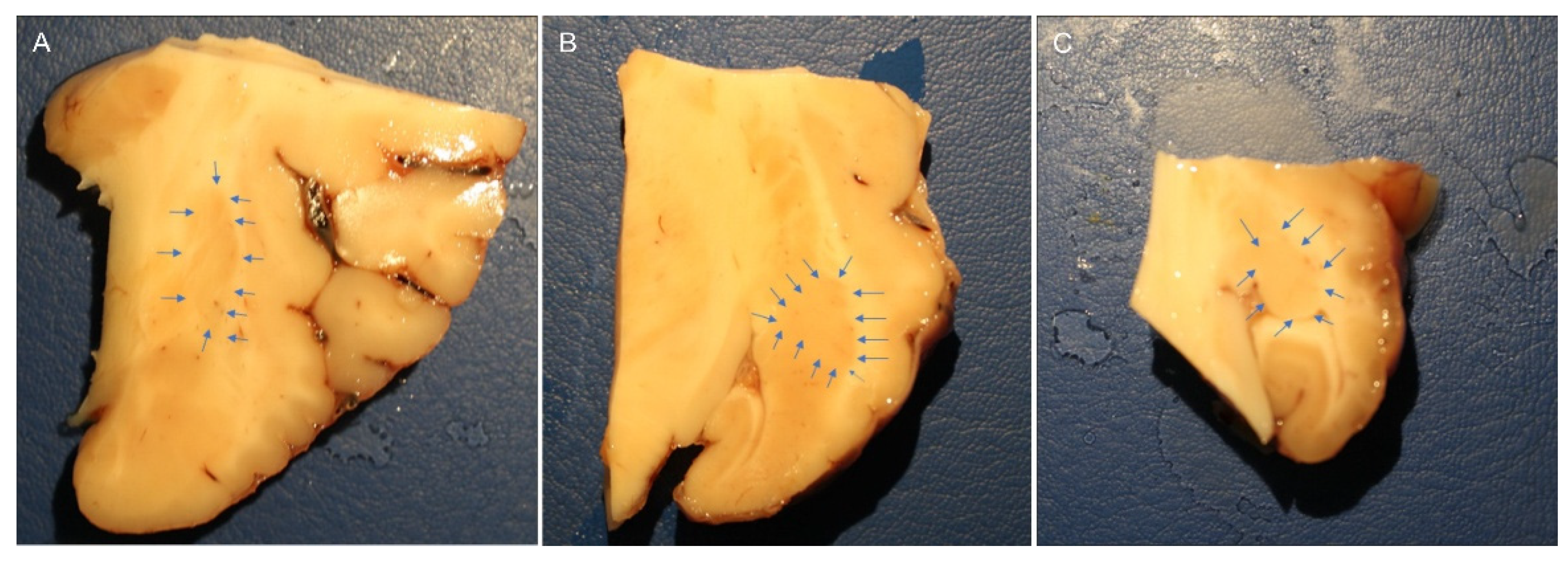
The brain halves were rapidly removed from the skulls of slaughtered horses and fixed for 48 hours at 4°C in 4% paraformaldehyde in phosphate buffer (0.1 M, pH 7.2), the piriform lobes containing the amygdales were then dissected to obtain smaller pieces of tissue fixed for an additional 24 hours. After a total fixation time of 72 hours, the tissues were rinsed in PBS (0.15M NaCl in 0.01M sodium phosphate buffer, pH 7.2) and stored at 4°C in PBS containing 30% sucrose and sodium azide (0.1%). On the following days, the tissues were transferred to a mixture of PBS-30% sucrose-azide and Optimal Cutting Temperature (OCT) compound (Sakura Finetek Europe, Alphen aan den Rijn, The Netherlands) in a 1:1 ratio for an additional 24 h before being embedded in 100% OCT in Cryomold®. Prior to embedding, each sample (approximately 2 x 3 cm) was divided into three parts along its longitudinal axis (rostral, central and caudal amygdala). In the current study, the central or rostral part of the amygdala (containing the lateral nucleus) was analyzed (Figure 7).
The sections were prepared by freezing the tissue in isopentane cooled in liquid nitrogen. Cryosections (14 μm thick) of the amygdala were cut on a cryostat and mounted on polylysinized slides. The cryosections were hydrated in PBS and processed for immunostaining. To block non-specific binding, the sections were incubated for 1 hour at RT (22-25°C) in a solution containing 20% normal donkey serum (Colorado Serum Co., Denver, CO, USA) , 0.5% Triton X-100g and bovine serum albumin (1%) in PBS. The cryosections were incubated overnight at RT in a humidity chamber with anti-CB1R, -CB2R, -TRPV1, and -PPARɣ; to identify the astrocytes, anti-glial fibrillary acidic protein (GFAP) antibodies were used (Table 1), diluted in a solution containing 20% normal donkey serum (Colorado Serum Co., Denver, CO, USA)f , 0.5% Triton X- 100g and bovine serum albumin (1%) in PBS. After washing in PBS (3 ×10 minutes), the sections were incubated for 1 hour at RT in a humidity chamber with secondary antibodies (Table 2) diluted in a solution containing 20% normal donkey serum (Colorado Serum Co., Denver, CO, USA), 0.5% Triton X-100g and bovine serum albumin (1%) in PBS. The cryosections were then washed in PBS (3 ×10 minutes) and processed for NeuroTrace® labeling (1:200) as a pan-neuronal marker in PBS for 40 minutes, followed by mounting in buffered glycerol at pH 8.6 with 4’,6-diamidino-2-phenylindole-DAPI (Santa Cruz Biotechnology, Santa Cruz, CA, USA).
4.3. Specificity of the Antibodies
The anti-CB1R, -TRPV1, and -PPARɣ antibodies had already been tested by the present research group using Western blot (Wb) analysis on horse tissue [80,81]. The rabbit anti-CB2R antibody (PA1-744) had already been tested by Wb analysis on horse tissues [82]. For negative controls, the sections were prepared by omitting primary antibodies and then incubating with the secondary antibodies. No stained cells were detected after omission of the primary antibodies.
4.4. Fluorescence Microscopy
The specimens were examined by the same observer using a Nikon Eclipse Ni microscope (Nikon Instruments Europe BV, Amsterdam, The Netherlands, Europe) equipped with the appropriate filter cubes. Images were captured using a DS-Qi1Nc digital camera and NIS Elements software BR 4.20.01 (Mountain View, Ottawa, ON, Canada). Minor contrast and brightness adjustments were made using Corel Photo Paint, while figure panels were prepared using Corel Draw.
4.5. Semiquantitative and Quantitative Analysis of the Immunofluorescence
The immunoreactivity of the antibodies was evaluated, and their cellular localization within the amygdala was reported. The intensity of expression was evaluated semi-quantitatively as weak, moderate and bright in images acquired at the same exposure times. The percentage of IR was evaluated quantitatively using ImageJ software and expressed as the percentage mean±SD calculated on eight sections (40x magnification) randomly selected within the rostro-caudal aspects of the lateral nucleus.
5. Conclusion
The present study is the first to demonstrate the genetic and immunohistochemical cellular distribution of the canonical CB1R, CB2R and two cannabinoid-related receptors (TRPV1, and PPARγ) in the equine amygdaloid complex. Due to their cellular localization, these receptors may be the target of many drugs (endocannabinoids and endocannabinoid-related molecules, non-psycoactive phytocannabinoids, synthetic cannabinoids, and various agonists and antagonists) which could potentially be used to improve anxiety, stress and pain in horses with behavioral problems and pathological conditions. These results should hopefully encourage the development of new molecular and preclinical studies to support the use of molecules already tested and used in humans and animals which could potentially reduce behavioral problems in horses. In-depth studies are needed in the equine species since it is known that cannabinoids can have different effects on anxiety depending on the species [83] .
The present results suggest that cannabinoids may reduce tonic inhibitory control over pyramidal cells in the lateral nucleus. Thus, the Authors suggest that the inhibition of the GABA release from the axon terminals of the local circuit GABAergic interneurons in the lateral nucleus by presynaptic CB1 receptors may be an important aspect of the neurobiological substrates of cannabinoid-induced emotional responses.
Author Contributions
R.C., G.S., C.B., and A.G. contributed to the study design. The mRNA analysis was carried out by A.Z. The immunohistochemical experiments were carried out by R.Z.C., M.D.S., C.T. and G.S. Acquisition of data: R.C., C.B. and A.G. All authors interpreted the data. Drafting of the manuscript: R.C., C.B., and G.S. All the authors contributed to the study execution and approved the final manuscript.
Funding
This study received a grant from NBF Lanes, Milan, Italy.
Acknowledgments
The authors would like to thank Fanny Mille for her valuable technical support in conducting immunohistochemical experiments.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Aggleton, J.P. The Amygdala: A Functional Analysis; Aggleton, J.P., Ed.; Oxford University Press: Oxford, 2000; ISBN 978-0-19-850501-3. [Google Scholar]
- Bombardi, C. Neuronal Localization of the 5-HT2 Receptor Family in the Amygdaloid Complex. Front. Pharmacol. 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Salamanca, G.; Tagliavia, C.; Grandis, A.; Graïc, J.M.; Cozzi, B.; Bombardi, C. Distribution of Vasoactive Intestinal Peptide (VIP) Immunoreactivity in the Rat Pallial and Subpallial Amygdala and Colocalization with γ-Aminobutyric Acid (GABA). The Anatomical Record 2024, 307, 2891–2911. [Google Scholar] [CrossRef] [PubMed]
- Pitkanen, A. Connectivity of the Rat Amygdaloid Complex. The amygdala, a functional analysis 2000. [Google Scholar]
- Guirado, S.; Real, M.Á.; Dávila, J.C. Distinct Immunohistochemically Defined Areas in the Medial Amygdala in the Developing and Adult Mouse. Brain Research Bulletin 2008, 75, 214–217. [Google Scholar] [CrossRef]
- Rambaldi, A. m.; Cozzi, B.; Grandis, A.; Canova, M.; Mazzoni, M.; Bombardi, C. Distribution of Calretinin Immunoreactivity in the Lateral Nucleus of the Bottlenose Dolphin (Tursiops Truncatus) Amygdala. The Anatomical Record 2017, 300, 2008–2016. [Google Scholar] [CrossRef]
- Sah, P.; Faber, E.S.L.; Lopez De Armentia, M.; Power, J. The Amygdaloid Complex: Anatomy and Physiology. Physiological Reviews 2003, 83, 803–834. [Google Scholar] [CrossRef]
- McGaugh, J.L. The Amygdala Modulates the Consolidation of Memories of Emotionally Arousing Experiences. Annu Rev Neurosci 2004, 27, 1–28. [Google Scholar] [CrossRef]
- LeDoux, J. The Amygdala. Current Biology 2007, 17, R868–R874. [Google Scholar] [CrossRef]
- Thompson, J.M.; Neugebauer, V. Amygdala Plasticity and Pain. Pain Research and Management 2017, 2017, 1–12. [Google Scholar] [CrossRef]
- Millan, M.J. The Neurobiology and Control of Anxious States. Progress in Neurobiology 2003, 70, 83–244. [Google Scholar] [CrossRef]
- Neugebauer, V.; Li, W.; Bird, G.C.; Han, J.S. The Amygdala and Persistent Pain. Neuroscientist 2004, 10, 221–234. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.C.; Shi, C.-Q.; Franaszczuk, P.J.; Crone, N.E.; Schretlen, D.; Ohara, S.; Lenz, F.A. Painful Laser Stimuli Induce Directed Functional Interactions within and between the Human Amygdala and Hippocampus. Neuroscience 2011, 178, 208–217. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, T.; Sugita, S. Histological Atlas and Morphological Features by Nissl Staining in the Amygdaloid Complex of the Horse, Cow and Pig. Journal of Equine Science 2007, 18, 13–25. [Google Scholar] [CrossRef]
- Hall, C.; Randle, H.; Pearson, G.; Preshaw, L.; Waran, N. Assessing Equine Emotional State. Applied Animal Behaviour Science 2018, 205, 183–193. [Google Scholar] [CrossRef]
- Kreitzer, F.R.; Stella, N. The Therapeutic Potential of Novel Cannabinoid Receptors. Pharmacology & Therapeutics 2009, 122, 83–96. [Google Scholar] [CrossRef]
- Lowin, T.; Pongratz, G.; Straub, R.H. The Synthetic Cannabinoid WIN55,212-2 Mesylate Decreases the Production of Inflammatory Mediators in Rheumatoid Arthritis Synovial Fibroblasts by Activating CB2, TRPV1, TRPA1 and yet Unidentified Receptor Targets. J Inflamm (Lond) 2016, 13, 15. [Google Scholar] [CrossRef]
- Ligresti, A.; De Petrocellis, L.; Di Marzo, V. From Phytocannabinoids to Cannabinoid Receptors and Endocannabinoids: Pleiotropic Physiological and Pathological Roles Through Complex Pharmacology. Physiol Rev 2016, 96, 1593–1659. [Google Scholar] [CrossRef]
- Morales, P.; Hurst, D.P.; Reggio, P.H. Molecular Targets of the Phytocannabinoids: A Complex Picture. In Phytocannabinoids; Kinghorn, A.D., Falk, H., Gibbons, S., Kobayashi, J., Eds.; Progress in the Chemistry of Organic Natural Products; Springer International Publishing: Cham, 2017; Vol. 103, pp. 103–131. ISBN 978-3-319-45539-6. [Google Scholar]
- Mlost, J.; Bryk, M.; Starowicz, K. Cannabidiol for Pain Treatment: Focus on Pharmacology and Mechanism of Action. IJMS 2020, 21, 8870. [Google Scholar] [CrossRef]
- McDonald, A.J.; Mascagni, F. Localization of the CB1 Type Cannabinoid Receptor in the Rat Basolateral Amygdala: High Concentrations in a Subpopulation of Cholecystokinin-Containing Interneurons. Neuroscience 2001, 107, 641–652. [Google Scholar] [CrossRef]
- McDonald, A.J. Expression of the Type 1 Cannabinoid Receptor (CB1R) in CCK-Immunoreactive Axon Terminals in the Basolateral Amygdala of the Rhesus Monkey (Macaca Mulatta). Neuroscience Letters 2021, 745, 135503. [Google Scholar] [CrossRef]
- Amaral, D.G. Anatomical Organization of the Primate Amygdaloid Complex. The amygdala: Neurobiological aspects of emotion, memory, and mental dysfunction 1992. [Google Scholar]
- Vásquez, C.E.; Reberger, R.; Dall’Oglio, A.; Calcagnotto, M.E.; Rasia-Filho, A.A. Neuronal Types of the Human Cortical Amygdaloid Nucleus. Journal of Comparative Neurology 2018, 526, 2776–2801. [Google Scholar] [CrossRef]
- McDonald, A.J. Chapter 1 - Functional Neuroanatomy of the Basolateral Amygdala: Neurons, Neurotransmitters, and Circuits. In Handbook of Behavioral Neuroscience; Urban, J.H., Rosenkranz, J.A., Eds.; Handbook of Amygdala Structure and Function; Elsevier, 2020; Vol. 26, pp. 1–38.
- McDonald, A.J.; Augustine, J.R. Nonpyramidal Neurons in the Primate Basolateral Amygdala: A Golgi Study in the Baboon (Papio Cynocephalus) and Long-Tailed Macaque (Macaca Fascicularis). Journal of Comparative Neurology 2020, 528, 772–786. [Google Scholar] [CrossRef]
- Muller, J.F.; Mascagni, F.; McDonald, A.J. Postsynaptic Targets of Somatostatin-Containing Interneurons in the Rat Basolateral Amygdala. Journal of Comparative Neurology 2007, 500, 513–529. [Google Scholar] [CrossRef] [PubMed]
- Mascagni, F.; McDonald, A.J. Immunohistochemical Characterization of Cholecystokinin Containing Neurons in the Rat Basolateral Amygdala. Brain research 2003, 976, 171–184. [Google Scholar] [CrossRef]
- Rainnie, D.G.; Mania, I.; Mascagni, F.; McDonald, A.J. Physiological and Morphological Characterization of Parvalbumin-Containing Interneurons of the Rat Basolateral Amygdala. J Comp Neurol 2006, 498, 142–161. [Google Scholar] [CrossRef] [PubMed]
- McDonald, A.J.; Pearson, J.C. Coexistence of GABA and Peptide Immunoreactivity in Non-Pyramidal Neurons of the Basolateral Amygdala. Neuroscience Letters 1989, 100, 53–58. [Google Scholar] [CrossRef]
- Degroot, A. Role of Cannabinoid Receptors in Anxiety Disorders. In Cannabinoids and the Brain; Köfalvi, A., Ed.; Springer US: Boston, MA, 2008; pp. 559–572. ISBN 978-0-387-74349-3. [Google Scholar]
- Zamith Cunha, R.; Zannoni, A.; Salamanca, G.; De Silva, M.; Rinnovati, R.; Gramenzi, A.; Forni, M.; Chiocchetti, R. Expression of Cannabinoid (CB1 and CB2) and Cannabinoid-Related Receptors (TRPV1, GPR55, and PPARα) in the Synovial Membrane of the Horse Metacarpophalangeal Joint. Frontiers in Veterinary Science 2023, 10. [Google Scholar] [CrossRef]
- Zamith Cunha, R.; Semprini, A.; Salamanca, G.; Gobbo, F.; Morini, M.; Pickles, K.J.; Roberts, V.; Chiocchetti, R. Expression of Cannabinoid Receptors in the Trigeminal Ganglion of the Horse. International Journal of Molecular Sciences 2023, 24, 15949. [Google Scholar] [CrossRef]
- Galiazzo, G.; De Silva, M.; Giancola, F.; Rinnovati, R.; Peli, A.; Chiocchetti, R. Cellular Distribution of Cannabinoid-related Receptors TRPV1, PPAR-gamma, GPR55 and GPR3 in the Equine Cervical Dorsal Root Ganglia. Equine Vet J 2022, 54, 788–798. [Google Scholar] [CrossRef]
- Chiocchetti, R.; Rinnovati, R.; Tagliavia, C.; Stanzani, A.; Galiazzo, G.; Giancola, F.; Silva, M.D.; Capodanno, Y.; Spadari, A. Localisation of Cannabinoid and Cannabinoid-Related Receptors in the Equine Dorsal Root Ganglia. Equine Vet J 2020. [Google Scholar] [CrossRef] [PubMed]
- Biscaia, M.; Marín, S.; Fernández, B.; Marco, E.M.; Rubio, M.; Guaza, C.; Ambrosio, E.; Viveros, M.P. Chronic Treatment with CP 55,940 during the Peri-Adolescent Period Differentially Affects the Behavioural Responses of Male and Female Rats in Adulthood. Psychopharmacology 2003, 170, 301–308. [Google Scholar] [CrossRef] [PubMed]
- Urigüen, L.; Pérez-Rial, S.; Ledent, C.; Palomo, T.; Manzanares, J. Impaired Action of Anxiolytic Drugs in Mice Deficient in Cannabinoid CB1 Receptors. Neuropharmacology 2004, 46, 966–973. [Google Scholar] [CrossRef]
- Pertwee, R.G. Pharmacology of Cannabinoid CB1 and CB2 Receptors. Pharmacology & Therapeutics 1997, 74, 129–180. [Google Scholar] [CrossRef]
- Rakhshan, F.; Day, T.A.; Blakely, R.D.; Barker, E.L. Carrier-Mediated Uptake of the Endogenous Cannabinoid Anandamide in RBL-2H3 Cells. J Pharmacol Exp Ther 2000, 292, 960–967. [Google Scholar] [CrossRef]
- Gunduz-Cinar, O. The Endocannabinoid System in the Amygdala and Modulation of Fear. Progress in Neuro-Psychopharmacology and Biological Psychiatry 2021, 105, 110116. [Google Scholar] [CrossRef]
- Gunduz-Cinar, O.; Castillo, L.I.; Xia, M.; Leer, E.V.; Brockway, E.T.; Pollack, G.A.; Yasmin, F.; Bukalo, O.; Limoges, A.; Oreizi-Esfahani, S.; et al. A Cortico-Amygdala Neural Substrate for Endocannabinoid Modulation of Fear Extinction. Neuron 2023, 111, 3053–3067.e10. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, A. Endocannabinoids Modulate Fear Extinction Controlled by a Cortical-Amygdala Projection. Neuron 2023, 111, 2948–2950. [Google Scholar] [CrossRef]
- Patel, S.; Cravatt, B.F.; Hillard, C.J. Synergistic Interactions between Cannabinoids and Environmental Stress in the Activation of the Central Amygdala. Neuropsychopharmacol 2005, 30, 497–507. [Google Scholar] [CrossRef]
- Marsicano, G.; Wotjak, C.T.; Azad, S.C.; Bisogno, T.; Rammes, G.; Cascio, M.G.; Hermann, H.; Tang, J.; Hofmann, C.; Zieglgänsberger, W.; et al. The Endogenous Cannabinoid System Controls Extinction of Aversive Memories. Nature 2002, 418, 530–534. [Google Scholar] [CrossRef]
- Azad, S.C.; Monory, K.; Marsicano, G.; Cravatt, B.F.; Lutz, B.; Zieglgänsberger, W.; Rammes, G. Circuitry for Associative Plasticity in the Amygdala Involves Endocannabinoid Signaling. J. Neurosci. 2004, 24, 9953–9961. [Google Scholar] [CrossRef] [PubMed]
- Rea, K.; Olango, W.M.; Harhen, B.; Kerr, D.M.; Galligan, R.; Fitzgerald, S.; Moore, M.; Roche, M.; Finn, D.P. Evidence for a Role of GABAergic and Glutamatergic Signalling in the Basolateral Amygdala in Endocannabinoid-Mediated Fear-Conditioned Analgesia in Rats. PAIN 2013, 154, 576–585. [Google Scholar] [CrossRef]
- Rubino, T.; Sala, M.; Viganò, D.; Braida, D.; Castiglioni, C.; Limonta, V.; Guidali, C.; Realini, N.; Parolaro, D. Cellular Mechanisms Underlying the Anxiolytic Effect of Low Doses of Peripheral Delta9-Tetrahydrocannabinol in Rats. Neuropsychopharmacology 2007, 32, 2036–2045. [Google Scholar] [CrossRef] [PubMed]
- Onaivi, E.S.; Ishiguro, H.; Gong, J.-P.; Pa℡, S.; Perchuk, A.; Meozzi, P.A.; Myers, L.; Mora, Z.; Tagliaferro, P.; Gardner, E.; et al. Discovery of the Presence and Functional Expression of Cannabinoid CB2 Receptors in Brain. Annals of the New York Academy of Sciences 2006, 1074, 514–536. [Google Scholar] [CrossRef]
- Cabral, G.A.; Raborn, E.S.; Griffin, L.; Dennis, J.; Marciano-Cabral, F. CB2 Receptors in the Brain: Role in Central Immune Function. British Journal of Pharmacology 2008, 153, 240–251. [Google Scholar] [CrossRef] [PubMed]
- Brusco, A.; Tagliaferro, P.; Saez, T.; Onaivi, E.S. Postsynaptic Localization of CB2 Cannabinoid Receptors in the Rat Hippocampus. Synapse 2008, 62, 944–949. [Google Scholar] [CrossRef]
- Roche, M.; Finn, D.P. Brain CB2 Receptors: Implications for Neuropsychiatric Disorders. Pharmaceuticals 2010, 3, 2517–2553. [Google Scholar] [CrossRef]
- Atwood, B.K.; Mackie, K. CB2: A Cannabinoid Receptor with an Identity Crisis. British Journal of Pharmacology 2010, 160, 467–479. [Google Scholar] [CrossRef]
- Bahi, A.; Al Mansouri, S.; Al Memari, E.; Al Ameri, M.; Nurulain, S.M.; Ojha, S. β-Caryophyllene, a CB2 Receptor Agonist Produces Multiple Behavioral Changes Relevant to Anxiety and Depression in Mice. Physiology & Behavior 2014, 135, 119–124. [Google Scholar] [CrossRef]
- Ramikie, T.S.; Patel, S. Endocannabinoid Signaling in the Amygdala: Anatomy, Synaptic Signaling, Behavior, and Adaptations to Stress. Neuroscience 2012, 204, 38–52. [Google Scholar] [CrossRef]
- García-Gutiérrez, M.S.; Manzanares, J. Overexpression of CB2 Cannabinoid Receptors Decreased Vulnerability to Anxiety and Impaired Anxiolytic Action of Alprazolam in Mice. J Psychopharmacol 2011, 25, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Manning, B.H.; Martin, W.J.; Meng, I.D. The Rodent Amygdala Contributes to the Production of Cannabinoid-Induced Antinociception. Neuroscience 2003, 120, 1157–1170. [Google Scholar] [CrossRef]
- Argue, K.J.; VanRyzin, J.W.; Falvo, D.J.; Whitaker, A.R.; Yu, S.J.; McCarthy, M.M. Activation of Both CB1 and CB2 Endocannabinoid Receptors Is Critical for Masculinization of the Developing Medial Amygdala and Juvenile Social Play Behavior. eNeuro 2017, 4. [Google Scholar] [CrossRef]
- Segev, A.; Akirav, I. Cannabinoids and Glucocorticoids in the Basolateral Amygdala Modulate Hippocampal–Accumbens Plasticity After Stress. Neuropsychopharmacol 2016, 41, 1066–1079. [Google Scholar] [CrossRef]
- Zschenderlein, C.; Gebhardt, C.; Halbach, O. von B. und; Kulisch, C.; Albrecht, D. Capsaicin-Induced Changes in LTP in the Lateral Amygdala Are Mediated by TRPV1. PLOS ONE 2011, 6, e16116. [Google Scholar] [CrossRef]
- Marsch, R.; Foeller, E.; Rammes, G.; Bunck, M.; Kössl, M.; Holsboer, F.; Zieglgänsberger, W.; Landgraf, R.; Lutz, B.; Wotjak, C.T. Reduced Anxiety, Conditioned Fear, and Hippocampal Long-Term Potentiation in Transient Receptor Potential Vanilloid Type 1 Receptor-Deficient Mice. J. Neurosci. 2007, 27, 832–839. [Google Scholar] [CrossRef]
- Santos, C.J.P.A.; Stern, C.A.J.; Bertoglio, L.J. Attenuation of Anxiety-Related Behaviour after the Antagonism of Transient Receptor Potential Vanilloid Type 1 Channels in the Rat Ventral Hippocampus. Behavioural Pharmacology 2008, 19, 357. [Google Scholar] [CrossRef] [PubMed]
- Marsicano, G.; Kuner, R. Anatomical Distribution of Receptors, Ligands and Enzymes in the Brain and in the Spinal Cord: Circuitries and Neurochemistry. In Cannabinoids and the Brain; Köfalvi, A., Ed.; Springer US: Boston, MA, 2008; pp. 161–201. ISBN 978-0-387-74349-3. [Google Scholar]
- Kirkedal, C.; Wegener, G.; Moreira, F.; Joca, S.R.L.; Liebenberg, N. A Dual Inhibitor of FAAH and TRPV1 Channels Shows Dose-Dependent Effect on Depression-like Behaviour in Rats. Acta Neuropsychiatrica 2017, 29, 324–329. [Google Scholar] [CrossRef] [PubMed]
- Frias, B.; Merighi, A. Capsaicin, Nociception and Pain. Molecules 2016, 21, 797. [Google Scholar] [CrossRef]
- Nagy, I.; White, J.P.M.; Paule, C.C.; Maze, M.; Urban, L. Functional Molecular Biology of the TRPV1 Ion Channel. In Cannabinoids and the Brain; Köfalvi, A., Ed.; Springer US: Boston, MA, 2008; pp. 101–130. ISBN 978-0-387-74349-3. [Google Scholar]
- Xiao, Y.; Chen, X.; Zhang, P.-A.; Xu, Q.; Zheng, H.; Xu, G.-Y. TRPV1-Mediated Presynaptic Transmission in Basolateral Amygdala Contributes to Visceral Hypersensitivity in Adult Rats with Neonatal Maternal Deprivation. Sci Rep 2016, 6, 29026. [Google Scholar] [CrossRef]
- Edwards, J.G. TRPV1 in the Central Nervous System: Synaptic Plasticity, Function, and Pharmacological Implications. Prog Drug Res 2014, 68, 77–104. [Google Scholar] [CrossRef] [PubMed]
- Zolezzi, J.M.; Santos, M.J.; Bastías-Candia, S.; Pinto, C.; Godoy, J.A.; Inestrosa, N.C. PPARs in the Central Nervous System: Roles in Neurodegeneration and Neuroinflammation. Biological Reviews 2017, 92, 2046–2069. [Google Scholar] [CrossRef] [PubMed]
- Cristiano, L.; Bernardo, A.; Cerù, M.P. Peroxisome Proliferator-Activated Receptors (PPARs) and Peroxisomes in Rat Cortical and Cerebellar Astrocytes. J Neurocytol 2001, 30, 671–683. [Google Scholar] [CrossRef] [PubMed]
- Moreno, S.; Farioli-Vecchioli, S.; Cerù, M.P. Immunolocalization of Peroxisome Proliferator-Activated Receptors and Retinoid X Receptors in the Adult Rat CNS. Neuroscience 2004, 123, 131–145. [Google Scholar] [CrossRef]
- Warden, A.; Truitt, J.; Merriman, M.; Ponomareva, O.; Jameson, K.; Ferguson, L.B.; Mayfield, R.D.; Harris, R.A. Localization of PPAR Isotypes in the Adult Mouse and Human Brain. Sci Rep 2016, 6, 27618. [Google Scholar] [CrossRef] [PubMed]
- Swanson, C.R.; Emborg, M.E. Expression of Peroxisome Proliferator-Activated Receptor-γ in the Substantia Nigra of Hemiparkinsonian Nonhuman Primates. Neurol Res 2014, 36, 634–646. [Google Scholar] [CrossRef]
- Bernardo, A.; Levi, G.; Minghetti, L. Role of the Peroxisome Proliferator-Activated Receptor-γ (PPAR-γ) and Its Natural Ligand 15-Deoxy-Δ12,14-Prostaglandin J2 in the Regulation of Microglial Functions. European Journal of Neuroscience 2000, 12, 2215–2223. [Google Scholar] [CrossRef]
- Roth, A.D.; Leisewitz, A.V.; Jung, J.E.; Cassina, P.; Barbeito, L.; Inestrosa, N.C.; Bronfman, M. PPAR Gamma Activators Induce Growth Arrest and Process Extension in B12 Oligodendrocyte-like Cells and Terminal Differentiation of Cultured Oligodendrocytes. J Neurosci Res 2003, 72, 425–435. [Google Scholar] [CrossRef]
- Gemma, C.; Stellwagen, H.; Fister, M.; Coultrap, S.J.; Mesches, M.H.; Browning, M.D.; Bickford, P.C. Rosiglitazone Improves Contextual Fear Conditioning in Aged Rats. Neuroreport 2004, 15, 2255–2259. [Google Scholar] [CrossRef]
- Gaspar, J.C.; Okine, B.N.; Dinneen, D.; Roche, M.; Finn, D.P. Effects of Intra-BLA Administration of PPAR Antagonists on Formalin-Evoked Nociceptive Behaviour, Fear-Conditioned Analgesia, and Conditioned Fear in the Presence or Absence of Nociceptive Tone in Rats. Molecules 2022, 27, 2021. [Google Scholar] [CrossRef]
- McDonald, A.J.; Mascagni, F. Localization of the CB1 Type Cannabinoid Receptor in the Rat Basolateral Amygdala: High Concentrations in a Subpopulation of Cholecystokinin-Containing Interneurons. Neuroscience 2001, 107, 641–652. [Google Scholar] [CrossRef] [PubMed]
- Evers, D.L.; Fowler, C.B.; Cunningham, B.R.; Mason, J.T.; O’Leary, T.J. The Effect of Formaldehyde Fixation on RNA. The Journal of Molecular Diagnostics 2011, 13, 282–288. [Google Scholar] [CrossRef] [PubMed]
- Jones, W.; Greytak, S.; Odeh, H.; Guan, P.; Powers, J.; Bavarva, J.; Moore, H.M. Deleterious Effects of Formalin-Fixation and Delays to Fixation on RNA and miRNA-Seq Profiles. Sci Rep 2019, 9, 6980. [Google Scholar] [CrossRef] [PubMed]
- Chiocchetti, R.; Rinnovati, R.; Tagliavia, C.; Stanzani, A.; Galiazzo, G.; Giancola, F.; Silva, M.D.; Capodanno, Y.; Spadari, A. Localisation of Cannabinoid and Cannabinoid-related Receptors in the Equine Dorsal Root Ganglia. Equine Vet J 2021, 53, 549–557. [Google Scholar] [CrossRef]
- Galiazzo, G.; Tagliavia, C.; Giancola, F.; Rinnovati, R.; Sadeghinezhad, J.; Bombardi, C.; Grandis, A.; Pietra, M.; Chiocchetti, R. Localisation of Cannabinoid and Cannabinoid-Related Receptors in the Horse Ileum. J Equine Vet Sci 2021, 104, 103688. [Google Scholar] [CrossRef]
- Kupczyk, P.; Rykala, M.; Serek, P.; Pawlak, A.; Slowikowski, B.; Holysz, M.; Chodaczek, G.; Madej, J.P.; Ziolkowski, P.; Niedzwiedz, A. The Cannabinoid Receptors System in Horses: Tissue Distribution and Cellular Identification in Skin. J Vet Intern Med 2022, 36, 1508–1524. [Google Scholar] [CrossRef]
- Haller, J. Anxiety Modulation by Cannabinoids—The Role of Stress Responses and Coping. International Journal of Molecular Sciences 2023, 24, 15777. [Google Scholar] [CrossRef]
Figure 1.
Gene expression of Cnr1 and Cnr2 in the equine fixed amygdala samples. The results are presented as ΔCt = (Ct RG– Ct IG), (ΔCt values less negative, higher expression). For each gene, mean ± SD are indicated by horizontal bars. No significant statistical difference (P < 0.05) was observed between Cnr1 and Cnr2 gene expression (Student’s t test). RG = Reference Gene; IG= Interest Gene.
Figure 1.
Gene expression of Cnr1 and Cnr2 in the equine fixed amygdala samples. The results are presented as ΔCt = (Ct RG– Ct IG), (ΔCt values less negative, higher expression). For each gene, mean ± SD are indicated by horizontal bars. No significant statistical difference (P < 0.05) was observed between Cnr1 and Cnr2 gene expression (Student’s t test). RG = Reference Gene; IG= Interest Gene.
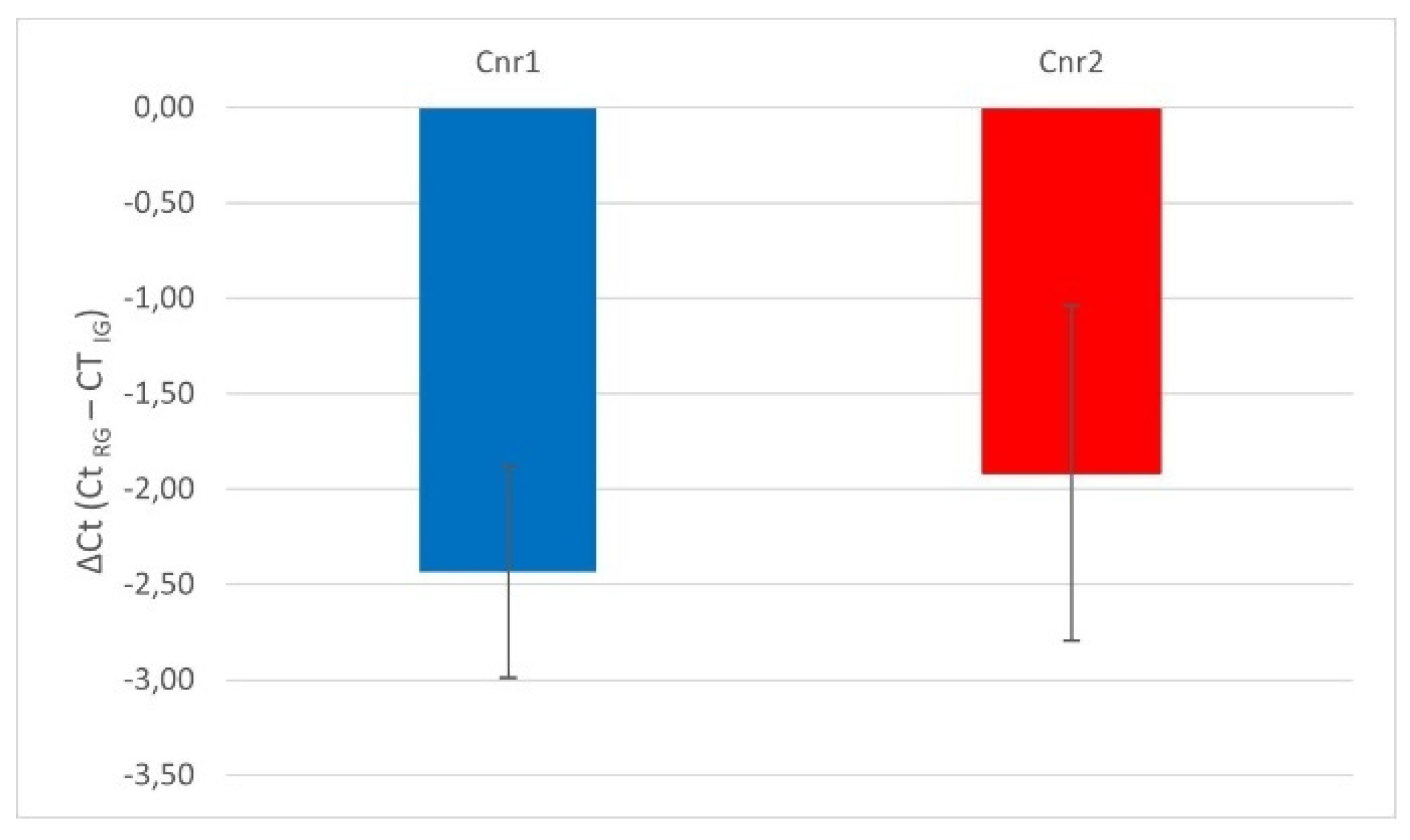
Figure 2.
Photomicrographs of the cryosections of the lateral nucleus of the amygdala of a horse showing bright cannabinoid receptor type 1 (CB1R) immunoreactivity in thin nerve fibers (white fibers) of the neuropil and varicosities (open arrows). The stars indicate some neuronal cell bodies labeled with Neurotrace (a, d) which were CB1R negative (b, e); the open arrows indicate baskets of bright CB1R immunoreactive varicosities encircling the neuronal cell bodies. The small white arrows (d-f) indicate neuropil fibers. c,f) Merged image. Scheme 50. m.
Figure 2.
Photomicrographs of the cryosections of the lateral nucleus of the amygdala of a horse showing bright cannabinoid receptor type 1 (CB1R) immunoreactivity in thin nerve fibers (white fibers) of the neuropil and varicosities (open arrows). The stars indicate some neuronal cell bodies labeled with Neurotrace (a, d) which were CB1R negative (b, e); the open arrows indicate baskets of bright CB1R immunoreactive varicosities encircling the neuronal cell bodies. The small white arrows (d-f) indicate neuropil fibers. c,f) Merged image. Scheme 50. m.
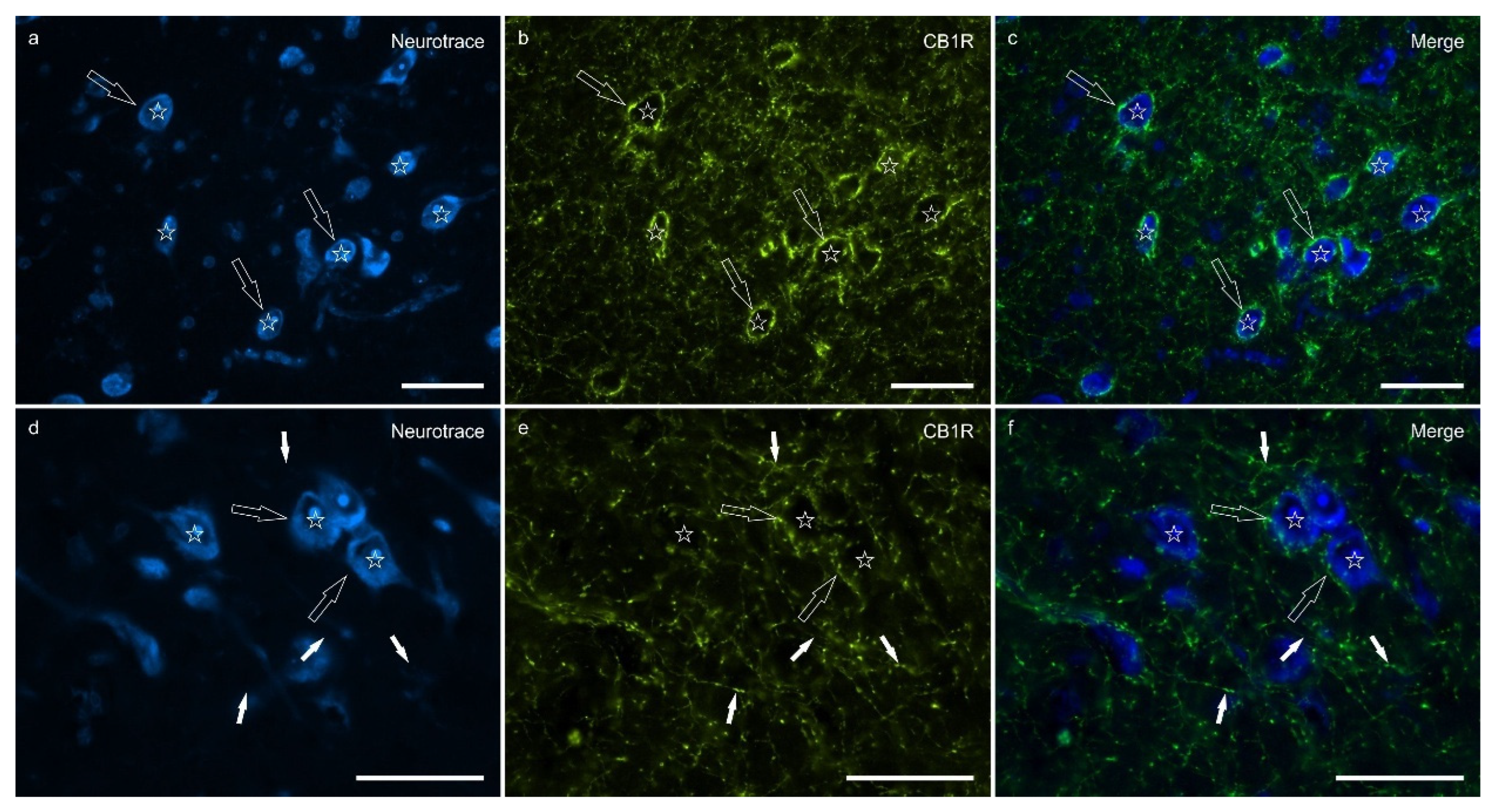
Figure 3.
Photomicrographs of the cryosections of the lateral nucleus of the amygdala of a horse showing weak cannabinoid receptor type 2 (CB2R) immunoreactivity in the neuropil varicosities. The stars indicate some neuronal cell bodies labeled with Neurotrace (a, d) which were CB2R negative (b, e); the white arrows indicate a deposit of autofluorescent pigment within the neuronal cell bodies. Note the diffuse punctate immunoreactivity. c,f) Merged image. Scale bar = 50µm.
Figure 3.
Photomicrographs of the cryosections of the lateral nucleus of the amygdala of a horse showing weak cannabinoid receptor type 2 (CB2R) immunoreactivity in the neuropil varicosities. The stars indicate some neuronal cell bodies labeled with Neurotrace (a, d) which were CB2R negative (b, e); the white arrows indicate a deposit of autofluorescent pigment within the neuronal cell bodies. Note the diffuse punctate immunoreactivity. c,f) Merged image. Scale bar = 50µm.
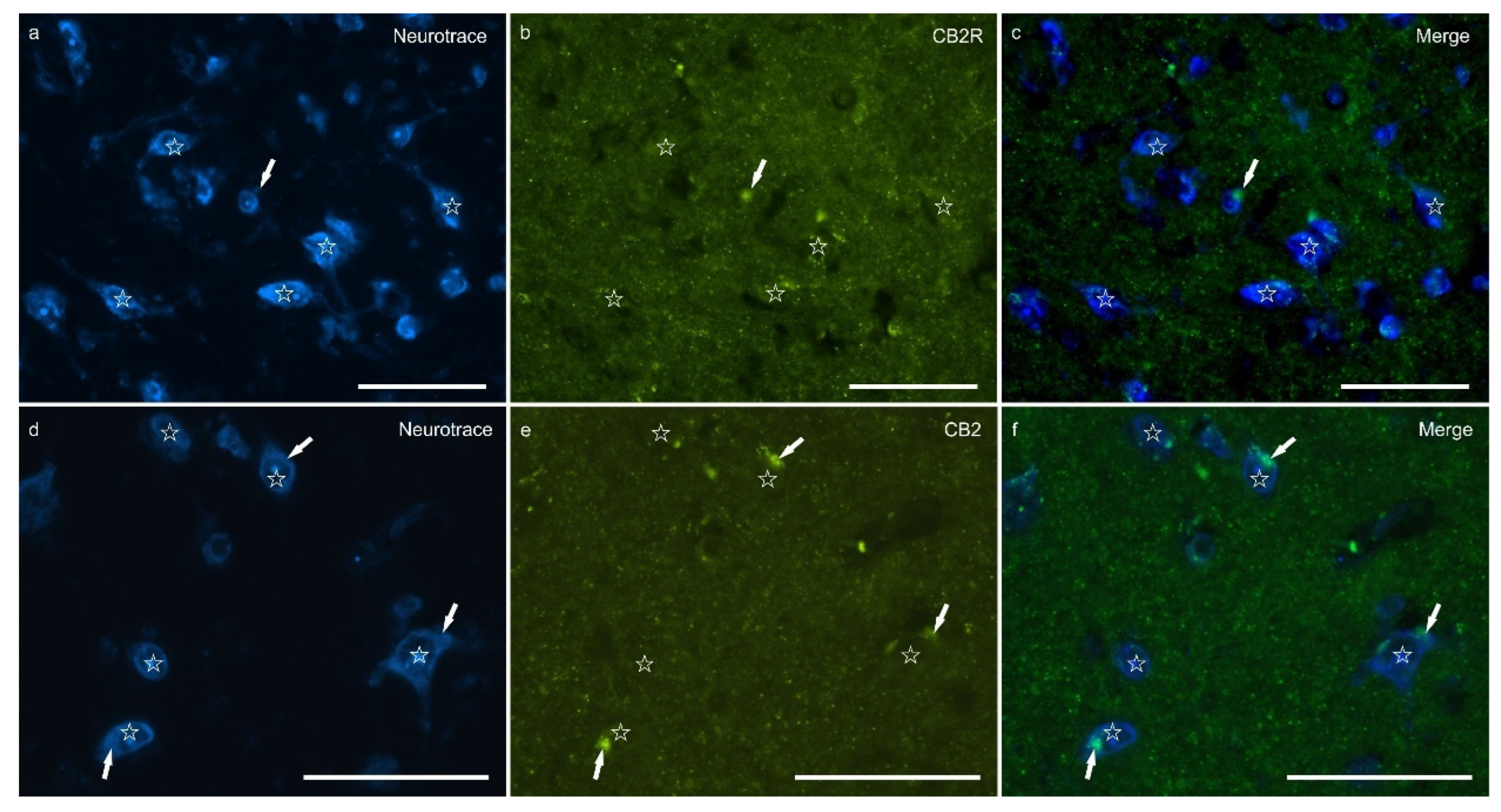
Figure 4.
Photomicrographs of the cryosections of the lateral nucleus of the amygdala of a horse showing moderate-to-bright transient receptor potential vanilloid 1 (TRPV1) immunoreactivity (b, e) in neurotrace labeled neuronal cell bodies (a, d) (stars), their nerve processes (open arrows) and the varicosities of the neuropil. c,f) Merged image. Scale bar = 50µm.
Figure 4.
Photomicrographs of the cryosections of the lateral nucleus of the amygdala of a horse showing moderate-to-bright transient receptor potential vanilloid 1 (TRPV1) immunoreactivity (b, e) in neurotrace labeled neuronal cell bodies (a, d) (stars), their nerve processes (open arrows) and the varicosities of the neuropil. c,f) Merged image. Scale bar = 50µm.
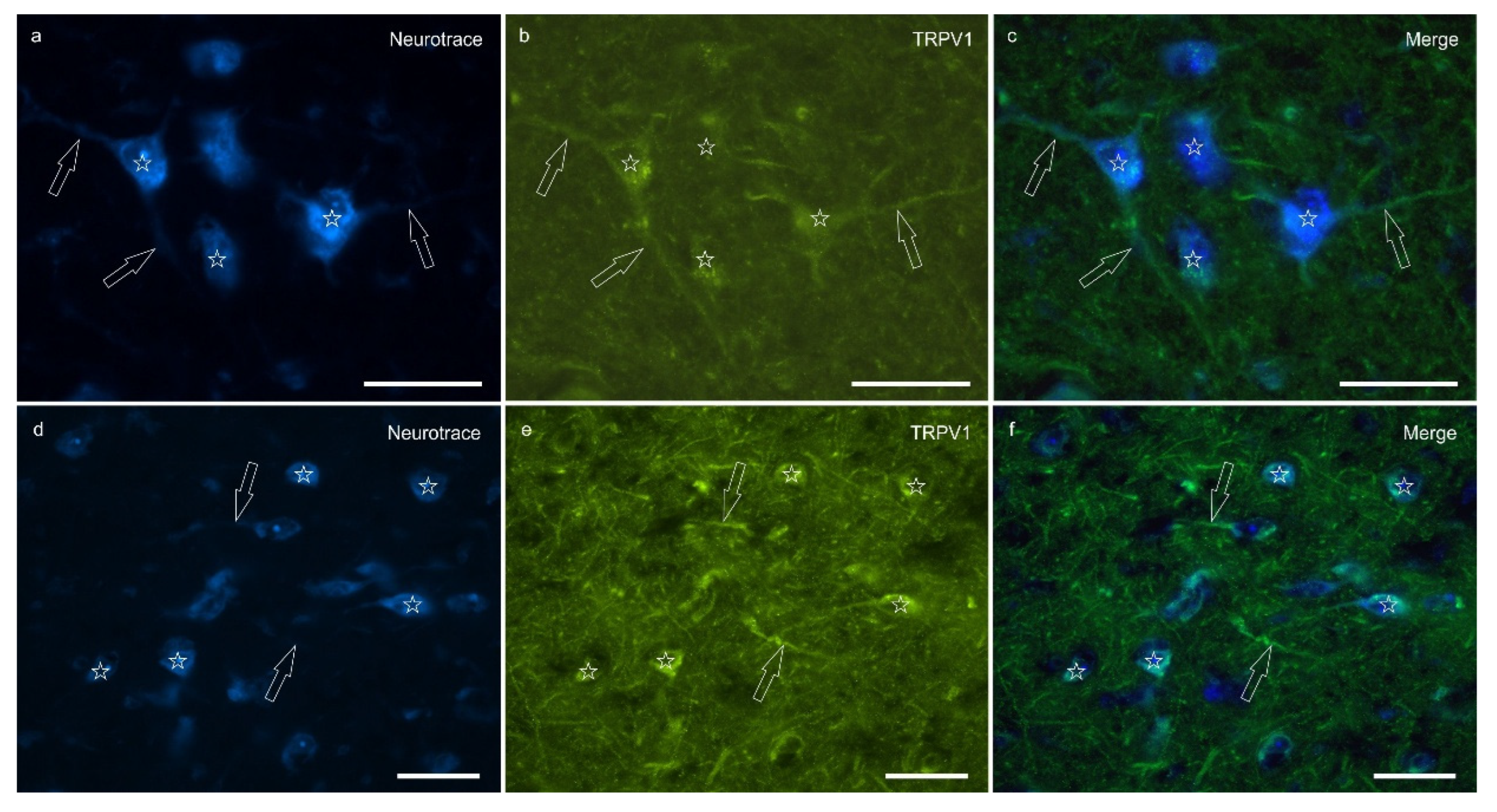
Figure 5.
Photomicrographs of the cryosections of the lateral nucleus of the amygdala of a horse. a-c) The stars indicate some neurotrace-labeled neurons (a) expressing bright nuclear peroxisome proliferator-activated receptor gamma immunoreactivity (PPARɣ) (b). The open arrows indicate moderate PPARɣ immunoreactivity in the smaller nuclei of the glial cells. d-f) The open arrows indicate two glial fibrillary acidic protein (GFAP) immunoreacrtive glial cells co-expressing moderate nuclear PPARɣ immunoreactivity (d). The arrows indicate the nuclei of GFAP negative cells. c,f) Merged image. Scale bar = 50µm.
Figure 5.
Photomicrographs of the cryosections of the lateral nucleus of the amygdala of a horse. a-c) The stars indicate some neurotrace-labeled neurons (a) expressing bright nuclear peroxisome proliferator-activated receptor gamma immunoreactivity (PPARɣ) (b). The open arrows indicate moderate PPARɣ immunoreactivity in the smaller nuclei of the glial cells. d-f) The open arrows indicate two glial fibrillary acidic protein (GFAP) immunoreacrtive glial cells co-expressing moderate nuclear PPARɣ immunoreactivity (d). The arrows indicate the nuclei of GFAP negative cells. c,f) Merged image. Scale bar = 50µm.
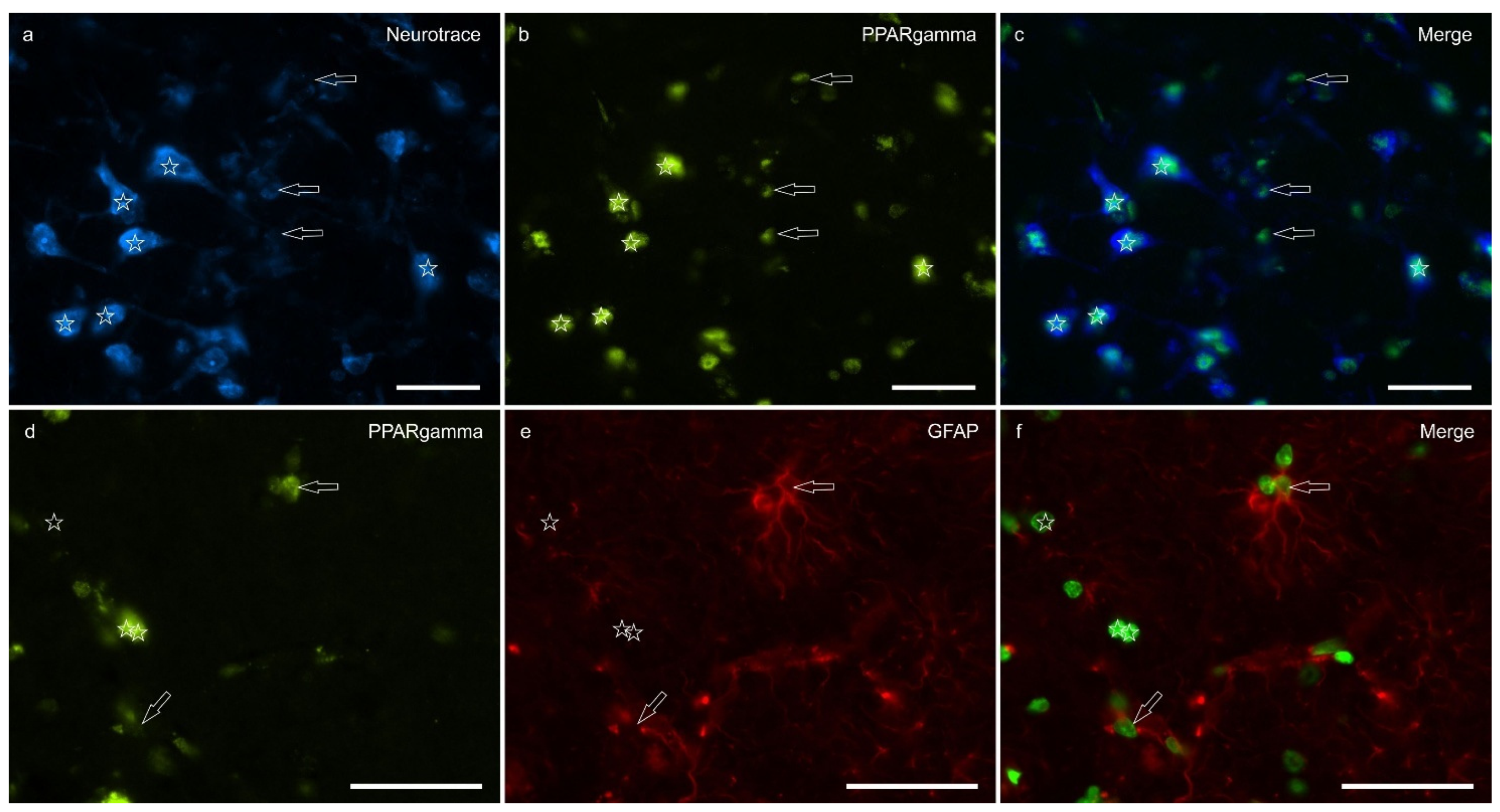
Figure 6.
Percentages of immunostaining within the lateral nucleus of the horse amygdala of CB1R, CB2R, TPRV1 and PPARɣ, calculated using ImageJ software.
Figure 6.
Percentages of immunostaining within the lateral nucleus of the horse amygdala of CB1R, CB2R, TPRV1 and PPARɣ, calculated using ImageJ software.
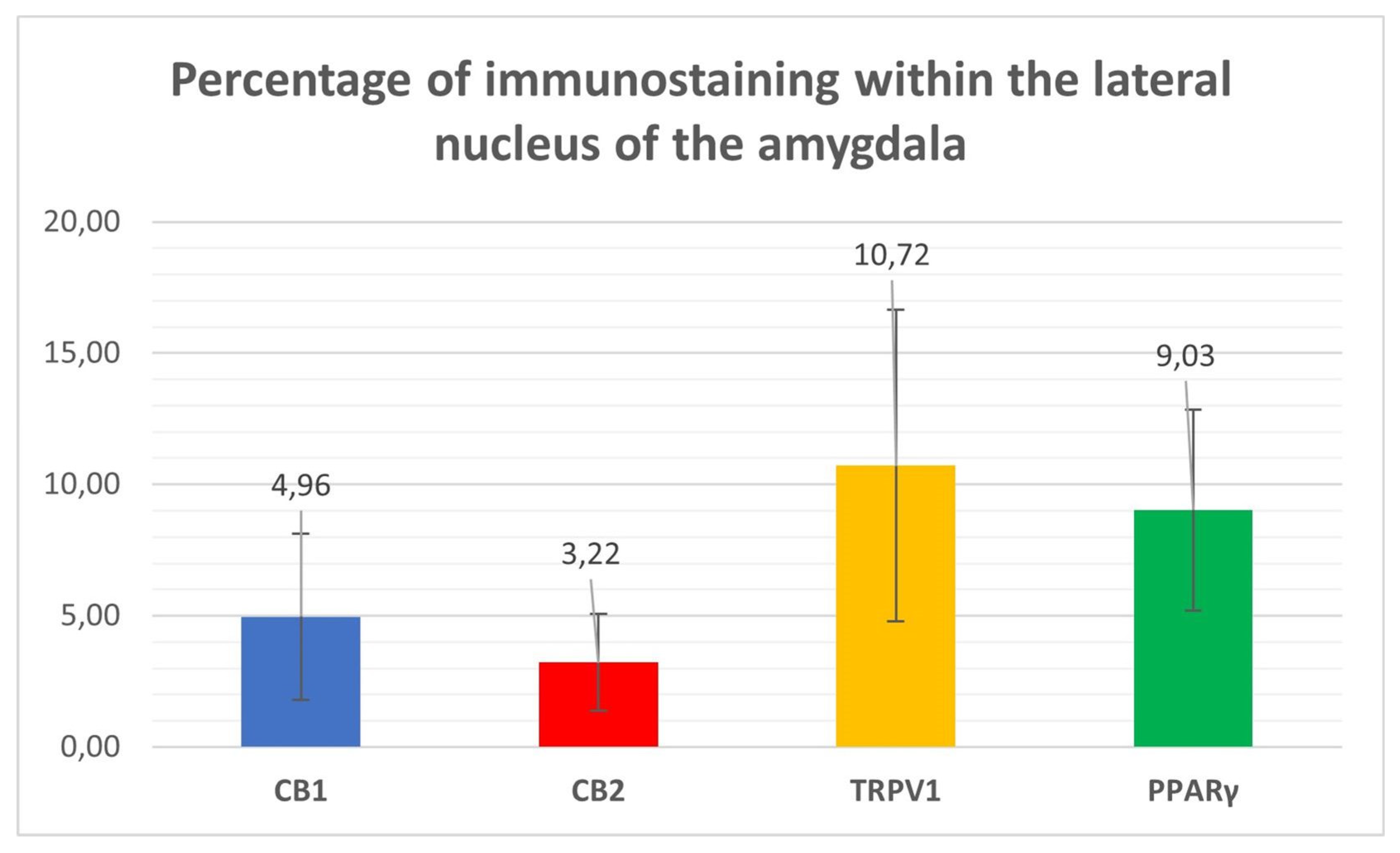
Figure 7.
Macroscopic view of the right horse amygdaloid complex in a coronal section at the rostral (A), middle (B), and caudal (C) levels. The lateral nucleus boundary is indicated by the arrows.
Figure 7.
Macroscopic view of the right horse amygdaloid complex in a coronal section at the rostral (A), middle (B), and caudal (C) levels. The lateral nucleus boundary is indicated by the arrows.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



